
Submitted:
22 July 2024
Posted:
23 July 2024
You are already at the latest version
Abstract
As environmental and health concerns increase, the trend of sustainable agriculture moves toward using biological agents. About 60% of all biological fungicides have Trichoderma species as the active ingredient, with T. harzianum as the most common species in these products. However, the name T. harzianum has often been incorrectly used in culture collections, databases, and the scientific literature due to the division of the Harzianum Complex Clade (HCC) into more than 95 cryptic species with only one being named T. harzianum. In this study, strains previously identified as T. harzianum in three surveys of Trichoderma species from soils in South and Central America were reanalyzed using phylogeny based on tef1α, rpb2, and ITS loci and combined with the corresponding species in two other studies, where the species were correctly identified. Based on the results of the five surveys, T. afroharzianum, T. lentiforme, T. endophyticum followed by T. azedevio and T. harzianum were found to be the dominant species of the HCC in South and Central America. This is the first report of identifying dominant Trichoderma species within the HCC in South and Central American soil based on multiple studies, which will be useful in selecting strains within the complex clade for formulation of biocontrol and biofertilizer products in the continent.
Keywords:
biocontrol agent
; biofertilizer
; sustainable agriculture
; Trichoderma species
; Harzianum Complex Clade
1. Introduction
A significant number of investigations documented beneficial microbes for disease suppression and plant growth enhancement. Species in the genus Trichoderma stand out for these plant-beneficial activities. Trichoderma as a genus was introduced in 1794 by Persoon [1]. The importance of Trichoderma in agriculture, specifically as a biocontrol agent (BCA) against fungal plant diseases has been known since the 1930s [2]. Then, in the 1980s, studies showed growth promotion of various crops by application of Trichoderma species [3,4]. However, only in the 1990s did commercial products with Trichoderma as an active ingredient become commercially available with reasonable success [5]. One of the most common species in those products is Trichoderma harzianum. Samuels and Hebbar [1] assembled a list of commercial Trichoderma biocontrol products that has T. harzianum as the active ingredient in 21 out of 55 products, which was higher than any other Trichoderma species, reflecting the importance of the species for biocontrol. Also, in a compiled list of publications, Zin and Badaluddin [6] showed investigations involving the effectiveness of Trichoderma species against fungal crop pathogens. Within the list, T. harzianum was the most studied species (11 out of 18) and showed high effectiveness against various crop diseases. Taxonomically, T. harzianum was only one of the nine aggregate species described by Rifai [7]. Aggregate species, per Rifai [7], means a group of more than one species that are morphologically identical but biologically different. Taxonomy based on DNA sequencing of specific markers started in the late 1990s which resulted in exponential expansion of the number of species in the genus of Trichoderma. The species, morphologically identified as T. harzianum, appeared to split into different clades. Those clades in some cases were marked by Roman numerals or Arabic numbers [8,9], without any coordination in numbering. These studies clearly showed that T. harzianum could represent several species that are morphologically indistinguishable. Therefore, the phrase Harzianum Complex Clade (HCC) started to replace T. harzianum. The confusion continued until 2015 when Chaverri et al. [10] described 14 species within the HCC, including a few that were already described. The number of species in the HCC continued to expand [11,12,13,14,15]. Unfortunately, the split of HCC did not resolve the confusion about the name completely. There are many sequences for strains deposited in databases including GenBank as T. harzianum, even though T. harzianum is only one of the uncommon species among more than 95 described species within the complex limiting the full value of the databases [10,16]. There is another problem with the HCC species. HCC boundaries are not clearly identified, and mistakes happen when new species are included in the HCC even though they are phylogenetically positioned outside the clade. For example, Chaverri et al. [10] did not include T. tawa, T. tomentosum, and T. velutinum within the complex clade of harzianum. However, Zheng et al. [13] included all the three species within the clade. There are several surveys exploring Trichoderma in soil in different parts of the world that have reported any species in HCC as T. harzianum.
In this study, strains identified as T. harzianum in three survey studies for isolation of Trichoderma strains from the soil of South and Central America were phylogenetically re-analyzed based on the available sequencing data of three loci namely, translation elongation factor 1α (tef1α), RNA polymerase subunit II (rpb2), and the Internal Transcribed Spacers (ITS). After re-identification, the number of strains for each species was added to the numbers of respective species in another two studies for which species in the HCC were correctly identified to determine the dominant soil resident species of the HCC in the continent. Knowing the exact Trichoderma species dominance in the soil from a geographical region could help biocontrol investigations to find species that may be Indigenous, compete well in soil, and may have the ability to establish endophytic relationships with plants, resulting in the better exploitation of plant beneficial activities by the Trichoderma species. Secondly, because of the importance of the species in HCC for biocontrol and plant growth promotion, accurate identification of Trichoderma species is critical as it reflects the properties associated with individual species.
2. Materials and Methods
2.1. Re-Identification of the Trichoderma Harzianum Strains Deposited in the GenBank
To evaluate the accuracy of identification of Trichoderma harzianum strains deposited at the National Center for Biotechnology Information (NCBI) GenBank (https://www.ncbi.nlm.nih.gov/genbank/) a search was carried out for “Trichoderma harzianum translation elongation factor” in the GenBank. The initial 100 sequence hits were downloaded in the FASTA file format. Sequences of sixteen ex-type species in HCC that were retrieved from GenBank were then added to this file as references. The FASTA file was aligned using Clustal Omega. (https://www.ebi.ac.uk/jdispatcher/msa/clustalo). The alignment file was adjusted manually using the software Mesquite version 3.81 [17]. Then after, the file was used to construct phylogenetic trees using two methods: 1) parsimony tree was obtained using PAUP version 4.0a (https://phylosolutions.com/paup-test/). The tree was produced using a heuristic search with a starting tree obtained by 1000 random stepwise addition of sequences, tree-bisection-reconnection (TBR) as the branch-swapping algorithm with MULTREES in effect. Gaps were treated as missing characters. Supports for branches were assessed with 1000 replicates of bootstrap. 2) Maximum Likelihood tree was obtained using MEGA X with the substitution model predetermined using MEGA X [18]. Support for the clades was assessed with 1000 bootstrap replicates.
2.2. Evaluation of the Dominant HCC Species from South and Central America
To determine the dominant HCC species in soils in South and Central America, strains identified based on translation elongation factor 1α (tef1α) in the studies of Hoyos-Carvajal et al., Smith et al., and Druzhinina et al. [19,20,21] as T. harzianum were re-analyzed phylogenetically based on the DNA sequencing data of three loci, tef1α, RNA polymerase subunit II (rpb2), and Internal Transcribed Spacers (ITS), respectively. The sequences for each locus were downloaded from the GenBank, and aligned with reference sequences particularly the ex-type species of known species in the clade described in Chaverri et al. [10] and del Carmen et al. [12] using the Clustal Omega (https://www.ebi.ac.uk/jdispatcher/msa/clustalo). All the strains used in the phylogenetic analysis are listed in Table 1. The alignment files for the three genes were concatenated and adjusted visually using the software Mesquite version 3.81 [17]. The alignment file was used to construct phylogenetic trees as described above. The trees obtained by both methods were essentially identical in topology and thus only the parsimony tree constructed by PAUP is presented.
After phylogenetic identification of the strains, the number of each species in the HCC from the three studies was added to respective species in two other studies by Inglis et al. and Barrera et al. [22,23] that had correctly identified species in the HCC. The results were tabulated to determine the most encountered species within the HCC in soils of South and Central America.
3. Results
3.1. Re-identification of the Trichoderma harzianum strains deposited in the GenBank
Initially, we searched GenBank for “Trichoderma harzianum translation elongation factor” and phylogenetically analyzed the first 100 sequences with the ex-type of T. harzianum strain CBS 226.95, GenBank accession number (AF348101). Based on the phylogenetic tree (Figure 1) only 22 out of 100 sequences clustered with the ex-type of T. harzianum strain CBS 226.95, GenBank accession number (AF348101); so, the respective 22 strains can be validated as T. harzianum. It also indicates that 78% of the sequences that were deposited had previously been erroneously identified.
3.2. Evaluation of the dominant HCC species from South and Central America
In the three survey studies by Hoyos-Carvajal et al., Smith et al., and Druzhinina et al. [19,20,21] for isolation of Trichoderma species in the soil of Central and South America, all the strains in the Harzianum complex clade (HCC) were reported as T. harzianum, which does not reflect the current taxonomy of the genus that subdivided the clade into more than 95 species [10,16]. Precise species names are critical as some but not all the species in HCC may be associated with biocontrol and biofertilizer activities. Re-analyses of sequencing data of strains identified as T. harzianum in the three survey studies (Figure 2, Table 1) showed that 25 strains clustered with the type species of T. lentiforme in Clade 1 with high bootstrap values. Therefore, these strains are identified as T. lentiforme. Two other reference sequences in Clade 1 were also used. The reference strain with GenBank accession number MK896667 was placed in the tree to confirm the identity of the species in the study by Inglis, et al. [22], and the reference strain with MK044089 was placed in the tree to show that T. lentiforme was obtained in regions other than South America as this accession number belongs to a strain from Africa.
In Clade 2, Figure 2, 10 strains from the three studies formed a highly supported clade with the type strain of T. afroharzianum. BLAST search for each isolate also confirmed that the most homologous sequences to the strains in this clade were sequences of T. afroharzianum. As a reference sequence, a sequence of the biocontrol strain known as T22 was aligned in this clade. T22 was re-identified as T. afroharzianum [10]. Also, the sequence of strain CEN414 from the study of Inglis et al. [22] fits in this clade, confirming the correct identification of this strain as T. afroharzianum.
In clade 3, Figure 2, one isolate from Peru [19], CIB T52 formed a clade with two species, T. pyramidale and T. pseudopyramidale. However, the CIB T52 stain is closer to T. pseudopyramidale than to T. pyramidale. The latter two species are closely related but one has been found only in Africa and the other in South America, respectively.
In clade 4, Figure 2, 6 strains formed a clade with the type species of T. endophyticum. However, the sequences of type species T. neotropicale and T. afarasin also fit into the clade. The type species T. neotropicle seems to be distantly related to the clade through a long branch, and T. afarasin is thus far only known to be an African fungus not found outside that continent. Therefore, it is most likely that the six isolates in the clade were T. endophytcum.
In Clade 5, Figure 2, two strains CIB T100 and TUB-F1078 from two different studies clustered with the type strain of T. harzianum CBS 226.95 with high bootstrap values by both methods, clearly identifying them as T. harzianum.
In clade 6, Figure 2, seven strains were nested with the ex-type strain of T. azevedoi (CEN1422 from Brazil); in most cases the sequences were identical. We also did a correction to the naming of a species. In the study of Barrera et al. [23], five strains from Argentina were identified as T. austroindianum. Four of the five strains have identical tef1α sequences. The other strain has 2 base differences with the other four isolates. T. austroindianum had sequences identical or highly homologous to T. azevedoi strains and fall in clade 6 and thus, we believe that the two species represent only one species. Thus, the strains of T. austroindianum were tabulated with the T. azevedoi as, on a priority basis, the latter species was described before T. austoindianum [24].
In clade 7, Figure 2, one isolate from Colombia CIB T52 formed a clade with the type species of T. rifaii and another reference strain of T. rifaii, suggesting the correct identification of the strain as T. rifaii.
The strain DAOM 234005 did not fit into any known clade and represents a distinct lineage distantly related to other species of the Harzianum complex. Moreover, based on BLAST search, the Tef1α sequence of this strain had no close homologous sequence to it in the GenBank; thus, we consider it a possible new species in the HCC. As to T. pedryi, this species was described as a new species in the HCC [22]. However, the position of this species in the phylogenetic tree (Figure 2) showed that this species is close to T. tomentosum, which is not included in the HCC [10]. Thus, we did not include this species in the tabulations of HCC species in Table 2.
The results of our re-identification of species of HCC were combined with the previously identified species in two other studies [22,23] and presented in Table 2 and summarized in Figure 3. T. afroharzianum is the most common species in Central and South America with 44 stains found in four out of five studies and in countries from North to South of the continent. This is a well-known biocontrol species and the strain T22 is an example [10,25].
T. lentiforme was the second most common species with 39 strains from four studies out of five. The strains were distributed in all the regions from North to South of the continent. Chaverri et al. [10] reported that this species was found only in South America and mainly as an endophyte. However, as the tree (Figure 2) and Table 2 show, this species is commonly isolated from soil and the strain E243 (accession MK044182) was also found as an endophyte in coffee plants in Cameroon, Africa.
The third most common encounter in soils of Central and South America is T. endophyticum with 18 stains that was found in four out of five studies. This species was reported to be found as endophytes of tropical trees [10]. However, this study showed that the species is a common encounter in soil as well. The fourth highest encounter with 15 strains is T. azevedoi which was first described in 2020 by Inglis et al [22]. However, based on BLAST search of Tef1α sequence of type species of T. azevedoi (accession # MK696660, strain CEN 1422) we found several sequences that are highly homologous and have been deposited under the same species name from the soil in Australia (e.g. strain BRIP 74284; accession number OR801290). T. harzianum is the fifth in order of most common species in Central and South America, with 8 strains found in three out of five studies. The strains were found in North Mexico, Colombia, and South Argentina. Even though this species is mainly found in north temperate regions, Europe, and North America [1], the results here show that it is also found in Central and South America. T. guizhouense with 5 strains made the 6th most common species in the HCC in Central and South America. However, the species was observed only in one out of five studies and found only in Argentina. This may indicate a biogeographic restriction of the species in the continent of South and Central America. Other species in order were T. neotrpicale (3 stains), T. syagri (2 strains), T. Hortense (1 strain), T. pseudopyramidale (1 strain) and one isolate remained unidentified and placed in the table as Trichoderma sp.
4. Discussion
Biocontrol programs are in place in South and Central America, notably in nations like Brazil, Argentina, Colombia, and Mexico, to battle plant diseases and encourage crop development with biological agents [26]. One of the most important fungal names in biocontrol products of plant diseases and / or plant health promotion are species in the genus of Trichoderma and in particular, the species name of T. harzianum [1,10,27]. However, confusion exists about the name T. harzianum being used for all the species within Harzianum Complex Clade (HCC) despite the revision of the phylogeny of the clade and description of more than 95 species that started in 2015 [10,16]. In fact, the problem is widespread and continuous in scientific publications, databases, and commercial platforms. As an example, after searching the GenBank for “Trichoderma harzianum translation elongation factor” and identifying the first 100 sequences (this study), it turned out that only 22 out of 100 clustered with the ex-type of T. harzianum strain CBS 226.95 GenBank accession number (AF348101) and could be identified as T. harzianum, indicating 78% misidentification in the name of strains deposited as T. harzianum. What is disturbing about this is the fact that 100 % of the sequences used in Figure 1 were deposited between 2018-2023, at least three years after the major revision of the HCC [10]. Precise naming of species of HCC is critical as beneficial properties of biocontrol and plant growth promotion are species specific or even strain specific. Due to improper usage of the name, there is a lack of studies exploring which species in the clade are dominant species in soil despite the HCC species being isolated in many survey investigations. We hypothesize that the dominance in the soil is an important criterion for selecting any biocontrol strain as it reflects the fungus’s high ability to compete, outgrow, and suppress other species, leading to a higher potential for the fungus to establish endophytic relationship with plants and possibly induce systemic resistance. Through inducing systemic resistance, the Trichoderma species provides other benefits to the plants such as resistance to abiotic stress and high efficiency in using nitrogen [5].
This study used data from five investigations to identify dominant Trichoderma species within the HCC in the soil of South and Central America. To determine the correct species name and identify the prevalent species in soils of Central and South America, in this study we re-analyzed with multi-locus phylogeny the data from three survey studies published in 2005-2013 [19,20,21]. The number of re-identified species was added to their respective species from two other studies where HCC species were identified correctly [22,23]. T. afroharzianum and T. lentiforme, respectively, were the top two dominant species in the continent and were obtained from north to south. T. afroharzianum is cosmopolitan and a well-known species of biocontrol agents, and the strain T22 is reported as the most known BCA [28]. T. lentiforme was reported [10] to be mainly an endophytic fungus. The dominance of T. lentiforme in soil is a new report of our findings. Having this ability in soil and the ability to establish endophytic relations with plants are top criteria for selection with biocontrol and biofertilizer properties. In a recent publication [29] T. lentifore was found to have biocontrol activity against Sclerotinia sclerotiorum and had growth stimulant property for cotton.
T. endophyticum and T. azevedoi are the third and fourth most common species, respectively, on the continent. T. endophyticum was shown to be exclusively endophytic based on the strains available for the study [10]. However, this study also showed that this species is a soil fungus as well (Table 2). T. azevedoi was described in the study of Inglis et al. [22] and has been obtained only in the South American continent. In this context, we corrected the name of a species described by Barrera et al. [23] as T. austroindianum. Five strains of T. austroindianum appear to have identical or highly homologous tef1α sequences to those of T. azevedoi and both clustered in one highly supported clade. Thus, we renamed all five strains of T. austroindianum as T. azevedoi in Table 1. Here we stress the importance of (1) BLAST search of the sequences of tef1α or rpb2 loci of unknown Trichoderma strains before describing them as new species to avoid duplication of species naming and (2) to include the most homologous species to it in the phylogenic analyses. The phylogenetic trees in Barrera et al. [23] did not include T. azevedoi in the analyses, which is probably the reason that the authors overlooked this error.
The boundaries of the species within the HCC are equally unclear. For example, Chaverri et al. [10] did not include T. tawa, T. tomentosum, and T. velutinum within the complex clade of Harzianum. However, Zheng et al. [13] included all the above three species within the clade. In another example, Chaverri et al. [10] excluded T. amazonicum and T. pleuroticola from HCC. Yet, Chen and Zhuang [30] placed both the species inside the HCC. In this regard, Inglis et al. [22] described a new species named T. peberdyi as part of the HCC. Based on BLAST search at NCBI GenBank, and our phylogenetic tree (Figure 2), this species is closely related to T. tomentosum. Based on Chaverri et al. [10], T. tomentosum is not part of the clade. Therefore, T. perbedyi is not part of the HCC clade and was excluded from our list of HCC species (Table 2, Figure 3).
We also attempted to compare the dominant species in Central and South America with other continents. However, there is currently insufficient data from survey studies that quantitatively show the number of strains of HCC within the total number of strains of Trichoderma isolated. Thus, comparing South and Central America with other continents was not feasible. Nevertheless, we found individual reports to which we could compare our results. For example, the study of a survey of Trichoderma isolates from the soil of India [31] showed that among 15 strains in HCC that were identified, 11 were T. afroharzianum. This data agrees with the high encounter of T. afroharzianum in the soil in South and Central America. On the other hand, a survey study from Iran showed that T. afroharzianum was not a dominant HCC species from soil in the western region of the country [32]. In fact, T. harzianum was the most frequently isolated species. In another study, Tang et al. [33] showed that species of HCC were the most encountered Trichoderma species from soil in China in the region of Zoige Alpine. Among species of the HCC, T. harzianum was the most dominant species representing about 72% of the species in HCC (37/51). This is different from what we obtained in South and Central America. T. harzianum is known to be a species of cold temperature region and the temperature range of soil in the region where samples were obtained is -10 C in winter to 15 C in summer [34]. This may have caused a bias in species dominance. In South Africa, du Plessis et al. [35] showed that species of T. afroharzianum, T. atrobrunneum and T. camerunense were the only species within HCC obtained from the soil in South Africa; however, the authors did not report the quantitative data of the respective strains.
5. Conclusions
Within the genus Trichoderma, T. harzianum is one of the most well-known species for biocontrol and plant growth promotion. The Harzianum Complex Clade’s (HCC) split into more than 95 species has led to the improper usage of this species’ name in scientific publications, databases, and commercial platforms. This study used data from five investigative reports to identify dominant Trichoderma species within the HCC in the soil of South and Central America. Based on the multilocus (tef1α, rpb2, and ITS) phylogenetic analyses of the five published surveys, T. afroharzianum, T. lentiforme, T. endophyticum, T. azedevio, and T. harzianum were found to be the dominant species of the HCC in South and Central America. However, the soil sampling in those five studies was not obtained in a statistical manner and order. Thus, we consider that the findings from this study are a rough approximation of the dominating species of HCC in Central and South America. Currently, the HCC is known to have roughly 95 species. Selecting strains or species for biocontrol and growth promotion from the HCC could be an expensive and difficult undertaking. Identifying the dominant strains can expedite the commercialization process by reducing the time and expense associated with strain selection. We further emphasize the need for additional research to be done in the future from different continents, as we were unable to locate any comparable data with which to compare our findings. Moreover, we emphasize the significance of accurately identifying Trichoderma species prior to entering them into the HCC database.
Author Contributions
A.I. conceptualization, data generation, analysis, interpretation, writing—original draft preparation and editing; D.K.L. conceptualization, analysis, interpretation, writing, and editing; P.P.J. data collection and analyses, and P.S. feedback, and formatting of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by USDA-ARS in-house projects 8042-21220-259-000D and 8042-22000-320-000D.
Data Availability Statement
Data is contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Samuels, G.J.; Hebbar, P.K. Trichoderma: Identification and Agricultural Applications; APS Press, 2015.
- Weindling, R. Trichoderma lignorum as a Parasite of Other Soil Fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Lindsey, D.L.; Baker, R. Effect of Certain Fungi on Dwarf Tomatoes Grown under Gnotobiotic Conditions. Phytopathology 1967, 57, 1262–1263. [Google Scholar]
- Chang, Y.-C.; Chang, Y.-C.; Baker, R.; Kleifeld, O.; Chet, I. Increased Growth of Plants in the Presence of the Biological Control Agent Trichoderma harzianum. Plant Dis. 1986, 70, 145–148. [Google Scholar] [CrossRef]
- Harman, G.E. Trichoderma—Not Just for Biocontrol Anymore. Phytoparasitica 2011, 39, 103–108. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological Functions of Trichoderma Spp. for Agriculture Applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Rifai, M.A. A Revision of the Genus Trichoderma. Mycol Pap. 1969, 116, 1–56. [Google Scholar]
- Chaverri, P.; Castlebury, L.A.; Samuels, G.J.; Geiser, D.M. Multilocus Phylogenetic Structure within the Trichoderma harzianum/Hypocrea lixii Complex. Mol. Phylogenet. Evol. 2003, 27, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Kubicek, C.P.; Bissett, J.; Druzhinina, I.; Kullnig-Gradinger, C.; Szakacs, G. Genetic and Metabolic Diversity of Trichoderma: A Case Study on South-East Asian Isolates. Fungal Genet. Biol. 2003, 38, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum Species Complex and the Re-Identification of Commercial Biocontrol Strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Wang, R.; Sun, Q.; Wu, B.; Sun, J.-Z. Four New Species of Trichoderma in the Harzianum Clade from Northern China. MycoKeys 2020, 73, 109–132. [Google Scholar] [CrossRef]
- del Carmen, H. Rodríguez, M.; Evans, H.C.; de Abreu, L.M.; de Macedo, D.M.; Ndacnou, M.K.; Bekele, K.B.; Barreto, R.W. New Species and Records of Trichoderma Isolated as Mycoparasites and Endophytes from Cultivated and Wild Coffee in Africa. Sci. Rep. 2021, 11, 5671. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Qiao, M.; Lv, Y.; Du, X.; Zhang, K.-Q.; Yu, Z. New Species of Trichoderma Isolated as Endophytes and Saprobes from Southwest China. J. Fungi 2021, 7, 467. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.-T.; Zhuang, W.-Y. Seven New Species of Trichoderma Hypocreales in the Harzianum and Strictipile Clades. Phytotax 2017, 305, 121–139. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Zhuang, W.-Y. New Species of Trichoderma in the Harzianum, Longibrachiatum and Viride Clades. Phytotaxa 2018, 379, 131–142. [Google Scholar] [CrossRef]
- Cao, Z.-J.; Qin, W.-T.; Zhao, J.; Liu, Y.; Wang, S.-X.; Zheng, S.-Y. Three New Trichoderma Species in Harzianum Clade Associated with the Contaminated Substrates of Edible Fungi. J. Fungi 2022, 8, 1154. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P. Mesquite: A Modular System for Evolutionary Analysis. Version 3.18. 2007. http://www.mesquiteproject.
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Genetic and Metabolic Biodiversity of Trichoderma from Colombia and Adjacent Neotropic Regions. Fungal Genet. Biol. 2009, 46, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Beltrán, C.A.; Kusunoki, M.; Cotes, A.M.; Motohashi, K.; Kondo, T.; Deguchi, M. Diversity of Soil-Dwelling Trichoderma in Colombia and Their Potential as Biocontrol Agents against the Phytopathogenic Fungus Sclerotinia sclerotiorum (Lib.) de Bary. J. Gen. Plant Pathol. 2013, 79, 74–85. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Kopchinskiy, A.G.; Komoń, M.; Bissett, J.; Szakacs, G.; Kubicek, C.P. An Oligonucleotide Barcode for Species Identification in Trichoderma and Hypocrea. Fungal Genet. Biol. 2005, 42, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Inglis, P.W.; Mello, S.C.; Martins, I.; Silva, J.B.; Macêdo, K.; Sifuentes, D.N.; Valadares-Inglis, M.C. Trichoderma from Brazilian Garlic and Onion Crop Soils and Description of Two New Species: Trichoderma azevedoi and Trichoderma peberdyi. PloS One 2020, 15, e0228485. [Google Scholar] [CrossRef] [PubMed]
- Barrera, V.A.; Iannone, L.; Romero, A.I.; Chaverri, P. Expanding the Trichoderma harzianum Species Complex: Three New Species from Argentine Natural and Cultivated Ecosystems. Mycologia 2021, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Taylor, J.W.; Wingfield, M.J.; Abaci, Ö.; Aime, C.; et al. The Amsterdam Declaration on Fungal Nomenclature. IMA Fungus 2011, 2, 105–111. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Steindorff, A.S.; Chenthamara, K.; Manganiello, G.; Henrissat, B.; Zhang, J.; Cai, F.; Kopchinskiy, A.G.; Kubicek, E.M.; Kuo, A.; et al. Evolution and Comparative Genomics of the Most Common Trichoderma species. BMC Genomics 2019, 20, 485. [Google Scholar] [CrossRef] [PubMed]
- van Lenteren, J. C.; Bueno, V. H. P.; Luna, M. G. Biological control in Latin America and the Caribbean: information sources, organizations, types and approaches in biological control. In J. C. V. Lenteren, V. H. P. Bueno, M. G. Luna, & Y. C. Colmeneraz (Eds.), Biological control in Latin America and the Caribbean: its rich history and bright future. (pp. 1-20). CABI 2020. [CrossRef]
- Xiao, Z.; Zhao, Q.; Li, W.; Gao, L.; Liu, G. Strain Improvement of Trichoderma harzianum for Enhanced Biocontrol Capacity: Strategies and Prospects. Front. Microbiol. 2023, 14, 1146210. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Wu, Z.; Zhong, Z.; Williams, J.; Jacobsen, S.E.; Sun, Z.; Tang, Y. Assessing the Biosynthetic Inventory of the Biocontrol Fungus Trichoderma afroharzianum T22. J. Agric. Food Chem. 2023, 71, 11502–11519. [Google Scholar] [CrossRef]
- Silva, L.G.; Camargo, R.C.; Mascarin, G.M.; Nunes, P.S. de O.; Dunlap, C.; Bettiol, W. Dual Functionality of Trichoderma: Biocontrol of Sclerotinia sclerotiorum and Biostimulant of Cotton Plants. Front. Plant Sci. 2022, 13, 983127. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhuang, W.-Y. Discovery from a Large-Scaled Survey of Trichoderma in Soil of China. Sci. Rep. 2017, 7, 9090. [Google Scholar] [CrossRef] [PubMed]
- Jambhulkar, P.P. , Singh, B., Raja, M. et al. Genetic diversity and antagonistic properties of Trichoderma strains from the crop rhizospheres in southern Rajasthan, India. Sci Rep 2024, 14, 8610. [Google Scholar] [CrossRef]
- Mirzaeipour, Z.; Bazgir, E.; Zafari, D.; Darvishnia, M. Isolation and identification of Harzianum clade species of Trichoderma from Khorramabad County. Mycologia Iranica 2023, 10, 67–78. [Google Scholar] [CrossRef]
- Tang, G.T.; Li, Y; Zhou, Y; Zhou, Y.; Zhu, Y.H.; Zheng, X.J.; Chang, X.L.; Zhang, S. R.; Gong, G.S. Diversity of Trichoderma species associated with soil in the Zoige alpine wetland of Southwest China. Sci. Rep. 2022, 12, 21709. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Lu, Q; Zhao, Q.; Wang, J.; Ouyang, H. Effects of alpine wetland landscapes on regional climate on the Zoige Plateau of China. Advances in Meteorology. 2013, 972430. 7pp. [CrossRef]
- du Plessis, I.L.; Druzhinina, I.S; Atanasova, L.; Yarden O.; Jacobs, K. The diversity of Trichoderma species from soil in South Africa, with five new additions, Mycologia, 2018. [CrossRef]
Figure 1.
Phylogenetic tree based on Tef1α sequence data for 100 strains retrieved from GenBank which were earlier deposited as Trichoderma harzianum. The leaves are identified by GenBank accession numbers. * Given above the branches indicate bootstrap values of 70% obtained via 1000 replications. The boldface indicates reference strains for ex-type species included to identify the clades. Strains in pink highlighted clade are correctly deposited in GenBank as T. harzianum.
Figure 1.
Phylogenetic tree based on Tef1α sequence data for 100 strains retrieved from GenBank which were earlier deposited as Trichoderma harzianum. The leaves are identified by GenBank accession numbers. * Given above the branches indicate bootstrap values of 70% obtained via 1000 replications. The boldface indicates reference strains for ex-type species included to identify the clades. Strains in pink highlighted clade are correctly deposited in GenBank as T. harzianum.
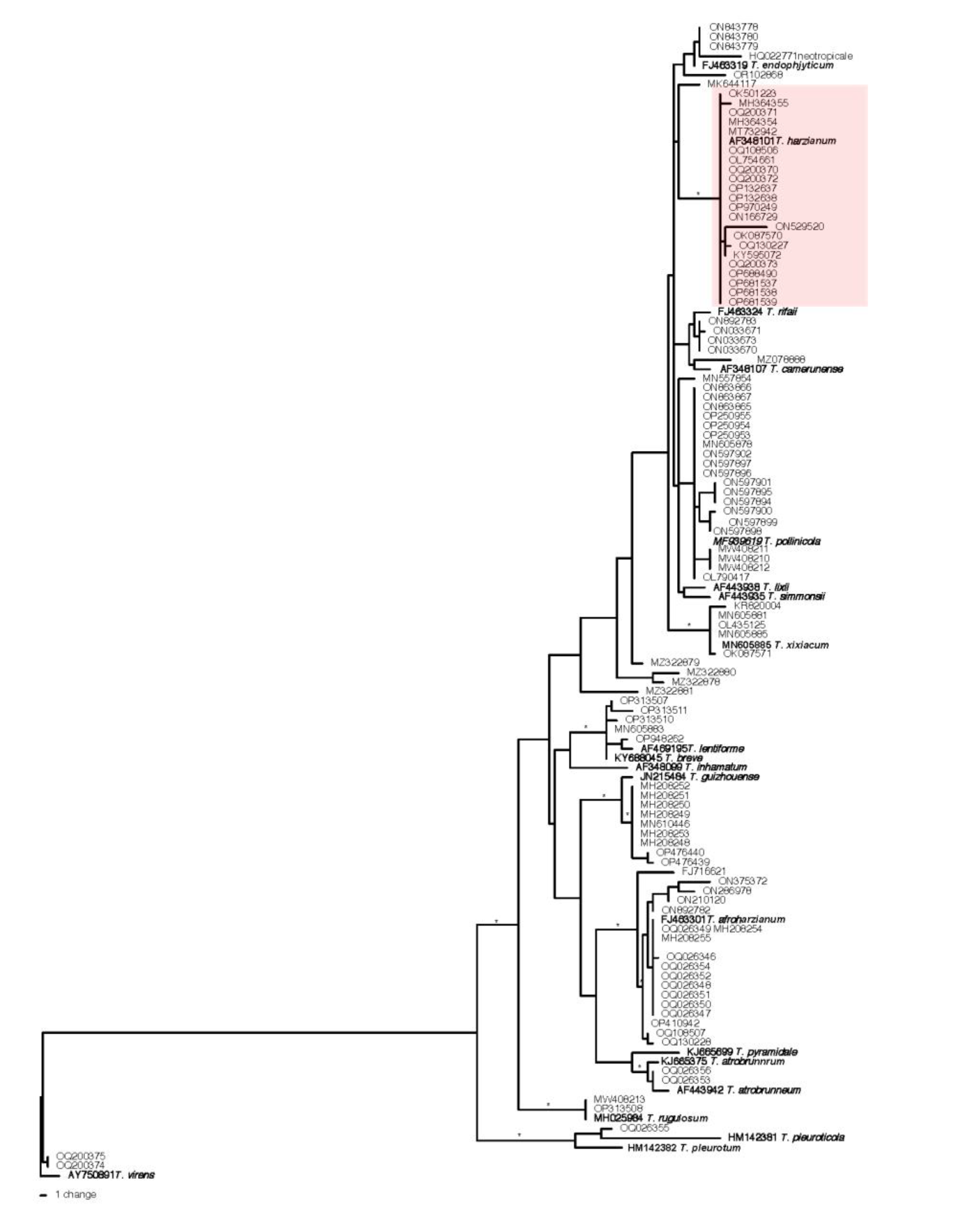
Figure 2.
One of the most parsimonious trees generated by phylogenetic analysis of combined DNA sequences of Tef1α, rpb2, and ITS. * Given above the branches indicate bootstrap values of 70% obtained via 1000 replications. Leaves are identified by Tef1α GenBank accession numbers followed by strain numbers. The boldface indicates reference strains included to identify the clades. Vertical lines with numbers C1-C7 identify important clades with bootstrap values of 70. * After species names indicate the ex-type species.
Figure 2.
One of the most parsimonious trees generated by phylogenetic analysis of combined DNA sequences of Tef1α, rpb2, and ITS. * Given above the branches indicate bootstrap values of 70% obtained via 1000 replications. Leaves are identified by Tef1α GenBank accession numbers followed by strain numbers. The boldface indicates reference strains included to identify the clades. Vertical lines with numbers C1-C7 identify important clades with bootstrap values of 70. * After species names indicate the ex-type species.
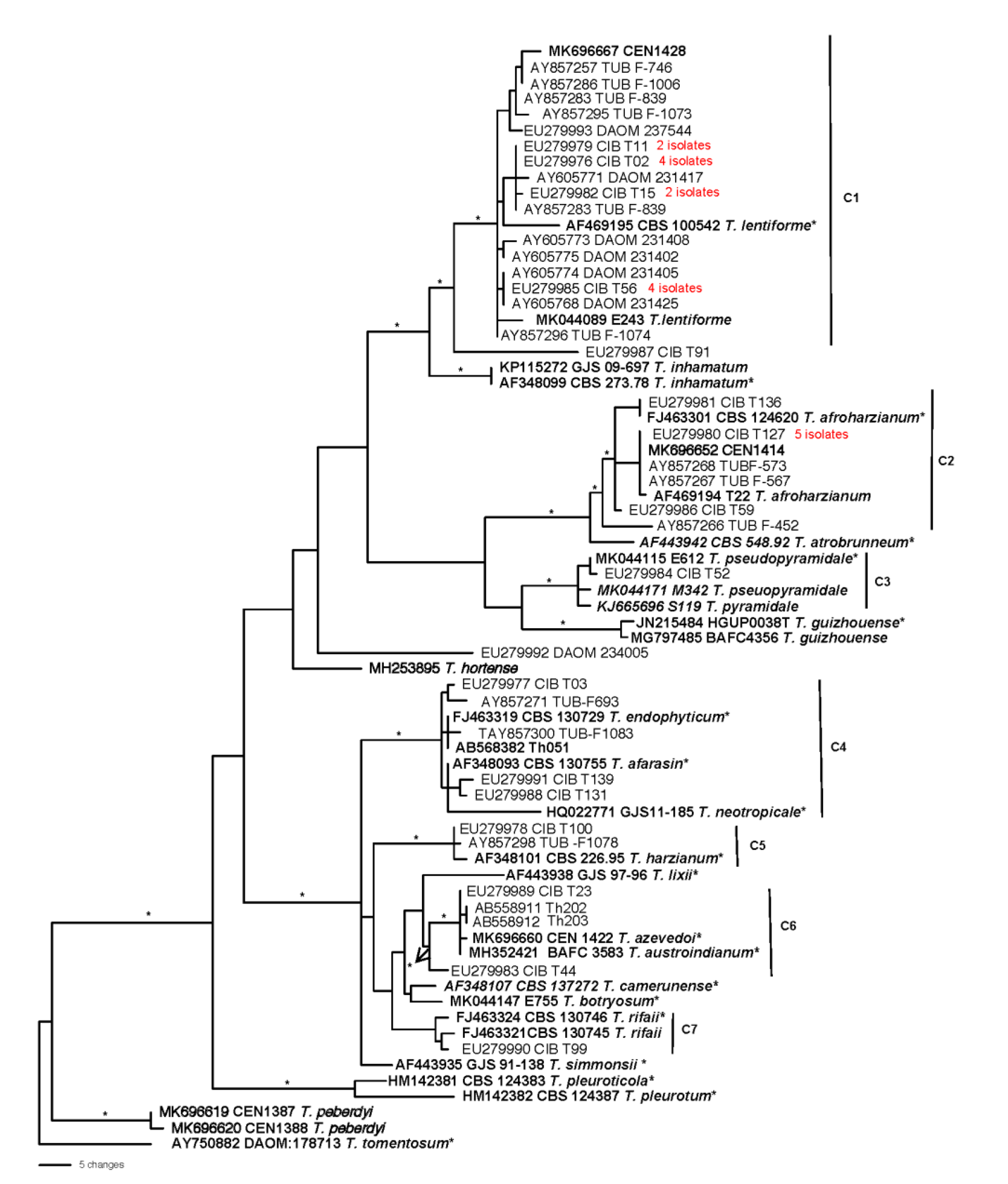
Figure 3.
Dominant species of Trichoderma in the Harazianum Complex Clade in soil in Central and South America. The numbers at the end of the bars represent the frequency of presence of a species in five studies.
Figure 3.
Dominant species of Trichoderma in the Harazianum Complex Clade in soil in Central and South America. The numbers at the end of the bars represent the frequency of presence of a species in five studies.
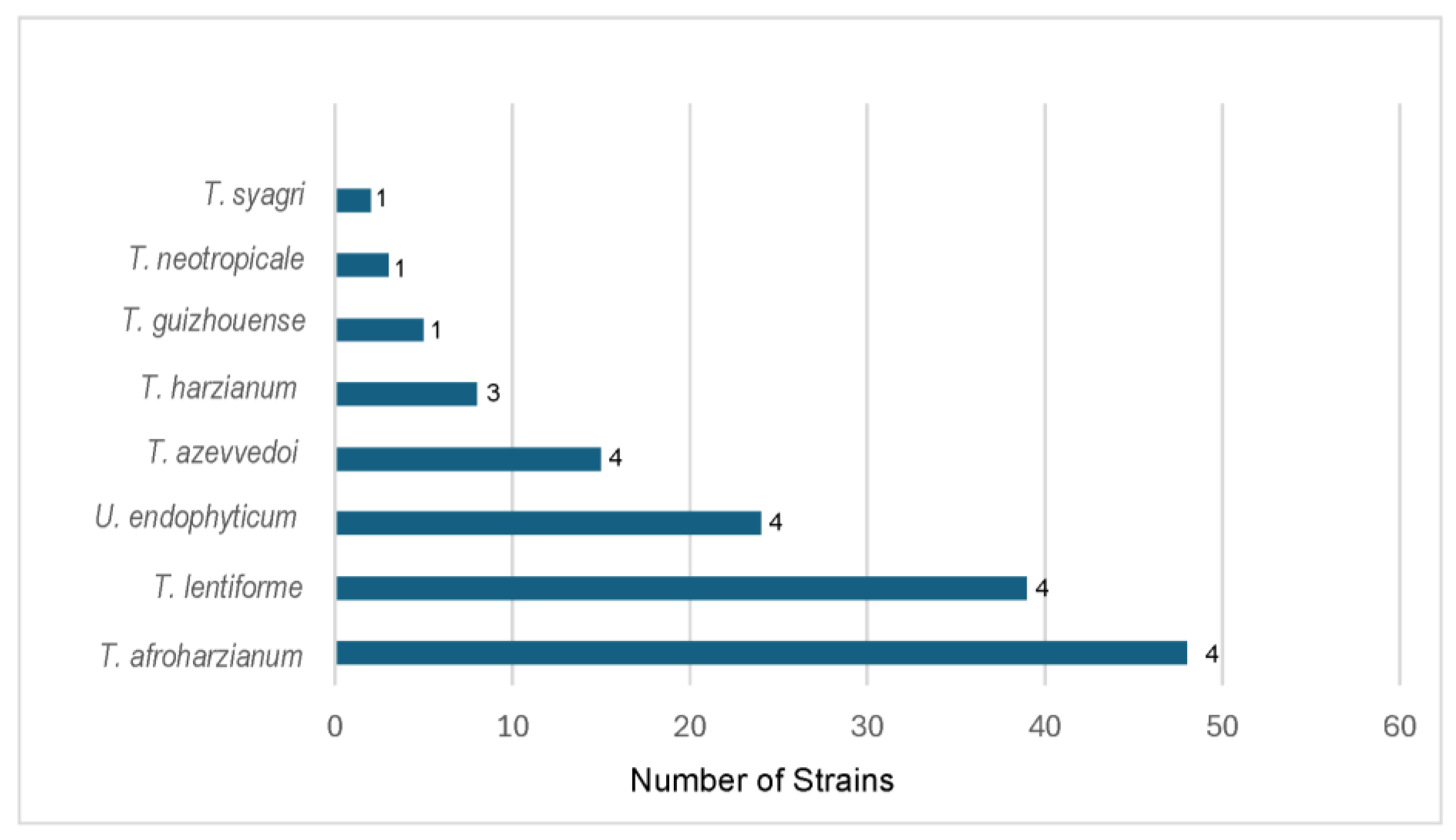

Table 1.
Strains with their origin, strain number, and their sequences GenBank accession number and number of strains with identical Tef1α, ITS and rpb2 GenBank accession numbers.
Table 1.
Strains with their origin, strain number, and their sequences GenBank accession number and number of strains with identical Tef1α, ITS and rpb2 GenBank accession numbers.
| Trichoderma species | origin | Strain # | Tef1α | ITS | rpb2 | No. of strains |
|---|---|---|---|---|---|---|
| T. lentiformea | Brazil | CEN1428 | MK666796 | MK714909 | MK696827 | |
| Brazil | TUB F-746 | AY857257 | AY857216 | X | ||
| Colombia | CIB T11 | EU279976 | EU280079 | X | 2b | |
| Colombia | CIB T02 | EU279976 | EU280079 | X | 4 | |
| Mexico | DAOM 231417 | AY605771 | AY605728 | X | ||
| Colombia | CIB T15 | EU279982 | EU280079 | X | 2 | |
| Mexico | DAOM 231408 | AY605773 | AY605730 | X | ||
| Mexico | DAOM 231402 | AY605775 | AY605732 | X | ||
| Mexico | DAOM 231439 | EU279994 | AY605728 | X | ||
| Colombia | CIB T91 | EU279987 | EU280079 | X | ||
| Mexico | DAOM 231405 | AY605774 | AY605731 | X | ||
| Colombia | CIB T56 | EU279985 | EU280079 | X | 4 | |
| Peru | DAOM 237544 | EU279993 | EU280133 | X | ||
| Mexico | DAOM 231425 | AY605768 | AY605725 | X | ||
| T. lentiforme | Cameroon | E243 | MK044089 | X | MK044182 | |
| Mexico | TUB F-839 | AY857283 | AY857231 | X | ||
| Brazil | TUB F-1073 | AY857295 | AY857247 | X | ||
| Brazil | TUB F-1006 | AY857286 | AY857235 | X | ||
| T. lentiforme* | French Guiana | CBS 100542 | AF469195 | AF469189 | X | |
| Argentina | TUB F-1074 | AY857296 | AY857248 | |||
| T. inhamatum | Peru | G.J.S. 09-697 | KP115272 | X | X | |
| T. inhamatum* | Colombia | CBS 273.78 | AF348099 | FJ442680 | FJ442725 | |
| Peru | DAOM 234005 | EU279992 | EU280091 | X | ||
| T. hortense* | Argentina | BAFC_cult_4291 | MH253895 | X | X | |
| Colombia | CIB T136 | EU279981 | EU280078 | X | ||
| Colombia | CIB T127 | EU279980 | EU280078 | X | 5 | |
| T. afroharzianum | Colombia | T22 | AF469194 | AF469188 | X | |
| T. afroharzianum | Brazil | CEN1414 | MK696652 | MK714894 | MK696813 | |
| T. afroharzianum* | Peru | CBS 124620 | FJ463301 | FJ442265 | FJ442691 | |
| Colombia | CIB T59 | EU279986 | EU280078 | |||
| T. pseudopyramidale* | COAD 2420 | MK044115 | X | MK044208 | ||
| T. pseudopyramidale | COAD 2439 | MK044171 | X | MK044264 | ||
| T. pyramidale | S119 | KJ665696 | X | X | ||
| T. atrobrunneum* | France | CBS 548.92 | AF443942 | AF443924 | X | |
| Peru | CIB T52 | EU279984 | EU280077 | |||
| T. guizhouense* | China | CBS 131803 | JN215484 | JN191311 | JQ901400 | |
| T. guizhouense | Argentina | BAFC 4356 | MG797485 | X | X | |
| Peru | TUB F-567 | AY857267 | AY857208 | X | ||
| Peru | TUB F-452 | AY857266 | AY857206 | X | ||
| Peru | TUB F-573 | AY857268 | AY857209 | X | ||
| Colombia | CIB T03 | EU279977 | EU280079 | X | ||
| T. endophyticum* | Ecuador | CBS 130729 | FJ463319 | FJ442243 | X | |
| T. afarasin* | Cameroon | CBS 130755 | AF348093 | AY027784 | X | |
| Guatemala | TUB F-693 | AY857271 | AY857211 | X | ||
| Mexico | TUB F-1083 | AY857300 | AY857253 | X | ||
| Colombia | CIB T139 | EU279991 | EU280075 | X | ||
| Colombia | Th051 | AB568382 | X | AB568476 | ||
| Colombia | CIB T131 | EU279988 | EU280075 | X | ||
| Colombia | CIB T100 | EU279978 | EU280079 | X | ||
| T. harzianum* | U.K. | CBS 226.95 | AF348101 | AJ222720 | AF545549 | |
| Mexico | TUB F-1078 | AY857298 | AY857250 | X | ||
| T. lixii** | Thailand | CBS 110080 | AF443938 | AF443920 | X | |
| T. camerunense* | Cameroon | CBS 137272 | AF348107 | AY027780 | X | |
| T. botryosum | Ethiopia | COAD 2526 | MK044147 | X | MK044240 | |
| Colombia | CIB T23 | EU279989 | EU280077 | X | ||
| Colombia | Th202 | AB558911 | X | AB558921 | ||
| Colombia | Th203 | AB558912 | X | AB558922 | ||
| T. austroindianum | Argentina | BAFC 3583 | MH352421 | X | X | |
| Colombia | CIB T44 | EU279983 | EU280077 | X | ||
| T. azevedoi | Brazil | CEN1422 | MK696660 | MK714901 | MK696821 | |
| T. rifaii* | Ecuador | CBS 130746 | FJ463324 | FJ442663 | X | |
| T. rifaii | Panama | CBS 130745 | FJ463321 | FJ442621 | FJ442720 | |
| Colombia | CIB T99 | EU279990 | EU280103 | X | ||
| T. simmonsii* | USA, MD | CBS 130431 | AF443935 | AF443917 | FJ442757 | |
| T. neotropicale* | Peru | G.J.S. 11-185 | HQ022771 | HQ022407 | X | |
| T. aggressivum | Northern Ireland | CBS 433.95 | AF348097 | FJ442605 | FJ442704 | |
| T. aggressivum | U.K. | CBS 100525 | AF348095 | AF057600 | AF545541 | |
| T. pleuroticola* | South Korea | CBS 124383 | HM142381 | HM142362 | HM142371 | |
| T. pleuroti* | South Korea | CBS 124387 | HM142382 | HM142363 | HM142372 | |
| T. peberdyi | Brazil | CEN1387 | MK696619 | MK714861 | MK696781 | |
| T. peberdyi | Brazil | CEN1388 | MK696620 | MK714862 | MK696782 | |
| T. tomentusom* | Canada | DAOM 178713a | AY750882 | EU330958 | AF545557 |
a , boldface strains are reference sequences. b numbers above one indicates more than one strain had the same tef1α GenBank accession number. *, ex-type species.
Table 2.
Trichoderma species in the Harzianum Complex Clade in soils of South and Central America based on data of five survey studies.
Table 2.
Trichoderma species in the Harzianum Complex Clade in soils of South and Central America based on data of five survey studies.
| Species | Strain number | Number of isolates | tef1α accession number | Country | Ref. |
|---|---|---|---|---|---|
| T. afroharzianum | CIB T136 | 1 | EU279981 | Colombia | 19 |
| CIB T07, CIB T63, CIB T61, CIB T53, CIB T127 | 5 | EU279980* | Colombia | 19 | |
| CIB 59 | 1 | EU279986 | Colombia | 19 | |
| CEN, 1410, CEN1414, CEN1417 | 3 | MK696648* | Brazil | 22 | |
| TUB F-567, TUB F-573, TUB F-452 | 3 | AY857267, AY857268, AY857266 | Peru | 21 | |
| BAFC 4374, BAF 4392, for the rest see the reference | 35 | MH395411, MH395415 | Argentina | 23 | |
| Total | 48 | ||||
| T. lentiforme | DAOM 237544 | 1 | EU279993 | Peru | 19 |
| CIB T02, CIB T112, CIB T35, JB M10-2 | 4 | EU279976* | Mexico, Colombia | 19 | |
| CIB T15, CIB T41 | 2 | EU279982* | Colombia | 19 | |
| DAOM 231417 | 1 | AY605771 | Mexico | 19 | |
| DAOM 231439 | 1 | EU279994 | Mexico | 19 | |
| DAOM 231408 | 1 | AY605773 | Mexico | 19 | |
| DAOM 231405 | 1 | AY605774 | Mexico | 19 | |
| DAOM 231425 | 1 | AY605768 | Mexico | 19 | |
| CIB T56, CIB T60, CIB T16,DAOM 229985 | 4 | EU279985* | Panama, Colombia | 19 | |
| CIB T91 | 1 | EU279987 | |||
| CIB T11, CIB T102 | 2 | EU279979* | Colombia | 19 | |
| DAOM 231402 | 1 | AY605775 | Mexico | 19 | |
| CEN1412, CEN1415, CEN1416, CEN1428, CEN1429 | 5 | MK696650, MK696653, MK696654, MK696668, MK696667 | Brazil | 22 | |
| TUB F-839, TUB F-1073, TUB F-1006, TUB F-746, TUB F-1074 | 5 | AY857283, AY857295, AY857286, AY857257, AY857296 | México, Brazil, Brazil, Brazil, Argentina | 21 | |
| BAFC 4391, BAFC 4394, for the rest see the reference | 9 | MH036883, MH036885 | Argentina | 23 | |
| Total | 39 | ||||
| T. endophyticuma | CIB T03, CIB T31, CIB T139 | 3 | EU279977, EU279988, EU279991 | Colombia | 19 |
| TUB F-1083, TUB F-693 | 2 | AY857300, AY857271 | Mexico, Guatemala | 21 | |
| Th051 | 1 | AB568382 | Colombia | 20 | |
| BAFC 4358, BAFC 4372, for the rest see the reference | 18 | MH371393, MH371397 | Argentina | 23 | |
| Total | 24 | ||||
| T. azevedoi | CIB T23, CIB T24, CIB T126, CIB T128 | 4 | EU279989* | Colombia | 19 |
| CIB T44 | 1 | EU279983 | Colombia | 19 | |
| CEN1422, CEN1423, CEN1403 | 3 | MK696638* | Brazil | 22 | |
| Th202, Th203 | 2 | AB558911* | Colombia | 20 | |
| BAFC 3583, BAFC 3844, GJS 08-128, GJS 08-181, VAB-T051 | 5 | MH352421, MG822709, MH352423, MH352422, MH352424 | Argentina | 23 | |
| Total | 15 | ||||
| T. harzianum | CIB T100, PER4-2 | 2 | EU279978* | Colombia, Peru | 19 |
| TUB F-1078 | 1 | AY857298 | Mexico | 21 | |
| GJS 08-172, GJS 08-173, VAB-T032, VAB-T052, VAB-T053 | 5 | KT275197, KT275198, KT275199, MH364354, MH364355 | Argentina | 23 | |
| Total | 8 | ||||
| T. guizhouense | BAFC 4356, BAFC 4370, GJS 08-102, GJS 08-121, VAB-T047 | 5 | MG797485, MG797486, MG797484, MG797483, MG797482 | Argentina | 23 |
| T. neotropicale | GJS 08-182, GJS 08-183, VAB-T049 | 3 | MG822718, MG822719, MG822720 | Argentina | 23 |
| T. syagri | BAFC 4357, BAFC 4371 | 2 | MG227714, MG227710 | Argentina | 23 |
| T. pseudopyramidale | CIB T52 | 1 | EU279984 | Peru | 19 |
| T. hortense | GJS 08-116 | 1 | MH253895 | Argentina | 23 |
| T. rifaii | CIB T99 | 1 | EU279990 | Colombia | 19 |
| T. sp. | JB PER6-2 | 1 | EU279992 | Peru | 19 |
*Strains have identical tef1α GenBank accession numbers.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



