
Submitted:
23 July 2024
Posted:
23 July 2024
Read the latest preprint version here
Abstract
Human development encompasses the integration of neuromotor, psychological, social-emotional, and cognitive processes across time. However, a better understanding of the interconnectedness of motor and cognitive development throughout the lifespan is needed. The integrative and dynamic nature of short-term motor skill acquisition and long-term developmental processes may foster executive functions. Unfortunately, assessments of motor competence have traditionally focused on skillfulness from a movement perspective, using restrictive task protocols that decontextualize performance and limit cognitive involvement. Additionally, traditional motor-cognitive dual-task assessments centered on cognitive involvement during specific movement protocols, have minimized impacts of task complexity and motor competence on cognitive performance. To gain a more ecologically valid understanding of the relation between motor and cognitive development, a novel assessment paradigm using an integrative approach to motor-cognitive dual-tasking is needed. The purpose of the present paper is to: 1) provide a conceptual bridge, based on multidisciplinary evidence, to effectively link the concurrent development of motor competence and executive functions via learning-related and exercise-related neurotrophic mechanisms, and 2) use this conceptual bridge to inform the development of novel motor-cognitive dual-task assessments that account for the role of task complexity, current levels of motor competence, and the continuous decision-making inherent in “real world” performance environments.
Keywords:
executive function
; perception-action
; dual-task
; motor competence
; motor development
; measurement
1. Introduction
The development of motor competence (MC) is essential for an individual to perceive, navigate, and explore the physical world and may be a critical antecedent for enhanced functioning in cognitive, psychological, social, and emotional domains [1,2]. Embodied learning theories encourage experiences that promote the exploration of novel and challenging bodily states [3], which inherently require the development of a foundation of competency in a variety of movement forms (i.e., MC) ranging from postural control and stability skills to locomotor and object control skills [4]. Likewise, core executive functions (EFs) play a critical role in the complex and continuous nature of decision-making and behavior regulation, which promote exploration in real-world environments. EFs are typically conceptualized as top-down processes, allowing an individual to regulate volitional behavior and successfully manage their actions in accordance with their task-goals [5]. Providing opportunities for children and adolescents that enable them to expand their capacity to perceive, navigate, and explore their physical, social-emotional, and sociocultural environments is critical for promoting the interconnected development of MC and EFs [4,6].
1.1. Executive Functions
EFs are employed in everyday situations to promote or suppress behaviors, switch between tasks, and adapt behavioral strategies [7,8]. Three established core EFs needed to build more complex EFs are: (1) Inhibitory Control – the ability to suppress interference or resist an automatic response in order to make a task-relevant response, (2) Working Memory – the active replacement of irrelevant information with newer task-relevant information to support the performance of goal-oriented behaviors, and (3) Cognitive Flexibility – the ability to switch between task sets or response rules, even in the presence of proactive interference or negative priming [9]. These three core cognitive processes function synergistically with the acquisition and long-term development of MC across the lifespan [5,9,10]. As EFs are used to regulate volitional behavior, they can be applied in various contexts across an emotional-motivational continuum. In cognitive psychology, EFs are routinely dichotomized into those applied in “cool” contexts with minimal incentive and/or emotional intensity and those applied in “hot” contexts where emotional-motivational aspects of behavior are intensified [11,12].
1.1. “Cool” Executive Functions
Traditionally, “cool” views on cognition have focused on the development and maintenance of mental processes that support goal-directed and future-oriented behaviors in relatively decontextualized, analytical, and emotionally detached testing conditions [9,13]. Clinical observations in pioneering research regarding focal brain damage documented differences in behavioral outcomes between damage to the frontal cortex and damage to posterior areas of the brain, revealing frontal patients lacked the ability to use basic cognitive functions to adapt their behaviors to meet future task-goals [14,15]. These findings narrowed investigations towards “cool” EFs, which centered on the relation between frontal lobe structures and anticipatory, goal-oriented behaviors essential to adaptive behavior [16]. In particular, researchers emphasized cognitive functions mediated by the prefrontal cortex and identified neural networks involved in cognitive control, such as the cingulo-opercular and fronto-parietal networks [17].
The cingulo-opercular network is believed to promote the maintenance of task-relevant goals and incorporate error-based adjustments to behavior; whereas the fronto-parietal network is associated with executive control during task execution and facilitates selective attention to task-relevant information [18]. Damage to the prefrontal cortex, a critical brain region within the fronto-parietal and cingulo-opercular networks, may not substantially impair EF performance in isolated conditions but could result in poor social regulation and increased impulsivity in social-decision-making [19,20]. The identification of frontal regions involved in socio-emotional regulation [21] is paralleled, at the behavioral research level, by an increased interest in the context-specific applications of EFs recruited in emotional-motivational aspects of risk-reward analyses and decision making, now commonly labeled as “hot” EFs [11].
1.2. “Hot” Executive Functions
The perception of emotionally salient stimuli significantly impacts cognitive performance [22]. Thus, the ability to selectively respond to relevant aspects of the environment while simultaneously inhibiting potential distractions from emotionally-charged cues and competing choices of action is critical for managing behaviors to achieve task-goals [23,24]. “Hot” EFs serve an important role in filtering out emotionally irrelevant information by means of emotional gating processes. Since representations of task-sets, goals, and other task-relevant information are continually updating, these gating processes serve to optimize the capacity of working memory [24,25]. Individuals commonly use “hot” EFs to flexibly apply emotion regulation strategies to adapt to situational demands [26]. More specifically, individuals recruit “hot” EFs in social environments to self-regulate responses to stimuli such as reward processing, social behavior, and affective decision-making that promote successful interactions with others [27].
The regulation of “hot” EFs is associated with activity in two distinct regions in the frontal lobe of the brain: the ventromedial prefrontal cortex and caudal orbitofrontal cortex [28]. Cerebral areas, including the medial prefrontal cortex, posterior cingulate cortex, and parietal regions, which contribute to the default mode network and are commonly referred to as the “task negative” network, are largely deactivated during the regulation of “hot” EFs [29,30]. Activation of the default mode network during “hot” EF tasks negatively impacts cognitive performance; however, the exact functions of this network are not fully understood as adolescents may recruit the default mode network during tasks that involve aspects of higher-level social cognition [31]. “Hot” EFs critically depend on appropriate levels of the ventromedial prefrontal cortex and caudal orbitofrontal cortex activity to regulate limbic functions in contexts where emotional-motivational aspects of behavior are intensified [12,32,33,34]. Labeling EFs as either “hot” or “cool” may provide unique insights into the functions of individual aspects of behavior; however, this reductionist approach neglects the interconnectedness and blending of EFs that occurs in “real world” situations.
1.3. A Hot–Cool Gradient
As information processing in the brain relies on complex reciprocal interactions within- and between-networks, the question of whether brain processes associated with “emotion” can be definitively separated from those associated with “cognition” has been debated as cognition includes both emotional and logical information [22,35,36]. Zelazo and Müller [12] suggested that “hot” and “cool” EFs exist on discernable ends of a continuum, allowing an individual to concurrently engage both “hot” and “cool” EFs at varying proportions, analogous to tempering water to attain a desired temperature. This tempering of “hot” and “cool” EFs facilitates exploration and meaningful interaction with an individual’s environment. Evidence supports that individuals engage core “cool” EFs to act on both emotional/motivational and logical information [12]. Moreover, conflict monitoring in both cognitive and emotional tasks can be detected using an electrophysiological marker known as the error-related negativity, which emanates from the anterior cingulate cortex [37]. Thus, the anterior cingulate cortex may have the role of a true nexus of emotional and cognitive processing involved in “hot” and “cool” EFs.
Evidence from meta-analyses of neuroimaging studies also supports the hypothesis of a “hot-cool” EF gradient, as traditionally “cool” tasks elicit activity in the ventrolateral prefrontal cortex and anterior insula which are commonly associated with emotions, suggesting functional relations between “hot” and “cool” brain regions [38,39]. In addition, the central autonomic network also suggests functional coupling between cognitive and emotional processing as this network crosses all local networks that subserve both “hot” and “cool” EFs to maintain and regulate the autonomic nervous system [40]. Integrating a cognitive-motivational perspective on this view, developments of Achievement Goal Theory incorporate affective and emotional aspects of motivation along with social cognitive processes and emphasize that goal-directed behaviors are influenced by emotional and motivational states (e.g., [41]). Therefore, tasks investigating “cool” EFs cannot be devoid of “hot” EFs as motivation and emotion are ever-present in goal-directed behaviors; thus, providing additional support towards the hypothesis of a “hot-cool” EF gradient.
2. How May Motor Learning Promote Executive Function Development?
With an understanding of how EFs support goal-directed behaviors, the interconnectedness of underlying brain regions, and their respective neural networks, a logical direction forward is to explore how motor learning processes foster the development of EFs. The human brain dynamically adapts both functionally, by adjusting patterns, strength, or efficiency of neural activity, and structurally, through anatomic changes by creating new synapses and generating new neurons to meet environmental demands. For example, the initial acquisition of movement coordination and control develops via functional changes in neural activity and may result in marked improvements in physical performance over the course of a single training session [42,43]. With continued motor skill practice, structural changes occur over time within the regions of the brain that are activated during motor skill acquisition and performance [42,44]. These structural changes involve the creation of new synapses and new neurons, processes commonly referred to as synaptogenesis and neurogenesis, respectively [44]. The brain employs these functional and structural connections to dynamically regulate neural processes within and between networks, to meet the specific demands imposed by the performer’s task-goal [45,46,47]. Promoting functional and structural adaptations across the brain is critical for the development of core EFs and increasingly complex levels of cognitive functioning over the lifespan [48]. Thus, a better understanding of the integrative and dynamic nature of motor learning and cognitive processes is needed to align MC assessments more effectively with cognitive outcomes.
2.1. Skill Acquisition and Functional Adaptations
Functional changes in neural activity associated with skill acquisition primarily occur in the fronto-parietal and default mode networks [49]. As previously mentioned, functions of the fronto-parietal and default mode networks are inversely related. Therefore, increased levels of neural activity in the default mode network may reflect a shift from more cognitively controlled to automatic task performance [50]. Increases in neural activity in the default mode network also represent an increase in cognitive resources that can be used to explore internal thoughts, memories, and future goals which are critical for decision-making [50]. Conversely, neural activity in the fronto-parietal network is likely used to allocate attentional resources to relevant task and environmental features [51,52]. Thus, neural activity within the default mode network is inversely related to task and environmental complexity [53,54,55]; suggesting that functional decreases in neural activity across nodes within the fronto-parietal network may serve as key neural markers for learning, automaticity, and decreased demands on cognitive control in movement execution. For example, when learning a visuo-motor task, higher level performers decrease neural activity in the fronto-parietal network and increase activity in the default mode network earlier than lower-level performers [56]. Therefore, functional shifts in neural activity from the fronto-parietal network to the default mode network may also reflect a decrease in challenges to EFs as cognitive resources are no longer allocated to explicitly coordinate and control movements. Functional changes in neural activity between networks may represent some EF-related benefits of skill acquisition, as finite cognitive resources are freed with increasing levels of automaticity and skillfulness. However, motor skill complexity and skill retention also promote structural changes which serve unique roles in improving EFs, fostering their development through continual challenges imposed by task and environmental demands [57].
2.2. Skill Retention and Structural Adaptations
Motor learning produces changes in the cerebellum [58,59], basal ganglia [60,61], hippocampus [62], and prefrontal cortex [58], which are due to the involvement of social-emotional, psychological, cognitive, and physical domains during skill learning. The acquisition stage of motor learning is associated with functional changes in neural activity within and between neural networks [42,43]. Meanwhile, the retention stage of motor learning is primarily associated with synaptogenesis [63] and neurogenesis [64]. Evidence from rodent models demonstrates that complex skill learning promotes increases in synaptogenesis within the cerebellum, basal ganglia, and prefrontal cortex [65]. Complex skill learning also impacts memory and learning processes in rodents by promoting the development of newly generated neurons in the hippocampus [64,66]. The acquisition and retention of complex motor skills requires effortful and sustained practice over time which may promote larger synaptogenic and neurogenic effects compared to less complex motor skills [64,67]. Complex motor skill practice requires individuals to actively attend to changing task and environmental information and adapt skill performance to effectively accomplish the task-goal which may challenge EFs more than less complex tasks. In addition, recent evidence suggests that an individual’s skill level may influence the effects of skill learning on one’s neural changes [68]. Therefore, motor learning tasks must maintain an optimal challenge point between the learner’s developmental level and the motoric complexity of the task to promote successful learning, performance, and motivation [69], as well as to foster EFs [70,71,72].
2.3. Exercise-Related Structural Adaptations
Investigations on exercise-related mechanisms that may affect cognition have shown positive effects on angiogenesis, which refers to the generation of new blood vessels, and neurogenesis [73]. However, recent evidence indicates that exercise intensity may moderate the magnitude of these angiogenic and neurogenic effects [74,75,76]. Exercises performed at moderate to vigorous intensities may stimulate angiogenesis and neurogenesis by increasing blood flow as well as energy and oxygen demands [77]. Evidence in mice further supports the moderating effects of exercise intensity on angiogenic responses as high-intensity exercise promoted greater levels of angiogenesis compared to moderate- and light-intensity exercise [76]. Similarly, neurogenic effects may be influenced by high-intensity exercise as research in humans has demonstrated more pronounced neurogenesis compared to moderate- and light-intensity exercise [74,75]. Because exercise promotes angiogenesis and neurogenesis, the moderating role of exercise intensity must be considered when investigating exercise-related effects on cognition. As mentioned in section 2.1, complex skill learning is deeply intertwined with cognitive development; however, evidence focusing on the interactions between movement complexity, exercise intensity, and EF-related outcomes is limited and warrants investigation.
3. Developmental Perspectives on Motor Competence and Executive Functions
The development of MC concurrently promotes the development of cognitive structures via learning- and exercise-related mechanisms. Learning-related mechanisms are influenced by movement task complexity while exercise-related mechanisms are influenced by the energetic demands of physical movement. To understand the integrative nature of MC and EFs from a developmental perspective, we examine how perception, action, and decision processes are deeply interconnected with physical movement and involve increasingly complex interactions within the individual-environment system.
To accomplish any movement-related task, an individual must identify relations between task and environmental features and the individual’s own movement capabilities to establish actionable qualities of the performance environment [78,79]. As such, the performance of any action is coupled with perceptions of constraints defined by the task, the environment, and the individual [79,80], commonly referred to as perception-action cycles. Embodied cognition encompasses these dynamic interactions between cognition and MC, spanning both micro and macro timescales, which are shaped by an individual’s discovery of novel perception-action couplings through repeated and meaningful interactions with their environment [81]. Aligned with this view, the Skilled Intentionality Framework [82] offers unique insight into “hot” EFs as these cognitive processes are intricately intertwined with bodily interactions, dynamically responsive to context-specific experiences, and linked to an individual’s emotional state. Furthermore, we contend that computational and representational theories of cognition offer complementary perspectives (i.e., hot-cool gradient) on symbolic representations, abstract reasoning, and mental imagery [83]; processes that rely on “cool” EFs to effectively facilitate the development of strategic solutions for both motor and cognitive challenges.
To enhance one’s capability to control their movements and explore the environment, an individual must regulate the complex interactions that occur within the individual-environment system. One method for adapting to complex task and environmental demands in early stages of movement development is to reduce or “freeze” degrees of freedom within the muscles, joints, and limbs to develop coordination strategies and optimize control to support goal-directed actions [84,85]. For example, at the onset of independent walking infants will “freeze” their arms in a high-guard position to decrease the complex physical demands needed for dynamic postural control [86]. As salient perception-action couplings become more frequent through effortful practice, the infant will begin to “un-freeze” degrees of freedom by adding oppositional arm movements. This will result in a more advanced locomotion which, in turn, will enhance the discovery of opportunities for action within the environment and insight into object properties and spatial relations [78,87]. The acquisition and advanced development of walking likely requires the infant to 1) Inhibit primitive neuro-motor synergies like the symmetrical coupling of the upper limbs [88], 2) Use working memory to enhance self-navigation using landmarks [1], and 3) Flexibly shift attention between interacting with objects, toys, other children, or adults and navigating the environment. Thus, the “education of attention” through active exploration within the task-environment and the search for task-relevant perceptual information not only promotes skill development but may also promote the development of core “cool” EFs [72]. In addition, infants may engage core “hot” EFs to initiate or regulate goal-directed behaviors, such as repeated attempts to walk despite failure, by referring to emotional signals provided by adults [89]. These cognitive processes are integrated within the sequential development of a variety of more complex locomotor and object control skills, like hopping, jumping, throwing, and kicking, in an intransitive and cumulative manner across childhood and adolescence.
Although there is still a general belief that competence in many skills is developed “naturally,” the inherent complexities involved in the developmental progression of skillfulness in a variety of motor skills (i.e., MC) requires repeated, effortful, and successful interactions with perceptually rich and dynamic environments [90]. Another commonly held belief is that breaking down complex tasks and tightly controlling environments benefits short-term skill acquisition [91]. However, promoting variability in task and environmental conditions is a hallmark of various motor learning and development theories as it optimizes learning and retention from a developmental perspective. Therefore, while more motorically complex tasks in dynamic environmental conditions incur slower acquisition rates, they promote higher levels of neural activity in the fronto-parietal network [92,93], synaptogenesis [63,67] and hippocampal neuron survival [64,66]; indicating that complex tasks are more effective for long-term development of MC and EFs [70,71].
Moreover, according to the exercise-related mechanisms introduced in section 2.3, the development of EFs is also enhanced through the physical effort and intensity generated by the prolonged practice of complex motor skills that are needed to continuously develop higher levels of competence. Specifically, the contexts surrounding the acquisition of motor skills during structured and unstructured play, sports, or specific skill practice provide individuals with opportunities for continued participation in moderate-to-vigorous physical activities that subsequently promote the development of cardiorespiratory and musculoskeletal fitness [94,95,96]. Physical activities and sports that inherently require complex skill performance also stimulate the development of musculoskeletal fitness via enhanced inter- and intra-muscular recruitment strategies [97]. Higher levels of exercise intensity are also related to more pronounced vascular and neurotrophic benefits [75] which may result in differential effects of complex skill learning for lower and higher skilled individuals. For example, higher skilled individuals generally have greater force production capabilities than lower skilled individuals [94,95,96,98] which is associated with higher levels of energy expenditure during object projection skills like throwing and kicking [99,100]. Thus, it is logical that higher skilled individuals may also experience greater exercise induced cognitive effects from complex skill training than lower skilled individuals due to the potential for more pronounced vascular and neurotrophic changes.
3.1. Linking Motor Competence Assessments to Cognitive Outcomes
In the sections above, we have emphasized how the development of EFs are intricately interwoven with complex skill learning and the concomitant integration of motor-cognitive processes in everyday life activities. For these reasons, we believe there is a need for assessments that effectively tap into the interactive processes of the motor and cognitive systems, as well as their individual contributions to overall motor-cognitive performance. In this section, we discuss a novel approach to MC-EF paradigms and offer exemplars of motor tasks that concurrently require a) greater motor complexity than current single- and dual-task paradigms, b) increase cognitive demands, and c) more effectively align with “real world” performance environments, thus exhibiting greater ecological validity. In addition, the application of novel assessments that are developmentally valid across childhood and adulthood allows for the simultaneous assessment of EFs across a wide age range of capabilities, facilitating the ability to assess and track the development of embodied cognition across time.
3.2. Limitations of Current Motor Competence Assessments and Their Mis/Alignment with Executive Functions
Restrictive assessment protocols in current MC assessments limit ecological validity and therefore limit their ability to capture motor and cognitive performance [101]. For example, discrete MC assessments like the standing long jump, supine-to-stand, and throwing for speed require individuals to perform discrete tasks in a way that limits the integration of salient environmental influences that impact skill performance. Further, these restrictive MC assessments decontextualize the motor skill and disregard the complex and continuous nature of decision-making in embodied performance settings. In this section, we discuss a novel approach to MC-EF paradigms and offer developmentally appropriate examples that embrace embodied decision-making.
Discrete MC assessment protocols require restrictive task demands which either purposefully or inadvertently limit participant decision-making and trivialize the effects of action dynamics on subsequent perception dynamics. The theoretical underpinnings of discrete decision-making tasks propose that decision-making is a sequential process [102,103]. This sequential process can be summarized as “decide then act”, in which, the decision-maker first perceives affordances within the assessment task and environment, subsequently chooses between predefined task solutions, and then acts on their decision [104]. Sequential decision-making processes underlying traditional discrete MC assessments rely on rigid task protocols which provide greater levels of experimental control and promote reproducible research. However, these tasks may have distorted our view of MC-EF relations by creating stable environments and predefined motor-cognitive solutions. This is exemplified when researchers provide participants with the “correct” coordination pattern for kicking a ball with maximum effort, in a controlled environment, without time or accuracy constraints. Alternatively, tasks that promote continuous decision-making provide a continuum of potential adaptations to motor-cognitive solutions that are uniquely affected by recent responses and continuously subject to change [105]. Expanding on the previous example, to enhance traditional task protocols, continuously kicking and receiving a ball from a wall with a time constraint allows an individual to explore and adapt motor-cognitive solutions (i.e., divergent movements) using real-time decision-making. Tasks that align with the dynamic nature of continuous decision-making open avenues for examining not only questions of coordination but also those of control and timing [105].
Current MC assessments can easily integrate continuous decision-making by promoting time-constrained tasks that require repeated performance of discrete motor skills or performance of continuous motor skills in more dynamic environments. In contrast to discrete and controlled decision-making, continuous decision-making is marked by two reciprocal processes which are “acting while deciding” and “deciding while acting” [104]. When “acting while deciding” the decision-maker initiates actions before completing the decision either to exploit options that would otherwise disappear, such as running towards defenders while searching for open gaps, or to buy time before initiating an action response [106]. Conversely, when “deciding while acting” the decision-maker may adapt their performance in real time as a result of perceiving novel affordances, gathering novel information, or reconsidering a previous decision [107]. Thus, one’s ability to apply “hot” and “cool” aspects of inhibitory control, working memory, and cognitive flexibility during “real world” continuous decision-making tasks is critically needed to effectively and efficiently adapt behaviors to achieve task-goals [5]. Further, motor skill performance and adjustment capabilities are impacted by an individual’s motor skill repertoire and the subsequent cognitive demands experienced by the individual during continuous decision-making tasks [108].
3.3. Integration of Executive Functions and Motor Tasks: Dual-Task Paradigms
Motor-cognitive dual-task paradigms provide an excellent “jumping off point” for our reconceptualization of motor-cognitive assessment paradigms as there is a need to more effectively a) capture cognitive demands concurrently associated with movement actions across various types of tasks, b) provide multiple combinations of potential task solutions, c) account for differences in skill levels, and d) represent dynamic everyday life contexts [109]. Traditional motor-cognitive dual-task paradigms focus on the application of EFs to flexibly allocate attentional resources between two distinct tasks such as simultaneously walking and counting [110]. These traditional approaches sacrifice ecological validity by oversimplifying the motoric context and thus, lowering the cognitive demands of the task. In addition, limited movement complexity may not provide the measurement sensitivity to capture differences in motor-cognitive capabilities based on current skill levels and varied motor repertoires across individuals.
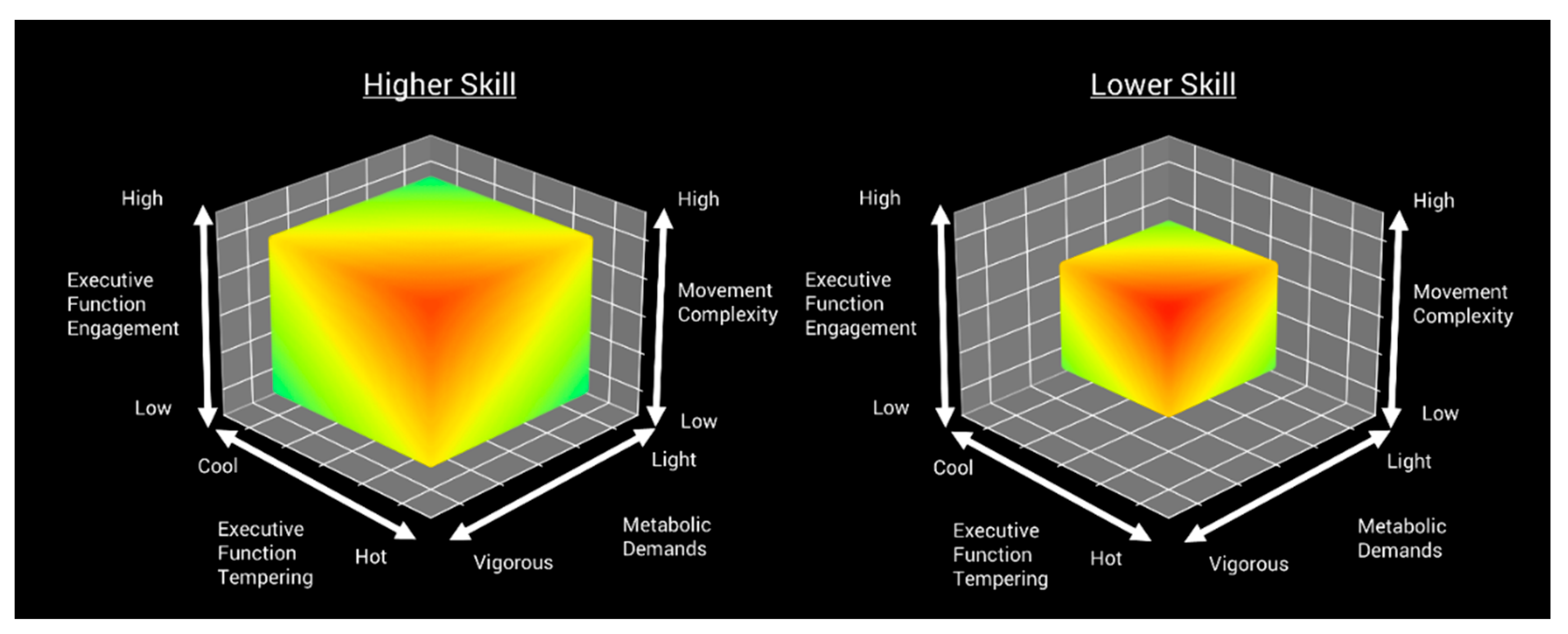
A novel and more ecologically valid approach to examining an individual’s cognitive function includes tasks that allow for choice between different motor-cognitive solutions or adaptations to a previously attempted motor-cognitive solution in continuous decision-making tasks. Such tasks require increased levels of core EFs to update feedback from previous performances, inhibit ineffective motor actions, and flexibly attend to both motor and cognitive aspects of task performance. This assessment scenario enhances real-time decision-making capabilities and speaks to the importance of having a diverse MC repertoire. As individual, task, and environmental constraints define the space within which motor-cognitive solutions can emerge, a diverse MC repertoire facilitates a broad spectrum of potential motor-cognitive solutions to achieve the task-goal. Figure 1 illustrates the spectrum of potential motor-cognitive solutions that may emerge from an individual's MC repertoire. These solutions are contingent upon the current task and environmental constraints, which, in turn, influence factors such as motoric complexity, metabolic demands, engagement of EFs, and the tempering of "hot-cool" EF processes. Advanced skill levels offer greater flexibility in motor-cognitive solutions that are predicated on an individual’s ability to find an acceptable solution to the task based on either success or failure of meeting the task-goal and, if necessary, modify performance based on continuous feedback from previous performances. Thus, the development and enhancement of an advanced MC repertoire, both within and across a broad foundation of skills, inherently requires repeated experiences with varied combinations of potential motor-cognitive solutions to meet context-specific task-goals.
Hulteen and colleagues [111] allude to this advancement in the dual-task paradigm by promoting two dynamic and continuous tasks in the context of MC assessment, the throw-catch [112] and supine-to-stand [and go]. Both tasks offer a continuum of coordinative and cognitive solutions that are inextricably linked to previous within-task responses and afford moment-to-moment adaptations in performance. Dual-task paradigms that promote continuous decision-making allow for considerable amounts of individual variation in coordination, control, and cognitive strategies as individuals seek to assemble, perform, and adapt their own motor-cognitive solutions [113]. Further, dual-task paradigms that afford continuous decision-making empower individuals to continuously regulate complex motoric and cognitive interactions within the individual-environment systems by exploring potential motor-cognitive solutions until an optimal performance state is achieved. Accumulating a broad MC repertoire and optimizing coordination and control, often referred to as skill, requires considerable experience, repeated active exploration of skill execution options, and effortful practice in context-specific environments. Increased exposure to diverse skill practice within dynamic environmental contexts and varying task constraints enhances an individual’s ability to quickly identify and optimally utilize task-relevant perceptual information. Ultimately, these experiences reduce the cognitive resources needed for skill execution and enhance an individual’s capacity to engage EFs to effectively adapt performance based on task-specific demands [114]. In Figure 2, we offer a conceptual representation that demonstrates the spectrum of potential motor-cognitive solutions available across different skill levels. Comparatively, individuals with lower skill levels have a more restricted range of potential motor-cognitive solutions, primarily due to their less developed MC repertoire. This novel view of the dual-task paradigm uniquely acknowledges the importance of an individual’s capacity to coordinate and control movements (i.e., skill level) and their accumulated context-specific experiences, while capturing learning- and exercise-related mechanisms inherent in the concurrent development of MC and EFs.
4. Conclusions
We hope this paper advances the understanding of interactions between motor and cognitive domains and provides insights into the mechanisms, through which physically effortful and complex motor skill learning can contribute to the joint development of MC and EFs. Second, we provide ideas for novel assessments to enhance the way we examine the relation between motor and cognitive development across the lifespan, while considering both “hot” and “cool” EFs. We contend that advancing the dual-task assessment paradigm, by accounting for the interactions between learning- and exercise-related mechanisms and the moderating roles of task complexity and an individual’s skill level, will more effectively capture the interconnectedness of motor and cognitive development. Most importantly, we provide a rationale to promote motor tasks that require continuous decision-making and more closely align with “real world” performance environments. Taken together, these advancements have the potential to provide new directions for embodied cognitive research across the lifespan.
Author Contributions
Conceptualization, T.C.A, C.P., and D.F.S; software, T.C.A.; writing—original draft preparation, T.C.A; writing—review and editing, R.D.M., C.P., A.DM., A.B., D.F.S.; visualization, T.C.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
While no data were analyzed in this manuscript, the code necessary to reproduce Figure 1 and Figure 2 is publicly available at https://github.com/PlayfulMaven/complexity-dual-task-conceptual.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Campos, J.J.; Anderson, D.I.; Barbu-Roth, M.A.; Hubbard, E.M.; Hertenstein, M.J.; Witherington, D. Travel Broadens the Mind. Infancy 2000, 1, 149–219. [Google Scholar] [CrossRef]
- Kenny, L.; Hill, E.; Hamilton, A.F. de C. The Relationship between Social and Motor Cognition in Primary School Age-Children. Front. Psychol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Skulmowski, A.; Rey, G.D. Embodied Learning: Introducing a Taxonomy Based on Bodily Engagement and Task Integration. Cogn. Res. Princ. Implic. 2018, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stodden, D.F.; Lakes, K.D.; Côté, J.; Aadland, E.; Brian, A.; Draper, C.E.; Ekkekakis, P.; Fumagalli, G.; Laukkanen, A.; Mavilidi, M.F.; et al. Exploration: An Overarching Focus for Holistic Development. Braz. J. Mot. Behav. 2021, 15. [Google Scholar] [CrossRef]
- Diamond, A. Executive Functions. Annu Rev Psychol 2013, 64, 135–168. [Google Scholar] [CrossRef] [PubMed]
- Pesce, C.; Stodden, D.F.; Lakes, K.D. Physical Activity “Enrichment”: A Joint Focus on Motor Competence, Hot and Cool Executive Functions. Front. Psychol. 2021, 12.
- Barkley, R.A. Executive Functions: What They Are, How They Work, and Why They Evolved; Guilford Press, 2012; ISBN 1-4625-0535-X.
- Goldstein, S.; Naglieri, J.A.; Princiotta, D.; Otero, T.M. Introduction: A History of Executive Functioning as a Theoretical and Clinical Construct. In The handbook of executive functioning; Springer: New York, 2014.
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The Unity and Diversity of Executive Functions and Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable Analysis. Cognit. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.; Ling, D.S. Conclusions about Interventions, Programs, and Approaches for Improving Executive Functions That Appear Justified and Those That, despite Much Hype, Do Not. Dev. Cogn. Neurosci. 2016, 18, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Zelazo, P.D.; Carlson, S.M. Hot and Cool Executive Function in Childhood and Adolescence: Development and Plasticity. Child Dev. Perspect. 2012, 6, 354–360. [Google Scholar] [CrossRef]
- Zelazo, P.D.; Müller, U. Executive Function in Typical and Atypical Development. In Blackwell handbook of childhood cognitive development; Blackwell Publishing, 2002; pp. 445–469.
- Peterson, E.; Welsh, M.C. The Development of Hot and Cool Executive Functions in Childhood and Adolescence: Are We Getting Warmer. In Handbook of Executive Functioning; Goldstein, S., Naglieri, J.A., Eds.; Springer New York: New York, NY, 2014; pp. 45–65. ISBN 978-1-4614-8106-5. [Google Scholar]
- Luria, A.R. Higher Cortical Functions in Man; New York: Basic, 1966.
- Tueber, H. The Riddle of Frontal Lobe Function in Man. In The frontal granular cortex and behavior; J. Warren, Akert, K., Ed.; New York: McGraw-Hill, 1964; pp. 410–440. [Google Scholar]
- Alvarez, J.A.; Emory, E. Executive Function and the Frontal Lobes: A Meta-Analytic Review. Neuropsychol. Rev. 2006, 16, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.P.; Robbins, T.W. The Role of Prefrontal Cortex in Cognitive Control and Executive Function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef]
- Dosenbach, N.U.F.; Fair, D.A.; Miezin, F.M.; Cohen, A.L.; Wenger, K.K.; Dosenbach, R.A.T.; Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E.; et al. Distinct Brain Networks for Adaptive and Stable Task Control in Humans. Proc. Natl. Acad. Sci. 2007, 104, 11073–11078. [Google Scholar] [CrossRef]
- Bechara, A. The Role of Emotion in Decision-Making: Evidence from Neurological Patients with Orbitofrontal Damage. Brain Cogn. 2004, 55, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Bechara, A.; Damasio, A.R.; Damasio, H.; Anderson, S.W. Insensitivity to Future Consequences Following Damage to Human Prefrontal Cortex. Cognition 1994, 50, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Firat, R.B. Opening the “Black Box”: Functions of the Frontal Lobes and Their Implications for Sociology. Front. Sociol. 2019, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Okon-Singer, H.; Hendler, T.; Pessoa, L.; Shackman, A.J. The Neurobiology of Emotion–Cognition Interactions: Fundamental Questions and Strategies for Future Research. Front. Hum. Neurosci. 2015, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Desimone, R.; Duncan, J. Neural Mechanisms of Selective Visual Attention. Annu. Rev. Neurosci. 1995, 18, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.K.; Cohen, J.D. An Integrative Theory of Prefrontal Cortex Function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Stout, D.M.; Shackman, A.J.; Johnson, J.S.; Larson, C.L. Worry Is Associated with Impaired Gating of Threat from Working Memory. Emotion 2015, 15, 6. [Google Scholar] [CrossRef]
- Sheppes, G.; Levin, Z. Emotion Regulation Choice: Selecting between Cognitive Regulation Strategies to Control Emotion. Front. Hum. Neurosci. 2013, 7, 179. [Google Scholar] [CrossRef] [PubMed]
- Leshem, R. Using Dual Process Models to Examine Impulsivity throughout Neural Maturation. Dev. Neuropsychol. 2016, 41, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Fuster, J.M. The Prefrontal Cortex—an Update: Time Is of the Essence. Neuron 2001, 30, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A Default Mode of Brain Function. Proc. Natl. Acad. Sci. 2001, 98, 676–682. [Google Scholar] [CrossRef]
- Pujol, J.; Reixach, J.; Harrison, B.J.; Timoneda-Gallart, C.; Vilanova, J.C.; Pérez-Alvarez, F. Posterior Cingulate Activation during Moral Dilemma in Adolescents. Hum. Brain Mapp. 2008, 29, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Barbas, H. Connections Underlying the Synthesis of Cognition, Memory, and Emotion in Primate Prefrontal Cortices. Brain Res. Bull. 2000, 52, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Otero, T.M.; Barker, L.A. The Frontal Lobes and Executive Functioning. In Handbook of Executive Functioning; Goldstein, S., Naglieri, J.A., Eds.; Springer New York: New York, NY, 2014; pp. 29–44. ISBN 978-1-4614-8106-5. [Google Scholar]
- Padoa-Schioppa, C.; Conen, K.E. Orbitofrontal Cortex: A Neural Circuit for Economic Decisions. Neuron 2017, 96, 736–754. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, L. On the Relationship between Emotion and Cognition. Nat. Rev. Neurosci. 2008, 9, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, P.; Castellanos, F.X. Top-down Dysregulation—from ADHD to Emotional Instability. Front. Behav. Neurosci. 2016, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Bush, G.; Luu, P.; Posner, M.I. Cognitive and Emotional Influences in Anterior Cingulate Cortex. Trends Cogn. Sci. 2000, 4, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Cieslik, E.C.; Mueller, V.I.; Eickhoff, C.R.; Langner, R.; Eickhoff, S.B. Three Key Regions for Supervisory Attentional Control: Evidence from Neuroimaging Meta-Analyses. Neurosci. Biobehav. Rev. 2015, 48, 22–34. [Google Scholar] [CrossRef]
- Nee, D.E.; Wager, T.D.; Jonides, J. Interference Resolution: Insights from a Meta-Analysis of Neuroimaging Tasks. Cogn. Affect. Behav. Neurosci. 2007, 7, 1–17. [Google Scholar] [CrossRef]
- Quadt, L.; Critchley, H.; Nagai, Y. Cognition, Emotion, and the Central Autonomic Network. Auton. Neurosci. 2022, 238, 102948. [Google Scholar] [CrossRef] [PubMed]
- Bj⊘rnebekk, G. Positive Affect and Negative Affect as Modulators of Cognition and Motivation: The Rediscovery of Affect in Achievement Goal Theory. Scand. J. Educ. Res. 2008, 52, 153–170. [Google Scholar] [CrossRef]
- Doyon, J.; Benali, H. Reorganization and Plasticity in the Adult Brain during Learning of Motor Skills. Curr. Opin. Neurobiol. 2005, 15, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Doyon, J.; Ungerleider, L.G. Functional Anatomy of Motor Skill Learning. In Neuropsychology of memory; Squire, L., Schacter, D., Eds.; The Guilford Press, 2002; Vol. 3, pp. 225–238.
- Dayan, E.; Cohen, L.G. Neuroplasticity Subserving Motor Skill Learning. Neuron 2011, 72, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.R.; D’Esposito, M. The Segregation and Integration of Distinct Brain Networks and Their Relationship to Cognition. J. Neurosci. 2016, 36, 12083–12094. [Google Scholar] [CrossRef] [PubMed]
- Mohr, H.; Wolfensteller, U.; Betzel, R.F.; Mišić, B.; Sporns, O.; Richiardi, J.; Ruge, H. Integration and Segregation of Large-Scale Brain Networks during Short-Term Task Automatization. Nat. Commun. 2016, 7, 13217. [Google Scholar] [CrossRef] [PubMed]
- Reijneveld, J.C.; Ponten, S.C.; Berendse, H.W.; Stam, C.J. The Application of Graph Theoretical Analysis to Complex Networks in the Brain. Clin. Neurophysiol. 2007, 118, 2317–2331. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Supekar, K.S.; Ryali, S.; Menon, V. Dynamic Reconfiguration of Structural and Functional Connectivity Across Core Neurocognitive Brain Networks with Development. J. Neurosci. 2011, 31, 18578–18589. [Google Scholar] [CrossRef] [PubMed]
- Hikosaka, O.; Nakamura, K.; Sakai, K.; Nakahara, H. Central Mechanisms of Motor Skill Learning. Curr. Opin. Neurobiol. 2002, 12, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Spreng, R.N.; Turner, G.R. Functional Brain Changes Following Cognitive and Motor Skills Training: A Quantitative Meta-Analysis. Neurorehabil. Neural Repair 2013, 27, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Cisek, P. Cortical Mechanisms of Action Selection: The Affordance Competition Hypothesis. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1585–1599. [Google Scholar] [CrossRef]
- Pezzulo, G.; Cisek, P. Navigating the Affordance Landscape: Feedback Control as a Process Model of Behavior and Cognition. Trends Cogn. Sci. 2016, 20, 414–424. [Google Scholar] [CrossRef]
- Gusnard, D.A.; Raichle, M.E. Searching for a Baseline: Functional Imaging and the Resting Human Brain. Nat. Rev. Neurosci. 2001, 2, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.S.; Roebroeck, A.; Maurer, K.; Linden, D.E. Specialization in the Default Mode: Task-induced Brain Deactivations Dissociate between Visual Working Memory and Attention. Hum. Brain Mapp. 2010, 31, 126–139. [Google Scholar] [CrossRef]
- McKiernan, K.A.; Kaufman, J.N.; Kucera-Thompson, J.; Binder, J.R. A Parametric Manipulation of Factors Affecting Task-Induced Deactivation in Functional Neuroimaging. J. Cogn. Neurosci. 2003, 15, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Hikosaka, O.; Miyauchi, S.; Takino, R.; Sasaki, Y.; Pütz, B. Transition of Brain Activation from Frontal to Parietal Areas in Visuomotor Sequence Learning. J. Neurosci. 1998, 18, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.; Ling, D.S. Review of the Evidence on, and Fundamental Questions about, Efforts to Improve Executive Functions, Including Working Memory. Cogn. Work. Mem. Train. Perspect. Psychol. Neurosci. Hum. Dev. 2020, 145–389. [Google Scholar] [CrossRef]
- Diamond, A. Close Interrelation of Motor Development and Cognitive Development and of the Cerebellum and Prefrontal Cortex. Child Dev. 2000, 71, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Doyon, J.; Song, A.W.; Karni, A.; Lalonde, F.; Adams, M.M.; Ungerleider, L.G. Experience-Dependent Changes in Cerebellar Contributions to Motor Sequence Learning. Proc. Natl. Acad. Sci. 2002, 99, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Leisman, G.; Braun-Benjamin, O.; Melillo, R. Cognitive-Motor Interactions of the Basal Ganglia in Development. Front. Syst. Neurosci. 2014, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Seidler, R.; Noll, D.; Chintalapati, P. Bilateral Basal Ganglia Activation Associated with Sensorimotor Adaptation. Exp. Brain Res. 2006, 175, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Saxe, M.D.; Gallina, I.S.; Gage, F.H. Adult-Born Hippocampal Dentate Granule Cells Undergoing Maturation Modulate Learning and Memory in the Brain. J. Neurosci. 2009, 29, 13532–13542. [Google Scholar] [CrossRef] [PubMed]
- Kleim, J.A.; Barbay, S.; Cooper, N.R.; Hogg, T.M.; Reidel, C.N.; Remple, M.S.; Nudo, R.J. Motor Learning-Dependent Synaptogenesis Is Localized to Functionally Reorganized Motor Cortex. Neurobiol. Learn. Mem. 2002, 77, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Shors, T.J. The Adult Brain Makes New Neurons, and Effortful Learning Keeps Them Alive. Curr. Dir. Psychol. Sci. 2014, 23, 311–318. [Google Scholar] [CrossRef]
- Sampaio, A.S.B.; Real, C.C.; Gutierrez, R.M.S.; Singulani, M.P.; Alouche, S.R.; Britto, L.R.; Pires, R.S. Neuroplasticity Induced by the Retention Period of a Complex Motor Skill Learning in Rats. Behav. Brain Res. 2021, 414, 113480. [Google Scholar] [CrossRef] [PubMed]
- Waddell, J.; Shors, T.J. Neurogenesis, Learning and Associative Strength. Eur. J. Neurosci. 2008, 27, 3020–3028. [Google Scholar] [CrossRef]
- Kleim, J.A.; Hogg, T.M.; VandenBerg, P.M.; Cooper, N.R.; Bruneau, R.; Remple, M. Cortical Synaptogenesis and Motor Map Reorganization Occur during Late, but Not Early, Phase of Motor Skill Learning. J. Neurosci. 2004, 24, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, L.; Larsen, M.N.; Madsen, M.J.; Grey, M.J.; Nielsen, J.B.; Lundbye-Jensen, J. Long-Term Motor Skill Training with Individually Adjusted Progressive Difficulty Enhances Learning and Promotes Corticospinal Plasticity. Sci. Rep. 2020, 10, 15588. [Google Scholar] [CrossRef] [PubMed]
- Guadagnoli, M.A.; Lee, T.D. Challenge Point: A Framework for Conceptualizing the Effects of Various Practice Conditions in Motor Learning. J. Mot. Behav. 2004, 36, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Moreau, D. Brains and Brawn: Complex Motor Activities to Maximize Cognitive Enhancement. Educ. Psychol. Rev. 2015, 27, 475–482. [Google Scholar] [CrossRef]
- Moreau, D.; Morrison, A.B.; Conway, A.R.A. An Ecological Approach to Cognitive Enhancement: Complex Motor Training. Acta Psychol. (Amst.) 2015, 157, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Pesce, C.; Croce, R.; Ben-Soussan, T.D.; Vazou, S.; McCullick, B.; Tomporowski, P.D.; Horvat, M. Variability of Practice as an Interface between Motor and Cognitive Development. Int. J. Sport Exerc. Psychol. 2019, 17, 133–152. [Google Scholar] [CrossRef]
- Hillman, C.H.; Erickson, K.I.; Kramer, A.F. Be Smart, Exercise Your Heart: Exercise Effects on Brain and Cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-Y.; Chen, F.-T.; Li, R.-H.; Hillman, C.H.; Cline, T.L.; Chu, C.-H.; Hung, T.-M.; Chang, Y.-K. Effects of Acute Resistance Exercise on Executive Function: A Systematic Review of the Moderating Role of Intensity and Executive Function Domain. Sports Med. - Open 2022, 8, 141. [Google Scholar] [CrossRef]
- Jeon, Y.K.; Ha, C.H. The Effect of Exercise Intensity on Brain Derived Neurotrophic Factor and Memory in Adolescents. Environ. Health Prev. Med. 2017, 22, 27. [Google Scholar] [CrossRef] [PubMed]
- Morland, C.; Andersson, K.A.; Haugen, Ø.P.; Hadzic, A.; Kleppa, L.; Gille, A.; Rinholm, J.E.; Palibrk, V.; Diget, E.H.; Kennedy, L.H.; et al. Exercise Induces Cerebral VEGF and Angiogenesis via the Lactate Receptor HCAR1. Nat. Commun. 2017, 8, 15557. [Google Scholar] [CrossRef]
- Hugues, N.; Pellegrino, C.; Rivera, C.; Berton, E.; Pin-Barre, C.; Laurin, J. Is High-Intensity Interval Training Suitable to Promote Neuroplasticity and Cognitive Functions after Stroke? Int. J. Mol. Sci. 2021, 22, 3003. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.J. The Theory of Affordances. The Ecological Approach to Visual Perception. In The People, Place and, Space Reader; Routledge New York and London, 1979; pp. 56–60.
- Newell, K. Constraints on the Development of Coordination. In Motor Development in Children: Aspects of Coordination and Control; Wade, M.G., Whiting, H.T.A., Eds.; Martinus Nijhoff: Dordrecht, 1986; pp. 341–360. [Google Scholar]
- Davids, K.; Button, C.; Bennett, S. Dynamics of Skill Acquisition: A Constraints-Led Approach; Human kinetics, 2008; ISBN 0-7360-3686-5.
- Mulder, H.; Oudgenoeg-Paz, O.; Hellendoorn, A.; Jongmans, M.J. How Children Learn to Discover Their Environment: An Embodied Dynamic Systems Perspective on the Development of Spatial Cognition. In Neuropsychology of space: Spatial functions of the human brain; Elsevier Academic Press: San Diego, CA, US, 2017; pp. 309–360. ISBN 978-0-12-801638-1. [Google Scholar]
- Rietveld, E.; Kiverstein, J. A Rich Landscape of Affordances. Ecol. Psychol. 2014, 26, 325–352. [Google Scholar] [CrossRef]
- Beckmann, P.; Köstner, G.; Hipólito, I. An Alternative to Cognitivism: Computational Phenomenology for Deep Learning. Minds Mach. 2023. [Google Scholar] [CrossRef]
- Davids, K.; Araújo, D.; Shuttleworth, R.; Button, C. Acquiring Skill in Sport: A Constraints-Led Perspective. Int. J. Comput. Sci. Sport 2003, 2, 31–39. [Google Scholar]
- Turvey, M.T. Coordination. Am. Psychol. 1990, 45, 938–953. [Google Scholar] [CrossRef] [PubMed]
- Bril, B.; Brenière, Y. Posture and Independent Locomotion in Early Childhood: Learning to Walk or Learning Dynamic Postural Control? In Advances in psychology; Elsevier, 1993; Vol. 97, pp. 337–358 ISBN 0166-4115.
- Gibson, E.J. Exploratory Behavior in the Development of Perceiving, Acting, and the Acquiring of Knowledge. Annu. Rev. Psychol. 1988, 39, 1–42. [Google Scholar] [CrossRef]
- Lazarus, J.C.; Todor, J.I. The Role of Attention in the Regulation of Associated Movement in Children. Dev. Med. Child Neurol. 1991, 33, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.; Lopez, L.D. Studying Hot Executive Function in Infancy: Insights from Research on Emotional Development. Infant Behav. Dev. 2022, 69, 101773. [Google Scholar] [CrossRef]
- Adolph, K.E.; Hoch, J.E. Motor Development: Embodied, Embedded, Enculturated, and Enabling. Annu. Rev. Psychol. 2019, 70, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Magill, R.A.; Lee, T.D. Motor Learning: Concepts and Applications; 6th ed.; WCB McGraw-Hill, 1998; ISBN 0-697-38953-7.
- O’Connell, M.A.; Basak, C. Effects of Task Complexity and Age-Differences on Task-Related Functional Connectivity of Attentional Networks. Neuropsychologia 2018, 114, 50–64. [Google Scholar] [CrossRef]
- Shashidhara, S.; Mitchell, D.J.; Erez, Y.; Duncan, J. Progressive Recruitment of the Frontoparietal Multiple-Demand System with Increased Task Complexity, Time Pressure, and Reward. J. Cogn. Neurosci. 2019, 31, 1617–1630. [Google Scholar] [CrossRef] [PubMed]
- Abrams, T.C.; Terlizzi, B.M.; De Meester, A.; Sacko, R.S.; Irwin, J.M.; Luz, C.; Rodrigues, L.P.; Cordovil, R.; Lopes, V.P.; Schneider, K.; et al. Potential Relevance of a Motor Skill" Proficiency Barrier" on Health-Related Fitness in Youth. Eur. J. Sport Sci. 2022, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cattuzzo, M.T.; dos Santos Henrique, R.; Ré, A.H.N.; de Oliveira, I.S.; Melo, B.M.; de Sousa Moura, M.; de Araújo, R.C.; Stodden, D. Motor Competence and Health Related Physical Fitness in Youth: A Systematic Review. J. Sci. Med. Sport 2016, 19, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Utesch, T.; Bardid, F.; Büsch, D.; Strauss, B. The Relationship between Motor Competence and Physical Fitness from Early Childhood to Early Adulthood: A Meta-Analysis. Sports Med. 2019, 49, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Stodden, D.F.; Gao, Z.; Goodway, J.D.; Langendorfer, S.J. Dynamic Relationships between Motor Skill Competence and Health-Related Fitness in Youth. Pediatr. Exerc. Sci. 2014, 26, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Stodden, D.F.; True, L.K.; Langendorfer, S.J.; Gao, Z. Associations Among Selected Motor Skills and Health-Related Fitness: Indirect Evidence for Seefeldt’s Proficiency Barrier in Young Adults? Res. Q. Exerc. Sport 2013, 84, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Sacko, R.S.; Nesbitt, D.; McIver, K.; Brian, A.; Bardid, F.; Stodden, D.F. Children’s Metabolic Expenditure during Object Projection Skill Performance: New Insight for Activity Intensity Relativity. J. Sports Sci. 2019, 37, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Sacko, R.S.; Utesch, T.; Bardid, F.; Stodden, D.F. The Impact of Motor Competence on Energy Expenditure during Object Control Skill Performance in Children and Young Adults. Braz. J. Mot. Behav. 2021. [Google Scholar] [CrossRef]
- Rudd, J.R.; Pesce, C.; Strafford, B.W.; Davids, K. Physical Literacy - A Journey of Individual Enrichment: An Ecological Dynamics Rationale for Enhancing Performance and Physical Activity in All. Front. Psychol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.A.; Pylyshyn, Z.W. Connectionism and Cognitive Architecture: A Critical Analysis. Cognition 1988, 28, 3–71. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S. Analogy, Cognitive Architecture and Universal Construction: A Tale of Two Systematicities. PLOS ONE 2014, 9, e89152. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.; Maselli, A.; Lancia, G.L.; Thiery, T.; Cisek, P.; Pezzulo, G. The Road towards Understanding Embodied Decisions. Neurosci. Biobehav. Rev. 2021, 131, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.B.M.; Hayden, B.Y.; Pearson, J.M. Continuous Decisions. Philos. Trans. R. Soc. B 2021, 376, 20190664. [Google Scholar] [CrossRef] [PubMed]
- Barca, L.; Pezzulo, G. Unfolding Visual Lexical Decision in Time. PloS One 2012, 7, e35932. [Google Scholar] [CrossRef] [PubMed]
- Resulaj, A.; Kiani, R.; Wolpert, D.M.; Shadlen, M.N. Changes of Mind in Decision-Making. Nature 2009, 461, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, I.; Davids, K.; Araújo, D.; Lucas, A.; Roberts, W.M.; Newcombe, D.J.; Franks, B. Evaluating Weaknesses of “Perceptual-Cognitive Training” and “Brain Training” Methods in Sport: An Ecological Dynamics Critique. Front. Psychol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-J.; Mercer, V.S. Dual-Task Methodology: Applications in Studies of Cognitive and Motor Performance in Adults and Children. Pediatr. Phys. Ther. Off. Publ. Sect. Pediatr. Am. Phys. Ther. Assoc. 2001, 13, 133–140. [Google Scholar]
- Walshe, E.A.; Patterson, M.R.; Commins, S.; Roche, R.A. Dual-Task and Electrophysiological Markers of Executive Cognitive Processing in Older Adult Gait and Fall-Risk. Front. Hum. Neurosci. 2015, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Hulteen, R.M.; Terlizzi, B.M.; Abrams, T.C.; Sacko, R.S.; De Meester, A.; Pesce, C.; Stodden, D.F. Reinvest to Assess: Advancing Approaches to Motor Competence Measurement Across the Lifespan. Sports Med. 2022. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, B.M.; Hulteen, R.M.; Rudd, J.; Sacko, R.S.; Sgrò, F.; Jaakkola, T.; Abrams, T.C.; Brian, A.; Nesbitt, D.; De Meester, A.; et al. A Pre-Longitudinal Screen of Performance in an Integrated Assessment of Throwing and Catching Competence. Phys. Educ. Sport Pedagogy 2024, 1–13. [Google Scholar] [CrossRef]
- Terlizzi, B.; Abrams, T.C.; Sacko, R.S.; Hand, A.F.; Silvey, K.; Stodden, D.F. The Relationship Between Functional Motor Competence and Performance on the Army Combat Fitness Test in Army Reserve Officer Training Corps Cadets. Mil. Med. 2022, 188, 1910–1917. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Yang, M.; Wymbs, N.F.; Grafton, S.T. Learning-Induced Autonomy of Sensorimotor Systems. Nat. Neurosci. 2015, 18, 744–751. [Google Scholar] [CrossRef]
Figure 1.
Conceptual model of potential motor-cognitive solutions.

Figure 2.
Comparison of potential motor-cognitive solutions between higher- and lower-skilled individuals.
Figure 2.
Comparison of potential motor-cognitive solutions between higher- and lower-skilled individuals.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



