
Submitted:
17 July 2024
Posted:
18 July 2024
You are already at the latest version
Abstract
Current fertilizer recommendations often neglect nutrient cycling across crop rotations. This study aimed to assess the decay rate and nutrient (N, P, K) release patterns of sorghum, black oat, and corn residues and their effects on corn yield in an integrated crop-livestock system. The experiment used factorial treatments based on two sward heights (high and low) and two nitrogen fertilization levels (N-Pasture at 200 kg N ha-1 and N-Corn at 0 kg N ha-1. Litter bags were collected at various intervals from each crop to measure nutrient release patterns and decomposition rates. The results showed that pasture height and nitrogen fertilization significantly influenced decomposition and nutrient release, affecting the subsequent grain crop phase. Potassium was released rapidly and in high amounts. Nitrogen fertilization during the pasture phase prevented nitrogen and phosphorus immobilization in black oat residue and reduced immobilization in corn residue.These findings highlight the importance of accounting for nutrient cycling and decomposition rates in fertilization strategies to enhance the sustainability of integrated crop-livestock systems.
Keywords:
System Fertilization
; Nutrient cycling
; Dry matter decomposition rates
1. Introduction
Nitrogen (N) fertilization is conventionally applied to grain crops, such as corn, within the crop cultivation systems of tropical and subtropical regions in Brazil. Predominantly used as a cover crop, grasses like black oat are subject to only low rates of N fertilization, even within integrated crop-livestock systems (ICLS). In such conditions, the litter produced exhibits a high carbon to nitrogen (C/N) ratio and significant lignin concentration, which can lead to decreased decomposition rates and nutrient immobilization, particularly of nitrogen and phosphorus [1].
Given the intricate dynamics of ICLS and the challenges of accurately interpreting soil fertility, existing fertilizer and liming recommendation models do not account for the nutrient cycling between different crop rotation phases. In Brazil, the perennial growth conditions allow for continuous nutrient cycling through soil, plant, and animal residues, which are crucial sources of nutrients, notably N, P, and K. Occasionally, the potassium released from residues surpasses the amounts typically applied through fertilization practices. System-level fertilization, a concept based on biological nutrient cycling across rotation phases, aims to enhance nutrient-use efficiency, reduce the dependence on mineral nutrient inputs, minimize losses, and sustain long-term soil fertility. This concept contrasts sharply with traditional fertilization paradigms focused on individual cash crops within a rotation, where the residual effects of fertilizers are often overlooked [2].
The synchronization of organically-bound nutrient release with crop nutrient demand is crucial for the sustainability of agrosystems. This synchronism, or the timing of nutrient release, is essential for maximizing the efficiency of nutritional resources. Elevated nutrient availability, especially of nitrogen resulting from previous fertilization, can significantly enhance the nutritional status of subsequent crops. For instance, in areas where pasture received nitrogen fertilization, soil N-mineral content tends to remain above critical levels even at the onset of grain crop cultivation, providing an optimal growth environment without nitrogen limitation. Conversely, corn plants grown in areas that did not receive nitrogen during the pasture phase may encounter a less favorable nutritional environment, potentially compromising yield components that are established early in the crop’s development. The critical role of nutrient timing and its impact on crop yield stability over a long-term period was highlighted in a 50-year study, which found that complete nutrient supply, especially the inclusion of nitrogen, was essential for maintaining yield stability across major European crops [3]. Furthermore, a 10-year term experiment on nutrient management in the rice-wheat system revealed that strategic nutrient applications could stabilize and enhance annual yields by aligning nutrient availability with critical growth phases of the crops [4].
Furthermore, research such as that by [5] emphasizes how sheep grazing and crop rotation impact soil acidification and the budget of key nutrients like P, K, Ca, and Mg in Southern Brazil, highlighting the importance of grazing management in ICLS for maintaining soil health and fertility. Similarly, provide insights into the nutrient dynamics in cattle grazing systems in the Amazon, where managing nutrient flows is vital to avoid soil degradation [6].
In Brazil, detailed studies on residue decomposition and nutrient release from pastures and catch crops are limited. This research aims to elucidate the decomposition rates and nutrient release patterns of sorghum, black oat, and corn residues, and their impact on corn yields within ICLS, as influenced by different stocking rates and nitrogen fertilizer applications. Studies like those by Vogel et al. (2021) and Camelo (2018) on nutrient flows in various Brazilian agroecosystems further support the need for this kind of integrative research [7,8].
2. Materials and Methods
The research was carried out in Abelardo Luz, state of Santa Catarina, southern Brazil (26° 31’ S, 51° 35’ W, at an elevation of 850 m). The study took place on a 20-hectare field which has been managed under a no-tillage Integrated Crop Livestock System since the year 2012. The sequence of experiments began with grazing of sorghum in the summer of 2012/13, followed by black oat grazing in 2013, and subsequent planting of corn for grain production during the summer of 2013/14. The soil at the experimental site is classified as an Oxisol with a clayey texture. The local climate falls under the Cfb category—humid subtropical, according to Köppen’s Climate Classification. The initial chemical properties of the soil at a depth of 0 - 0.2 m were determined: pH measured with CaCl was 4.6, organic matter (OM) content was 36.3 g dm, phosphorus (P) content was 4.6 mg dm, potassium (K) was 88 mg dm, calcium (Ca) was 3.9 cmol(c) dm, magnesium (Mg) was 2.0 cmol(c) dm, base saturation was 56%, and cation exchange capacity (CEC) was 15.9 cmol(c) dm.
The experimental pasture phase was initiated on November 7, 2012, with the no-till sowing of sorghum hybrid BMR - ADV 2800. The drill spacing was set at 45 cm with a final stand achieving 450,000 plants per hectare. Fertilization at sowing included 42 kg N ha, 106 kg PO ha, and 106 kg KO ha, following the soil analysis and the guidelines provided by the Brazilian Commission for Chemical and Soil Fertility [9]. The sorghum pasture was desiccated on April 22, 2013, and subsequently, on April 25, 2013, black oat (Avena strigosa) was planted using a no-till method. The seeding rate was 100 kg ha with a row spacing of 17 cm. Black oat was also fertilized with 75 kg PO ha and 75 kg KO ha at sowing.
The experimental design was structured as a randomized complete block with three replications, adopting a 2 x 2 bi-factorial arrangement in a split-plot model. The design incorporated combinations of two sward heights and two N-fertilization timings. Grazing management of the sorghum and black oat pastures employed continuous stocking, utilizing a variable number of weaned Nelore/Charolais steers. Each experimental unit included three "tester" steers designated to monitor weight gain in the pasture, and additional "put and take" steers were used to maintain the desired sward height. The black oat pasture was grazed until September 6, 2013.
In the first experimental pasture phase (Phase I), treatments consisted of a factorial arrangement of two sward heights x two N-fertilization timings. The targeted sward heights were 30 cm (low sward height) and 60 cm (high sward height) for sorghum, and 15 cm (low sward height) and 30 cm (high sward height) for black oat. Sward heights were measured weekly using a grazing stick marked in centimeters. After the removal of cattle at the end of the pasture phase, plant residues were treated with 1.5 L ha of glyphosate. Corn, hybrid ’Maximus,’ was then sown on October 10, 2013, in a no-till system with a row spacing of 0.8 m. The entire experimental area was fertilized with 32 kg N ha, 80 kg PO ha.
In the second experimental phase, referred to as the corn phase, following the removal of cattle at the conclusion of the pasture phase, plant residues were treated with a desiccation application of 1.5 L ha of glyphosate. Subsequently, corn hybrid ’Maximus’ was planted on October 10, 2013, utilizing a no-tillage system and maintaining a row spacing of 0.8 m. The entire experimental area received an application of formulated N-P-K fertilizer, comprising 32 kg N ha, 80 kg PO ha, and 80 kg KO ha.
The experimental variable, N-Fertilization Time, is designed to represent system-level fertilization by applying nitrogen fertilizer exclusively during one phase of the Integrated Crop Livestock System. This variable was operationalized through two treatments: N-Pasture fertilization and N-Corn fertilization. The plots receiving N-Pasture fertilization were applied with 200 kg N ha each on the sorghum pasture and the black oat crop. Conversely, the plots designated as N-Corn fertilization did not receive nitrogen during these pasture phases. However, during the corn phase, these plots were supplied with 200 kg N ha, whereas the N-Pasture fertilization plots did not receive any nitrogen application during this phase.
The overall experimental design was a randomized complete block with three replications, and was divided into two crop phases: the pasture phase and the corn phase.
2.1. Dry Matter Decomposition and Nutrient Release Experiment
Dry matter decomposition and N, P, and K release from sorghum, black oat, and corn residues were evaluated using litter bags placed on the soil surface corresponding to different combinations of N-Fertilization Time rates and Sward height in subsequent crop areas. The experimental design was a randomized complete block with three replications in a tri-factorial arrangement. Treatments were structured in a split-plot model, with main plots assigned to combinations of 2 Sward heights x 2 N-Fertilization Times and sub-plots designated for Days of Deposition.
Residues from sorghum, black oat, and corn were collected at the end of each respective cropping phase. Stubble Dry Matter (DM) samples weighing 10 g were enclosed in 20 x 20 cm nylon-screen litter bags with 2 mm mesh openings. These litter bags were sealed and positioned on the soil surface to mimic no-tillage conditions, placed seven days post-sowing of each crop.
For sorghum, six litter bags per experimental unit were retrieved at incubation days 15, 35, 55, 75, 100, and 135. For black oat, eight litter bags were retrieved at 15, 27, 42, 62, 78, 96, 120, and 150 days; for corn, seven bags were collected at 12, 28, 49, 84, 110, 136, and 182 days. All litter bag contents were subsequently dried in a forced-air oven at 55°C.
Decomposition rates and nutrient release were quantified from weight changes and nutrient concentrations over the incubation periods. Litter samples were ground to pass a 0.841 mm sieve and then subjected to sulfuric acid digestion. The resultant digest was analyzed for total N using the Kjeldahl method, for P via photo-colorimetry, and for K by flame photometry [10].
Data on DM, N, P, and K remaining at each retrieval interval were modeled non-linearly for each replication to generate decomposition characteristics, which were analyzed using analysis of variance. To depict the rates of decomposition and nutrient release, percentages of remaining dry biomass and nutrient contents were adjusted to nonlinear decay models using Statgraphics Plus 4.1:
where RDM represents the remaining dry matter, RN the remaining nutrients after time t (days), and is the decay constant for the rapidly decomposable compartment (A). The model selected was the one with the highest coefficient of determination (), indicating a strong correlation between fitted values and observed data.
The decomposition model assumes that litter DM and nutrients are segmented into two fractions. In the model specified in Equation (1), the fraction labeled (A) decomposes exponentially at a consistent rate over time. The second fraction (100 - A) is considered more resistant and remains largely unchanged within the observed period.
Half-life (), or the time required for 50% of a compartment to decompose, was calculated using the decay constants derived for each compartment based on an equation proposed by [11]:
Cumulative N-P-K release was estimated by the difference between the initial and remaining N-P-K quantities in the residues, calculated by multiplying the percent N-P-K concentration by the DM remaining, as obtained from the decay model.
2.2. Statistical Procedures
All experimental results were subjected to an analysis of variance (ANOVA), with data transformation applied where necessary. For quantitative factors, non linear models were fitted. Model selection was based on statistical significance (less than 5%) and the coefficient of determination ().
In cases of significant interactions, varying nitrogen rates were individually evaluated under each unique scenario. The interactions between situations were analyzed within each rate of N-Fertilization Time or Sward height. Where interactions were non-significant, individual factors were analyzed independently.
All statistical analyses were conducted using the GENES statistical software [12] and SigmaPlot® version 12.5 (Systat Software, San Jose, CA).
3. Results
3.1. Initial Characterization of Residue Dry Matter
Among the crop residues studied, corn residue, which was not grazed, exhibited the highest value of dry matter remaining on the soil post-harvest, averaging 6773 kg DM ha (Table 1). In contrast, black oat residue had the lowest value, averaging 1385 kg DM ha.
Black oat residue had the highest concentrations of nitrogen (N), phosphorus (P), and potassium (K) due to its classification as a C3 plant (Table 1). The initial nitrogen concentration in black oat residue was significantly influenced by the interaction between N-Fertilization Time and Sward Height (P = 0.0066). The highest nitrogen concentration was observed in the N-Pasture/Low Height (NPLH) plots, with a value of 38.52 g kg.
Neutral Detergent Fiber (NDF) and Acid Detergent Fiber (ADF) values varied among residues. Corn residue had the highest values for both NDF and ADF, particularly in the N-Corn fertilization/High Sward Height plots, which exhibited NDF at 76.57% and ADF at 47.40%. This indicates higher levels of structural carbohydrates compared to sorghum and black oat residues, which showed lower NDF and ADF percentages overall.
The lowest ADF values were seen in sorghum residue from the N-Pasture fertilization plots, which had ADF percentages as low as 28.16%. Black oat residue exhibited intermediate NDF and ADF values, varying by sward height and fertilization timing. This variation in NDF and ADF values underscores the differing fiber composition across crop residues, with implications for decomposition rates and nutrient cycling.
3.2. Dry Matter Decomposition of Sorghum, Black Oat and Corn
The dry matter active fraction (Ac.Fra) that decomposed ranged from 36.8% to 80.2% (Table 2). Consequently, the resistant fraction varied from 14.2% to 18.8%. The highest active fraction values were observed in black oat residue grazed at high sward height and in the N-Corn plots. Conversely, the smallest active fraction value was seen in corn residue plots that were grazed at high sward height and received N-Corn fertilization. The time required to decompose 50% of the initial residue (half-life) varied between 15 days (Black oat - low sward height, N-Pasture) and 76 days (Corn - high sward height, N-Corn).
Dry matter loss was well-described by the single exponential decay model for all three plant residues (Figure 1). The model had an average fit of 90.5% ± 7.8%.
The average residual dry matter of sorghum, after grazing, was 4909 kg ha. The decomposition of sorghum residue was not affected by N-Fertilization Time or sward height.
In black oat, the active fraction ranged from 71% (NC-LH) to 80% (NC-HH). The highest decay constant of the active fraction was observed in the NPLH plot (0.04483), meaning that 15 days were required to decompose 50% of the initial residue in the active fraction. In NCHH, the observed half-life was 31 days.
After 42 days of incubation, the greatest residue amount remaining was found in the NCHH and NPHH plots (715 kg DM ha), while the smallest was observed in the NPLH plot (431 kg DM ha) due to a combination of low initial dry matter quantity and rapid decomposition.
The residual dry matter after the corn harvest was 6772.9 kg ha. Corn residue in plots that received the highest stocking rate before planting (LH) decomposed faster. The highest decay constant of the active fraction was observed with low sward height and N-fertilizer applied to the pasture phase (NPLH = 0.02132), requiring 33 days to decompose 50% of the initial residue. The active fraction was also higher in plots that received the highest stocking rate (LH) (45%).
3.3. Nitrogen Release of Sorghum, Black Oat and Corn Residue
All nutrient releases followed a similar exponential trend as observed for dry matter decomposition. Potassium was released the fastest and to the greatest extent. Phosphorus exhibited rapid release in the initial stages, followed by a slower release phase and occasional immobilization episodes.
Nitrogen release in sorghum and especially black oat residue tended to be quicker when 200 kg N ha was applied as N-Pasture fertilization (Figure 2). The nitrogen release from corn residue was not significantly affected by N-fertilization timing or stocking grazing.
The release of nitrogen from sorghum residue was faster in plots that received N-Pasture fertilization. On average, across sward heights, 81% of the nitrogen remained after 35 days of incubation. In these cases, approximately 53 kg N ha was released from sorghum residue. This period coincided with the black oat tillering phase, a critical period of nitrogen demand. Due to the higher initial residue quantity in NPHH plots (5204.7 kg DM ha), these plots exhibited the greatest nitrogen release at 35 days of incubation (57 kg N ha). In N-Corn plots grazed at high sward height (NCHH), 80% of nitrogen remained after 35 days of incubation.
At 75 days post-deposition, nitrogen immobilization was observed in sorghum residue plots that had not received nitrogen fertilization, regardless of pasture height. An increase in nitrogen remaining was seen in NCHH (106%) and NCLH (102%). Immobilization continued up to 100 days post-deposition but was limited to NCLH plots. After this point, nitrogen release resumed from the residue.
The highest nitrogen active fraction was observed in black oat residue (75% on average). For sorghum and corn residues, the active fractions were 32% and 37%, respectively (Table 2).
In the NPLH plots, 42 days after deposition, 35 kg N ha was released from black oat residue, while only 25 kg N ha was released in NGHH plots (Figure 2). At this point, the corn crop was in the V5 vegetative growth stage and required significant nitrogen.
Nitrogen release from corn residue was not significantly affected by the treatments, and on average, the nitrogen active fraction was 38%, with a half-life of 7 days. However, due to the effect of sward height on dry matter decomposition, plots previously grazed at low height released more nitrogen. At 28 days post-deposition, on average, 73 kg N ha was released in LH plots compared to 47 kg N ha in HH plots.
No nitrogen immobilization episodes were observed in corn residue, even in N-Pasture plots that had not received N-fertilization during the corn phase. This suggests that prior N-Pasture fertilization ensured nitrogen supply for both crops and soil microorganisms.
3.4. Phosphorous Release of Sorghum, Black Oat, and Corn Residue
Phosphorus release during the study is shown in Figure 3. Generally, phosphorus release in the initial stages was rapid, followed by a slower phase. For sorghum residue, 15 days after deposition, an average of 35% of phosphorus had been released. At this time, the NPHH plots released 30.4 kg P ha, while the lowest phosphorus release was observed in the NCLH plots (9.5 kg P ha). Nitrogen fertilization of sorghum resulted in substantial early phosphorus release. After this period, phosphorus content in the residue remained the same, indicating no further release, possibly due to immobilization by microorganisms during decomposition.
Phosphorus release from black oat residue followed a similar pattern to that of sorghum. However, initial phosphorus amounts released were smaller than those for sorghum due to the smaller amount of residual black oat dry matter (1385 kg DM ha). Fifteen days after deposition, an average of 34% of phosphorus had been released. At this time, NPHH plots had released 11.6 kg P ha, while other treatments released an average of 4.2 kg P ha.
Afterward, a more intense immobilization process was observed in black oat residue compared to sorghum. By 42 days after deposition, phosphorus concentrations in black oat residue had increased.
Phosphorus release from corn residue also showed a rapid initial release followed by an immobilization period. At 49 days post-deposition, an average of 22% of phosphorus had been released. At this time, the NCLH plots released 9.0 kg P ha, while the lowest phosphorus release was observed in the NPHH plots (3.4 kg P ha). Nitrogen fertilization in corn (N-Corn) resulted in substantial early phosphorus release. Following this period, a strong immobilization process began. At 136 days after deposition, an average of 3.5 kg P ha was immobilized in corn residue.
3.5. Potassium Release of Sorghum, Black Oat, and Corn Residue
Potassium was released the fastest and in the greatest amount compared to nitrogen and phosphorus. There were fewer differences among treatments in the active fraction and the non-linear decay constant for potassium across all crop residue types (Figure 4). The average potassium active fraction and half-life for all residues were 83% and 13 days, respectively.
At 35 days after deposition of sorghum residue, 82.4% of potassium had been released, corresponding to a release of 270 kg K ha. At 42 days, black oat residues released 95 kg K ha, while NCHH plots released only 33 kg K ha.
The consistent pattern of potassium release across all residues is shown in Figure 5. The results of fitting a non-linear regression model describe the relationship between remaining potassium (%) and the independent variable "Days after deposition." The fitted model equation is:
K-remaining = 90.2347 * exp + 9.77
The active fraction of potassium has a half-life of 11 days. The R-squared statistic indicates that the model explains 83.8% of the variability in potassium release.
As potassium plays a minor role in organic compounds, its cycle tends to be shorter and faster. To accelerate potassium release from residues, the following crop should be planted immediately after cover crop management to minimize leaching through the soil profile.
4. Discussion
4.1. Dry Matter Decomposition of Sorghum, Black Oat, and Corn Residue
The average days required to decompose 50% of the initial active fraction of sorghum residue (66%) was 33. [13] evaluated the decomposition of different pasture residues and reported a 75-day litter half-life. Similarly, [14] found a 118-day half-life for sorghum residue used as a cover crop. In our study, after 135 days, the remaining sorghum dry matter averaged 1913 kg DM ha (Figure 1).
[1] reported that the time required to decompose 50% of dual-purpose wheat residue ranged from 52 to 99 days, significantly declining with increased grazing periods.
The shorter half-life of the active fraction in black oat is likely due to the higher regrowth in pastures under a higher stocking rate (LH) and the application of N-fertilizer in the pasture phase (NP). Greater stocking rates also affected the cellulose and lignin content (% ADF). The shorter active fraction half-life may have resulted from the increased presence of younger plant material due to more intensive grazing.
A larger proportion of younger plant material was present in the lower height (LH) plots because of the increased presence of black oat tillers and regrowth. [15] found that grazed Lolium had a lower fiber content and lignin:N ratio than ungrazed forage. In comparisons between fresh and senesced plant residues, decomposition was faster in fresh residues due to lower lignin concentration, higher nitrogen content, and higher soluble sugar levels [16].
N fertilization in black oat pasture (NP plots) was particularly evident in plots with a higher stocking rate (LH). [17] also observed that nitrogen application in grazed pastures resulted in a higher rate of straw decomposition.
In corn plants that were not grazed, the stocking rate influenced the decomposition of corn residue. Residues in plots with a higher previous stocking rate (LH) decomposed faster. Additionally, plots with a higher stocking rate had a greater active fraction.
Prior grazing history can either accelerate or slow down nutrient release from litter by altering the soil environment for decomposition [18,19,20]. Herbivory can also impact organic matter decomposition and nutrient cycling rates by altering the quality of litter entering the soil via above- and belowground pathways [21].
[22] found that plants from grazed sites produced litter with lower fiber content (higher cell solubles) and lower lignin:N ratios than plants from ungrazed sites, contributing to accelerated decomposition [23,24]. As shown in Table 1, corn residue fiber content (FDA) was lower in plots that had received N fertilization during the pasture phase (P = 0.0262).
In our study, prior grazing and N fertilization may have changed the soil microbial community due to higher available nitrogen levels, reflecting greater nutrient returns in feces, urine, decomposable plant litter, and higher net mineralization rates, leading to faster litter breakdown of corn residue.
4.2. N, P, and K release of Sorghum, Black Oat, and Corn Residue
Nitrogen Release
Nitrogen release from sorghum residue showed a rapid initial release when fertilized with 200 kg N ha (N-pasture). The results indicate that this practice ensures a consistent nitrogen supply and improves nitrogen availability during critical crop growth periods. This observation aligns with Shariff [18], who found that grazing and prior nitrogen applications significantly enhance nutrient cycling. Similarly, [25,26] showed that grazing nitrogen-fertilized cover crops improves subsequent maize yield without requiring additional nitrogen topdressing.
A recent study by [27] confirms that optimal nitrogen fertilization timing enhances nitrogen release and uptake in subsequent crops. The nitrogen immobilization episodes observed in the sorghum residue confirm the three-phase nitrogen release model described by [28], indicating that rapid nitrogen release is followed by microbial immobilization and nitrogen loss through mineralization. The modification in nitrogen release due to prior fertilization aligns with findings by [29], where substrate composition and exogenous nitrogen affect nitrogen dynamics.
The rapid nitrogen release from black oat residue in N-pasture plots supports the idea that fertilization and grazing can accelerate nutrient release by altering the decomposition environment [19,20]. Moreover, slow-release nitrogen fertilizers combined with biochar and organic amendments have been shown to synchronize nitrogen release with plant nutrient demand [4].
Phosphorus Release
Phosphorus release showed a pattern of rapid initial release, followed by immobilization. This pattern is consistent with other research, where phosphorus immobilization by microorganisms is linked to residue composition and soil phosphorus availability. In phosphorus-deficient soils, microbial immobilization can be beneficial by increasing phosphorus uptake and preventing adsorption to soil particles [30,31].
Recent research by [32] has demonstrated that phosphorus availability is further influenced by cover crops and microbial interactions that enhance soil phosphorus retention. In particular, the immobilization phases observed for black oat and corn residue confirm the importance of optimizing phosphorus fertilization to account for both microbial activity and soil adsorption dynamics.
Potassium Release
Potassium was released quickly from all residues, with an active fraction and half-life indicating rapid uptake by subsequent crops. This finding is consistent with global insights that potassium cycling is faster than nitrogen and phosphorus, leading to shorter availability periods and a higher risk of leaching. To minimize potassium losses, subsequent crops should be planted soon after cover crop management [33].
The consistent pattern of potassium release across residues aligns with other research, suggesting that maintaining potassium levels requires efficient crop rotation to reduce the impact of leaching. [34] recently highlighted the importance of potassium fertilizer application timing, as well as integrating organic amendments to optimize potassium uptake and reduce environmental losses.
5. Conclusions
The effects of pasture height and nitrogen (N) fertilization during the pasture phase significantly influenced dry matter decomposition and nutrient release throughout the grain crop phase. This finding indicates that these factors have a lasting impact on the integrated crop-livestock system and should be carefully considered when developing fertilization recommendations. Modifications in decomposition and nutrient release rates can alter soil fertility and nutrient availability, emphasizing the need for adaptive nutrient management practices.
Potassium (K) was released faster and in greater amounts than other nutrients, and the kinetic release of K was similar across all residues. This relationship was well described by the equation (). This uniform kinetic release pattern demonstrates the need to optimize potassium fertilization timing to reduce leaching losses and maximize crop uptake.
Applying N fertilizer during the pasture phase prevented the immobilization of N and phosphorus (P) in black oat residue and minimized immobilization in corn residue. This highlights the importance of optimizing N fertilization timing, as well as integrating effective grazing and residue management practices to enhance nutrient cycling and ensure sustainable crop production. These findings provide valuable insights for developing nutrient management strategies that improve soil fertility and crop yields while reducing environmental impacts.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, Tangriani Simioni Assmann; Marcos Antonio de Bortolli; Angela Bernadon and Marcieli Maccari; methodology, Tangriani Simioni Assmann; Marcos Antonio de Bortolli; Alan J. Franzluebbers; software, Betania Brum de Bortolli; Igor Kieling Severo and Jorge Jamhour; validation,Tangriani Simioni Assmann; Betania Brum de Bortolli and André Brugnara Soares; formal analysis, Betania Brum de Bortolli; investigation,Tangriani Simioni Assmann; Marcos Antonio de Bortolli; resources, Tangriani Simioni Assmann; Marcos Antonio de Bortolli; Angela Bernadon and Marcieli Maccari; data curation, Betania Brum de Bortolli and Igor Kieling Severo; writing—original draft preparation,Tangriani Simioni Assmann; Betania Brum de Bortolli and Igor Kieling Severo; writing—review and editing, Tangriani Simioni Assmann; Betania Brum de Bortolli and Igor Kieling Severo; visualization, Tangriani Simioni Assmann; Betania Brum de Bortolli and Igor Kieling Severo; supervision, Tangriani Simioni Assmann; Betania Brum de Bortolli and André Brugnara Soares; project administration, Tangriani Simioni Assmann; and Marcos Antonio de Bortolli; funding acquisition, Tangriani Simioni Assmann. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed in part by the Coordination of Improvement of Higher-Level Personnel (CAPES, Brasília, Brazil) - Finance Code 001.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge the LabSolos and LAQUA from the Universidade Tecnológica Federal do Paraná (UTFPR), Câmpus Pato Branco, for the analytical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Assmann, T.S.; de Bortolli, M.A.; Assmann, A.L.; Soares, A.B.; Pitta, C.S.R.; Franzluebbers, A.J.; et al. Does cattle grazing of dual-purpose wheat accelerate the rate of stubble decomposition and nutrients released? Agriculture, Ecosystems & Environment, 2014, 37–42.
- Maccari, M.; Assmann, T.S.; Bernadon, A.; Soares, A.B.; Franzlubbers, A.; Bortolli, M.; et al. Grazing intensity effects on litter decomposition and soil nitrogen mineralization. European Journal of Agronomy, 2021, 126317.
- Ahrends, H.; et al. Nutrient supply affects the yield stability of major European crops—a 50 year study. Environmental Research Letters, 2020, 014003.
- Zhang; Li; Wang; Qi Biochar and Slow-Release Nitrogen Fertilizers Improve Nutrient Synchronization in Crops. Nutrient Management Review, 2022, 321–335.
- L. Alves and others. Soil acidification and P, K, Ca and Mg budget as affected by sheep grazing and crop rotation in a long-term integrated crop-livestock system in southern Brazil. Geoderma, 2019.
- B. L. Rueda and others. Nutrient status of cattle grazing systems in the western Brazilian Amazon. Cattle Systems Research, 2020, 001–010. [CrossRef]
- E. Vogel and others. Environmental and economic performance of paddy field-based crop-livestock systems in Southern Brazil. Agricultural Systems, 2021, 103–109. [CrossRef]
- Germana Gomes dos Santos Camelo. Nutrient flows in small-scale farm production systems from Northeastern Brazil. Journal of Agricultural Science, 2018, 134–145. [CrossRef]
- CQFS-RS/SC C. Manual de adubação e calagem para os estados do Rio Grande do Sul e Santa Catarina. Porto Alegre: Sociedade Brasileira de Ciência do Solo Porto Alegre; 2004.
- Tedesco M, Gianello C, Bissani C, Bohnen H, Volkweiss S. Análise de solo, plantas e outros materiais. 2nd ed. Porto Alegre; 1995.
- Paul, Eldor Alvin and Clark, Francis Eugene. Soil microbiology and biochemistry. Soil Microbiology and Biochemistry, 1996.
- Cruz CDa. GENES - a software package for analysis in experimental statistics and quantitative genetics. Acta Scientiarum, 2013, 35(3):271-6.
- Barbosa Teixeira, Michelle AND Loss, Arcãngelo AND Gervasio Pereira, Marcos AND Pimentel, Carlos. Decomposição e ciclagem de nutrientes dos resíduos de quatro plantas de cobertura do solo. Idesia (Arica), 2012, 55–64. [CrossRef]
- Torres, José Luiz Rodrigues and Pereira, Marcos Gervasio. Dinâmica do potássio nos resíduos vegetais de plantas de cobertura no Cerrado. Revista Brasileira de Ciência do Solo, 2008, 1609–1618. [CrossRef]
- Semmartin, María and Garibaldi, Lucas A and Chaneton, Enrique J. Grazing history effects on above-and below-ground litter decomposition and nutrient cycling in two co-occurring grasses. Plant and Soil, 2008, 177–178.
- Sanaullah, Muhammad and others. How does plant leaf senescence of grassland species influence decomposition kinetics and litter compounds dynamics? Nutrient Cycling in Agroecosystems, 2010, 159–171.
- Jaramillo, David Mirabedini and others. Litter mass, deposition rate, and decomposition in nitrogen-fertilized or grass–legume grazing system. Crop Science, 2021, 2176–2189. [CrossRef]
- Shariff AR, Biondini ME, Grygiel CE. Grazing intensity effects on litter decomposition and soil nitrogen mineralization. Rangeland Ecology & Management/Journal of Range Management Archives, 1994, 444–449.
- Bardgett, Richard D and others. Linking above-ground and below-ground interactions: how plant responses to foliar herbivory influence soil organisms. Soil Biology and Biochemistry, 1998, 1867–1878.
- Sankaran, Mahesh and Augustine, David J. Ecology, 2004, 1052–1061.
- Olofsson, Johan and Oksanen, Laur. Role of litter decomposition for the increased primary production in areas heavily grazed by reindeer: a litterbag experiment. Oikos, 2002, 507–515.
- Semmartin, Maria and Oyarzabal, Mariano and Loreti, Juan and Oesterheld, Martin. Controls of primary productivity and nutrient cycling in a temperate grassland with year-round production. Austral Ecology, 2007, 416–428.
- Aber, John D and others. Factors controlling nitrogen cycling and nitrogen saturation in northern temperate forest ecosystem. Ecological Applications, 1991, 305–315.
- Vivanco, Lucía and Austin, Amy T. Intrinsic effects of species on leaf litter and root decomposition: a comparison of temperate grasses from North and South America. Oecologia, 2006, 97–107.
- Assmann, Tangriani Simioni and others. Rendimento de milho em área de integração lavoura-pecuária sob o sistema plantio direto, em presença e ausência de trevo branco, pastejo e nitrogênio. Revista Brasileira de Ciência do Solo, 2003, 675–683.
- Sartor, Laércio Ricardo and others. Effect of swine residue rates on corn, common bean, soybean and wheat yield. Revista Brasileira de Ciência do Solo, 2012, 661–669.
- Teixeira, Marcos and Oliveira, Flavia and Santos, Rafael. Optimized Nitrogen Timing Enhances Nutrient Release and Uptake in Subsequent Crops. Journal of Sustainable Agriculture, 2021, 567–578.
- Berg, B and Staaf, H. Leaching, accumulation and release of nitrogen in decomposing forest litter. Ecological Bulletins, 1981, 163–178.
- Pastor, John and Naiman, Robert J and Dewey, Brad. A hypothesis of the effects of moose and beaver foraging on soil nitrogen and carbon dynamics, Isle Royale. Alces, 1987, 107–124.
- Conte, Elaine and Anghinoni, Ibanor and Rheinheimer, Danilo dos Santos. Fósforo da biomassa microbiana e atividade de fosfatase ácida após aplicação de fosfato em solo no sistema plantio direto. Revista Brasileira de Ciência do Solo, 2002, 925–930.
- Rheinheimer, Danilo dos Santos and others. Comparação de métodos de determinação de carbono orgânico total no solo. Revista Brasileira de Ciência do Solo, 2008, 435–440.
- Basso, Andrea and Silva, João. Cover Crop and Microbial Interactions Enhance Soil Phosphorus Retention. Soil Science Advances, 2023, 89–101.
- Montgomery, Douglas C and Runger, George C. Applied statistics and probability for engineers. John Wiley & Sons, 2010.
- Wang, Xin and Lee, Yong. Optimizing Potassium Fertilizer Application Timing for Sustainable Agriculture. Agriculture & Environment, 2022, 45–58.
Figure 1.
Remaining dry matter (% and Mg DM ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time (N-Fert time, Sward Height and Days after deposition.
Figure 1.
Remaining dry matter (% and Mg DM ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time (N-Fert time, Sward Height and Days after deposition.
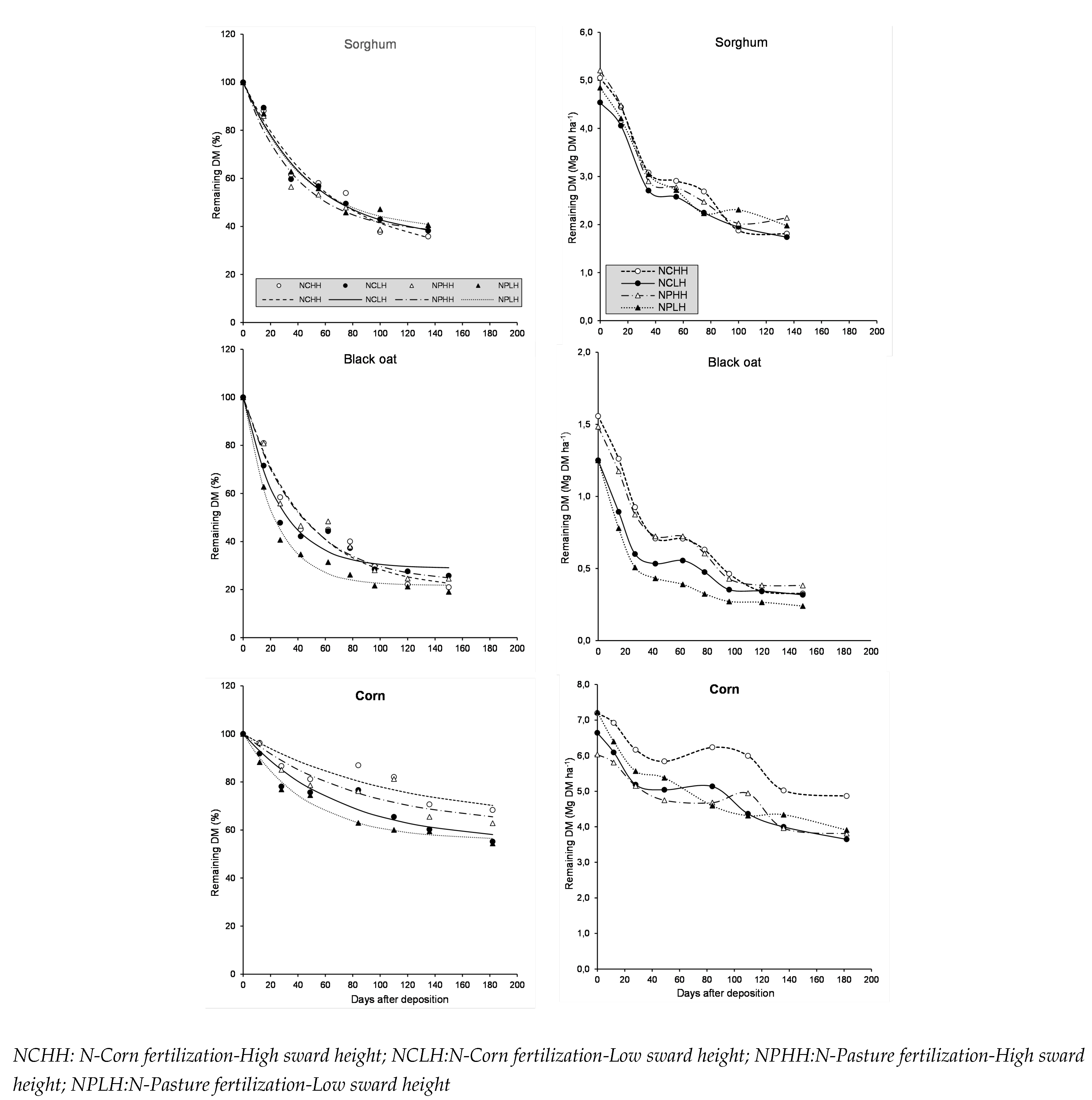
Figure 2.
Remaining nitrogen (%) and Released N (kg N ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time, Sward Height and Days after deposition.
Figure 2.
Remaining nitrogen (%) and Released N (kg N ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time, Sward Height and Days after deposition.
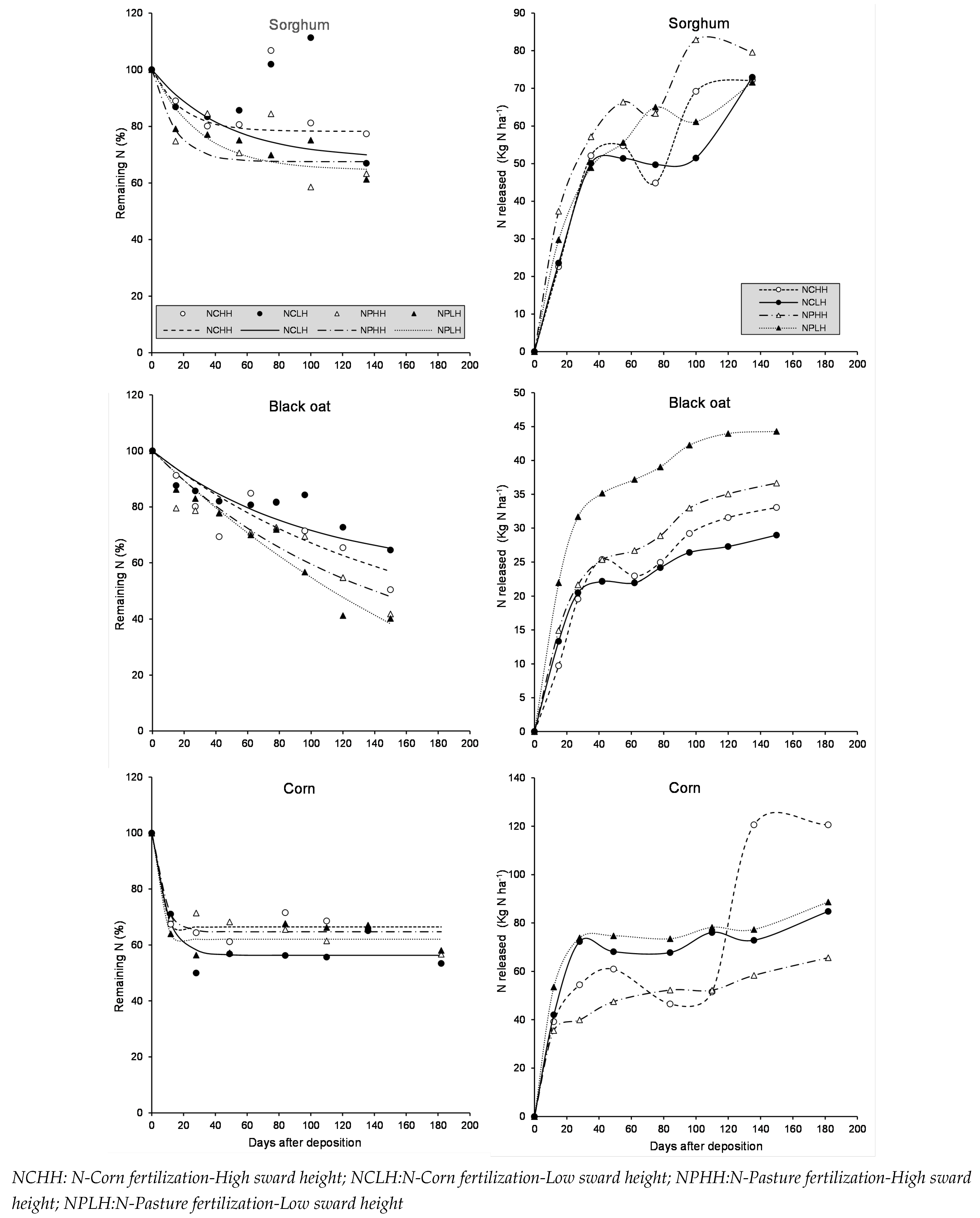
Figure 3.
Remaining phosphorus (%) and Released P (kg P ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time, Sward Height and Days after deposition.
Figure 3.
Remaining phosphorus (%) and Released P (kg P ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time, Sward Height and Days after deposition.
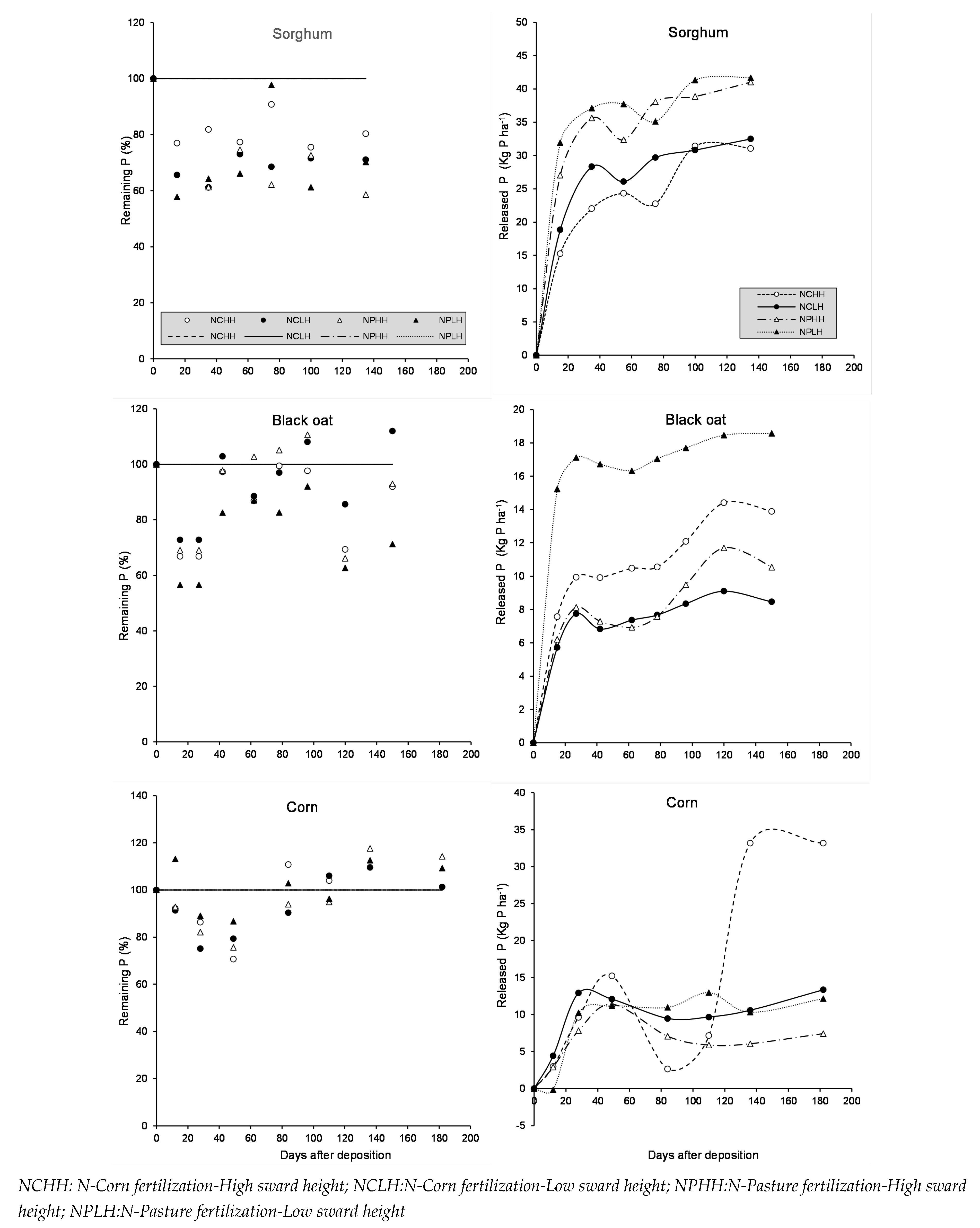
Figure 4.
Remaining potassium (%) and Released K (kg K ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time, Sward Height and Days after deposition.
Figure 4.
Remaining potassium (%) and Released K (kg K ha) of sorghum, black oat and corn residue during litter-bag exposure in the field as affected by N-Fertilization Time, Sward Height and Days after deposition.
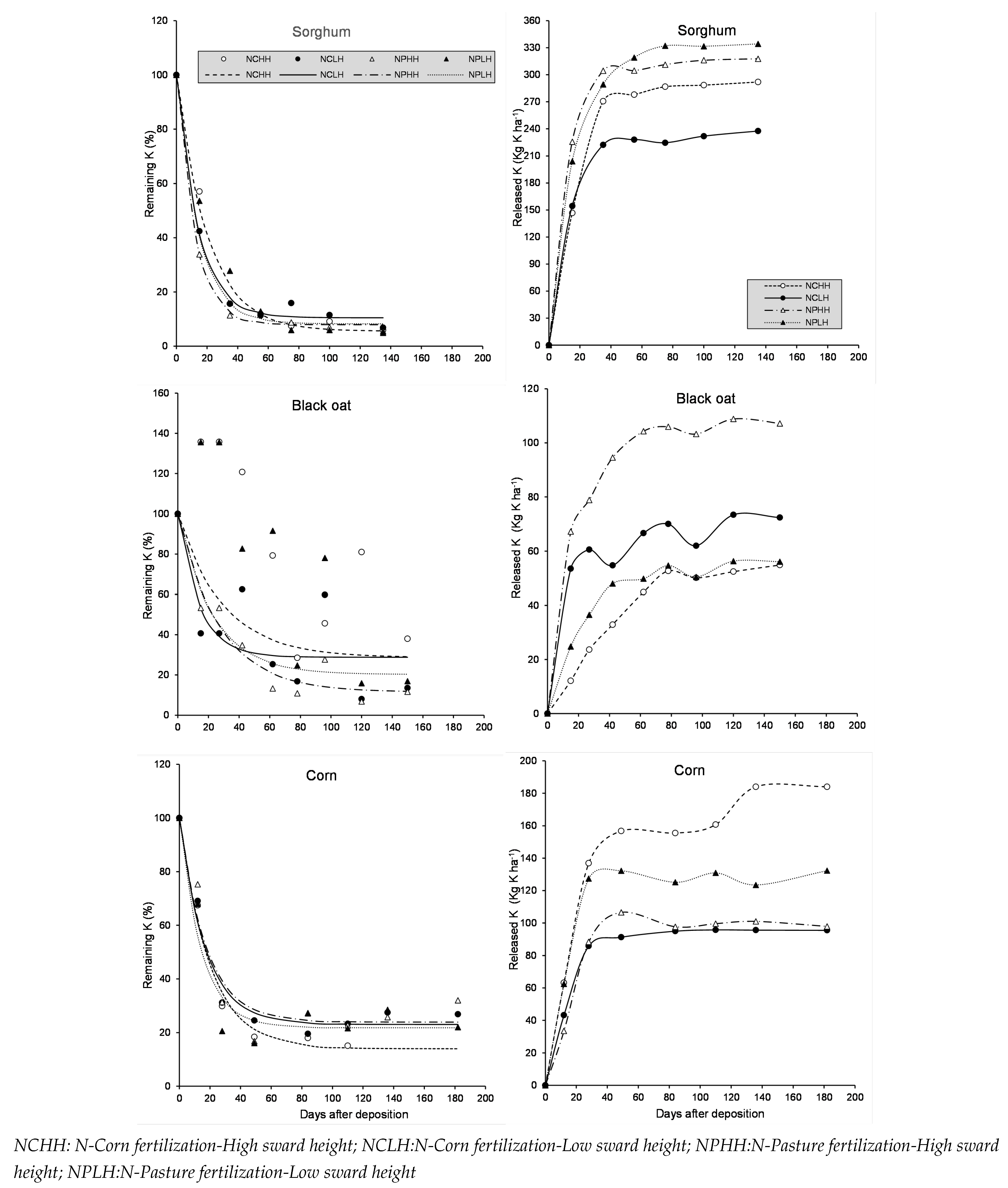
Figure 5.
Single model of remaining potassium of sorghum, black oat and corn residue during litter-bag exposure in the field. Each point is average of three observations, n=27.
Figure 5.
Single model of remaining potassium of sorghum, black oat and corn residue during litter-bag exposure in the field. Each point is average of three observations, n=27.
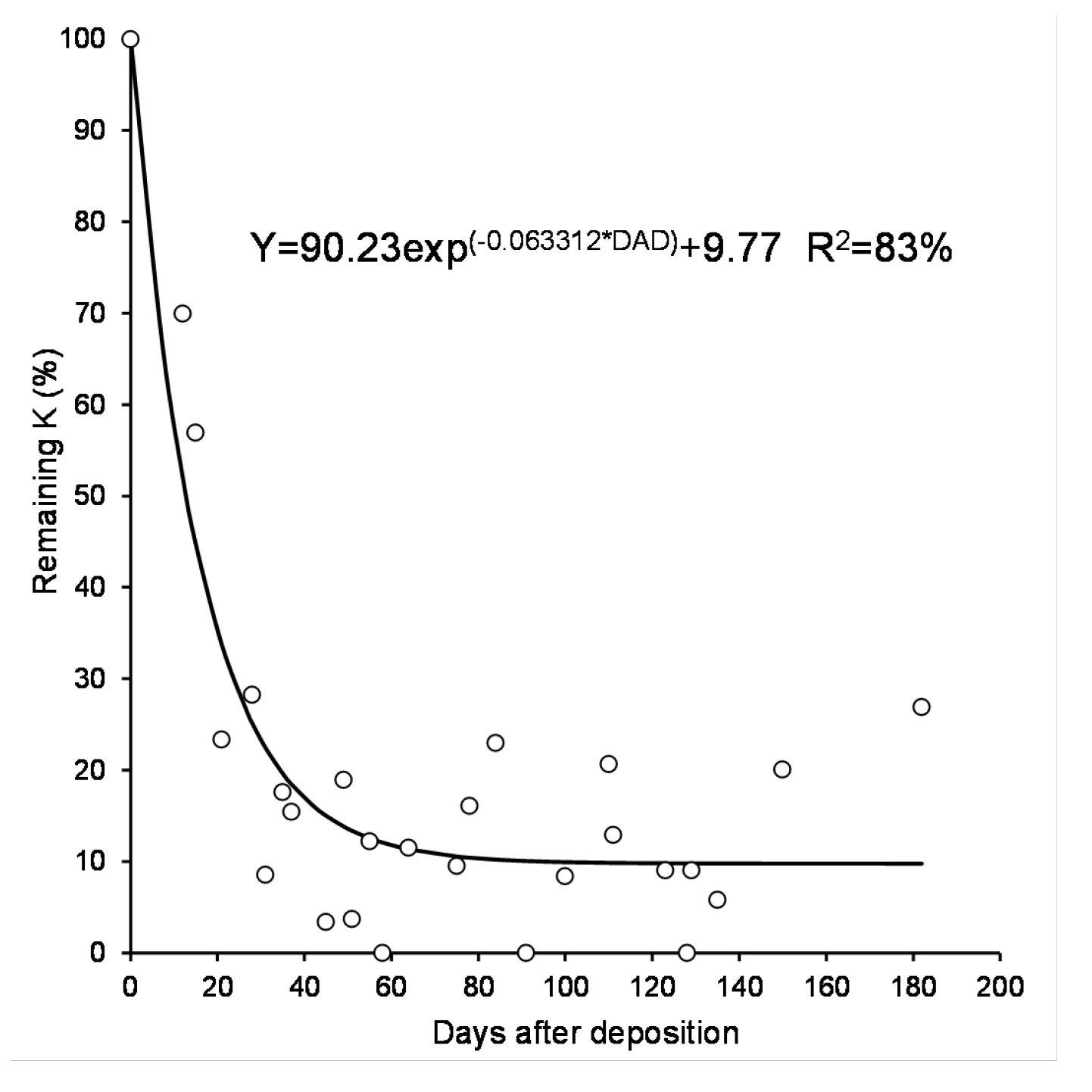

Table 1.
Initial residue characterization of sorghum, black oat and corn residue: Dry matter DM; N, P, and K concentration; Neutral Detergent Fibre NDF and Acid Detergent Fiber FDA in function of N-Fertilization Time (N-Fert) and Sward Height (SH).
Table 1.
Initial residue characterization of sorghum, black oat and corn residue: Dry matter DM; N, P, and K concentration; Neutral Detergent Fibre NDF and Acid Detergent Fiber FDA in function of N-Fertilization Time (N-Fert) and Sward Height (SH).
| N-Fert | SH | DM | N | P | K | FDN | FDA |
|---|---|---|---|---|---|---|---|
| kg | g kg | % | |||||
| Sorghum | |||||||
| NC | HH | 5048.6 | 19.83 +-3.54 | 8.53 +-1.16 | 59.17 +-5.92 | 52.97 +-1.85 | 30.87 +-1.20 |
| NC | LH | 4537.2 | 21.53 +-3.54 | 9.78 +-1.12 | 53.67 +-4.07 | 54.55 +-2.98 | 32.63 +-1.43 |
| NP | HH | 5204.7 | 20.40 +-2.94 | 10.43 +-0.64 | 61.83 +-7.64 | 55.46 +-0.49 | 31.40 +-0.44 |
| NP | LH | 4847.4 | 19.83 +-2.60 | 11.54 +-6.99 | 71.67 +-27.66 | 47.70 +-7.15 | 28.16 +-2.48 |
| Black oat | |||||||
| NC | HH | 1557.0 | 26.79 +-6.83 | 10.62 +-1.90 | 39.67 +-21.89 | 43.95 +-6.60 | 24.67 +-3.59 |
| NC | LH | 1250.3 | 21.96 +-6.19 | 9.54 +-0.31 | 60.17 +-3.33 | 46.62 +-3.25 | 25.70 +-1.29 |
| NP | HH | 1400.0 | 27.76 +-3.54 | 9.70 +-0.56 | 83.17 +-45.14 | 41.53 +-3.86 | 24.26 +-2.67 |
| NP | LH | 1250.7 | 38.52 +-4.28 | 16.27 +-10.55 | 47.67 +-24.58 | 47.76 +-1.11 | 28.11 +-1.22 |
| Corn | |||||||
| NC | HH | 7195.1 | 16.63 +-1.11 | 4.49 +-0.78 | 26.05 +-6.08 | 76.57 +-2.30 | 47.40 +-3.76 |
| NC | LH | 6639.0 | 18.30 +-2.19 | 4.42 +-0.89 | 17.33 +-5.06 | 74.66 +-4.20 | 46.55 +-5.53 |
| NP | HH | 6030.6 | 17.03 +-1.11 | 4.58 +-0.63 | 20.83 +-3.76 | 72.95 +-3.17 | 42.30 +-3.90 |
| NP | LH | 7227.0 | 18.03 +-1.25 | 4.17 +-0.92 | 21.00 +-3.50 | 72.95 +-2.82 | 39.42 +-1.13 |
| NC=N-Corn fertilization; NP=N-Pasture fertilization; HH=High Sward Height; LH=Low Sward Height. | |||||||
Table 2.
Parameters of non-linear model fitted to dry matter and nutrients (N and K) during field exposure of sorghum, black oat and corn residue as affected by N-Fertilization Time (N-Fert time and Sward Height (SH).
Table 2.
Parameters of non-linear model fitted to dry matter and nutrients (N and K) during field exposure of sorghum, black oat and corn residue as affected by N-Fertilization Time (N-Fert time and Sward Height (SH).
| Dry Matter | Nitrogen | Potassium | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N-Fert. Time |
Sward Height |
Ac. Fra % |
kA | T1/2 days |
R2 | Ac. Fra % |
kA | T1/2 days |
R2 | Ac. Fra % |
kA | T1/2 days |
R2 | ||
| Sorghum | |||||||||||||||
| NC | HH | 72.3 | 0.01670 | 42 | 90.1 | 21.8 | 0.05318 | 13 | 94.5 | 0.04878 | 14 | 91.5 | |||
| NC | LH | 65.7 | 0.02051 | 34 | 90.7 | 31.8 | 0.02168 | 32 | 89.5 | 0.07104 | 10 | 94.5 | |||
| NP | HH | 63.2 | 0.02572 | 27 | 93.5 | 32.5 | 0.07132 | 10 | 92.1 | 0,08466 | 8 | 97.0 | |||
| NP | LH | 61.9 | 0.02309 | 30 | 96.1 | 35.6 | 0.03256 | 21 | 91.8 | 0.06991 | 10 | ||||
| Black oat | |||||||||||||||
| NC | HH | 80.2 | 0.02235 | 31 | 95.1 | 71.0 | 0.00622 | 111 | 71.4 | 0.03382 | 20 | ||||
| NC | LH | 71.2 | 0.03753 | 18 | 95.5 | 44.4 | 0.01018 | 68 | 71.2 | 0.07131 | 10 | ||||
| NP | HH | 77.1 | 0.02426 | 29 | 91.1 | 78.4 | 0.00725 | 96 | 88.6 | 0.03634 | 19 | ||||
| NP | LH | 78.3 | 0.04483 | 15 | 96.0 | 133.4 | 0.00414 | 168 | 79.8 | 0.04324 | 16 | ||||
| Corn | |||||||||||||||
| NC | HH | 36.8 | 0.00911 | 76 | 69.1 | 33.6 | 0.29575 | 14 | 86.0 | 0.05045 | 14 | 93.2 | |||
| NC | LH | 45.3 | 0.01422 | 49 | 89.2 | 43.7 | 0.11133 | 6 | 77.0 | 0.05861 | 12 | 88.1 | |||
| NP | HH | 38.9 | 0.01196 | 58 | 83.0 | 35.3 | 0.14364 | 5 | 76.1 | 0.05701 | 12 | 87.6 | |||
| NP | LH | 44.5 | 0.02132 | 33 | 96.9 | 37.9 | 0.26027 | 3 | 78.2 | 0.06908 | 10 | 85.9 | |||
| NC=N-Corn fertilization; NP=N-Pasture fertilization; HH=High Sward Height; LH=Low Sward Height. | |||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



