Submitted:
16 July 2024
Posted:
17 July 2024
You are already at the latest version
Abstract
Glioblastoma is a highly aggressive brain tumor and is considered to be the most common primary one. Recurrence after treatment is a major problem. The recurrence of GBM is related to different cellular mechanisms and molecular signaling. Therapies to prolong survival are not completely effective. Epigenetics and non-coding genetic elements play a crucial role in gliomagenesis. Non-coding RNAs are novel regulatory RNAs that play key roles in different physiological processes such as gene regulation, cell differentiation, and proliferation. Interestingly, some LncRNAs have a tumor suppressor role while others are oncogenic. In this review article, we are going to illustrate the role of a well-known lncRNA HOX transcript antisense intergenic RNA (HOTAIR) in glioma and highlight the possible functions in glioma in both pre-clinical and clinically evident studies.
Keywords:
HOTAIR
; ncRNAs
; GBM
; high grade glioma
; epigenetics
Introduction
Glioblastoma (GBM) is an aggressive primary brain tumor [1]. Recurrence is a major problem with a survival rate after one year of about 39.7% [2]. The recurrence of GBM is a complex multifactorial process. Most clinical trials failed to promote prolonged survival, the best outcome was reported with the European Organization for Research and Treatment of Cancer (EORTC) and National Cancer Institute of Canada (NCIC) clinical trials in 2005 [3] Epigenetics and non-coding genetic elements are extensively involved in the virulence of this cancer. Many factors contribute to treatment failure such as the heterogeneity of the GBM microenvironment, repository of stem cells with great regenerative potential, and developing resistance to common therapies.
Non-coding RNAs (ncRNAs) are recent classes of RNA molecules that play essential roles in different physiological processes as gene regulation, cell differentiation ,and growth[4]. The non-coding elements represent a big portion of the human genome, however, its main functions are still poorly understood [5,6]. The mechanism through which ncRNA regulates certain biological processes needs to be more elucidated. Non-coding RNA is classified into small and long non-coding RNA according to the nucleotide sequence. Small ncRNAs (20-200 nucleotides) include microRNAs (miRNAs), small nuclear RNAs (snRNAs), small interfering RNAs (siRNAs), small nucleolar RNAs (snoRNAs), Piwi-interacting RNAs (piRNAs) [7].
Long non-coding RNAs are composed of more than 200 nucleotides and could control genes that regulate the cell cycle, apoptosis, and cellular growth in various tissues [8]. Mounting research suggests the role of LncRNAs in different cancers including glioma [9]. For example, a lncRNA nuclear enriched abundant transcript 1 (NEAT1) through manipulating miRNA-449b-5p could enhance invasion of GBM cells and inhibit apoptosis [9]. Another lncRNA H19 was found to correlate with glioma grade and control invasiveness [10]. The tumor suppressor candidate 7 (TUSC7), is a LncRNA that suppresses invasion and migration of glioma cells and correlates with prognosis [10,11]. Some LncRNAs could work as tumor suppressor factors while others are oncogenic. In this review article, we are going to highlight the role of a well-known LncRNA HOX transcript antisense intergenic RNA (HOTAIR) in glioma and highlight the possible biological roles in glioma in both pre-clinical and clinically evident studies.
Mechanism of HOTAIR function:
The lncRNA HOX transcript antisense intergenic RNA (HOTAIR) was the first lncRNA to be identified [12]. It is one of the homeobox superfamilies and comprises 2158 nucleotides. It is transcribed from the HOXC locus on chromosome 12q13.13 [13]. Chromatin modifying complex as polycomb-repressive complex 2 (PRC2) is a binding target for HOTAIR [14]. PRC2 complex induces lysine methylation on histone H3. H3K27-methylation is a form of gene silencing and is assisted by histone methyltransferase EZH2 (Enhancer of Zeste homolog 2) [15]. HOTAIR can silence different genes through interaction with histone lysine-specific demethylase (KDM1) [16]. KDM1 can be combined with REST (RE1-Silencing Transcription factor) and CoREST to promote gene silencing [16]. Histone methylation and demethylation is a dynamic epigenetic feature that controls gene regulation. In early embryo life, HOTAIR is expressed in the genital bud, hind limb bud, and posterior trunk [17]. HOTAIR also can regulate the cell cycle proteins, by adjusting the expression of cyclin-dependent kinase 2 (CDK2), CDK4, and cyclin D1 [17]. Aberrant HOTAIR expression has been reported in various tumors, and HOTAIR expression has been correlated with growth, progression, recurrence, and poor prognosis by affecting downstream targets [18,19,20].
Molecular Interactions involving HOTAIR in GBM:
HOTAIR has oncogenic potential in different cancers like breast and renal cancer by enhancing cell proliferation, suppressing apoptosis, and promoting cellular invasion [21,22]. We will highlight the potential mechanisms that involve HOTAIR as a preoncogenic candidate in glioma. HOTAIR was found to be highly expressed in glioma compared to normal brain tissues [23]. A certain study showed that HOTAIR knockdown disrupted mouse intracranial GBM model [24]. A group of investigators observed that HOTAIR is highly expressed in both classic and mesenchymal glioma subtypes compared to neural and proneural subtypes [25]. They identified HOTAIR as a marker that correlates for tumor grade and outcome given the fact that low-grade glioma has lower expression levels of HOTAIR compared with high-grade tumors [25]. Different studies investigating the role of HOTAIR in GBM are summarized in Table 1.
Several transcription factors control HOTAIR expression in glioma. HOXA 9 stimulates the expression of HOTAIR in glioma [23]. The upregulation of HOXA9 was associated with abnormaly aggressive behavior [26]. As mentioned before, HOTAIR can induce gene silencing depending on EZH2, meanwhile, HOXA9 is regulated by the PI3K pathway and the inhibition of EZH2-mediated histone methylation creating a loop between HOTAIR and HOXA9 [23,27].
Another study evaluated the role of programmed cell death protein 4 (PDCD4) in the progression of GBM and found that overexpression of PDCD4 in glioma cells down regulated cellular proliferation suggesting that PDCD4 could function as a tumor suppressor [28]. Lower expression levels of PDCD4 are associated with a high level of histone h3 methylation, which is mediated by HOTAIR [28].
Exposure of glioma cells to a BET inhibitor (I-BET151) downregulated the expression of HOTAIR and stopped cell proliferation through cell cycle arrest. Moreover, the upregulation of HOTAIR abolished the anti-cancer effect of I-BET151 on glioma cells. [29]. The role of HOTAIR as a tumor suppressor gene needs further evaluation [30].
HOTAIR can influence cell-cycle related genes in GBM:
LncRNAs can regulate the cell cycle through several ways Figure 2 [31,32]. For example, ANRIL downregulates p15INK4B expression, and MALAT1 controls B-MYB that controls cell cycle progression [33,34]. Downregulating HOTAIR resulted in cell cycle G0/G1 stage arrest [35]. The downregulation of cyclin D1, cyclin E, cyclin-dependent kinase (CDK)2, CDK4, and the enhanced expression of certain cell cycle-related proteins such as p21 and p16 was associated with HOTAIR suppression in LN229 and U87 cells [25]. HOTAIR regulates about 18 genes that form a cell-cycle related mRNA network [24]. HOTAIR controls glioma cell cycle by regulating FoxM1 and AURKB that are involved in mitosis [24]. Several genes such as ASPM, NCAPG, CDC6, CHEK1, CEP55 play a role in gliomagenesis, through their effect on cell cycle progression [36,37,38,39]. In a certain study, HOTAIR affected the expression of some cell-cycle related genes such as CDC6, NCAPG, CENPE, and PLK4 [24]. As mentioned earlier, HOTAIR can induce gene silencing depending on EZH2 through histone methylation[40]. EZH2 inhibition was reported to stop cell cycle progress at the G0/G1 phase of GBM cells which may favor it as a therapeutic target [40].
Prominent Interactions of HOTAIR with micro-RNA in GBM:
LncRNAs can interact with several mRNAs and affect their activities [41]. LncRNAs can compete with micro RNAs displacing them from binding sites and they are termed competing endogenous RNAs (ceRNAs) [42]. In breast cancer, HOTAIR miR-7 relation is a clear example and in gastric cancer, its pro oncogenic effect was through competing with miR-331-3p [42,43].
HOTAIR/miR-326:
A study has shown that the expression of miR-326 is downregulated in glioma tissues [35]. Knocking down HOTAIR resulted in the upregulation of miR-326 which suppressed the expression of fibroblast growth factor 1 (FGF1) Figure 1 in U87 cells impacting cellular proliferation[35].
HOTAIR/miR-15b:
A study found that HOTAIR reduced miR- 15b expression in glioma cells which may have oncogenic potential [44]. miR-15b could upregulate the tumor suppressor gene p53 expression [44]. HOTAIR, miR-15b, and p53 is a closed loop that controls glioma progression.
HOTAIR/miR-125a:
miR-125a-5p was reported to inhibit glioblastoma cell proliferation, and HOTAIR has been demonstrated to reduce miR-125a expression [45,46]. Schisandrin B, a herbal extract, reduced HOTAIR expression and increased miR-125a-5p expression in glioma cell lines by targeting the mammalian target of rapamycin (mTOR) expression [47].
HOTAIR/miR-219:
miR-219-5p acts as a glioma suppressor by targeting tyrosine kinase and EGFR [48]. HOTAIR has been also shown to inhibit miR-219 in U87 cells, resulting in enhanced cell proliferation and increased cyclin D1 levels [49].
HOTAIR and angiogenesis:
Angiogenesis is controlled by hypoxia mediators; the most well known ones are HIF and VEGF [50,51]. Both HIF and VEGF work together to promote a vascular platform for glioma cells. In nasopharyngeal carcinoma cells, HOTAIR was observed to enhance angiogenesis by activating the transcription promoter of VEGFA [52]. It may act through the formation of extracellular vesicles as it was noticed in the supernatant of GBM culture [53]. Comprehensive studies are needed to evaluate the role of HOTAIR in terms of glioma vascularization.
Potential Use of HOTAIR as a diagnostic marker in GBM:
An absolute need for a non-invasive accurate marker for prognostic implications in patients diagnosed with high-grade gliomas is demanding. The possibility for certain body fluid markers to be used for clinical prediction of glioma is still under investigation. Markers that can monitor response to therapy are essential especially for an aggressive disease like GBM. Differentiating true GBM recurrence from pseudoprogression seems difficult and technically challenging. Conventional MRIs could not easily pick the exact differences between both conditions. A serum biomarker could be a promising tool to aid in the clinical differentiation between both situations.
GFAP, lactate, miR-504, have been reported as potential candidates for diagnosing GBM [54,55,56]. The stability of LncRNAs secondary structures makes them favorable potential biomarkers [57]. HOTAIR also has been identified as a possible serum marker in certain cancers [58,59]. HOTAIR concentration was lower after the surgical treatment of a recurrent GBM and the reduction was more noticeable further weeks after surgery [57]. Further experimental and clinical work should be implemented to evaluate the sensitivity and predictability of HOTAIR as a novel serum biomarker in patients diagnosed with GBM.
HOTAIR as a potential therapeutic target in GBM:
As discussed earlier, HOTAIR can regulate glioma progression in an EZH2-dependent manner through epigenetic role. Therefore, therapeutic targeting of HOTAIR-EZH2 interaction may be utilized as a possible targeting approach. A certain compound AC1Q3QWB that targeted HOTAIR-EZH2, was found to inhibit glioma cell proliferation, with a resultant increase in the tumor suppressor candidate CWF19L1[60,61].
The bromodomain and extraterminal (BET) proteins are epigenetic modulators that have emerged as therapeutic tools for some cancers enriched with epigenetic changes [62]. In a published study, I-BET151 treatment and BRD4 depletion reduced the overexpression of HOTAIR in glioma cells through an effect on transcription and elongation factors [63].
RNAi are possible tools that could be used for inhibiting specific genes, including short interfering RNAs (siRNAs) which are short double-stranded RNAs targeting complementary RNA molecules, resulting in RNA silencing and gene suppression [64]. Carriers of nucleic acids could be used to deliver these siRNAs into the tumor cells. Due to their high stability, iron oxide nanoparticles and specifically SPIONs have been used widely in the delivery [65]. A study has demonstrated the successful delivery of siHOTAIR that subsequently inhibited glioma stem cell proliferation[66]. In a study by Zhang L et al., deleting the HOTAIR regulatory element improved the sensitivity of glioma cells to Temozolomide. [67].
In Temozolomide-resistant GBM cells, HOTAIR was upregulated, while temozolomide resistance was enhanced upon the exosome-mediated transfer of HOTAIR by a mechanism involving miR-519a-3p downregulation [68]. One of the therapeutic challenges is the poor penetration of the blood-brain barrier and achieving a maximal intratumoral concentration. HOTAIR knockdown resulted in improving brain-tumor barrier permeability by a mechanism involving the miR-148b-3p targeting. miR-148b-3p affects the microvascular endothelial cells which control the expression of proteins involved in BBB integrity as ZO-1,claudin-5, and occludin [69].

Table 1.
a sample of experimental studies investigating HOTAIR in glioblastoma.
| Role of HOTAIR | Reference |
|---|---|
|
- HOTAIR inhibits the transcription of NLK in U87 GBM cells, regulate Wnt/β-catenin pathway, inhibit cell cycle arrest and promote cell migration. |
[70] |
| - HOTAIR mRNA levels are increased in A172 glioma cells compared to normal astrocytes. . |
[71] |
| - miR-141 directly binds to the 3UTR of HOTAIR in U251 and U87 glioma cells, inhibiting its expression. |
[72] |
| - miR-148b-3p downregulates the expression of tight junction-related proteins including ZO-1, clauidin-5, and occludin |
[69] |
| - HOTAIR rs920778 and rs12826786 frequencies do not differ between glioma patients and controls |
[73] |
| - HOTAIR levels positively correlate with MMP-7, MMP-9, and VEGF levels in human glioma |
[74] |
| - HOTAIR upregulates the expression of hexokinase 2 by downregulating miR-125 |
[75] |
| - HOTAIR is upregulated in temozolomide-resistant GBM cells - Serum exosome HOTAIR levels are higher in GBM patients’ resistant to temozolomide compared with responders. |
[68] |
Conclusions
There is a compelling need for extensive clinical studies uncovering the HOTAIR role in GBM. Therapies to prolong survival in patients diagnosed with GBM are traditional and their effect on survival is not remarkable. More understanding of the biology of HOTAIR will enable researchers to develop new therapeutic strategies and diagnostic markers that will eventually apply in clinical trials.
Credit Author Statement
Conceptualization, Methodology, Validation, Writing - Original Draft, Writing - Review & Editing and Supervision: Dr Azab..
Conflict of Interest
The authors certify that there is no conflict of interest with any financial organization about the material described in the manuscript.
Funding
The authors' did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
- Bush, N.A.O.; Chang, S.M.; Berger, M.S. Current and future strategies for treatment of glioma. Neurosurg. Rev. 2016, 40, 1–14. [Google Scholar] [CrossRef]
- Sherrod, B.A.; Gamboa, N.T.; Wilkerson, C.; Wilde, H.; Azab, M.A.; Karsy, M.; Jensen, R.L.; Menacho, S.T. Effect of patient age on glioblastoma perioperative treatment costs: a value driven outcome database analysis. J. Neuro-Oncology 2019, 143, 465–473. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA Maps Reveal New RNA Classes and a Possible Function for Pervasive Transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef]
- Wahlestedt, C. Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat. Rev. Drug Discov. 2013, 12, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef]
- Gomes, A.Q.; Nolasco, S.; Soares, H. Non-Coding RNAs: Multi-Tasking Molecules in the Cell. Int. J. Mol. Sci. 2013, 14, 16010–16039. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Nowakowski, T.J.; Pollen, A.A.; Lui, J.H.; Horlbeck, M.A.; Attenello, F.J.; He, D.; Weissman, J.S.; Kriegstein, A.R.; Diaz, A.A.; et al. Single-cell analysis of long non-coding RNAs in the developing human neocortex. Genome Biol. 2016, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dong, X. , et al., Knockdown of long non-coding RNA ANRIL inhibits proliferation, migration, and invasion but promotes apoptosis of human glioma cells by upregulation of miR-34a. J Cell Biochem, 2018. 119(3): p. 2708-2718.
- Shi, Y.; Wang, Y.; Luan, W.; Wang, P.; Tao, T.; Zhang, J.; Qian, J.; Liu, N.; You, Y. Long Non-Coding RNA H19 Promotes Glioma Cell Invasion by Deriving miR-675. PLOS ONE 2014, 9, e86295. [Google Scholar] [CrossRef]
- Shang, C.; Guo, Y.; Hong, Y.; Xue, Y.-X. Long Non-coding RNA TUSC7, a Target of miR-23b, Plays Tumor-Suppressing Roles in Human Gliomas. Front. Cell. Neurosci. 2016, 10, 235. [Google Scholar] [CrossRef]
- Bhan, A.; Mandal, S.S. LncRNA HOTAIR: A master regulator of chromatin dynamics and cancer. Biochim. et Biophys. Acta (BBA) - Rev. Cancer 2015, 1856, 151–164. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Morales, D.R.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Mandal, S.S. Long Noncoding RNAs: Emerging Stars in Gene Regulation, Epigenetics and Human Disease. ChemMedChem 2014, 9, 1932–1956. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Mozdarani, H.; Ezzatizadeh, V.; Parvaneh, R.R. The emerging role of the long non-coding RNA HOTAIR in breast cancer development and treatment. J. Transl. Med. 2020, 18, 1–15. [Google Scholar] [CrossRef]
- Shi, J.; Dong, B.; Cao, J.; Mao, Y.; Guan, W.; Peng, Y.; Wang, S. Long non-coding RNA in glioma: signaling pathways. Oncotarget 2017, 8, 27582–27592. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Qin, Y.; Zhi, Q.; Wang, J.; Qin, C. Knockdown of long non-coding RNA HOTAIR inhibits cisplatin resistance of gastric cancer cells through inhibiting the PI3K/Akt and Wnt/β-catenin signaling pathways by up-regulating miR-34a. Int. J. Biol. Macromol. 2017, 107, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, C.; Yong, W.; Ye, Y.; Huang, Z. Calycosin and Genistein Induce Apoptosis by Inactivation of HOTAIR/p-Akt Signaling Pathway in Human Breast Cancer MCF-7 Cells. Cell. Physiol. Biochem. 2015, 35, 722–728. [Google Scholar] [CrossRef]
- Liang, H.; Huang, W.; Wang, Y.; Ding, L.; Zeng, L. Overexpression of MiR-146a-5p Upregulates lncRNA HOTAIR in Triple-Negative Breast Cancer Cells and Predicts Poor Prognosis. Technol. Cancer Res. Treat. 2019, 18. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Zheng, Y.; You, L.; Kuang, D.; Liu, T. Suppressed expression of long non-coding RNA HOTAIR inhibits proliferation and tumourigenicity of renal carcinoma cells. Tumor Biol. 2014, 35, 11887–11894. [Google Scholar] [CrossRef]
- Xavier-Magalhães, A. , et al., The long non-coding RNA HOTAIR is transcriptionally activated by HOXA9 and is an independent prognostic marker in patients with malignant glioma. Oncotarget, 2018. 9(21): p. 15740-15756.
- Huang, K.; Sun, J.; Yang, C.; Wang, Y.; Zhou, B.; Kang, C.; Han, L.; Wang, Q. HOTAIR upregulates an 18-gene cell cycle-related mRNA network in glioma. Int. J. Oncol. 2017, 50, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X. , et al., HOTAIR, a cell cycle-associated long noncoding RNA and a strong predictor of survival, is preferentially expressed in classical and mesenchymal glioma. Neuro Oncol, 2013. 15(12): p. 1595-603.
- Pojo, M.; Gonçalves, C.S.; Xavier-Magalhães, A.; Oliveira, A.I.; Gonçalves, T.; Correia, S.; Rodrigues, A.J.; Costa, S.; Pinto, L.; Pinto, A.A.; et al. A transcriptomic signature mediated by HOXA9 promotes human glioblastoma initiation, aggressiveness and resistance to temozolomide. Oncotarget 2015, 6, 7657–7674. [Google Scholar] [CrossRef] [PubMed]
- Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature, 2008. 455(7216): p. 1061-8.
- Chen, Y.; Bian, Y.; Zhao, S.; Kong, F.; Li, X. Suppression of PDCD4 mediated by the long non-coding RNA HOTAIR inhibits the proliferation and invasion of glioma cells. Oncol. Lett. 2016, 12, 5170–5176. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Ji, R.; Zhan, W. Long noncoding RNA papillary thyroid carcinoma susceptibility candidate 3 (PTCSC3) inhibits proliferation and invasion of glioma cells by suppressing the Wnt/β-catenin signaling pathway. BMC Neurol. 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Xiao, D.; Cui, X.; Wang, X. LncRNA PTCSC3 inhibits cell proliferation in laryngeal squamous cell carcinoma by down-regulating lncRNA HOTAIR. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Visconti, R.; Della Monica, R.; Grieco, D. Cell cycle checkpoint in cancer: A therapeutically targetable double-edged sword. J. Exp. Clin. Cancer Res. 2016, 35, 153. [Google Scholar] [CrossRef] [PubMed]
- Bucher, N.; Britten, C.D. G2 checkpoint abrogation and checkpoint kinase-1 targeting in the treatment of cancer. Br. J. Cancer 2008, 98, 523–528. [Google Scholar] [CrossRef]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Kitagawa, M.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15INK4B tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar] [CrossRef]
- Tripathi, V.; Shen, Z.; Chakraborty, A.; Giri, S.; Freier, S.M.; Wu, X.; Zhang, Y.; Gorospe, M.; Prasanth, S.G.; Lal, A.; et al. Long Noncoding RNA MALAT1 Controls Cell Cycle Progression by Regulating the Expression of Oncogenic Transcription Factor B-MYB. PLOS Genet. 2013, 9, e1003368. [Google Scholar] [CrossRef]
- Ke, J.; Yao, Y.-L.; Zheng, J.; Wang, P.; Liu, Y.-H.; Ma, J.; Li, Z.; Liu, X.-B.; Li, Z.-Q.; Wang, Z.-H.; et al. Knockdown of long non-coding RNA HOTAIR inhibits malignant biological behaviors of human glioma cells via modulation of miR-326. Oncotarget 2015, 6, 21934–21949. [Google Scholar] [CrossRef]
- Stangeland, B.; Mughal, A.A.; Grieg, Z.; Sandberg, C.J.; Joel, M.; Nygård, S.; Meling, T.; Murrell, W.; Mo, E.O.V.; Langmoen, I.A. Combined expressional analysis, bioinformatics and targeted proteomics identify new potential therapeutic targets in glioblastoma stem cells. Oncotarget 2015, 6, 26192–26215. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.-L.; Hsieh, T.-H.; Ng, K.-H.; Tsai, Y.-N.; Tsai, C.-F.; Chao, M.-E.; Liu, D.-J.; Chu, S.-S.; Chen, W.; Liu, Y.-R.; et al. Downregulation of miR-137 and miR-6500-3p promotes cell proliferation in pediatric high-grade gliomas. Oncotarget 2016, 7, 19723–19737. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Dai, Y.; Grant, S.; Dent, P. Enhancing CHK1 inhibitor lethality in glioblastoma. Cancer Biol. Ther. 2012, 13, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Varambally, S.; Dhanasekaran, S.M.; Zhou, M.; Barrette, T.R.; Kumar-Sinha, C.; Sanda, M.G.; Ghosh, D.; Pienta, K.J.; Sewalt, R.G.A.B.; Otte, A.P.; et al. The polycomb group protein EZH2 is involved in progression of prostate cancer. Nature 2002, 419, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R. , et al., Downregulation of Ezh2 expression by RNA interference induces cell cycle arrest in the G0/G1 phase and apoptosis in U87 human glioma cells. Oncol Rep, 2012. 28(6): p. 2278-84.
- Jalali, S. , et al., Systematic transcriptome wide analysis of lncRNA-miRNA interactions. PLoS One, 2013. 8(2): p. e53823.
- Liu, X.-H.; Sun, M.; Nie, F.-Q.; Ge, Y.-B.; Zhang, E.-B.; Yin, D.-D.; Kong, R.; Xia, R.; Lu, K.-H.; Li, J.-H.; et al. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331-3p in gastric cancer. Mol. Cancer 2014, 13, 92–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, K.; Wang, J.; Wang, X.; Cheng, K.; Shi, F.; Jiang, L.; Zhang, Y.; Dou, J. MiR-7, Inhibited Indirectly by LincRNA HOTAIR, Directly Inhibits SETDB1 and Reverses the EMT of Breast Cancer Stem Cells by Downregulating the STAT3 Pathway. STEM CELLS 2014, 32, 2858–2868. [Google Scholar] [CrossRef]
- Sun, G.; Wang, Y.; Zhang, J.; Lin, N.; You, Y. MiR-15b/HOTAIR/p53 form a regulatory loop that affects the growth of glioma cells. J. Cell. Biochem. 2018, 119, 4540–4547. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Xiao, G.; Peng, G.; Liu, D.; Wang, Z.; Liao, Y.; Liu, Q.; Wu, M.; Yuan, X. MiRNA-125a-5p inhibits glioblastoma cell proliferation and promotes cell differentiation by targeting TAZ. Biochem. Biophys. Res. Commun. 2015, 457, 171–176. [Google Scholar] [CrossRef]
- Tang, L.; Shen, H.; Li, X.; Li, Z.; Liu, Z.; Xu, J.; Ma, S.; Zhao, X.; Bai, X.; Li, M.; et al. MiR-125a-5p decreases after long non-coding RNA HOTAIR knockdown to promote cancer cell apoptosis by releasing caspase 2. Cell Death Dis. 2016, 7, e2137–e2137. [Google Scholar] [CrossRef]
- Jiang, Y. , et al., Schisandrin B inhibits the proliferation and invasion of glioma cells by regulating the HOTAIR-micoRNA-125a-mTOR pathway. Neuroreport, 2017. 28(2): p. 93-100.
- Rao, S.A.M.; Arimappamagan, A.; Pandey, P.; Santosh, V.; Hegde, A.S.; Chandramouli, B.A.; Somasundaram, K. miR-219-5p Inhibits Receptor Tyrosine Kinase Pathway by Targeting EGFR in Glioblastoma. PLOS ONE 2013, 8, e63164. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guan, C. HOTAIR inhibits the proliferation of glioblastoma cells by targeting miR-219. Cancer Biomarkers 2020, 28, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Plate, K.H.; Breier, G.; Weich, H.A.; Mennel, H.D.; Risau, W. Vascular endothelial growth factor and glioma angiogenesis: Coordinate induction of VEGF receptors, distribution of VEGF protein and possible In vivo regulatory mechanisms. Int. J. Cancer 1994, 59, 520–529. [Google Scholar] [CrossRef]
- A Azab, M.; Alomari, A.; Azzam, A.Y. Featuring how calcium channels and calmodulin affect glioblastoma behavior. A review article. Cancer Treat. Res. Commun. 2020, 25, 100255. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.-M.; Lu, Y.-F.; Hu, B.-G.; Liang, W.-C.; Zhu, X.; Yang, H.-D.; Li, G.; Zhang, J.-F. Long noncoding RNA hotair mediated angiogenesis in nasopharyngeal carcinoma by direct and indirect signaling pathways. Oncotarget 2015, 7, 4712–4723. [Google Scholar] [CrossRef]
- Subramaniam, S.R.; Federoff, H.J. Targeting Microglial Activation States as a Therapeutic Avenue in Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 176–176. [Google Scholar] [CrossRef] [PubMed]
- Cata, J.; Bhavsar, S.; Hagan, K.; Arunkumar, R.; Grasu, R.; Dang, A.; Carlson, R.; Arnold, B.; Popat, K.; Rao, G.; et al. Intraoperative serum lactate is not a predictor of survival after glioblastoma surgery. J. Clin. Neurosci. 2017, 43, 224–228. [Google Scholar] [CrossRef]
- Jin, Z.; Jin, R.-H.; Ma, C.; Li, H.-S.; Xu, H.-Y. Serum expression level of miR-504 can differentiate between glioblastoma multiforme and solitary brain metastasis of non-small cell lung carcinoma. . 2017, 22, 474–480. [Google Scholar]
- Vietheer, J.-M.; Rieger, J.; Wagner, M.; Senft, C.; Tichy, J.; Foerch, C. Serum concentrations of glial fibrillary acidic protein (GFAP) do not indicate tumor recurrence in patients with glioblastoma. J. Neuro-Oncology 2017, 135, 193–199. [Google Scholar] [CrossRef]
- Tan, S.K.; Pastori, C.; Penas, C.; Komotar, R.J.; Ivan, M.E.; Wahlestedt, C.; Ayad, N.G. Serum long noncoding RNA HOTAIR as a novel diagnostic and prognostic biomarker in glioblastoma multiforme. Mol. Cancer 2018, 17, 1–7. [Google Scholar] [CrossRef]
- Cantile, M.; Scognamiglio, G.; Marra, L.; Aquino, G.; Botti, C.; Falcone, M.R.; Malzone, M.G.; Liguori, G.; Di Bonito, M.; Franco, R.; et al. HOTAIR role in melanoma progression and its identification in the blood of patients with advanced disease. J. Cell. Physiol. 2017, 232, 3422–3432. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; He, X.; Zheng, Z.; Ma, X.; Hu, X.; Wu, D.; Wang, M. Serum HOTAIR as a novel diagnostic biomarker for esophageal squamous cell carcinoma. Mol. Cancer 2017, 16, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ren, Y.; Wang, Y.; Tan, Y.; Wang, Q.; Cai, J.; Zhou, J.; Yang, C.; Zhao, K.; Yi, K.; et al. A Compound AC1Q3QWB Selectively Disrupts HOTAIR-Mediated Recruitment of PRC2 and Enhances Cancer Therapy of DZNep. Theranostics 2019, 9, 4608–4623. [Google Scholar] [CrossRef] [PubMed]
- Shi, J. , et al., HOTAIR-EZH2 inhibitor AC1Q3QWB upregulates CWF19L1 and enhances cell cycle inhibition of CDK4/6 inhibitor palbociclib in glioma. Clin Transl Med, 2020. 10(1): p. 182-198.
- Filippakopoulos, P.; Knapp, S. Targeting bromodomains: epigenetic readers of lysine acetylation. Nat. Rev. Drug Discov. 2014, 13, 337–356. [Google Scholar] [CrossRef] [PubMed]
- Pastori, C.; Kapranov, P.; Penas, C.; Peschansky, V.; Volmar, C.-H.; Sarkaria, J.N.; Bregy, A.; Komotar, R.; St Laurent, G.; Ayad, N.G.; et al. The Bromodomain protein BRD4 controls HOTAIR, a long noncoding RNA essential for glioblastoma proliferation. Proc. Natl. Acad. Sci. USA 2015, 112, 8326–8331. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Chen, Y. Targeting long non-coding RNAs in cancers: Progress and prospects. Int. J. Biochem. Cell Biol. 2013, 45, 1895–1910. [Google Scholar] [CrossRef] [PubMed]
- Bruniaux, J.; Allard-Vannier, E.; Aubrey, N.; Lakhrif, Z.; Ben Djemaa, S.; Eljack, S.; Marchais, H.; Hervé-Aubert, K.; Chourpa, I.; David, S. Magnetic nanocarriers for the specific delivery of siRNA: Contribution of breast cancer cells active targeting for down-regulation efficiency. Int. J. Pharm. 2019, 569, 118572. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Liu, P.; Dong, S.; Guo, Y.; Cui, X.; Zhu, X.; Li, X.; Jiang, L.; Liu, T.; Wu, Y. Magnetofection based on superparamagnetic iron oxide nanoparticle-mediated low lncRNA HOTAIR expression decreases the proliferation and invasion of glioma stem cells. Int. J. Oncol. 2016, 49, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, A.; Chen, B.; Bi, J.; Chen, J.; Guo, D.; Qian, Y.; Wang, W.; Shi, T.; Zhao, Z.; et al. A HOTAIR regulatory element modulates glioma cell sensitivity to temozolomide through long-range regulation of multiple target genes. Genome Res. 2020, 30, 155–163. [Google Scholar] [CrossRef]
- Yuan, Z. , et al., Exosome-Mediated Transfer of Long Noncoding RNA HOTAIR Regulates Temozolomide Resistance by miR-519a-3p/RRM1 Axis in Glioblastoma. Cancer Biother Radiopharm, 2020.
- Sa, L.; Li, Y.; Zhao, L.; Liu, Y.; Wang, P.; Liu, L.; Li, Z.; Ma, J.; Cai, H.; Xue, Y. RETRACTED: The Role of HOTAIR/miR-148b-3p/USF1 on Regulating the Permeability of BTB. Front. Mol. Neurosci. 2017, 10, 194. [Google Scholar] [CrossRef]
- Zhou, X.; Ren, Y.; Zhang, J.; Zhang, C.; Zhang, K.; Han, L.; Kong, L.; Wei, J.; Chen, L.; Yang, J.; et al. HOTAIR is a therapeutic target in glioblastoma. Oncotarget 2015, 6, 8353–8365. [Google Scholar] [CrossRef]
- Wang, G.; Li, Z.; Tian, N.; Han, L.; Fu, Y.; Guo, Z.; Tian, Y. miR-148b-3p inhibits malignant biological behaviors of human glioma cells induced by high HOTAIR expression. Oncol. Lett. 2016, 12, 879–886. [Google Scholar] [CrossRef]
- Bian, E.-B.; Ma, C.-C.; He, X.-J.; Wang, C.; Zong, G.; Wang, H.-L.; Zhao, B. Epigenetic modification of miR-141 regulates SKA2 by an endogenous ‘sponge’ HOTAIR in glioma. Oncotarget 2016, 7, 30610–30625. [Google Scholar] [CrossRef]
- Xavier-Magalhães, A.; Oliveira, A.I.; de Castro, J.V.; Pojo, M.; Gonçalves, C.S.; Lourenço, T.; Viana-Pereira, M.; Costa, S.; Linhares, P.; Vaz, R.; et al. Effects of the functional HOTAIR rs920778 and rs12826786 genetic variants in glioma susceptibility and patient prognosis. J. Neuro-Oncology 2017, 132, 27–34. [Google Scholar] [CrossRef]
- Zhao, W.-H.; Yuan, H.-Y.; Ren, X.-Y.; Huang, K.; Guo, Z.-Y. Association between expression of HOTAIR and invasiveness of gliomas, and its predictive value. Adv. Clin. Exp. Med. 2019, 28, 1179–1183. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Gao, Y.; Liang, H. HOTAIR/miR-125 axis-mediated Hexokinase 2 expression promotes chemoresistance in human glioblastoma. J. Cell. Mol. Med. 2020, 24, 5707–5717. [Google Scholar] [CrossRef]
- Hanisch, A.; Silljé, H.H.W.; A Nigg, E. Timely anaphase onset requires a novel spindle and kinetochore complex comprising Ska1 and Ska2. EMBO J. 2006, 25, 5504–5515. [Google Scholar] [CrossRef]
- Wang, G.; Li, Z.; Tian, N.; Han, L.; Fu, Y.; Guo, Z.; Tian, Y. miR-148b-3p inhibits malignant biological behaviors of human glioma cells induced by high HOTAIR expression. Oncol. Lett. 2016, 12, 879–886. [Google Scholar] [CrossRef]
- Chen, L.; Han, L.; Wei, J.; Zhang, K.; Shi, Z.; Duan, R.; Li, S.; Zhou, X.; Pu, P.; Zhang, J.; et al. SNORD76, a box C/D snoRNA, acts as a tumor suppressor in glioblastoma. Sci. Rep. 2015, 5, 8588. [Google Scholar] [CrossRef]
- Chen, L.; Han, L.; Wei, J.; Zhang, K.; Shi, Z.; Duan, R.; Li, S.; Zhou, X.; Pu, P.; Zhang, J.; et al. SNORD76, a box C/D snoRNA, acts as a tumor suppressor in glioblastoma. Sci. Rep. 2015, 5, 8588. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed]
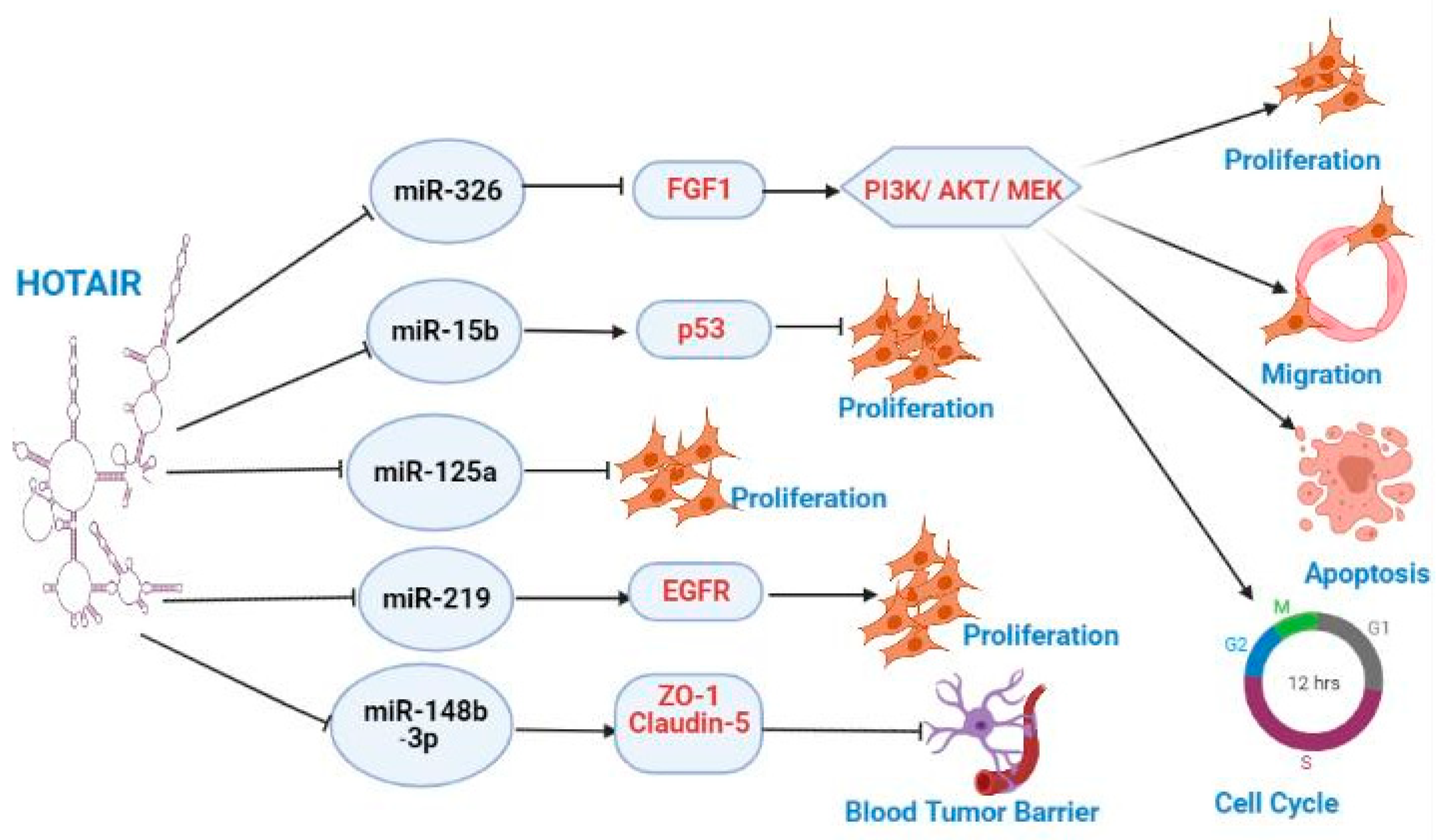
Figure 1.
HOTAIR can interact with several other non-coding mRNAs and affect their activities which eventually affect cellular proliferation, apoptosis and cell cycle. MiR-326 inhibition resulted in decreased FGF1 that mediates the activity of the pathway PI3K/AKT/MEK which affects proliferation, migration and apoptosis. MiR-15 inhibition by HOTAIR will affect the level of p53 which is a tumor suppressor gene. Inhibition of miR-148b-3p by HOTAIR results in decreased expression of tight junction proteins which affects the tumor-brain barrier permeability.
Figure 1.
HOTAIR can interact with several other non-coding mRNAs and affect their activities which eventually affect cellular proliferation, apoptosis and cell cycle. MiR-326 inhibition resulted in decreased FGF1 that mediates the activity of the pathway PI3K/AKT/MEK which affects proliferation, migration and apoptosis. MiR-15 inhibition by HOTAIR will affect the level of p53 which is a tumor suppressor gene. Inhibition of miR-148b-3p by HOTAIR results in decreased expression of tight junction proteins which affects the tumor-brain barrier permeability.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



