
Submitted:
19 July 2024
Posted:
19 July 2024
You are already at the latest version
Abstract
Since the 20th century, numerous studies have detected or isolated parasites from the Trypanosomatidae family in various tick species. However, the status of ticks as vectors of medically or veterinary important Trypanosoma and Leishmania remains unclear. We conducted a systematic review and meta-analysis to pro-vide new insights into the potential vector status of Leishmania and Trypanosoma, which have medical and veterinary significance. We searched three databases (PubMed, Google Scholar, and Web of Science) from 1912 to June 30, 2023, resulting in 94 and 86 papers included in the qualitative and quantitative analyses, respectively. All identified field studies were conducted in endemic areas and investigated the presence of Trypanosoma and Leishmania parasite, DNA, or antigen in ticks. We recorded a pooled prevalence of Trypa-nosomatidae detection in ticks at 15.48% [7.99-24.61%], with significant variations depending on the year, detection method, and geographical area. Most of positive tick species belonged to the genera Amblyomma, Hyalomma, Ixodes, and Rhipicephalus. Experimental laboratory work on transmission routes proved potential vector competence in both the Argasidae and Ixodidae tick families. Although our systematic review and meta-analysis provide unambiguous evidence of the natural infection of ticks by Trypanosomatidae para-sites with some evidence of non-traditional transmission routes, it does not provide conclusive evidence on the role of ticks as biological or mechanical vectors for veterinary and medically interest protozoan Trypa-nosomatidae species. All these unambiguously demonstrates the need of additional investigations to address this point.
Keywords:
Trypanosomatidae
; Trypanosoma
; Leishmania
; Ticks
; Ixodidae
; Argasidae
; vector borne disease
1. Introduction
Ticks (Parasitiformes: Ixodida) are obligate hematophagous ectoparasites of all vertebrate classes and are vectors or reservoirs of human and animal pathogenic virus, bacteria, protozoa and fungi fungi [1]. Two families of ticks of numerous genuses possess considerable significance in public health and veterinary matters [2]. Currently, there are 996 known tick species worldwide, which are divided into three families: ∼774 hard tick species (Ixodidae), ∼221 soft tick species (Argasidae), and the monotypic family of Nuttalliellidae, mixing features of both families [3,4]. The identification of tick fossils in amber has led to the description of two novel families, the Deinocrotonidae and Khimairidae, while many unique genera are described, including Compluriscututla, Cornupalpatum, Deinocroton, and Khimaira [5]. Protozoan pathogens carried by ticks and affecting mammals of medical or veterinary interest are classified under the order Piroplasmida (e.g., genera Babesia, Theileria, and Cytauxzoon) or haemogregarines of the Hepatozoon genus (Adeleorina: Hepatozoidae) [6].
The Trypanosomatidae family consists of unicellular eukaryotes, including pathogens of humans and animals from the genera Trypanosoma and Leishmania, as well as Endotrypanum and Porcisia. The life cycle of these organisms is characterized by the involvement of arthropod vectors from the Hemiptera and Diptera orders, mainly, and for some Trypanosomes from the Siphonoptera orders and for fish Trypanosome of the Arhynchobdellida (Hirudinidae, leech) order. Two Trypanosoma subspecies of T. brucei (i.e., Trypanosoma brucei gambiense, T. brucei rhodesiense), and atypical human trypanosomiasis T. cruzi, and T. rangeli, along with at least 23 species of Leishmania, including the recently described Leishmania (Mundinia) chancei species, are pathogenic for humans. They cause human African trypanosomiasis (HAT or sleeping sickness), Chagas disease (CD), and cutaneous (CL), mucocutaneous (MCL), or visceral (VL) Leishmaniases [7,8,9,10,11,12,13]. These diseases also affect domestic, feral, or wild animals. Canine visceral Leishmaniases (CVL) are primarily caused by L. infantum infection and occasionally by L. donovani or L. tropica [14]. Trypanosoma congolense, T. evansi, T. b. brucei, T. vivax, T. simiae, T. suis, T. theleiri, and more rarely, T. godfreyi infect livestock. Trypanosoma equiperdum infects equids [15,16]. In addition, Trypanosoma theileri which is considered as non-pathogenic, is found in cattle, buffalo, and antelope worldwide [17]. It has been unfrequently linked to disease resembling nagana in specific cases: a calf [18], cattle [19], or a cow [20]. Trypanosoma theileri can cause illness in cattle under severe stress due to concurrent diseases or poor nutrition [21]. Trypanosoma caninum was described in 2014 as a new Trypanosoma species infecting dogs, in asymptomatic cases with low humoral immune response [22]. Additionaly, T. lewisi, that infect Rattus sp. Is also an opportistic bllod parasites of human and share common vertebrate hosts with T. Cruzi [23]. Altogether, more than 30 million people are infected, and over 48 million cattle are at risk of contracting animal trypanosomiasis in Africa, causing about 3 million deaths in cattle every year [24].
Since the early 20th century, the idea that ticks can transmit protozoan parasites from the genera Leishmania and Trypanosoma has been investigated, and more recently, molecular biology techniques have been used to revisit this hypothesis (e.g., Next-Generation Sequencing) [25,26]. There is still an ongoing debate within the scientific community regarding ticks' ability to transmit Trypanosomatidae parasites of medical or veterinary interest. To provide an updated perspective on this current debate, we undertook a systematic review and meta-analysis with the following objectives: (i) to gather published field and experimental data on the detection in ticks of Trypanosomatidae with medical and veterinary interests, including T. theleiri and T. caninum, and to assess their capacity to act as vectors for these pathogens; (ii) to explore factors associated with the transmission of these pathogens by ticks.
he introduction should briefly place the study in a broad context and highlight why it is important. It should define the purpose of the work and its significance. The current state of the research field should be carefully reviewed and key publications cited. Please highlight controversial and diverging hypotheses when necessary. Finally, briefly mention the main aim of the work and highlight the principal conclusions. As far as possible, please keep the introduction comprehensible to scientists outside your particular field of research. References should be numbered in order of appearance and indicated by a numeral or numerals in square brackets—e.g., [1] or [2,3], or [4,5,6]. See the end of the document for further details on references.
2. Materials and Methods
2.1. Protocol and Registration
The current study was conducted following the Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) guidelines [166]. The protocol was neither registered nor published. The checklist for meta-analysis is provided as Supplementary Material (Check List S1).
2.2. Information Source
The systematic screening of existing literature was performed using PubMed, Google Scholar, and Web of Science (WOS) databases. Publish or Perish (Harzing.com), a software that retrieves and analyses academic citations was used to retrieve relevant articles from Google Scholar.
2.3. Search
A set of keywords was used: “Trypanosomatidae” and all species of “Trypanosoma” or “Leishmania”, known to be pathogenic for humans or animals and having medical or veterinary interest, including T. theleiri and T. caninum, in combination with “ticks” “Ixodidae”, “Argasidae”, and selected tick genus (e.g., “Rhipicephalus”, “Hyalomma”, “Ornithodoros”). Database search was done between January 1900 and June 30, 2023, without language restrictions. Articles reporting “Trypanosomatidae” in ticks by direct examination (e.g., culture, microscopy, immunohistochemistry “IHC”) or molecular method (e.g., conventional Polymerase chain reaction “PCR”, real-time PCR “RT-PCR”, capillary or next generation sequencing “NGS”) were included in the study. We exclude studies that doesn’t fall in the scope of our study and reporting, i.e., those whose scope and objective are “Vaccine”, “Virus”, “Drug”, “Treat”, “Pest”, “Acaricide”, “Immunology”, “Serology”, “Serum”, “ELISA”, “Antigen”.
2.4. Eligibility Criteria and Study Selection
We undertook the review with current recommendations reported in 2015 and reported our findings as per the PRISMA guidelines, taking into account the remarks for “biological” meta-analyses [167]. Two authors, TK and DS, independently conducted a preliminary review of the articles by examining their titles and abstracts. Articles selected by at least one reviewer were retrieved, and duplicated papers were excluded. Two dependent reviewers (TK and DS) performed a second selection based on full-text analysis, and disagreements were resolved through discussion with a third reviewer (BM). Studies were considered eligible based on established inclusion or exclusion criteria.
The selection of eligible articles was based on previously established criteria: (1) articles dealing with the detection or transmission of Trypanosomatidae parasites of medical or veterinarian interest, including T. theleiri and T. caninum, in ticks; (2) field or experimental studies; (3) with no restrictions on the host of origin; and (4) without language restriction. The exclusion criteria included (1) literature categories (letters, books, and reviews); (2) studies not focusing on ticks as vectors for Trypanosomatidae (e.g., cellular immune responses of ticks, host immunogenetic influences on tick resistance); (3) studies not focusing on Trypanosomatidae of medical or veterinary interest; (4) studies lacking information on tick collection place and origin; and (5) studies not stating the sample size, the number of positive cases, and the identity of tick and Trypanosomatidae species.
2.5. Data Collection
We employed a pre-existing template to obtain data from imported articles, and two of the authors (TK and DS) compiled them in a Microsoft Excel® spreadsheet. The information collected included the author’s name, publication year, study's subregion/country, tick family and species, identification methods, number of tested and positive tick samples, and detection methods.
2.6. Quality Assessment
The quality of the selected publications was assessed following the Grading of Recommendations Assessment, Development, and Evaluation (GRADE) [168]. The total score of each article was calculated using the following seven items: (1) the tick family and species identified; (2) the Trypanosomatidae species identified; (3) the detection method stated; (4) the number of tested ticks; (5) the number of positive ticks; (6) the prevalence of Trypanosomatidae parasites in ticks; and (7) the identity of the host where positive tick(s) originate stated. Items 1-5 are quoted for 2 points, while items 6 and 7 for 1. Based on this score, each publication was classified as of high quality (score = 8-12), medium quality (score = 5-7), or poor quality (score = 0-4).
2.7. Statistical Analysis
For data related to field studies, we performed a meta-analysis of proportions [169] using the 'meta' and 'metafor' packages in R software version 4.3.1. We performed a Freeman-Tukey transformation with double arc sine (PFT) to convert the proportions before meta-analysis. This transformation is beneficial for standardizing and stabilizing the distribution variance [170] (dat<-escalc(measure="PFT", xi=xi, ni=ni, data=dat). Because of the high heterogeneity expected in the meta-analysis, a random-effects model was performed to combine the total size effect and subgroup analysis. Cochrane statistics I2 and Q (expressed as X2 and P, respectively) were used to assess and quantify heterogeneity. I2 < 50% corresponds to low heterogeneity, whereas I2 > 50% indicates high heterogeneity. Meta-analysis statistics were visualized using a Forest plots representation. The Egger test and Funnel plots were performed to assess publication bias, and the results were further submitted to a stability analysis. This later evaluates the impact of deleting data extracted from an article on the results of the remaining papers.
We carried out subgroup analyses of potential risk factors, namely: sampling year (before 2000, 2001-2009, 2010-2019, 2020 and after), continent (Europe, Asia, Africa, South America, and Iceland), host family (Canidae, Bovidae, etc.), tick genus, tick species, detection methods, parasite type, and the location of the tick’s organs. We also performed a meta-regression with the parameters studied used as co-variables to address possible heterogeneity sources.
Concerning the experimental studies, because in a large number of papers, quantitative data are lacking (number of ticks used for the study), we performed a meta-analysis on semi-quantitative data, i.e., detection or not of pathogens in ticks (0: absence, 1: presence). Using this methodology, we also analyzed the detection of pathogens after a blood meal, mechanical transmission, transmission following the injection of homogenate ticks, and vertical transmission. Statistical tests mentioned above were calculated, and subgroup analyses of potential risk factors were performed on the tick family, tick species, donor host, receiving host, family of the parasite tested, and parasite species.
3. Results
3.1. Selection Process Overview: Curating Datasets for Systematic Review
The systematic search on multiple databases, including PubMed, Google Scholar, MEDLINE®, SciELO, and WOS, a total of 29,592 articles were retrieved (Figure 1). After the removal of duplicates (n= 21,007) and ineligible papers (n = 8,423), 166 published studies were selected (last updated 30 June 2023). After analysis of their titles, abstracts, and detailed contents, out of those 166 articles, 94 satisfied the eligibility criteria to be included in the systematic review, 86 articles were eligible for the meta-analysis, of which 49 dealt with field studies and 37 with experimental studies. In addition, 46 are focused on the transmission of Leishmania, while 40 are focused on Trypanosoma. Full-text Portable Document Format (PDF) files not freely accessible online were obtained through the French Development Research Institute (IRD) library. Publications in French, German, Spanish, Portuguese, Russian, and Turkish languages were handled by both authors (TK and DS) and/or native language colleagues.
3.2. Exploring the Historical Presence in Tick of Trypanosomatidae Infecting Human and Animals of Veterinary Interests
The Arthropoda phylum hosts monoxenous and dixenous Trypanosomatidae [27]. Sand flies (Diptera; Phlebotominae) are biological vectors of Leishmania, with some exceptions [28]. Diptera (e.g., Tsetse flies, Tabanids Stomoxes), Hemiptera (e.g., triatomine ‘reduviid’ bugs), Siphonaptera (e.g., fleas), and Arhynchobdellida (Hirudinidae, leech) are biological vectors of Trypanosoma [29,30]. Although most Trypanosomatidae colonize members of the Insecta class, Blastocrithidia sp. has been reported in ticks (Arachnida class) [31]. Since the late 19th and early 20th centuries, field trials and experiments have been conducted to detect these parasites in ticks (Figure 2). The presence of developmental forms of Trypanosoma spp. in ticks by O'Farrell (1913) and the demonstration that ticks can transmit parasites of the Leishmania genus are documented long ago [32]. Over the past several years, improved molecular detection of Trypanosomatidae DNA in ticks has generated renewed interest in the role of ticks in the transmission of Trypanosoma and Leishmania. Specifically, since 2010, the number of studies investigating this topic has increased significantly.
3.2.1. Ticks as Vectors of Trypanosoma: Historical Aspects
The hypothesis that ticks can act as reservoirs and/or vectors of human and animal Trypanosomes has been investigated as early as the beginning of the 20th century [33,34,35,36,37,38,39,40,41,42,43,44,45,46,47]. In 1972, Hoare reviewed existing data on ticks acting as vectors of trypanosomes and inferred that ticks could not transmit them because of the absence of infective forms, the trypomastigotes [29]. Nevertheless, Burgdorfer et al. (1973) and Shastri and Deshpande (1981) described the presence of various trypanosomes developmental stages in ticks (i.e., amastigotes, sphaeromastigotes, epimastigotes, and trypomastigotes) [48,49].
3.2.2. Leishmania: a Parasite Transmitted by Ticks?
The idea that ticks contribute to the transmission of Leishmania emerged in the 20th century. Early experiments by Blanc and Caminopetros suggested that ticks could be vectors for Leishmania causing human and canine kala-azar (L. donovani/L. infantum complex) around the Mediterranean. However, this idea was questioned by Malamos in 1938, that Leishmania could survive in the tick gut for more or less extended periods, which called into question the concept of direct transmission [50]. This hypothesis was questioned in 1972 by Rioux et al. who provided evidence contradicting the involvement of ticks in the cycle of L. infantum [51]. In the mid-1980s, McKenzie's contradicted this idea by proving the transmission of Leishmania during ticks' blood meal [52]. Subsequent field and laboratory studies identified Leishmania RNA or DNA within tick organs (including the salivary glands and ovaries) at different development stages (from larvae to nymphs and adults), further supporting the role of ticks in Leishmania transmission [53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77].
3.3. Observations of Trypanosomatidae of Medical and Veterinary Interest in Field-Collected Tick Specimens
As detailed in the following sections, many field studies were conducted to investigate the prevalence of pathogenic Trypanosomatidae in ticks by detecting the parasite by direct examination (e.g., culture, microscopy), molecular tools like Polymerase Chain Reaction (PCR), quantitative Polymerase chain reaction (qPCR), capillary sequencing, or Next-generation Sequencing (NGS), or immunological tools (Immuno histochromatography).
3.3.1. Trypanosoma Carriage and Prevalence
Traces of the presence of Trypanosomes of medical/veterinary interest were reported in ticks; these include Trypanosoma cruzi, T. vivax, T. evansi, T. theileri, T. theileri-like, T. congolense, T. caninum [36,78,87,88,89,90,91,79,80,81,82,83,84,85,86]. All these studies were performed on hard tick species (Ixodidae). Most tick species positive belonged to four genera, Amblyomma, Hyalomma, Ixodes, and Rhipicephalus, including A. cajennense [83,90], A. longirostrum [36], A. variegatum [78], H. detritum [88], H. marginatum [80], I. ricinus [86,88], Rhipicephalus (Boophilus) microplus [81,87,90], R. (Boophilus) spp. [85], R. sanguineus [82,88], R. sanguineus s.L.[89,91], Rhipicephalus sp. [84]. Conversely, other species were negative for Trypanosoma sp. (Table 1) [84,85,88,89,90,91,92]. Ticks collected on dogs and cattle were most frequently infected by Trypanosoma compared to those collected on other domestic animals (e.g., sheep, goats, camels, etc.) or wild animals (e.g., foxes, boars, etc.).
- Trypanosoma cruzi
Since the 1940s, the prevalence of T. cruzi in field-collected ticks has remained uncertain. Most of the published studies focus on ticks taken from infected animals. A study by Pifano (1941) on ectoparasites of Cercolabidae in the State of Yaracuy, Venezuela, documented A. longirostrum, a parasite of the porcupine, naturally infected with T. cruzi [36]. In 2013-2015, ticks were collected from 148 dogs in the urban area of Campo Grande, state of Mato Grosso do Sul, Brazil. This study revealed that R. sanguineus and A. cajennense were not infected by T. cruzi [92]. Recently, Opazo et al. (2022) investigated the presence of T. cruzi in dogs and their ectoparasites in a rural area of Central Chile. 57% of blood samples were infected by T. cruzi, and 5.4% of the ticks were positive by PCR, particularly R. sanguineus s.l. (5/82), whereas all A. tigrinum specimens (0/11) were negative [89]. The same year, a study focused on ticks' utility as xenosurveillance sentinels to survey circulating pathogens in Kenyan drylands. Ticks collected from wild ungulates, carnivores, domestic animals, and Boran cattle were screened for pathogens using metagenomics. Trypanosoma cruzi DNA was detected in 3/46 (6.5%) pools of Rhipicephalus spp. but not in Amblyomma spp. [84]. Nevertheless, careful examination of the sequences provides evidence that they belong to the Schizotrypanum genus but cannot definitively identify them as T. cruzi.
- Trypanosoma vivax
Trypanosoma vivax infects wild domestic ungulates and is transmitted mechanically via tabanids and other blood-sucking insects in the Americas [83]. The search for T. vivax’s presence has only recently been engaged. Postoyan et al. (2001) analyzed two tick species, A. cajennense (Fabricius) sensu stricto (s.s.) and R. (Boophilus) microplus (Canestrini) (Acari: Ixodidae), collected from cattle (Bos taurus) on two farms in Cuba. The gut contents and hemolymph of these ticks were smear-examined for flagellates. In both species, living T. vivax forms were observed in the gut for 96 hours after repletion but not in the hemolymph [90]. In 2013, Bolivar et al. conducted a study in which 285 R. (Boophilus) microplus ticks were collected from cattle in the livestock areas of the state of Merida, Venezuela. It was found that 7.7% of the samples tested were positive using PCR [81]. In the same year, Cominetti et al. examined 63 specimens of R. sanguineus collected from dogs in the city of Campo Grande, Brazil, for the presence of Trypanosoma spp. using PCR. 15 (23.8%) were positive for T. vivax [82]. Dyonisio et al. (2021) looked for T. vivax in both A. cajennense sensu stricto and R. (Boophilus) microplus collected on cattle. Of the 48 and 45 specimens of each species, 6.25% (3/48) of A. cajennense s.s. and 4.5% (2/45) of R. (Boophilus) microplus were positive for T. vivax. The sequences obtained were 99% identical to those of bovine T. vivax from north-eastern Brazil [83].
- Trypanosoma evansi
Trypanosoma evansi is primarily transmitted by tabanids and Stomoxys spp. It affects a wide range of hosts, including livestock, camelids, equids, carnivores, rodents, and humans, and produces variable clinical symptoms depending on the host and the geographic region [91]. In a study conducted by Monod et al. (1986) on the seasonal dynamics of R. sanguineus in dogs in the urban areas of western Algeria, several hemolymph smears were prepared to search for potential microorganisms. Five of the 250 R. sanguineus examined were infected with trypanosomes (four females and one male), representing 2% of the total. It is worth noting that two infected females displayed a pathological reaction (globular body and milky hemolymph) that may be related to the infection. Based on the observed form and small size of the protozoan, Monod et al. hypothesized that it could be T. berberum (evansi), a parasite of the Camelidae that can also cause deadly trypanosomiasis in dogs [88]. This hypothesis is corroborated earlier by Cominetti et al. (2013), who found 15 out of 63 dogs in the city of Campo Grande, MS, Brazil, positive for T. vivax and 7 for T. evansi through PCR [82]. In 2021, Said et al. found that 0.2% of the 352 ticks collected from the environment and wild Tunisian ruminants were positive for T. evansi by PCR [91]. Krishnamoorthy et al. (2021) reported a prevalence of T. evansi in R. sanguineus s.l of 0.2%. PCR and sequencing of 240 pooled tick DNA samples display that the prevalence of T. evansi was null in Karnataka and 8.3% in Kerala, India [85].
- Trypanosoma theileri and T. theileri-like
Multiple studies document the presence of trypanosomes in tick hemolymph [31,48,79,88,93]. Most of these studies identified T. theileri, which is typically transmitted by tabanids [94]. Trypanosoma theileri was first reported in field-collected ticks in a study published in 1979 in Switzerland. In this study, 0.19% (5 out of 2501) of Ixodes ricinus were found to carry trypanosomes morphologically similar to T. theileri. [79]. Between 1979 and 1982, the presence of T. theileri in the hemolymph of ticks from Switzerland and Algeria was confirmed: 0.24% (37/1570) of H. detritum and I. ricinus ticks collected from bovines tested positive [88]. In 2008, Martins reported the presence of Trypanosomatidae epimastigote forms in the hemolymph of cattle tick R. (Boophilus) microplus from Rio Grande do Sul, Southern Brazil. Again, the microscopic examination suggested that these forms were similar to those of T. theileri [87].
- Trypanosoma congolense
Animal trypanosomosis is a complex disease caused by one or more species of pathogenic trypanosomes, including T. congolense. Clinical symptoms are characterised by intermittent fever, parasitemia, anemia, lymphadenopathy, jaundice, progressive emaciation, weakness, and reduced productivity [95]. Trypanosomes transmitted by tsetse flies (Glossina spp.) are responsible for animal African trypanosomoses in livestock in sub-Saharan Africa [96]. In 2016, trypanosomes were detected in engorged adult A. variegatum collected from cattle in the Unguwan Rimi and Kaduna state areas in northwestern Nigeria. Of the 33 samples examined under a microscope, 14 were positive for T. congolense, representing a prevalence rate of 42.4%. On the first day of parasitemia follow-up, 10 (30.3%) samples were microscopically positive for T. congolense, while 23 (69.7%) were negative [78].
- Trypanosoma caninum
Trypanosoma caninum, the most recent species within the Trypanosomatidae family, was reported to infect dogs in Brazil [22]. Despite its recent identification, 67 cases of natural infection in dogs have been documented in areas where canine visceral Leishmaniosis is endemic [22,97]. Luu et al. (2020) isolated and partially characterised a novel trypanosome from I. ricinus collected in Slovakia, and the resulting sequences suggest that this trypanosome, referred to as Trypanosoma sp. Bratislava1 could be a new species closely related to several trypanosomes isolated from, or detected in, ticks in South America and Asia, including T. caninum isolated in Brazil [86].
3.3.2. Leishmania Prevalence in Field Collected Ticks
The presence of Leishmania is well documented in ticks, including Leishmania infantum/L. donovani , L. major, L. chagasi (syn L. infantum), L. braziliensis, L. guyanensis, and L. martiniquensis [55,56,57,58,59,60,62,63,64,66,67,68,69,74,76,77,92,98,99,100,101,102,103,104,105,106]. Most of the field studies focus on ticks that parasitise domestic animals in urban areas, like R. sanguineus [55,57,58,62,65,67,68,69,74,76,77,82,98,99,100,101,103,104,106]. Ticks collected from more wild ecosystems highlight the diversity of tick species that are positive for the presence of Leishmania, including A. sabanerae [105], Amblyomma spp. [63], A. tigrinum [58], A. variegatum [56], Hyalomma aegyptium, H. dromedarii [60], Ixode ricinus [62,64,67,102,104], Ixodes spp. [104], I. ventalloi [62,104], R. (Boophilus) microplus [56,63,105], R. pusillus [62,104], R. sanguineus s.l. and R. turanicus [60] (Table 2).
- Leishmania infantum and L. chagasi (Syn L. infantum)
The hypothesis that ticks can be vectors of L. infantum was first proposed in the early 20th century [113]. Domestic dogs play a central role as reservoirs of L. infantum in the peridomestic zoonotic transmission cycle. The brown dog tick, R. sanguineus (Latreille 1806), has been the subject of extensive research due to its prevalence among urban dogs [114]. The overall L. infantum prevalence given by molecular techniques in R. sanguineus collected on dogs [54,57,65,66,67,74,77,100,115] or cats [62,104] ranged from 2.5% to 70.3% in Brazil and Italy. Significant L. infantum presence is recorded in species of the Rhipicephalus genus, including 9.2% of R. sanguineus s.l. ticks collected with the flagging technique that collects questing ticks and 23.5% of R. turanicus collected from dogs, hedgehogs, and tortoises in Israel [60]. Additionally, 10.9% and 17.6% of infections were reported in R. pusillus from cats in Italy. Studies reported the detection of L. infantum in I. ricinus (1.5% to 50%) [62,64,67,102,104], I. ventalloi (19% and 62%) [62,104], Ixodes sp. (10.9%) [98], H. aegyptium (38.7%), and H. dromedarii (55.6%) [60], collected from dogs, horses, cats, bovines, tortoises, camels, and humans in Italy and Israel. In the majority of these studies, L. infantum DNA was detected in whole ticks and, in some cases, in the gut or salivary glands [57,74]. Leishmania chagasi (Syn L. infantum), which causes american visceral leishmaniasis, is widely distributed in Latin America [116]. It is thought that L. chagasi (syn L. infantum) was introduced in South America along with conquistadors' dogs [117]. In Campo Grande, state of Mato Grosso do Sul, Brazil, de Almeida et al. (2013) identified two tick species, R. sanguineus and A. cajennense, on dogs in urban areas. Tick samples from 36 dogs were positive for L. chagasi, and all were R. sanguineus [92].
- Leishmania major
Zoonotic Cutaneous Leishmaniasis (ZCL), caused by Leishmania major, is a public health problem in several countries, affecting a large number of people. Azarmi et al. (2022) suspected various arthropods, including ticks, to act as secondary vectors of Leishmania. They found one nymph of R. sanguineus positive for L. major in the rodent in Segzi plain in Esfahan Province, Iran [98].
- Leishmania braziliensis
American cutaneous leishmaniasis (ACL) is caused mainly by Leishmania braziliensis. De Morais et al. (2013) proposed that difficulties in controlling ACL might be related to the complex epidemiology of this disease, which involves the participation of various vector species, including ticks. Two rural areas in Pernambuco, northeastern Brazil, where ACL is endemic, were investigated for canine ectoparasites. Genomic DNA was extracted from 75 R. sanguineus ticks; 32 of them (42.67%) were positive for L. braziliensis using conventional PCR and real-time PCR (RT-qPCR) [76].
- Leishmania guyanensis
Leishmania guyanensis is a causal agent of American Tegumentary Leishmaniasis (ATL). In 2017, 81 R. (Boophilus) microplus and Amblyomma collected from three Tapirus terrestris and three Pecari tajacu in Madre de Dios, Perú, disclosed the presence of DNA belonging to the Leishmania (Viannia) subgenus after kDNA (kinetoplast DeoxyriboNucleic Acid) amplification [55]. Leishmania (Viannia) kDNA was detected in three R. microplus collected from a collard peccary (P. tajacu) hunted in the forests of Madre de Dios. The HRM-PCR (High Resolution Melting PCR) indicates that one positive sample displays a kDNA melting curve compatible with L. (V) guyanensis [63].
- Leishmania martiniquensis
Leishmania martiniquensis was first isolated in 1995; its taxonomical position was established in 2002, and it was named in 2014 [11,118,119,120]. The vectors and reservoirs have not yet been described, although biting midges are suspected to be involved in transmission of Leishmania belonging to the Mundinia subgenus [118,121]. In 2020, a high-throughput microfluidic real-time PCR system suitable for screening parasites at the genus-level was applied to 132 adult specimens of A. variegatum and 446 of R. microplus collected in Guadeloupe and Martinique. It was found that 0.7% of the R. microplus ticks from Martinique were positive for Leishmania spp., and the sequences identified L. martiniquensis [56].
- Other/unidentified Leishmania parasites
In 2010, the presence of Leishmania promastigotes forms in ticks parasitizing domestic dogs was reported in the Municipality of São Vicente Férrer, located in the Northern Agreste of Pernambuco, Brazil [106]. In 2015 and 2016, the presence of Leishmania was reported in the intestines, ovaries, and salivary glands by immunohistochemistry (IHC) but also by real-time PCR in R. sanguineus from dogs in Brazil. IHC detected Leishmania in 98% of the intestines, 14% of the ovaries, and 8% of the salivary glands. Real-time PCR indicates that the intestines, ovaries, and salivary glands were mostly positive [68,69]. Leishmania DNA (most likely belonging to the L. donovani complex) was detected in 11 out of 17 pools (64.7%) of A. tigrinum were collected from eight foxes (six grey foxes, Pseudalopex griseus and two culpeo foxes P. culpaeus) captured in Argentinean Patagonia [58]. The fact that sandflies were trapped only 2000 km and 750 km north of the study area suggests that their distribution may be wider than currently believed or that Leishmania is able to persist in another vector in this area of transmission. In 2022, 95.7% of R. (Boophilus) microplus and 90% of A. sabanerae collected from Pecari tajacu and Chelonoidis denticulata of the Leishmaniasis endemic zones in the Peruvian Amazon, with a load of 34.1 and 5428.6 parasites per arthropod [122].
3.4. Experimental infection and Transmission of Trypanosomatidae by Ticks
In the early 20th century, laboratory infections were conducted to probe the vector competence of ticks for trypanosomes. Several reports have demonstrated the presence of different forms of Trypanosoma or Leishmania, including trypomastigotes, amastigotes, epimastigotes, promastigotes, and sphaeromastigotes, in ixodides and argasides. The transmission routes for infestation were investigated from the bite of the tick and its blood meal to other alternative routes such as ingestion of infected ticks, passage between ticks at different life stages and their hosts (transstadial passage), or ovarian infection (transovarial passage).
3.4.1. Transmission of Trypanosoma following tick bites and feeding
The role of ticks as vectors of trypanosomes has been increasingly strengthened by recent studies that have provided additional evidence that ticks can transmit distinct clades of trypanosomes to various mammalian species [26]. Reports on the detection of parasites of the genus Trypanosoma after blood feeding on an infected animal in an experimental setting are available [40,43,45,46,123,124]. Trypanosomes of medical or veterinary interest were detected in hard and soft ticks from three tick genera (Rhipicephalus, Hyalomma, and Ornithodoros) and four species: R. sanguineus, H. anatolicum anatolicum, O. crossi, and O. lahorensis. Experimental infection of ticks following a bite or a meal was reported for T. cruzi, T. evansi, and T. theileri [125].
- Trypanosoma cruzi
The first available reports on dog infection by T. cruzi-infected R. sanguineus ticks date back to 1913 [40]. Mayer and Roca-Lima later attempted to infect rats with infected soft ticks of the genus Ornithodoros, but the results were negative [41]. Several other studies have failed to show the transmission of T. cruzi by soft ticks in laboratories (i.e., O. moubata, O. talaje, and O. turicata) [35,39,42,47].
- Trypanosoma evansi
Investigations into the biological transmission of T. evansi by ticks were initiated at the dawn of the 20th century. Studies have given clues on the capacity of a soft tick, O. crossi, infected by T. evansi to transmit the parasite after a blood meal to non-infected animals [43,44,46]. But, in 1924, Yorke and Macfie did not confirm the transmission of this protozoan by O. crossi [45]. In 1976, Taylor-Lewis also reported the non-transmission of T. evansi by H. dromedarii to rodents after a blood feeding [126]. Recently, Mahmoud et al. (2020) did not detect T. evansi DNA after blood-feeding infected O. savignyi on uninfected rodents [127].
- Trypanosoma theileri and Trypanosoma theileri-like
Shastri and Deshpande (1981) conducted experimental work on cattle and attempted unsuccessfully to infect calves with H. a. anatolicum carrying T. theileri [49]. A few years later, Morzaria succeeded in infecting calves and rats with H. a. anatolicum ticks infected with T. theileri-like [123]. In the early 2000s, Latif et al. reported the ability of Hyalomma ticks to transmit T. theileri but could not confirm its transmission to calves [124].
3.4.2. Transmission of Leishmania Following Tick Bites and Feeding
To date, sandflies (Phlebotominae) are recognized biological vectors of Leishmania [128], but the role of ticks, fleas, and leaches as potential secondary vectors has been evoked [71,129]. Five laboratory studies have attempted to infect ticks by allowing them to feed on vertebrates experimentally infected with Leishmania [32,51,52,70,130]. All experiments were performed on hard ticks (Ixodidae), and only ticks of the Rhipicephalus genus were involved because of their close relationship with dogs. Ticks were fed on dogs or rodents, and two species were used: R. turanicus and R. sanguineus. R. sanguineus was the only one to have been infected by Leishmania, referred to as L. infantum, and Leishmania sp. [52,70].
- Leishmania infantum
The transmission of L. infantum by ticks was first reported in 1930 by Blanc and Caminopétros, who infected squirrels after exposition to infected R. sanguineus ticks infected with what they named “Leishmania kala azar” at the time, which today most probably corresponds to parasites of the L. donovani complex [32]. Nevertheless, this was not confirmed by Joyeux and Sautet in 1938 [131] nor by Rioux et al. (1972) who failed to establish experimentally the transmission of Leishmania to rats by R. sanguineus [51]. More recently, de Almeida et al. (2016) were able to infect 14 hamsters with R. sanguineus infected with L. infantum [70]. In contrast, Rakhshanpour et al. (2017) could not confirm the transmission of L. infantum by R. sanguineus ticks collected from experimentally infected dogs [130].
3.4.3. Host Infection Following Ingestion of Infected Ticks or Injection with Trypanosoma-Infected Ticks
Documented transmission of trypanosomes following the ingestion of an infected invertebrate suggests that they may also be transmitted through the ingestion of infected ticks [29,132,133]. To date, nine studies on the Argasidae family and five on the Ixodidae family have investigated the transmission of trypanosomes in the laboratory through ingestion of infected ticks or infection through injection of an infected material [35,37,38,39,41,42,45,47,48,49,93,123,124,126]. Of these 14 studies, nine confirmed transmission by the injection of infected tick materials and only one by the ingestion of infected ticks. However, it is difficult to draw clear conclusions from these results that have not been obtained on the same tick genera or species, parasites, and hosts on which ticks have been fed or exposed to infected ticks. Seven tick species were tested, O. moubata, O. crossi, O. venezuelensis, O. turicata, O. furcosus, R. pulchellus, and H. a. anatolicum excavatum. Five Trypanosoma species were tested.
- Trypanosoma cruzi
Experimental T. cruzi transmission through ingestion or injection of tick infected material began in the 20th century. Mayer and Roca-Lima (1914) reported that injection of O. moubata infected with T. cruzi caused rodent infection [41]. The same author confirmed their findings in 1918 and mentioned that laboratory rats also became infected after injection of O. moubata infected material [42]. Later reports confirmed the infection by injection of T. cruzi infected O. venezuelensis, O. turicata, O. furcosus, and O. moubata ticks into rodents [35,37,39,47].
- Trypanosoma theileri and T. theileri-like
In 1973, Burgdorfer and colleagues unsuccessfully attempted to transmit T. theileri by injecting infected hard tick material collected from cattle into rodents [48]. On the other hand, Krinsky and Burgdorfer (1976) showed that T. theileri can be transmitted by injecting O. moubata infected material [93], as Shastri and Deshpande (1981) showed that injection of T. theleiri H. a. anatolicum infected material causes infection in cattle [49]. In 1986, Morzaria et al. reproduced the work of Shastri and Deshpande, but could not demonstrate the transmission of T. theileri-like by this route. Latif and colleagues recently succeeded in infecting cattle by injecting H. a. anatolicum infected material [117, 118].
- Other Trypanosoma
As early as 1924, Yorke and Macfie attempted to transmit T. rhodesiense to rodents by injecting them with infected O. crossi material [45]. In 1948, Packchanian did not succeed in infecting animals with T. brucei using the same route [38]. Finally, in 1976, Taylor-Lewis did not succeed in infecting rodents by injecting H. a. excavatum-T. lewisi infected material [126].
3.4.4. Infection after Ingestion or Injection with Leishmania- Infected Ticks Matertial
Several studies reported the presence of L. infantum in R. sanguineus and I. ricinus [57,65,67,74,99] and the transmission of other protozoa (Hepatozoon canis) in dogs after ticks ingestion [134]. Six studies experimentally investigated the transmission of Leishmania via the oral route and the infective capacity of injecting material from infected ticks [50,51,52,71,73,74,75]. Of these, five investigated infection by injecting, and one examined oral transmission. There were five confirmed infections in these seven laboratory reports (four after injection and one after ingestion). All these studies focused exclusively on ticks of the Ixodidae family, specifically R. sanguineus, which were infected with various Leishmania species and fed on rodents or dogs.
- Leishmania infantum and Leishmania chagasi (Syn: L. infantum)
In 2010, Dantas-Torres et al. reported an experimental infection of L. infantum by injecting infected R. sanguineus material [74], which was also supported by the work of Coutinho and colleagues on dog ticks infected with L. chagasi (syn: L. infantum) in rodents [71].
- Leishmania
In 1984, McKenzie studied the infection of two hunting dogs, crossbred between a German Shepherd and a Labrador Retriever, by injecting them with material from crushed, infected R. sanguineus ticks. Leishmania sp. was detected by several diagnostic methods in the recipient dogs, indicating the possibility of infection through the injection of tick infected material [52].
3.5. Transstadial Transmission in Ticks
The ability of a microorganism to survive the shedding process and persist for an extended period within the tick is crucial for successful transstadial transmission. Weilgama reported the transstadial transmission of the parasite Trypanosoma thylacis in the native tick I. tasmani and then to the Australian short-nosed bandicoot (Isoodon macrourus) [135].
3.5.1. Transstadial transmission of Trypanosoma
Shastri and Deshpande (1981) observed the transstadial passage of T. theileri in H. a. anatolicum ticks. Larvae and nymphs feeding on infected calves were found to be positive after passage to the adult stage. However, their vectorial competence was not confirmed [49]. The study by Latif et al. (2004) also reported the transstadial passage of T. theireri in ticks of the genus Hyalomma [124].
3.5.2. Transstadial Transmission of Leishmania
Transstadial transmission of L. infantum into R. sanguineus has been documented since the 1930s [32]. Recent work again provides evidence of the persistence of L. infantum kDNA in R. sanguineus nymphs and adults that had fed on infected dogs before molting [61]. However, the presence of the parasites could not be confirmed by microscopic examination or culture of infected tick material [61].
3.6. Transovarial Passage
3.6.1. Transovarial Passage of Trypanosoma
The detection of flagellates and Trypanosomes in tick ovary was reported as early as 1961 [48,93,136]. Therefore, a transovarial transmission has been proposed as an alternative route for maintaining the parasite in the wild. The first experimental reports describing the transovarial transmission of Trypanosoma in ticks are the work of Mayer and Rico-Lima in Germany, who worked with T. cruzi in the early 1900s [41]. In 1972, Lapierre attempted to reproduce Mayer's work but could not detect transovarian transmission in O. moubata [39]. One year later, Burgdorfer et al. (1973) also failed to observe transovarian transmission in Rhipiciphalus hard ticks [48]. To demonstrate the likelihood of T. theileri transovarian transmission, Shastri and Deshpande (1981) conducted an experiment on H. a. anatolicum but were unable to yield a conclusive outcome [41]. It wasn't until 2004 that a laboratory study provided evidence of pathogenic trypanosomes being transmitted through the ovaries of ticks [124]. Latif et al. discussed the transovarial transmission of T. theileri in H. a. anatolicum. Their findings implied that the transmission of T. theileri in ticks was only sporadic. Subsequently, Vergne et al. tried to establish ovarian transmission in R. sanguineus without conclusive evidence collected [137].
3.6.2. Transovarial passage of Leishmania
Recent reports have confirmed the transovarial passage of Leishmania in R. sanguineus, but McKenzie first pointed out this mode in the early 1980s [52]. Experimental studies by Paz et al. (2010) and Dantas-Torres and colleagues (2010, 2011) demonstrated the presence of L. chagasi and L. infantum kDNA, respectively, in R. sanguineus larvae four months after experimental infection of females [61,75,113]. Coutinho et al. (2005) did not confirm the transovarial passage of L. chagasi (syn L. infantum) [71] unlike Dabaghmanesh et al. (2016), who reported the occurrence of such mechanism [72]. Finally, Bilgiç et al. failed to demonstrate L. major transovarial passage [138].
3.7. Other Circumstantial Evidence
Significative associations between tick infestation prevalence and the presence of anti-Leishmania antibodies in dogs in endemic areas for canine Leishmaniosis are reported [73,111,139,140,141]. According to Paz et al. (2010), the prevalence of R. sanguineus infestation was significantly higher (p = 0.04) among seropositive dogs (38.5%) compared with their seronegative counterparts (29.0%). The probability of seropositivity for Leishmania is 1.5 times higher in tick-infested dogs than in non-infested animals [61]. Such findings were corroborated by Bernardino et al. (2020) and Nakkoud et al. (2022) [140,141]. On the contrary, the odds infection ratio does not significantly differ between non-infested and R. sanguineus-infested dogs in the study of Paz et al. (2013) [111].
Finally, the almost perfect correlation recorded between Leishmania detection in dogs by fine-needle aspiration biopsy of the lymph node and the detection of Leishmania in the tick’s intestine by immunohistochemistry is additional circumstantial evidence [142].
4. Meta-analyses Results
4.1. Field Detection
4.1.1. Overview of the Meta-Analysis
For the field studies, we identified 42 publications for meta-analysis and meta-regression (Figure 3). The use of a random-effects model is justified by the P and I2 statistics, which showed significantly high heterogeneity (X2=3551.8296 and I2=98.99%, p=0.0001; Figure 3). We identified publication bias in the selected studies based on visual inspection of the graph's asymmetry (Figure 4), which is confirmed by Egger's test (t= 4.15, p= 0.0002) (Supplementary File, Table S1). The elimination, one by one or simultaneously, of the four publications that show bias (Figure 4) did not affect the cumulative prevalence recorded.
The 42 studies selected included approximately 12,000 ticks collected worldwide. However, in some publications, it was not possible to determine the exact number of ticks used for the experiment. Therefore, the data we used for our analysis was the number of pools, which may overestimate the prevalence. Our results depicted an overall cumulative prevalence for Trypanosomatidae detection of 15.48% (95% CI: 7.99-24.61%) (Figure 3).
We then examined factors associated with the detection of Leishmania and Trypanosoma having medical/veterinary interest in ticks. Factors examined included the sampling decade, study area (continent), animal host (family), tick genus or species, the detection method, parasite species, and location in the tick’s organs (Table 3). We recorded high heterogeneity in all subgroups; therefore, the pooled seroprevalence estimate for each subgroup was calculated using the random-effects model.
4.1.2. Detection Method
Molecular biology yielded the highest detection rates, reaching 17.55% (with a confidence interval of 9.31–27.44%), surpassing the rates detected by microscopic methods, which stand at a mere 0.1% (confidence interval: 0–0.23%). Despite the specificity of microscopy in identifying Trypanosomatidae [16,143] they demand skilled personnel for accurate parasite detection. The efficacy of these methods varies with the type of sample and generally falls short of the sensitivity and specificity offered by PCR and cell culture methods [144]. Due to their superior sensitivity and specificity, molecular techniques are regarded as more effective [16,145].
4.1.3. Temporal Analysis
There were statistically significant differences across the collection decade (Table 3). The highest annual prevalence recorded was 14.92% (95% CI: 7.7-23.77%, 518/3043) between 2011 and 2019, and the lowest at 0.29% (95% CI: 0.16-0.45%, 19/6470) was reported before the 2000s. This increasing prevalence trend since 2000 is likely due to advances in the performance of molecular detection methods [74,125] and/or the impacts of climate change on vector populations and behaviors [146,147].
4.1.4. Ticks Geographical Origin
We also assessed differences according to geographical origin (continent) (Table 3). The highest prevalence, 25.17% (95% CI: 14.27-37.83%, 626/1986) was recorded in South America, while the lowest was in Africa (p = 0.0056). The cumulative prevalence statistics by host animal on which ticks were collected, displayed a significant difference (p=0.0012) between animal hosts, with the highest cumulative prevalence being on Canidae (27.92% [14.51-43.57%], 595/2184).
4.1.5. Tick’s Genus
The prevalence of tick infection significantly varies according to the tick’s taxonomic status (p=0.0001), from 19.06% (CI: 9.64-30.39%, 668/3271) in Rhipicephalus to 8.78% (95% CI: 0-29.41%, 27/328) in Amblyomma or 1.91% and 1.20% in Ixodes and Hyalomma, respectively, to 0.0% in Dermacentor and Haemaphysalis (Table 3).
4.1.6. Detection Method, Genus and Organ Location
We found statistical differences according to the detection method (p>0.05). The prevalence was higher at 17.55% (95% CI: 9.31, 27.44) with molecular methods and lowest at 0.1% (95% CI: 0-0.23) using microscopy. Moreover, the infection rate was significantly higher (p= 0.0046) at 18.87% [9.12-30.75] for Leishmania than for the Trypanosoma genus at 4.62% [1.14-9.79]. Infection rate also varied significantly (p=0.000) by organ in ticks: the highest rate is 74.32% (95% CI: 28.24-99.99, 164/209) in the digestive tract, followed by 44.48% [37.27-52.53] in the ovaries, 31.79% [23.31-40.91%] in the salivary glands, and the lowest rate of infection is 0.29% [95% CI: 0.16-0.45%] in the hemolymph (Table 3).
4.2. Experimental Studies
For the experimental studies, 37 publications were selected. Data were extracted from these research papers. Quantitative data such, as the number of ticks used for experimental infections, were not considered in the meta-analysis. They were transformed into semi-quantitative ones (presence or absence of parasites following experimental infection or transmission). Since no heterogeneity was detected in the data set, the common-effect model was chosen for the meta-analysis.
4.2.1. Ingestion after Blood Feeding on an Infected Host
The meta-analysis was performed on 36 studies. The analysis showed that the presence of parasites in ticks after their blood meal was reported in 91% [72-100%] of the studies (Supplementary File, Figure S1). The factors associated with the detection of Leishmania and Trypanosoma in the blood meal were analysed taking into account each actor involved in the transmission cycle, tick, animal host and parasite detected (Table 4). No heterogeneity was detected in the subgroups. Rate estimates pooled for each subgroup were calculated using a common-effects model.
- Ticks Family and Genus
For tick families, positivity rates were 100% (95% CI: 81.65-100) for Argasidae and 78.01 (95% CI: 48.9-98.84%) for Ixodidae, with no statistical difference according to tick family (p > 0.05). Slight variations were recorded for the genus, with no statistically significant difference. For the genera Ornithodoros and Amblyomma, all publications reported positive detection after the blood meal; for the genera Rhipicephalus and Hyalomma, detection rates were 74.57% and 82.18%, respectively. A large variation in prevalence is recorded at the species level, mainly related to the low number of studies that were published (Table 4).
- Parasite genus
For parasite genera, the highest detection rate is 99.11% (95% CI: 80.73-100) for Trypanosoma, whereas Leishmania is detected in 69.10% (95% CI: 62.64-97.17) of the studies. However, this difference is not statistically significant (P > 0.05) (Table 4).
- Host family
No statistical differences were recorded (Table 4).
4.2.2. Transmission by injection of tick-infected material
The meta-analysis includes 22 scientific papers. The common-effects model was used to analyze associated factors. About 72% (95% CI: 42-95%) of articles show positive transmission results. In 1984, McKenzie studied the infection of two hunting dogs, crossbred between a German shepherd and a Labrador retriever, by injecting them with material from crushed, infected R. sanguineus ticks by injection of ticks infected material (Supplementary File Figure S2).
- Tick's family, genus and species
The infection rate elicited via the injection of Argasidae-infected material is statistically higher than that of Ixodidae-infected ones (rate of 85.25% vs. 55.23%). The highest rate, 85.25%, was recorded for Ornithodoros, followed by Rhipicephalus and Hyalomma. However, if all these observations are not statistically significant, they interestingly point to some specificity according to the tick genus (Table 4).
- Parasite's genus and species
No significant difference was recorded within parasite genera and species subgroups (p > 0.05). Although we observed a generally higher infection rate with T. cruzi, T. theileri, Leishmania. sp., L. chagasi (Syn L. infantum), and L. infantum infected tick material.
- Host family
These analyses disclosed that tick material collected from infected Camelidae was more likely to initiate infection when injected into a noninfected recipient and that Canidae appears to be more susceptible to infection when injected with infected tick material.
4.2.3. Transmission Through Tick’s Blood Feeding on a Non-Infected Host
Tick-borne pathogen transmission can occur via mechanical or biological means. In mechanical transmission, ticks act as carriers, transferring pathogens between hosts without mandatory pathogen development within the tick. In contrast, biological transmission involves the pathogen undergoing necessary biological changes or replication within the tick, completing part of its life cycle before infecting the next host. We performed a meta-analysis on data extracted from 21 publications dealing with experimental transmission by ticks via blood feeding (Table 4). This analysis showed that 34% (95% CI: 8-64%) of the studies confirmed transmission by ticks of medical and veterinary interest Trypanosomatidae, for which data can be gathered during our study (Supplementary Figure S3). The common-effects model was used to analyze factors associated with experimental transmission.
- Ticks family
The Ixodidae appeared to be more able to transmit Trypanosomatidae than Argasidae, with rates of 34.33% (95% CI: 0-94%) and 23.7% (95% CI: 0-66.4%), respectively (Table 4). Regarding the genus, minor variations were recorded without significant differences (p > 0.05). Regarding tick species, R. sanguineus, O. lahorensis, O. crossi, and H. a. anatolicum seemed more able to transmit Trypanosomatidae parasites of medical and veterinary interest. However, the sample size is too small to get insight into the statistical significance of these observations.
- Parasite genus
Although the success rate of transmission attempts is higher with Leishmania (82.18% CI:35.16-100%) than with Trypanosoma (12.42% CI: 0-70.74%), the meta-analysis does not record a statistically significant difference between Leishmania and Trypanosoma (p>0.05) (Table 4).
- Donor and receiver host family
Surprisingly, the donor host appears to be a factor influencing the subsequent transmission of the pathogen during tick blood feeding (p<0.05) Table 4. The infection rate for ticks varied from 100% to 95% (CI: 21.26-100%) in Camelidae, to 59.68% for Canidae and 50.0% for Bovidae. The eight studies that focused on ticks collected on infected rodents reported no transmission. The host recipient also showed heterogeneity, with Canidae and Bovidae being more frequently infected at a rate of 72.14% [2.45-100%] than rodents (8.01%, [0-41.38%], when infected and allowed to be fed.
4.2.4. Vertical Transmission
Our analysis includes only 13 studies focusing on the vertical transmission of Trypanosomatidae by ticks. The findings revealed that 55% presented evidence supporting the vertical transmission of parasites within tick populations, as detailed in the supplementary file (Supplementary Figure S4). The factors associated with this transmission were evaluated using the common-effect model. Despite the small sample size, vertical transmission is recorded at 61.27% [22.41-94.58] for ticks of the Ixodidae family and 0% for the Argasidae, although only one study deals with this issue. No significant difference exists according to genus and species (Table 4). Concerning genus and parasite species, it seems that parasites belonging to the Leishmania genus are better adapted to vertical transmission (82.18 [35.16-100]) than those of the Trypanosoma genus (12.42 [0-70.74]). Parasites from the L. donovani complex (L. donovani or L. infantum) appear to be adapted to vertical transmission. Also, the host family from which ticks feed plays a significant role in variability (p=0.0425). Ticks feeding on dogs (Canidae) have the highest vertical transmission rate at 92.94% [46.20-100%], followed by those feeding on Bovida at 27.86% [0-97.55], and lastly, rodents with no vertical transmission of Trypanosomatidae in ticks following blood meal on rodent infected host.
5. Discussion
Biologists have been particularly interested in insects, vectors of Trypanosomatidae, because of the significant infectious impact of these parasite protists on animal species, including humans [147]. While almost all Trypanosomatidae are transmitted by insects, a single publication reported the case of an avian trypanosome vectorized by a non-traditional vector, an arachnid belonging to the subclass of acarians [148]. This anomaly prompted further investigations into the presence of Trypanosomatids within ticks, as evidenced by the early work of Novy et al. (1907), who identified Trypanosomes in ticks [149]. This has led to an increased focus on understanding the potential role of ticks in harboring and transmitting these pathogenic protozoans.
To determine the prevalence of Trypanosomatidae of medical and veterinary interest in wild-caught ticks from endemic areas, both molecular (PCR, qPCR, PCR-HRM) and parasitological/immunological (microscopic examination, IHC) methods were used. Molecular methods that amplify genomic DNA, which can last after parasite death, are more sensitive than parasitological/immunological methods which target living parasites or their proteic determinants that are rapidly degrade upon parasite death. Statistical differences in detection rates were observed according to the method employed. Specifically, the highest prevalence (17.55%) was recorded with molecular methods compared to microscopic detection of parasites (0.1%). The discovery of DNA or parasites in the digestive system of blood-fed ticks collected from hosts has limited predictive value in inferring a vectorial role. The vast majority of publications included in our systematic review and meta-analysis dealt with ticks collected directly from hosts, focusing on detecting pathogens from host blood deposited in tick bodies. DNA detection alone is insufficient, especially in the case of ticks, due to their digestive capacity and other metabolic peculiarities. To further analyze the prevalence of Trypanosomatidae in field-collected tick samples, it will be necessary to find the parasites themselves in these ticks and/or detect parasite-specific mRNA, which is a better indicator of parasite survival in the digestive tract and other tick organs.
Data on tick infection disclose that the detection of Trypanosomatidae of medical or veterinary interest in ticks varies between 0.01-1.26% to 14.27-37.83% depending on countries or continents (Table 3). In many studies, pools are used to ascertain the prevalence, which greatly overestimates the results. These provide the first clues on the high contact frequency between infected hosts and ticks, a crucial element to engage transmission by arthropod vectors. Overall, in many places where infections with Trypanosomatidae are common, there's no available information on the infection rate by these pathogenic agents in their proven vectors and/or in ticks. For example, the Mediterranean area has a high rate of visceral Leishmaniasis [150], but no information on the carriage of Leishmania by ticks in countries like Egypt, Libya, and Morocco is available. Latin America and the Caribbean are affected by Chagas disease, caused by T. cruzi [151], but no data on its presence in ticks is available, which makes it difficult to draw a global analysis of the seasonal activity patterns of ticks in relation to the incidence of Trypanosomatidae pathogen infections in hosts but also on the geographical overlap between tick populations and human or animal infection by Trypanosomatidae pathogens.
Our systematic review and meta-analysis disclose that if 996 species (774 hard ticks and 221 soft ticks) are described worldwide [3,4], with 25 species acting as vectors of major diseases, the presence of Trypanosomatidae pathogens having medical or veterinary interests was investigated on twenty tick species. In South America, 137 species of hard ticks, from 5 genera and 87 species of soft ticks are reported [152,153]. All studies we collected focused on the Ixodidae family, with no field data available on the soft ticks of the Argasidae family. The detection rate in members of the Rhipicephalus genus is the highest at 17.49%, followed by Amblyomma, Hyalomma, and Ixodes with infection rates of 11.47%, 2.68%, and 1.87%, respectively. Ticks belonging to the Dermacentor and Haemaphysali genera were negative. Variability in field studies concerning the detection of Trypanosomatidae in ticks can often be attributed to a host effect. Specifically, ticks harvested from Canidae, which are alost exclusively non questing ones and belong to the Rhipicephalus genus, exhibit a higher pathogens detection rate compared to ticks collected from other mammals (P=0.0007). This discrepancy may be due in part to the close relationship between dogs and humans and their heightened susceptibility to Trypanosoma cruzi, which may increase the risk of tick infection [154]. Additionally, dogs serve as a significant reservoir for several Leishmania species, such as L. infantum, L. peruviana, and L. donovani [155]. The extent of the geographical distribution of the ticks belonging to the genus Rhipicephalus, the broad range of their animal hosts, their known vector competence, and their diverse morphology provide arguments for this meta-analysis on their potential role in the transmission of Trypanosomatidae [156]. R. sanguineus is a typical representative of this genus in so far as the close association of this tick species with domestic dogs, which constitute a known reservoir for Leishmania, signs its likely involvement in the persistence and transmission of these parasites in natural habitats [157].
The concept of vector competence is defined as the innate ability of an arthropod to harbor and transmit microbial agents [158,159]. Laboratory studies were conducted to establish whether ticks can acquire and maintain the infection after feeding on infected hosts. Therefore, establishing the vector capacity of a tick consists of confirming its ability to become infected during a blood meal on a host, facilitating the multiplication of the pathogen before its subsequent transmission through saliva, and maintaining the pathogen throughout the tick development stages for potential vertical transmission [160]. Regarding parasite acquisition, the meta-analysis reveals that in experimental settings, tick infection after blood feeding is efficient, with 31 successes out of 37 attempts (Table 4). Although no significant factors were associated with parasite acquisition from an infected host (donor host), the analysis does indicate some tendencies that warrant discussion. Firstly, Argasidae ticks are likely to become infected compared to Ixodidae ticks when feeding on an infected host (P=0.0976). The host’s nature does not appear to affect tick infection rates. However, the type of parasite does influence tick infection rates, with a higher rate of infection observed when ticks feed on a Trypanosoma-infected host compared to a Leishmania-infected host; tick infection is more efficient when they take a blood meal from a host infected with Trypanosoma (P=0.0794) Table 4. Trypanosoma cruzi and T. evansi are particularly likely to infect ticks following blood feeding. These observations may be related to the intrinsic ability of Trypanosomes to multiply in the blood of the infected host, making them more available to ticks during the blood-feeding process. Additionally, such disparity in infection rates might be attributed to the feeding behaviors differing for the two tick families: Ixodidae ticks attach to the hosts' skin and suck blood slowly over several days, whereas Argasidae ticks rapidly ingest large volumes of blood in a short time (20–70 minutes), facilitating a more efficient uptake of infectious agents, especially under experimental conditions [161]. The rapid feeding behavior of Argasidae ticks makes them more efficient at acquiring blood-circulating parasites like T. brucei or T. evansi, compared to the long-feeding Ixodidae ticks, which may be less effective at acquiring tissue-located parasites. Finally, the prolonged feeding process of Ixodidae ticks, which lasts up to two weeks, subjects engorged pathogens to the full spectrum of host defense mechanisms, including specific acquired immunity that may impact parasites of the Trypanosomatidae family’s survival [162]. Information gathered on the efficiency of infection through the injection of contaminated tick material provides insight into the presence of infectious parasitic stages in infected ticks. Our meta-analysis did not identify any significant factors related to the donor or receiver host, ticks, or parasite identity that play a role in the transmission of Trypanosomatidae by ticks. While Argasidae ticks appear to be more efficient in such transmission than Ixodidae ticks (7 successes out of 9 attempts compared to 7 successes out of 13 attempts), these differences are not statistically significant (P=0.2884). Because ticks feed only once at each life stage, vertical transmission of pathogens is an important factor to consider when addressing the vectorial status of ticks for Trypanosomatidae parasites. Our literature survey indicates that vertical transmission is detected in experimental settings and only in Ixodidae ticks. Although not statistically significant, Leishmania appears to be more likely to be vertically transmitted in R. sanguineus than Trypanosoma species (P=0.0634). Interestingly, protozoan of the Babesia genus can undergo transovarial transmission in ticks, which suggest potential similar behavior for protozoan of the trypanosomatidae family [163]. However, the limited number of studies on vertical transmission prevents further discussion on the influence of tick identity or parasite species on vertical transmission.
Trypanosoma evansi and T. vivax are mechanically transmitted pathogens, meaning they are transmitted from host to host without biological replication within their vector. This transmission mode does not align well with the feeding habits of ticks, except in cases of interrupted feeding [164]. There is no available data on T. vivax. However, a relatively good success rate (3 successes out of 6 attempts) was recorded for transmitting T. evansi via tick blood feeding. Therefore, the transmission of T. evansi or T. vivax after interrupted blood feeding should be more thoroughly investigated.
Salivarian Trypanosomes, such as T. brucei, T. cruzi, T. evansi, T. congolense, T. vivax, and Leishmania parasites, are transmitted through blood-feeding by the injection or regurgitation of saliva during the feeding process. Stercocarian trypanosomes such as T. cruzi and T. rangeli infect their host are deposited on the host's skin during blood-feeding and subsequently infect the host through the scratching of the infected blood onto the host's mucosa. Infection by ingestion is also documented for T. cruzi [165]. Considering this aspect and examining the results of the meta-analysis, we noticed that attempts to transmit Trypanosomatidae pathogens by blood-feeding of infected ticks are more successful with Salivarian Trypanosomatidae pathogens, such as T. evansi, T. lewisi, and T. theileiri, than with Stercorarian Trypanosomatidae pathogens. Specifically, there were five attempts for two successes for Leishmania, 11 attempts for five successes for Salivarian Trypanosomes, and 6 attempts for one success for the Stercocarian T. cruzi Trypanosome. Data collected indicate a general trend of better infection rates for Salivarian Trypanosomatidae during tick blood meals. However, these differences are not statistically significant, so we cannot draw a definitive conclusion about the impact of the transmission route.
Overall, this systematic review and meta-analysis provide an updated overview of the vector status of ticks for Trypanosomatidae. This updated perspective reveals that partial information is available regarding the presence of Trypanosomatidae infecting humans and animals of veterinary importance in field-collected specimens or experimental studies aimed at probing the vector competence of ticks. Specifically, data are available for 20 species out of 774 recognized hard ticks and 221 soft ticks. Of the 23 Leishmania and 11 Trypanosoma species of medical or veterinary interest, we gathered information on 6 species belonging to the Leishmania and 9 to the Trypanosoma genus. Surprisingly, we did collect data on the presence of Trypanosomes involved in human African Trypanosomiasis (T. b. gambiense and T. b. rhodesiense) in field-collected ticks. Altogether, we could not provide conclusive quantitative evidence regarding the vectorial role of ticks for Leishmania and Trypanosoma parasites in medical or veterinary contexts. However, for Rhipcephalus sanguineus the documentation of the presence of Trypanaosomatidae infecting humans and animals is the larger one (21 studies), the capacity to engorge parasites during blood feeding on an infected host has been successfully attempted (11 successes over 15 experiments), and the presence of the infective parasite stage has been probed through the injection of tick infected material (7 attempts, and 5 successes), as well as re-transmission through infected tick blood feeding (6 attempts and 3 successes). Interestingly and importantly, data on vertical transmission have also been collected (9 attempts 7 successes). The majority of these collected data are for L. infantum, but altogether, these data argue for the vectorial competence of R. sanguineus, which needs now to be more thoroughly demonstrated using advanced molecular methods in field-collected specimens and with additional experimental information. In addition, our analysis suggests that the presence in ticks of parasites from the Trypanosomatidae family having medical or veterinary interests is not infrequent and that ticks of the Argasidae family might play a role in the transmission of Trypanosomatidae. The exact role of ticks in sustaining both the parasitic developmental and epidemiological cycles of Trypanosomatidae requires further investigation and continuous scrutiny.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization and methodology, T.K. and D.S.; software, B.M.; validation, T.K. and D.S.; formal analysis, T.K., B.M. and D.S.; investigation, T.K.; resources, N.E. and D.S.; data curation, T.K.; writing—original draft preparation, T.K., B.M. and D.S.; writing—review and editing, N.E., P.H., B.M., D.S. and T.K.; visualization, T.K. and D.S.; supervision, D.S.; project administration, N.E. and D.S.; funding acquisition, N.E. and D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by LeiSHeild-RISE MATI project, Grant Agreement N°778298.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and Supplementary Material.
Acknowledgments
We acknowledge the French Development Research Institute (IRD) library who provided us with the various articles, thesis and books and even from other libraries. Additionally, I extend my gratitude to UMR INTERTRYP, IRD, Montpellier, France, for welcoming me into their unit.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Brites-Neto, J.; Duarte, K.M.R.; Martins, T.F. Tick-Borne Infections in Human and Animal Population Worldwide. Vet. world 2015, 8, 301. [Google Scholar] [CrossRef]
- Rajakaruna, R.S.; Eremeeva, M.E. Eco-Epidemiology of Tick-Borne Pathogens. One Heal. Human, Anim. Environ. Triad 2023, 325. [Google Scholar] [CrossRef]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Pena, A.; Horak, I.G.; Shao, R.F.; Barker, S.C. The Argasidae, Ixodidae and Nuttalliellidae (Acari: Ixodida) of the World: A List of Valid Species Names. Zootaxa 2010, 1–28. [Google Scholar] [CrossRef]
- Mans, B.J.; Featherston, J.; Kvas, M.; Pillay, K.-A.; Klerk, D.G. de; Pienaar, R.; Castro, M.H. de; Schwan, T.G.; Lopez, J.E.; Teel, P. Argasid and Ixodid Systematics: Implications for Soft Tick Evolution and Systematics, with a New Argasid Species List. Ticks Tick. Borne. Dis. 2019, 10, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J. Paradigms in Tick Evolution. Trends Parasitol. 2023, 39, 475–486. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, P. Haemoprotozoa: Making Biological Sense of Molecular Phylogenies. Int. J. Parasitol. Wildl. 2017, 6, 241–256. [Google Scholar] [CrossRef]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef] [PubMed]
- Brun, R.; Blum, J.; Chappuis, F.; Burri, C. Human African Trypanosomiasis. Lancet 2010, 375, 148–159. [Google Scholar] [CrossRef]
- Kwakye-Nuako, G.; Mosore, M.-T.; Boakye, D.; Bates, P.A. Description, Biology, and Medical Significance of Leishmania (Mundinia) chancei n. sp.(Kinetoplastea: Trypanosomatidae) from Ghana and Leishmania (Mundinia) procaviensis n. sp.(Kinetoplastea: Trypanosomatidae) from Namibia. J. Parasitol. 2023, 109, 43–50. [Google Scholar] [CrossRef]
- Moncayo, Á.; Silveira, A.C. Current Epidemiological Trends of Chagas Disease in Latin America and Future Challenges: Epidemiology, Surveillance, and Health Policies. Am. Trypanos. Chagas Dis. 2017, 59–88. [Google Scholar] [CrossRef]
- Sereno, D. Leishmania (Mundinia) spp.: From Description to Emergence as New Human and Animal Leishmania Pathogens. New Microbes New Infect. 2019, 30, 100540. [Google Scholar] [CrossRef]
- Bayão T de S, Cupertino M do C, Mayers NAJ, Siqueira-Batista R. 2020. A systematic review of the diagnostic aspects and use of Trypanosoma rangeli as an immunogen for Trypanosoma cruzi infection. Rev Soc Bras Med Trop 53:e20190608. [CrossRef]
- Truc, P.; Büscher, P.; Cuny, G.; Gonzatti, M.I.; Jannin, J.; Joshi, P.; Juyal, P.; Lun, Z.-R.; Mattioli, R.; Pays, E. Atypical Human Infections by Animal Trypanosomes. PLoS Negl. Trop. Dis. 2013, 7, e2256. [Google Scholar] [CrossRef]
- Maurıcio, I.L.; Stothard, J.R.; Miles, M.A. The Strange Case of Leishmania chagasi. Parasitol. today 2000, 16, 188–189. [Google Scholar] [CrossRef] [PubMed]
- Desquesnes, M.; Gonzatti, M.; Sazmand, A.; Thévenon, S.; Bossard, G.; Boulangé, A.; Gimonneau, G.; Truc, P.; Herder, S.; Ravel, S. A Review on the Diagnosis of Animal Trypanosomoses. Parasit. Vectors 2022, 15, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Desquesnes, M.; Sazmand, A.; Gonzatti, M.; Boulangé, A.; Bossard, G.; Thévenon, S.; Gimonneau, G.; Truc, P.; Herder, S.; Ravel, S. Diagnosis of Animal Trypanosomoses: Proper Use of Current Tools and Future Prospects. Parasit. Vectors 2022, 15, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Itard, J.; Morel, P.C. Manual of Tropical Veterinary Parasitology; CAB International, 1989; ISBN 085198584X. [Google Scholar]
- Doherty ML, Windle H, Voorheis HP, Larkin H, Casey M, Clery D, Murray M. 1993. Clinical disease associated with Trypanosoma theileri infection in a calf in Ireland. Vet Rec 132:653–656. [CrossRef]
- Monzon, C.M.; Mancebo, O.A.; Jara, G.; Hoyos, C.B. Trypanosoma Theileri (Laveran, 1902) En Bovino de La Provincia de Formosa: Aislamiento, Cultivo y Alteraciones Hemáticas. 1993.
- Seifi HA. 1995. Clinical trypanosomiasis due to Trypanosoma theileri in a cow in Iran. [CrossRef]
- Radostits OM, Blood DC, Gay CC. Veterinary medicine. A textbook of the diseases of cattle, sheep, pigs, goats and horses. 8th Edition, Bailliere, Tindall, London, 1994, 1015-1026.
- Madeira, M.F.; Almeida, A.B.P.F.; Barros, J.H.S.; Oliveira, T.S.F.; Sousa, V.R.F.; Alves, A.S.; Miranda, L.F.C.; Schubach, A.O.; Marzochi, M.C.A. Trypanosoma Caninum, a New Parasite Described in Dogs in Brazil: Aspects of Natural Infection. J. Parasitol. 2014, 100, 231–234. [Google Scholar] [CrossRef]
- de Sousa, M.A. On Opportunist Infections by Trypanosoma Lewisi in Humans and Its Differential Diagnosis from T. Cruzi and T. Rangeli. Parasitol. Res. 2014, 113, 4471–4475. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.; Jenner, C.; Pender, J.; Rogers, D.; Slingenbergh, J.; Wint, W. The Programme against African Trypanosomiasis Information System (PAATIS). African Trypanos. 2002, 11–24. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Ticks as Vectors of Leishmania Parasites. Trends Parasitol. 2011. [Google Scholar] [CrossRef]
- Koual, R.; Buysse, M.; Grillet, J.; et al. Phylogenetic Evidence for a Clade of Tick-Associated Trypanosomes. Parasites and vector. 2023. [Google Scholar] [CrossRef]
- McGhee, R.B.; Cosgrove, W.B. Biology and Physiology of the Lower Trypanosomatidae. Microbiol. Rev. 1980, 44, 140–173. [Google Scholar] [CrossRef] [PubMed]
- Becvar, T.; Vojtkova, B.; Siriyasatien, P.; Votypka, J.; Modry, D.; Jahn, P.; Bates, P.; Carpenter, S.; Volf, P.; Sadlova, J. Experimental Transmission of (Mundinia) Parasites by Biting Midges (Diptera: Ceratopogonidae). PLoS Pathog. 2021, 17, e1009654. [Google Scholar] [CrossRef] [PubMed]
- Hoare, C.A. The Trypanosomes of Mammals. A Zoological Monograph. Trypanos. mammals. A Zool. Monogr. 1972. [Google Scholar]
- Wenyon, C.M. The Transmission of Leishmania Infections: A Review. Trans. R. Soc. Trop. Med. Hyg. 1932, 25, 319–348. [Google Scholar] [CrossRef]
- O’Farrell, W.R. Hereditary Infection, with Special Reference to Its Occurrence in Hyalomma aegyptium Infected with Crithidia Hyalommae. Ann. Trop. Med. Parasitol. 1913, 7, 545–562. [Google Scholar] [CrossRef]
- Blanc, G.; Caminopetros, J. Transmission of Mediterranean EA by a Tick. Compte Rendu l’Academie des Sci. 1930, 191. [Google Scholar]
- Swellengrebel, N.H.; Strickland, C. The Development of Trypanosoma lewisi Outside the Vertebrate Host1. Parasitology 1910, 3, 360–389. [Google Scholar] [CrossRef]
- Brumpt, E. Le Trypanosoma Cruzi Évolue Chez Conorhinus Megistus, Cimex Lectularius, Cimex Boueti et Ornithodorus Moubata. Cycle Évolutif de Ce Parasite. Bull. la Société Pathol. Exot. 1912, 5, 360–367. [Google Scholar]
- Wheeler, C.M. Experimental Infection of Ornithodoros turicata (Duges) with a Brazilian Strain of Trypanosoma cruzi Chagas. Proc. Soc. Exp. Biol. Med. 1938, 38, 191–193. [Google Scholar] [CrossRef]
- Pifano, F. Amblyomma longyrostrum Infected in Nature by T. cruzi. Gac. Med. Caracas 1941, 48. [Google Scholar]
- Mazzotti, L.; Osorio, M.T. Experiments in the Transmission of T. cruzi to Four Species of Ornithodoros. Rev. Inst. Salubr. Enferm. Trop. 1943, 4, 163–165. [Google Scholar]
- Packchanian, A.A. The Fate of Some Pathogenic Trypanosomes in Triatoma and Ornithodorus. Am. J. Trop. Med. 1948, 28. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, J. Evolution et Conservation de Trypanosoma cruzi Chez Ornithodorus moubata. Ann. Parasitol. Hum. comparée 1972, 47, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Neiva, A. Transmission of T. Cruzi by R. Sanguineus. Bras. Med. 1913, 27. [Google Scholar]
- Mayer, M.; Roca-lima, H.D. On the Behaviour of Schizotrypanum Cruzi in Warm-Blooded Animals and Arthropods. Beihefte zum Arch. fur Schiffs-und … 1914.
- Mayer, M. On the Length of Infection of Ornithodorus Moubata by S. Cruzi. Arch. fur Schiffs-und Tropenhygiene 1918, 22. [Google Scholar]
- Cross, H.E.; Patel, P.G. A Note on the Transmission of Surra by Ticks. Vet. J. 1922, 78, 469–471. [Google Scholar] [CrossRef]
- Cross, H.E. A Further Note on Surra Transmission Experiments with Tabanus Albimedius and Ticks. Trans. R. Soc. Trop. Med. Hyg. 1923, 16. [Google Scholar] [CrossRef]
- Yorke, W.; Macfie, J.W.S. Trypanosoma evansi and Ornithodorus crossi. Ann. Trop. Med. Parasitol. 1924, 18, 125–126. [Google Scholar] [CrossRef]
- Kahan, S. A Further Note on Surra Transmission Experiments with Ticks. A Furth. Note Surra Transm. Exp. with Ticks. 1925. [Google Scholar]
- Dunn, L.H. Attempts to Transmit Trypanosoma cruzi Chagas with Ticks of the Genus Ornithodoros. Am. J. Trop. Med. Hyg. 1934, s1-14, 283–289. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Schmidt, M.L.; Hoogstraal, H. Detection of Trypanosoma Theileri in Ethiopian Cattle Ticks. Acta Trop. 1973, 30, 340–346. [Google Scholar] [PubMed]
- Shastri, U. V.; Deshpande, P.D. Hyalomma anatolicum anatolicum (Koch, 1844) as a Possible Vector for Transmission of Trypanosoma theileri, Laveran, 1902 in Cattle. Vet. Parasitol. 1981, 9, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Malamos, B. Experiments with Leishmania. IV. Experiments in the Transmission of Kala-Azar by Ticks, R. Sanguineus. Arch. fur Schiffs-und Tropenhygiene 1938, 42. [Google Scholar]
- Rioux, J.-A.; Lanotte, G.; Croset, H.; Houin, R.; Guy, Y.; Dedet, J.-P. Ecologie Des Leishmanioses Dans Le Sud de La France-3.-Réceptivité Comparée de Phlebotomus ariasi Tonnoir, 1921 et Rhipicephalus turanicus Pomerancev et Matikasvili, 1940 Vis-à-Vis de Leishmania donovani (Laveran et Mesnil, 1903). Ann. Parasitol.L. Hum. Comparée 1972, 47, 147–157. [Google Scholar] [CrossRef]
- McKenzie, K.K. A Study of the Transmission of Canine Leishmaniasis by the Tick, Rhipicephalus Sanguineus, and an Ultrastructural Comparison of the Promastigote. Oklahoma State Univ. Stillwater 1984. [Google Scholar]
- Azarmi, S.; Zahraei-ramazani, A.; Mohebali, M.; Rassi, Y.; Ahmad, A. Original Article PCR Positivity of Gerbils and Their Ectoparasites for Leishmania spp . in a Hy- Perendemic Focus of Zoonotic Cutaneous Leishmaniasis in Central Iran. 2022, 16, 124–135. [Google Scholar]
- Campos, J.H.F.; Costa, F.A.L. Participação Do Rhipicephalus Sanguineus No Ciclo Infeccioso Da Leishmaniose Visceral Canina Em Teresina, Piauí, Brasil. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 297–300. [Google Scholar] [CrossRef]
- Gonçalves, L.R.; Filgueira, K.D.; Ahid, S.M.M.; Pereira, J.S.; Vale, A.M. do; Machado, R.Z.; André, M.R. Study on Coinfecting Vector-Borne Pathogens in Dogs and Ticks in Rio Grande Do Norte, Brazil. Rev. Bras. Parasitol. Veterinária 2014, 23, 407–412. [Google Scholar] [CrossRef]
- Gondard, M.; Delannoy, S.; Pinarello, V.; Aprelon, R.; Devillers, E.; Galon, C.; Pradel, J.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Upscaling the Surveillance of Tick-Borne Pathogens in the French Caribbean Islands. Pathogens 2020, 9, 1–37. [Google Scholar] [CrossRef]
- Medeiros-Silva, V.; Gurgel-Gonçalves, R.; Nitz, N.; D’Anduraim Morales, L.E.; Cruz, L.M.; Sobral, I.G.; Boité, M.C.; Ferreira, G.E.M.; Cupolillo, E.; Romero, G.A.S. Successful Isolation of Leishmania infantum from Rhipicephalus sanguineus Sensu Lato (Acari: Ixodidae) Collected from Naturally Infected Dogs. BMC Vet. Res. 2015, 11. [Google Scholar] [CrossRef]
- Millán, J.; Travaini, A.; Zanet, S.; López-Bao, J.V.; Trisciuoglio, A.; Ferroglio, E.; Rodríguez, A. Detection of Leishmania DNA in Wild Foxes and Associated Ticks in Patagonia, Argentina, 2000 Km South of Its Known Distribution Area. Parasites and Vectors 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, K.Y.; Arslan-Akveran, G.; Aydogdu, S.; Karasartova, D.; Koşar, A.; Savci, U.; Keskin, A.; Taylan-Ozkan, A. Pathogens in Ticks Collected in Israel: II. Bacteria and Protozoa Found in Rhipicephalus sanguineus Sensu Lato and Rhipicephalus turanicus. Ticks Tick. Borne. Dis. 2022, 13, 101986. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, K.Y.; Arslan-akveran, G.; Aydogdu, S.; Karasartova, D.; Kosar, N.; Semra, A.; Shacham, B.; Taylan-ozkan, A. Ticks and Tick-Borne Diseases Pathogens in Ticks Collected in Israel : I. Bacteria and Protozoa in Hyalomma aegyptium and Hyalomma dromedarii Collected from Tortoises and Camels. Ticks Tick. Borne. Dis. 2022, 13, 101866. [Google Scholar] [CrossRef] [PubMed]
- Paz, G.F.; Ribeiro, M.F.B.; Michalsky, É.M.; Da Rocha Lima, A.C.V.M.; França-Silva, J.C.; Barata, R.A.; Fortes-Dias, C.L.; Dias, E.S. Evaluation of the Vectorial Capacity of Rhipicephalus sanguineus (Acari: Ixodidae) in the Transmission of Canine Visceral Leishmaniasis. Parasitol. Res. 2010, 106, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, M.G.; Persichetti, M.F.; Serrano, L.; Altet, L.; Reale, S.; Gulotta, L.; Solano-Gallego, L. Ticks and Associated Pathogens Collected from Cats in Sicily and Calabria (Italy). Parasites and Vectors 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Jaimes, J.E.; Correa-Núñez, G.H.; Rojas-Palomino, N.; Cáceres-Rey, O. Detección de Leishmania (V) guyanensis En Ejemplares de Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) Recolectados En Pecaríes de Collar (Pecari tajacu). Biomedica 2017, 37, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, D.; Aureli, S.; Baldelli, R.; Di Francesco, A.; Tampieri, M.P.; Galuppi, R. Analisi Molecolare Di Leishmania infantum in Zecche Ixodes Ricinus Da Cani e Gatti in Italia. Vet. Ital. 2014, 50, 307–312. [Google Scholar] [CrossRef]
- Colombo, F.A.; Odorizzi, R.M.F.N.; Laurenti, M.D.; Galati, E.A.B.; Canavez, F.; Pereira-Chioccola, V.L. Detection of Leishmania (Leishmania) infantum RNA in Fleas and Ticks Collected from Naturally Infected Dogs. Parasitol. Res. 2011, 109, 267–274. [Google Scholar] [CrossRef]
- Trotta, M.; Nicetto, M.; Fogliazza, A.; Montarsi, F.; Caldin, M.; Furlanello, T.; Solano-Gallego, L. Detection of Leishmania infantum, Babesia canis, and Rickettsiae in Ticks Removed from Dogs Living in Italy. Ticks Tick. Borne. Dis. 2012, 3, 294–297. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Rossi, L.; Scroccaro, A.M.; Montarsi, F.; Caldin, M.; Furlanello, T.; Trotta, M. Detection of Leishmania infantum DNA Mainly in Rhipicephalus sanguineus Male Ticks Removed from Dogs Living in Endemic Areas of Canine Leishmaniosis. Parasit. Vectors 2012, 5, 1–6. [Google Scholar] [CrossRef]
- Viol, M.A.; Guerrero, F.D.; Oliveira, B.C.M. de; Aquino, M.C.C. de; Loiola, S.H.; Melo, G.D. de; Gomes, A.H. de S.; Kanamura, C.T.; Garcia, M.V.; Andreotti, R. Identification of Leishmania spp. Promastigotes in the Intestines, Ovaries, and Salivary Glands of Rhipicephalus sanguineus Actively Infesting Dogs. Parasitol. Res. 2016, 115, 3479–3484. [Google Scholar] [CrossRef] [PubMed]
- Viol, M.A. Considerações sobre a presença de formas promastigotas de Leishmania spp. em intestinos, ovários e glândulas salivares de carrapatos Rhipicephalus sanguineus em cães de áreas endêmicas. repositorio.unesp.br, 2015.
- Almeida, V. dos A.; Hora, T.N. da; Junior, N.F.L.; Carvalho, F.S.; Silva, A.L. da; Wenceslau, A.A.; Albuquerque, G.R.; Silva, F.L. Detection of Leishmania infantum DNA in Hamsters Infested with Ticks Collected from Naturally Infected Dogs. Rev Bras Med Vet 2016, 38, 329–333. [Google Scholar]
- Coutinho, M.T.Z.; Bueno, L.L.; Sterzik, A.; Fujiwara, R.T.; Botelho, J.R.; De Maria, M.; Genaro, O.; Linardi, P.M. Participation of Rhipicephalus sanguineus (Acari: Ixodidae) in the Epidemiology of Canine Visceral Leishmaniasis. Vet. Parasitol. 2005, 128, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Dabaghmanesh, T.; Asgari, Q.; Moemenbellah-Fard, M.D.; Soltani, A.; Azizi, K. Natural Transovarial and Transstadial Transmission of Leishmania infantum by Naïve Rhipicephalus sanguineus Ticks Blood Feeding on an Endemically Infected Dog in Shiraz, South of Iran. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 408–413. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; de Paiva-Cavalcanti, M.; Figueredo, L.A.; Melo, M.F.; da Silva, F.J.; da Silva, A.L.; Almeida, E.L.; Brandão-Filho, S.P. Cutaneous and Visceral Leishmaniosis in Dogs from a Rural Community in Northeastern Brazil. Vet. Parasitol. 2010, 170, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Lorusso, V.; Testini, G.; De Paiva-Cavalcanti, M.; Figueredo, L.A.; Stanneck, D.; Mencke, N.; Brandão-Filho, S.P.; Alves, L.C.; Otranto, D. Detection of Leishmania infantum in Rhipicephalus sanguineus Ticks from Brazil and Italy. Parasitol. Res. 2010, 106, 857–860. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Martins, T.F.; de Paiva-Cavalcanti, M.; Figueredo, L.A.; Lima, B.S.; Brandão-Filho, S.P. Transovarial Passage of Leishmania infantum KDNA in Artificially Infected Rhipicephalus sanguineus. Exp. Parasitol. 2010, 125, 184–185. [Google Scholar] [CrossRef]
- de Morais, R.C.S.; Gonçalves-de-Albuquerque, S. da C.; Silva, Rô.P. e.; Costa, P.L.; Silva, K.G. da; Silva, F.J. da; Brandão-Filho, S.P.; Dantas-Torres, F.; Paiva-Cavalcanti, M. de Detection and Quantification of Leishmania braziliensis in Ectoparasites from Dogs. Vet. Parasitol. 2013, 196, 506–508. [Google Scholar] [CrossRef] [PubMed]
- de Morais, R.C.S.; Gonçalves, S. da C.; Costa, P.L.; da Silva, K.G.; da Silva, F.J.; Silva, R.P.E.; de Brito, M.E.F.; Brandão-Filho, S.P.; Dantas-Torres, F.; de Paiva-Cavalcanti, M. Detection of Leishmania infantum in Animals and Their Ectoparasites by Conventional PCR and Real Time PCR. Exp. Appl. Acarol. 2013, 59, 473–481. [Google Scholar] [CrossRef]
- Abdullahi, M.A.; Abubakar, Z.T.; Umar, L.M.; Sabo, H.M.; Onaolapo, A.Y.; Jibril, H.; Abubakar, A.; Haruna, I.L.; Muhammed, A.S.; Tasie, P.C. Amblyomma variegatum as Bioindicators for the Presence of Trypanosoma congolense in Semi-Extensively Managed White Fulani Cattle in Kaduna Metropolis, Kaduna State, Nigeria. IOSR J. Agric. Vet. Sci. 2016, 9, 42–45. [Google Scholar]
- Aeschlimann, A.; Burgdorfer, W.; Matile, H. Aspects Nouveaux Du Role De Vecteur Joue Par Ixodes Ricinus L. En Suisse. Note Preliminaire. Acta Trop. 1979, 36, 181–191. [Google Scholar] [PubMed]
- Bacellar, F.; Núncio, M.S.; Filipe, A.R. Forms of Trypanosoma in Tick Haemolymph. Rev. Port. Ciências Veterinárias 1990, 85, 158–159. [Google Scholar]
- Bolivar, A.M. Detection of Anaplasma Marginale and Trypanosoma Vivax in Ticks Collected from Cattle Using the Polimerase Chain Reaction. REDVET 2013, 14. [Google Scholar]
- Cominetti, M.C.; Almeida, R.F.C. de; Csordas, B.G.; Andreotti, R. Ticks as Bioindicators the Presence of Trypanosoma spp. in Campo Grande City, MS, Brazil; 2013.
- Dyonisio, G.H.S.; Batista, H.R.; Silva, R.E. da; Azevedo, R.C. de F. e; Costa, J. de O.J.; Manhães, I.B. de O.; Tonhosolo, R.; Gennari, S.M.; Minervino, A.H.H.; Marcili, A. Molecular Diagnosis and Prevalence of Trypanosoma Vivax (Trypanosomatida: Trypanosomatidae) in Buffaloes and Ectoparasites in the Brazilian Amazon Region. J. Med. Entomol. 2021, 58, 403–407. [Google Scholar] [PubMed]
- Ergunay, K.; Mutinda, M.; Bourke, B.; Justi, S.A.; Caicedo-Quiroga, L.; Kamau, J.; Mutura, S.; Akunda, I.K.; Cook, E.; Gakuya, F.; et al. Metagenomic Investigation of Ticks From Kenyan Wildlife Reveals Diverse Microbial Pathogens and New Country Pathogen Records. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, P.; Sudhagar, S.; Goudar, A.L.; Molecular survey and phylogenetic analysis of tick-borne pathogens in ticks infesting cattle from two South Indian states, Vet. Parsit. Reg. Stud. 2021. 25. [CrossRef]
- Luu, L.; Bown, K.J.; Palomar, A.M.; Kazimirova, M.; Bell-Sakyi, L. Isolation and Partial Characterisation of a Novel Trypanosoma from the Tick Ixodes ricinus. Ticks Tick. Borne. Dis. 2020, 11. [Google Scholar] [CrossRef]
- Martins, J.R.; Leite, R.C.; Doyle, R.L. Tripanosomatides like Trypanosoma theileri in the Cattle Tick Boophilus microplus. Rev. Bras. Parasitol. Vet. 2008, 17, 113–114. [Google Scholar] [CrossRef]
- Monod, R.Y.; Aeschlimann, A.; Derscheid, J.M. 3 Infections by Trypanosomes Observed in Hyalomma detritum, Ixodes ricinus and Rhipicephalus sanguineus (Acarina: Ixodidae). Schweiz. Arch. Tierheilkd. 1986, 128, 243–254. [Google Scholar]
- Opazo, A.; Bacigalupo, A.; Urrutia, S.; Chavez, G. Detection of Trypanosoma cruzi Infection by PCR in Canis lupus familiaris and Their Ectoparasites in Chile. Med. Vet. Entomol. 2022, 36, 88–96. [Google Scholar] [CrossRef]
- Postoyan, S.R. Сoхранение Жизнеспoсoбнoсти Trypanosoma Vivax Ziemann, 1905 в Клещах. Հայաստանի կենսաբանական հանդես 2001, 53, 128–130. [Google Scholar]
- Said, Y.; Lahmar, S.; Dhibi, M.; Rjeibi, M.R.; Jdidi, M.; Gharbi, M. First Survey of Ticks, Tick-Borne Pathogens (Theileria, Babesia, Anaplasma and Ehrlichia) and Trypanosoma evansi in Protected Areas for Threatened Wild Ruminants in Tunisia. Parasitol. Int. 2021, 81. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, R.F.; Garcia, M.V.; Cunha, R.C.; Matias, J.; e Silva, E.; Cepa Matos, M. de F.; Andreotti, R. Ixodid Fauna and Zoonotic Agents in Ticks from Dogs: First Report of Rickettsia Rickettsii in Rhipicephalus Sanguineus in the State of Mato Grosso Do Sul, Mid-Western Brazil. Exp. Appl. Acarol. 2013, 60, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Krinsky, W.L.; Burgdorfer, W. Trypanosomes in Amblyomma Americanum from Oklahoma. J. Parasitol. 1976. [Google Scholar] [CrossRef]
- Brotánková, A.; Fialová, M.; Čepička, I.; Brzoňová, J.; Svobodová, M. Trypanosomes of the Trypanosoma theileri Group: Phylogeny and New Potential Vectors. Microorganisms 2022, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Oluwafemi, R.A.; Ilemobade, A.A.; Laseinde, E.A.O. The Impact of African Animal Trypanosomosis and Tsetse on the Livelihood and Wellbeing of Cattle and Their Owners in the BICOT Study Area of Nigeria. Sc Res Essays 2007, 2, 380–383. [Google Scholar]
- Awuoche, E.O.; Weiss, B.L.; Mireji, P.O.; Vigneron, A.; Nyambega, B.; Murilla, G.; Aksoy, S. Expression Profiling of Trypanosoma congolense Genes during Development in the Tsetse Fly Vector Glossina morsitans morsitans. Parasit. Vectors 2018, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.H.S.; Almeida, A.; Figueiredo, F.B.; Sousa, V.R.F.; Fagundes, A.; Pinto, A.G.S.; Baptista, C.; Madeira, M.F. Occurrence of Trypanosoma caninum in Areas Overlapping with Leishmaniasis in Brazil: What Is the Real Impact of Canine Leishmaniasis Control? Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 419–423. [Google Scholar] [CrossRef]
- Azarmi, S.; Zahraei-Ramazani, A.; Mohebali, M.; Rassi, Y.; Akhavan, A.A.; Azarm, A.; Dehghan, O.; Elikaee, S.; Abdoli, R.; Mahmoudi, M. PCR Positivity of Gerbils and Their Ectoparasites for Leishmania spp. in a Hyperendemic Focus of Zoonotic Cutaneous Leishmaniasis in Central Iran. J. Arthropod. Borne. Dis. 2022, 16, 124. [Google Scholar] [CrossRef]
- Campos, J.H.F.; Costa, F.A.L. Participation of Ticks in the Infectious Cycle of Canine Visceral Leishmaniasis, in Teresina, Piauí, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 297–300. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Rhipicephalus Sanguineus E a Epidemiologia Da Leishmaniose Visceral Canina No Estado De Pernambuco 2009, 96.
- Giraud, P. Transmission of KA in the Mediterranean Region, with Special Reference to Ticks. Bull. la Société Pathol. Exot. 1934, 27. [Google Scholar]
- Magri, A.; Caffara, M.; Fioravanti, M.; Galuppi, R. Detection of Leishmania Sp . KDNA in Questing Ixodes Ricinus ( Acari , Ixodidae ) from the Emilia - Romagna Region in Northeastern Italy. 2022, 3331–3336. [CrossRef]
- Odorizzi, R.M.F.N. Investigação Do Papel de Vetores Secundários Da Leishmania (Leishmania) Infantum No Município de Mirandópolis-SP, Brasil; Universidade de São Paulo, 2010. [CrossRef]
- Persichetti, M.F.; Solano-Gallego, L.; Serrano, L.; Altet, L.; Reale, S.; Masucci, M.; Pennisi, M.G. Detection of Vector-Borne Pathogens in Cats and Their Ectoparasites in Southern Italy. Parasites and Vectors 2016, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Jaimes, J.; Correa-Núñez, G.H.; Donayre, L.; Lescano, A.G. Quantitative Detection of LeishmaniaLeishmania in Rhipicephalus microplus and Amblyomma sabanerae in the Peruvian Amazon Basin. Trop. Med. Infect. Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Silva, O.A. da; Braga, G.M. da S. The Role of Rhipicephalus Sanguineus in the Transmission of Visceral Leishmaniasis Dogs: Epidemiological Aspects. PUBVET 2010, 4. [Google Scholar]
- Rojas-jaimes, J.; Zarate-rendon, D.; Montoya-arauco, G. Aislamiento e Identificación de Leishmania sp de Amblyomma naponense y Amblyomma humerale Colectadas de Pecari tajacu , Tayassu pecari y Chelonoide Denticulata Isolation and Identification of Leishmania sp of Amblyomma naponense and Amblyomma humerale Col. Amblyomma humerale 2022, 19, 189–192. [Google Scholar] [CrossRef]
- Satta, G.; Chisu, V.; Cabras, P.; Fois, F.; Masala, G. Pathogens and Symbionts in Ticks: A Survey on Tick Species Distribution and Presence of Tick-Transmitted Micro-Organisms in Sardinia, Italy. J. Med. Microbiol. 2011, 60, 63–68. [Google Scholar] [CrossRef]
- Karasartova, D.; Gureser, A.S.; Gokce, T.; Celebi, B.; Yapar, D.; Keskin, A.; Celik, S.; Ece, Y.; Erenler, A.K.; Usluca, S.; et al. Bacterial and Protozoal Pathogens Found in Ticks Collected from Humans in Corum Province of Turkey. PLoS Negl. Trop. Dis. 2018, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, Q.; Liu, J.Q.; Xu, B.L.; ... Tick-Borne Pathogens and Associated Co-Infections in Ticks Collected from Domestic Animals in Central China. Parasites \& … 2014. [CrossRef]
- Paz, G.F.; Reis, I.A.; Avelar, D.M.; Ferreira, E.C. d. M.; Werneck, G.L. Ectoparasites and Anti-Leishmania Antibodies: Association in an Observational Case-Control Study of Dogs from a Brazilian Endemic Area. Prev. Vet. Med. 2013, 112, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhang, J.; Shi, Z.; Song, C.; Zheng, X.; Zhang, Y.; Hao, Y.; Dong, H.; Wei, L.; El-Mahallawy, H.S.; et al. Molecular Detection of Vector-Borne Agents in Dogs from Ten Provinces of China. Parasites and Vectors 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Latrofa, M.S.; Otranto, D. Quantification of Leishmania infantum DNA in Females, Eggs and Larvae of Rhipicephalus Sanguineus. Parasit. Vectors 2011, 4, 1–4. [Google Scholar] [CrossRef]
- Dantas-Torres, F. The Role of Dogs as Reservoirs of Leishmania Parasites, with Emphasis on Leishmania (Leishmania) infantum and Leishmania (Viannia) braziliensis. Vet. Parasitol. 2007, 149, 139–146. [Google Scholar] [CrossRef]
- Odorizzi, Rosa Maria Ferreira Noguerol. Investigação do papel de vetores secundários da Leishmania (Leishmania) infantum no Município de Mirandópolis-SP, Brasil. 2010. Thèse de doctorat. Universidade de São Paulo. [CrossRef]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A Review. F1000Research 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Sereno, D.; Akhoundi, M.; Dorkeld, F.; Oury, B.; Momen, H.; Perrin, P. What Pre-Columbian Mummies Could Teach Us about South American Leishmaniases? Pathog. Dis. 2017, 75, ftx019. [Google Scholar] [CrossRef] [PubMed]
- Desbois, N.; Pratlong, F.; Quist, D.; Dedet, J.-P. Leishmania (Leishmania) martiniquensis n. sp.(Kinetoplastida:Trypanosomatidae), Description of the Parasite Responsible for Cutaneous Leishmaniasis in Martinique Island (French West Indies). Parasite 2014, 21. [Google Scholar] [CrossRef] [PubMed]
- Noyes, H.; Pratlong, F.; Chance, M.; Ellis, J.; Lanotte, G.; Dedet, J.-P. A Previously Unclassified Trypanosomatid Responsible for Human Cutaneous Lesions in Martinique (French West Indies) Is the Most Divergent Member of the Genus Leishmania Ss. Parasitology 2002, 124, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Dedet, J.P.; Roche, B.; Pratlong, F.; Cales-Quist, D.; Jouannelle, J.; Benichou, J.C.; Huerre, M. Diffuse Cutaneous Infection Caused by a Presumed Monoxenous Trypanosomatid in a Patient Infected with HIV. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 644–646. [Google Scholar] [CrossRef] [PubMed]
- Seblova, V.; Sadlova, J.; Vojtkova, B.; Votypka, J.; Carpenter, S.; Bates, P.A.; Volf, P. The Biting Midge Culicoides sonorensis (Diptera: Ceratopogonidae) Is Capable of Developing Late Stage Infections of Leishmania enriettii. PLoS Negl. Trop. Dis. 2015, 9, e0004060. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Jaimes, J.; Correa-Núñez, G.H.; Donayre, L.; Lescano, A.G. Quantitative Detection of Leishmania in Rhipicephalus microplus and Amblyomma sabanerae in the Peruvian Amazon Basin. Trop. Med. Infect. Dis. 2022, 7, 358. [Google Scholar] [CrossRef] [PubMed]
- Morzaria, S.P.; Latif, A.A.; Jongejan, F.; Walker, A.R. Transmission of a Trypanosoma sp to cattle by the tick Hyalomma anatolicum anatolicum. Vet. Parasitol. 1986, 19, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Latif, A.A.; Bakheit, M.A.; Mohamed, A.E.; Zweygarth, E. High Infection Rates of the Tick Hyalomma Anatolicum Anatolicum with Trypanosoma Theileri. ONDERSTEPOORT J. Vet. Res. 2004, 71, 251–256. [Google Scholar] [CrossRef]
- Krige, A.-S.; Thompson, R.C.A.; Clode, P.L. ‘Hang on a Tick’–Are Ticks Really the Vectors for Australian Trypanosomes? Trends Parasitol. 2019, 35, 596–606. [Google Scholar] [CrossRef]
- Taylor-lewis, E.G. Investigations on the Transmission of Trypanosoma Lewisi and T. Evansi through 5 Different Species of Hard Ticks. Investig. Transm. Trypanos. lewisi T. evansi through 5 Differ. species hard ticks. 1976. [Google Scholar]
- Mahmoud, N.E.; Habeeb, S.M.; Abdelrahman, K.A.; Ashry, H.M.; Hassan, M.R. Role of Tampan Tick Ornithodoros savignyi (Acari: Argasidae) in Transmitting Trypanosoma evansi in Laboratory Animals. Int. J. Vet. Sci. 2020, 9, 273–278. [Google Scholar] [CrossRef]
- Dujardin, J.-C.; Campino, L.; Cañavate, C.; Dedet, J.-P.; Gradoni, L.; Soteriadou, K.; Mazeris, A.; Ozbel, Y.; Boelaert, M. Spread of Vector-Borne Diseases and Neglect of Leishmaniasis, Europe. Emerg. Infect. Dis. 2008, 14, 1013. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, M.T.Z.; Linardi, P.M. Can Fleas from Dogs Infected with Canine Visceral Leishmaniasis Transfer the Infection to Other Mammals? Vet. Parasitol. 2007, 147, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Rakhshanpour, A.; Malmasi, A.; Mohebali, M.; Nabian, S.; Mirhendi, H.; Zarei, Z.; Dalimi, A.; Mohammadiha, A.; Akhoundi, B.; Azarm, A. Transmission of Leishmania Infantum by Rhipicephalus Sanguineus (Acari: Ixodidae) in Dogs. Iran. J. Parasitol. 2017, 12, 482. [Google Scholar] [PubMed]
- Joyeux, C.; Sautet, J. Observations Sur La Leishmaniose Canine Méditerranéenne. Bull. la Société Pathol. Exot. 1938, 31. [Google Scholar]
- Arganaraz, E.R.; Hubbard, G.B.; Ramos, L.A.; Ford, A.L.; Nitz, N.; Leland, M.M.; Vandeberg, J.L.; Teixeira, A.R.L. Blood-Sucking Lice May Disseminate Trypanosoma cruzi Infection in Baboons. Rev. Inst. Med. Trop. Sao Paulo 2001, 43, 271–276. [Google Scholar] [CrossRef]
- Votýpka, J.; Szabova, J.; Radrova, J.; Zídková, L.; Svobodova, M. Trypanosoma culicavium Sp. Nov., an Avian Trypanosome Transmitted by Culex Mosquitoes. Int. J. Syst. Evol. Microbiol. 2012, 62, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G.; Samish, M.; Shkap, V. Life Cycle of Hepatozoon canis (Apicomplexa: Adeleorina: Hepatozoidae) in the Tick Rhipicephalus sanguineus and Domestic Dog (Canis familiaris). J. Parasitol. 2007, 93, 283–299. [Google Scholar] [CrossRef]
- Weilgama, D.J. Cyclical Development of Trypanosomes in the Tick and Their Transmission. Haemoprotozoan Dis. Domest. Anim. proceedings, Semin. Asian/Australian Reg. Haryana Agric. Univ., Hissar, India, Oct. 27-Nov. 1, 1980/edited by OP Gautam, RD Sharma, S. Dhar 1980.
- Arifdzhanov, K.A.; Nikitina, R.E. Detection of Crithidia Hyalomma (O’Farrel 1913) in Hyalomma a. Anatolicum (Koch, 1844). Zool. Cheskii 1961, 40, 20–24. [Google Scholar]
- Vergne, T.; Kamyinkird, K.; Desquesnes, M.; Jittapalapong, S. Attempted Transmission of Trypanosoma evansi to Rats and Mice by Direct Ingestion of Contaminated Blood and via Engorged Ticks. ACTA Protozool. 2011, 50, 133–136. [Google Scholar] [CrossRef]
- Bilgiç, H.B.; Bakirci, S.; Köse, O.; Aksulu, A.; Hacilarlioglu, S.; Karagenç, T. Determination the Role of Rhipicephalus sanguineus for Transmission of Leishmania major to Reservoir Animals. Türkiye Parazitolojii Derg. 2016, 40, 179. [Google Scholar] [CrossRef] [PubMed]
- Paz, G.F.; Ribeiro, M.F.B.; Magalhães, D.F. de; Sathler, K.P.B.; Morais, M.H.F.; Fiúza, V.O.P.; Brandão, S.T.; Werneck, G.L.; Fortes-Dias, C.L.; Dias, E.S. Association between the Prevalence of Infestation by Rhipicephalus sanguineus and Ctenocephalides felis felis and the Presence of Anti-Leishmania Antibodies: A Case–Control Study in Dogs from a Brazilian Endemic Area. Prev. Vet. Med. 2010, 97, 131–133. [Google Scholar] [CrossRef]
- Nakkoud, J.R.; Santos, C.M. dos; Aquino, D.R.R.R.A.; Favacho, A.R. de M. Um Olhar Para as Populações de Ectoparasitas Em Cães Com Leishmaniose Visceral Canina (LVC) Em Mato Grosso Do Sul–Potenciais Vetores de Transmissão Para Essa Doença. Ensaios e Ciência C Biológicas Agrárias e da Saúde 2022, 26, 43–47. [Google Scholar] [CrossRef]
- Bernardino, M. das G. da S.; Angelo, D.F. do S.; Silva, R.B.S.; Silva, E.G. da; Silva, L.F.F.; Vaz, A.F. de M.; Melo, M.A. de; Santos, C. de S.A.B.; Alves, C.J.; Azevedo, S.S. de High Seroprevalence and Associated Factors for Visceral Leishmaniasis in Dogs in a Transmission Area of Paraíba State, Northeastern Brazil. Rev. Bras. Parasitol. Veterinária 2020, 29. [Google Scholar] [CrossRef]
- Santos-Doni, T.R.; Viol, M.A.; Lima, V.M.F.; Oliveira, B.C.M.; Matos, L.V.S.; Costa, A.J. da; Gomes, J.F.; Bresciani, K.D.S. Canine Visceral Leishmaniasis and Rhipicephalus sanguineus: Evaluation and Comparison of Classical Techniques. Vet. Res. Commun. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Piyasiri, S.B.; Dewasurendra, R.; Samaranayake, N.; Karunaweera, N. Diagnostic Tools for Cutaneous Leishmaniasis Caused by Leishmania donovani: A Narrative Review. Diagnostics 2023, 13, 2989. [Google Scholar] [CrossRef]
- Kariyawasam, K.; Edirisuriya, C.S.; Senerath, U.; Hensmen, D.; Siriwardana, H.; Karunaweera, N.D. Characteisation of Cutaneous Leishmaniasis in Matara District, Southern Sri Lanka: Evidence for Case Clustering. Pathog. Glob. Health 2015, 109, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Reimão, J.Q.; Coser, E.M.; Lee, M.R.; Coelho, A.C. Laboratory Diagnosis of Cutaneous and Visceral Leishmaniasis: Current and Future Methods. Microorganisms 2020, 8, 1632. [Google Scholar] [CrossRef]
- Caminade, C.; McIntyre, K.M.; Jones, A.E. Impact of Recent and Future Climate Change on Vector-borne Diseases. Ann. N. Y. Acad. Sci. 2019, 1436, 157–173. [Google Scholar] [CrossRef]
- de Angeli Dutra, D.; Salloum, P.M.; Poulin, R. Vector Microbiome: Will Global Climate Change Affect Vector Competence and Pathogen Transmission? Parasitol. Res. 2023, 122, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Macfie, J.W.S.; Thomson, J.G. A Trypanosome of the Canary (Serinus canarius Koch). Trans. R. Soc. Trop. Med. Hyg. 1929, 23. [Google Scholar] [CrossRef]
- Novy, F.G.; MacNeal, W.J.; Torrey, H.N. The Trypanosomes of Mosquitoes and Other Insects. J. Infect. Dis. 1907, 223–276. [Google Scholar] [CrossRef]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; Boer, M. den; Team, W.H.O.L.C. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS One 2012, 7, e35671. [Google Scholar] [CrossRef]
- Browne, A.J.; Guerra, C.A.; Alves, R.V.; Da Costa, V.M.; Wilson, A.L.; Pigott, D.M.; Hay, S.I.; Lindsay, S.W.; Golding, N.; Moyes, C.L. The Contemporary Distribution of Trypanosoma cruzi Infection in Humans, Alternative Hosts and Vectors. Sci. Data 2017, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Nava, S.; Robbins, R.G. Neotropical Hard Ticks (Acari: Ixodida: Ixodidae); Springer, 2021; ISBN 3030723526. [Google Scholar]
- Barros-Battesti, D.M.; Ramirez, D.G.; Landulfo, G.A.; Faccini, J.L.H.; Dantas-Torres, F.; Labruna, M.B.; Venzal, J.M.; Onofrio, V.C. Immature Argasid Ticks: Diagnosis and Keys for Neotropical Region. Rev. Bras. Parasitol. Veterinária 2013, 22, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Gürtler, R.E.; Cardinal, M.V. Reservoir Host Competence and the Role of Domestic and Commensal Hosts in the Transmission of Trypanosoma cruzi. Acta Trop. 2015, 151, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Alemayehu, B.; Alemayehu, M. Leishmaniasis: A Review on Parasite, Vector and Reservoir Host. Heal. Sci. J. 2017, 11, 1. [Google Scholar] [CrossRef]
- Sánchez-Montes, S.; Salceda-Sánchez, B.; Bermúdez, S.E.; Aguilar-Tipacamú, G.; Ballados-González, G.G.; Huerta, H.; Aguilar-Domínguez, M.; Mora, J.D. la; Licona-Enríquez, J.D.; Mora, D.D. la Rhipicephalus Sanguineus Complex in the Americas: Systematic, Genetic Diversity, and Geographic Insights. Pathogens 2021, 10, 1118. [Google Scholar] [CrossRef]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press, 2000; ISBN 0521480086. [Google Scholar]
- Kahl, O.; Gern, L.; Eisen, L.; Lane, R.S. Ecological Research on Borrelia burgdorferi sensu lato: Terminology and Some Methodological Pitfalls. 2002.
- Nuttall, P.A. Molecular Characterization of Tick-Virus Interactions. Front Biosci 2009, 14, 2466–2483. [Google Scholar] [CrossRef]
- Gargili, A.; Estrada-Peña, A.; Spengler, J.R.; Lukashev, A.; Nuttall, P.A.; Bente, D.A. The Role of Ticks in the Maintenance and Transmission of Crimean-Congo Hemorrhagic Fever Virus: A Review of Published Field and Laboratory Studies. Antiviral Res. 2017, 144, 93–119. [Google Scholar] [CrossRef] [PubMed]
- Lazzari, C.R.; Fauquet, A.; Lahondère, C.; Araújo, R.N.; Pereira, M.H. Soft Ticks Perform Evaporative Cooling during Blood-Feeding. J. Insect Physiol. 2021, 130, 104197. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J.; Andersen, J.F.; Francischetti, I.M.B.; Valenzuela, J.G.; Schwan, T.G.; Pham, V.M.; Garfield, M.K.; Hammer, C.H.; Ribeiro, J.M.C. Comparative Sialomics between Hard and Soft Ticks: Implications for the Evolution of Blood-Feeding Behavior. Insect Biochem. Mol. Biol. 2008, 38, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Antunes, S.; Rosa, C.; Couto, J.; Ferrolho, J.; Domingos, A. Deciphering Babesia-Vector Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 429. [Google Scholar] [CrossRef] [PubMed]
- Tahir D, Meyer L, Fourie J, Jongejan F, Mather T, Choumet V, Blagburn B, Straubinger RK, Varloud M. 2020. Interrupted blood feeding in ticks: causes and consequences. Microorganisms 8:910. [CrossRef]
- Velásquez-Ortiz, N.; Ramírez, J.D. Understanding the Oral Transmission of Trypanosoma Cruzi as a Veterinary and Medical Foodborne Zoonosis. Res. Vet. Sci. 2020, 132, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; ... & Moher, D.(2021). The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Bmj 2021. [CrossRef]
- Nakagawa, S.; Noble, D.W.A.; Senior, A.M.; Lagisz, M. Meta-Evaluation of Meta-Analysis: Ten Appraisal Questions for Biologists. BMC Biol. 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.; Hill, S. Supporting Implementation of Cochrane Methods in Complex Communication Reviews: Resources Developed and Lessons Learned for Editorial Practice and Policy. Heal. Res. Policy Syst. 2019, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.J. The Inverse of the Freeman–Tukey Double Arcsine Transformation. Am. Stat. 1978, 32, 138. [Google Scholar] [CrossRef]
- Freeman, M.F.; Tukey, J.W. Transformations Related to the Angular and the Square Root. Ann. Math. Stat. 1950, 607–611. [Google Scholar] [CrossRef]
Figure 1.
PRISMA Flowchart of the systematic review and meta-analysis.
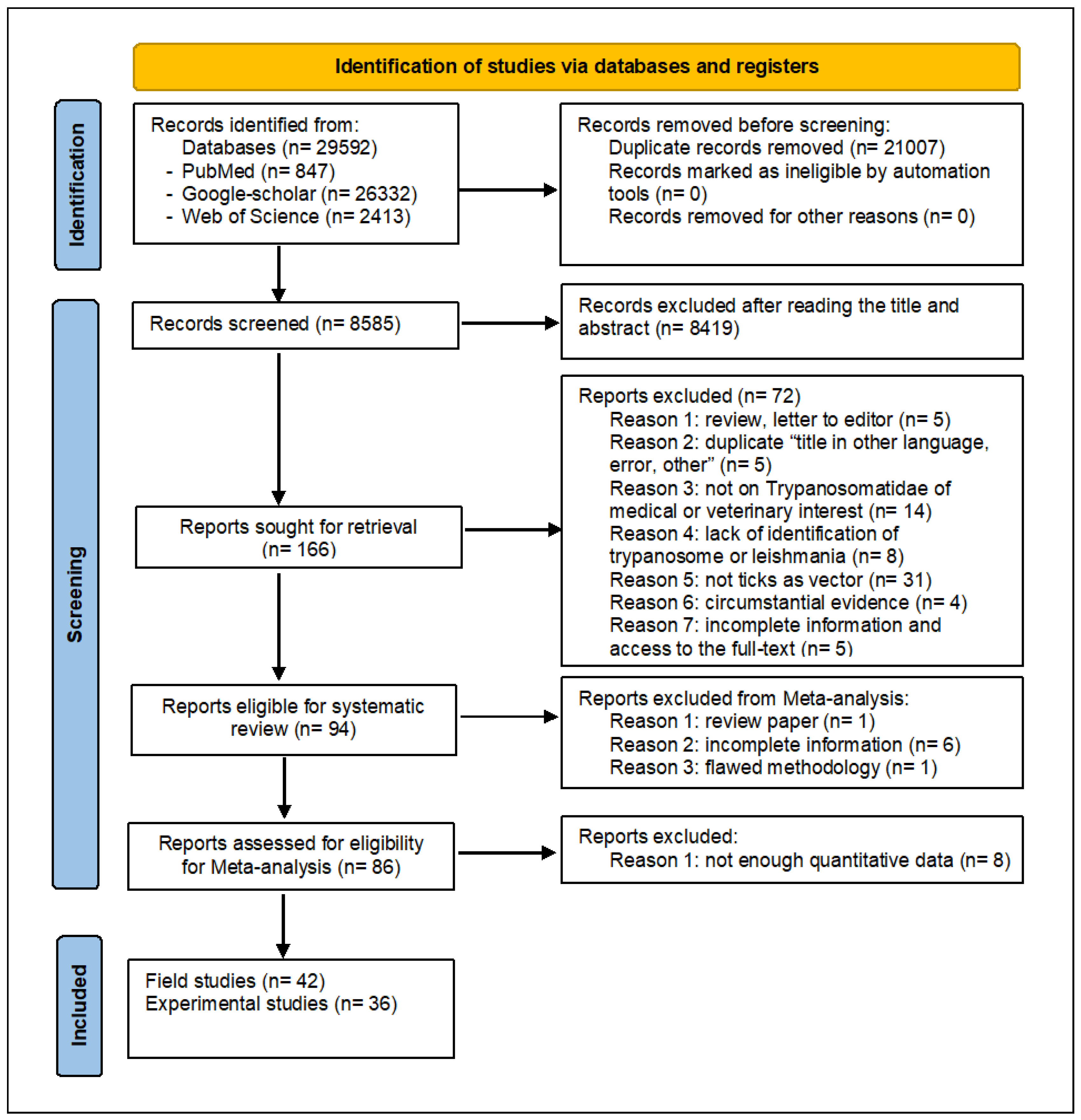
Figure 2.
Distribution of the collected and selected scientific papers.
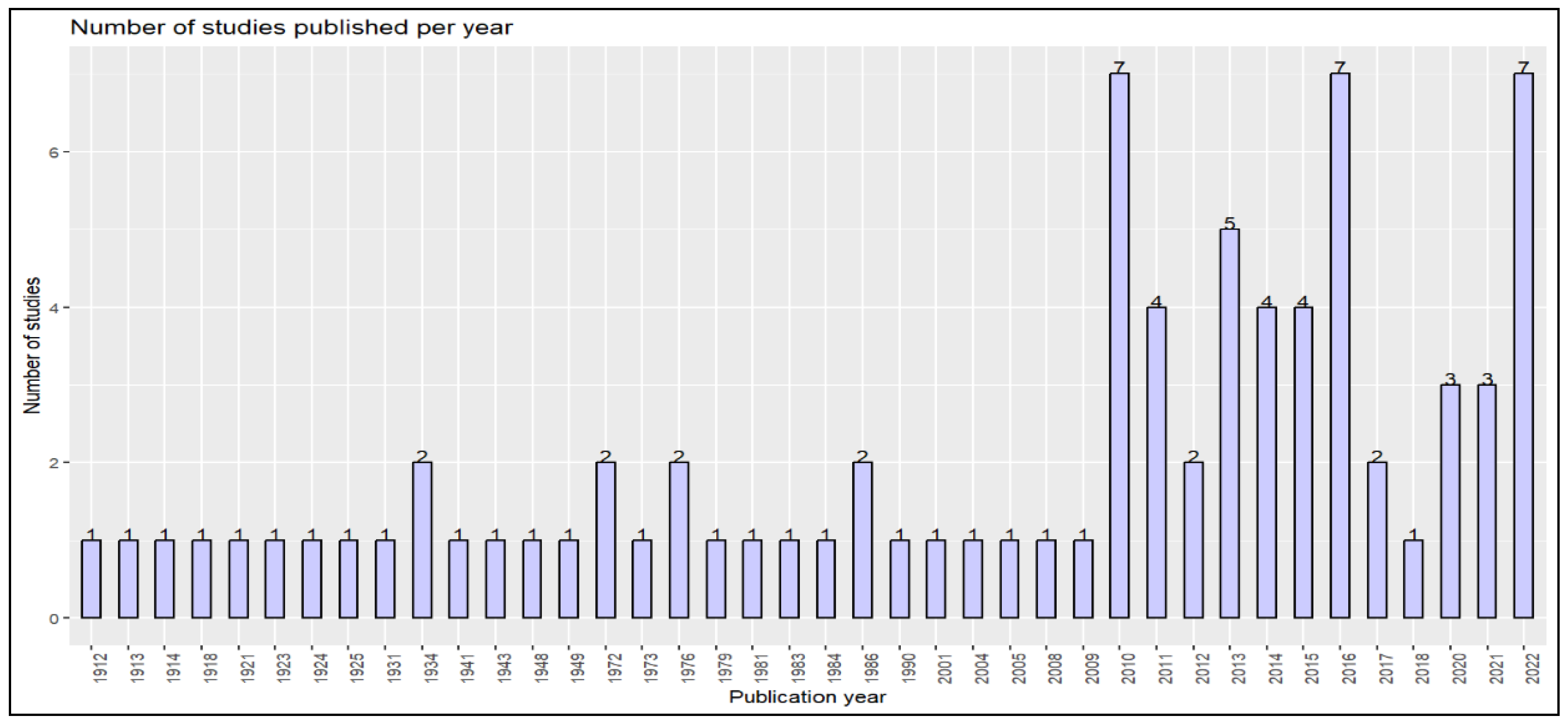
Figure 3.
Forest plot of prevalence of Trypanosomatidae detection in field ticks. The length of the horizontal line represents the 95% CI; the diamond represents the summarized effect.
Figure 3.
Forest plot of prevalence of Trypanosomatidae detection in field ticks. The length of the horizontal line represents the 95% CI; the diamond represents the summarized effect.
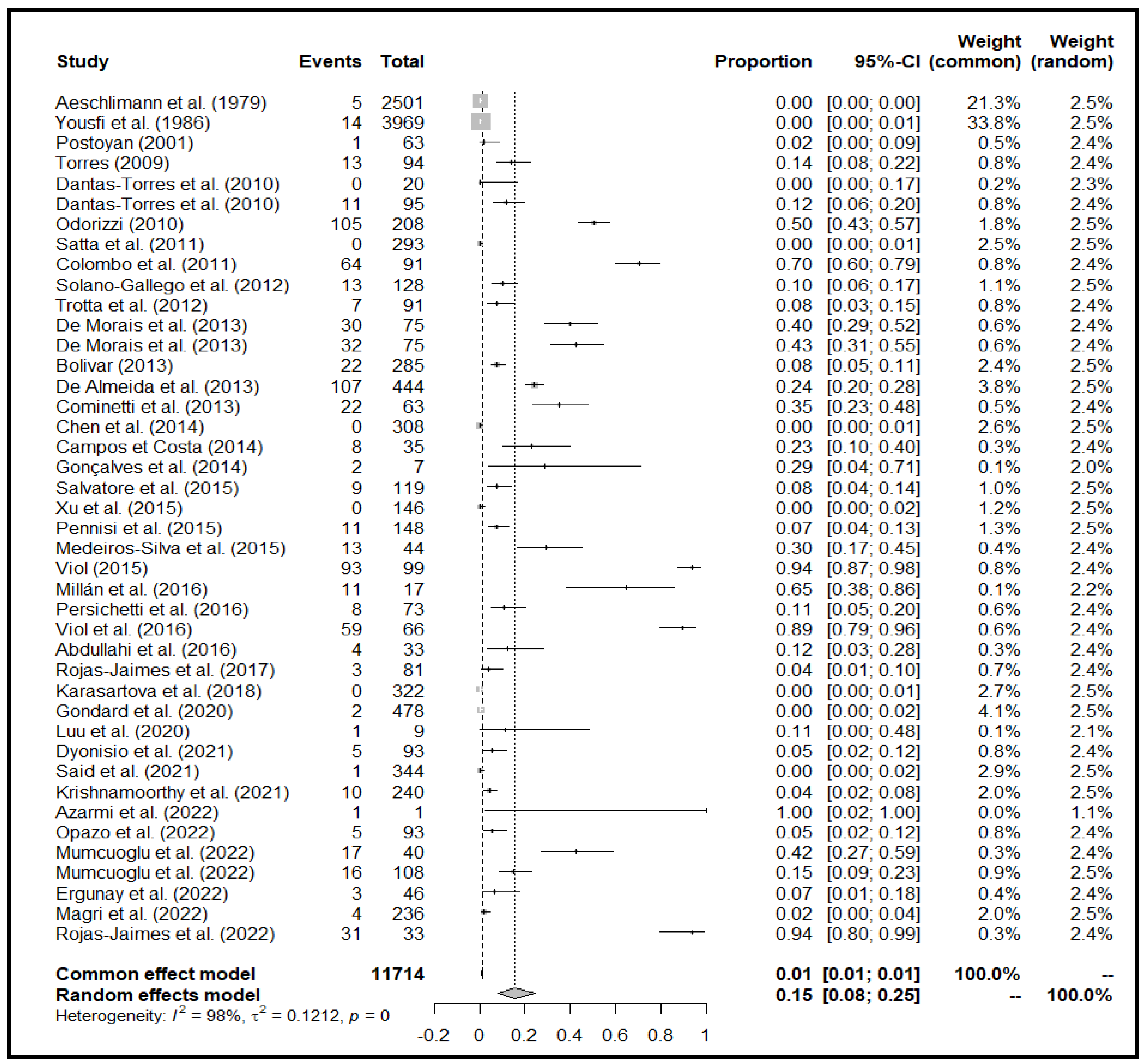
Figure 4.
Funnel plot with 95% confidence limit intervals for the examination of publication bias.
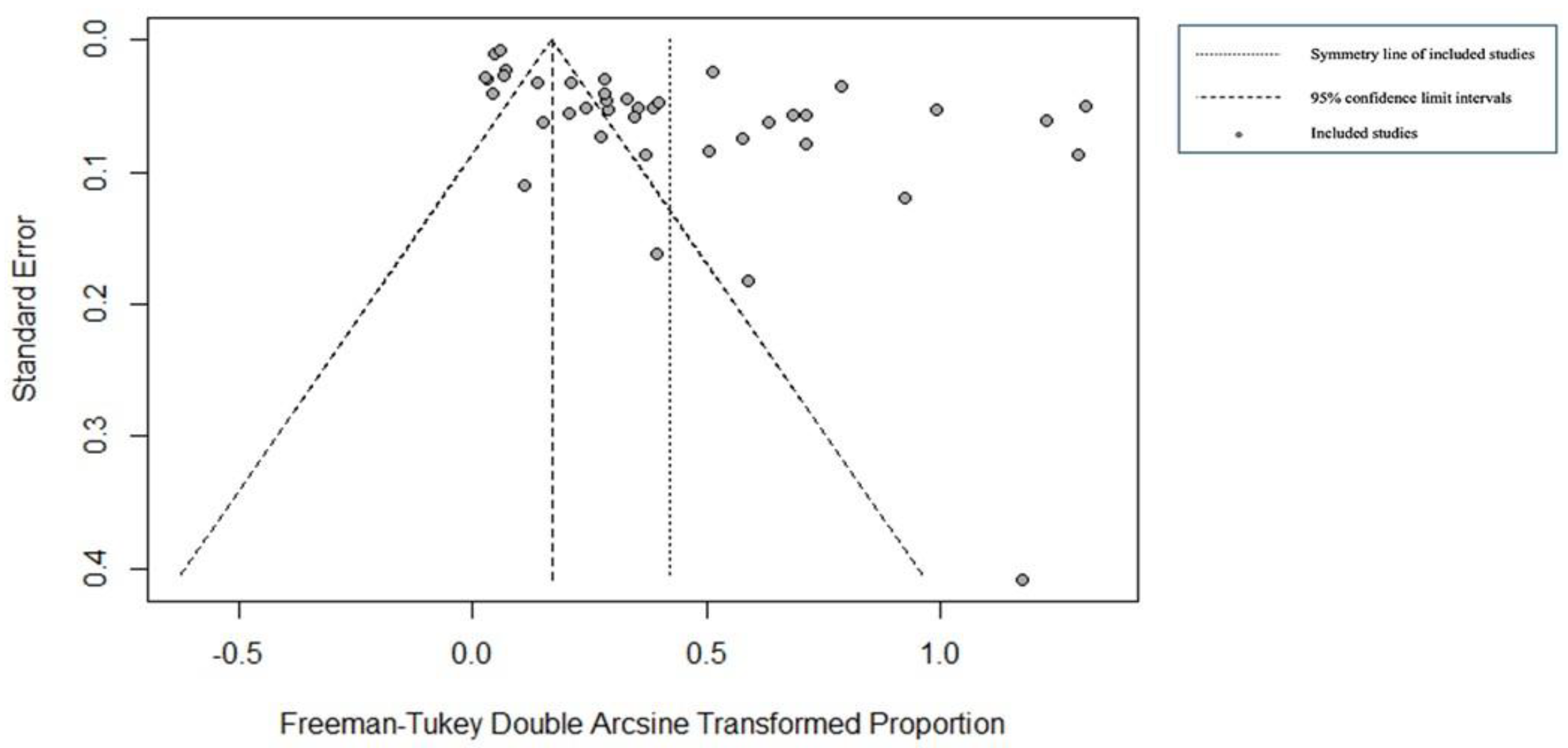

Table 1.
Evidences of presence of Trypanosoma of medical and veterinary interest in field-collected ticks.
Table 1.
Evidences of presence of Trypanosoma of medical and veterinary interest in field-collected ticks.
| Species | Sampling year | Trypansoma species | Tick statut (P/N) |
Localization in the tick | Tick Detection method |
Host (Species) | Country | Continent | Author(s) |
|---|---|---|---|---|---|---|---|---|---|
| Amblyomma cajennense | 2013 | T. cruzi | N | H ; Gr | Microscopy, PCR | Dog (Canis lupus) | Brazil | South America | [92] |
| NS | T. vivax | N | H | Microscopy | Bovine (Bos taurus) | Cuba | South America | [90] | |
| NS | T. vivax | P | G | Microscopy | Bovine (Bos taurus) | Cuba | South America | [90] | |
| NS | T. vivax | P | Gr | PCR, Sequencing | buffalos | Brazil | South America | [83] | |
| Amblyomma longirostrum | NS | T. cruzi | P | NS | NS | Cercolabidae (Arboreal porcupine) | Venezuela | South America | [36] |
| Amblyomma spp. | 2013-2019 | T. cruzi | N | Gr | Metagenomics | * Wild ungulates Carnivores | Kenya | Africa | [84] |
| Amblyomma tigrinum | 2013 | T. cruzi | N | Gr | PCR, Sequencing | Canidae (Canis lupus familiaris) | Chile | South America | [89] |
| Amblyomma variegatum | NS | T. congolense | P | Gr | Parasitological analysis, PCR | Cattle | Nigeria | Africa | [78] |
| Haemaphysalis spp. | 2017-2018 | T. evansi | N | Gr | PCR, Sequencing | Cattle | India | Asia | [85] |
| Hyalomma detritum | 1982 | T. theileri | P | H | Microscopy | Bovine (Bos taurus) Dog (Canis lupus) | Algeria | Africa | [88] |
| Hyalomma dromedarii | 2015-2016 | T. evansi | N | Gr | PCR | **Wild ruminants species | Tunisia | Africa | [91] |
| Hyalomma excavatum | 2015-2016 | T. evansi | N | Gr | PCR | **Wild ruminants species | Tunisia | Africa | [91] |
| 1982 | T. theileri, T. evansi | N | H | Microscopy | Bovine (Bos taurus) Dog (Canis lupus ) | Algeria | Africa | [88] | |
| Hyalomma lusitanicum | 1982 | T. theileri, T. evansi | N | H | Microscopy | Bovine (Bos taurus); Dog (Canis lupus) | Algeria | Africa | [88] |
| Hyalomma marginatum | 2015-2016 | T. evansi | N | Gr | PCR | **Wild ruminants species | Tunisia | Africa | [91] |
| NS | T. theileri | P | H | Microscopy | Bovine (Bos taurus) | Portugal | Europe | [80] | |
| 1982 | T. theileri, T. evansi | N | H | Microscopy | Bovine (Bos taurus) Dog (Canis lupus) | Algeria | Africa | [88] | |
| Hyalomma spp. | 2017-2018 | T. evansi | N | Gr | PCR, Sequencing | Cattle | India | Asia | [85] |
| Ixodes ricinus | 2013 | T. caninum | P | Gr | Culture, PCR, Sequencing | Vegetation | Slovakia | Europe | [86] |
| 1979-1982 | T. theileri | P | H | Microscopy | Bovine (Bos taurus) Dog (Canis lupus) | Switzerland | Europe | [88] | |
| 2015-2016 | T. evansi | N | Gr | PCR | **Wild ruminants species | Tunisia | Africa | [91] | |
| Rhipicephalus (Boophilus) microplus | NS | T. theileri-like | P | H | ND | Bovine (Bos taurus) | Brazil | South America | [87] |
| NS | T. vivax | N | H | Microscopy | Bovine (Bos taurus) | Cuba | South America | [90] | |
| NS | T. vivax | P | G | Microscopy | Bovine (Bos taurus) | Cuba | South America | [90] | |
| NS | T. vivax | P | Gr | PCR | Bovine (Bos taurus) | Venezuela | South America | [81] | |
| NS | T. vivax | P | Gr | PCR, Sequencing | buffalos | Brazil | South America | [83] | |
| Rhipicephalus (Boophilus) spp. | 2017-2018 | T. evansi | P | Gr | PCR, Sequencing | Cattle | India | Asia | [85] |
| Rhipicephalus bursa | 2015-2016 | T. evansi | N | Gr | PCR | **Wild ruminants species | Tunisia | Africa | [91] |
| Rhipicephalus sanguineus | 2013 | T. cruzi | N | H ; Gr | Microscopy, PCR | Dog (Canis lupus) | Brazil | South America | [92] |
| 1982 | T. evansi | P | H | Microscopy | Bovine (Bos taurus) Dog (Canis lupus) | Algeria | Africa | [88] | |
| NS | T. evansi | P | Gr | PCR | Dog (Canis lupus) | Brazil | South America | [82] | |
| NS | T. vivax | P | Gr | PCR | Dog (Canis lupus) | Brazil | South America | [82] | |
| Rhipicephalus sanguineus sensu lato | 2013 | T. cruzi | P | Gr | PCR, Sequencing | Canidae (Canis lupus familiaris) | Chile | South America | [89] |
| 2015-2016 | T. evansi | P | Gr | PCR | Wild ruminants species | Tunisia | Africa | [91] | |
| Rhipicephalus spp. | 2013-2019 | T. cruzi | P | Gr | Metagenomics | * * * Wild ungulates Carnivores Regular domestic | Kenya | Africa | [84] |
| Rhipicephalus turanicus | 1982 | T. theileri, T. evansi | N | H | Microscopy | Bovine (Bos taurus) Dog (Canis lupus) | Algeria | Africa | [88] |
NS : unspecified ; P : positive ; N : negative ; F: faeces; G: gut; Gr: whole body; H: haemolymph; O: ovaries; SG: salivary glands ; PCR : conventional Polymerase chain reaction, Sequencing : Capillary sanger sequencing.* Wild ungulates [Black rhinoceros (Diceros bicornis), white rhinoceros (Ceratotherium simum), buffalo (Syncerus caffer), elephant (Loxodonta africana), giraffe (Giraffa camelopardalis), Grévy’s zebra (Equus grevyi), plains zebra (Equus quagga), hartebeest (Alcelaphus buselaphus), impala (Aepyceros melampus)]; Carnivores [leopard (Panthera pardus), lion (Panthera leo), spotted hyena (Crocuta crocuta), wild dog (Lycaon pictus)]; Regular domestic [farmed Boran (Bos indicus) and cattle (Bos taurus)]. **Wild ruminants species : Scimitar-horned oryx; Addax antelope; Barbary red deer; Dorcas gazelle*** Wild ungulates [plains zebra (Equus quagga)]; Carnivores [lion (Panthera leo), spotted hyena (Crocuta crocuta)].
Table 2.
Evidences of presence of Leishmania of medical and veterinary interest in field-collected ticks.
Table 2.
Evidences of presence of Leishmania of medical and veterinary interest in field-collected ticks.
| Species | Sampling year | Leishmania species | Tick statut (P/N) |
Localisation in the tick | Tick Detection method |
Host (Species) | Host Detection method | Country | Continent | Author(s) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Amblyomma cajennense | 2013 | L. chagasi (Syn L. infantum) | N | H ; Gr | Microscopy, PCR | Dog (Canis lupus) | ND | Brazil | South America | [92] | |
| Amblyomma ovale | 2008 | L. infantum | N | Gr | qPCR | Dog (Canis lupus) | IFAT | Brazil | South America | [73] | |
| Amblyomma sabanerae | NS | Leishmania sp. | P | Gr | qPCR | Collared peccary (Pecari tajacu) Tortoise (Chelonoidis denticulata) | ND | Peru | South America | [107] | |
| Amblyomma spp. | 2012 | L. guyanensis | P | Gr | PCR ; HRM-PCR | Tapiridae (Tapirus terrestris) Tayassuidae (Pecari tajacu) | ND | Peru | South America | [63] | |
| Amblyomma tigrinum | 2010-2013 | Leishmania sp. | P | Gr | PCR | Wild foxes (Pseudalopex griseus) | PCR, qPCR | Argentina | South America | [58] | |
| Amblyomma variegatum | 2014-2015 | L. martiniquensis | P | Gr | HT-qPCR | Cattle | ND | Guadeloupe | Island | [56] | |
| Dermacentor marginatus | 2007-2008 | L. infantum | N | Gr | PCR | Wild boars | ND | Italy | Europe | [108] | |
| 2007 | L. infantum | N | Gr | qPCR | Dog (Canis lupus) | ELISA, PCR | Italy | Europe | [66] | ||
| 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | ||
| Haemaphysalis longicornis | 2012 | L. infantum | N | Gr | PCR | sheep, cattle and dogs | ND | China | Asia | [110] | |
| Haemaphysalis parva | 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | |
| Haemaphysalis punctata | 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | |
| Haemaphysalis sulcata | 2007-2008 | L. infantum | N | Gr | PCR | Sheep; Goats | ND | Italy | Europe | [108] | |
| 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | ||
| Hyalomma aegyptium | 2015-2018 | L. infantum | P | Gr | PCR ; Sequencing | Tortoise (Testudo graeca) Arabian camels (Camelus dromedarius) | ND | Israel | Asia | [59] | |
| 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | ||
| Hyalomma dromedarii | 2015-2018 | L. infantum | P | Gr | PCR, Sequencing | Tortoise (Testudo graeca) Arabian camels (Camelus dromedarius) | ND | Israel | Asia | [59] | |
| Hyalomma excavatum | 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | |
| Hyalomma marginatum | 2007-2008 | L. infantum | N | Gr | PCR | Cattle | ND | Italy | Europe | [108] | |
| 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | ||
| Hyalomma spp. | 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | |
| Ixode ricinus | 2007 | L. infantum | P | Gr | qPCR | Dog (Canis lupus) | ELISA, PCR | Italy | Europe | [66] | |
| 2007-2008; 2010 | L. infantum | P | Gr | PCR | Dogs, horses, cat, bovine, humans | ND | Italy | Europe | [64] | ||
| 2011 and 2013 | L. infantum | P | Gr | PCR | Cat (Felis catus) | ND | Italy | Europe | [62] | ||
| Ixode ricinus(Cont.) | 2012-2013 | L. infantum | P | Gr | qPCR | Cat (Felis catus) | PCR, qPCR | Italy | Europe | [104] | |
| 2010 | L. infantum | P | Gr | PCR ; Sequencing | Flagging | ND | Italy | Europe | [102] | ||
| 2014 | Leishmania sp. | P | Gr | PCR | Human | ND | Turkey | Asia | [109] | ||
| Ixodes spp. | 2011 and 2013 | L. infantum | N | Gr | PCR | Cat (Felis catus) | ND | Italy | Europe | [62] | |
| 2012-2013 | L. infantum | P | Gr | qPCR | Cat (Felis catus) | PCR, qPCR | Italy | Europe | [104] | ||
| Ixodes ventalloi | 2011 and 2013 | L. infantum | P | Gr | PCR | Cat (Felis catus) | ND | Italy | Europe | [62] | |
| 2012-2013 | L. infantum | P | Gr | qPCR | Cat (Felis catus) | PCR, qPCR | Italy | Europe | [104] | ||
| Rhipicephalus (Boophilus) microplus | 2012 | L. guyanensis | P | Gr | PCR ; HRM-PCR | Tapiridae (Tapirus terrestris) Tayassuidae (Pecari tajacu) | ND | Peru | South America | [63] | |
| 2012 | L. infantum | N | Gr | PCR | Sheep, cattle and dogs | ND | China | Asia | [110] | ||
| Leishmania sp. | P | Gr | qPCR | Collared peccary (Pecari tajacu); Tortoise (Chelonoidis denticulata) | ND | Peru | South America | [105] | |||
| 2014-2015 | L. martiniquensis | P | Gr | HT-qPCR | Cattle | ND | Guadeloupe | Island | [56] | ||
| 2014-2015 | L. martiniquensis | P | Gr | HT-qPCR | Cattle | ND | Martinique | Island | [56] | ||
| Rhipicephalus bursa | 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | |
| 2007-2008 | L. infantum | N | Gr | PCR | Sheep, goats,cattle, horses, deer | ND | Italy | Europe | [108] | ||
| Rhipicephalus pusillus | 2011 and 2013 | L. infantum | P | Gr | PCR | Cat (Felis catus) | ND | Italy | Europe | [62] | |
| 2007-2008 | L. infantum | N | Gr | PCR | Hedgehogs | ND | Italy | Europe | [108] | ||
| 2012-2013 | L. infantum | P | Gr | qPCR | Cat (Felis catus) | PCR, qPCR | Italy | Europe | [104] | ||
| Rhipicephalus sanguineus | 2012-2013 | L. infantum | P | Gr | qPCR | Cat (Felis catus) | PCR, qPCR | Italy | Europe | [104] | |
| 2016-2017 | Leishmania major | P | Gr | PCR, Sequencing | Rodents (Rhombomys opimus; Nesokia indica) | ND | Iran | Asia | [98] | ||
| 2013 | L. chagasi (syn L. infantum) | P | H ; Gr | Microscopy, PCR | Dog (Canis lupus) | ND | Brazil | South America | [92] | ||
| NS | L. braziliensis | P | Gr | PCR, qPCR | Dog (Canis lupus) | ND | Brazil | South America | [76] | ||
| 2007 | L. infantum | P | Gr | PCR ; qPCR | Dog (Canis lupus) | IFAT, PCR, qPCR | Brazil | South America | [100] | ||
| 2007 | L. infantum | P | Gr, SG | PCR ; qPCR ; sequencing | Dog (Canis lupus) | ND | Italy | Europe | [74] | ||
| 2008 | L. infantum | P | Gr, SG | PCR; qPCR; sequencing | Dog (Canis lupus) | ND | Brazil | South America | [74] | ||
| 2007-2008 | L. infantum | P | Gr | PCR | Dog (Canis lupus) | ELISA | Brazil | South America | [103] | ||
| 2007-2008 | L. infantum | N | Gr | PCR | Dog (Canis lupus) | ND | Italy | Europe | [108] | ||
| 2008-2009 | L. infantum | P | Gr | PCR, RT-PCR, Sequencing | Dog (Canis lupus) | ELISA, PCR | Brazil | South America | [65] | ||
| Rhipicephalus sanguineus | 2007 | L. infantum | P | Gr | qPCR | Dog (Canis lupus) | ELISA, PCR | Italy | Europe | [66] | |
| 2006-2007 | L. infantum | P | Gr | qPCR | Dog (Canis lupus) | ELISA, PCR | Italy | Europe | [67] | ||
| 2011-2012 | L. infantum | N | NS | Infestation | Dog (Canis lupus) | IFAT, ELISA | Brazil | South America | [111] | ||
| NS | L. infantum | P | Gr | PCR, qPCR, seuqencing | Dog (Canis lupus) | PCR, qPCR | Brazil | South America | [77] | ||
| 2012 | L. infantum | P | Gr | PCR | Dog (Canis lupus) | IFAT, ELISA | Brazil | South America | [54] | ||
| 2013 | L. infantum | P | Gr | PCR, Sequencing | Dog (Canis lupus) | PCR, Sequencing | Brazil | South America | [55] | ||
| 2011 | L. infantum | P | G | PCR, RFLP, seuqencing, Parasit culture | Dog (Canis lupus) | DPP, IFAT, ELISA, PCR | Brazil | South America | [57] | ||
| 2011 | L. infantum | P | SG | PCR, RFLP, Sequencing, Parasit culture | Dog (Canis lupus) | DPP, IFAT, ELISA, PCR | Brazil | South America | [57] | ||
| 2011 and 2013 | L. infantum | P | Gr | PCR | Cat (Felis catus) | ND | Italy | Europe | [62] | ||
| 2002 | Leishmania sp. | P | G | Microscopy | Dog (Canis lupus) | IFAT | Brazil | South America | [106] | ||
| 2009 | Leishmania sp. | N | NS | present of tick or no | Dog (Canis lupus) | IFAT | Brazil | South America | [61] | ||
| 2012-2014 | Leishmania sp. | N | Gr | qPCR | Dog (Canis lupus) | qPCR | China | Asia | [112] | ||
| NS | Leishmania sp. | P | G ; O ;SG | IHC, qPCR, IHC | Dog (Canis lupus) | ND | Brazil | South America | [69] | ||
| NS | Leishmania sp. | P | G ; O ; SG | IHC, qPCR, IHC | Dog (Canis lupus) | ND | Brazil | South America | [68] | ||
| NS | L. kala azar£ | P | NS | NS | Human | NS | France | Europe | [101] | ||
| 2008 | L. infantum | N | Gr | qPCR | Dog (Canis lupus) | IFAT | Brazil | South America | [73] | ||
| Rhipicephalus sanguineus s.L. | NS | L. infantum | P | Gr | PCR, Sequencing | $Flagging & *Animals | ND | Israel | Asia | [59] | |
| Rhipicephalus turanicus | 2007-2008 | L. infantum | N | Gr | PCR | Sheep; Goats | ND | Italy | Europe | [108] | |
| 2014 | Leishmania sp. | N | Gr | PCR | Human | ND | Turkey | Asia | [109] | ||
| NS | L. infantum | P | Gr | PCR, Sequencing | $Flagging & *Animals | ND | Israel | Asia | [59] | ||
NS : unspecified ; P : positive ; N : negative ; F: faeces; G: gut; Gr: crushing; H: haemolymph; O: ovaries; SG: salivary glands ; PCR : conventional Polymerase chain reaction, RFLP : restriction fragment length polymorphism ; qPCR : real time PCR; RT-PCR : reverse transcriptase PCR; HRM-PCR : High Resolution Melting PCR ; HT-qPCR : High-throughput microfluidic PCR; Sequencing : Capillary sanger sequencing, IFAT : Indirect Fluorescent Antibody Test ; ELISA : Enzyme Linked Immunosorbent Assay ; DPP : rapid immunochromatographic test ; IHC : immunohistochemistry * Animals : dogs (Canis familiaris); Tortoise (Testudo graeca); hedgehogs (Erinaceus concolor); badger. * L. kala azar refers to members of the L. donovani complex (L. infantum and L. donovani) without any other information on parasite typing at the time of the study. We therefore use the term proposed by the author. $Flagging: refers to the use of a method to collect questing ticks.
Table 3.
Pooled prevalence of parasites detections in ticks by sampling year, continent, host, tick genus/species, method, pariste genus and species, localization.
Table 3.
Pooled prevalence of parasites detections in ticks by sampling year, continent, host, tick genus/species, method, pariste genus and species, localization.
| Category | Variable | No. of studies | No. of tested | No. of positive | % [95% CI] | Heterogeneity | Univariate meta regression | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ꭕ2 | P-value | I2 (%) | P-value | R2 (%) | I2-res (%) | ||||||
| Sampling year |
2000 or before 2001-2010 2011-2019 2020 or after |
2 5 23 12 |
6470 480 3043 1721 |
19 130 518 96 |
0.22 [0.16-0.45] 12.19 [0.87-32.08] 21.13 [7.7-23.77] 11.65 [0.3-30.84] |
1.14 126.37 818.31 289.5 |
<0.0001 <0.0001 <0.0001 <0.0001 |
12.0 96.8 97.3 96.2 |
0.000*** |
0.00 | 98.74 |
| Continent |
Africa Asia Europe Island South America |
4 7 10 1 20 |
4392 1165 3693 478 1986 |
22 44 69 2 626 |
2.44 [0.00-9.54] 3.11 [0.00-20.83] 4.36 [1.49-9.06] 0.42 [0.01-1.26] 33.35 [14.27-37.83] |
23.24 131.61 158.52 0.000 828.68 |
< 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 |
85.8 95.4 94.3 -- 96.1 |
0.0056** |
23.77 | 97.78 |
| Host (Family) |
Several# Bovidae Canidae Felidae Other |
6 6 20 2 7 |
4894 1192 2184 221 722 |
43 44 595 19 57 |
2.83 [0.0-8.60] 3.92 [1.20-7.85] 27.92 [14.51-43.57] 8.47 [5.07-12.60] 21.23 [0.0-62.24]0 |
80.46 24.26 1071.56 0.81 252.480 |
< 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 |
93.8 88.2 98.2 0.0 97.6 |
0.0007** |
0.00 | 98.34 |
| Tick Genus |
Amblyomma Dermacentor Haemaphysalis Hyalomma Ixodes Rhipicephalus |
10 3 4 6 9 33 |
328 23 360 639 6906 3271 |
27 0 00 17 42 668 |
8.78 [0-29.41] 0.00 [0-5.25] 0.00 1.20 [0-14.55] 1.91 [0-10.06] 19.06 [9.64-30.39] |
100.12 0.41 1.58 70.55 71.41 1412.59 |
< 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 |
91.0 0.0 0.0 92.9 88.8 97.7 |
0.0001*** |
2.20 | 97.09 |
| Tick species |
A. cajennense A. ovale A. sabanerae A. tigrinum A. variegatum Amblyomma spp. D. marginatus H. aegyptium H. detritum H. dromedarii H. excavatum H. lusitanicum H. marginatum Hyalomma sp. Ha. longicornis Ha. parva Ha. punctata Ha. sulcata Haemaphysalis. sp. I. ricinus I. ventalloi Ixodes sp. R. (Boophilus) R. bursa R. pusillus R. sanguineus R. turanicus Rhipicephalus sp. |
1 1 1 2 2 2 3 2 1 2 3 1 4 2 1 1 1 2 1 8 1 1 6 3 2 21 4 1 |
131 10 10 28 165 43 21 32 1 45 291 3 171 81 308 41 6 4 13 6580 62 5 1025 23 18 2222 158 43 |
0 0 9 0 11 4 0 12 1 5 0 0 0 0 0 0 0 0 0 32 3 0 62 0 3 583 10 3 |
0.00 [0-7.04] 0.00 [0-16.52] 90.00 [61.91-100] 24.86 [0-95.91] 3.09 [0-24.26] 0.00 [0-0.64] 0.00 [0-5.25] 32.69 [11.66-56.75] 100 [0-100] 16.56 [0-88.80] 0.00 0 [0-50] 0.00 0.00 [0-2.36] 0.00 [0-0.58] 0.00 [0-4.15] 0.00 [0-26.80] 0.00 [0-78.74] 0.00 [0-12.82] 1.36 [0-10.39] 6.45 [1.42-14.19] 0.00 [0-31.73] 0.00 [0-3.92] 13.54 [0-47.12] 17.65 [2.57-39.97] 25.15 [11.67-41.19] 2.48 [0-1467] 6.98 [0.9-16.93] |
3.86 0.00 0.00 16.89 0.43 11.54 0.59 0.46 0.00 17.10 1.45 0.00 2.04 0.01 0.00 0.00 0.00 0.07 0.00 64.66 0.00 0.00 0.51 185.13 0.02 1092.57 23.70 0.00 |
< 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 |
-- 0.0 -- 94.1 91.30.0 0.0 0.0 -- 94.2 0.0 -- 0.0 0.0 -- -- -- 0.0 -- 87.6 -- -- 0.0 22.2 0.0 97.9 87.3 -- |
0.0001*** |
3.29 | 97.50 |
| Detection method |
Microscopy Molecular |
3 39 |
6533 5181 |
20 743 |
0.1 [0.0-0.23] 17.55 [9.31-27.44] |
3.47 1937.17 |
< 0.0001 < 0.0001 |
42.4 98.1 |
0.000*** | 5.37 | 98.84 |
| Parasite genus |
Leishmania Trypanosoma |
30 12 |
4569 7739 |
719 93 |
18.87 [9.12-30.75] 4.62 [1.14-9.79] |
1642.76 203.03 |
< 0.0001 < 0.0001 |
98.294.6 |
0.0046** | 7.10 | 98.88 |
| Parasite species |
L. braziliensis L. chagas (syn L. infantum) L. guyanensis L. infantum L. major L. martiniquensis Leishmania sp. T. caninum T. congolense T. cruzi T. evansi T. theileri T. vivax |
1 1 1 18 1 1 6 1 1 3 4 2 3 |
75 444 81 2124 1 578 683 9 33 583 892 6220 441 |
32 107 3 323 1 2 194 1 4 8 38 14 42 |
42.67 [31.64-64.66] 24.10 [20.23-28.19] 12.12 [2.79-25.84] 15.44 [7.18-25.89] 100 [0-100] 0.35 [0-1.04] 51.82 [8.42-93.57] 11.11 [0-41.77] 3.70 [0.47-9.18] 2.39 [0-10.25] 6.44 [0-23.33] 0.20 [0.12-0.36] 10.88 [2.91-22.73] |
0.00 0.00 0.00 679.35 0.08 0.00 0.00 0.00 0.00 895.52 13.00 73.71 22.82 |
< 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 |
-- -- -- 97.5 -- -- 94.4 -- -- 91.2 95.9 0.0 84.6 |
0.000*** | 9.25 |
98.97 |
| Localization in ticks |
H G GR GR-SG H-GR SG O |
2 3 35 1 1 2 2 |
6470 209 4532 95 444 110 165 |
19 164 478 11 107 35 74 |
0.29 [0.16-0.45] 74.32 [28.48-99.99] 13.45 [6.62-21.87] 11.58 [5.82-18.88] 24.10 [20.23-28.19] 31.79 [23.31-40.91] 44.48 [37.27-52.53] |
1.14 312.09 1234.08 0.00 0.00 0.00 0.67 |
< 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 < 0.0001 |
12.0 97.3 97.2 -- -- 0.0 0.0 |
0.000*** | 0.00 | 98.89 |
G: gut; Gr: whole body; H: haemolymph; O: ovaries; SG: salivary glands *0.05; **0.01; ***0.001, # In these studies, various tick species were allowed to feed on diverse infected host ( Swiss mice, Meadow voles, Rats, Mice, Rabbits, Camel, Dog, Guinea pig or White rat). In these papers, it was not possible to retrieve the information on the host origin of the ticks that were tested. .
Table 4.
Pooled rate of parasite detections in ticks in experimental transmission experiments.
| Acquisition of pathogens via tick blood feeding | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Category | Variable | No. of studies | No. of positive | % [95% CI] | Heterogeneity | Univariate meta regression | |||||||||||||
| ꭕ2 | P-value | I2 (%) | P-value | R2 (%) | I2-res (%) | ||||||||||||||
| Ticks Family | 0.0976 | 0.00 | 0.00 | ||||||||||||||||
| Argasidae | 16 | 16 | 100 [81.65-100] | 0.00 | 0.9966 | 0.0 | |||||||||||||
| Ixodidae | 21 | 15 | 78.01 [48.91-98.34] | 15.86 | 0.9966 | 0.0 | |||||||||||||
| Ticks Genus | 0.3905 | 0.00 | 0.00 | ||||||||||||||||
| Amblyoma | 1 | 1 | 100 [0-100] | 0.00 | 0.9938 | -- | |||||||||||||
| Hyalomma | 4 | 3 | 82.18 [16.67-100] | 2.78 | 0.9938 | 0.0 | |||||||||||||
| Ornithodoros | 15 | 15 | 100 [80.68-100] | 0.00 | 0.9938 | 0.0 | |||||||||||||
| Rhipicephalus | 16 | 11 | 74.57 [40.89-98.60] | 12.72 | 0.9938 | 0.0 | |||||||||||||
| Ticks species | 0.6118 | 0.00 | 0.00 | ||||||||||||||||
| A. americanum | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| B. decoloratus | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| D. andersoni | 2 | 2 | 100 [21.62-100] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| H. A. anatolicum | 3 | 2 | 72.14 [2.45-100] | 2.47 | 0.9989 | 18.9 | |||||||||||||
| H. A. excavatum | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| H. dromaderii | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| H. impressum | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| O. amblus | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| O. crossi | 4 | 4 | 100 [48.72-100] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| O. furcosis | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| O. hermsi | 2 | 0 | 0 [0-78.74] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| O. lahorensis | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| O. moubata | 7 | 7 | 100 [65.46-100] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| O. parkeri | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| O. savignyi | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| O. talaje | 1 | 0 | 0 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| O. turanicus | 2 | 2 | 100 [21.62-100] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| O. venzualensis | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| R. sanguneus | 13 | 11 | 91.99 [58.62-100] | 6.26 | 0.9989 | 0.0 | |||||||||||||
| R. sanguneus s.I. | 2 | 0 | 0 [0-78.74] | 0.00 | 0.9989 | 0.0 | |||||||||||||
| R. pulchellus | 1 | 1 | 100 [0-100] | 0.00 | 0.9989 | -- | |||||||||||||
| Parasite genus | 0.0794 | 0.00 | 0.00 | ||||||||||||||||
| Leishmania | 14 | 9 | 69.10 [62.64-97.17] | 11.90 | 0.9974 | 0.0 | |||||||||||||
| Trypanosoma | 22 | 21 | 99.11 [80.73-100] | 3.53 | 0.9974 | 0.0 | |||||||||||||
| Parasite Species | 0.7192 | 0.00 | 0.00 | ||||||||||||||||
| L. donovani | 1 | 1 | 100 [0-100] | 0.00 | 0.9928 | -- | |||||||||||||
| L. chagasi (syn L.infantum) | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9928 | 0.0 | |||||||||||||
| L. infantum | 5 | 4 | 87.58 [29.26-100] | 2.96 | 0.9928 | 0.0 | |||||||||||||
| L. kala azar£ | 4 | 1 | 17.82 [0-83.24] | 2.78 | 0.9928 | 0.0 | |||||||||||||
| L. major | 1 | 1 | 100 [0-100] | 0.00 | 0.9928 | -- | |||||||||||||
| Leishmania sp. | 1 | 1 | 100 [0-100] | 0.00 | 0.9928 | -- | |||||||||||||
| T. bruci | 1 | 1 | 100 [0-100] | 0.00 | 0.9928 | -- | |||||||||||||
| T. cruzi | 8 | 8 | 100 [68.71-100] | 0.00 | 0.9928 | 0.0 | |||||||||||||
| T. evansi | 7 | 6 | 92.94 [46.20-100] | 3.17 | 0.9928 | 0.0 | |||||||||||||
| T. gambiense | 1 | 1 | 100 [0-100] | 0.00 | 0.9928 | -- | |||||||||||||
| T. lewisi | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9928 | 0.0 | |||||||||||||
| T. rhodesciense | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9928 | 0.0 | |||||||||||||
| T. theileri | 4 | 3 | 82.18 [16.67-100] | 2.78 | 0.9928 | 0.0 | |||||||||||||
| T. theileri like | 1 | 1 | 100 [0-100] | 0.00 | 0.9928 | -- | |||||||||||||
| Donor host family | 0.9455 | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 4 | 3 | 82.18 [16.67-100] | 2.78 | 0.9471 | 0.0 | |||||||||||||
| Canidae | 13 | 9 | 75.34 [37.81-99.71] | 10.25 | 0.9471 | 0.0 | |||||||||||||
| Camelidae | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9471 | 0.0 | |||||||||||||
| Rodentia | 15 | 14 | 93.73 [63.97-100] | 42.76 | 0.9471 | 0.0 | |||||||||||||
| ND | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9471 | 0.0 | |||||||||||||
| Other | 1 | 1 | 100 [0-100] | 0.00 | 0.9471 | -- | |||||||||||||
| Ticks | 1 | 1 | 100 [0-100] | 0.00 | 0.9471 | -- | |||||||||||||
| Infection via the injection of ticks infected material | |||||||||||||||||||
| Ticks Family | 0.2884 | 0.00 | 0.00 | ||||||||||||||||
| Argasidae | 9 | 7 | 85.25 [41.62-100] | 5.76 | 0.6062 | 0.0 | |||||||||||||
| Ixodidae | 13 | 7 | 55.23 [18.49-89.66] | 11.96 | 0.6062 | 0.0 | |||||||||||||
| Ticks Genus | 0.5954 | 0.00 | 0.00 | ||||||||||||||||
| Hyalomma | 4 | 2 | 50.0 [0-100] | 3.70 | 0.5378 | 18.9 | |||||||||||||
| Ornithodoros | 9 | 8 | 85.25 [41.62-100] | 5.76 | 0.5378 | 0.0 | |||||||||||||
| Rhipicephalus | 7 | 4 | 59.68 [11.06-99.18] | 6.34 | 0.5378 | 5.4 | |||||||||||||
| Ticks species | 0.2229 | 0.00 | 0.00 | ||||||||||||||||
| B. decoloratus | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| D. andersoni | 2 | 0 | 0 [0-78.74] | 0.00 | 0.8803 | 0.0 | |||||||||||||
| H. a. anatolicum | 3 | 2 | 72.14 [2.45-100 | 2.47 | 0.8803 | 18.9 | |||||||||||||
| H. a. excavatum | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| H. dromaderii | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| H. impressum | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. amblus | 1 | 1 | 100 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. crossi | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. furcosus | 1 | 1 | 100 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. hermsi | 2 | 0 | 0 [0-78.74] | 0.00 | 0.8803 | 0.0 | |||||||||||||
| O. moubata | 4 | 4 | 100 [48.72-100] | 0.00 | 0.8803 | 0.0 | |||||||||||||
| O. perkeri | 1 | 1 | 100 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. talaje | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. turanicus | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| O. turicata | 2 | 1 | 50.0 [0-100] | 1.85 | 0.8803 | 46.0 | |||||||||||||
| O. venzualensis | 1 | 1 | 100 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| R. pulchellus | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| R. sanguineus | 6 | 5 | 90.82 [38.83-100] | 3.08 | 0.8803 | 0.0 | |||||||||||||
| R. sanguineus s.I. | 1 | 0 | 0 [0-100] | 0.00 | 0.8803 | -- | |||||||||||||
| Parasite genus | 0.7666 | 0.00 | 0.00 | ||||||||||||||||
| Leishmania | 7 | 5 | 78.10 [27.34-100] | 5.29 | 0.5774 | 0.0 | |||||||||||||
| Trypanosoma | 14 | 9 | 69.10 [32.64-97.17] | 11.9 | 0.5774 | 0.0 | |||||||||||||
| Donor host Family | 0.7557 | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 4 | 2 | 50.0 [0-100] | 3.70 | 0.5180 | 18.9 | |||||||||||||
| Camelidae | 1 | 1 | 100 [0-100] | 0.00 | 0.5180 | -- | |||||||||||||
| Canidae | 5 | 4 | 87.58 [29.26-100] | 2.96 | 0.5180 | 0.0 | |||||||||||||
| Rodentia | 11 | 8 | 68.26 [27.33-98.66] | 9.42 | 0.5180 | 0.0 | |||||||||||||
| Receiver host Family | 0.5494 | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 3 | 2 | 72.14 [2.45-100] | 2.47 | 0.5387 | 18.9 | |||||||||||||
| Canidae | 1 | 1 | 100 [0-100] | 0.00 | 0.5387 | -- | |||||||||||||
| Other | 1 | 0 | 0 [0-100] | 0.00 | 0.5387 | -- | |||||||||||||
| Rodentia | 15 | 11 | 63.49 [28.31-93.43] | 13.32 | 0.5387 | 0.0 | |||||||||||||
| Ticks | 2 | 2 | 100 [21.26-100] | 0.00 | 0.5387 | 0.0 | |||||||||||||
| Parasite species | 0.2614 | 0.00 | 0.00 | ||||||||||||||||
| L. chagasi (Syn L. infantum) | 1 | 1 | 100 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| L. infantum | 1 | 1 | 100 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| L. kala azar£ | 4 | 2 | 50.0 [0-100] | 3.70 | 0.8397 | 18.9 | |||||||||||||
| Leishmania sp. | 1 | 1 | 100 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| T. brucei | 1 | 0 | 0 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| T. cruzi | 6 | 6 | 100 [61.37-100] | 0.00 | 0.8397 | 0.0 | |||||||||||||
| T. evansi | 1 | 0 | 0 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| T. lewisi | 1 | 0 | 0 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| T. rhodeseinse | 1 | 0 | 0 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| T. theileri | 4 | 3 | 82.18 [16.76-100] | 2.78 | 0.8397 | 0.0 | |||||||||||||
| T. theileri like | 1 | 0 | 0 [0-100] | 0.00 | 0.8397 | -- | |||||||||||||
| Transmission through blood feeding | |||||||||||||||||||
| Ticks Family | 0.4962 | 0.00 | 0.00 | ||||||||||||||||
| Argasidae | 10 | 3 | 23.71 [0-66.41] | 7.77 | 0.5314 | 0.9 | |||||||||||||
| Ixodidae | 11 | 5 | 34.33 [0.0-94] | 10.09 | 0.5314 | 0.0 | |||||||||||||
| Ticks Genus | 0.7745 | 0.00 | 0.00 | ||||||||||||||||
| Hyalomma | 4 | 2 | 50.0 [0-100] | 3.70 | 0.4677 | 18.9 | |||||||||||||
| Ornithodoros | 10 | 3 | 23.71 [0-66.41] | 7.77 | 0.4677 | 0.0 | |||||||||||||
| Rhipicephalus | 7 | 3 | 40.32 [0.48-88.94] | 6.32 | 0.4677 | 5.4 | |||||||||||||
| Parasite genus | 0.5011 | 0.00 | 0.00 | ||||||||||||||||
| Leishmania | 5 | 2 | 36.51 [0-92.81] | 4.44 | 0.9952 | 9.9 | |||||||||||||
| Trypanosoma | 16 | 8 | 33.22 [5.25-67.40] | 13.88 | 0.9952 | 0.0 | |||||||||||||
| Donnor host family | 0.0405* | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 4 | 2 | 50.0 [0-100] | 3.70 | 0.9017 | 18.9 | |||||||||||||
| Camelidae | 2 | 2 | 100 [21.26-100] | 0.00 | 0.9017 | 0.0 | |||||||||||||
| Canidae | 7 | 4 | 59.68 [11.06-99.18] | 6.34 | 0.9017 | 5.4 | |||||||||||||
| Rodentia | 8 | 0 | 0 [0-31.29] | 0.00 | 0.9017 | 0.0 | |||||||||||||
| Receiver host family | 0.0678 | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 3 | 2 | 72.14 [2.45-100] | 2.47 | 0.8461 | 18.9 | |||||||||||||
| Canidae | 3 | 2 | 72.14 [2.45-100] | 2.47 | 0.8461 | 18.9 | |||||||||||||
| Other | 2 | 2 | 100 [21.26-100] | 0.00 | 0.8461 | 0.0 | |||||||||||||
| Rodentia | 13 | 4 | 8.01 [0-41.38] | 6.26 | 0.8461 | 0.0 | |||||||||||||
| Ticks species | 0.6077 | 0.00 | 0.00 | ||||||||||||||||
| B. decoloratus | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| D. andersoni | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| H. a. anatolicum | 3 | 2 | 72.14 [2.45-100] | 2.47 | 0.6280 | 18.9 | |||||||||||||
| H. a. excavatum | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| H. dromaderii | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| H. impressum | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| O. crossi | 4 | 3 | 82.18 [16.67-100] | 2.78 | 0.6280 | 0.0 | |||||||||||||
| O. hermsi | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| O. lahorensis | 2 | 2 | 100 [21.26-100] | 0.00 | 0.6280 | 0.0 | |||||||||||||
| O. moubata | 3 | 0 | 0 [0-61.92] | 0.00 | 0.6280 | 0.0 | |||||||||||||
| O. savigny | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| O. talaje | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| O. turanicus | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| O. turicata | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| O. venzualensis | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| R. pulchellus | 1 | 0 | 0 [0-100] | 0.00 | 0.6280 | -- | |||||||||||||
| R. sanguineus | 6 | 3 | 50.0 [2.78-97.22] | 5.55 | 0.6280 | 9.9 | |||||||||||||
| Parasite species | 0.5529 | 0.00 | 0.00 | ||||||||||||||||
| L. infantum | 2 | 1 | 50.0 [0-100] | 1.85 | 0.5302 | 46.0 | |||||||||||||
| L. kala azar£ | 2 | 0 | 0 [0-78.74] | 0.00 | 0.5302 | 0.0 | |||||||||||||
| Leishmania sp. | 1 | 1 | 100 [0-100] | 0.00 | 0.5302 | -- | |||||||||||||
| T. cruzi | 6 | 1 | 9.18 [0-61.17] | 3.08 | 0.5302 | 0.0 | |||||||||||||
| T. evansi | 6 | 3 | 50.0 [2.78-97.22] | 5.55 | 0.5302 | 9.9 | |||||||||||||
| T. lewisi | 1 | 0 | 0 [0-100] | 0.00 | 0.5302 | -- | |||||||||||||
| T. theileri | 3 | 1 | 27.86 [0-97.55] | 2.47 | 0.5302 | 18.9 | |||||||||||||
| T. theileri like | 1 | 1 | 100 [0-100] | 0.00 | 0.5302 | -- | |||||||||||||
| Vertical transmission | |||||||||||||||||||
| Ticks Family | 0.2809 | 0.00 | 0.00 | ||||||||||||||||
| Argasidae | 1 | 0 | 0 [0-100] | 0.00 | 0.4606 | -- | |||||||||||||
| Ixodidae | 12 | 7 | 61.27 [22.41-94.58] | 10;79 | 0.4606 | 0.0 | |||||||||||||
| Ticks Genus | 0.5422 | 0.00 | 0.00 | ||||||||||||||||
| Hyalomma | 2 | 1 | 50.0 [0-100] | 1.85 | 0.3787 | 46.0 | |||||||||||||
| Ornithodoros | 1 | 0 | 0 [0-100] | 0.00 | 0.3787 | -- | |||||||||||||
| Rhipicephalus | 10 | 6 | 63.49 [21.05-97.58] | 8.88 | 0.3787 | 0.0 | |||||||||||||
| Ticks species | 0.4394 | 0.00 | 0.00 | ||||||||||||||||
| H. a. anatolicum | 2 | 1 | 50.0 [0-100] | 1.85 | 0.4143 | 46.0 | |||||||||||||
| O. moubata | 1 | 0 | 0 [0-100] | 0.00 | 0.4143 | -- | |||||||||||||
| R. pulchellus | 1 | 0 | 0 [0-100] | 0.00 | 0.4143 | -- | |||||||||||||
| R. sanguineus | 9 | 7 | 72.14 [27.07-100] | 7.40 | 0.4143 | 0.0 | |||||||||||||
| Parasite genus | 0.0634 | 0.00 | 0.00 | ||||||||||||||||
| Leishmania | 8 | 7 | 82.18 [35.16-100] | 5.55 | 0.6668 | 0.0 | |||||||||||||
| Trypanosoma | 5 | 1 | 12.42 [0-70.74] | 2.96 | 0.6668 | 0.0 | |||||||||||||
| Donor host family | 0.0425* | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 3 | 1 | 27.86 [0-97.55] | 2.47 | 0.8446 | 18.9 | |||||||||||||
| Canidae | 7 | 6 | 92.94 [46.20-100] | 3.17 | 0.8446 | 0.0 | |||||||||||||
| Rodentia | 3 | 0 | 0 [0-61.92] | 0.00 | 0.8446 | 0.0 | |||||||||||||
| Receiver host family | 0.3582 | 0.00 | 0.00 | ||||||||||||||||
| Bovidae | 2 | 1 | 50.0 [0-100] | 1.85 | 0.4748 | 46.0 | |||||||||||||
| Canidae | 1 | 1 | 100 [0-100] | 0.00 | 0.4748 | -- | |||||||||||||
| ND | 4 | 3 | 82.18 [16.67-100] | 2.78 | 0.4748 | 0.0 | |||||||||||||
| Rodentia | 5 | 1 | 12.42 [0-70.74] | 2.69 | 0.4748 | 0.0 | |||||||||||||
| Ticks | 1 | 1 | 100 [0-100] | 0.00 | 0.4748 | -- | |||||||||||||
| Parasite species | 0.3655 | 0.00 | 0.00 | ||||||||||||||||
| L. chagasi (Syn L. infantum) | 2 | 1 | 50.0 [0-100] | 1.85 | 0.5046 | 46.0 | |||||||||||||
| L. infantum | 3 | 3 | 100 [38.08-100] | 0.00 | 0.5046 | 0.0 | |||||||||||||
| L. major | 1 | 0 | 0 [0-100] | 0.00 | 0.5046 | -- | |||||||||||||
| L. kala azar£ | 1 | 1 | 100 [0-100] | 0.00 | 0.5046 | -- | |||||||||||||
| Leishmania sp. | 1 | 1 | 100 [0-100] | 0.00 | 0.5046 | -- | |||||||||||||
| T. cruzi | 1 | 0 | 0 [0-100] | 0.00 | 0.5046 | -- | |||||||||||||
| T. evansi | 1 | 0 | 0 [0-100] | 0.00 | 0.5046 | -- | |||||||||||||
| T.theileri | 3 | 1 | 27.86 [0-97.55] | 2.47 | 0.5046 | 18.9 | |||||||||||||
*0.05; **0.01; ***0.001 £L. kala azar refers to members of the L. donovani complex (L. infantum and L. donovani) without any other information on parasite typing at the time of the study. We therefore use the term proposed by the author. # ticks (receiver host) were infected via the inoculation of A. americanum-infected hemolymph (donor host).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



