Submitted:
21 July 2024
Posted:
23 July 2024
You are already at the latest version
Abstract
Antibiotic resistance is a global health problem. The human gut microbiome is implicated in the dynamics of antibiotic resistance acquisition and transmission, with the gut microbiota thought to play a crucial role. This study aimed to determine the potential influence of human gut bacteria microbiota on the gut resistome and the relationship between gut background microbiota and Escherichia coli resistome. The PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines was used to systematically review studies that characterised gut microbiota and resistome using metagenomic analysis and/or those that reported gut E. coli resistome in healthy individuals. Changes in the diversity and abundance of bacterial gut microbiota and the resistome across different groups and the microbiota composition of the gut harbouring antibiotic-resistant E. coli were summarised. Findings suggest that lower bacterial microbiota diversity is likely associated with an increased abundance of the overall gut resistome. Age-related differences were observed, with younger infants exhibiting lower microbiota diversity and higher ARG abundance compared to older infants and adults. Studies that reported correlation revealed positive correlations between the compositional relative abundance of Proteobacteria and ARGs abundance, mainly driven by members within the Enterobacteriaceae family, particularly E. coli. This study also revealed that human gut microbiome studies that investigate gut resistome using metagenomic sequencing approaches in apparently healthy individuals are at least uncommon.
Keywords:
Gut microbiota
; Gut resistome
; Antibiotic resistance genes
; Escherichia coli
1. Introduction
Antibiotic resistance (AR) is a global health challenge that has resulted in continuous reduced antibiotic effectiveness and increasing morbidity and mortality due to bacterial infections [1]. The Sustainable Development Goal (SDG) indicator on antimicrobial resistance intends to lower antimicrobial-resistant infections, including those caused by Escherichia coli resistant to third-generation cephalosporins [2]. Knowledge of the factors that increase the emergence and spread of AR is essential in developing strategies and informing policies and procedures for AR stewardship, working toward the SDG goal of increasing well-being and reducing infectious diseases by 2030 [3].
The gut microbiome is an integral part of humans that significantly influences health [4,5,6]. It comprises a varied ecosystem of microorganisms that co-exist, including bacteria, fungi, and viruses; however, it is primarily dominated by bacteria, most of which are beneficial [7,8]. On the other hand, the gut can operate as a repository of antibiotic-resistant bacteria and genes [7,9,10] that can be disseminated to susceptible bacteria and the environment [11]. Antibiotic-resistant bacteria and genes have been reported in the gut microbiome, even in healthy populations, including infants who had never been exposed to antibiotics [12,13,14,15,16].
Understanding the gut bacteria resistome is crucial in the context of AR and its implications for human health. The gut bacteria resistome comprises genes conferring antibiotic resistance carried by commensals, including opportunistic pathogens. Therefore, gut bacteria should influence the ARGs composition and abundance. However, how the gut microbiota influences the gut resistome remains unclear. It is also unknown if varying microbiota contribute uniformly to AR enrichment. Unravelling these associations between the gut bacteria microbiota and the resistome would be useful in identifying potential targets for controlling antibiotic resistance in humans. It may also serve as a foundation for formulating guidelines and strategies to address antibiotic resistance, including interventions that modify the gut microbiota. Based on these, we hypothesised that the diversity of the bacteria gut microbiota influences the abundance of gut ARGs.
Next-generation sequencing (NGS), particularly metagenomic sequencing, has improved microbiome characterisation and analysis of the total gut resistome pool. Among the normal gut flora, Escherichia coli can be used as an indicator organism in healthy populations to understand the risk factors and trends in gut antibiotic resistance (AR). This is because E. coli (i) is less fastidious and, therefore, can easily be cultivated in the laboratory, (ii) is opportunistic, causing not only gastrointestinal infections but also extraintestinal infections at several sites of the human body [17,18,19,20,21], most of which are due to translocation from the gut, (iii) these infections require antibiotic treatment, and a broad range of antibiotics have been used for its treatment [22] and (iv) E. coli AR has been reported in many E. coli-associated infections in humans, with geographical variations in resistance patterns. Furthermore, the bacterium is versatile, zoonotic, and has been isolated from animals, including livestock, pets, and other primates, which are sources of acquisition for humans [23,24,25]. Given the significance of E. coli, it can be used as a proxy to elucidate the association between the gut microbiome and the resistome. Therefore, we also hypothesise that there is a relationship between the composition of the background microbiota and the E. coli resistome. This review aimed to determine the (i) potential influence of gut bacterial microbiota on the bacterial resistome and (ii) the relationship between microbiota composition and E. coli resistome. To achieve these, we reviewed published studies investigating gut microbiota and resistome and those investigating gut microbiota and E. coli resistome in apparently healthy individuals.
2. Methodology
2.1. Search Strategy
The Preferred Reporting Items for Systematic Reviews and Meta-analysis (PRISMA) approach was used to conduct the review except for meta-analysis. To identify potentially relevant articles investigating the human gut microbiota and gut resistome, or human gut microbiota and E. coli resistome, an electronic literature search was performed on Web of Science, Scopus, and PubMed databases. Separate searches were performed to address the two objectives:
Search category 1
Web of Science: Human gut microbiome OR human gut microbiota OR human gut microflora OR human gut bacteria OR human intestinal bacteria AND resistome OR antibiotic resistance genes OR ARG. Scopus: “Human gut microbiome” OR “human gut microbiota” OR “human gut microflora” OR “human gut bacteria” OR “human intestinal bacteria” AND “resistome” OR “antibiotic resistance genes” OR “ARG” PubMed: # (i) Human gut microbiome OR human gut microbiota OR human gut microflora OR human gut bacteria OR human intestinal bacteria; # (i) resistome OR antibiotic resistance genes OR ARG. Final algorithm, #(i) AND #(ii).
Search category 2.
Web of Science: Human gut microbiome OR human gut microbiota OR human gut microflora OR human gut bacteria OR human intestinal bacteria AND resistome OR antibiotic resistance genes OR antibiotic resistance OR multiple antibiotic resistance OR bacteria resistance OR multidrug resistance AND Escherichia coli OR E. coli. Scopus: “Human gut microbiome” OR “human gut microbiota” OR “human gut microflora” OR “human gut bacteria” OR “human intestinal bacteria” AND “resistome” OR “antibiotic resistance genes” OR “antibiotic resistance” OR “multiple antibiotic resistance” OR “bacteria resistance” OR “multidrug resistance” AND “Escherichia coli” OR “E. coli”. PubMed: search #(i), human gut microbiome OR human gut microbiota OR human gut microflora OR human gut bacteria OR human intestinal bacteria; #(ii), resistome OR antibiotic resistance genes OR antibiotic resistance OR multiple antibiotic resistance OR bacteria resistance OR multidrug resistance; #(iii), Escherichia coli OR E. coli. Final algorithm, # iv, #(i) AND #(ii) AND #(iii).
All fields were searched, and no date restriction was employed. The last search was performed on 25 March 2024. Bibliographic search outcomes were imported to Mendeley desktop v. 1.19.5, and duplicates were removed.
2.2. Selection of Studies
Eligible studies for inclusion were those that (i) were human studies involving healthy participants that investigated gut bacteria microbiota and resistome or gut microbiota and E. coli resistome (ii) identified gut microbiota and resistome by metagenomic sequencing, and (iii) were primary research articles published in English. Studies of the following nature were excluded: (i) those that did not characterise the microbiota by metagenomic sequencing, (ii) did not report E. coli resistome/resistance or gut resistome, (iii) targeted only specific bacteria as microbiota (iv) involved unhealthy or critically ill participants (v) involved high-risk populations likely to carry ARGs such as studies involving mass administration of antibiotics, (vi) involved participants with undefined health status and (v) preterm infants. However, only data from full-term infants, if described as part of the latter, was extracted and included in the analysis. Also, studies for which late preterm participants (33-36 weeks) constituted less than 10% of the total sample were included. Reviews, book chapters, case reports, studies in languages other than English and those that utilised publicly available data (metadata) were also excluded.
2.3. Data Extraction and Synthesis
Variables were extracted from all eligible studies following the inclusion and exclusion criteria into a predefined data extraction sheet. This included author(s), year of publication, title of study, objective, design, country of study, sample size, age of participants, methodology, sequencing platforms, bioinformatic pipelines, alpha diversity measures, microbiota composition, and E. coli resistome. Data for full-term infants was extracted and reported in cases where the studies included preterm and full-term infants. If only a subset of a study was analysed for microbiota and resistome, only the sample size of the subset was captured. Supplementary sheets of included studies were consulted for missing data in the original articles. Information not specified in a study or supplementary material was captured as data not reported (NR), and if not applicable to the specific case, was labelled not applicable (NA). Figure 1 represents the PRISMA flow diagram used in sourcing, identifying, and selecting studies included in the analysis, and Table 1 shows a summary of the characteristics of the included studies.
2.4. Outcome Measures
To determine the potential influence of gut microbiota on gut resistome, outcome measures include (i) significant changes in the microbiota alpha diversity and abundance over different time points and (ii) significant changes in the microbiota alpha diversity and abundance between comparative groups. As a secondary measure, (iii) associations/correlations between microbiota and resistome load, if available, were included.
To determine the potential association between the gut background microbiota and E. coli resistome, the composition of the background bacteria microbiota, and E. coli resistome composition was recorded, and homogenous data was summarised. A secondary measure was correlations between microbiota and E. coli ARGs/ resistance patterns.
2.5. Risk of Assessment Bias and Critical Appraisal of Eligible Studies
To limit possible bias and errors, the first author (FJ) performed the data search, and a second (RM) performed an independent search, examination, and confirmation. Two independent authors also performed preliminary screening based on titles and abstracts and data extraction from full-text articles. Any discrepancies were discussed, and the authors agreed upon a consensus. The Joanna Briggs Institute (JBI) critical appraisal checklist (https://jbi.global/critical-appraisal-tools) was used to appraise the longitudinal and cross-sectional studies before final inclusion in the analysis (Supplementary material, S1 and S2).
3. Results
3.1. Characteristics of Studies Included in the Analysis
The electronic search to identify articles to assess the potential influence of the human gut microbiota on the gut resistome yielded 9175 studies and 7640 after excluding duplicates. Following screening, only a small proportion of the studies (0.1%, 8/7640) fulfilled the study eligibility (Figure 1). Five of the eight (62.5%) eligible studies were longitudinal out of which four involved child participants within the first five years of life. Three (37.5%) studies were cross-sectional. Three studies were conducted in the USA, two in China, and one in Norway, Denmark, and Vietnam. Majority (75%, 6/8) of the studies used the MetaPhlan bioinformatic database in microbiota profiling. The gut resistome was profiled using various databases; however, protein markers were primarily sourced from the Comprehensive Antibiotic Resistance Database (CARD) (75%, 6/8) for gut resistome characterisation. The approaches used for gut microbiota and resistome characterisation in the eligible studies are shown in Table 1.
The literature search performed to source articles to determine the relationship between the bacteria gut microbiota and E. coli resistome yielded 9816 studies, with 7786 remaining after duplicate removal. Screening also revealed a small proportion (0.09%, 7/ 7786) of eligible articles to address the second objective. These constituted four longitudinal and 3 cross-sectional studies (Table 1). Gut E. coli resistome was identified from these studies by shotgun metagenomic sequence analysis (E. coli ARGs) (n = 5), functional selection using specific antibiotics (n = 1), and the use of both the Kirby Bauer disc diffusion technique and PCR detection targeting 47 ARGs in gut isolates.
Generally, four out of the eight studies eligible for objective one were also among the seven studies eligible for objective 2 (Figure 1). The publication dates of the eligible studies ranged from 2015 to 2023.

Table 1.
Summary of characteristics of included studies, testing methodology and sequencing strategies.
Table 1.
Summary of characteristics of included studies, testing methodology and sequencing strategies.
| S/N | Country of Study | Study Design | Study Objective/Hypothesis | Age of Participant and Study Time Points | Sample Size | DNA Extraction and Quantification | Sequencing Technology and Platform | Database and Bioinformatic Pipelines for Microbiota and ARG Detection | Reference | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1ab | Norway | Longitudinal | Determine resistome and mobilome across gestational ages and microbiota-modifying treatment. | 7days, 28days, 120days, 365days. |
10 | NorDiag Arrow Stool DNA Extraction kit (+bead beating) Qubit +nanodrop |
Shotgun Illumina Miseq |
Bowtie 2 MetaPhlAn3 (based on CHOCOPhlAn) MetaSPAdes and MetaQUAST (from QUAST) ShortBRED based on CARD NanoARG HUMAnN |
[26] | |
| 2ab | USA | Longitudinal | Determine factors associated with early life resistome development. | 6weeks & 1year. | 195 | Fecal DNA extraction kit Qubit |
Shotgun | MetaPhlAn PanPhlAn HUMANn2 ShotBRED based on CARD |
[27] | |
| 3ab | USA | Longitudinal | Determine potential sources of infant and maternal ARGs. | Mother-child 1month, 6months. |
10 | InviMag® Stool DNA Kit Qubit, Nanodrop |
Shotgun Illumina NextSeq |
Bowtie2 MetaPhlan2 CARD RestFinder PlasmidFinder |
[28] | |
| 4ab | Denmark | Longitudinal | Characterise the ARGs acquired during the first year of life and assess the impacts of diverse environmental exposures on ARG load. | 1 year 1 week 1 month 1 year 4 years 5 years |
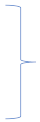
|
662- shotgun 660-16S rRNA |
PowerMagSoil DNA isolation kit | Shotgun Illumina NovaSeq 16SrRNA sequencing- Illumina Miseq |
SPAdes Humann2 MetaPhlAn MetaWRAP MetaBAT2 Bowtie2 QllME2 CARD |
[29] |
| 5a | China | Longitudinal | To understand the characteristics of the gut microbial composition | 18-69 years (Mean= 28.6) |
7 followed for 1 year (12 time points) | QIAamp Fast DNA stool minikit |
Shotgun Illumina HiSeq |
HUMAnN3, UniRef 90, KEGG, Kraken2.0, ResFinder and SPAdes | [30] | |
| 6a | Vietnam | Cross-sectional | Healthy human gut in Vietnam is a source of ARGs transferable to gut pathogens. | 0-23months 2-5years > 18yrs, |
42 | FastDNA soil kit | Shotgun Illumina |
Bowtie2 Kraken2 Bracken ARGANNOT database |
[31] | |
| 7a | USA | Cross-sectional | To characterise the microbiome and resistome of dairy workers | Mean age dairy workers, 38.4 community controls, 49.5 |
16(10 dairy workers and 6 nondairy workers | MoBio DNeasy PowerLyzer PowerSoil Kit | Shotgun Illunina HiSeq |
MetaPhlAn3, ChocoPhlAn, Anvio, Centrifuge, MEGAHIT, ABRicate, MetaCherchant, Kraken2, CARD | [32] | |
| 8a | China | Cross-sectional. | Determine antibiotic resistome shared between chicken farms and Life poultry markets workers and those with no contact with life poultry markets. | NR | 36 (18 life poultry market workers & 18 non-workers) | DNeasy PowerSoil Pro Kit agarose gel electrophoresis Qubit dsDNA assay kit |
Shotgun Illumina NovaSeq PE150. |
MEGAHIT MetaGeneMark MetaPhlAn2 CARD, ResFinder |
[33] | |
| 9b | Malaysia | Cross-sectional | Profile the gut resistome of Malaysians and investigate its association with demographic and lifestyle variables. | ≤ 90 yrs Lower boundary NR |
200 | QIAamp PowerFecal Pro DNA Kit | Shotgun Illumina NovaSeq |
BioBakery3 KneadData MetaPhlAn3 ARGs-OAP |
[34] | |
| 10b | USA | Cross-sectional | Characterise fecal, oral, and skin bacterial microbiome and resistome of the Yanomami Amerindians with no previous contact with Western people. | 4- 50yrs old | 12 | Powersoil DNA isolation kit | V4 region of the 16SrRNA Illumina HiSeq |
PICRUSt STAMP KEGG CONCOCT PARFuMS Resfams |
[35] | |
| 11b | Saudi Arabia | Cross-sectional | To assess pregnancy induced gut microbiome composition and antimicrobial resistome in Saudi females. | Mean age NP 39.1±7.7 First trimester 25.4±4.1 Third trimester 33.3±7.3 |
24 (8 NP 8 first trimester 8 third trimester) |
QIAamp Fast DNA Stool Mini Kit | 16S rRNA Illumina MiSeq |
NR | [36] | |
Superscripts a-Eligible studies for objective 1; b- Eligible studies for objective 2; ab Eligible studies for objective 1 and 2.
3.2. The Potential Influence of the Gut Microbiome on Gut Resistome
To determine the potential influence of gut microbiota on gut bacterial resistome, changes in the alpha diversity in gut microbiota between participant groups or longitudinal sample time points were summarised comparable to the changes observed in the resistome between these groups. Correlations between microbiota and resistome from individual studies were also extracted with the aim of performing a meta-analysis on similar populations. However, due to high dissimilarity in populations based on age, and other significant heterogeneity among the studies, a robust meta-analysis was not conducted. Therefore, the results from individual studies were summarised, and observed trends and consistent findings across studies were reported as a narrative synthesis.
Shannon’s diversity index was the primary metric utilised to describe alpha diversity in six of the eight studies, Simpson’s index was utilised in one study, and OTU (Operational Taxonomic Unit) richness was used in one. Changes in diversity of the gut microbiome and abundance of ARGs are shown in Table 2. Except for one of the studies whereby an adult group was compared to data from the human microbiome project, all other studies that showed a lower diversity of bacteria microbiota compared to a comparison group also reported a relatively higher resistome abundance [26,27,28,29,31,33]. Therefore, lower gut microbiota diversity is likely associated with higher gut ARGs gene load. This association was clearly visible when considering age or longitudinal study time points [26,27,31]. Neonates and younger infants had a less diverse microbiota, which matures over time [31], particularly within the first two years, compared to older infants (Table 2). Concurrently, the gut ARGs were relatively abundant in the younger than older participants. This association was reported between 5 groups in four studies [26,27,28,31] (Table 2).
Based on the composition of various microbiota taxa, there was evidence of positive significant correlations between microbiota compositional structure and ARGs gene load [26,27,28,29]. A strong positive correlation between the compositional relative abundance of Proteobacteria and gut ARG abundance was reported in two studies [27,28]. Also, amongst the Proteobacteria, E. coli was reported to have the strongest positive association with resistome abundance, [26,27,28,29,34,36], followed by other members of Enterobacteriaceae, including Klebsiella pneumoniae, K. aerogenes, Citrobacter and Enterobacter [26] E. coli was identified as the strongest predictor of ARGs in infants, and according to Li et al. [29], it haboured over 51% (68/133) of the different ARGs in the gut of young infants. Two of the four studies. Conversely, there was a negative correlation between Bifidobacterium and ARG load.
Although the potential influence of external factors was not considered as part of the core objective of this study, two studies involved healthy participants who were exposed to poultry [33] and dairy [36], respectively, which are environments that may enhance antibiotic resistance acquisition. Although both the exposed and unexposed to live poultry markets were healthy residents within the same geographical area, the unexposed group exhibited a higher phylogenetic diversity and lower corresponding ARG diversity and abundance than the exposed similar to the previous trend described (Table 2).
3.3. The Potential Association between Gut Microbiota and E. coli Resistome
To examine if there was an association between gut background microbiota composition and E. coli resistome, using E. coli resistome as a proxy for gut resistome, seven studies reported resistance in E. coli and characterised the gut microbiota by metagenomic sequencing. Generally, there was a significantly low level of detail or comprehensive description provided for gut E. coli ARGs and gut microbiota composition and high heterogeneity in microbiota and corresponding E. coli resistome data (Table 3 For example, one study detailed the E. coli resistome and gut bacteria composition in 7-day-old full-term infants [26], another [34] presented microbiota only for ARG-containing species, while the detailed microbiota composition was unavailable in the study by Lebeaux et al. [27]. Also, Clemente et al. [35] reported the full gut E. coli resistome in a significantly different population, which are older children and adults (4-50 years), and resistance was based on the functional selection of ARGs using specific antibiotics as opposed to the other studies that used shotgun metagenomics for E. coli resistome detection. Also, Khan et al. [36] reported the microbiota composition and resistance profiles of pregnant and non-pregnant women. Although the study reported varying levels of E. coli resistance, there was no differentiation between ARGs associated with E. coli and other gut bacteria, making it difficult in definitively attributing all the ARGs solely to E. coli.
Despite the significant variations and heterogeneity in the data from the 7 studies, which reduced the feasibility of drawing concrete conclusions about the relationship between gut microbiota composition and the E. coli resistome, we managed to summarise the composition of E. coli resistome and the associated bacterial microbiota based on a subset of more homogeneous data. Bifidobacterium, followed by Escherichia, was identified as the most abundant genera in guts of participants less than 6 months old. The genera Bifidobacterium, Prevotella, Collinsella, Eubacterium and Ruminococcus belonging to Actinobacteria, Bacteroides and Firmicutes were the most abundant in the adult-dominated group. It was observed that while Escherichia and other members of the Enterobacteriaceae form part of the top five most abundant genera in infant groups, Escherichia, particularly Escherichia coli, was not part of the top five in the adult-dominated group. Khan et al. [36] also presented bacteria families in non-pregnant adult women where families belonging to the Proteobacteria phylum were not among the most abundant five. Table 4 summarises the most abundant genera in the gut of infants and adults that display E. coli AR.
Summary findings from the studies involving 0-1-year-old participants revealed the multidrug resistance efflux pumps as the most diverse and abundant class of E. coli ARGs. This was followed by ARGs encoding resistance to polypeptides, Beta-lactams and Fosfomycins. Table 5 summarises the composition of E. coli resistome identified from the studies.
4. Discussion
The current emergence, acquisition and spread of antibiotic resistance are alarming and need urgent attention. The SDG indicator on AMR intends to lower antimicrobial-resistant infections, working towards achieving the SDG goal of increasing well-being and reducing infectious diseases by 2030 [3]. The gut microbiome, particularly that of the developing infant, is crucial to human health. Belonging to this dynamic consortium, gut resistome studies are vital to continuously unravel important data that aid in developing strategies to reduce the spread of antibiotic resistance. This study provides insights into the gut bacteria microbiota that potentially influences the abundance of gut resistome.
A healthy human gut is thought to have a properly balanced bacteria microbiota, and the core microbiota has co-evolved with humans [8,37,38]. It is considered harmless and plays a vital role in various activities beneficial to health [39]. The findings of this review showed that a decrease in the diversity of gut bacteria microbiota was concurrent with an increased relative abundance of gut resistome; an increase in the abundance of gut resistome is caused mainly by the increase in the relative abundance of Proteobacteria, particularly the Enterobacteriaceae; and among the Enterobacteriaceae, E. coli was the primary source ARGs. Also, due to the low gut microbiota maturity and resulting lower diversity in infants, a higher relative abundance of Proteobacteria compared to older age groups is prevalent and associated with abundant ARGs that reduced over time. Generally, following delivery, the neonatal gut is abundant in oxygen; therefore, the presence of strict anaerobes is unlikely. Compared to most gut enterotypes that are obligate anaerobes, Proteobacteria are facultative anaerobes and, therefore, tolerate the oxygen-rich neonatal gut, justifying the disparity in gut Proteobacteria abundance between newborn infants and older children and adults. Proteobacteria are thought to be essential for the consumption of oxygen and the reduction of redox potential. Thus, they prepare the gut for colonisation by the more stable strict anaerobes [40]. It is worth noting that Proteobacteria, particularly those of the Gamma-proteobacteria group, are also significantly involved in conjugation, hence horizontal gene transfer [41]. This characteristic increases the odds of disseminating antibiotic resistant genes to other gut bacteria species, increasing the gut resistome. Therefore, the prevalence of gut ARGs in neonates could be reduced by regulating the relative abundance of Proteobacteria, particularly E. coli, thus reducing further dissemination of ARGs to other gut bacteria and the environment.
The findings of this study of Enterobacteriaceae as significant contributor to ARGs load, mirrors WHO report of Global deaths attributed to and associated with bacterial antimicrobial resistance, which ranks antibiotic-resistant E. coli as the leading pathogen associated with global deaths [42]. Also, out of the approximately 700,000 deaths worldwide that are attributed to antibiotic-resistant infections, approximately 200,000 of these occur in neonates in the first four weeks of life [39]. Increased global deaths are due to E. coli that are resistant to third generation cephalosporins, fluoroquinolone-resistant E. coli, and other Gamma Proteobacteria such as carbapenem-resistant Acinetobacter baumannii, carbapenem-resistant Klebsiella pneumoniae, and third-generation cephalosporin-resistant Klebsiella pneumoniae [42]
Although our findings reveal Proteobacteria and particularly E. coli as the major gut bacteria strains harbouring ARGs, the possibility of bias cannot totally be refuted, which may also account for a proportion of the identified ARGs linked to E. coli. Like other databases, ARG databases may overrepresent markers frequently identified in certain bacterial species, such as those encoding resistance in E. coli. Historically, E. coli has been extensively studied and is still a research focus, and its associated ARGs are undoubtedly inclusive with available genomic data. As a result, the database may contain more known markers for ARGs associated with E. coli than for other gut bacteria.
Various environmental exposures may influence the gut microbiota, consequently influencing the resistome. Two studies examined the microbiome and overall gut resistome in participants exposed to dairy and poultry. One [32] found no significant difference in taxonomic abundance between dairy and non-dairy workers and lower ARGs load in dairy compared to non-dairy workers, attributed likely to the lower sequencing depth. On the other hand, the other [34] reported significantly less diverse but higher ARGs of live poultry market workers compared to the control group, probably due to environmental exposures. It is worth noting that poultry is one of the agricultural sectors with high usage of antibiotics as growth promoters, and a correlation has been shown between the abundance of human gut ARGs and antibiotic exposure in animals [43]. These antibiotics increase microbiome dysbiosis, ultimately decreasing diversity leading to a less stable gut microbiota composition and increased ARGs, primarily due to Proteobacteria enrichment. A previous study reported an increase in the diversity and abundance of ARGs in participants exposed to antibiotics within six months prior to the commencement of the study compared to a lower diversity in the antibiotic-unexposed individuals [44]. Specifically, the relative abundance of E. coli, at 0.1% in the antibiotic-unexposed group, increased to nearly 10% in the antibiotic-exposed group.
The gut bacterial population of healthy humans is mainly dominated by the phyla Firmicutes and Bacteroidetes, with a lesser abundance of Actinobacteria, Proteobacteria, and other minor phyla [44,45]. However, of the Actinobacteria phylum, members of the genus Bifidobacterium are the most abundant, dominating the gut of healthy infants, similar to what we observed in participants guts harbouring AR E. coli. However, this is almost always followed by abundance in Escherichia or other Proteobacteria genera in infants, as opposed to Actinobacteria or Bacteroidetes genera in adults. Although indicating resistance, a healthy gut dominated by Bifidobacterium spp. highlights its importance in modulating and enhancing metabolic and mitochondrial activities [46]. Bifidobacteria function in vitamin and protein synthesis, digestion supplementation, immune system stimulation and suppression of the growth of exogenous organisms [47]. In a previous study by Gagnon and colleagues [48], two strains of Bifidobacterium isolated from infant faeces were shown to be resistant to bile, acid, and lysozyme and inhibited enterohemorrhagic Escherichia coli serotype O157:H7 in vitro, decreasing its adherence to human enterocyte-like CaCo-2 cells. Inhibition of the potentially pathogenic E. coli serotype was associated with increased concentration of the Bifidobacterium species [48]. This shows that an increased relative abundance of Bifidobacterium would be beneficial to human health as it suppresses the abundance of the E. coli population, resulting in a reduction in E. coli antibiotic resistance and overall gut resistance.
The current study also identified efflux pump associated ARGs as the significant contributor to E. coli resistome. The mechanisms by which efflux pumps confer resistance vary from intrinsic to overexpression of the pumps or mutations in the repressor genes or other encoding genes [49,50]. These mechanisms successfully lower intracellular concentrations of several antibiotics, a major source of the multidrug resistance traits observed. For example, the AcrAB efflux pumps confer resistance to multiple antibiotics, including tetracycline, quinolones, and fluoroquinolones [51]. E. coli 's central involvement as a multidrug resistance pathogen that increases the gut antibiotic resistance gene pool and subsequent spread. This could be related to its ranking as the leading pathogen associated with global mortality due to resistance [42].
Generally, antimicrobial resistance ranks among the most significant risks to global public health linked to approximately five million deaths in 2019 [42]. The WHO report is particular about generating antimicrobial resistance programs and initiatives especially in low- and medium-income countries (LMICs). However, it alludes to the fact that these countries are the most severely challenged by the lack of well-characterised data and having huge knowledge gaps that hamper the effective control of antibiotic resistance [52]. In this review, over 7000 articles were screened to answer the core objectives, and approximately only 0.1% of studies were eligible to address each with minimal representation from LMICs, highlighting a huge dearth of data, which aligns to the WHO report. Also, since the development of NGS in the early to mid-2000s [53], it has been applied in many biological scientific fields, yet the earliest article in the current review was published in 2015, approximately two decades further. It is worth mentioning that unlike culture-based techniques, metagenomic sequencing has the added advantage of profiling the total gut microbiome and resistome and can also reveal novel ARGs and plasmids carrying these genes. The low global uptake indicates that research using innovative technology to generate antibiotic-resistant, well-characterised data sets is still uncommon. Therefore, this research area needs to be fostered to provide valuable data for strategic intervention. Although not undermining the significance of nosocomially-acquired antibiotic-resistant infections, clinical cases most often stem from community carriers, likewise antibiotic resistance. If human gut resistome community data are available, including data on how the microbiota evolves with the overall antibiotic resistance or its evolution with the resistome of a targeted species such as E. coli and other confounding factors that may mitigate gut resistance, interventions will be evidence-directed. These interventions would be aimed at regulating identified drivers of antibiotic resistance, thus, reducing reported morbidity and mortality, and other negative impact of human, environmental and economic consequences of increasing antibiotic resistance [54,55,56].
From knowledge of this field, coupled with the finding of this review that revealed the enrichment of ARGs by the increase in the abundance of specific bacteria taxa, it is important to consider the modification of the gut microbiota, such as the use of specific probiotics and prebiotics therapies as future strategies in addressing antibiotic-resistant in humans. Probiotics have been shown to decrease the abundance of antibiotic-resistant bacteria through competitive exclusion and production of antibacterial compounds, improve immunity through immune system modulation, and restore a balanced microbiota after gut dysbiosis following antibiotic treatment [57,58,59,60,61]. Probiotic research is ongoing, particularly knowledge of its use as a complementary therapy. The efficacy has been shown to depend on various factors, including individual variations in the microbiome of each person, the specific probiotic strain(s) involved, and the human health status and individual characteristics [62]
4.1. Conclusion and Future Directives
In conclusion, the bacteria gut microbiota is associated with the gut resistome. Lower diversity in microbiota is likely a significant contributor to higher gut resistome load and vice versa. Also, a higher relative abundance of Proteobacteria, especially members within the Enterobacteriaceae family, particularly E. coli, can significantly contribute to resistome enrichment. Despite high heterogeneity and low granularity in the data, there was evidence that Bifidobacterium was the most abundant genera in healthy participants carrying E. coli ARGs. In infants, this was followed by Escherichia or other Proteobacteria genera, as opposed to genera of the Actinobacteria or Bacteroidetes families in adults. There is a dearth of studies that use metagenomic sequencing technologies to explore human gut microbiota and resistome, the outcome of which could be used to identify drivers in the spread and dissemination of antibiotic resistance.
A critical component of the solution to antibiotic resistance is high-quality and innovative research that produces large and well-characterised prospective data sets. Such data are essential in developing evidence-driven policy briefs that strengthen regulations and foster effective antibiotic resistance stewardship programs. Therefore, more studies should be undertaken in this area. Exploring publicly available gut metagenomic datasets, which were not initially set to examine the resistome, also holds the potential to yield valuable insights into the gut resistome. Such analysis would contribute to narrowing the substantial research gap, complementing our understanding of the microbiota influence on the resistome and assist in identifying other microbiome drivers associated with gut ARG enrichment. Additionally, research could also focus on investigating the fundamental mechanisms that link gut microbiota diversity to the abundance of antibiotic resistance genes. This could involve exploring metabolic pathways or microbial interactions that facilitate the transfer of resistance genes with particular attention to critical periods in early life when the gut microbiota is most susceptible to resistome enrichment.
4.1. Strengths and Limitations of the Study
This study unveils the potential association between bacteria microbiota and gut resistome by looking at studies that employed metagenomic sequencing, which offers a more in-depth perspective of the microbiota and resistome compared to conventional culture-based approaches or other molecular-based techniques, a significant strength in this review. Despite this strength, the observations here should be taken in the context of the limitation that no cut-off restrictions were made regarding the number of study participants for the eligibility of the studies. Appropriate sample size increases precision and higher power to determine minor effects. Also, studies included were limited and from a limited number of countries despite the comprehensive search, which may limit the generalizability of the findings.
Author Contributions
Conceptualisation, J.F. and P.O.B; Methodology J.F. and P.O.B.; validation, J.F., M.R., L.G.M., and P.O.B.; formal analysis, J.F., M.R., L.G.M. and P.O.B.; writing—original draft preparation, F.J; writing—review and editing, F.J., M.R., M.L.G. and P.O.B.; supervision P.O.B.; project administration, L.G.M and P.O.B.; funding acquisition, P.O.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the South African Medical Research Council (SAMRC) through its Division of Research Capacity Development under the SAMRC Extramural Postdoctoral Programme. The content and findings reported are the sole deduction, view and responsibility of the researchers and do not reflect the official position and sentiments of the SAMRC.
Data availability statement
Not applicable.
Acknowledgments
The authors acknowledge the support of the University of Venda, South Africa, host to the SAMRC-UNIVEN Antimicrobial Resistance and Global Health Research Unit.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organisation. Antibiotic resistance. WHO, 2023a. Available online: https://www.who.int/news-room/fact-sheets/detail/antibioticresistance#:~:text=Bacteria%2C%20not%20humans%20or%20animals,hospital%20stays%2C%20and%20increased%20mortality (accessed on 18 September 2023).
- World Health Organisation. Proportion of bloodstream infection due to Escherichia coli resistant to third generation cephalosporins. WHO, 2023b. Available online: https://www.who.int/data/gho/data/indicators/indicator-details/GHO/sdg-3.d.2--proportion-of-bloodstream-infections-due-to-selected-antimicrobial-resistant-organisms--median- (accessed on 3 November 2023).
- The United Nations. The Sustainable Development Goal Report, 2023. UN 2023. Available online: https://unstats.un.org/sdgs/report/2023/The-Sustainable-Development-Goals-Report-2023.pdf. (accessed on 3 November 2023).
- Million, M.; Diallo, A.; and Raoult, D. Gut microbiota and malnutrition. Microb pathog 2017, 106, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Bhattacharjee, R.; Nandi, A.; Sinha, A.; Kar, S.; Manoharan, N.; Mitra, S.; Mojumdar, A.; Panda, P.K.; Patro, S.; Dutt, A. Phage delivered CRISPR-Cas system to combat multidrug-resistant pathogens in gut microbiome. Biomed. Pharmacother. 2022, 151, 113122. [Google Scholar] [CrossRef] [PubMed]
- Priadko, K.; Romano, L.; Olivieri, S.; Romeo, M.; Barone, B.; Sciorio, C.; Spirito, L.; Morelli, M.; Crocetto, F.; Arcaniolo, D.; Mirone, V. Intestinal microbiota, intestinal permeability and the urogenital tract: is there a pathophysiological link? J. Physiol. Pharmacol 2022, 73. [Google Scholar]
- Moore, A.M.; Patel, S.; Forsberg, K.J.; Wang, B.; Bentley, G.; Razia, Y.; Qin, X.; Tarr, P.I.; Dantas, G. Pediatric fecal microbiota harbor diverse and novel antibiotic resistance genes. PloS one 2013, 8, e78822. [Google Scholar] [CrossRef] [PubMed]
- Purkait, D.; Hameed, S.; Fatima, Z. Gut microbiome: Current development, challenges, and perspectives. New and Future Developments in Microbial Biotechnology and Bioengineering, Gupta V.K Ed. Elsevier: Amsterdam, Netherlands, 2020; pp.227-241.
- Penders, J.; Stobberingh, E.E.; Savelkoul, P.H.; Wolffs, P.F. The human microbiome as a reservoir of antimicrobial resistance. Front. in Microbiol. 2013, 4, 87. [Google Scholar] [CrossRef] [PubMed]
- von Wintersdorff, C.J.; Wolffs, P.F.; van Niekerk, J.M.; Beuken, E.; van Alphen, L.B.; Stobberingh, E.E.; Oude Lashof, A.M.; Hoebe, C.J.; Savelkoul, P.H.; Penders, J. Detection of the plasmid-mediated colistin-resistance gene mcr-1 in faecal metagenomes of Dutch travellers. J. Antimicrob. Chemother. 2016, 71, 3416–3419. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Ellabaan, M.M.H.; Charusanti, P.; Munck, C.; Blin, K.; Tong, Y.; Weber, T.; Sommer, M.O.A.; Lee, S.Y. Dissemination of antibiotic resistance genes from antibiotic producers to pathogens. Nat. Commun 2017, 8, 15784. [Google Scholar] [CrossRef] [PubMed]
- DeFrancesco, A.S.; Tanih, N.F.; Samie, A.; Guerrant, R.L.; Bessong, P.O. Antibiotic resistance patterns and beta-lactamase identification in Escherichia coli isolated from young children in rural Limpopo Province, South Africa: The MAL-ED cohort. S. Afr. Med. J. 2017, 107, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Li, B.; Jiang, X.; Yang, Y.; Wells, G.F.; Zhang, T.; Li, X. Antibiotic resistome in a large-scale healthy human gut microbiota deciphered by metagenomic and network analyses. Environ. Microbiol. 2018, 20, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, A.J.; Wang, B.; Sun, X.; Kennedy, E.A.; Hernandez-Leyva, A.; Ndao, I.M.; Tarr, P.I.; Warner, B.B.; Dantas, G. Persistent metagenomic signatures of early-life hospitalisation and antibiotic treatment in the infant gut microbiota and resistome. Nat. Microbiol. 2019, 4, 2285–2297. [Google Scholar] [CrossRef]
- Chong, C.W.; Alkatheeri, A.H.S.; Ali, N.; Tay, Z.H.; Lee, Y.L.; Paramasivam, S.J.; Jeevaratnam, K.; Low, W.Y.; Lim, S.H.E. Association of antimicrobial resistance and gut microbiota composition in human and non-human primates at an urban ecotourism site. Gut pathogens 2020, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.J.; Sands, K.; Thomson, K.; Portal, E.; Mathias, J.; Milton, R.; Gillespie, D.; Dyer, C.; Akpulu, C.; Boostrom, I.; Hogan, P. Antibiotic resistance genes in the gut microbiota of mothers and linked neonates with or without sepsis from low-and middle-income countries. Nat. Microbiol. 2022, 7, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Narayan, S.; Nayak, A.; King, C. L. Inflammatory pseudotumor of the liver with Escherichia coli in the sputum. Case Reports in Medicine 2015, 1, 249210. [Google Scholar]
- Kobayashi, T.; Ikeda, M.; Okada, Y.; Higurashi, Y.; Okugawa, S.; Moriya, K. Clinical and microbiological characteristics of recurrent Escherichia coli bacteremia. Microbiol. Spectr. 2021, 9, e01399–21. [Google Scholar] [CrossRef] [PubMed]
- Geurtsen, J.; de Been, M.; Weerdenburg, E.; Zomer, A.; McNally, A.; Poolman, J. Genomics and pathotypes of the many faces of Escherichia coli. FEMS Microbiol. Rev. 2022, 46, fuac031. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, A.S.; Alsakini, A.H.; Ali, M.R. Outbreak of drug resistance Escherichia coli phylogenetic F group associated urinary tract infection. Iran J Microbiol 2022, 14, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, R.N.; Juboory, Y.H.O.; Noomi, B.S. Antibiotic Susceptibility Profile and Virulence Factors Profile of E. coli Isolated from Otitis Media. Ann. Romanian Soc. Cell Biol. 2023, 27, 105–111. [Google Scholar]
- Ryan (2023). Escherichia coli (E. coli) Infections Medication. MedScape, Drugs and diseases. Available online: https://emedicine.medscape.com/article/217485-medication?form=fpf. (accessed on 3 June 2023).
- Campbell, T.P.; Sun, X.; Patel, V.H.; Sanz, C.; Morgan, D.; Dantas, G. The microbiome and resistome of chimpanzees, gorillas, and humans across host lifestyle and geography. The ISME journal 2020, 14, 1584–1599. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.T.; Oliveira, R.P.; Silva, B.F.; Monteiro, G.P.; Saut, J..PE., Costa, L.R.M., Dias, S.D.C., Rossi, D.A. Phylogeny and Virulence Factors of Escherichia coli Isolated from Dogs with Pyometra. Vet Sci, 2022, 9,158.
- Sajeev, S.; Hamza, M.; Rajan, V.; Vijayan, A.; Sivaraman, G.K.; Shome, B.R.; Holmes, M.A. Resistance profiles and genotyping of extended-spectrum beta-lactamase (ESBL)-producing and non-ESBL-producing E. coli and Klebsiella from retail market fishes. Infect. Genet. Evol. 2023, 112, 105446. [Google Scholar] [CrossRef] [PubMed]
- Bargheet, A.; Klingenberg, C.; Esaiassen, E.; Hjerde, E.; Cavanagh, J.P.; Bengtsson-Palme, J.; Pettersen, V.K. Development of early life gut resistome and mobilome across gestational ages and microbiota-modifying treatments. EBioMedicine 2023, 92. [Google Scholar] [CrossRef] [PubMed]
- Lebeaux, R.M.; Coker, M.O.; Dade, E.F.; Palys, T.J.; Morrison, H.G.; Ross, B.D.; Baker, E.R.; Karagas, M.R.; Madan, J.C.; Hoen, A.G. The infant gut resistome is associated with E. coli and early-life exposures. BMC Microbiol. 2021, 21, 201. [Google Scholar] [CrossRef] [PubMed]
- Pärnänen, K.; Karkman, A.; Hultman, J.; Lyra, C.; Bengtsson-Palme, J.; Larsson, D.J.; Rautava, S.; Isolauri, E.; Salminen, S.; Kumar, H.; Satokari, R. Maternal gut and breast milk microbiota affect infant gut antibiotic resistome and mobile genetic elements. Nat. commun. 2018, 9, 3891. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Stokholm, J.; Brejnrod, A.; Vestergaard, G.A.; Russel, J.; Trivedi, U.; Thorsen, J.; Gupta, S.; Hjelmsø, M.H.; Shah, S.A.; Rasmussen, M.A. The infant gut resistome associates with E. coli, environmental exposures, gut microbiome maturity, and asthma-associated bacterial composition. Cell Host & Microbe 2021, 29, 975–987. [Google Scholar]
- Zhang, W.; Han, N.; Zhang, T.; Qiang, Y.; Peng, X.; Li, X.; Kan, B. The spatial features and temporalchanges in the gut microbiota of a healthy Chinese population. Microbiol. Spectr. 2022, 10, pp.e01310-22.
- Pereira-Dias, J.; Nguyen Ngoc Minh, C.; Tran Thi Hong, C.; Nguyen Thi Nguyen, T.; Ha Thanh, T.; Zellmer, C.; Chung The, H.; Pike, L.; Higginson, E.E.; Baker, S. The gut microbiome of healthy Vietnamese adults and children is a major reservoir for resistance genes against critical antimicrobials. J. Infect. Dis. 2021, 224 (Supplement_7), S840–S847. [Google Scholar] [CrossRef] [PubMed]
- Trinh, P.; Roberts, M.C.; Rabinowitz, P.M.; Willis, A.D. Differences in gut metagenomes between dairy workers and community controls: a cross-sectional study. bioRxiv. 2023 (Preprint).
- Wang, Y.; Lyu, N.; Liu, F.; Liu, W.J.; Bi, Y.; Zhang, Z.; Ma, S.; Cao, J.; Song, X.; Wang, A.; Zhang, G. More diversified antibiotic resistance genes in chickens and workers of the live poultry markets. Environ. Int. 2021, 153, 106534. [Google Scholar] [CrossRef] [PubMed]
- Dwiyanto, J.; Huët, M.A.L.; Hussain, M.H.; Su, T.T.; Tan, J.B.L.; Toh, K.Y.; Lee, J.W.J.; Rahman, S.; Chong, C.W. Social demographics determinants for resistome and microbiome variation of a multiethnic community in Southern Malaysia. npj Biofilms and Microbiomes 2023, 9, 55. [Google Scholar] [CrossRef]
- Clemente, J.C.; Pehrsson, E.C.; Blaser, M.J.; Sandhu, K.; Gao, Z.; Wang, B.; Magris, M.; Hidalgo, G.; Contreras, M.; Noya-Alarcón, Ó.; Lander, O. The microbiome of uncontacted Amerindians. Sci. Adv. 2015, 1, e1500183. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Yasir, M.; Farman, M.; Kumosani, T.; AlBasri, S.F.; Bajouh, O.S.; Azhar, E.I. Evaluation of gut bacterial community composition and antimicrobial resistome in pregnant and non-pregnant women from Saudi population. Infect. drug Res. 2019, pp.1749-1761.
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; Bertalan, M. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Kleerebezem, M. The human gut microbiome: are we our enterotypes? Microb. Biotechnol. 2011, 4, 550. [Google Scholar] [CrossRef]
- Lozupone, C.A, Stombaugh, J.I, Gordon, J.I, Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220-30.
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Ellabaan, M.M.H.; Charusanti, P.; Munck, C.; Blin, K.; Tong, Y.; Weber, T.; Sommer, M.O.A.; Lee, S.Y. Dissemination of antibiotic resistance genes from antibiotic producers to pathogens. Nat. Commun 2017, 8, 15784. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; Johnson, S.C. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, X.; Lu, N.; Zhu, B. The abundance of antibiotic resistance genes in human guts has correlation to the consumption of antibiotics in animal. Gut microbes 2014, 5, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Afridi, O.K.; Ali, J.; Chang, J.H. Fecal microbiome and Resistome profiling of healthy and diseased Pakistani individuals using next-generation sequencing. Microorganisms 2021, 9, 616. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P. B.; Bik, E. M.; Bernstein, C. N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D. A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Luo, L.; Liang, W.; Yin, Q.; Guo, J.; Rush, A.M.; Lv, Z.; Liang, Q.; Fischbach, M.A.; Sonnenburg, J.L.; Dodd, D. Bifidobacterium alters the gut microbiota and modulates the functional metabolism of T regulatory cells in the context of immune checkpoint blockade. PNAS 2020, 117, 27509–27515. [Google Scholar] [CrossRef] [PubMed]
- Mitsuoka, T. Bifidobacteria and their role in human health. J. Ind. Microbiol. 1990, 6, 263–267. [Google Scholar] [CrossRef]
- Gagnon, M.; Kheadr, E.E.; Le Blay, G.; Fliss, I. In vitro inhibition of Escherichia coli O157: H7 by bifidobacterial strains of human origin. nt. J. Food Microbiol 2004, 92, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Yamasaki, S.; Nakashima, R.; Zwama, M.; Hayashi-Nishino, M. Function and inhibitory mechanisms of multidrug efflux pumps. Front. Microbiol. 2021, 12, 737288. [Google Scholar] [CrossRef] [PubMed]
- Nuonming, P.; Khemthong, S.; Dokpikul, T.; Sukchawalit, R.; Mongkolsuk, S. Characterization and regulation of AcrABR, a RND-type multidrug efflux system, in Agrobacterium tumefaciens C58. Microbiol. Res. 2018, 214, 146–155. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Global research agenda for antimicrobial resistance in human health. Policy brief. June 2023. WHO 2023c. Available online: https://cdn.who.int/media/docs/default-source/antimicrobial-resistance/amr-spc-npm/who-global-research-agenda-for-amr-in-human-health---policy-brief.pdf?sfvrsn=f86aa073_4&download=true (accessed on 4 November 2023).
- Barba, M.; Czosnek, H.; Hadidi, A. Historical perspective, development and applications of next-generation sequencing in plant virology. Viruses 2014, 6, 106–136. [Google Scholar] [CrossRef] [PubMed]
- Marchant, J. When antibiotics turn toxic. Nature 2018, 555, 431–433. [Google Scholar] [CrossRef]
- Naylor, N.R.; Pouwels, K.B.; Hope, R.; Green, N.; Henderson, K.L.; Knight, G.M.; Atun, R.; Robotham, J.V.; Deeny, S.R. The health and cost burden of antibiotic resistant and susceptible Escherichia coli bacteraemia in the English hospital setting: A national retrospective cohort study. PloS one 2019, 14, e0221944. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.; Olson, L.; Khu, D.T.; Linnros, S.; Le, N.K.; Hanberger, H.; Hoang, N.T.; Tran, D.M.; Larsson, M. Multiple antibiotic resistance as a risk factor for mortality and prolonged hospital stay: a cohort study among neonatal intensive care patients with hospital-acquired infections caused by gram-negative bacteria in Vietnam. PloS one, 2019, 14, e0215666. [Google Scholar] [CrossRef]
- Collado, M.C., Gueimonde, M. and Salminen, S. Probiotics in adhesion of pathogens: mechanisms of action. In Bioactive foods in promoting health Academic Press. 2010, 353–370.
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente,, C.; Gil, A. Probiotic mechanisms of action. Annals of Nutrition and Metabolism, 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M. and Rokana, N., 2021. Dietary management by probiotics, prebiotics and synbiotics for the prevention of antimicrobial resistance. Sustainable Agriculture Reviews 49: Mitigation of Antimicrobial Resistance Vol 2. Natural and Synthetic Approaches, pp.33-56.
- Nataraj, B.H. and Mallappa, R.H., 2021. Antibiotic resistance crisis: an update on antagonistic interactions between probiotics and methicillin-resistant staphylococcus aureus (MRSA). Current microbiology, 78(6), pp.2194-2211.
- Mazziotta, C., Tognon, M., Martini, F., Torreggiani, E. and Rotondo, J.C., 2023. Probiotics mechanism of action on immune cells and beneficial effects on human health. Cells, 12(1), p.184.
- Pot, B. and Vandenplas, Y., 2021. Factors that influence clinical efficacy of live biotherapeutic products. European Journal of Medical Research, 26(1), pp.1-10.
Figure 1.
PRISMA flow diagram indicating the screening and final selection of studies used to examine the potential influence of gut microbiota on gut/ E. coli resistome.
Figure 1.
PRISMA flow diagram indicating the screening and final selection of studies used to examine the potential influence of gut microbiota on gut/ E. coli resistome.
Table 2.
Bacteria microbiota and potential influence on gut ARGs.
| Groups | Metric | Microbiota Diversity | Taxonomic Abundance | ARGs Diversity | ARG Abundance | Significant Associations and Prediction between Microbiota and Resistome | Other Comments | Ref. |
|---|---|---|---|---|---|---|---|---|
| Full-term infants at 7 days, vs 28days, vs 120days, vs 365days |
α, Shannon Diversity |
lowest at 7 days and increases to 365 days. | Based on various time points, Bifidobacterium is highest at 28 days old > 120 > 7 > 365. Escherichia; 7 days > 120 > 28. Bacteroides: 365 days> lower time points. Klebsiella: highest in 7 days similar to 120days > 28days and least in 365days. |
NA | Higher ARGs of full-term infants at 28 days compared to 120 days. Median 7days > 28 days > 120 days. |
Escherichia coli was associated with the highest number of ARGs, followed by Klebsiella pneumoniae and Klebsiella aerogenes. |
[26] | |
| 6weeks vs 1yr | α, Shannon & Simpson | NR | NR | More even distribution of unique ARGs at 6 weeks, compared to 1-year samples, which were more dominated by one or two specific ARGs. | Higher | Positive correlation of compositional relative abundance of Proteobacteria with resistome composition Amongst the Proteobacteria, there was a strong positive correlation between E. coli relative abundance and resistome load. |
[27]. | |
| 0- 23m vs 2-5yrs | α, Shannon | Lower | Proteobacteria and Actinobacteria are > in children than adults. Bacteroides and Firmicutes A is > in adults than children. |
ARG encoding Tetracycline resistance had the greatest diversity. | Higher | NR | Although more diverse, there was a significant overlap of microbial signatures between 0-23 months and 2-5 years. |
[31] |
| 0- 23m vs > 18 | α, Shannon | lower | NR | Higher | ||||
| Infant 1month vs Mother and infant at 6month vs mother. |
α, Simpson | Lower diversity in infants than mothers. | Higher Gammaproteobacteria. Higher E. coli |
Higher | Higher in 1month infants compared to mother. Higher in 6 months infants compared to mother. |
Strong correlation between microbiome struc- ture and ARGs (r M, ≥ 0.5, p ≤ 0.001). Strong positive correlation between Gammaproteobacteria and resistome load. Strong positive correlation between E. coli and overall resistance gene load. E. coli was reliably indicated as the strongest predictor of ARGs in infants. Negative correlation between Bifidobacterium with resistance load. |
The most abundant ARGs in mothers and infants were those encoding tetracycline resistance. All resistant gene classes, except tetracycline, MLS and trimethoprim resistance, were more abundant in infants compared to mothers. |
[28] |
| NA | OTU richness For E. coli |
NR | Higher mean Relative abundance of E. coli from one week, lowering to 1yr | NR | Higher in the first year of life and lowers towards an equilibrium. | Higher in Proteobacteria. Higher in Enterobacteriaceae. Highest in E. coli |
Gut E. coli Abundance was associated with gut microbiome immaturity (High gut E. coli abundance was associated with low gut maturity and vice versa). |
[29] |
| 18-69 years olds chinese vs HMP data set |
α, Shannon | Higher | NR | NR | Higher (91% of total ARGs in were detected in Bacteriodetes, Proteobacteria and Firmicutes) |
. | Diffrence in gut microbiota were observed in samples from diffent region, location, individuals and time points. ARGs encoding tetracyclines were the most abundnant. ARGs encoding fosfomycin (9 genes) and quinolone ARGs (5 genes) were only identified in the Proteobacteria family. | [30] |
| Poultry vs. non-poultry workers |
α, Simpson | Lower | NR | Higher | Higher | LPM workers were enriched with beta-lactam and lincosamide resistance genes. Antibiotic inactivation mechanisms were higher in LPM workers. Microbiota of LPM workers was significantly different from the control group. |
[33] | |
| Dairy vs. non-dairy workers | α, Shannon | NR | No significant difference |
NR | Lower (probably due to lower sequencing depth in this group) |
Higher abundance of tetracycline and cephamycin genes in poultry workers. Evidence of commensal bacteria association with plasmid-mediated tetracycline resistance genes in both groups (including Faecalibacterium prausnitzii, Ligilactobacillus animalis, and Simiaoa sunii | [32] |
Table 3.
Microbiota composition and abundance and E. coli resistome detected in healthy individuals.
Table 3.
Microbiota composition and abundance and E. coli resistome detected in healthy individuals.
| Characteristics of the Group Considered | Microbiota Composition |
E. coli Resistome Profile | Comments | Ref. |
|---|---|---|---|---|
| 7-day old full-term infants |
Bifidobacterium breve* Bifidobacterium longum Escherichia coli Bifidobacterium bifidum Klebsiella pneumoniae Vellionella parvula Bacteroides dorei Klebsiella veriicola Bacteroides fragilis Enterococcus faecalis, Staphylococcus epidermis Klebsiella pneumoniae |
acrA acrD acrE acrF ampH bacA EC.15 emrA eptA evgA gadW gadX marA mdtE mdtF mdtG mdtH mdtO OmpA pmrF tolC yoji |
[26] | |
| 6weeks to 1year old healthy infants |
EF-Tu rpoB UhpT SoxR murA folP SoxS GlpT gyrB emrE acrR marR mdfA ompF nfSA |
The genes encode resistance to Pulvomycin Rifampicin Fosfomycin Sulfonamides Aminocoumarin Multidrug antibiotic resistance Nitrofurantoin Betalactams Resistome abundance was correlated with Proteobacteria (78.9%) and E. coli (62.2%) |
[27] | |
| 1 month and 6months olds | 1month Bifidobacterium* Escherichia Lactobacillus Bacteroides Streptococcus Staphylococcus Blautia 6months Bifidobacterium* Escherichia Blautia Bacteroides Lactobacillus Eubacterium Akkermansia Subdoligranulum |
E. coli was the highest predictor of ARG abundance | The strong correlation between the presence of E. coli and total ARG abundance in 1-month and six months olds | [28] |
| 1year – 5years |
Based on highest abundance of ARGs Escherichia coli Citrobacter werkmanii Klebsiella pneumoniae Enterobacter himalayensis Klebsiella oxytoca Citrobacter sp001037495 Enterobacter cloacae Bacteroides fragilis Bacteroides dorei Faecalibacterium prausnitzii Ruminococcus bromii Bifidobacterium longum Bifidobacterium breve Haemophilus parainfluenzae Morganella morganii Faecalicatena gnavus Tyzzerella nexilis Blautia wexlerae Ruminococcus bicirculans Flavonifractor plautii Veillonella seminalis Erysipelatoclostridium ramosum (Thomasclavelia ramosa) Agathobacter rectalis Staphylococcus epidermidis Collinsella sp003487125 Bifidobacterium pseudocatenulatum Bacteroides uniformis Bacteroides ovatus Bacteroides thetaiotaomicron Parabacteroides distasonis Alistipes putredinis Prevotella buccae |
68 out of 133 unique types of ARGs in Proteobacteria came from E. coli | Bacteria microbiota is based on the relative abundance of ARG-containing species. | [29] |
| ≤ 90 yrs |
20 most abundant Bifidobacterium adolescentis Prevotella copri Bifidobacterium longum Collinsella aerofaciens Bifidobacterium bifidum Eubacterium rectale Ruminococcus bromii Escherichia coli Bifidobacterium pseudocatenulatum Lactobacillus ruminis Faecalibacterium prausnitzii Blautia obeum Bacteroides vulgatus Bacteroides uniformis Fusicatenibacter saccharivorans Roseburia faecis Dorea longicatena Alistipes putredinis Blautia wexlerae Eubacterium hallii |
(not exclusive) mdfA emrE ampC ß-lactamase |
E. coli positively correlated with 36 ARGs. A strong association between E. coli and the Shannon resistome diversity |
[34] |
| 4-50yrs |
Prevotella* Ruminococcus Clostridaceae Bacteroides Succinovibrio Bacteroideles S24-7 Oscillospira Phascolarctobacterium Ruminobacter Desulfovibrio Helicobacter Oxalobacter formigenes |
Functional E. coli ARG detection ampC* mdfA bcr mdlB mdlA SoxS Classes Beta-lactam* ABC-transporter MFS -transporter AraC-family transcriptional regulator |
Antibiotics used for functional selection were Penicillin Piperacillin Piperacillin-tazobactam Cefotaxime Ceftazidime Cefepime Meropenem Aztreonam Chloramphenicol Tetracycline Tigecycline Gentamicin Ciprofloxacin Colistin |
[35] |
|
Non Pregnant (NP) Pregnant 1st-Trim (P1) Pregnant 3rd-Trim (P3) |
Based on 16S metagenomics Lower species diversity in pregnant compared to NP Phylum Bacteroidetes* Firmicutes Proteobacteria Actinobacteria Firmicutes* Bacteroidetes Proteobacteria Actinobacteria Firmicutes* Bacteroidetes Actinobacteria Proteobacteria |
E. coli was the most prevalent AR species E. coli isolates were resistant to Kanamycin Gentamicin Metronidazole Oxytetracycline Cycloserine Chloramphenicol Cefixime Trimethoprim/sulfamethoxazole Azithromycin Ampicillin Amoxicillin |
Majority of ARG containing species belonged to Proteobacteria in NP and Firmicutes in pregnant women. ARG rich families were Enterobacteriaceae, Enterococcaceae, and Streptococcaceae. |
[36] |
*Most abundant; AST; NP= non-pregnant.
Table 4.
Genus-level microbiota composition of the most abundant genera identified in the gut of apparently healthy participants carrying AR E. coli.
Table 4.
Genus-level microbiota composition of the most abundant genera identified in the gut of apparently healthy participants carrying AR E. coli.
| Infant Groups | Adult-Dominated Group | ||
|---|---|---|---|
| 7-Day Old | 1 Month | 6 Months | < 11years (2%) 11-20years (22%) 20-90years (76%) |
| BifidobacteriumA | BifidobacteriumA | BifidobacteriumA | BifidobacteriumA |
| EscherichiaP | EscherichiaP | EscherichiaP | PrevotellaA |
| KlebsiellaP | LactobacillusF | BlautiaBa | CollinsellaB |
| VellionellaBa | BacteroidesB | BacteroidesB | EubacteriumF |
| BacteroidesB | StreptococcusF | LactobacillusF | RuminococcusF |
| EnterococcusF | StaphylococcusF | EubacteriumP | EscherichiaP |
| StaphylococcusF | BlautiaBa | AkkermansiaV | LactobacilluF |
The superscripts indicate the bacteria Phyla: A- Actinobacteria; P-Proteobacteria; F-Firmicutes; B-Bacteroides; Ba-Bacillota; V-Verrucomicrobia.
Table 5.
Summary of the composition of gut E. coli resistome in 7 days to one-year-old infants.
| Antibiotic Class | ARGs |
|---|---|
| MDR-Efflux pump system |
acrA, acrD, acrE, acrF, acrR, mdfA ,mdtE, mdtF, mdtG, mdtH, mdtO, emrA, emrE mdfA, marA, marR, gadW, gadX, SoxS, SoxR, tolC, |
| Betalactam | ompA, ompF, ampH |
| Polypeptide | BacA, eptA, evgA, pmrF |
| Fosfomycins | murA, glpT, UhpT |
| Rifampicin | RpoB |
| Nitrofurans | nfSA |
| Aminocoumarins | GyrB |
| Beta-lactam | EC-15 |
| Folate pathway antagonists | FolP |
| Peptides | Yojl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



