
Submitted:
18 July 2024
Posted:
19 July 2024
You are already at the latest version
Abstract
Livestock production in the Cajamarca, La Libertad and Amazonas basins is based on pasture-based feeding with "Cajamarquino ecotype" ryegrass under monoculture or in association with white clover. The objective of this research work was the morphological characterization and evaluation of the productive yield and cutting time of annual ryegrass - Lolium multiflorum L. Six local genotypes of Lolium multiflorum in six locations were characterised, using the multiplicative interaction effect model and additive main effects for the analysis of interaction and variability of the genotypes and locations. Ryegrass “Cajamarquino ecotype” is semi-erect with intermediate growth, with a dark green colouring, 0.8 cm broadleaf, and can reach an average stem length of 46 cm, up to 1.6 cm.day, achieving the growth of the fourth leaf after 28 days under adequate management conditions. Despite the differentiated characteristics, according to the BLASTn evaluation, the ITS1 sequences showed an identification greater than 99.9% similar to Lolium multiflorum L. It was determined that the Cajamarquino ecotype produces from 3.52 to 4.98 Mg.ha.biomass cutting. Likewise, the pasture's quality is better when used at 30 days with 14.84% protein and at 45 days, it achieves 12.75% protein with a biomass of 4.05 Mg.ha.cut. With these morphological and yield characteristics, the "Cajamarca ecotype" ryegrass is a well-adapted species for local livestock farming. These species are adapted to the Sierra Norte area of Peru.
Keywords:
Lolium multiflorum L.
; Cajamarquino ecotype
; Andean pastures
; drought tolerance
; dairy farming
1. Introduction
Grasslands constitute over 70% of the world's agricultural land and play a pivotal role in food security, the economy, and global ecology due to their adaptability and functionality [1]. It is anticipated that climate change will intensify weather events, which will, in turn, lead to an increase in the frequency of droughts. This will harm forage production while exposing agricultural and silvopastoral systems to a heightened risk of imminent hazards [2]. Implementing measures designed to enhance genetic heterogeneity, topography, and grassland diversity can improve stress tolerance, mitigate damage, and facilitate recovery following periods of drought [3].
Lolium multiflorum, a grass species cultivated annually worldwide, stands out for its exceptional adaptability to diverse soil types and various climatic conditions. Its rapid establishment, high palatability, good digestibility, high yields of quality forage, and tolerance to diverse grazing methods make it a preferred choice for many farmers and producers. Its ability to reseed reduces costs and facilitates soil restoration, providing a reassuring solution in the face of climate change [4,5].
Ryegrass, in conjunction with other forage species (clover, vetch), is the primary source of feed for livestock. This is particularly true in hillside areas, where prolonged droughts impact pasture production, leading to a reversible or irreversible disruption of the plant's functioning and structure. This, in turn, affects the economy and livelihood of the livestock community in the northern highlands of Perú [6,7,8].
Lolium multiflorum L. is a grass species widely used in forage production due to its rapid growth and high productivity. This plant is adapted to altitudes between 2400 and 3200 m asl and temperatures between 12 and 18 °C. In addition to exhibiting resistance to trampling, it is considered a primary source of winter forage [9]. However, in highland areas, where acid soils are prevalent, the yield of Lolium spp. Varieties are adversely affected by low nutrient availability, yield deficit, and environmental stress [10]. The ryegrass-soil interaction is significant due to the exposure to such soils, given the grass's high adaptability, absorption, and fixation capacity. This substantially balances crowns and roots, thereby ensuring the grass's vigour against these challenging soils [11]. The Lolium family has demonstrated remarkable resistance and adaptability, enabling grasses in the northern highlands of Peru to thrive in low temperatures. This suggests that they possess a valuable source of frost and drought tolerance genes, as evidenced by their ability to activate the cold acclimatisation process [7].
Genetic improvement of Lolium spp. has resulted in the generation of specific genetic mutations, which have enabled the species to behave as a perennial under managed conditions due to its high competitiveness, genetic diversity, and seed production [12,13]. In recent decades, research has concentrated on enhancing yield, dry matter, and forage quality, thereby facilitating more effective crop management and enabling a more significant number of harvests per year. These productivity gains are contingent upon optimal humidity and temperature, an adequate pH level, organic matter, potassium, and phosphorus, which are essential for plant development and maturity. Nitrogen is also a crucial element, as it is vital for seedling production [14,15].
Understanding the germination behaviour of ryegrass seeds is crucial for effective integrated weed management strategies. The type of dormancy enriches seed ecology, the species' life cycle, and seasonal changes in environmental conditions, including temperature, precipitation, and soil salinity, from dispersal to germination [16]. Ryegrass seeds initially exhibit dormancy, which may be broken and result in germination occurring at different times throughout the growing season. This knowledge empowers effective weed management strategies [17].
Lolium multiflorum L., a species with a rapid growth rate, is characterised by its long and thin foliage with a glossy appearance and a distinctive erect and hollow stem. The root system is characterised by high fibrousness and superficiality, which enables rapid absorption of nutrients and water. In response to environmental cues, the plant initiates an adaptive process to prolong its vegetative stage, manifested in various morphological changes. These include a reduction in the height of the panicle, a decrease in the length of intact leaves, and an increase in the number of growing leaves [18].
A reduction in grazing intensity was associated with an enhanced leaf elongation rate, reaching 3.5 cm.day, and a diminished tillering density, with 150 tillers.m². Additionally, plant height, green forage yield, and dry matter ranged from 115.0 to 130.7 cm, 1407 to 3240 kg. ha, and 392 to 976 kg. ha, respectively. These results significantly impacted the attributes of ryegrass (Lolium multiflorum L.), indicating that moderate to light grazing increases spatial heterogeneity and productivity [19,20].
The morphological variation of Lolium multiflorum L. is interpreted based on its high phenotypic plasticity, the ability of an organism to change its phenotype in response to changes in the environment. This is the ability to alter its physiological characteristics in response to environmental variations and its hybridization (such as L. multiflorum x L. perenne). This highlights the need for effective localised control methods for this genus's different species and hybrids, thanks to its competitive nature, which makes it prone to developing resistance to herbicides. Applying pesticides that are harmful to forage and soil while reducing the occurrence of crown rot under different management conditions has resulted in variations in height ranging between 115.0 and 130.7 cm and green forage yields ranging between 1407 and 3240 kg. Ha, and dry matter production ranging between 392 and 976 kg. Ha [13,21].
It has been demonstrated that annual ryegrass can exhibit enhanced quality when subjected to water stress. Landscape heterogeneity indicates that, in specific locations, adding nutrients and soil amendments can significantly regulate pH and create favourable conditions for increased forage production. This is supported by evidence from studies by Abraha et al. (2015) [22] and Bridges et al. (2019) [23].
The Poaceae, including Lolium multiflorum L., play a significant role in the remediation of degraded soils. It mitigates these processes' deleterious effects on the environment. It facilitates the restoration of soils that can be utilised for agricultural production in the context of progressive soil degradation and depletion [24,25]. Cultivated as a cover crop, they have been demonstrated to enhance soil fertility, mitigate erosion, and curtail nutrient loss [26,27]. The sustainable management of soil offers solutions to the conservation and reduction of pollution by utilising plants adapted to the local climate and specific pollutants. These plants demonstrate effective responses to drought and salinity stress at the morphological, physiological, and biochemical levels. Consequently, the Poaceae are indispensable not only for sustainable food production but also for protecting the environment and managing ecosystems [28,29].
The appropriate requirement is fertiliser in ryegrass ranging between 100-150 kg.ha of N, 40-60 kg.ha of P, and 60-80 kg/ha of K. The correct application is crucial to increase yields and improve soil quality by maximising the yield and nutrient absorption of Lolium multiflorum L. in acid soils (pH 5.1 - 6.1). These practical recommendations can be readily implemented, thereby optimising milk production and farmers' profits while reinforcing the significance of proper soil management for attaining more efficient and sustainable agricultural practices [30,31].
The findings of this study are of significant value as they provide invaluable insights into the productive performance of Lolium multiflorum L. 'Cajamarquino ecotype' under acidic soil conditions in the northern highlands of Peru. The knowledge gained in this study can be used to inform recommendations on genotype management practices and cutting times, as well as the importance of yield differentiation under different climatic conditions.
2. Results
2.1. Yield Performance
Table 1 shows the superiority of the LM-58 genotype over LM-43 in green forage yield (20.97 Mg. ha), dry matter or biomass (4.49 Mg. ha), seed yield (259.23 Kg. ha) and protein value (13.48%); the LM-58 genotype shows a higher production capacity in terms of forage quantity and nutritional value in most locations. 48%); the LM-58 genotype shows a higher production capacity in terms of forage quantity and nutritional value in most locations, which represents a more advantageous option for optimising both agricultural productivity and forage quality.
2.2. Cutting time
Table 2 shows the crucial interactions between cutting days and biomass and protein production, highlighting the significance of our results that by increasing the cutting interval from 30 to 60 days, biomass produced increases significantly from 18.87 to 49.10 Mg. ha. yr (p < 0.001); similarly, protein production increases from 2.8 to 4.9 Mg. ha. yr (p = 0.0405). These results underline the direct influence of cutting time in optimising biomass and protein production, suggesting that a 60-day cutting interval may be more beneficial for maximising both parameters in the crops evaluated.
The effect of the interaction between cutting time (C) and location (L) on biomass and protein of Lolium multiflorum L. Figure 1 shows in detail the interaction between C x L, highlighting their effect on biomass and protein indicators of Lolium multiflorum L. Panel (A) shows the biomass production expressed in Mg. ha. year. At the same time, panels (B), (C) and (D) show the protein ratios in percentage (%), Mg ha. cut and Mg ha. year, respectively, for the genotypes LM-58 and LM-43. This analysis shows that the Baños del Inca site had the best yields in terms of biomass and protein content in Mg. ha. yr. These results highlight the importance of taking into account both the timing of cutting to optimise yield and quality of Lolium multiflorum L. crops, providing relevant information for farm management and improvement of cultivation practices. Taking into account the interaction for protein percentage (p= 0.0036)
2.3. Morphological characteristics
Table 3 shows the differentiation and morphological characterization of Lolium multiflorum L. for genotypes LM-58 and LM-43. The results highlight the superiority of the LM-58 genotype, which showed better results in the evaluated characters compared to the LM-43 genotype, which obtained lower results in all these parameters.
Table 4 shows a comprehensive morphological characterization of the qualitative characteristics of the genotype LM-58 of Lolium multiflorum L. This analysis reveals the diversity and complexity of the morphological characters of the genotype, which provides a detailed and multifaceted view of its behaviour and physical attributes throughout its growth and development.
Table 5 shows the evaluation of the qualitative characteristics of the genotype Ryegrass LM - 58, which was carried out with meticulous attention to detail, the results showed total uniformity in several aspects, where no atypical plant showed a different colour for the "colour of the lemma", which indicates uniformity in this character. Similarly, we observed uniformity and absence of atypical seeds for the presence of "kernel ridge", with no variation in the level of expression. This assessment was carried out visually by observing 30 plants, confirming their homogeneous classification in both assessment seasons.
Table 6 shows that the vegetative cycle of Ryegrass LM-58 exhibits an impressively early expression at 105 days, with coefficients of variability of 2.80% and 2.73% for the first and second seasons, respectively. Similarly, the "time of flower initiation" demonstrated an early expression at 40 days, with coefficients of variability of 2.91% and 2.59% for the first and second seasons, respectively. The classification of the variety as uniform according to UPOV regulations is supported by the observation that both characteristics exhibit coefficients below the permitted limit of 3% for self-pollinated varieties.
The analysis carried out to identify the mitochondrial DNA sequences of Lolium multiflorum L., genotype LM-58, for which a bioinformatic analysis was carried out where the ITS1 sequence haplogroup (ITS1 = INIA LM-58) differs in three mutational steps from the reference sequence of Lolium multiflorum L. (ecotype Cajamarquino). As can be seen in supplementary material S1.
3. Discussion
In this study, we evaluated the morphological differentiation, productive yield, and cutting time of Lolium multiflorum' Cajamarquino ecotype' under the challenging acid soil conditions of the northern highlands of Peru. Our impressive results indicate that the Cajamarquino ecotype adapts to unfavourable conditions. This adaptability is reflected in its morphological characteristics and productive yield. Even when faced with the primary source of abiotic stress, drought, which can severely impact plant growth and development, the Lolium multiflorum ecotype Cajamarquino produced favourable results regarding green forage and dry matter yields.
3.1. Performance yield
Green forage yield is influenced and conditioned by several factors, such as climate, grazing frequency and intensity, and cutting time [32,33]. Ryegrass is grown in winter because of its adaptability to various soils; this cover crop is valued for its cold hardiness, extensive root system and high biomass production. It is essential to remember that plant productivity is closely linked to the mineralization of soil nutrients and their uptake and utilisation by plants. This process involves intense interactions between roots, microflora and soil fauna [26,34].
It is worth noting that Lolium multiflorum L. boasts a higher nutritional value than Lolium perenne. Its rich concentration of vitamins and antioxidants makes it a highly sought-after species in the market, offering significant health benefits to consumers [35].
The dry matter yield of ryegrass is influenced by several factors, including soil moisture level, type and method of planting, number of harvests and fertilisation. Also, the soil texture and the competitive ability of Lolium multiflorum to grow on different soil types, on which the dry matter productivity will depend [36,37].
It is noteworthy that in the results obtained and as stated by the authors, a particular genotype of Lolium multiflorum L. exhibits superior performance in terms of green fodder yield. Although both genotypes, LM-58 and LM-43, are Cajamarquino ecotypes, the LM-58 genotype exhibits significantly higher yield compared to the LM-43 genotype. This dominance can be attributed to the superior characteristics exhibited by the LM-58 genotype, including greater leaf length and width, intense and dark leaf colour, and greater plant height after vernalisation, among others. These characteristics, which are corroborated in detail in Table 3 and Table 4, indicate not only a higher yield in terms of biomass produced, but also a higher efficiency in nutrient uptake. In addition, the LM-58 genotype has shown greater resilience to adverse climatic and soil conditions, which is vital for its adaptation and productivity in the locations evaluated. The above characteristics make the LM-58 genotype a more viable and promising option for farmers seeking to maximise green forage production.
As for the locations evaluated, despite being in geolocations with similar conditions, green forage yields varied from location to location; this could be due to differences in cropping seasons across years and evaluation periods. Factors such as microclimatic variations, specific agronomic practices, and soil quality may have influenced these results, however, the Baños del Inca location showed the highest yield in both production seasons, as detailed in Table 8 and Table 1. This consistency suggests that Baños del Inca offers more favourable conditions for growing Lolium multiflorum L. than the other locations. These findings highlight the importance of carefully selecting locations to maximise crop production and suggest that Baños del Inca could serve as a model for successful agricultural practices in the region.
3.4. Cutting time
The analyses revealed that the forage quality of the Cajamarquino ecotype is determined by the moment of cutting according to the dry matter (DM) yield per hectare and the percentage of protein. Three groups of analysis were formed with samples every 30, 45 and 60 days, showing that at 30 days, the best result was obtained with a protein content of 14.84%, as opposed to 45 days, while biomass increases in this period, protein content decreases to 12.75%, suggesting that the optimum cutting time to maximise forage quality is at 30 days to obtain better utilisation.
The study showed that 90% of ryegrass seedlings emerging on the first day compared to other species was due to the mowing period's ability to manage ryegrass dominance. The time to first clip and repeat clipping on seedling growth depends on the impact of mowing height, as defoliating according to soil height minimises damage to the tillers' growing points. It was also found that 22-50% of the variability in growth at 60 days was due to these initial treatments [38,39].
This study shows that cutting times are critical factors in determining the quality and quantity of Lolium multiflorum L. forage. A 30-day cutting interval is optimal for maximising protein content, while adequate cutting height can significantly improve grass recovery and growth. Furthermore, the variability in green forage yield between sites underlines the importance of taking local conditions into account in agronomic practices. Finally, it is noteworthy that the genotype LM-58, in interaction with the sites, obtained a significant value (p = 0.0036) for protein percentage. These data provide valuable information for genetic improvement and selection in forage production and show that this genotype outperformed the other local genotypes, as confirmed in Table 2.
3.3. Morphological differentiation
Morphological differentiation among local genotypes of Lolium multiflorum L. is essential to understanding its adaptation and performance under various environmental conditions. The study results indicate that genotype LM-58 has superior characteristics to genotype LM-43. In the parameters evaluated in growth habit, LM-58 exhibited semi-erect growth and a greater intensity of dark green colour in its leaves, contrasting with the more upright growth and light green colour of LM-43. In addition, LM-58 showed a greater width of the last leaf (0.8 cm vs. 0.6 cm for LM-43) and a higher number of seeds per spikelet (13 vs. 10 by LM-43). The growth rate of LM-58 was also higher, reaching 1.6 cm per day compared to 1.2 cm per day for LM-43, allowing faster recovery after cutting and higher forage production in a shorter period. Age at the fourth leaf was earlier in LM-58 (28 days) than LM-43 (35 days), indicating a higher earliness of LM-58, which may benefit faster production cycles.
Bioinformatic analysis of mitochondrial DNA sequences shows the identity of Lolium multiflorum L. and identified two distinct haplotypes. The LM-58 genotype clustered into one haplotype together with other samples under study, differing from the reference sequence of the LM-43 genotype. Cross-pollinated species, such as ryegrass, show high genetic variability within and between populations. Thus, L. multiflorum and the wind dispersal mating system promote high genetic variability within populations but low genetic differentiation between populations [40,41]. This genetic variability facilitates that the haplotypic network constructed shows two unique haplotypes: haplotype 1 containing the three samples under study and haplotype 2 containing the reference sequence, thus reinforcing the genetic differentiation of the LM-58 genotype, as can be seen in the supplementary material S1. Finally, the LM-58 genotype showed increased resilience to adverse climatic and soil conditions, vital for its adaptation and productivity in the locations evaluated. These characteristics make LM-58 a more viable and promising option for maximising green forage production.
4. Materials and Methods
4.1. Place and duration of study
This research was conducted in the Cajamarca Region - Peru, which had two experimental stages. The first phase was developed in the agricultural campaigns 2017 - 2018; the second campaign was created in the periods 2018 - 2019, where the productive yield parameters (Green Forage, Dry Matter, and time of cutting) were evaluated in two local genotypes LM - 43 and LM - 58, as detailed in Table 8, which was developed in 6 locations located in the region of Cajamarca, department of Cajamarca - Peru, with average annual temperature of 15.6°C, average relative humidity of 79% and an average annual rainfall 2963 mm/year [42], this research was evaluated in December, a suitable time for installing this forage species.
4.2. Experimental units
To determine the productive yield of Lolium multiflorum L., a Completely Randomised Block Design (CRBD) was used in 6 locations in the Cajamarca Region. This design, known for its robustness and reliability, was the perfect choice for our experiment. Rye Grass LM - 58 and LM - 43 genotypes were evaluated; each variety was sown in 6 m x 5 m plots, installing six plots in each location, with two replications randomly.
The treatments were divided according to the phenological stage of the Rye Grass Lolium multiflorum L., where T1 Genotype LM - 58 and T2 = Genotype LM - 43. It is essential to mention that the sowing system was broadcast in the 6 locations with a sowing density of 30 kg. ha of seed. Fertilisation was carried out in two seasons, each with two key fertilisation moments; in the first fertilisation at sowing, agricultural Urea (N) = 80 kg.ha.year, Triple Calcium Superphosphate (P2O5) = 130.4 kg.ha.year, Potassium Chloride KCl (K2O) = 66.4 kg.ha.year and in the second tillering fertilisation consisted only of Urea = 80 kg.ha.year, using the same fertilisation rates for both campaigns.
4.3. Experimental material
The experimental material was derived from a backcrossing process involving a population of previously selected elite genotypes. The experimental treatments comprised the genotypes LM-58 and LM-43 of Lolium multiflorum L., which have undergone two selection processes. The initial phase involved 620 accessions, while the subsequent phase encompassed 21 exceptional emerging lines selected over 25 years.
Table 8 presents the detailed periods and critical activities of the study, such as the initial selection based on growth habit, anthesis, and pest and disease resistance, followed by propagation and polycrossing to enhance desirable traits. The research, which began in 1995 with the collection and germination of seedlings, progressed through several phases of characterization, selection, and field trials. After numerous tests, the genotype LM-58 was identified as an elite of Lolium multiflorum L., Cajamarca ecotype, marking a significant milestone in our research.
4.4. Sampling
We evaluated the morphological differentiation, yield, and cutting time of the Lolium multiflorum L. genotypes LM—58 and LM—43 with utmost precision. We obtained the samples using a square metre, manually cut the forage with a sickle, and weighed it fresh on an electronic balance. A 100g sample was then taken to the laboratory for further analysis, including determining the percentage of dry matter and calculating Biomass yield.
The amount of seed harvested was recorded per square metre, while plant height was measured with a ruler, averaging the values for each after forage cutting. Six samples were taken in each treatment to determine the best cutting time according to the phenological stage of the association. These samples were left to wilt for 12 hours, reduced to 5 cm in size, and placed in 50 kg polyethene bags.
For the chemical analysis, we followed the rigorous AOAC methodologies (AOAC, 1990) to ensure the accuracy and reliability of our results. The samples were dried in a forced air oven, ground to a specific particle size, and then analysed for Crude Protein (CP), Crude Fibre (CF), Ethereal Extract (EE), ash, and Free Nitrogen Extract (FN).
For seed production, a uniform cut was made before flowering to achieve uniform and synchronised growth between flowering and fruiting.
4.5. Evaluation characteristics
Following the comprehensive guidelines of the Union for the Protection of New Varieties of Plants (UPOV), the evaluation process was conducted at a single location and over two similar vegetation periods. Under these stringent conditions, the local lines LM-58 and LM-43 underwent a thorough evaluation in two cycles of identification trials.
- − Cycle 1: In the 2017 - 2018 season.
- − Cycle 2: During the 2018 - 2019 campaign.
According to the regulations for generating new cultivars established in Decision 345 in Article 10° and as established by the Head Resolution N° 047-2000-INIA, Chapter VII, Article 12.- A variety is considered distinct if one or several distinctive characteristics are notoriously different from any other commonly known variety. It is sufficient that only one characteristic is distinct for the variety to be considered distinct, Article 21.- The Distinctness test, a crucial part of the evaluation process, must include the commonly known varieties with a more remarkable similarity to the variety for which protection is sought. For quantitative characteristics, the hypothesis of equality will be evaluated, and only if this hypothesis is rejected with a significance level of 5% probability will it be possible to conclude that the varieties are different. If the characteristic is qualitative, the variety to be protected must show a different characteristic status compared to other varieties.
The evaluations of the genotypes LM - 58 and LM - 43 were carried out according to the UPOV Codes: LOLIU_PER; LOLIU_MUL_ITA; LOLIU_MUL_WES; LOLIU_BOU; LOLIU_RIG. These codes, internationally recognized standards for the distinctness, uniformity, and stability (DUS) testing of plant varieties, were applied through the parameters for cultivating semi-perennial and perennial forage grasses. The criteria established by the International Union for the Protection of New Varieties of Plants [43] were used as a reference, and the focus was on the morphological characterization of Lolium multiflorum L., local cultivars LM - 58 and LM - 43. The evaluations unveiled these grasses' fast-growing and robust nature, signalling their promising adaptability to various soil types and climatic conditions. They also demonstrated efficient nutrient uptake and soil anchorage, further bolstering their potential.
Table 9 presents a detailed evaluation of the qualitative morphological characteristics of the genotypes LM-58 and LM-43 of Lolium multiflorum L., covering various physical and developmental attributes of the genotypes evaluated. This characterization highlights both differences and similarities between the genotypes, providing valuable information for the implementation of breeding programs and optimised agronomic practices.
4.5. Statistical analysis
Prior to undertaking the statistical analysis of differences, a comprehensive test for normality of the data (Levene, p < 0.05) and a thorough check for homogeneity of variances (Kolmogorov-Smirnov, p < 0.05) were conducted. Subsequently, Duncan's test was employed to ascertain the statistical significance of the factors and to identify the most prominent line. The data were analysed using Infostat software (version 2020).
5. Conclusions
The results obtained in the research have fundamental implications for agriculture in regions with acid soils due to the extraordinary capacity of Lolium multiflorum "cajamarquino ecotype" to maintain an excellent productive yield under adverse conditions, which suggests that it is a viable option to improve forage production in the northern highlands of Peru. It will contribute to the sustainability of diverse agricultural practices and improve rural communities' sustainability, resilience and economy. Correctly managing cutting time and fertilisation is critical to maximise forage yield and quality. The study of Lolium multiflorum L., ecotype 'Cajamarquino,' under acid soil conditions in the northern highlands of Peru has yielded essential results with important implications for agriculture in these regions. The ecotype 'Cajamarquino' LM-58 showed remarkable adaptability to the different geographical conditions of the six sites studied, maintaining high green fodder and dry matter yields and suggesting its viability for improving fodder production in these areas. The detailed analysis of the morphological characteristics of the LM-58 and LM-43 genotypes revealed valuable differences and similarities for the implementation of breeding programs and optimised agronomic practices, highlighting the superiority of the LM-58 genotype for its greater resilience and efficiency in the production of significant biomass yields, with a protein content of 14.84% at 30 days and 12.75% at 45 days. In addition, bioinformatic analysis of mitochondrial DNA sequences identified two distinct haplotypes, reinforcing the genetic differentiation of LM-58 and consolidating it as a promising option for farmers seeking to maximise green fodder production. In conclusion, the LM-58 genotype of the 'Cajamarquino' ecotype of Lolium multiflorum L. is a species well adapted to local livestock production, capable of maintaining excellent productive performance, and represents a viable alternative for improving forage production in the highlands of northern Peru.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Suplementary 1: Genomic evaluation on Lolium multiflorum L.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, W.C.-Ch., M.C.-P. and W.A.-G.; methodology, W.C.-Ch., M.C.-P., L.M., H.V. and W.A.-G.; software, L.M., Y.M.-V. and W.A.-G.; validation, W.C.-Ch., M.C.-P., Y.M.-V., C.Q.and H.V.; formal analysis, L.M., C.Q., H.V. and W.A.-G.; investigation, W.C.-Ch., M.C.-P., D.C.N.-M., H.V. and W.A.-G.; resources, W.C.-Ch., M.C.-P., C.Q., H.V; data curation, L.M., Y.M.-V. and W.A.-G.; writing—original draft preparation, W.C.-Ch., Y.M.-V., L.M., C.Q., H.V. and W.A.-G.; writing—review and editing, L.M., C.Q., D.C.N.-M. and W.A.-G.; visualization, L.M., Y.M.-V., C.Q., D.C.N.-M., H.V. and W.A.-G.; supervision, M.C.-P., C.Q., D.C.N.-M. and W.A.-G.; project administration, W.C.-Ch., D.C.N.-M. and H.V; funding acquisition, W.C.-Ch, C.Q., D.C.N.-M. and H.V. All authors have read and agreed to the published version of the manuscript.”
Funding
This work was financed with resources from Project CUI 2432072: ‘Mejoramiento de la disponibilidad de material genético de ganado bovino con alto valor a nivel nacional. 7 departamentos’ of the Ministry of Agrarian Development and Irrigation - Peru.
Data Availability Statement
Data sharing is not applicable to the article as datasets were generated or analysed during the current study.
Acknowledgments
In this section, we thank P. Mantilla-Rudas for field support and sample collection.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Loka, D.; Harper, J.; Humphreys, M.; Gasior, D.; Wootton-Beard, P.; Gwynn-Jones, D.; Scullion, J.; Doonan, J.; Kingston-Smith, A.; Dodd, R.; et al. Impacts of Abiotic Stresses on the Physiology and Metabolism of Cool-Season Grasses: A Review. Food and Energy Security 2019, 8, e00152. [Google Scholar] [CrossRef]
- Finn, J.A.; Suter, M.; Haughey, E.; Hofer, D.; Lüscher, A. Greater Gains in Annual Yields from Increased Plant Diversity than Losses from Experimental Drought in Two Temperate Grasslands. Agriculture, Ecosystems & Environment 2018, 258, 149–153. [Google Scholar] [CrossRef]
- Mroncz, A.; Isselstein, J.; Komainda, M.; Leuschner, C. Microtopography Causes Small-Scale Variation in Harvest and Forage Quality of High-Yielding Silage Grassland in Northern Germany. Grassland Research 2024, 3, 33–42. [Google Scholar] [CrossRef]
- Stewart, A.V.; Goldson, S.L.; Marris, J.W.; Rubenstein, J.M.; Rolston, M.P. Analytical Purity of Old New Zealand Forage Seed Samples and Detection of Fungal and Insect Contaminants Including Ryegrass Endophyte and Argentine Stem Weevil. New Zealand Journal of Agricultural Research 2023, 66, 270–283. [Google Scholar] [CrossRef]
- Victoria Arellano, A.D.; Silva, G.M. da; Guatimosim, E.; Dorneles, K. da R.; Moreira, L.G.; Dallagnol, L.J. Las Semillas Recubiertas Con Trichoderma Atroviride y El Suelo Modificado Con Silicio Mejoran La Resistencia de Lolium Multiflorum Frente a Pyricularia Oryzae. Biological Control 2021, 154, 104499. [Google Scholar] [CrossRef]
- Ambus, J.V.; Awe, G.O.; Faccio Carvalho, P.C. de; Reichert, J.M. Integrated Crop-Livestock Systems in Lowlands with Rice Cultivation Improve Root Environment and Maintain Soil Structure and Functioning. Soil and Tillage Research 2023, 227, 105592. [Google Scholar] [CrossRef]
- Augustyniak, A.; Perlikowski, D.; Rapacz, M.; Kościelniak, J.; Kosmala, A. Conocimiento Del Proteoma Celular de Las Formas de Introgresión de Lolium Multiflorum/Festuca Arundinacea Para Descifrar Los Mecanismos Cruciales de La Aclimatación al Frío En Gramíneas Forrajeras. Plant Science 2018, 272, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Vallejos-Cacho, R.; Vallejos-Fernandez, L.A.; Alvarez-García, W.Y.; Tapia-Acosta, E.A.; Saldanha-Odriozola, S.; Quilcate-Pairazaman, C.E. Sustainability of Lolium Multiflorum L. “Ecotype Cajamarquino”, Associated with Trifolium Repens L., at Three Harvest Times in the Northern Highlands of Peru 2024.
- Cepeda Cepeda, E.B. Efecto del cultivo mixto de bacterias y levaduras sobre las características físicoquímicas y biomasa de Rye grass (Lolium multiflorum) en Andisoles, parroquia Columbe cantón Colta. 2022.
- Vanek, S.J.; Meza, K.; Ccanto, R.; Olivera, E.; Scurrah, M.; Fonte, S.J. Participatory Design of Improved Forage/Fallow Options across Soil Gradients with Farmers of the Central Peruvian Andes. Agriculture, Ecosystems & Environment 2020, 300, 106933. [Google Scholar] [CrossRef]
- Prerostova, S.; Zupkova, B.; Petrik, I.; Simura, J.; Nasinec, I.; Kopecky, D.; Knirsch, V.; Gaudinova, A.; Novak, O.; Vankova, R. Respuestas Hormonales Asociadas Con La Aclimatación al Estrés Por Congelación En Lolium Perenne. Environmental and Experimental Botany 2021, 182, 104295. [Google Scholar] [CrossRef]
- Perlikowski, D.; Augustyniak, A.; Masajada, K.; Skirycz, A.; Soja, A.M.; Michaelis, Ä.; Wolter, G.; Kosmala, A. Alteraciones Estructurales y Metabólicas En Sistemas Radiculares Bajo Condiciones Hídricas Limitadas En Gramíneas Forrajeras Del Complejo Lolium-Festuca. Plant Science 2019, 283, 211–223. [Google Scholar] [CrossRef]
- Wu, C.; Song, M.; Zhang, T.; Zhou, C.; Liu, W.; Jin, T.; Zhao, N. La Mutación En El Sitio Diana y El Citocromo P450 Confieren Resistencia a Múltiples Herbicidas En Raigrás Italiano (Lolium Multiflorum Lam.) de China. Crop Protection 2022, 161, 106068. [Google Scholar] [CrossRef]
- Kim, M.; Sung, K. Comparison of Causality of Temperature and Precipitation on Italian Ryegrass (Lolium Multiflorum Lam.) Yield between Cultivation Fields via Multi-Group Structural Equation Model Analysis in the Republic of Korea. Agriculture 2019, 9, 254. [Google Scholar] [CrossRef]
- Navarro-Zamora, L.A.; Villalobos-Villalobos, L.A. Composición morfológica del forraje ryegrass anual (Lolium multiflorum) cv. jumbo en respuesta a tres fórmulas nitrogenadas. Nutrición Animal Tropical 2021, 15, 99–122. [Google Scholar] [CrossRef]
- Maleki, K.; Soltani, E.; Seal, C.E.; Colville, L.; Pritchard, H.W.; Lamichhane, J.R. The Seed Germination Spectrum of 486 Plant Species: A Global Meta-Regression and Phylogenetic Pattern in Relation to Temperature and Water Potential. Agricultural and Forest Meteorology 2024, 346, 109865. [Google Scholar] [CrossRef]
- Schaeffer, A.H.; Silveira, D.C.; Schaeffer, O.A.; Lângaro, N.C.; Vargas, L. Seed Germination Behavior of Glyphosate-Resistant and Susceptible Italian Ryegrass (Lolium Multiflorum Lam.). Weed Biology and Management 2021, 21, 3–10. [Google Scholar] [CrossRef]
- Faisca, L.D.; Peres, M.T.P.; Fernandes, S.R.; Bonnet, O.J.F.; Batista, R.; Deiss, L.; Monteiro, A.L.G. A New Insight about the Selection and Intake of Forage by Ewes and Lambs in Different Production Systems on Pasture. Small Ruminant Research 2023, 221, 106949. [Google Scholar] [CrossRef]
- Yüce, İ.; Tatar, M.; Kökten, K.; Sarikaya, M.F.; Çi̇lesi̇z, Y.; Karaköy, T. Determination of Herbage Yield and Quality of Some Italian Ryegrass Varieties in Sivas Ecological Conditions. ISPEC Journal of Agricultural Sciences 2024, 8, 36–44. [Google Scholar] [CrossRef]
- Nunes, P.A. de A.; Bredemeier, C.; Bremm, C.; Caetano, L.A.M.; de Almeida, G.M.; de Souza Filho, W.; Anghinoni, I.; Carvalho, P.C. de F. Grazing Intensity Determines Pasture Spatial Heterogeneity and Productivity in an Integrated Crop-Livestock System. Grassland Science 2019, 65, 49–59. [Google Scholar] [CrossRef]
- Brooker, R.; Brown, L.K.; George, T.S.; Pakeman, R.J.; Palmer, S.; Ramsay, L.; Schöb, C.; Schurch, N.; Wilkinson, M.J. Active and Adaptive Plasticity in a Changing Climate. Trends in Plant Science 2022, 27, 717–728. [Google Scholar] [CrossRef]
- Abraha, A.B.; Truter, W.F.; Annandale, J.G.; Fessehazion, M.K. Forage Yield and Quality Response of Annual Ryegrass (Lolium Multiflorum) to Different Water and Nitrogen Levels. African Journal of Range & Forage Science 2015, 32, 125–131. [Google Scholar] [CrossRef]
- Bridges, K.M.; Fultz, L.M.; Alison, M.W.; Han, K.-J.; Macoon, B.; Pitman, W.D. Quantifying Soil Health in a Topographically Diverse Warm-Season Perennial Pasture over-Seeded with a Mix of Cool-Season Annuals. Agriculture, Ecosystems & Environment 2019, 282, 58–68. [Google Scholar] [CrossRef]
- Cui, E.; Cui, B.; Fan, X.; Li, S.; Gao, F. Los Cultivos Intercalados de Raigrás (Lolium Multiflorum L.) y Mostaza de La India (Brassica Juncea L.) Pueden Mejorar La Fitorremediación de Los Antibióticos y Los Genes de Resistencia a Los Antibióticos, Pero No de Los Metales Pesados. Science of The Total Environment 2021, 784, 147093. [Google Scholar] [CrossRef] [PubMed]
- Joniec, J.; Oleszczuk, P.; Jezierska-Tys, S.; Kwiatkowska, E. Effect of Reclamation Treatments on Microbial Activity and Phytotoxicity of Soil Degraded by the Sulphur Mining Industry. Environmental Pollution 2019, 252, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.; Wang, J.; Condron, L.M.; Guo, B.; Liu, J.; Qiu, G.; Li, H. Impact of Ryegrass Cover Crop Inclusion on Soil Phosphorus and pqqC- and phoD-Harboring Bacterial Communities. Soil and Tillage Research 2023, 234, 105823. [Google Scholar] [CrossRef]
- Han, K. j.; Smith, D.J.; Pitman, W. d. Potential of Cool-Season Species as Cover Crops and Forage in the Southeastern United States. Crop, Forage & Turfgrass Management 2018, 4, 170038. [Google Scholar] [CrossRef]
- Pandey, V.C.; Gajić, G.; Sharma, P.; Roy, M. 4 - Soil and Phytomanagement for Adaptive Phytoremediation Practices. In Adaptive Phytoremediation Practices; Pandey, V.C., Gajić, G., Sharma, P., Roy, M., Eds.; Elsevier, 2022; pp. 135–179 ISBN 978-0-12-823831-8.
- Shi, G.; Hu, J.; Zhang, S.; Ni, G.; Shi, W.; Hu, C.; Zhao, X. La Aplicación de Se Exógeno Mejoró La Eficiencia de Remediación de Lolium Multiflorum Lam Cultivado En Suelos Cocontaminados Con Nonilfenol-Cd. Journal of Environmental Chemical Engineering 2022, 10, 108962. [Google Scholar] [CrossRef]
- Gao, R.; Duan, Y.; Zhang, J.; Ren, Y.; Li, H.; Liu, X.; Zhao, P.; Jing, Y. Effects of Long-Term Application of Organic Manure and Chemical Fertilizer on Soil Properties and Microbial Communities in the Agro-Pastoral Ecotone of North China. Front. Environ. Sci. 2022, 10. [Google Scholar] [CrossRef]
- Liu, M.; Che, Y.; Wang, L.; Zhao, Z.; Zhang, Y.; Wei, L.; Xiao, Y. Rice Straw Biochar and Phosphorus Inputs Have More Positive Effects on the Yield and Nutrient Uptake of Lolium Multiflorum than Arbuscular Mycorrhizal Fungi in Acidic Cd-Contaminated Soils. Chemosphere 2019, 235, 32–39. [Google Scholar] [CrossRef]
- Oliva, M.; Valqui Valqui, L.; Meléndez Mori, J.; Milla, M.; Leiva, S.; Collazos, R.; Maicelo, J. Influence of Arboreal Native Species on Silvopastoral Systems on the Yield and Nutritional Value of Lolium Multiflorum and Trifolium Repens. Scientia Agropecuaria 2018, 9, 579–583. [Google Scholar] [CrossRef]
- Paye, W.S.; Szogi, A.A.; Shumaker, P.D.; Billman, E.D. Annual Ryegrass (Lolium Multiflorum Lam.) Growth Response to Nitrogen in a Sandy Soil Amended with Acidified Manure and Municipal Sludge after “Quick Wash” Treatment. Agronomy 2023, 13, 2655. [Google Scholar] [CrossRef]
- Hu, J.; Hassi, U.; Gebremikael, M.T.; Dumack, K.; De Swaef, T.; Wesemael, W.; Sleutel, S.; De Neve, S. Root Traits Explain Multitrophic Interactions of Belowground Microfauna on Soil Nitrogen Mineralization and Plant Productivity. Soil Biology and Biochemistry 2023, 184, 109093. [Google Scholar] [CrossRef]
- Montenegro, L.F.; Descalzo, Adriana. M.; Rizzo, S.; Rossetti, L.; García, P.T.; Pérez, C.D. Mejora Del Estado Antioxidante, Las Vitaminas Liposolubles, La Composición de Ácidos Grasos y La Estabilidad Lipídica En La Carne de Carpa Herbívora (Ctenopharyngodon Idella Val) Alimentada Con Raigrás Fresco (Lolium Multiflorum Lam). Aquaculture 2022, 553, 738067. [Google Scholar] [CrossRef]
- Fernández-Habas, J.; Real, D.; Vanwalleghem, T.; Fernández-Rebollo, P. LANZA® Tedera Is Strongly Suppressed by Competition from Lolium Multiflorum and Is Best Adapted to Light-Textured Soils. Agronomy 2023, 13, 965. [Google Scholar] [CrossRef]
- Rupngam, T.; Messiga, A.J.; Karam, A. Phosphorus Mobility in Heavily Manured and Waterlogged Soil Cultivated with Ryegrass (Lolium Multiflorum). Agronomy 2023, 13, 2168. [Google Scholar] [CrossRef]
- Brede, A.D.; Brede, J.L. Establishment Clipping of Tall Fescue and Companion Annual Ryegrass. Agronomy Journal 1988, 80, 27–30. [Google Scholar] [CrossRef]
- Ollerenshaw, J.H.; Incoll, L.D. Leaf Photosynthesis in Pure Swards of Two Grasses (Lolium Perenne and Lolium Multiflorum) Subjected to Contrasting Intensities of Defoliation. Annals of Applied Biology 1979, 92, 133–142. [Google Scholar] [CrossRef]
- Mavlyutov, Y.; Kostenko, S.; Shamustakimova, A.; Klimenko, I. Análisis de Variabilidad Genética de Cultivares Rusos de Raigrás (Lolium) a Partir de Marcadores SCoT. Journal of Genetic Engineering and Biotechnology 2022, 20, 163. [Google Scholar] [CrossRef]
- Beckie, H.J.; Jasieniuk, M. Chapter 12 - Lolium Rigidum and Lolium Multiflorum. In Biology and Management of Problematic Crop Weed Species; Chauhan, B.S., Ed.; Academic Press, 2021; pp. 261–283 ISBN 978-0-12-822917-0.
- SENAMHI - Pronostico Meteorologico. Available online: https://www.senamhi.gob.pe/?p=pronostico-meteorologico (accessed on 6 June 2024).
- UPOV Unión Internacional Para La Protección de Las Obtenciones Vegetales (UPOV). Available online: https://www.upov.int/portal/index.html.es (accessed on 27 June 2024).
Figure 1.
Interaction between cutting time and (A) annual biomass (Mg. ha. year), (B) protein percentage (%), (C) protein yield per cut (Mg. ha. cut), and (D) annual protein yield (Mg. ha. year) for Lolium multiflorum L. genotypes, LM - 58 and LM - 43, respectively.
Figure 1.
Interaction between cutting time and (A) annual biomass (Mg. ha. year), (B) protein percentage (%), (C) protein yield per cut (Mg. ha. cut), and (D) annual protein yield (Mg. ha. year) for Lolium multiflorum L. genotypes, LM - 58 and LM - 43, respectively.
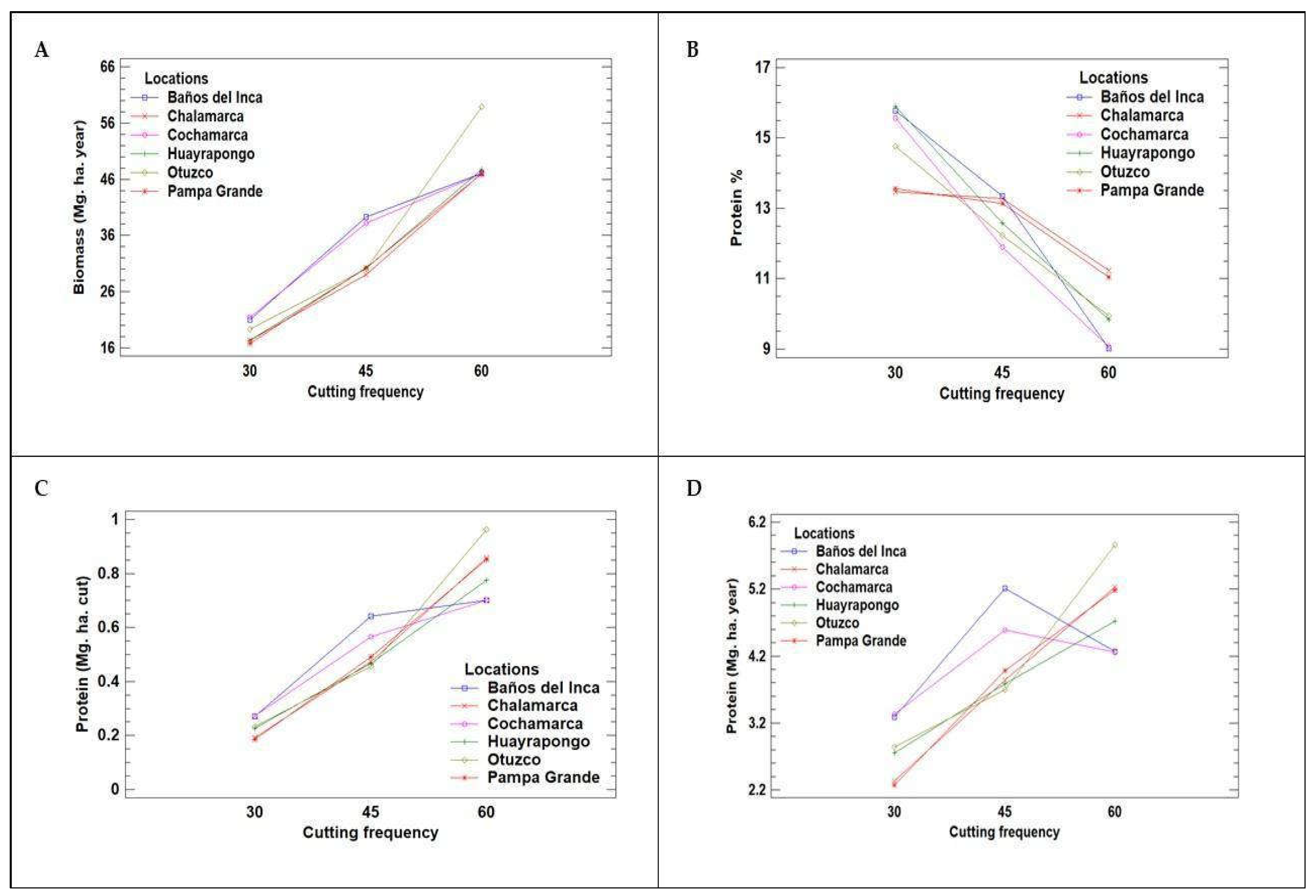

Table 1.
Yield performance of the two lolium lines at the six locations.
| Factors | Green forage (Mg. ha) | Biomass (Mg. ha) | Seeds (kg. ha) | Protein % |
|---|---|---|---|---|
| Genotype | ||||
| LM-43 | 20.15 b | 3.64 b | 189.95 b | 11.95 b |
| LM-58 | 20.97 a | 4.49 a | 259.23 a | 13.48 a |
| Standar error | 0.21 | 0.06 | 13.56 | 0.29 |
| p-value | 0.0104 | <0.001 | 0.0047 | 0.0037 |
| Locations | ||||
| Baños del Inca | 22.19 a | 4.3 a | 251.98 | 13.31 |
| Chalamarca | 18.83 c | 3.95 b | 218.24 | 12.50 |
| Cochamarca | 21.26 ab | 3.77 b | 207.95 | 13.04 |
| Huayrapongo | 22.10 a | 4.35 a | 217.40 | 12.18 |
| Otuzco | 20.71 b | 4.06 ab | 254.75 | 13.34 |
| Pampa Grande | 18.26 c | 3.95 b | 197.20 | 11.94 |
| Standar error | 0.36 | 0.11 | 23.49 | 0.48 |
| p-value | <0.001 | 0.0061 | 0.8743 | 0.7580 |
*a,b in the columns show differences (Duncan test, p<0.05).
Table 2.
Biomass and protein by cutting times.
| Factors | Biomass | Protein | |||
|---|---|---|---|---|---|
| Mg. ha. cut | Mg. ha. year | % | Mg. ha. cut | Mg. ha. year | |
| Cutting time | |||||
| 30 days | 1.55 c | 18.87 c | 14.84 a | 0.23 c | 2.80 b |
| 45 days | 4.05 b | 32.83 b | 12.74 b | 0.52 b | 4.19 a |
| 60 days | 8.07 a | 49.10 a | 10.03 c | 0.81 a | 4.91 a |
| SE | 0.24 | 1.60 | 0.44 | 0.036 | 0.26 |
| p-value | |||||
| Cuts (C) | 0.0000 | 0.0000 | 0.0001 | 0.000 | 0.0006 |
| Locations (L) | 0.4008 | 0.5376 | 0.9755 | 0.9599 | 0.8896 |
| C x L | 0.6303 | 0.6192 | 0.0036 | 0.2076 | 0.1817 |
SE: Standar error. *a,b in the columns show differences (Duncan test, p<0.05).
Table 3.
Morphological differentiation of Lolium multiflorum L., Rye Grass LM - 58 and LM – 43.



Table 4.
Qualitative morphological characterization of the assessed qualitative characteristics - LM 58.
Table 4.
Qualitative morphological characterization of the assessed qualitative characteristics - LM 58.
| Character | Observation period | States | Score |
|---|---|---|---|
| Growth habit | Plant | Semi-erect | 3 |
| Leaf length in vegetative state (33 to 38) | Leaflet | Long | 7 |
| Leaf width in vegetative state (0.8-1.0) | Limbo | Media | 5 |
| The intensity of the green colour | Leaflet | Dark | 7 |
| Width after vernalization (basal coverage) | Plant | Medium | 5 |
| Height (after vernalization) | Plant | High (60 - 70) | 7 |
| Tendency to form inflorescences (without vernalization) | Plant | Medium | 5 |
| Flag leaf length | Leaflet | Medium | 5 |
| Width | Leaflet | Wide (0.6 to 0.8) | 7 |
| Length/width ratio | Leaflet | Media | 5 |
| Length of most extended stem, including inflorescences (when fully developed) | Stem | Long (45 to 55) | 7 |
| Length including inflorescences (when fully developed) | Panicle | Long (25 to 30) | 7 |
| Number of spikelets | Panicle | Very high (>11) | 9 |
| Number of spikelets density | Panicle | Medium | 5 |
| External length of basal spikelet | Glumes | Long (0.8 to 1.0) | 7 |
| Length of basal spikelet, excluding the ridge | Glumes | Long (1.5 to 2.0) | 7 |
| Growth rate (cm/day) | Plant | High (1.4 to 1.8) | 3 |
| Form | Leaflet | Lanceolate medium | 3 |
| Length at 30 days | Plant | Long (33 to 39) | 7 |
| Presence of atria | Leaflet | Media | 5 |
| Expression of the ligule | Limbo | Regularly noted | 7 |
| Age at fourth leaf (cm) | Leaflet | High (35 to 40) | 5 |
| Number of seeds per spikelet | Grain | High (12 to 14) | 7 |
| Short slogan | Grain | Dark cream | 5 |
| Length of the locker | Grain | Cut | |
| Time of beginning of flowering (green fodder) | Flowering | High (60 to 70 days) | 5 |
| Growing season (grain production) | Harvest | High (100 to 110 days) | 7 |
| Presence of edges | Grain | Present | 3 |
Table 5.
Evaluation of homogeneity, lemma colour and grain ridge presence of Ryegrass LM - 58.
| Campaign | Characters | |||||
|---|---|---|---|---|---|---|
| Colour of the slogan | Grain presence of edge | |||||
| Level of expression | Off-type plants | Maximum number | Level of expression | Off-type plants | Maximum number | |
| First | Dark cream | 0 | 3 | Present | 0 | 3 |
| Second | Dark cream | 0 | 3 | Present | 0 | 3 |
* Maximum number of non-typical plants (UPOV).
Table 6.
Evaluation of uniformity, vegetative cycle and time of beginning of flowering (days).
| Campaign | Characters | |||||||
|---|---|---|---|---|---|---|---|---|
| Vegetative cycle | Start of flowering | |||||||
| Level of expression |
Average (days) |
DS. | CV. | Level of expression |
Average (days) |
DS. | CV. | |
| First | Semi-early | 105.0 | 2.94 | 2.80% | Precoz | 40.2 | 1.17 | 2.91% |
| Second | Semi-early | 105.8 | 2.87 | 2.73% | Precoz | 39.7 | 1.03 | 2.59% |
Table 7.
Geographical location and soil chemical composition of the locations for the yield evaluation of the genotypes LM - 58 and LM - 43 of Lolium multiflorum L.
Table 7.
Geographical location and soil chemical composition of the locations for the yield evaluation of the genotypes LM - 58 and LM - 43 of Lolium multiflorum L.
| Locations |
Agricultural campaign |
Altitude | District | Geographical coordinates | Chemical composition of the soil | |||||
|---|---|---|---|---|---|---|---|---|---|---|
|
P (ppm) |
K (ppm) |
pH |
OM (%) |
Al (mEq/100g) |
||||||
| Otuzco | 2018-2019 | 2680 | Baños del Inca | -7.131402 | -78.469430 | 16.22 | 225 | 4.5 | 1.54 | 0.10 |
| Cochamarca | 2016-2017 | 2820 | Gregorio Pita | -7.181967 | -78.467969 | 8.11 | 195 | 4.3 | 2.13 | 0.90 |
| Huayrapongo | 2015-2016 | 2650 | Baños del Inca | -7.181967 | -78.467970 | 33.70 | 271 | 6.1 | 3.82 | 0.00 |
| Pampa Grande | 2014-2015 | 2680 | Cajabamba | -7.611059 | -78.070150 | 6.20 | 170 | 3.8 | 3.55 | 0.50 |
| Chalamarca | 2014-2015 | 2760 | Chota | -7.279996 | -78.218141 | 18.22 | 186 | 4.9 | 4.12 | 0.70 |
| Baños del Inca | 2014-2015 | 2650 | Cajamarca | -7.159426 | -78.461260 | 10.90 | 330 | 7.0 | 2.25 | 0.00 |
* P=phosphorus; K=potassium; OM=organic matter; Al=aluminum; pH=hydrogen ion potential.
Table 8.
Timeline of key activities in the development of elite Lolium multiflorum L. genotypes.
| Period (years) | Activity | Description |
|---|---|---|
| 1995 | Collection and installation of experimental material. | Sowing (germination beds). |
| 1996 – 1998 | Characterisationof genotypes | Selection and sowing of 680 seedlings (genotypes). |
| 1999 – 2000 | Selection multiplication | Characterisation of genotypes (growth habit, anthesis and pests and diseases). |
| 2001 – 2003 | Multiplication selection | Characterisation and selection of the best genotypes. |
| 2004 – 2006 | Research | Selection and evaluation of elite genotypes in Forage green, Dry matter, and seed. |
| 2006 – 2008 | Research | Polycrosses between the 10 elite genotypes, in experimental field. To fix superior characters. |
| 2009 - 2011 | Field testing by growers | Selection and evaluation of 10 superior genotypes in forage production, nutritive value and tolerance to pests and diseases. |
| 2011 – 2016 | Adaptation and efficiency trials in growers' fields. | First campaign - Producers' field adaptation and efficiency trials. |
| 2016 – 2019 | Adaptation and efficiency trials in growers' fields. | Second campaign - Adaptation and efficiency trials in growers' fields. |
Table 9.
Qualitative morphological evaluation characteristics of Lolium multiflorum L. genotypes LM - 58 and LM – 43.
Table 9.
Qualitative morphological evaluation characteristics of Lolium multiflorum L. genotypes LM - 58 and LM – 43.
| Parts | Features |
|---|---|
| Plant | Vegetative growth habit (without vernalization); Tendency to form inflorescences (without vernalization); Width after vernalization (basal area in cm) Height after vernalization Length of most extended stem, including inflorescences (when fully developed) |
| Leaflet | Leaf length (in a vegetative state); Leaf width (in a vegetative state); The intensity of the green colour Flag leaf length (last leaf); Width (previous sheet); Length/width ratio (previous leaf); Leaf shape; Presence of atria; Expression of the ligule; Motto colour. |
| Inflorescence | Length, including inflorescences (when fully developed); Number of spikelets; Number of spikelets density; Length of outer glume of basal spikelet; Length of basal spikelet, excluding the ridge; Number of seeds per spikelet; Grain presence of edge; Time of beginning of flowering. |
| Other | Growth rate per day; Plant length at 30 days; Growing season for grain-seed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



