
Submitted:
19 July 2024
Posted:
22 July 2024
You are already at the latest version
Abstract
Selenium nanoparticles (SeNPs) can mitigate cadmium (Cd2+) toxicity in crop cultivation and aquaculture. This study investigates the impact of SeNPs on Cd-induced toxicity in Chlamydomonas reinhardtii, a model organism, aiming to mitigate Cd damage in crops. SeNPs were synthesized using sodium carboxymethyl cellulose (CMC) as a dispersant and vitamin C to reduce H2SeO3. Characterization by transmission electron microscopy (TEM) and nanoparticle granulator revealed uniform, stable spherical SeNPs. Cd2+ and SeNPs of varying concentrations were introduced into culture media of wild type (CC125mt+) and cell wall defect type (CC400mt+) Chlamydomonas reinhardtii. Growth curves, photosynthetic pigment content, enzyme activities (T-SOD and CAT), MDA content, and ROS levels were assessed. Fluorescence induction kinetics, QA oxidation kinetics, and Cd2+ content in cells were measured to evaluate SeNPs' effect on Cd toxicity. Results showed SeNPs exacerbated Cd2+ toxicity in CC125mt+ by inhibiting growth and increasing catalase activity, indicating enhanced oxidative damage. SeNPs intensified PSII damage by Cd2+ and increased intracellular Cd2+ content, possibly exacerbating toxicity. SeNPs had no effect on Cd2+ toxicity in CC400mt+. Overall, SeNPs exacerbated Cd2+ toxicity in wild type CC125mt+ but not in cell wall defective CC400mt+. These findings caution against indiscriminate use of nano-selenium to mitigate Cd damage, stressing the importance of careful target selection, dosage, and range considerations.
Keywords:
SeNPs
; cell wall
; enhanced toxicity
; selenium doses
; nanomaterial safety
1. Introduction
The safety of Marine environment affects all aspects of human life. Microalgae is the most important primary producer in the water environment ecosystem, which is widely existed in the water environment and plays an important role in maintaining the stability of the ecosystem. Heavy metal pollutants entering the water environment through various ways will cause physiological changes in their growth process. Microalgae are the lowest nutrient level in the water environment, and they accumulate harmful substances such as heavy metals into the life network, so it is urgent to solve the problem of heavy metal pollution of microalgae.
Cadmium, a toxic element, may contribute bad influence to aquatic plants. Cadmium stress causes oxidative stress in microalgae. Excessive Reactive Oxygen Species, such as superoxide anion free radicals (O2·), hydroxyl radicals (OH·), and hydrogen peroxide (H2O2), generated from oxidative stress, can lead to lipid peroxidation in microalgae, damage the structures of protein and nucleic acid, interfere the synthesis of photosynthetic pigments, inhibit photosynthesis and respiration and finally cause loss of normal growth and metabolism function of microalgae and change the composition of microalgae species[1]. In addition, the photosynthetic system of algae will also be adversely affected by cadmium stress. The basic processes of energy conversion and carbon assimilation in photosynthesis occurs in Chloroplast.The thylakoid membrane in Chloroplast is a lipid bilayer membrane required for energy conversion during photosynthesis. Photosystem I (PSI), Photosystem II (PSII), ATP synthase complex and cytochrome b6f complex (Cyt b6f) are the four membrane-bound supramolecular complexes which directly participate in photosynthesis. The toxic effect of cadmium on microalgae is mainly manifested as the inhibition of the reaction center and electron transport of PSII. This inhibition causes toxicity to both the electron donor side and the electron acceptor side of PSII, ultimately inhibiting photosynthesis[2].
Selenium is one of the most important microelement for human and animals. Selenium is an important component of glutathione peroxidase, which is closely related to antioxidant and immunity in vivo. Selenium can remove free radicals and inhibit the production of Reactive Oxygen Species[3,4,5]. Lack of selenium may lead to cardiovascular disease, accelerated aging, compromised immune systems and other problems, but consuming too much may bring toxic reactions. There are a number of clinical studies show that selenium has the function of alleviating heavy metal toxicity in plants[6,7,8] and animals[9,10]. However, the range between nutritional and toxic doses of inorganic and organic selenium in nature is narrow, which brings difficulties in the practical application of selenium. Research has shown that zero-valent elemental SeNPs have unique physical and chemical properties, showing high biological activity, good bioavailability, and low acute toxicity[11,12]. SeNPs have been found being able to scavenge free radicals and chelate metal ions, thus altering the pathway of cell protection and effectively improving the toxicity induced by cadmium chloride. In addition, SeNPs also has certain therapeutic effects on anticancer [13,14,15], anti-inflammatory [16], antibacterial[17], antiviral[18] and therapeutic effects on Alzheimer's disease[19]. Many studies have shown that nanomaterials can reduce the cytotoxicity of cadmium to algae[20] and inhibit the accumulation of cadmium in algae [21].
Nanomaterials can reduce the accumulation of cadmium in organisms within a certain concentration, but nanomaterials themselves can also cause oxidative stress in organisms [22,23,24,25].
Some research suggest that the shading effect, generation of reactive oxygen species, membrane damage and disintegration of pigments are the main reasons for nanoparticle toxicity to algae cells[26]. These toxic effects particularly affect the photosynthetic system of algae, including decrement of chlorophyll content[27] and disturbance of light reaction[28]. Therefore, it is very important to grasp the concentration of cadmium in the study of alleviating cadmium toxicity by nanomaterials.
The manufacture methods of SeNPs are mainly divided into chemical synthesis and biosynthesis.The chemical method is fast and efficient in preparation which is generally to reduce selenium dioxide, selenite or selenate with reducing agent, and introduce dispersant or stabilizer in the nucleation stage of the reaction to control the particle size within the nanometer range. Commonly used reducing agents are ascorbic acid, sodium sulfite, sodium thiosulfate, glutathione, hydrazine, etc., dispersant or stabilizer carbohydrates, proteins, polyphenols and so on. The preparation of SeNPs is usually done by polysaccharide soft template method. Commonly used polysaccharide soft template media mainly include chitosan[29,30], sodium carboxymethyl cellulose[31], sdium lauryl sulfate[32] et al. Sodium carboxymethyl cellulose (CMC) is a water-soluble polyanionic compound, which is widely and most used cellulose in the world. After rigorous biological and toxicological studies and experiments, FAO and WHO have approved the use of pure CMC in food products. Due to its safe and reliable nature, the amount of CMC is not restricted by the national food hygiene standard ADI. CMC is widely used in the food industry because of its good emulsification and dispersibility [33]. In the pharmaceutical industry, it is used as a drug carrier, and in the daily chemical industry, it is used as a binder and anti-reagglutination agent, etc.
Microalgae are the most important primary producers in the aquatic ecosystem. They exist widely in the water environment and play an important role in maintaining the stability of the ecosystem[34]. As the source of the food chain, microalgae easily introduce toxic substances or secondary metabolites from the environment into the food chain [35]. However, the study on microalgae as the research object to explore the toxic effects of SeNPs on cadmium pollution is still lacking. Chlamydomonas reinhardtii is a low grade plant with high light utilization efficiency and abundant nutrients. It is sensitive to toxins, small in size and fast in reproduction, so it is widely used in scientific research as a Model organism [36]. SeNPs has strong particularity compared with traditional selenium because of its nanometer structure. Therefore, the study of the effect of SeNPs on cadmium toxicity in algae cells is not only a new research direction in the field of environmental science and ecotoxicology, providing theoretical basis for reducing cadmium damage in crops, but also a new idea in the field of materials science to evaluate the biological effects of SeNPs.
2. Materials and Methods
2.1. Preparation and Property Determination of Sodium Carboxymethyl Cellulose - SeNPs Particles (CMC-SeNPs)
2.1.1. Preparation of CMC-SeNPs
Ten millilitres Se(IV) reserve solution (0.01 mol/L) and 10 mL CMC (0.5%(w/w)) reserve solution were mixed and stirred for 30 min, 10 mL Vitamin C (Vc) reserve solution (0.07mol/L) was added for reduction, and the solution was dissolved to 100 mL with ultra-pure water, and then stirred for 30 min until a stable red solution was obtained. That is, sodium carboxymethyl cellulose - SeNPs system. The final concentrations of each component in the reaction system were CMC 0.5‰ (w/w), Se(0) 0.001 mol/L, Vc 0.007 mol/L. The samples were stored at 4℃ for future use. The preparation of CMC-SeNPs is as Figure 1 shows.
2.1.2. Property Determination of CMC-SeNPs
Transmission electron microscopy (TEM) characterization and energy dispersive spectroscopy (EDS) analysis: After the preparation of the SeNPs system solution, 10 μL sample was dropped on the carbon-supported copper net, and after standing for 2 min, the excess sample solution on the copper net was absorbed by filter paper. After the copper net sample was dried naturally, the transmission electron microscopy (FEI Tecnai G2 F30) was used for observation under the accelerated voltage of 300 kV. The image acquisition detector was HAADF probe, and EDS was used for element analysis.
Characterization of SeNPs by dual wavelength method: The scanning pattern of the following components was determined after the baseline correction of the UV-VIS spectrophotometer: (1) CMC+Vc+H2SeO3; (2) CMC+Vc; (3) Vc+H2SeO3; (4) Vc; (5) Curve scanning is performed on the CMC. Two millilitres sample solution was added to the cupola and it was put into the sample room. Full wavelength scanning (UV3150, Shimadzu, Japan) at 200 ~ 800nm with step size of 1 nm was performed. The reaction system was quickly timed after mixing. In the experiment, 410 nm and 490 nm were selected as the measurement wavelengths of SeNPs solution. The absorbance of the system at 410 nm and 490 nm was tracked and measured by UVvisible spectrophotometer (UV5300, Shanghai Yuan Analysis Instrument Co., LTD.) within 6 h. The ratio of A410 to A490 was used to judge the change of particle size over time, and the ratio of absorbance of the two sites (A410/A490) was used as the basis to characterize the change of particle size of the product.
Particle size determination and Zeta potential analysis of SeNPs: Two millilitres SeNPs solution was added into the sample pool, and the outer wall of the sample pool was kept dry. The particle size was determined using Marvin Zetasizer Nano ZS90. Then Two millilitres SeNPs solution was added into the potential pool, and the preceding steps above were repeated. Samples were taken in triplicates.
2.2. Strain and Culture
Chlamydomonas reinhardtii were purchased from the Chlamydomonas Genetic Center of Duke University and the Chlamydomonas Genetic Center of Minnesota as wild type CC125mt+ (hereinafter referred to as "CC125") and cell wall defect type CC400mt+ (hereinafter referred to as "CC400") respectively. The purpose to choose these two kind of Chlamydomonas reinhardtii is that CC400 are more likely to ingest substances and the wild type are used as contrast. The culture of Chlamydomonas reinhardtii was performed using TAP (Tris-Acetate-Phosphate) liquid medium. TAP solid medium containing 1.5% agar was used for subculture and preservation of algae pure culture. The cell culture of Chlamydomonas reinhardtii was based on the culture method of Harris[37], that is, wild type CC125 and cell wall defective type CC400 were cultured respectively. On the 3rd day of logarithmic growth of Chlamydomonas reinhardtii in TAP liquid medium, the strain was inoculated onto solid medium and the initial cell count was controlled to 5×104/mL. After inoculation, the culture bottle was placed in an artificial climate incubator with white light at 25℃ and 100 μmol/m2/s for alternating light and dark (14 h light: 10 h dark), and the culture was shaken every 6 hours.
2.3. Determination of EC50
Based on the semi-maximum effect concentration (EC50) value of Chlamydomonas reinhardtii growth, Cd2+ concentration was identified as 30, 40, 50, 60, 70, 80 (μmol/L), SeNPs concentration was 0.02, 0.1, 0.5, 1, 5, 10 (μmol/L). Chlamydomonas reinhardtii were inoculated in TAP liquid culture medium at logarithmic stage (the third day), and the initial cell number was controlled to be consistent. The corresponding groups were added with different concentrations of Cd2+ or different concentrations of SeNPs. After algal cells were inoculated , 2 mL cell culture medium was sampled every 24 h in aseptic operation, and the absorbance value was measured at 680 nm by optical density method[38]. Measure was taken continuously for 4 days and the biomass of Chlamydomonas reinhardtii was calculated. (CK: algal fluid, but no Cd2+ and SeNPs).
2.4. Measurement of Growth Curves
There are four groups involved in this procedure:Control group,Cd2+ intervention group,SeNPs intervention group and Cd2+ and SeNPs combined intervention group. According to the EC50 concentration of Cd2+ obtained in the previous step, the Cd2+ concentration in this step was determined. According to the SeNPs obtained in the previous step, the EC50 concentration of the two kinds of algae was selected from low to high 5 concentrations to determine the growth curve. Chlamydomonas reinhardtii cells were inoculated in the logarithmic phase (day 3) of TAP liquid culture medium to control the same initial cell number, and Cd2+ and different concentrations of CMC-SeNPs were added to the corresponding groups. After inoculation of algae seeds, 2 mL was sampled every 24 h in aseptic operation. The absorbance value was determined at 680 nm by optical density method for 4 consecutive days.
2.5. Determination of Chlorophyll Content
After Chlamydomonas reinhardtii have been cultivated for 72h, 0.4 mL algae fluid (liquid contains a mixture of SeNPs algae) was taken to mix with 1.6 mL 80% acetone, and the mixture were shocked vortically after blending, then it was stood at 4 ℃, dark place away from light for 24 h. After that, the sample was centrifuged at 4 ℃ in 5000 RPM centrifugal for 5 min. Then 200 μL supernatant was taken, 80% acetone was used as reference solution, and the light absorption values at 663 nm, 646 nm and 470 nm were measured by enzyme label.
2.6. Oxidative Stress and Inflammation
After Chlamydomonas reinhardtii cells were treated as described in the “strain and culture” subsection, 10 mL of the algae cell liquid cultured for 72 hours was transferred to a 15 mL centrifuge tube and centrifuged at 6000 rpm for 10 min in a frozen centrifuge at 4℃. After centrifugation, the supernatant was quickly removed, and 1 mL of 0.05mol /L phosphoric acid buffer with pH 7.0 pre-cooled at 4℃ was added. The cells were re-suspended by shaking and transferred to a 2 ml centrifuge tube with a pipetting gun. The cells were broken in an ultrasonic cell fragmentation apparatus under the condition of ice bath for 5 s each time and 10 s at intervals for 5 min. The supernatant is absorbed and transferred to another 2 ml centrifuge tube, and it is stored at 4℃ for testing after being centrifuged at 4℃ and 3500 rpm for 5 min. Protein concentration in the supernatant was measured using a BCA Protein Assay Kit (Beyotime Biotechnology). Similarly, T-SOD, MDA, CAT and ROS were assayed using commercial assay kits (Beyotime Biotechnology) according to the manufacturer’s directions.
2.7. Fast Chlorophyll Fluorescence Induced Kinetics Curve (OJIP Curve)
Chlamydomonas reinhardtii cells were treated as described in the “strain and culture” subsection. The cell fluid of Chlamydomonas reinhardtii cultured for 72 h was dark acclimated for 5 min and placed in the measuring chamber of FL3500 chlorophyll fluorescence instrument, and the saturation light intensity was set as 100%. The changes of chlorophyll fluorescence within 1 s were measured and recorded by the instrument. The data was recorded every 10 μs for the first 2 ms and every 1 ms after 2 ms. Chlorophyll fluorescence induction kinetics was analyzed according to JIP-test.
2.8. QA- Determination of Reoxidation Kinetics
The sample was darkened for 5 min before determination. The flash time range measured by QA-reoxidation kinetics curve was 200 μs~10 s. Both the actinic light (30 μs) and the measurement light (2.5 μs) were red LED lights, and the saturation light intensity set by the measurement light was 100%. QA-reoxidation kinetic curve data were calculated and analyzed according to Li's method[39].
2.9. Determination of Cadmium and Selenium Content in Chlamydomonas Reinhardtii Cells
Chlamydomonas reinhardtii cells cultured in the subsection of “strain and culture” for 72 h were measured and their biomass was recorded. Forty millilitres Chlamydomonas reinhardtii cells were taken to centrifuge under freezing conditions at 6000 rpm/min for 10 min. Then the supernatant fluid of algal cells were rapidly obtained to mixed with 10mL 10mM EDTA-2Na, and the mixture were taken to centrifuge under freezing conditions at 6000rpm/min for 10 min again. The algal cells were eluted twice and the supernatant was removed to obtain algal cells. Ten millilitres of mixed acid was added for digestion on a 110℃ electric heating plate. The solution to be tested was obtained by constant volume of 1% nitric acid solution to 10 mL, and stored at 4℃ [40]. Cadmium and selenium content in the Chlamydomonas reinhardtii cells were determined by ICP-MS instrument (rf power was 1550 w, the cooler the argon gas flow velocity in 14 L/min, atomizer argon flow rate of 1 ml/min, auxiliary velocity of 0.8 L/min, the sampling depth is 5 mm, atomizing chamber temperature was 3 ℃, collision gas for high purity helium (purity>99.999%), the flow rate was 4.2mL/min, the injection speed was 40rpm, the resident time was 0.02s, and the scanning times were 20.) After the instrument was stabilized, the standard solution, digested blank solution and sample solution were measured in sequence, and the working curve was drawn.
2.10. Data Processing and Analysis
SPSS 20.0 was used for one-way analysis of variance (ANOVA) and Duncan's multiple comparison test was used for significance analysis (P<0.05), Origin Pro 2017 was used for plotting. Results are expressed as mean ± standard deviation.
3. Results
3.1. Preparation and Characterization of SeNPs
SeNPs has attracted the attention of many researchers because its surface activity is higher than other selenium compounds, so it can be better absorbed by intestinal tract. In our study, we wanted to establish a convenient method to prepare SeNPs with high biological activity and low toxicity. In this paper, SeNPs was prepared by chemical method which is that sodium carboxymethyl cellulose was used as dispersant, ascorbic acid was used as reducing agent and SeNPs was used as target. The morphology was observed by transmission electron microscopy, and then the particle size, potential and other indicators were determined to characterize it, and the stability was analyzed by dual-wavelength colorimetry.
SeNPs prepared with sodium carboxymethyl cellulose as dispersant and ascorbic acid as reducing agent is a bright orange red solution with no precipitation and can be stably preserved (Figure 2A). Under TEM ( transmission electron microscope), the SeNPs was well dispersed in sodium carboxymethyl cellulose, and there was no obvious agglomeration phenomenon, which was uniform spherical with a particle size of about 70nm (Figure 2B). The size of Chlamydomonas reinhardtii single cell is about 10 μm,so SeNPs can be totally absorbed by Chlamydomonas reinhardtii. EDS analysis showed that the elements with peaks were C, O, Se and Cu, in which C and O were the C and O elements in sodium carboxymethyl cellulose, and Cu were the copper elements in the copper mesh carried by transmission electron microscope, indicating that SeNPs was Se elemental(Figure 2C). According to the colloidal solution of double wavelength colorimetry [41], when the wavelength λ1 and λ2 were fixed, if the A2/A1 absorbance ratio remains unchanged, colloid particle size remains the same in a stable state[42]. The mixed solution with Vc had an obvious absorption peak at 260 nm (Figure 2D), while the sodium carboxymethyl cellulose solution had no characteristic absorption within the measured wavelength range. Therefore, in order to avoid the absorption of Vc and facilitate the determination, 410 nm and 490 nm[32] were selected as the determination wavelengths of SeNPs solution. The larger the ratio of A410/A490 is, the smaller the particle size of colloidal particles will be. When the ratio is unchanged, the particle size of colloidal particles will not change[31]. UV-vis absorption spectra showed that the absorbance of the solution at 410 nm and 490 nm gradually increased from the mixing of the reaction system, and the ratio of A410/A490 at the two wavelengths continued to fluctuate, and gradually stabilized after 2 h with a ratio of about 2 (Figure 2E). This indicates that the size of the SeNPs particles formed at this time does not change and is in a uniform and stable state. The particle size analysis of SeNPs using Malvern nanoscale analyzer shows that the spectral line of particle size distribution presents a single peak, and the peak shape is narrow, reaching the highest point at 186nm (Figure 2F). Therefore, 186nm is the most frequent particle size of nano-system system, indicating that the particle size of SeNPs is uniform and stable. Zeta Potential is one of the indicators to detect the stability of particle dispersion in the system, and it is a measure of mutual repulsion or attraction strength between particles. It not only determines the surface structure of ancient particles, but also depends on dispersant, so it can be used to predict the stability of dispersion solution. The smaller the molecule or dispersed particle is, the lower the absolute value (positive or negative) of the Zeta potential will reach, and the more likely it is to condense or condense. Table 1 shows the relationship between Zeta potential and stability. The measured zeta potential of SeNPs solution is -44.20±3.10, indicating that the SeNPs distribution system had a good stability according to Table 1.

Table 1.
The relationship between Zeta potential and stability.
| Zeta potential(mV) | Colloid stability |
|---|---|
| 0~±5 | Rapid coagulation or condensation |
| ±10~±30 | Start to get unstable |
| ±30~±40 | Poor stability |
| ±40~±60 | Good stability |
| >±61 | Excellent stability |
Table 2.
Particle size and Zeta potential of CMC-SeNPs.
| Particle size under TEM(nm) | 70 |
| Particle size in hydrology(nm) | 186.13±1.40 |
| Zeta potential(mV) | -44.20±3.10 |
3.2. Effects of SeNPs on the Growth of Chlamydomonas Reinhardtii under Cadmium Stress
Selenium can reduce the toxicity of heavy metals. SeNPs has attracted wide attention due to its higher biological activity and lower toxicity. One study showed that SeNPs can ameliorate the nonspecific immune function and antioxidant capacity decline of tilapia caused by cadmium stress to a certain extent[43]. However, there are few reports on the effect of SeNPs on heavy metals in algae. In this paper, different concentrations of SeNPs were applied to the wild-type Chlamydomonas rheinhardtii CC125 under cadmium stress and the cell wall deficient type Chlamydomonas rheinhardtii CC400, to explore the changes in biomass and chlorophyll content of Chlamydomonas rheinhardtii caused by cadmium toxicity in the presence of SeNPs, so as to observe that in the presence of SeNPs, direct effects of cadmium toxicity on the growth of Chlamydomonas reinhardtii.
Table 3 shows the growth curve of Chlamydomonas reinhardtii in 96 h under different Cd2+ concentrations. The area surrounded by each concentration and the coordinate axis are calculated according to the formula. The fitting equation takes the area as the abscissa and the Cd2+ concentration as the ordinate. According to the equation, the semi-maximum effect concentration EC50 of Cd2+ on the two species of Chlamydomonas reinhardtii was calculated (i.e. the Cd2+ concentration corresponding to the biomass was suppressed by half). The results showed that the EC50 concentration of Cd2+ was about 53.95±0.36 μmol/L against the wild type CC125 and 56.39±0.39 μmol/L against the cell wall defective type CC400. This experiment determined the Cd2+ concentration used in subsequent experiments, which were all the EC50 concentration of Cd2+. Table 4 shows the semi-maximum effect concentration EC50 of SeNPs on two species of Chlamydomonas reinhardtii. The results showed that the EC50 concentration of SeNPs was about 1.88±0.08 μmol/L for wild type CC125 and 0.53±0.03 μmol/L for cell wall defective type CC400. The concentration of SeNPs used in subsequent experiments was determined according to this experiment. It can also be seen from the comparison of the above two algae data that Cd2+ has a slightly lower EC50 concentration than CC400 for wild-type Chlamydomonas reinhardtii (no significant difference), while SeNPs has a higher EC50 concentration than CC400 for wild-type Chlamydomonas reinhardtii CC125. In the experiment, it was found that the range from the active concentration of Cd2+ and SeNPs to the lethal concentration was narrower in CC400 than in CC125. This may be related to the small size and defective cell wall of CC400.
In Figure 3A, (a) and (b) show the growth curves of CC125 and CC400 within 96 h, respectively. The solid lines are CK group (algae only) and SeNPs group (algae+SeNPs). The dashed lines are Cd2+ group (alga +Cd2+) and SeNPs+Cd2+ group (algae+SeNPs+Cd2+). In CC125 cells, the biomass of SeNPs group was not inhibited at the concentrations of 0.005, 0.01 and 0.2 μmol/L, which was not significantly different from that of CK group. The biomass of Cd2+ group was inhibited, which was about 50% of that of CK group. The biomass of SeNPs+Cd2+ groups with 0.005 μmol/L concentration increased slightly, while that of SeNPs+Cd2+ groups with 0.01, 0.2, 1 and 2 concentrations did not increase significantly, and the growth was very slow. After 48 hours of culture, cells entered the logarithmic growth phase, in which the biomass increased logarithmically on the basis of transitional growth and accumulation, and the distinction between groups became more and more obvious. The biomass of low concentration SeNPs group was slightly higher than that of CK group, and the biomass of 2μmol/L, which is close to the EC50 concentration of SeNPs, only was half suppressed, which was close to that of Cd2+ group. The growth curve of SeNPs+Cd2+ group was lower than that of Cd2+ group, and the growth inhibition was very significant. These results showed that low concentration of SeNPs had no significant inhibitory effect on the growth of wild type CC125, but high concentration of SeNPs had inhibitory effect on the growth of Chlamydomonas reinhardtii. Cd2+ itself can inhibit the growth of Chlamydomonas reinhardtii, and the addition of SeNPs can aggravate this inhibitory effect. The growth of CC400 cells was not significantly inhibited within 48 h of the transition period in every group, showing a concentration-dependent relationship, and the biomass growth was slightly different but not significant. After entering the logarithmic growth phase, the differentiation between groups became more and more obvious. The biomass of SeNPs group showed an extremely significant logarithmic growth, and the biomass was obviously concentration-dependent, while the biomass of Cd2+ group was inhibited, about 50% of that of CK group. SeNPs+Cd2+ group and Cd2+ group had no significant difference. The results showed that the growth inhibition of SeNPs+Cd2+ in CC400 cells with cell wall defect was not significantly different from that in Cd2+ group. From the overall trend, at SeNPs concentration without growth inhibition effect on Chlamydomonas reinhardtii, in the presence of Cd2+, the growth of CC125 was inhibited and the biomass of CC125 was significantly lower than that of Cd2+ group. The simultaneous presence of SeNPs and Cd2+ showed no significant difference in the growth inhibition of CC400 compared with that of Cd2+ alone. The possible reason for this phenomenon is that in CC125, the addition of SeNPs enhanced the toxicity of Cd2+, while in CC400, SeNPs did not have this effect.
Chlorophyll is the main pigment in plant photosynthesis. It is a kind of lipid pigment and plays a core role in light absorption in photosynthesis so chlorophyll content in algae cells can reflect the growth and physiological conditions of algae. Figure 3B to Figure 3G show the contents of chlorophyll a, chlorophyll b and carotenoid of CC125 and CC400 after 72 h growth in different treatment groups (CK group, SeNPs group, Cd2+ group and SeNPs+Cd2+ group).
In Figure 3B(a), the change of chlorophyll a content showed concentration-dependent relationship at different concentrations of SeNPs, among which 0.005, 0.01 and 0.2μmol/L groups were slightly higher than CK group, possibly due to the low toxicity excitatory effect. Low concentration of SeNPs promoted the synthesis of chlorophyll in algal cells, resulting in increased content. It may be because that algal cells open their own protective mechanism, which greatly increases energy consumption and respiration, prompting algal cells to synthesize more chlorophyll, thus ensuring the supply of substrate for respiration[44]. In Figure 3B (b), the chlorophyll a content in group Cd2+ is about 50% of that in group CK, while that in group SeNPs+Cd2+ is relatively low. The chlorophyll a of the lowest SeNPs concentration of 0.005μmol/L was significantly decreased compared with that of the Cd2+ group, and the growth of algal cells was severely inhibited at higher SeNPs concentration with extremely low chlorophyll contents. The contents of chlorophyll b and carotenoid in Fig. 3C and Fig. 3D were also concentration-dependent in CK group and SeNPs group, and the single action of SeNPs showed low toxicity excitation below 0.2μmol/L. The contents of chlorophyll b and carotenoid in SeNPs+Cd2+ group were significantly lower than those in Cd2+ group, which showed the same trend as that in biomass between SeNPs+Cd2+ group and Cd2+ group.
In Figure 3E(a), SeNPs with different concentrations showed concentration-dependent changes in chlorophyll a content. In Figure 3E(b), compared with the CK group, the chlorophyll a content in Cd2+ group was about 50% of that in CK group, while the chlorophyll a content in SeNPs+Cd2+ group was slightly reduced but had no significant difference from that in Cd2+ group. The contents of chlorophyll b and carotenoid in Fig 3F and Fig 3G were also concentration-dependent in CK group and SeNPs group. The contents of chlorophyll b and carotenoid in SeNPs+Cd2+ group were decreased compared with those in Cd2+ group, but with the increase of concentration, there was no obvious differentiation among several concentrations. The results showed the same trend as those of biomass. In summary, the 96 h growth curve of CC125 showed that 0.005 and 0.01 μmol/L SeNPs had no inhibitory effect on the growth of Chlamydomonas reinhardtii, but the inhibitory effect was obvious in the presence of Cd2+. The chlorophyll content of CC125 was determined and the same phenomenon was found. However, there was no such phenomenon in CC400 with cell wall defects, and there was no significant difference between the growth inhibition of Chlamydomonas reinhardtii in the presence of SeNPs and Cd2+ and the growth inhibition of CC400 by Cd2+ itself. The results of chlorophyll content showed the same trend.
3.3. Effects of SeNPs on Physiology of Chlamydomonas Rheinensis under Cadmium Stress
Reactive oxygen refers to some oxygen metabolites and their derivatives which contain oxygen atoms but have stronger oxidation capacity than oxygen. In normal growth cells, a very low level of Reactive Oxygen Species will not cause harm, and the production and removal of Reactive Oxygen Species in algal cells are generally in a dynamic equilibrium state. Trace Reactive Oxygen Species play an important role in the regulation of some physiological phenomena, but when the dynamic balance of Reactive Oxygen Species is broken, it may cause harm to the body [45]. The objective of this study was to investigate the enzyme activities of total superoxide dismutase (T-SOD) and catalase (CAT), the content of malondialdehyde (MDA) and the relative content of Reactive Oxygen Species (ROS) in the cells of two kinds Chlamydomonas reinhardtii under different treatments. The change of enzyme activities of total superoxide dismutase (T-SOD) and catalase (CAT), the content of malondialdehyde (MDA) and the relative content of Reactive Oxygen Species (ROS) were compared in two kinds of Chlamydomonas reinhardtii with and without cadmium.
According to the results of the experiment in the previous chapter, the concentration range of SeNPs in the experiment was further narrowed, and only two lower concentrations with no toxic effect on Chlamydomonas reinhardtii were selected: 0.005 and 0.01 μmol/L.
Figure 4A (a) shows the T-SOD activity of CC125. There is no significant difference between the CK and SeNPs groups, and the two concentrations in SeNPs+Cd2+ group are significantly increased compared with CK and Cd2+ groups. Compared with CK group, SeNPs+Cd2+ in 0.01 μmol/L group was nearly doubled. The reason may be that SeNPs+Cd2+ increased the content of superoxide anion radical in the cells, and promoted the production of a large amount of superoxide dismutase in the enzyme system in order to maintain the intracellular balance, indicating that the addition of SeNPs and Cd2+ at the same time had a greater toxic effect on CC125 algae cells. Figure 4A (b) shows the T-SOD activity of CC400. There is no significant difference in T-SOD activity between CK and SeNPs groups at 0.005 μmol/L concentration, but it is slightly higher in 0.01 μmmol/L SeNPs group. The activity of T-SOD in Cd2+ and SeNPs+Cd2+ groups was significantly increased compared with that in CK group, while there was no significant difference between the two concentrations in SeNPs+Cd2+ and Cd2+ groups. These results indicated that although Cd2+ was toxic, but the simultaneous addition of SeNPs and Cd2+ did not produce more toxic effects on CC400 algal cells.
Figure 4B (a) shows the CAT enzyme activity of CC125. There is no significant difference between the CK and SeNPs groups, and the two concentrations of SeNPs+Cd2+ group are significantly increased compared with CK and Cd2+ groups. The activity of CAT enzyme in 0.01 μmol/L SeNPs+Cd2+ group was about twice that in CK group. When SeNPs and Cd2+ were added at the same time, the external stress increased the hydrogen peroxide in CC125 cells and promoted the activity of CAT enzyme, thus reducing the formation of oxygen free radicals and reducing the toxic effect. These results indicated that the simultaneous addition of SeNPs and Cd2+ destroyed the homeostasis of the body cells, resulting in a certain degree of cell damage. Figure 4B (b) shows the CAT enzyme activity of CC400. There is no significant difference in CAT enzyme activity between CK group and 0.005μmol /L SeNPs group, while catalase is slightly increased in 0.01 μmol/L SeNPs+Cd2+ group, but there is no significant difference among the three groups. Therefore, it cannot be inferred that SeNPs increase Cd2+ toxicity in CC400.
Figure 4C (a) shows the MDA content of CC125. There is no significant difference in MDA content between CK group and SeNPs group. 0.005μmol/L and 0.01 μmol/L SeNPs alone do not produce toxicity. MDA content in SeNPs+Cd2+ group was significantly increased compared with CK and Cd2+ groups. The results indicated that the simultaneous addition of SeNPs and Cd2+ had a significant toxic effect on CC125 algal cells. The addition of SeNPs and Cd2+ at the same time produced significant toxic effects on CC125 algal cells resulting in the increase of oxygen free radicals. Lipid peroxidation occurs between oxidizing free radicals and polyunsaturated fatty acids in biofilms, resulting in the increase of lipid peroxides. Figure 4C (b) shows the MDA content of CC400, and there is no significant difference between the CK group and the three SeNPs groups. Compared with CK group, MDA contents in Cd2+ group and SeNPs+Cd2+ group were significantly increased, but there was no significant difference between the two concentrations in SeNPs+Cd2+ group and Cd2+ group.
Figure. 4D (a) shows the relative ROS content of CC125. There is no significant difference between the ROS relative content of CK group and that of SeNPs group, and the two concentrations of SeNPs+Cd2+ group have extremely significant increases compared with CK group and Cd2+ group. It may be due to the toxic effects of SeNPs+Cd2+ that lead to the destruction of the defense enzyme system in Chlamydomonas reinhardtii cells. Excessive oxygen free radicals cannot be removed in time in the cell, which breaks the ROS balance. Electron transfer in the photosynthetic system or respiratory system is blocked, and electrons are transferred to molecular oxygen O2 to form O2-, thus leading to ROS increase. Figure 4D (b) shows the ROS relative content of CC400. The ROS relative content of Cd2+ and SeNPs+Cd2+groups is significantly increased compared with that of CK group, while the ROS relative content of 0.005μmol /L between CK and SeNPs groups has no significant difference.
3.4. Effects of SeNPs on Photosynthetic System and Intracellular Cadmium Content of Chlamydomonas Reinhardtii under Cadmium Stress
Photosynthesis is a unique physiological function of plants. It is the process of converting solar energy into chemical energy on the largest scale on Earth, synthesizing inorganic substances such as carbon dioxide and water into organic matter while releasing oxygen. Chloroplasts are the organelles that carry out the core reactions of photosynthesis. The basic processes of energy conversion and carbon assimilation in photosynthesis are carried out in chlorophylls. The thylakoid membrane is a lipid bilayer membrane required for energy conversion in photosynthesis. It contains the pigment protein and enzyme system necessary for capturing light and converting light energy into chemical energy in photosynthesis. Many different complexes are involved in the photosynthesis process, most of which are embedded in the thylakoid membrane, and four membrane-bound supramolecular complexes are directly involved in photosynthesis: photosystem I (PSI), photosystem II (PS II), ATP synthase complex, and cytochromatin b6f complex (Cyt b6f) [46]. The structure of PSI and PSII are as Figure 5 shows.
The PSI complex consists of photoreaction center pigment (P700), light trapping complex (LHC I), and electron acceptor, and its function is to transfer electrons from Plastocyanin (PC) to Ferredoxin (Fd). When the pigment molecules in the reaction center of PSI absorb light energy and are excited, they transfer electrons to various electron acceptors, and by Fd, NADP+ is reduced to NADPH with the participation of NADP reductase. NADPH participates in the dark reaction stage of photosynthesis and carries out solid carbonization reaction. Specific transmission route: LHCI transfers the absorbed light energy to P700→ primary electron acceptor A0 (Chla)→ primary electron acceptor A1 (possibly folloquinone VK1)→ iron-sulfur center (Fx→FA→FB)→ ferredodoredoin Fd.
PSII contains the PSII reaction center (PSII), light-harvesting complex II (LHCII), and oxygen-evolving complex (OEC). Its function is to use the energy absorbed from light to split water and transfer its released electrons to plastiquinone, while establishing an H+ proton gradient on both sides of the thylakoid membrane through oxidation of water and reduction of PQB2-. PSII transfers the light energy absorbed by LCHII to the PSII reaction center complex, causing it to produce a high-energy electron, which is passed to the primary electron acceptor. This process creates a positively charged donor (P680+) and a negatively charged primary electron acceptor (Pheo-). P680+ can be used as an oxidizing agent to trigger the photolysis of water, resulting in the transfer of electrons released by water oxidation to PSII. Pheo- can be used as a reducing agent to lose an electron, causing electron transfer to QA and QB[47].
The light reaction takes place on the chloroplast thylakoid membrane. The electrons produced by cracking water molecules in the oxygen-releasing complex pass through PSII, Cyt b6f, and PSI, and are finally transferred to nicotinamide adenine dinucleoside phosphate (NADP) to form NADPH. The transmembrane proton gradient generated during this process drives ATP synthase to form ATP. Such electron transport flow is called linear electron flow, and the ATP formation process coupled with it is called non-cyclic photophosphorylation (NCPSP). If the electron is not transferred to NADP after PPI, but is returned to PSI via Cyt b6f or plastoquinone (PQ), only a transmembrane proton gradient is formed during the transfer process, and ATP is synthesized. Such electron transport around PSI is called cyclic electron flow (CEF), and the ATP synthesis process coupled with CEF is called cyclic photophosphorylation (CPSP)[48].
So, as all above, photosynthesis involves multiple electron transfers and energy conversion, so it is crucial for cell growth in autotrophs, especially algae cells.
According to the change of chlorophyll fluorescence intensity caused by the effect of SeNPs on the toxicity of cadmium-induced Chlamydomonas reinhardtii, rapid fluorescence kinetic analysis was carried out to obtain the OJIP kinetic curve and QA-reoxidation kinetic curve. OJIP-test was analyzed and relevant photosynthetic parameters were deduced, so as to determine the changes in the donor side, recipient side and reaction center of PSII under the conditions of SeNPs and cadmium treatment[49], and thus the influence on the photosynthetic system of Chlamydomonas reinhardtii was known. The toxicity mechanism of heavy metals to algae is mainly the destruction of cell membrane and the damage of antioxidant enzyme system of algae, and algae have a certain biological accumulation of heavy metals[50]. The possible mechanism of SeNPs enhancing Cd2+ toxicity could be further determined by the determination of cadmium and selenium contents in algal cells.
When plants or parts of tissues with photosynthetic organs transfer from dark conditions to visible light, the green tissues of plants emit a dark red fluorescence with ever-changing intensity, which increases with the change of time, rises to a maximum value, then declines, and finally reaches a stable value. This phenomenon is known as chlorophyll fluorescence induction phenomenon, also known as Kautsky effect[51]. The curve of fluorescence over time is called chlorophyll fluorescence induced kinetic curve. Generally, the lowest fluorescence measured just after exposure to light is O point, and the maximum fluorescence is defined as P point. The fast chlorophyll fluorescence induction kinetics curve refers to the fluorescence change process of plant leaves or cells from O point to P point under different measured light irradiation[52]. It mainly reflects the changes of the primary photochemical reaction of PSII, the structure and state of photosynthetic apparatus, etc. The descending stage mainly reflects the changes of photosynthetic carbon metabolism, and the fluorescence intensity gradually decreases with the increase of photosynthetic carbon metabolism rate[53]. Typical fast chlorophyll fluorescence induction kinetics curves generally include phases such as O-J-I-P, which records the whole process from initial fluorescence (F0) to maximum fluorescence (Fp or Fm)[54].
After being treated with SeNPs and Cd2+ at different concentrations for 72 h, the OJIP test was carried out (Fig.6A). As can be seen from the curve in Figure 6A (a), the curve of CK group and SeNPs group in CC125 increased significantly, and FJ, FI and FM were higher than those in Cd2+ and SeNPs+Cd2+ groups, indicating that Cd2+ and SeNPs+Cd2+ groups had an inhibitory effect on PSII activity of Chlamydomonas reinhardtii. Among them, there was no significant difference between the two concentrations of SeNPs group and CK group, while the addition of Cd2+ reduced the fluorescence intensity successively, and there was no significant difference between CK group and SeNPs groups, indicating that under the SeNPs concentration that had no effect on Chlamydomonas reinhardtii, after the addition of Cd2+, the inhibitory effect of cadmium on PSII activity of Chlamydomonas reinhardtii was enhanced in different degrees, and the trend was concentration dependent. As can be seen from the curve in Figure 6A (b), the curve of CK group and SeNPs group increased significantly in CC400, among which the CK group was the highest and the two groups 0.005 and 0.01 μmol/L decreased successively, indicating that the PSII activity of SeNPs groups were slightly inhibited compared with CK group. Cd2+ group and SeNPs+Cd2+ groups were significantly lower than CK group and SeNPs groups, but the difference between groups was not significant. The results indicated that Cd2+ and SeNPs+Cd2+ groups inhibited the PSII activity of Chlamydomonas reinhardtii, but the addition of SeNPs could not enhance the inhibitory effect of Cd2+.
A large amount of original data can be obtained from the chlorophyll fluorescence induction curve, including a large amount of information about the primary photochemical reaction of the PSII reaction center. By analyzing the data according to parameters related to the chlorophyll fluorescence induction curve, it can be known that the structure and function of the plant photosynthetic system are affected under external stress conditions[55,56]. PIABS is a performance index of light energy absorption, which can comprehensively reflect the electron transport efficiency of the acceptor side, the number of active reaction centers and the maximum photochemical efficiency of PSII, and can more accurately reflect the influence of stress on the state and functional activity of PSII [57,58]. As shown in Fig.6B (a), PIABS values in Cd2+ group and SeNPs+Cd2+ group of CC125 were significantly lower than those in CK and SeNPs groups, indicating that the presence of Cd2+ had a strong toxic effect on PSII, and the addition of SeNPs at the same time would promote this effect. In Fig.6B (b), the CK group and SeNPs group of CC400 were different from the Cd2+ group and SeNPs+Cd2+ group, and there was no significant difference within the groups, indicating that the concurrent addition of SeNPs would not promote the toxic effect of Cd2+ on PSII. Mo reflects the maximum rate at which QA is reduced and reflects the changes on the receptor side of PSII reaction center [59,60]. When the downward electron transfer rate of QA- decreases, the reduction rate of QA is accelerated, so Mo correspondingly increases, that is, the increase of Mo indicates that the downward electron transfer of QA- is inhibited [61]. In Fig. 6B (a), Mo values of CC125 in Cd2+ group and SeNPs+Cd2+group were slightly higher than those in CK and SeNPs groups, indicating that electron transport in Cd2+ group and SeNPs+Cd2+ group was inhibited to a certain extent, but the difference was not significant. In Fig 6B (b), CC400 is similar to CC125. VJ reflects the openness of the reaction center, that is, the amount of QA- accumulation. The increase of VJ indicates that the amount of QA- accumulation in the photosynthetic electron transport chain increases, while the relative electron transport capacity of PSII decreases. In Figure 6B (a), the VJ values of CC125 in Cd2+ group and SeNPs+Cd2+ group were slightly higher than that in CK and SeNPs groups, indicating that the relative electron transport capacity of PSII in Cd2+ group and SeNPs+Cd2+ group were lower, and the difference was not significant. In Figure 6B (b), there is no significant difference between the groups of CC400, indicating that the QA-to-QB electron transfer of CC400 is not affected by Cd2+ and SeNPs. ABS/RC, TRo/RC, ETo/RC and DIo/RC respectively reflect the light energy absorbed by unit reaction center, the energy captured by unit reaction center for reducing QA, the energy captured by unit reaction center for electron transfer and the energy dissipated by unit reaction center. The difference in these values was not significant between the two algal cells.
QA-reoxidation kinetics was used to further investigate the effect of SeNPs and Cd2+ treatments at different concentrations on the PSII accepter side electron transport efficiency of Chlamydomonas reinhardtii. In Fig 6C (a), there was no significant difference between CK group and SeNPs group in CC125, while the Cd2+ group and SeNPs+Cd2+ group decreased successively. These results indicated that Cd2+ group and SeNPs+Cd2+ group improved the sensitivity of electron acceptor side of PSII and inhibited the reoxidation process of QA-. In Fig. 6C (b), there was no significant difference between CK group and SeNPs group in CC400 at 0.005 μmol/L, but the 0.01 μmol/L in SeNPs group was slightly decreased, indicating that the reoxidation process of QA- was also inhibited at 0.01 μmol/L. There was no significant difference between the Cd2+ group and SeNPs+Cd2+ group at 0.005 μmol/L, but the SeNPs+Cd2+ group at 0.01 μmol/L was lower.
Figure 6D shows the effect of SeNPs on the intracellular Cd content of Chlamydomonas reinhardtii. Fig.6D(a) shows the intracellular cadmium content of CC125 under conditions of Cd2+, 0.005 μmol/LSeNPs+Cd2+, and 0.01 μmol/LSeNPs+Cd2+. It can be seen that, compared with the Cd2+ group, the addition of SeNPs increased the intracellular Cd2+ content, and the difference was significant, and showed a concentration dependent trend with the increase of SeNPs concentration. Figure 6D(b) shows the determination result of cadmium content in CC400 algae cells. Different from the results of CC125, the intracellular Cd2+ content of CC400 algae was not significantly increased due to the presence of SeNPs, and there was no significant difference in Cd2+ content among the three groups.
The SeNPs concentrations in Chlamydomonas reinhardtii culture are about 0.395µg/L and 0.79µg/L. Because the concentration is too low and is too close to the detection limit of the instrument, the error of the measurement results is large, and the trend is covered up. Therefore, the results of the determination of the intracellular selenium content by ICP-MS cannot be trusted.
4. Discussion
In this paper, the preparation of SeNPs and the effect of SeNPs on cadmium toxicity in the growth, physiological status and photosynthetic system of Chlamydomonas reinhardtii were preliminarily investigated. It was found that the biomass, photosynthetic pigment content, physiological indexes and photosynthetic parameters of Chlamydomonas reinhardtii cells were changed in different degrees when cultured under different concentrations of SeNPs, Cd2+ and SeNPs+Cd2+.
Elemental selenium is very unstable when left alone and is difficult to exist at the nanoscale. This requires the selenium to be dispersed in a stabilizer to control its particle size in the nanometer range. Using sodium carboxymethyl cellulose as dispersing agent, ascorbic acid as reducing agent and selenite as selenium source, SeNPs with high biological activity and low toxicity can be conveniently prepared. Sodium carboxymethyl cellulose is like soft template so that SeNPs is evenly distributed in it, the particle size is uniform and stable, suitable for application as experimental materials.
In previous studies, researches have proved that metallic nanoparticles can be used as stressors or regulators of microalgae growth and metabolism. Metals at small concentrations are indispensable for microalgae cells to perform cellular functions[62]. Furthermore, some studies have shown that metal nanoparticles can mitigate the toxic effects of cadmium. The adsorption of nano-titanium dioxide can reduce the concentration of Cd2+ in water, thereby reducing the toxicity of Cd2+ to algae. The presence of nano-titanium dioxide can slow down the cytotoxicity of Cd2+ to chlamydomonas. It may be that the adsorption reduces the concentration of Cd2+ in water, thus reducing the bioaccumulation and toxicity of Cd2+ to algal cells. It may also be that the electrostatic repulsion and potential steric hindrance between the coated nano-titanium dioxide and algal cells prevent direct contact with each other. In addition, it may be the protective effect of the cell wall, which needs to be further studied[20]. Carbon nanotubes were found to inhibit cadmium enrichment in duckweed, but the inhibition decreased with the increase of cadmium concentration[21]. Nanometer manganese dioxide can reduce the accumulation of heavy metals in rice seedlings[22]. Nano zinc oxide can inhibit the toxicity of CdCl2 in mice, and the combined exposure of nano zinc oxide and CdCl2 increases the contents of GSH and MDA and the activities of SOD and GSH-Px in liver and kidney of mice, which can inhibit oxidative damage[23]. The silver nanoparticles can promote the removal of Cd2+ from the solution of P. chrysospora, and the promotion effect increases with the increase of the concentration of silver nanoparticles in a certain concentration range[24]. The accumulation of cadmium in cucumber and wheat, the inhibitory effect of cadmium on seed germination and the oxidative damage in seedlings were significantly reduced by 6 nm ferric oxide nanoparticles[25].
Similarly, previous studies have found that selenium can reduce the toxicity of cadmium in plants and animals, but our study found that the presence of SeNPs did not reduce the toxicity of cadmium in algae, but to some extent increased the toxicity of cadmium. The reason why this phenomenon occurs is worth exploring. Other researches also found the toxicity of metal nanoparticles. For example, as the positive effects mentioned above, there are negative effects exists simultaneously in the mentioned studies earlier. Although nano-manganese dioxide can reduce the accumulation of heavy metals in rice seedlings, nano-manganese dioxide itself can cause strong oxidative stress reaction. Earthworms were taken as experimental materials, nano-manganese dioxide can also significantly reduce the accumulation of cadmium in earthworms, but nano-manganese dioxide can stimulate the stress response of the antioxidant enzyme system in earthworms, raising the cadmium toxicity from toxic level to high toxic level[22]. In addition, the application concentration of metal nanoparticles needs to be very careful. Although nano silver particles can promote the removal of Cd2+ from the solution, the promotion effect will gradually disappear when the concentration of nano silver exceeds 1mg/L, and the inhibition effect on P. flavescens appears[24].
In our study, red elemental selenium dispersed by sodium carboxymethyl cellulose did not show good biological effects in Chlamydomonas reinhardtii cells, but showed significantly enhanced toxicity to Cd2+ in CC125 cells, while CC400 mutant strain with cell wall defects did not show this property. That is,SeNPs may enhance Cd2+ toxicity in CC125, but not significantly in CC400. In the presence of SeNPs, the enhanced Cd2+ toxicity of CC125 algae was related to the amount of Cd2+ entering the cells. In the presence of SeNPs, the Cd2+ content in CC125 cells increased significantly, which showed a concentration-dependent trend with the increase of SeNPs concentration. There was no significant difference in intracellular cadmium content among CC400 groups. The reason for the difference may be due to the difference in cell wall and individual size between the two kinds of algae cells. If it is boldly assumed that SeNPs can enter CC125 cells, the increase of intracellular Cd2+ content is related to the amount of selenium entering the cells. The higher the concentration of selenium, the higher the amount of Cd2+ entering the cells, the stronger the toxicity to CC125 algae. On the contrary, there was no significant difference in Cd2+ content in CC400 algal cells, which may be due to the small size of CC400 cells, SeNPs could not enter CC400 algal cells. If the SeNPs cannot enter CC125 algal cells, the interaction between the SeNPs and the cell wall surface may result in different Cd2+ content entering the cells due to the difference in cell wall between the two algal cells.
Combined exposure of SeNPs and Cd2+ not only inhibited the growth of CC125, but also showed toxic effects on physiological indexes and the most important photosynthetic system of Chlamydomonas reinhardtii. Combined exposure promoted various oxidative stress reactions in cells, resulting in a series of physiological changes. The photosynthetic system was affected by the reduction of the number of reactive centers of the photosynthetic system and the reduction of photosynthetic efficiency caused by the reduction of electron transport efficiency. The same phenomenon was not seen in CC400 cells.
As for CC125, the enhanced toxicity of Cd2+ caused by SeNPs is is thought to be caused by these factors below: (i) Causing cellular oxidative stress (ii) Damaging the photosynthetic system (iii) Changing ultrastructure.
Metal nanoparticles have been reported to have inhibitory effects on many freshwater and Marine microalgae strains, and their inhibitory activity is thought to be due to the production of reactive oxygen species (ROS)[63,64] or mechanical damage caused by the nanoparticles themselves. The activity of superoxide dismutase (SOD) increased with the increase of ROS species in cells[65]. Lipid peroxidation and plasmolysis may result if antioxidant mechanisms are insufficient to minimize oxidative stress[66].
The main mechanism of SeNPs' influence on the photosynthetic system may be that cells under the stress of nanoparticles produce too much ROS to damage the PSII light-collecting complex and reaction center or NPQ's compensatory protection against toxic effects is weakened, preventing electron transfer and causing pigment degradation[26]. In addition, the positively charged nanoparticle interacts with the negatively charged component of the cell wall and aggregates around the cell. It reduces light availability causing a shading effect and decreases photosynthesis.
The interaction between nanoparticles can significantly damage the ultrastructure of cells. Studies have shown that NPs may damage the cell surface and cause the cell to contract. After AgNPs treatment for 48 h, the morphology and structure of microcystis were changed, and the cells were deformed and shrunken[67,68,69]. The interaction of TiO2 NPs with the cell forms heteroaggregation, resulting in mechanical destruction of the cell membrane and a reduction in the amount of available light, as well as the accumulation of NPs in the cells[65,70].
The difference between CC125 and CC400 is the existence or vanish of cell wall. One of the reason why CC400 escape for the enhanced toxicity is may because it does not have a negatively charged cell wall and cannot interact with the positively charged SeNPs to gather around the cell, so there’s nothing to reduce the cell's photosynthesis. The cell wall of Chlamydomonas Rhine is rich in glycoproteins containing hydroxyproline. Hydroxyproline has more tendency to chelate metal so the other reason that CC400 are less susceptible to the combined exposure of SeNPs and Cd2+ is that it doesn’t have cell wall rich in hydroxyproline so it has less possibilities to chelate the mixture.
This provides some guidance for future research on the effect of SeNPs on alleviating the toxicity of heavy metals in algae. In addition, we found that the presence of SeNPs can enhance the toxicity of cadmium in CC125 of Chlamydomonas reinhardtii, but the specific mechanism of action remains to be studied. The comparison between normal CC125 cells and cell wall defective CC400 cells on the effects of SeNPs on cadmium toxicity can provide a possibility for the mechanism study. The addition of SeNPs in CC125 increased the toxicity of Cd2+, and the mechanism of toxic action was related to the amount of Cd2+ entering cells, while the amount of SeNPs was extremely small. It was difficult to determine whether the increase of Cd2+ entering cells was due to SeNPs entering cells or SeNPs acting on the cell surface. Further research can be conducted on the amount and morphological changes of SeNPs entering algal cells based on the detection method of trace substances.
Author Contributions
Sun Yuanyuan - Conceptualization; Visualization; Results discussion; Writing and editing manuscript. Gao Min - Methodology; Experiment design; Data analysis; Metabolite quantification; Writing the first draft of manuscript. Zhu Yi - Conceptualization; Funding acquisition; Supervision; Manuscript editing.
Funding
Supported by the National Natural Science Foundation of China (31101263).
Data Availability Statement
All data generated or analysed during this study are included in this published article. If you need more information, please contact corresponding author by zhuyi@cau.edu.cn.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that may have an influence on the work reported in this paper.
References
- Sabatini SE, Juárez ÁB, Eppis MR, Bianchi L, Luquet CM, Molina MdCRd. Oxidative stress and antioxidant defenses in two green microalgae exposed to copper. Ecotoxicology and Environmental Safety. 2009;72. [CrossRef]
- Ran XF, Liu R, Fang B, Shi JQ, Wu ZX. Response of microcystis growth and photosynthetic system ⅱ to Cadmium. Journal of Hydrobiology. 2015.
- K B, B H, J H, Z H, R T. Preparation and antioxidant properties of Selenium Nanoparticles-loaded chitosan microspheres. International Journal of Nanomedicine. 2017;Volume 12.
- Chang L, Yuanting F, Chang-E L, Tianfeng C, Xiaoling L. Phycocyanin-Functionalized Selenium Nanoparticles Reverse Palmitic Acid-Induced Pancreatic β Cell Apoptosis by Enhancing Cellular Uptake and Blocking Reactive Oxygen Species (ROS)-Mediated Mitochondria Dysfunction. Journal of agricultural and food chemistry. 2017;65.
- Xiao Y, Huang Q, Zheng Z, Guan H, Liu S. Construction of a Cordyceps sinensis exopolysaccharide-conjugated Selenium Nanoparticles and enhancement of their antioxidant activities. International Journal of Biological Macromolecules. 2017;99. [CrossRef]
- Li HY, Tang S, Wang YQ, et al. Effects of selenium on Cadmium content and subcellular distribution in rice.Journal of Ecological Environment. 2016.
- Qu CJ, Wu ZC, Zhao XH, et al. Cadmium accumulation, photosynthetic characteristics and antioxidant activity of different genotypes of Rapeseed under cadmium stress by selenium influence. Journal of Agricultural Resources and Environment. 2015.
- Gao XY, Zhang JS, Zhang LD, et al. Acute toxicity and bioavailability of nano-red elemental selenium. Health research. 2000.
- Ren CM. Study on the protective effect of Selenium on cadmium-induced reproductive toxicity in male mice and its mechanism:[Doctoral dissertation] Nanjing Medical University. 2012.
- Wang Y. Protective effect of selenium on kidney injury induced by cadmium in mice and its mechanism:[Masteral dissertation] Nanjing Medical University. 2013.
- Deng XY, Liu YM, Zhao XY,et al. Clinical research progress of nano-selenium. International Journal of Laboratory Medicine. 2013.
- Fang Y, Yang CY, Chen MQ, et al. Nanotechnology and nanomaterials-Introduction to nanotechnology and nanomaterials. Domestic chemical industry. 2003.
- Yanzhou C, Lizhen H, Zhibin L, Lilan Z, Zhenhuan S, Penghui L, et al. Designing Core-Shell Gold and Selenium Nanocomposites for Cancer Radiochemotherapy. ACS nano. 2017;11.
- Feng C, Hong ZX, Dan HX, Dang LP, Qian ZH. The effects of combined Selenium Nanoparticles and radiation therapy on breast cancer cells in vitro. Artificial cells, nanomedicine, and biotechnology. 2018;46.
- Rao L, Rao M, Qiu HH, et al. Effect and mechanism of nano-selenium on apoptosis of pancreatic β cells. General Journal of Biotechnology. 2016.
- A E-GM, N F, E R, A E-B, A KS. Anti-inflammatory effect of Selenium Nanoparticles on the inflammation induced in irradiated rats. Canadian journal of physiology and pharmacology. 2017;95.
- Nguyen THD, Vardhanabhuti B, Lin M, Mustapha A. Antibacterial properties of Selenium Nanoparticles and their toxicity to Caco-2 cells. Food Control. 2017;77. [CrossRef]
- Zhengfang L, Yinghua L, Min G, Misi X, Changbing W, Mingqi Z, et al. Inhibition of H1N1 influenza virus by Selenium Nanoparticles loaded with zanamivir through p38 and JNK signaling pathways. RSC Advances. 2017;7.
- Mustafa N, Salina M, Laszlo P. Nanoparticles as potential clinical therapeutic agents in Alzheimer's disease: focus on Selenium Nanoparticles. Expert review of clinical pharmacology. 2017;10.
- Wang JJ, Wang XN, Dai H, et al. Research progress on joint toxicity of titanium dioxide nanoparticles and heavy metals. Journal of Ecotoxicology. 2015.
- You HP. Effect of nanoparticles on heavy metals enrichment in water by red Duckweed:[Masteral dissertation] Fujian Normal University. 2014.
- Huang YF. Characteristics and safety evaluation of adsorption of heavy metal copper and cadmium ions by nano-sized manganese dioxide:[Masteral dissertation] Chinese Academy of Agricultural Sciences. 2016.
- Gao S. Study on synergistic toxicity of nanometer zinc oxide and cadmium chloride:[Masteral dissertation] University of Science and Technology of China. 2014.
- Zuo YN. Study on the effect of Silver nanoparticles on the removal of Cadmium by Anoderma chrysospora and its migration and transformation in the system:[Masteral dissertation] Hunan University. 2015.
- Alexandre K. Physiological effects of magnetic ferric oxide nanoparticles on cucumber and wheat and their mitigation effects on heavy metal toxicity:[Doctoral dissertation] China Agricultural University. 2017.
- Mukesh K, Sneha S, Vishambhar S, Mukesh M, D RV, Tatiana M, et al. The mechanism of nanoparticle toxicity to cyanobacteria. Archives of microbiology. 2022;205.
- Hazeem LJ, Kuku G, Dewailly E, Slomianny C, Barras A, Hamdi A, et al. Toxicity Effect of Silver Nanoparticles on Photosynthetic Pigment Content, Growth, ROS Production and Ultrastructural Changes of Microalgae Chlorella vulgaris. Nanomaterials. 2019;9.
- Abdallah O, Stephanie P, François P, Radovan P. Temperature influence on silver nanoparticles inhibitory effect on photosystem II photochemistry in two green algae, Chlorella vulgaris and Dunaliella tertiolecta. Environmental science and pollution research international. 2012;19.
- Wang HY, Zhang SY, Liu MZ, et al. Preparation of nano-selenium by chitosan template method. Applied chemistry. 2004.
- EH. H. Chlamydomonas as a model organism [Review]. Annual Review of Plant Physiology and Plant Molecular Biology. 2001.
- Wang HY, Zhang SY. Hao HW, et al. Preparation of nano-selenium by sodium carboxymethyl cellulose template method. Functional material. 2004.
- Hua PM. Preparation and application of nano-selenium:[Masteral dissertation] Hebei University. 2009.
- Yang JZ. Study on the application of Sodium carboxymethyl cellulose in food industry. Agricultural product processing. 2014.
- Zheng H. Effect of Chlamydomonas rheinaceans on the distribution and removal efficiency of copper oxide nanoparticles in water environment:[Masteral dissertation] Shandong Agricultural University. 2016.
- Liu G, Chai X, Shao Y, Hu L, Xie Q, Wu H. Toxicity of copper, lead, and cadmium on the motility of two marine microalgae Isochrysis galbana and Tetraselmis chui. Journal of Environmental Sciences. 2011;23. [CrossRef]
- Jiang YB, Ji HB. Bioremediation of heavy metal polluted water by algae. Progress in geography. 2007.
- EH. H. Chlamydomonas as a model organism [Review]. Annual Review of Plant Physiology and Plant Molecular Biology. 2001;52.
- Sager R, Granick S. Nutritional Studies with Chlamydomanas-Reinhardi. Annals of the New York Academy of Sciences. 1953;56.
- Li L, Chen X, Zhang D, Pan X. Effects of insecticide acetamiprid on Photosystem II (PSII) activity of Synechocystis sp. (FACHB-898). Pesticide Biochemistry and Physiology. 2010;98. [CrossRef]
- Wang YN, Shi YP, Li JD. Determination of lead and cadmium in Spirulina by ICP-MS. Analytical laboratory. 2004.
- Shibata S, Furukawa M, Nakashima R. Dual-Wavelength Spectrophotometry .6. Determination of Phenol in Industrial-Waste and Derermination of 2,4-Dichlorophenol and 2,4,6-Trichlorophenol in Mixtures by 1St Derivative Spectra. Analytica Chimica Acta. 1976;81.
- Hong HW. Determination of Ca2+ in natural water by dual wavelength spectrophotometry. Shanghai Environmental Science. 1992.
- Qin FJ, Jin J, Gu HJ, et al. Effects of nano-selenium on the nonspecific immune and antioxidant functions of Gifu Tilapia under cadmium stress. Journal of Agro-Environmental Sciences. 2011.
- Yuan HY, Liu JQ, Lu XP. Effects of lead stress on chlorophyll content and antioxidant enzyme activity of Cauliflower. Anhui agricultural Sciences. 2010;23.
- Liu XL, Teng L, Li SF. Research progress in the mechanism of producing Reactive Oxygen Species in algae. Journal of Huangshi Institute of Technology. 2007.
- Du LF. Some advances in photosynthesis research. World scientific and technological research and development. 1999:01.
- Yu Y, Weng J, Xu CH. Research progress on the structure and function of plant photosystem II oxygen-releasing complex periproteins in vitro. Journal of Plant physiology. 2001;06.
- Xue X, Xu HM, Wu HY. Research progress of cyclic electron transport in plant photosynthesis. Journal of Plant physiology. 2017;02.
- Zhang RJ, Ren F, Bai YB, et al. Research progress on the effects of stress on Psii based on rapid chlorophyll fluorescence induction kinetics. Anhui agricultural Sciences. 2012.
- Liu L. Interaction of heavy metal lead and cadmium with two species of freshwater algae:[Masteral dissertation] Anhui Normal University. 2014.
- Kautsky H. Influence of Eutrophication On the Distribution of Phytobenthic Plant and Animal Commuities. Internationale Revue Der Gesamten Hydrobiologie. 1991;76.
- Krause G H, Weis E. Chlorophyll Fluorescence and Photosynthesis-the Basics. Annual Review of Plant Physiology and Plant Molecular Biology. 1991;42.
- Li PM, Gao HY, Reto J. Strasser. Application of kinetic analysis of rapid chlorophyll fluorescence induction in the study of photosynthesis. Journal of Plant Physiology and Molecular Biology. 2005.
- Stirbet A, Govindjee. Chlorophyll a Fluorescence Induction: A Personal Perspective of the Thermal Phase, the J-I-P Rise. Photosynthesis Research,. 2012;113. [CrossRef]
- R. VHPD, J. SR, J. KGH. Reduction of dark chilling stress in N-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics. Physiologia plantarum. 2004;121.
- Hermans C, Smeyers M, Rodriguez RM, Eyletters M, Strasser RJ, Delhaye J-P. Quality assessment of urban trees: A comparative study of physiological characterisation, airborne imaging and on site fluorescence monitoring by the OJIP-test. Journal of Plant Physiology. 2003;160. [CrossRef]
- Gert S, Alaka S, J SR. Characterization of the 820-nm transmission signal paralleling the chlorophyll a fluorescence rise (OJIP) in pea leaves. Functional plant biology : FPB. 2003;30.
- Heerden PDRV, Tsimilli-Michael M, Krüger GHJ, Strasser RJ. Dark chilling effects on soybean genotypes during vegetative development: parallel studies of CO2 assimilation, chlorophyll a fluorescence kinetics O-J-I-P and nitrogen fixation. Physiologia Plantarum. 2003;117.
- Bueno M, Fillat M F, Strasser R J, et al. Effects of Lindane On the Photosynthetic Apparatus of the Cyanobacterium Anabaena - Fluorescence Induction Studies and Immunolocalization of Ferredoxin-NADP(+) Reductase. Environmental Science and Pollution Research. 2004;11.
- DemmigAdams B, Adams W W. Xanthophyll Cycle and Light Stress in Nature: Uniform Response to Excess Direct Sunlight Among Higher Plant Species. Planta. 1996;198. [CrossRef]
- Appenroth K J, Stockel J, Srivastava A, et al. Multiple Effects of Chromate On the Photosynthetic Apparatus of Spirodela Polyrhiza as Probed by OJIP Chlorophyll a Fluorescence Measurements. Environmental Pollution. 2001;115. [CrossRef]
- Miazek K, Iwanek W, Remacle C, Richel A, Goffin D. Effect of Metals, Metalloids and Metallic Nanoparticles on Microalgae Growth and Industrial Product Biosynthesis: A Review. International Journal of Molecular Sciences. 2015;16. [CrossRef]
- Xia B, Chen B, Sun X, Qu K, Ma F, Du M. Interaction of TiO 2 nanoparticles with the marine microalga Nitzschia closterium : Growth inhibition, oxidative stress and internalization. Science of the Total Environment. 2015;508. [CrossRef]
- Suman TY, Rajasree SRR, Kirubagaran R. Evaluation of zinc oxide nanoparticles toxicity on marine algae chlorella vulgaris through flow cytometric, cytotoxicity and oxidative stress analysis. Ecotoxicology and Environmental Safety. 2015;113. [CrossRef]
- Aruoja V, Dubourguier H-C, Kasemets K, Kahru A. Toxicity of nanoparticles of CuO, ZnO and TiO 2 to microalgae Pseudokirchneriella subcapitata. Science of the Total Environment. 2008;407.
- Hall, Bradley, Moore, Kuykindall, Minella. Acute and chronic toxicity of nano-scale TiO2 particles to freshwater fish, cladocerans, and green algae, and effects of organic and inorganic substrate on TiO2 toxicity. Nanotoxicology. 2009;3.
- Chen L, Zhou L, Liu Y, Deng S, Wu H, Wang G. Toxicological effects of nanometer titanium dioxide (nano-TiO 2 ) on Chlamydomonas reinhardtii. Ecotoxicology and Environmental Safety. 2012;84.
- Yanık F, Vardar F. Toxic Effects of Aluminum Oxide (Al2O3) Nanoparticles on Root Growth and Development in Triticum aestivum. Water, Air, & Soil Pollution. 2015;226. [CrossRef]
- Duong TT, Le TS, Tran TTH, Nguyen TK, Ho CT, Dao TH, et al. Inhibition effect of engineered silver nanoparticles to bloom forming cyanobacteria. Advances in Natural Sciences: Nanoscience and Nanotechnology. 2016;7.
- Ji J, Long Z, Lin D. Toxicity of oxide nanoparticles to the green algae Chlorella sp. Chemical Engineering Journal. 2010;170. [CrossRef]
Figure 1.
Preparation principle of SeNPs.
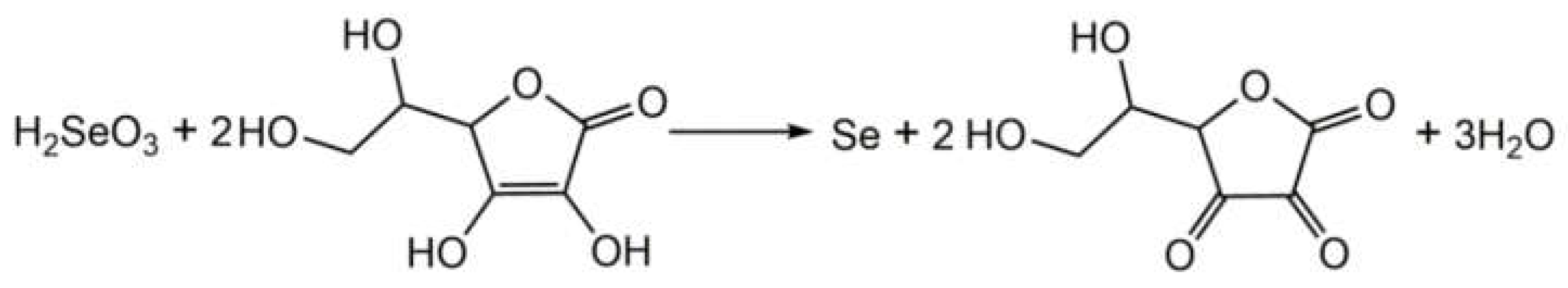
Figure 2.
Morphology and characterization of selenium nanoparticles.(A) Light photograph of CMC-SeNPs solutions. (B) The TEM image of CMC-SeNPs (a) ruler 1 μm;(b) ruler 0.5 μm; (c) ruler 100 nm; (d) ruler 0.5 nm. (C) The EDS spectrum of CMC-SeNPs. (D) Mixed system UV-vis absorption spectrum(1.mixed of ascorbic acid and CMC solution 2.mixed of ascorbic acid and selenite 3.ascorbic acid solution 4.SeNPs solution 5.CMC solution). (E) Absorbance of SeNPs reaction solution variation with time. (F) Nanoparticle size distribution intensity spectrum of SeNPs.
Figure 2.
Morphology and characterization of selenium nanoparticles.(A) Light photograph of CMC-SeNPs solutions. (B) The TEM image of CMC-SeNPs (a) ruler 1 μm;(b) ruler 0.5 μm; (c) ruler 100 nm; (d) ruler 0.5 nm. (C) The EDS spectrum of CMC-SeNPs. (D) Mixed system UV-vis absorption spectrum(1.mixed of ascorbic acid and CMC solution 2.mixed of ascorbic acid and selenite 3.ascorbic acid solution 4.SeNPs solution 5.CMC solution). (E) Absorbance of SeNPs reaction solution variation with time. (F) Nanoparticle size distribution intensity spectrum of SeNPs.
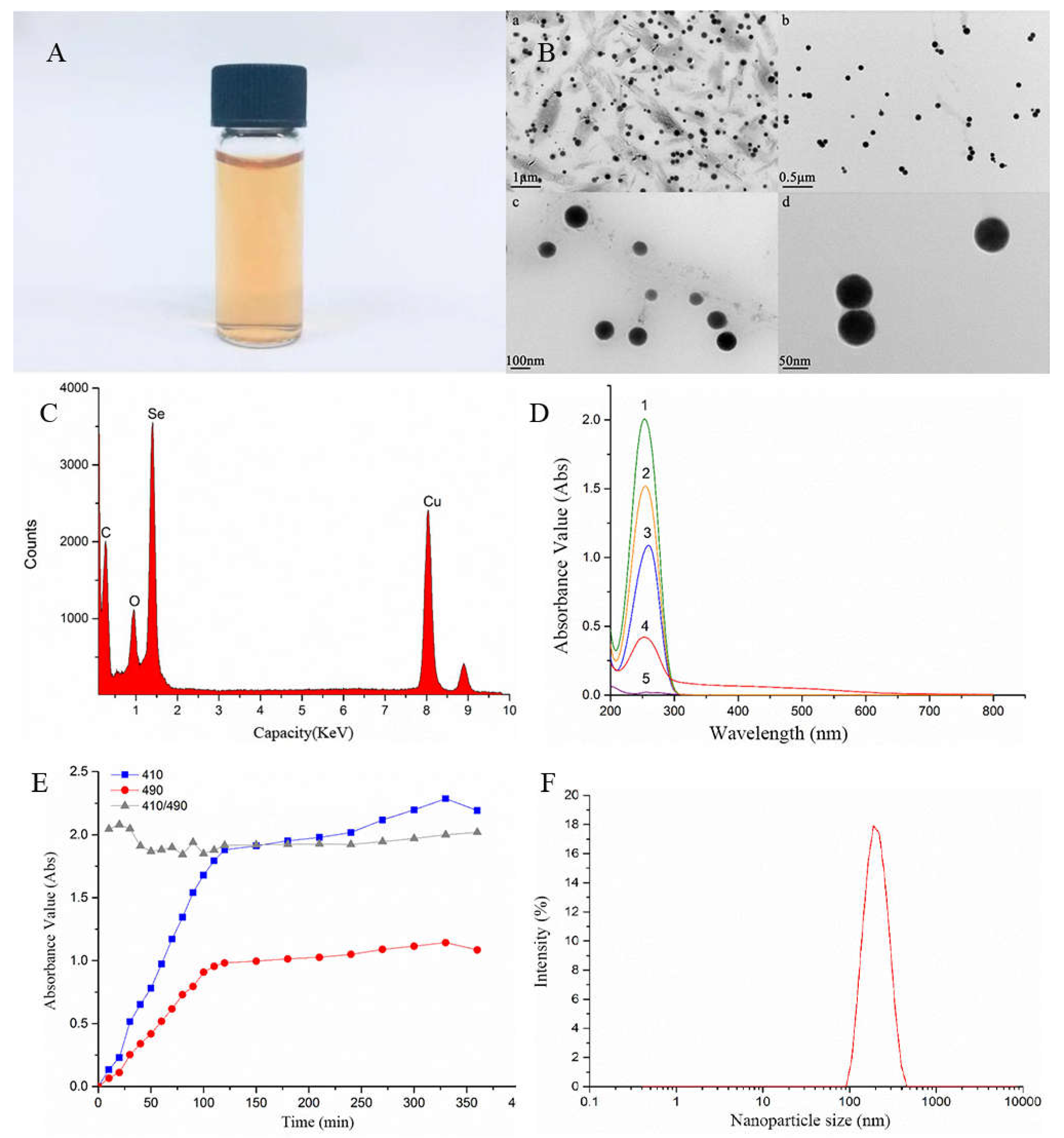
Figure 3.
Growth curve and chlorophyll change of Chlamydomonas rheinis. (A)Growth of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+ (a)CC125;(b)CC400. (B) Contents of chlorophyll a of CC125 at different concentrations with or without Cd2+ (P<0.05). (a) without Cd2+;(b)with Cd2+. (C) Contents of chlorophyll b of CC125 at different concentrations with or without Cd2+ (P<0.05). (a) without Cd2+;(b)with Cd2+. (D) Contents of carotenoid of CC125 at different concentrations with or without Cd2+ (P<0.05). (a)without Cd2+;(b)with Cd2+. (E) Contents of chlorophyll a of CC400 at different concentrations with or without Cd2+ (P<0.05). (a)without Cd2+;(b)with Cd2+. (F) Contents of chlorophyll b of CC400 at different concentrations with or without Cd2+ (P<0.05). (a) without Cd2+;(b)with Cd2+. (G) Contents of carotenoid of CC400 at different concentrations with or without Cd2+(P<0.05). (a)without Cd2+;(b)with Cd2+.
Figure 3.
Growth curve and chlorophyll change of Chlamydomonas rheinis. (A)Growth of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+ (a)CC125;(b)CC400. (B) Contents of chlorophyll a of CC125 at different concentrations with or without Cd2+ (P<0.05). (a) without Cd2+;(b)with Cd2+. (C) Contents of chlorophyll b of CC125 at different concentrations with or without Cd2+ (P<0.05). (a) without Cd2+;(b)with Cd2+. (D) Contents of carotenoid of CC125 at different concentrations with or without Cd2+ (P<0.05). (a)without Cd2+;(b)with Cd2+. (E) Contents of chlorophyll a of CC400 at different concentrations with or without Cd2+ (P<0.05). (a)without Cd2+;(b)with Cd2+. (F) Contents of chlorophyll b of CC400 at different concentrations with or without Cd2+ (P<0.05). (a) without Cd2+;(b)with Cd2+. (G) Contents of carotenoid of CC400 at different concentrations with or without Cd2+(P<0.05). (a)without Cd2+;(b)with Cd2+.
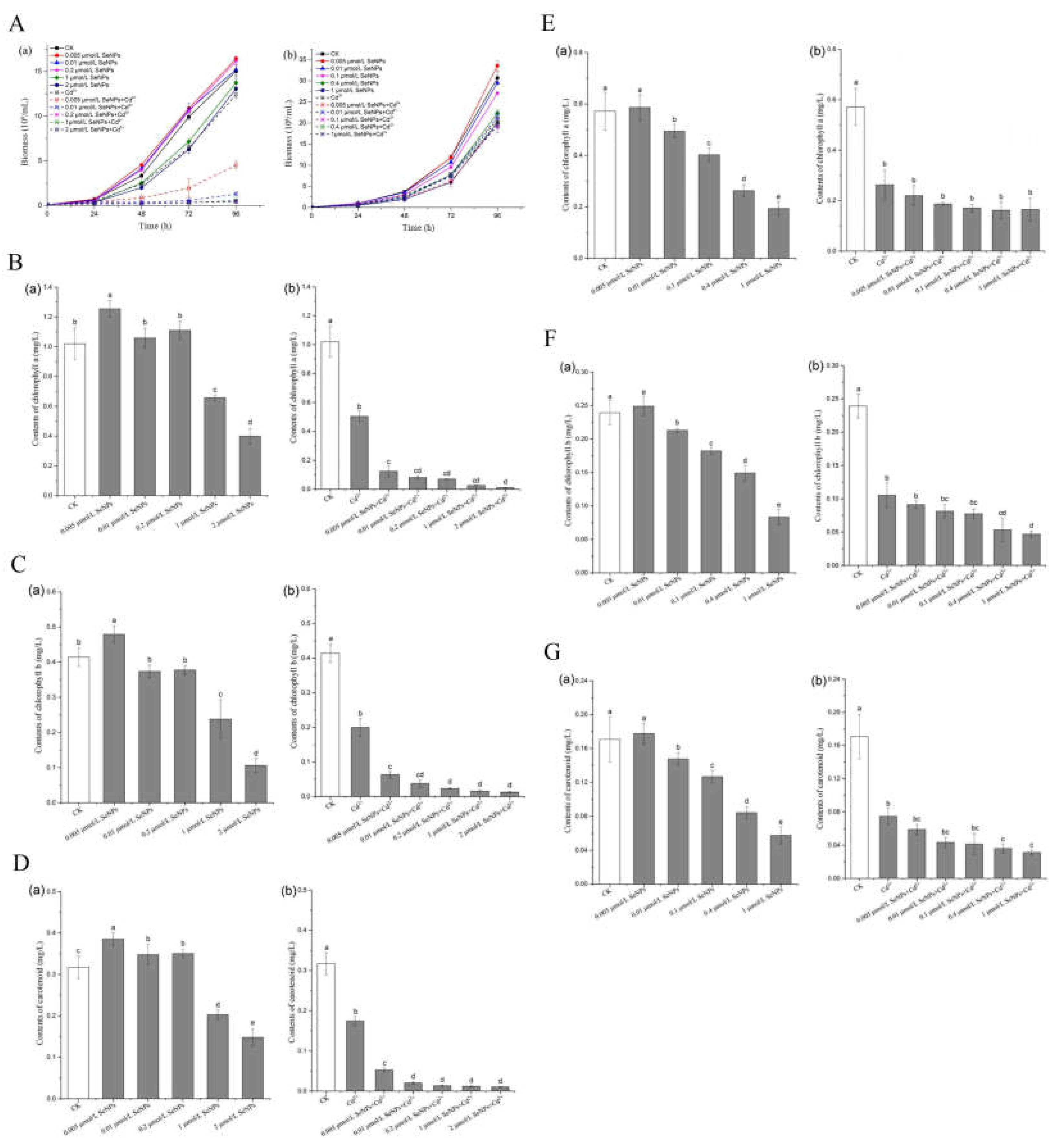
Figure 4.
Changes in levels of oxidative stress-related factors in Chlamydomonas reinhardtii. (A) Activity of T-SOD of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05).(a)CC125;(b)CC400. (B) Activity of CAT of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05). (a)CC125;(b)CC400. (C)Activity of MDA of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05).(a)CC125;(b)CC400. (D) Relative contents of ROS of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05). (a)CC125;(b)CC400.
Figure 4.
Changes in levels of oxidative stress-related factors in Chlamydomonas reinhardtii. (A) Activity of T-SOD of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05).(a)CC125;(b)CC400. (B) Activity of CAT of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05). (a)CC125;(b)CC400. (C)Activity of MDA of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05).(a)CC125;(b)CC400. (D) Relative contents of ROS of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+(P<0.05). (a)CC125;(b)CC400.
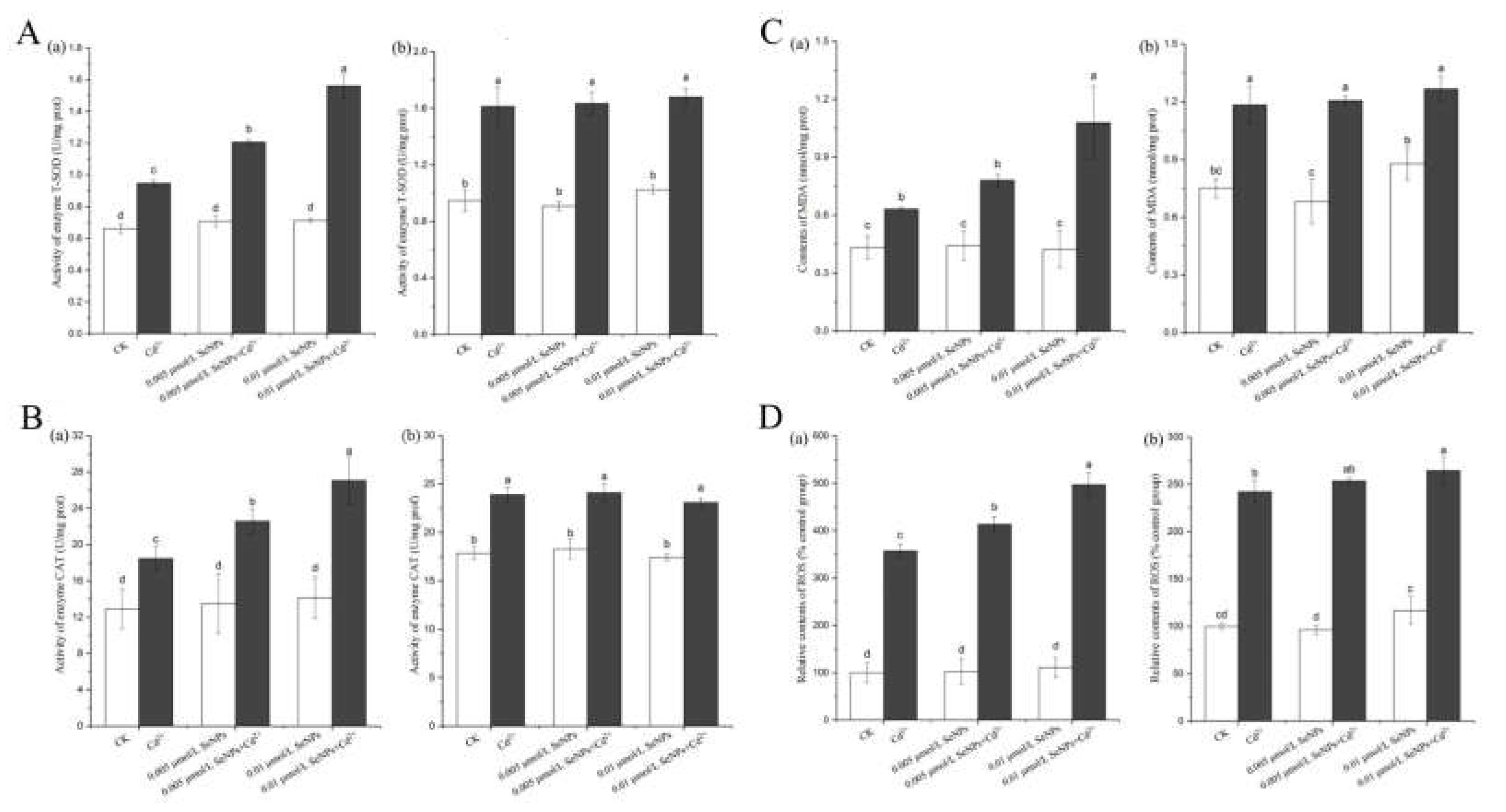
Figure 5.
Core complex structure of PSII and PSI. ©Jon Nield, Mechanistic and Structural Biology, SBCS,Queen Mary, University of London 2007 -2010.
Figure 5.
Core complex structure of PSII and PSI. ©Jon Nield, Mechanistic and Structural Biology, SBCS,Queen Mary, University of London 2007 -2010.
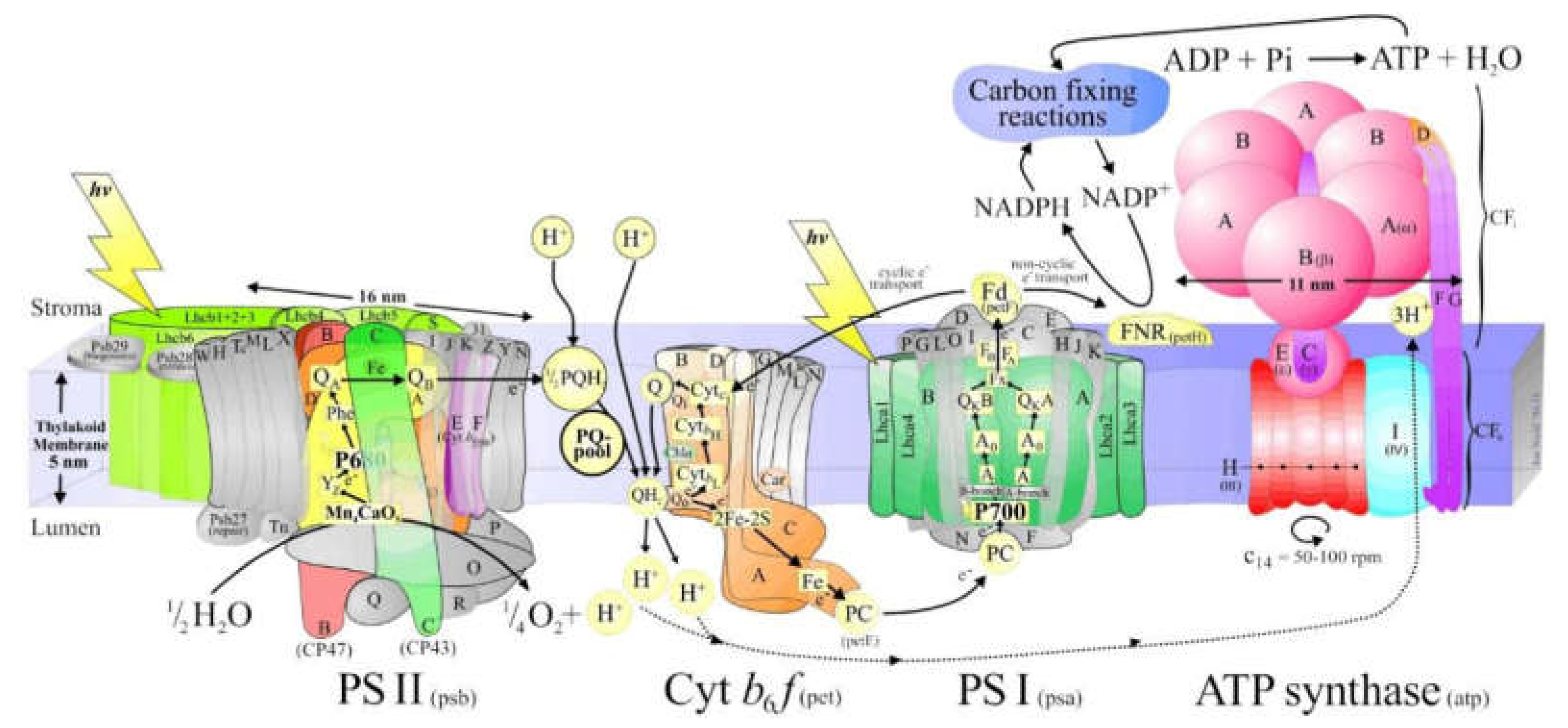
Figure 6.
Changes of photosynthetic system and intracellular cadmium content in Chlamydomonas reinhardtii. (A) Fast fluorescence rise transient of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+.(a)CC125;(b)CC400. (B) Photosynthetic parameters deduced with the JIP-test of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+.(a)CC125;(b)CC400. (C) QAreoxidation kinetics of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+.(a)CC125;(b)CC400. (D) Bioaccumulation of Cd2+ associated with different concentrations of SeNPs in Chlamydomonas reinhardtii (P<0.05). (a)CC125;(b)CC400.
Figure 6.
Changes of photosynthetic system and intracellular cadmium content in Chlamydomonas reinhardtii. (A) Fast fluorescence rise transient of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+.(a)CC125;(b)CC400. (B) Photosynthetic parameters deduced with the JIP-test of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+.(a)CC125;(b)CC400. (C) QAreoxidation kinetics of Chlamydomonas reinhardtii at different concentrations of SeNPs with or without Cd2+.(a)CC125;(b)CC400. (D) Bioaccumulation of Cd2+ associated with different concentrations of SeNPs in Chlamydomonas reinhardtii (P<0.05). (a)CC125;(b)CC400.
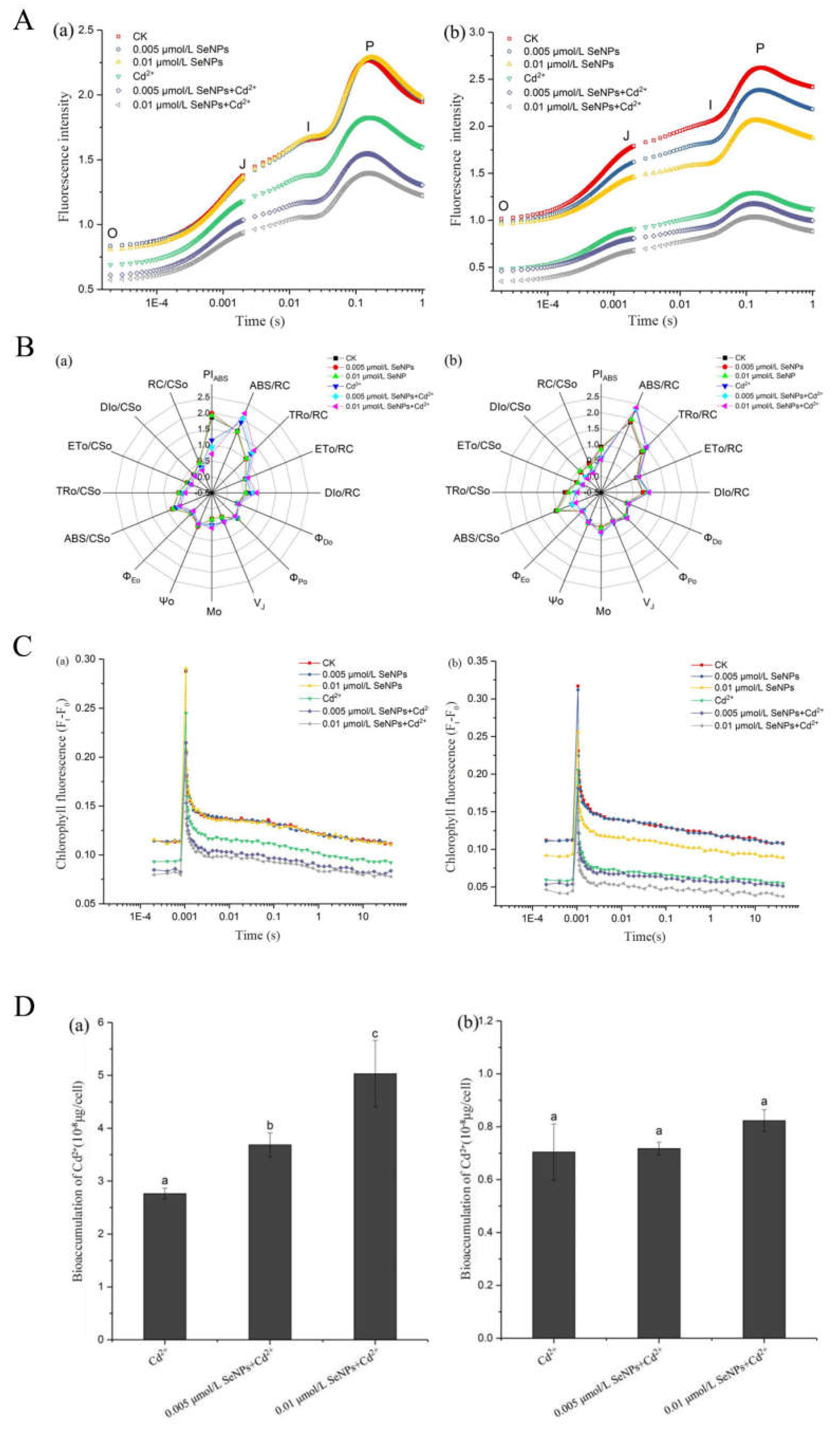
Table 3.
Regression analysis of Cd2+ inhibit the growth of Chlamydomonas reinhardtii.
| Algal cell | Fitting equation | Correlation coefficient | EC50(μmol/L) |
|---|---|---|---|
| CC125 | y=0.0023x2-0.5841x+72.283 | R²=0.8377 | 53.95 ±0.36 |
| CC400 | y=0.0236x2-1.8948x+81.089 | R²=0.9444 | 56.39 ±0.39 |
Values are mean ± standard deviation (n=3).
Table 4.
Regression analysis of SeNPs inhibit the growth of Chlamydomonas reinhardtii.
| Algal cell | Fitting equation | Correlation coefficient | EC50(μmol/L) |
|---|---|---|---|
| CC125 | y=63.364e-0.094x | R²=0.8899 | 1.88 ±0.08 |
| CC400 | y=5.4425e-0.08x | R²=0.9010 | 0.53 ±0.03 |
Values are mean ± standard deviation (n=3).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



