
Submitted:
18 July 2024
Posted:
19 July 2024
You are already at the latest version
Abstract
Vitamin D receptor (VDR) is an important candidate gene in musculoskeletal phenotypes. Polymorphisms in VDR have been previously associated with several pathologies and muscular strength in athletes and elderly people, however literature reported contradictory results. The object of this research is to verify the association between the most studied VDR variants (rs2228570, rs7975232, and rs1544410) and the increase of muscle mass in elite young soccer players. A sample of 55 soccer players (15-18 years old) form a professional team was selected for this study. DNA was extracted by salting out method and polymorphisms were genotyped by PCR-RFLP, followed by 2% agarose gel electrophoresis. To test the effect of the three SNPs, logistic regression analysis was applied. Body composition was carried out through skinfold thickness method, and muscular area of arm and lower limb were calculated using Frisancho formulas. All three polymorphisms met Hardy-Weinberg equilibrium (p > 0.05) and their frequencies fell within the worldwide variability. A significant correlation between rs1544410 and increase of calf muscle mass was observed. Individuals carrying A allele showed higher calf muscular mass than those carrying the G allele (p = 0.034). Moreover, haplotype analysis applied to the two SNPs in linkage disequilibrium (rs7975232 and rs1544410), showed that AG haplotype resulted negatively correlated to calf muscle area. In conclusion, we confirm an association between VDR polymorphisms and muscular mass that could encourage the genetic screening of VDR gene to identify a potential risk of injury and for individual nutritional interventions.
Keywords:
vitamin D receptor
; SNPs
; genotype
; muscle mass
1. Introduction
Vitamin D, also called calciferol, is a fat-soluble molecule belonging to the family of steroid hormones and plays an important role in the regulation of calcium metabolism, bone [1], and muscle development. Vitamin D is involved in the mechanism of muscle contraction [2] and a strong relation between Vitamin D level and sarcopenia has been demonstrated [3]. Moreover, it has been shown that sufficient levels of vitamin D enable the maintenance and development of athletic performance; consequently, reduced levels of the vitamin inhibit muscle relaxation required after training and increase muscle pain. This leads to a major risk of injury and stress fractures, causing loss of muscle power and reducing bone mineral density [4].
Vitamin D can either be assumed with the diet or synthesized by the skin: UVB rays, by penetrating the epidermis, convert 7-dehydrocholesterol (7DHC) into vitamin D3 (cholecalciferol). Two subsequent hydroxylation steps occurring, respectively, in the liver and in the kidney convert vitamin D3 into 1α-25-dihydroxyvitamin D (1,25(OH)2D or calcitriol), which is the biologically active molecule. Calcitriol enters the cell where it binds to the cytoplasmic vitamin D receptor (VDR), a protein constituted by 427 amino acids in humans, and then the hormone-receptor complex migrates to the nucleus, where it binds to the retinoid X receptor α (RXR α); finally, this trimeric complex binds to DNA, affecting the expression of several vitamin D target genes, orchestrating various biological processes [5].
VDR gene is located on human chromosome 12q13.11 and comprises 11 exons, encompassing about 75 kb. Exons 2 and 3 are involved in DNA binding, whereas exons 4 and 9 are involved in ligand binding [6].
It has been suggested that different polymorphisms in VDR gene can determine different response to strength training [7] and muscle injury [8], but results are still controversial [9]. In addition, VDR overexpression has been linked to the hypertrophic response to resistance exercise in humans [10].
Among the more than one hundred SNPs (Single Nucleotide Polymorphisms) present in the VDR gene, the most common and studied are rs228570 (FokI) located in exon 2, rs1544410 (BsmI) and rs7975232 (ApaI) both located in intron 8.
The FokI polymorphism is located at the 5′ end of exon 2, corresponding to the start codon. This mutation involves the replacement of a guanine with an adenine, guanine being the ancestral allele. The A allele was found to be correlated to the maintenance of bone homeostasis [11,12,13]. The same allele has been significantly associated with total Fat Free Mass (FFM), appendicular FFM, and relative appendicular FFM, demonstrating a correlation with sarcopenia [14]; accordingly, males carrying GG genotype had significantly less FFM, appendicular fat-free mass and skeletal muscle index (SMI) compared to A allele carriers, and two-fold higher risk of being sarcopenic when compared to carriers of the A allele.
Other studies demonstrated an association between FokI polymorphism and muscle phenotypes with male GG homozygotes showing significantly lower values of knee extensor (KE) [15] and handgrip (HG) strength [16] than A allele carriers. On the contrary, a study conducted by [17] found AA individuals displaying significantly greater HG strength than G carriers.
Finally, a study conducted on soccer players [18] showed that analysis of the FokI polymorphism could help select young athletes who possess the most favorable genetic potential to succeed in soccer. Indeed, it was observed that the homozygous AA genotype of the FokI polymorphism was significantly more represented in young soccer players than in the control group.
BsmI, located at the 3′ end of the VDR gene, shows a change from A to G, with the G allele representing the ancestral variant. Functional implications of this SNP include the possibility of altering splice sites involved in mRNA transcription or affecting intronic regulatory elements of the VDR gene.
Its relationship with risk of osteoporosis has been widely debated with conflicting results: a metanalysis conducted on 26 studies [19] reported the possible association between BsmI polymorphism and the risk of bone fracture, suggesting a protective role for GG for osteoporosis risk. A different metanalysis conducted on 41 studies [20] did not confirm such an association.
Studies on the association between BsmI polymorphism and muscle mass and strength also highlighted contrasting results. While several research denied any association between rs1544410 polymorphism and muscle mass phenotype [17], a work displayed higher quadriceps and grip strength for elderly with G allele [16]. In contrast, an association between AA genotype and higher hamstring strength was demonstrated in a sample of elderly women [21,22], suggesting a higher knee extensor strength for AA genotypes when compared with GG and AG genotypes.
ApaI, also located at the 3′ end, shows a change from A to C, with the C allele representing the ancestral variant. The VDR ApaI is associated with increased bone mass concomitant with higher calcium intake [23], and it seems to influence the severity of musculoskeletal injuries in professional football players [8]. A recent meta-analysis [24] highlighted an increased risk for osteoporotic fracture for AA genotype in the European population but not in the overall sample that included individual from the Americas (northern, central and southern).
Several studies are based on the analysis of the association between VDR polymorphisms and bone mineral density (BMD). Limited research focused on the association with muscular mass and strength and has led to contrasting results. A work by Iki [25] negated any difference in developing muscular strength for the different genotypes. On the contrary, an association between genotypes and hand grip strength has been suggested in research on elderly from Taiwan [26]. Moreover, a study by Wang [27] found lower knee and elbow concentric or eccentric peak torque among individuals carrying the ApaI A allele.
The aim of the present research is to verify the association between the most studied VDR variants (rs2228570, rs7975232, and rs1544410) and the increase of lean and muscle mass in young soccer players.
2. Materials and Methods
2.1. Sampling
55 male elite soccer players, 15 to 18 years old (mean: 16.65 ± 1.55 years), were selected for this study. All players included in the study were from the same professional team, participated in the Official National Football Championship (Primavera, Allievi, and Giovanissimi), and self-reported to be Italians for at least 3 generations. All study participants read and signed an informed consent, which was signed by the parents of underage athletes. This study was approved by the Ethics Committee of Azienda Ospedaliera Universitaria (AOU) of Cagliari University (Italy).
2.2. DNA Analyses
A buccal swab was taken by using a sterile cytobrush from each participant and DNA was extracted using the salting-out method.
Concentration and quality of the extracted DNA were measured using the NanoDrop spectrophotometer (Thermo Fisher Scientific).
DNA amplification was performed by classical PCR method using the following primers for the three polymorphisms:
ApaI forward: 5′-CAGAGCATGGACAGGGAGCAAG-3′; reverse: 5′- CAACTCCTCATGGCTGAGGTCT-3′;
BsmI forward 5′-CAACAAGACTACAAGTACCGCGTCAGTGA-3′; reverse 5′- AACCAGCGGAAGAGGTCAAGGG-3′;
FokI forward 5′-GATGCCAGCTGGCCCTGGCACTG-3′; reverse 5′-ATGGAAACACCTTGCTTCTTCTCCCTC-3′.
Each reaction mix (total 25 µl) for all three polymorphisms included 2µl of DNA, 0.2µl of the forward and reverse primers, 10.6µl of water, and 12µl of NZYSupreme qPCR Green Master Mix 2x (Nzytech, Portugal).
Enzymatic digestion was performed with restriction enzymes ApaI, BsmI, and FokI for allele recognition.
Genotyping was performed by electrophoresis technique, with a 2% agarose gel for ApaI and BsmI and a 10% polyacrylamide gel for FokI, observing the following DNA fragments: for ApaI (A:740 bp, C:515+225 bp); for BsmI (A: 825 bp, G:650+175 bp); for FokI (G:272 bp, A:198+74 bp).
2.3. Body Composition Determination
For each subject height and weight were measured according to standard procedure [28].
Body composition parameters were calculated using the skinfold equations. Two different formulas were used to calculate the percentage of Fat Mass (FM) and Fat Free Mass (FFM): Slaughter’s formula [29], designed specifically for boys (up to age 18) using measurements of the tricipital skinfold and subscapular skinfold; and Reilly’s formula [30], specifically validated for young soccer players (age 16.67 ± 0.5 years) by [31], that uses measurements of the thigh, abdominal, triceps, and calf skinfold.
The muscle area of the arm (AMA), calf (CMA), and thigh (TMA) were calculated according to Frisancho formula [32]. For AMA correspondence to proper percentile for each participant was also verified.
2.4. Statistical Analyses
Allele and genotype frequencies, as well as Hardy-Weinberg were calculated with Genepop software (ver.4.4.3).
One-way ANOVA was used to evaluate the hypothesis of equality of means among genotypes in relation to all body composition measures. Analyses were performed with the R 4.2.3 program.
Genotypic association analysis with fat-free mass and muscle mass was performed with software SNPstats (https://snpstats.net/)
Given the variability of the three polymorphisms across 1000 genomes, the population branch statistic (PBS), a summary statistic that exploits pairwise FST values among three populations, was calculated to check for evidence of natural selection within the VDR gene [33]. A population’s PBS value represents the amount of allele frequency change at a given locus in the history of this population.
The VCF (Variant Call Format) file of chromosome 12 was downloaded by the 1,000 Genomes Database Phase 3. From the VCF file of the whole gene, only SNPs with Minimum Allele Frequency (MAF) ≥ 0.05 were selected, using Plink 1.9 [34]; next, FST index was calculated among three potential population pairs: YRI, CEU, and CHB, using the program VCFtools [35]; finally, PBS was calculated and the two-dimensional graph created using R Studio [36].In order to obtain further information regarding the possible natural selection of the VDR gene, Tajima’s D was calculated using VCFtools.
3. Results
Allele and genotype frequencies are shown in Table 1. Hardy-Weinberg test revealed that all samples were in equilibrium for all loci (p > 0.05). The three SNPs showed a high worldwide genetic variability (1,000 Genomes dataset), and allele frequencies of our sample fell within the worldwide range; ApaI and BsmI frequencies were also included within the European range (Table 1).
Considering the high variability of allele frequency and the correlation of the gene with several diseases, we examined, through PBS, possible traces of natural selection using data from 1000 genomes. Some SNPs had PBS values near or equal to the limits of the distribution, but not the specific polymorphisms we focus on. They fell on both lines and had consistently low PBS values, suggesting the absence of selection (Figure 1).
The absence of selection was confirmed by D’ Tajima test: positive values were obtained for the three SNPs (2.486 for ApaI, 1.957 for BsmI, and 2.113 for FokI), indicating low levels of low- and high-frequency polymorphisms.
One-way ANOVA test was used to assess any difference in fat-free mass between the different genotypes. The data showed no significant difference between genotypes for any considered variables (p > 0.05, Table 2). It is noteworthy a weak trend for increasing CMA values from CC to AA for Apa I (88.09, 94.27, 95.72 for CC, AC, and AA, respectively).
The logistic regression analyses revealed a significant association between BsmI polymorphism and CMA under the dominant model: athletes carrying the A allele developed higher calf muscle mass than GG homozygous (Table 3). Moreover, although not statistically significant, we found higher values of CMA and AMA in individuals with FokI AA genotype under the recessive model (97.32 vs 93.49 and 54.22 vs 49.71 respectively).
To test the combined effects of rs7975232 and rs1544410 SNPs, we apply logistic regression analysis to the haplotypes. The linkage disequilibrium of these variants for European population was previously verified through the LDlink program [37], that attested a strong linkage between ApaI and BsmI. This result was confirmed in our sample applying D statistics through SNPstats program (https://www.snpstats.net/start.htm).
A significant association has been found between CMA and haplotype AG (Table 4): the negative value of difference (95% CI) suggests significant low muscle mass for individuals carrying AG haplotype.
In addition, despite not reaching the level of significance, haplotype AG showed lower muscle mass also for arm and thigh area (data not shown).
4. Discussion
Vitamin D has been proven to influence muscle activity and performance [38]. Literature shows some evidence on the association between Vitamin D receptor polymorphisms and muscle strength, but the issue is still under debate [5]. Most of the literature deals with the relationship between VDR polymorphisms and decrease muscle mass with aging or the analysis of muscular trait in young athletes [17].
In this study, we analyzed the association between VDR polymorphisms and muscle mass, in a sample of elite young soccer players from a professional Italian team, to verify if the increase of muscular mass can be influenced by VDR variants.
PBS analysis showed no evidence of selection for the three polymorphisms: their high level of variability is likely due to random evolutive factors, such as genetic drift.
Previous research has focused on the correlation between VDR polymorphisms, muscle mass and sarcopenia in elderly [14,39] and found that individuals with FokI GG genotype were more likely to have lower muscle mass and to develop sarcopenia than AA and AG genotypes. Our sampling includes only young athletes and therefore we could not detect correlation with sarcopenia; however, our data are in good agreement with previous reports, suggesting that FokI rs 2228570 can lead to different response to training by increasing muscle size and strength [20] even in young soccer players. Indeed, logistic regression highlighted higher values of CMA in individuals with AA genotype, suggesting that the AA genotype is a favorable factor to develop muscle mass and protect from sarcopenia [14,18], confirming the role of FokI rs2228570 in developing muscular mass. The molecular and physiological basis for this association is still uncertain, but previous studies suggested that the G allele produces an isoform with a decreased ability to induce transcription compared to the isoform coded by the A allele variant [40], and, consequently, a reduced muscle mass.
Moreover, our study suggested a significant correlation between BsmI polymorphism rs1544410 and the development of calf muscle mass. The dominant model shows a statistically significant increase of CMA among individuals carrying A allele. This result partially confirms previous studies, that demonstrated higher values of hamstring strength in individuals with AA genotype, although the authors did not find any correlation with muscle mass [21,22].
Finally, ApaI rs7975232 did not show any statistical association with muscle mass, even though a weak trend versus a higher increase of CMA was evident in CC carriers respect to AA carriers. To the best of our knowledge only one study found similar results: a research carried out on elderly from Taiwan showed an association between hand grip strength and homozygous CC females [26].
The role of rs7975232 ApaI and rs1544410 BsmI SNPs in VDR activity is not fully understood, but it has been hypothesized that, despite being located in a not translated region of the gene, these two SNPs could have a role in mRNA stability, due to their proximity to the poly-A tail [41].
To verify the combined effect of the two SNPs in linkage disequilibrium on muscle mass, we analyzed the correlation between haplotype and muscle mass, and, to our knowledge, this is the first time that this approach was used. Haplotype AG appeared negatively correlated with muscular mass of arm, calf, and thigh, reporting a significant value in the correlation with CMA. This datum is particularly useful for its possible application in the field of sports science, since individuals carrying the AG haplotype, given the same amount of training, seem to develop less muscle mass. It would be important to be able to confirm whether this datum also determines earlier sarcopenia in the elderly age.
5. Conclusions
This study supports the importance of studying the simultaneous effect of different allelic variants to better define the relationship between genotype and phenotype.
Our data confirm the role of FokI rs2228570 in developing muscular mass: AA homozygotes are more likely to develop muscle mass and therefore to be protected from sarcopenia. In addition, the association found between BsmI rs1544410 and AG haplotype (ApaI rs7975232 and BsmI rs1544410) with increase of calf muscle mass suggests the potential use of genetic screening for the VDR gene in athletes or in elderly individuals, to identify a potential risk of injury and/or for individual nutritional interventions, such as vitamin D supplementation, as suggested by previous papers [38,42].
Although the sample size is not large, we believe that the highly homogeneous population tested may be considered adequate for a pilot study. However, analysis of a much larger sample size is required to validate our conclusions.
Author Contributions
Conceptualization, C.M.C. and L.F.; Writing—original draft preparation, R.R., L.F. and M.M.; Writing—review and editing, P.F. and R.R. Methodology: F.T. and M.M. data Collection: M.S. and L.F. formal analysis, L.F. and L.D.; supervision, R.R. and C.M.C. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors wish to thank the Cagliari Calcio’s society and the players who participated in this study. This work was financially supported by F.I.R. (Fondo Integrativo per la Ricerca) from the University of Cagliari to CMC and RR.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Saponaro, F.; Saba, A.; Zucchi, R. An Update on Vitamin D Metabolism. Int. J. Mol. Sci. 2020, 21, 6573. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, M.; Pacini, S. Chronic kidney disease and vitamin D: how much is adequate? Kidney Int. 2009, 76, 931–933. [Google Scholar] [CrossRef]
- Kuwabara, A.; Tsugawa, N.; Kondo, H.; Ao, M.; Fujiwara, H.; Hosokawa, N.; Matsumoto, S.; Tanaka, K.; Nakano, T. Associations between serum 25-hydroxyvitamin D 3 level and skeletal muscle mass and lower limb muscle strength in Japanese middle-aged subjects. Osteoporos. Sarcopenia 2017, 3, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.; Begerow, B.; Minne, H.W. Vitamin D and Muscle Function. Osteoporos Int. 2002, 13, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Bollen, S.E.; Bass, J.J.; Wilkinson, D.J.; Hewison, M.; Atherton, P.J. The impact of genetic variation within the vitamin D pathway upon skeletal muscle function: A systematic review. J. Steroid Biochem. Mol. Biol. 2023, 229, 106266. [Google Scholar] [CrossRef]
- Bulgay, C.; Bayraktar, I.; Kazan, H.H.; Yıldırım, D.S.; Zorba, E.; Akman, O.; Ergun, M.A.; Cerit, M.; Ulucan, K.; Eken, Ö.; Ceylan, H.İ.; Badicu, G.; Grosz, W.R.; Mijaică, R. Evaluation of the Association of VDR rs2228570 Polymorphism with Elite Track and Field Athletes’ Competitive Performance. Healthcare 2023, 11, 681. [Google Scholar] [CrossRef] [PubMed]
- Rabon-Stith, K.M.; Hagberg, J.M.; Phares, D.A.; Kostek, M.C.; Delmonico, M.J.; Roth, S.M.; Ferrell, R.E.; Conway, J.M.; Ryan, A.S.; Hurley, B.F. Vitamin D receptor FokI genotype influences bone mineral density response to strength training, but not aerobic training. Exp. Physiol. 2005, 90, 653–661. [Google Scholar] [CrossRef]
- Massidda, M.; Corrias, L.; Bachis, V.; Cugia, P.; Piras, F.; Scorcu, M.; Calò, C.M. Vitamin D receptor gene polymorphisms and musculoskeletal injuries in professional football players. Exp. Therap. Med. 2015, 9, 1974–1978. [Google Scholar] [CrossRef]
- Krasniqi, E.; Boshnjaku, A.; Wagner, K.-H.; Wessner, B. Association between Polymorphisms in Vitamin D Pathway-Related Genes, Vitamin D Status, Muscle Mass and Function: A Systematic Review. Nutrients 2021, 13, 3109. [Google Scholar] [CrossRef]
- Bass, J.J.; Nakhuda, A.; Deane, C.S.; Brook, M.S.; Wilkinson, D.J.; Phillips, B.E.; Philp, A.; Tarum, J.; Kadi, F.; Andersen, D.; Garcia, A.M.; Smith, K.; Gallagher, I. J.; Szewczyk, N.J.; Cleasby, M.E.; Atherton, P.J. Overexpression of the vitamin D receptor (VDR) induces skeletal muscle hypertrophy. Mol. Metab 2020, 42, 101059. [Google Scholar] [CrossRef]
- Arai, H.; Miyamoto, K.-I.; Taketani, Y.; Yamamoto, H.; Iemori, Y.; Morita, K.; Tonai, T.; Nishisho, T.; Mori, S.; Takeda, E. A Vitamin D Receptor Gene Polymorphism in the Translation Initiation Codon: Effect on Protein Activity and Relation to Bone Mineral Density in Japanese Women. J. Bone Min. Res. 1997, 12, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Colin, E.M.; Weel, A.E.A.M.; Uitterlinden, A.G.; Buurman, C.J.; Birkenhäger, J.C.; Pols, H.A.P.; Van Leeuwen, J.P.T.M. Consequences of vitamin D receptor gene polymorphisms for growth inhibition of cultured human peripheral blood mononuclear cells by 1,25-dihydroxyvitamin D 3. Clin. Endocrinol. 2000, 52, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.; Krishnan, A.V.; Malloy, P.J.; Eccleshall, T.R.; Zhao, X.-Y.; Feldman, D. The Vitamin D Receptor Gene Start Codon Polymorphism: A Functional Analysis of Fok I Variants. J. Bone Min. Res. 1998, 13, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.M.; Zmuda, J.M.; Cauley, J.A.; Shea, P.R.; Ferrell, R.E. Vitamin D receptor genotype is associated with fat-free mass and sarcopenia in elderly men. J. Gerontol. A Biol. Sci. Med. Sci. 2004, 59, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Hopkinson, N.S.; Li, K.W.; Kehoe, A.; Humphries, S.E.; Roughton, M.; Moxham, J.; Montgomery, H.; Polkey, M.I. Vitamin D receptor genotypes influence quadriceps strength in chronic obstructive pulmonary disease. Am. J. Clin. Nutr. 2008, 87, 385–390. [Google Scholar] [CrossRef]
- Xia, Z.; Man, Q.; Li, L.; Song, P.; Jia, S.; Song, S.; Meng, L.; Zhang, J. Vitamin D receptor gene polymorphisms modify the association of serum 25-hydroxyvitamin D levels with handgrip strength in the elderly in Northern China, Nutrition 2019, 57, 202–207. [CrossRef]
- Gussago, C.; Arosio, B.; Guerini, F.R.; Ferri, E.; Costa, A.S.; Casati, M.; Bollini, E.M.; Ronchetti, F.; Colombo, E.; Bernardelli, G.; Clerici, M.; Mari, D. Impact of vitamin D receptor polymorphisms in centenarians. Endocrine 2016, 53, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Micheli, M.L.; Gulisano, M.; Morucci, G.; Punzi, T.; Ruggiero, M.; Ceroti, M.; Marella, M.; Castellini, E.; Pacini, S. Angiotensin-Converting Enzyme/Vitamin D Receptor Gene Polymorphisms and Bioelectrical Impedance Analysis in Predicting Athletic Performances of Italian Young Soccer Players. J. Strength Cond. Res. 2011, 25, 2084–2091. [Google Scholar] [CrossRef]
- Jia, F.: Sun, R.-F.; Li, Q.-H.; Wang, D.-X.; Zhao, F.; Li, J.-M.; Pu, Q.; Zhang, Z.-Z.; Jin, Y.; Liu, B.-L.; Xiong, Y. Vitamin D Receptor Bsm I Polymorphism and Osteoporosis Risk: A Meta-Analysis from 26 Studies. Genet. Test. Mol. Biomark. 2013, 17, 30–34. [CrossRef]
- Qin, G.; Dong, Z.; Zeng, P.; Liu, M.; Liao, X. Association of vitamin D receptor BsmI gene polymorphism with risk of osteoporosis: a meta-analysis of 41 studies. Mol. Biol. Rep. 2013, 40, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Bahat, G.; Saka, B.; Erten, N.; Ozbek, U.; Coskunpinar, E.; Yildiz, S.; Sahinkaya, T.; Akif Karan, M. BsmI polymorphism in the vitamin D receptor gene is associated with leg extensor muscle strength in elderly men. Aging Clin. Exp. Res. 2010, 22, 198–205. [Google Scholar] [CrossRef]
- Grundberg, E.; Brandstrom, H.; Ribom, E.; Ljunggren, O.; Mallmin, H.; Kindmark, A. Genetic variation in the human vitamin D receptor is associated with muscle strength, fat mass and body weight in Swedish women. Eur. J. Endocrinol. 2004, 150, 323–328. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Oono, F.; Iida, K.; Wang, P.-L.; Tachi, Y. Relationship between vitamin D receptor gene polymorphisms (BsmI, TaqI, ApaI, and FokI) and calcium intake on bone mass in young Japanese women. BMC Women’s Health 2021, 21, 76. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Liu, B.; Chen, B.; Zhu, W.; Ye, X.-H.; Li, H.; He, X. Evaluation of Association Studies and an Updated Meta-Analysis of VDR Polymorphisms in Osteoporotic Fracture Risk. Front. Genet. 2022, 12. [Google Scholar] [CrossRef]
- Iki, M.; Saito, Y.; Dohi, Y.; Kajita, E.; Nishino, H.; Yonemasu, K.; Kusaka, Y. Greater Trunk Muscle Torque Reduces Postmenopausal Bone Loss at the Spine Independently of Age, Body Size, and Vitamin D Receptor Genotype in Japanese Women. Calcified Tissue Intern. 2002, 71, 300–307. [Google Scholar] [CrossRef]
- Wu, F.-Y., Liu, C.-S.; Liao, L.-N.; Li, C.-I.; Lin, C.-H.; Yang, C.-W.; Meng, N.-H.; Lin, W.-Y.; Chang, C.-K.; Hsiao, J.-H.; Li, T.-C.; Lin, C.-C. Vitamin D receptor variability and physical activity are jointly associated with low handgrip strength and osteoporosis in community-dwelling elderly people in Taiwan: the Taichung Community Health Study for Elders (TCHS-E). Osteoporosis Int. 2014, 25, 1917–1929. [CrossRef]
- Wang, P.; Ma, L.; Wang, H.; Zhang, W.; Tian, Q.; Cao, D.; Zheng, G.; Sun, Y. Association between Polymorphisms of Vitamin D Receptor Gene ApaI, BsmI and TaqI and Muscular Strength in Young Chinese Women. Int. J. Sports Med. 2005, 27, 182–186. [Google Scholar] [CrossRef]
- Lohman, T.G.; Roche, A.F.; Martorell, R. Anthropometric standardization reference manual. Ed. Human Kinetics Books, Champaign, IL. 1988. [Google Scholar]
- Slaughter, M.H.; Lohman, T.G.; Boileau, R.A.; Horswill, C.A.; Stillman, R.J.; Van Loan, M.D.; Bemben, D.A. Skinfold equations for estimation of body fatness in children and youth. Hum. Biol. 1988, 60, 709–723. [Google Scholar]
- Reilly, T.; George, K.; Marfell-Jones, M.; Scott, M.; Sutton, L.; Wallace, J. How Well do Skinfold Equations Predict Percent Body Fat in Elite Soccer Players? Int. J. Sports Med. 2009, 30, 607–613. [Google Scholar] [CrossRef]
- Núñez, F.J.; Munguía-Izquierdo, D.; Suárez-Arrones, L. Validity of Field Methods to Estimate Fat-Free Mass Changes Throughout the Season in Elite Youth Soccer Players. Front. Physiol. 2020, 11, 16. [Google Scholar] [CrossRef]
- Frisancho, A. Anthropometric Standards for the Assessment of Growth and Nutritional Status. 1st ed.; Ann Arbor, University of Michigan Press, Michigan (USA) 1990. [CrossRef]
- Yi, X.; Liang, Y.; Huerta-Sanchez, E.; Jin, X.; Cuo, Z.X.P.; Pool, J.E.; Xu, X.; Jiang, H.; Vinckenbosch, N.; Korneliussen, T.S.; et al. Sequencing of 50 Human Exomes Reveals Adaptation to High Altitude. Science 2010, 329, 75–78. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; Sham, P.C. ). PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; McVean, G.; Durbin, R. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2022. https://www.R-project.org/. (n.d.).
- Machiela, M.J.; Chanock, S.J. LDlink: a web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B. Vitamin D and muscle function. J. Steroid Biochem. Mol. Biol. 2017, 173, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Yang, L.; Li, M.; Xiao, H. Relationship of vitamin D receptor gene polymorphism with sarcopenia and muscle traits based on propensity score matching. J. Clin. Lab. Anal. 2020, 34. [Google Scholar] [CrossRef]
- Kerr Whitfield, G.; Remus, L.S.; Jurutka, P.W.; Zitzer, H.; Oza, A.K.; Dang, H.T.L.; Haussler, C.A.; Galligan, M.A.; Thatcher, M.L.; Dominguez, C.E.; Haussler, M.R. Functionally relevant polymorphisms in the human nuclear vitamin D receptor gene. Molecular and Cellular Endocrinology, 177(1–2), 145–159. [CrossRef]
- Vuolo, L.; Di Somma, C.; Faggiano, A.; Colao, A. Vitamin D and Cancer. Front. Endocrinol. 2012, 3. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, L.-J.; Zhou, Y.; Badr, R.; Watson, P.; Ye, A.; Zhou, B.; Zhang, J.; Deng, H.-W.; Recker, R.R.; Lappe, J. M. SNP rs11185644 of RXRA gene is identified for dose-response variability to vitamin D3 supplementation: a randomized clinical trial. Sci. Rep. 2017, 7, 40593. [Google Scholar] [CrossRef]
Figure 1.
PBS representation of VDR gene. Dashed and horizontal lines represent the 99.5th and 99.9th percentiles of the empirical distribution, respectively. SNPs under scrutiny are indicated with a square.
Figure 1.
PBS representation of VDR gene. Dashed and horizontal lines represent the 99.5th and 99.9th percentiles of the empirical distribution, respectively. SNPs under scrutiny are indicated with a square.
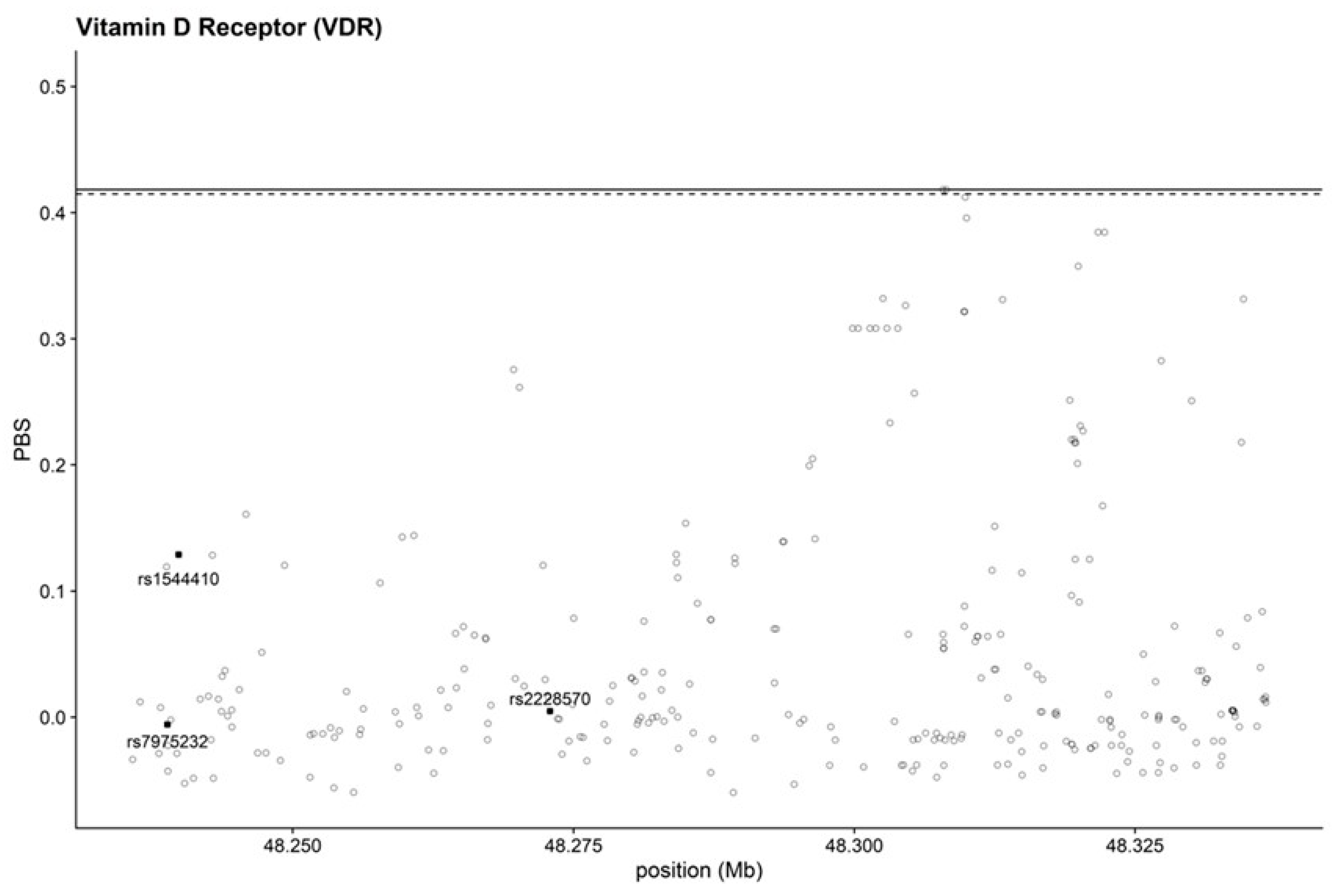

Table 1.
Allele and genotype frequencies. Values are indicated in percentages, while absolute values are shown in parentheses.
Table 1.
Allele and genotype frequencies. Values are indicated in percentages, while absolute values are shown in parentheses.
| VDR ApaI rs7975232 (N = 55) | ||||
|---|---|---|---|---|
| A | C | AA | AC | CC |
| 58.18 | 41.82 | 30.91 (17) | 54.54 (30) | 14.55 (8) |
| VDR BsmI rs1544410 (N = 54) | ||||
| G | A | GG | AG | AA |
| 55.56 | 44.44 | 35.18 (19) | 40.74 (22) | 24.08 (13) |
| VDR FokI rs2228570 (N = 54) | ||||
| A | G | AA | AG | GG |
| 25.93 | 74.07 | 3.71 (2) | 44.44 (24) | 51.85 (28) |
Table 2.
p values calculated for ANOVA test.
| ApaI | BsmI | FokI | ||||
|---|---|---|---|---|---|---|
| F value | p | F value | p | F value | p | |
| FFM Slaughter | 0.997484 | 0.375752 | 0.105224 | 0.900317 | 1.016038 | 0.369237 |
| FFM Reilly | 0.538281 | 0.586968 | 0.046829 | 0.954292 | 0.290343 | 0.749235 |
| AMA | 1.022569 | 0.366788 | 0.08582 | 0.917892 | 1.449376 | 0.244221 |
| CMA | 1.132188 | 0.330143 | 2.263845 | 0.1143 | 0.100998133 | 0.90411506 |
| TMA | 0.536284 | 0.588118 | 1.382144 | 0.260282 | 0.536284 | 0.588118 |
F value: the test statistic from the F test. p: the p-value of the F statistic.
Table 3.
BsmI association with CMA.
| Model | Genotype | n | Response means (s.e.) | Difference (95% CI) | p-value | AIC | BIC |
|---|---|---|---|---|---|---|---|
| Codominant | GG | 19 | 89.04 (2.58) | 0.00 | 0.11 | 420 | 430 |
| AG | 22 | 95.37 (2.93) | 6.93 (0.03–13.82) | ||||
| AA | 13 | 97.18 (2.34) | 6.79 (−1.18–14.77) | ||||
| Dominant | GG | 19 | 89.04 (2.58) | 0.00 | 0.034 | 418 | 426 |
| AG – AA | 35 | 96.05 (2.02) | 6.88 (0.68–13.08) | ||||
| Recessive | GG – AG | 41 | 92.44 (2.02) | 0.00 | 0.4 | 422 | 430 |
| AA | 13 | 97.18 (2.34) | 3.16 (−4.15–10.46) | ||||
| Overdominant | GG -AA | 32 | 92.34 (1.92) | 0.00 | 0.19 | 421 | 429 |
| AG | 22 | 95.37 (2.93) | 4.26 (−1.99–10.52) | ||||
| Log-additive | 3.71 (−0.25–7.67) | 0.072 | 419 | 427 |
AIC: Akaike Information Criterion. BIC: Bayesian Information Criterion.
Table 4.
Haplotype association with response with CMA.
| ApaI | BsmI | Frequency | Difference (95% CI) | p-value |
|---|---|---|---|---|
| A | A | 0.4138 | 0.00 | --- |
| C | G | 0.3807 | −3.68 (−8.74–1.39) | 0.16 |
| A | G | 0.168 | −6.49 (−12.59–−0.4) | 0.04 |
| C | A | 0.0375 | −11.09 (−27.62–5.45) | 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



