
Submitted:
22 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
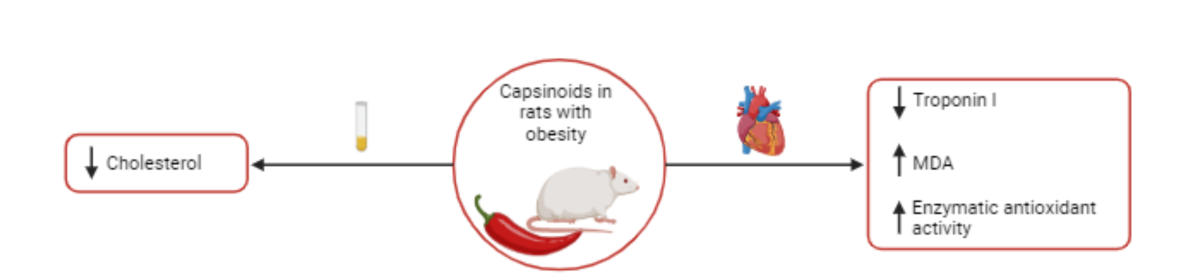
Capsinoids (CAP) are potential antioxidant agents capable of reducing oxidative damage and the resulting complications triggered by obesity. Thus, the aim of this study was to investigate the effects of CAP on adiposity and biomarkers of cardiac oxidative stress in rats with obesity induced by a high-fat diet. Male Wistar rats were exposed to a high-fat diet for 27 consecutive weeks. After characterization of obesity (week 19), some of the obese (Ob) animals began to receive capsinoids (10 mg/kg/day) by orogastric gavage. Adiposity and comorbidities were assessed. In the heart, remodeling, injury and biomarkers of oxidative stress were determinated. The treatment did not reduce the adiposity induced d by obesity, but they were efficient in reducing cholesterol levels. CAP treatment did not cause a difference in heart and LV mass, despite having reduced troponin I concentrations. Furthermore, capisinoids did not reduce the increase in advanced oxidation of proteins products (AOPP) and carbonylated proteins (CBO) caused by obesity in cardiac tissue. Besides that, it was observed that ObCap rats presented high levels of malondialdehyde and greater activity of antioxidant enzymes compared to Ob. In conclusion, treatment with capsinoids did not lead to significant positive effects in reducing obesity-induced oxidative stress.
Keywords:
capsinoids
; cardiac remodeling
; oxidative stress
; antioxidant capacity
; obesity
1. Introduction
Obesity, defined as excessive accumulation of body fat, is considered a risk factor for impairment of health and mortality in the population [1,2,3,4,5,6]. This disease affects around 810 million people and is related to more than 10 million deaths around the word, being considered an important global public health problem [5].
In obesity, the imbalance between energy consumption and expenditure promotes hypertrophy and hyperplasia of white adipose tissue (WAT)[7,8,9]. However, the stimulus of the expansion of these adipocytes drives the entry of cells from the immune system, influencing the construction of the inflammatory and dysfunctional profile of WAT [9]. These changes may induce the increase in morbidity biomarkers, such as increased reactive species, induction of insulin resistance and hyperleptinemia, decreased glucose tolerance, lipotoxicity, ectopic lipid deposition and inflammation [7,8,9].
As previously stated, obesity is related to an increase in reactive species (RE) in the organism, as superoxide radical (O2 -.), hydrogen peroxide (H2O2), hydroxyl radical (. OH), nitric oxide (NO) and peroxynitrite (ONOO-) [10]. Naturally, our body is capable of producing such species [11], which at controlled levels, they are related to the control of body homeostasis, and contribute to processes such as immunological defense, cellular proliferation and differentiation, defense against pathogens, signaling, among others [10,12,13]. However, some external sources, such as high-calorie diets, can induce its increase and exacerbation, triggering toxic and harmful effects on the body and the oxidative stress state [10,11].
The oxidative stress, is a state of imbalance between oxidants and antioxidants in favor of oxidants, and which leads to a dysregulation of signaling and redox control and/or molecular damage [14]. Studies highlight that excessive fat accumulation contributes to the release of pro-oxidant and proinflammatory in the organism [1,7]. In this way, overstimulation of RE production and release can cause changes in protein structures and functions, damage to nucleic acids and lipids, carcinogenesis and inflammation [10,11].
Specifically, regarding changes occurring in the heart, obesity causes a reduction in metabolic flexibility and a decrease in cardiac efficiency [15]. In this context, the increase in circulating free fatty acids can increase the mitochondrial uncoupling and reduce the efficiency of oxygen use from this substrate, causing deficiency in energy production and increased release of reactive species in cardiac tissue [15]. In turn, reactive species can lead to cardiac hypertrophy and dysfunction, becoming relevant for the development of myocardial dysfunction and consequent heart failure, among other cardiovascular complications [15,16].
In this context, as well as the high incidence of morbidity and mortality and the high cost of obesity treatment and cardiovascular diseases (CVDs), searching for alternatives for the treatment and prevention of these diseases becomes relevant [17,18] . Thus, literature has searched alternatives to the treatment and prevention of obesity and its complications. In this scenario, studies with capsinoids (CAP) have been used; the CAP consist of alkaloid molecules that are vanillic alcohol esters with fatty acids present in peppers of the Capsicum genus [19,20,21]. The CAP group has compounds such as capsiate, dihydrocapsiate and nordihydrocapsiate [19,21]. The CAP have demonstrate effects such as promotion of thermogenesis, stimulation of energy and lipid metabolism, promotion of body mass loss and reduction of adipose tissue. Furthermore, they can improve glucose tolerance, have antihyperlipidemic activity, and anti-inflammatory and antioxidant properties [19,21,22,23].
Thus, there is evidence that capsinoids may have effects on obesity and its consequences. However, the majority studies do not take into account the obesity associated with CVD. Therefore, considering the physiological property of CAP and the absence of studies on cardiac stress on obesity, the current study was performed to test the hypothesis that chronic administration of capsinoids positively modulates biomarkers of oxidative stress in the heart in the obesity, resulting in a reduction in products derived from oxidative damage and prevention of pathological cardiac remodeling.
2. Materials and Methods
Animal care and experimental protocol
After approval by the ethics committee on the use of animals at the Federal University of Espírito Santo, Brazil (08/2022), male Wistar rats (n = 44) were subjected to a protocol with 27 consecutive weeks, which it was divided into two moments: exposure to experimental diets per 19 weeks (obesity induction and maintenance), and period of treatment with capsinoids by 8 weeks. Initially, the rats were randomized into two groups: SD (standard diet) and HFD (high-fat diet). The standard diet was based on the AIN 93 recommendations, being composed of corn starch, casein, dextrinized starch, sucrose, soy oil, microcrystalline cellulose, mineral mix, vitamin mix, L-cystine, BHT and bitartrate choline (Pragsoluções biociências ®, São Paulo, Brazil). This diet was composed of 9.45% lipids, 75.66% carbohydrates and 14.89% proteins (Pragsoluções biociências ®, São Paulo, Brazil). For the composition of the high-fat diet, the same recommendations were followed, but with the addition of lard (Pragsoluções biociências ®, São Paulo, Brazil). The high-fat diet was composed of 45.33% lipids, 40.29% carbohydrates and 14.38% proteins (Pragsoluções biociências ®, São Paulo, Brazil).
During the 27-week protocol, 40 g of food and water ad libitum were offered daily. Energy density of each diet was also important to estimate daily caloric intake [24] and feed efficiency (FE) [FE = total body weight gain of the animals (g) / total energy ingested (kcal)] [25,26].
The characterization of obesity occurred when the HFD group showed a significant increase in body weigth (BW) in relation to the SD group. This moment was demonstrated in previous studies [27,28], 2020). After the initial moment of obesity (week 16), SD and HFD groups were renamed as control (C) and obese (Ob) groups, respectively (Figure 1). Afterwards, the rats were maintained in obesity maintenance for 4 weeks. Thus, at the end of the 19th week of the protocol, the rats were randomized into four different groups according to the absence and/or presence of capsinoid treatment: C (standard diet), CCap (standard diet with capsinoids), Ob (obese) and ObCap (obese with capsinoids)(Figure 1). However, considering that the aim of this study was only to evaluate the effects of treatment with capsinoids on obesity, the CCap rats were excluded from the current study and used in other studies of our laboratory (Figure 1).
Administration of Capsinoids
ObCap group was supplemented daily with capsinoids (CAP; Infinity Pharma, Brazil) by orogastric gavage (10 mg of capsinoids/kg of BW diluted in 1 ml of water/kg of body weight) for 8 weeks. The capsinoid dose was adjusted weekly according to the change in BW to maintain a constant CAP dose throughout the study. Similarly, groups C and Ob received vehicle gavage in the same amounts.
Euthanasia
At the end of the experimental protocol (27 weeks), following an 6-hour fasting period, the animals were heparinized and anesthetized with a solution containing ketamine hydrochloride (90 mg/kg) and xylazine hydrochloride (10 mg/kg)[29]. In cases were animals still exhibited signs of nociceptive reflex after anesthetic induction, an anesthetic overdose (lethal dose) was administered, consisting of three times the doses of ketamine hydrochloride and xylazine hydrochloride used during the animal’s anesthetic induction [29]. Following euthanasia, the animals submitted a median thoracotomy to collect blood and tissue samples.
Body weight and adiposity
To evaluate characterize the obesity in the rats, the body weight and fat, as well as adiposity index (AI) were analyzed. In addition, the evolution of body mass was measured weekly. Regarding the amount of body fat, it was determined through the sum of epididymal, retroperitoneal and visceral fat pads. Finally, the AI was calculated by the formula: [AI = amount of body fat / final BW x 100] [30,31].
Visceral adipose tissue morphology
For visceral adipose tissue morphometry, collected samples were fixed in 4% paraformaldehyde. They were then dehydrated in ethanol, clarified in xylene and embedded in paraffin. After inclusion, sections with a thickness of 5 μm were obtained using a rotating microtome (Leica ® RM 2125 RTS, Wetzar, Germany) and placed on slides for histology. Then, the histological sections were stained with hematoxylin and eosin (HE). For adipocyte area analysis, 10 fields from each adipose tissue slide were checked. The morphometric analysis was performed blindly. And the images were captured using a 10x objective lens, with the aid of a video camera (LAS EZ®, ICC50 HD – 51112061, Leica, Germany) coupled to an optical microscope (Leica®, RM 2125 RTS, Wetzar, Germany). The measurement was carried out using specific software (Image J Pro-Plus®, Media Cybernetics, Silver Spring, Maryland, USA).
Comorbidities Associated with Obesity
To evaluate the possible metabolic and hormonal changes induced by obesity, glucose tolerance test (GTT), homeostatic model assessment for insulin resistance (HOMA-IR) index, lipid and hormonal profiles were evaluated.
In the last week of the protocol (weel 27), the animals were subjected to a glucose tolerance test. They remained fasting for 6h, to analyze glycemic levels in basal conditions and after glucose overload (50% glucose; ip) at times 30, 60, 90 and 120 minutes [32]. The area under the curve for glucose (AUC) was also evaluated to identify presence or absence of tolerance to glucose.
Homeostatic model assessment for insulin resistance (HOMA-IR) index was used to determinate the insulin resistance, being calculated by following formula: [fasting insulin concentration (μU/mL) x fasting blood glucose (mmol/L) / 22.5] [33].
For lipid and hormonal profiles analysis, blood samples were collected in Falcon tubes and centrifuged at 10,00 rpm for 10 minutes and then stored at -80°C. The insulin (Elabscience Biotechnology, USA), leptin (R&D Systems, USA), adiponectin (Elabscience Biotechnology, USA) and glucagon (R&D Systems, USA) ware determined by enzyme-linked immunosorbent assay (ELISA) using specific kits. Plasmatic concentrations of triglycerides (TG), total cholesterol (TC), high-density lipoproteins (HDL) and low-density lipoproteins (LDL) was determined using specific kits (Bioclin®, Belo Horizonte, Brazil) and analyzed with the BS-200 automated biochemical apparatus.
Cardiac remodeling and injury
The cardiac remodeling process was evaluated by analyzing the total mass of the heart, left ventricle (LV) and their relationship with tibia length [34]. Cardiac injury was measured by troponin I concentration, determined using a specific ELISA kit (AFG Scientific, USA).
Analysis of the production of reactive species
The determination of ROS was carried out using the dihydroethidium (DHE) fluorescence method, which allows analyzing the “in situ” production of the superoxide radical. Transverse sections of the left ventricle (8 μm) were placed on gelatinized slides. Afterwards, the slides were washed and incubated with Krebs’s solution, in a light-protected (2 μM DHE) humidified chamber at 37ºC for 30 min, to detect superoxide. The intensity of fluorescence was detected at 585 nm, using a confocal fluorescence microscope (Leica DM 2500 TI, Nikon Instruments Inc., Melville, NY, USA). The fluorescence intensity was quantified using specific software (Image J Pro-Plus®, Media Cybernetics, Silver Spring, Maryland, USA). Analysis of 4 fields per sample were performed.
Biomarkes of cardiac oxidants and antioxidants
Fragments of LV were homogenized in 1:10 sodium phosphate buffer (SPB) and transferred to Eppendorf microcentrifuge tubes. Then, the samples were centrifuged at 3500 rpm at 4°C for 10 minutes.
The supernatant was used to measure oxidant biomarkers, such as malondialdehyde (MDA), carbonyl proteins (CBO) and advanced oxidation protein products (AOPP), and antioxidant biomarkers such as the antioxidant capacity (FRAP) and the activity of the enzymes catalase (CAT) and superoxide dismutase (SOD).
To evaluate the peroxidation of membrane lipids, the concentrations of malondialdehyde (MDA) was performed using the thiobarbituric acid reactive substances method (TBARS). For this purpose, the homogenates were centrifuged together with a solution containing thiobarbituric acid, trichloroacetic acid 2M, hydrochloric acid and distilled water for 10 min at 25°C. The supernatants were collected and boiled for 45 min. Then the reading was carried out at 532 nm and 600 nm. The MDA concentration was obtained taking into account the difference between the absorbances. The concentration of MDA was obtained through the molar extinction coefficient (1.56 × 105 M−1 cm−1) and the absorbances of the samples and the final result was expressed in nmol/g of protein.
The CBO were measured using the 2,4-dinitrophenylhydrazine (DNPH) derivatizer and spectrophotometric detection of proteins modified by the carbonylation process (MESQUITA et. al, 2014). To this end, solutions containing DNPH and HCL (2M) and another with NaOH and distilled water were prepared. For the analyses, the homogenate diluted in a solution containing DNPH and HCL (2M) was used. Afterwards, the samples were incubated for 10 minutes at room temperature, then NaOH solution (6 M) and distilled water were added. Another 10 minutes of incubation at room temperature and protected from light were necessary until reading at 450 nm in a microplate reader, with the result obtained through the following calculation: [(sample abs. / 22000) x 1000000] / abs. of the protein. AOPP were measured using a technique that uses potassium iodide (1,16 M) and glacial acetic acid, and a curve prepared with chloramine, with the reading at 340 nm.
The antioxidant capacity was measured using the FRAP technique (Ferric reducing antioxidant power), using the working reagent which contained acetate buffer, TPTZ/HCl solution, and ferric chloride. In a plate, the sample was added with the working solution. The reading was performed at 594 nm after 30 min of waiting.
The CAT activity was evaluated by following the decrease in the levels of hydrogen peroxide. For analysis, the samples were diluted in 10% Triton solution. This preparation was left on ice for 15 min to release the catalase. Then, 10 mM potassium phosphate buffer was added at room temperature. Then the medium with hydrogen peroxide was added. After 2 min, a reading was carried out at 240 nm for 3'30'', reading every 30 seconds.
To determine the activity of the SOD, in the sample wells, glycine and catalase buffer were added. Afterwards, a solution with adrenaline and HCL PA was added. Immediately afterwards, a reading was taken at 412 nm for 20 min, reading every 40 seconds.
Statistical analysis
Data distribution was assessed using the Kolmogorov-Smirnov normality test. The results were expressed as the mean ± standard deviation. All comparisons (SD vs. HFD, as well as C vs. Ob and Ob vs. ObCap) were performed using the student’s t test for independent samples. However, to analyze the evolution of body mass during the period of exposure to experimental diets, two-way ANOVA for independent samples was used, complemented with Bonferroni's post hoc test. During the capsinoid treatment period, two-way ANOVA for independent samples was used for body mass and GTT evaluation, complemented with Tukey's post hoc test. The significance level adopted was 5%. The statistical analyses and graphics were conducted using GraphPad Prism 9.0 software (GraphPad, San Diego, CA, USA).
3. Results
3.1. Exposure to Experimental Diets
The Figure 2 demonstrates the evolution of body mass during the period of exposure to experimental diets. As indicated, from the 16th week onwards, the HFD group (601 ± 71 g) presented a significantly higher (+ 8.5%) BW then SD group (554 ± 56 g), characterizing the initial moment of obesity. The period between the 16th and 19th week was called the period of obesity maintenance and the groups were renamed in groups C and Ob, respectively. At the end of this period (19th week), it was verified that the FBW was higher in the Ob group (606 ± 67 g) compared to C (551 ± 53 g), representing an elevation of 10%. However, there was no significant difference in body mass gain between the groups (C: -2.68 ± 14.13 g; Ob: 4.39 ± 17.60 g).
3.2. Exposure to Capsinoids Treatment
Considering the nutritional profile, during the chronic administration of capsinoides, the Ob group (15.1 ± 1.5 g/day) presented a reduction (- 22.2%) in food consumption, when compared to C group (19.4 ± 2.4 g/day), but this parameter was similar to ObCap group (15.3 ± 2.2 g/day). In addition, no significant differences were observed between Ob and ObCap in relation to caloric intake (Ob: 72.8 ± 7.3 kcal/day vs. ObCap: 73.6 ± 10.7 kcal/day) and feed efficiency (Ob: - 0.29 ± 0.96% vs. ObCap: 0.08 ± 0.91%), respectively.
Regarding the body weight at week 19, the results indicate that the initial body weight (IBW) was higher (+9%) in Ob rats than C without statistical difference in relation to ObCap (Figure 3A). Additionally, after 8 weeks of treatment with capsinoids, the FBW in the Ob group was similar in relation to C and ObCap, respectively, indicating that there was a stabilization of body mass gain in the animals.
Finally, the results showed that Ob rats presented high values for all parameters in relation to C, demonstrating the efficiency of high-fat diet in the development of obesity (Figure 3C-F). Nevertheless, the capsinoids treatment was not able to reduce and/or to prevent the accumulation of total fat and increase in fat pads in obesity, since ObCap animals presented higher values than Ob epidydimal fat (+ 15.5%), retroperitoneal fat (+ 34.5%) and total body fat (+23.5%) (Figure 3C, D and F). However, when total body fat values were normalized by body weight, the results showed that there was no difference between Ob and ObCap (Figure 3G).
Furthermore, in relation to visceral adipose tissue, no difference was observed in adipocity area among groups, revealing the existence of heterogeneous adipocytes with regard to their size in both groups (Figure 4).
Table 1 illustrates the lipid, glycemic and hormonal profiles after treatment with capsinoids. The results showed that Ob group had increased in cholesterol levels in relation to C group, representing an elevation of 69.3%. One important result is related to effect of capsinoids treatment that it was able to reduce this parameter (-18.8%) (ObCap < Ob, p< 0,05). In addition, there were no difference between Ob and the other groups in relation to triglycerides, HDL and LDL.
Regarding glycemic parameters, the results showed that the baseline glucose obtained from GTT was increased (15.7%) in ObCap rats in relation to the Ob, but the glycemic values after the administration of glucose overload were similar among groups (data not show). However, there were no alterations in the AUC for glucose among the groups (Table 1) between the groups. The insulin levels were elevated in Ob rats, when compared to C (+128%), however, the treatment with capsinoids did not prevent this increase (p > 0.05). The same behavior was observed in relation to the HOMA-IR index, in which the Ob group presented values 144% higher than C group, indicating an insulin resistance induced by obesity. However, the treatment with capsinoids was also not able to improve this behavior.
Regarding the hormonal profile, it was found that there was an increase in leptin levels in the Ob group (+60.5%) compared to C, but without statistical difference in relation to ObCap. Furthermore, there was no difference in glucagon and adiponectin levels among the groups (Tabe 1).
Table 2 summarizes the cardiac injury and remodeling after treatment with capsinoids. The results indicate that Ob group presented an increase in heart (+17.8%) and LV (+37.2%) in relation to the C group, respectively. However, after normalization by tibia length, this difference did not remain, with no difference between the groups. However, it's worth noting that capsinoids treatment was not able to prevent the cardiac remodeling induced by obesity.
Despite that, the results showed that the treatment with capsinoids in obesity was able to reduce the troponin I (-33%) in Obcap when compared to Ob group, indicating that the treatment triggered a reduction in this marker of cardiac injury (Table 2).
Furthermore, the production of superoxide radical in situ in LV was verified using the DHE as a marker. The results showed that there was no difference among the three groups for this parameter (Figure 5).
In addition, when the cardiac biomarkers of oxidative stress and antioxidant enzymes where checked (Figure 6), the data show that for there was no difference between Ob and C for MDA, a marker of lipid peroxidation. However, MDA levels in ObCap rats were higher than Ob group (+76.9%), indicating that the treatment had a negative effect on lipid peroxidation in the animal’s hearts (Figure 5A). In addition to this finding, the levels of carbonyl proteins (CBO) and advanced oxidation protein products (AOPP) were increased in Ob group in relation to C group (+265% and +76.8%, respectively); however, no difference were observed, when compared to ObCap (Figure 6B,C). In addition, a statistical difference in antioxidant capacity (FRAP) was observed between C and Ob, but there was no difference between Ob groups (Figure 6D). These findings highlight that treatment with capsinoids was no efficient in reversing oxidative changes and damage triggered by obesity.
Finally, it was demonstrated that the SOD and CAT were elevated in Obcap than Ob group (+62.5% and +636.4%, respectively), but no differences were observed between groups C and Ob. In the other words, the stimulus for SOD and CAT activity in both groups were reduced ((Figure 6E,F).
4. Discussion
The aim of this study was to investigate the effects of capsinoid treatment on biomarkers of oxidative stress in the hearts of rats with obesity induced by high-fat diet. The main findings were that chronic treatment with capsinoids in obesity was not able to reduce body weight and to prevent excessive accumulation of adiposity, as well as significantly to reversing the metabolic changes. Specifically in the heart, it was observed that treatment with capsinoids was able to prevent the cardiac injury and to elevate the antioxidant enzymes in the heart. In addition, the chronic administration of capsinoids was unable to reverse the oxidative stress induced by obesity. Furthermore, the treatment with capsinoids promoted greater lipid peroxidation in the hearts of animals belonging to the ObCap group.
Experimental models of obesity have been extensively used in research to study the response to various forms of the disease [35]. In experiments where obesity is induced by a high-fat diet, progression to the obese phenotype tends to be a process that occurs slowly, just as it occurs in humans [35]. The literature indicates that diets that provide 30-60% of total calories from fat favor the development of obesity and associated comorbidities [36,37]. The increase in body mass can occur from 16-20 weeks after starting to use the diet, with an increase of 20-30% in body mass being possible in relation to a control group [35]. In the current study, obesity was characterized from the 16th week onwards, followed by an increase of 11.8% in body weight in HFD group when compared to SD group. Previous studies from our laboratory have already shown a body weight increased when using a high-fat diet for 20 weeks [37,38]. However, research with shorter intervention time (8 weeks) also managed to observe the efficiency of different high-fat diets in increasing body mass [38,39].
Considering body fat pads, our results demonstrate that the high-fat diet induced an increase in epididymal, visceral, retroperitoneal fat and total body fat in relation to the group that received a standard diet, characterizing obesity. In this context, capsinoids (CAP) emerging as compounds with potential action for controlling and reducing obesity [3,4,40,41,42]. This is because the compounds in this group are capable of causing an increase in energy expenditure, increased expression of proteins related to lipid metabolism, increased oxygen consumption, reduced body mass and fat deposits [21,41,43]. Despite that, in the current study, the treatment with capsinoids was not efficient in reducing the obesity induced by obesity, on the contrary, these values were increased. However, there was no alteration in adiposity index. These results indicate the inefficiency of the treatment in reducing the obesity phenotype, which suggests that this effect may be related to the increase in circulating leptin in the treated group [44]. Despite the increase in adiposity, when the morphology of visceral adipose tissue was verified, there was no difference in the adipocyte area between the groups, suggesting that the increase in adipose mass was driven by adipocyte hyperplasia instead than hypertrophy.
Furthermore, taking into account that the model used was effective in inducing increased adiposity in animals, it becomes relevant to understand the biochemical and physiological disorders related to the disease. In this context, when there is a positive energy balance and high consumption of saturated fatty acids or simple carbohydrates, there may be a greater release of lipoproteins and molecules carrying triglycerides and cholesterol into the circulatory system [45]. In a previous study carried out with enzymatically synthesized capsiate and dihydrocapsiate, the effect of the treatment was found to reduce serum cholesterol and serum lipids in animals [46]. In line with this, in this research, although obesity only induced isolated hypercholesterolemia in animals, a beneficial effect of capsinoids was observed to reverse this condition. In this context, a possible explanation for this effect would be that capsinoids may have similar effects to their capsaisinoid analogues, which are capable of inducing a reduction in HMG-CoA reductase mRNA and increasing CYP7A1 mRNA in the liver, decreasing cholesterol synthesis and increasing their conversion to bile acids, respectively [47]. However, specific studies on the effects of capsinoids on lipid metabolism are necessary to confirm this assumption.
Additionally, the oxidation of lipids as a substrate is stimulated in obesity, however this oxidation ends up being smaller than the amount of lipids that enter the adipose cell, resulting in an increase in adipocytes [48]. This effect causes hypoxia in adipose tissue and triggers changes in the production, release and activity of hormones secreted by adipose tissue [37,48]. For example, the release of leptin becomes highly stimulated, leading to a state of leptin resistance, altering the regulation of appetite and energy expenditure [8,9,48]. Furthermore, adiponectin secretion tends to be reduced, triggering effects such as increased inflammation and blood triglyceride levels [8,45,49]. In disagreement, in the current study was not observed leptin resistance induced by obesity. In addition, the diet and/or exposure time were not efficient in causing changes in serum adiponectin levels. Furthermore, no benefit was observed in reducing leptin and improving these hormonal parameters when animals were subjected to treatment with capsinoids. One of the reasons why capsinoids did not change the pattern of secretion of these hormones is that the treatment did not induce a reduction in body fat mass, so that the increase in tissue can continue to stimulate the distorted secretion of leptin and adiponectin [44]. Furthermore, the increased insulin levels in the group treated with capsinoids may have had an effect on the greater expression and release of leptin [44].
Furthermore, it is worth noting that obesity can trigger direct or indirect changes in cardiac function [50,51]. In this context, increased body adiposity induces hemodynamic changes in the cardiovascular system, such as increased blood volume and cardiac output and reduced vascular resistance [51]. As a consequence, there is an increase in volume and pressure overload, which often triggers concentric hypertrophy of the LV, dilation and degradation of the tissue matrix and cardiac fibrosis, leading to heart dysfunction and failure [50,51]. Additionally, visceral obesity plays an important role in cardiac changes, as it can induce cardiomyocyte hypertrophy, leading to myocardial fibrosis and the activation of inflammatory pathways in the tissue [51]. Besides that, the ectopic accumulation of lipids, in addition to increasing circulatory demand, also induces the release of pro-atherogenic factors, and can cause stress on the heart wall and lead to myocardial injury, triggering concentric hypertrophy and LV remodeling, and failure cardiac [51,52].
Considering this context, it was expected that the hearts of rats with obesity would present hypertrophy. However, although the hearts and LV of obese animals showed an increase in mass, when the values were normalized by the tibia lenght, no difference was observed. Given these consequences, treatment with capsinoids also did not produce any effect in reducing the mass of the heart or ventricle. These effects may be due to the inefficiency of the diet or the time to induce comorbidities associated with obesity, even though an increase in adiposity, plasma cholesterol and insulin resistance have been observed in Ob animals. Gasparini (2021) [53] also did not observe changes in the mass of the heart and ventricle in obese rats after 20 weeks of obesity induction, even though they used a high-fat diet different from that used in this study. However, Cavalera, Wang and Frangogiannis (2014) [50], indicate that genetic models of obesity may be more effective in inducing fibrosis and cardiac hypertrophy. The authors also mention that in models of obesity induced by high-fat diets, the establishment of cardiac fibrosis may be late [50]. Furthermore, the severity of these changes may depend on variables of the obesity model, such as species, age of the animals, mechanism of obesity and the presence of other pathophysiological conditions [50].
Considering the effects of capsinoids on cardiac remodeling, Wang et al. (2014) [54] previously demonstrated that TRPV1 can mediate the hypertrophic response triggered by pressure overload in the heart, and that the anti-hypertrophic action of its analog capsaicin depends precisely on the presence of TRPV1 channels. This effect appears to be mediated by the induction of PPAR-δ expression and inhibition of NF-κB [55]. In this regard, despite the first being more potent, it is known that both capsaicin and the capsinoid group are TRPV1 receptor agonists [19,21,23]. However, unlike capsaicin, capsinoids are quickly hydrolyzed in the gastrointestinal tract, possibly hindering the subsequent activation of TRPV1 receptors present in other tissues, such as the heart [20,56]. Thus, it is worth noting that the effects of TRPV1 activation on cardiac hypertrophy have not yet been completely established, and more studies are needed to understand this issue [54].
Linked to the changes mentioned above, in heart failure it is also possible to observe an increase of cardiac troponin, a marker of myocardial injury that can be increased as a consequence of myocardial stretching resulting from volume overload, subendocardial ischemia, elevated intracardiac pressure and/or or arrhythmias [57]. Therefore, in the current study, this marker was also evaluated. However, in disagreement with our initial hypothesis, the model of obesity was not efficient in inducing myocardial injury. However, the treatment with capsinoids was effective and prevented the elevation of this marker. Horiuchi et al. (2023) [58] previously identified that in humans, higher Body-mass-index (BMI) was associated with lower troponin levels compared to overweight or normal weight people, however, the reasons behind this effect are still poorly understood. It is worth mentioning that in our study, although BMI was not checked in the animals, the obese group treated with capsinoids showed increased values of body adiposity, which could be a possible explanation for the low troponin I levels in the treated group. Related to reduction of troponin I by CAP, it has already been demonstrated that TRPV1 knockout mice had higher plasma troponin concentrations after myocardial infarction compared to animals without TRPV1 silenced [59]. In this way, it is suggested that the protective effect of capsinoids on cardiac injury is possibly related to the expression and activation of TRPV1 in the heart.
In addition, several studies have demonstrated that there is an increase in oxidative stress induced by obesity on heart [15,16,60]. Such as, in obesity and associated comorbidities, greater production of hydrogen peroxide and superoxide in cardiac cells has been observed [52]. Also, a reduction in the expression and/or activity of antioxidant enzymes in the heart and circulation of obese people is commonly observed [15]. In this context, systemic changes occurring in obesity such as hyperglycemia, insulin resistance, hyperlipidemia, activation of the sympathetic nervous system and the renin-angiotensin-aldosterone system are important in increasing the generation of reactive species in the myocardium [16,50,52]. Furthermore, local changes such as increased oxidation of fatty acids as an energy substrate, mitochondrial dysfunction, glucose autoxidation, cardiac tissue stretching and hypoxia also tend to increase the release of reactive species in the heart [15,16,52]. These effects cause consequences in the structure, size, and in cardiac function constituting a damaging process for the myocardium [16,52,60].
Regarding the oxidative damage induced by obesity and the potential treatment with capsinoids and their analogues, it was demonstrated that capsiate and its analogues can promote protection against lipid oxidation [61]. It was also found that the activation of TRPV1 by its agonists can increase the expression of uncoupling protein 2 (UCP2), important for reducing the generation of reactive species in the electron transport chain in mitochondria [62]. In addition, TRPV1 can exert a protective effect on the myocardium, by being able to reduce markers of endogenous peroxynitrite formation [55].
In relation to oxidative biomarkers, the literature indicates that direct measurement of reactive oxygen species is very difficult due to the low concentrations and short lifetime of the species. However, fluorescent indicators such as dihydroethide (DHE), which produces fluorescence when reacting with superoxide anium, can be used for this purpose [63]. In our study, however, no increase in superoxide levels was observed in obese animals and no changes were observed caused by treatment with capsinoids. This result may be due to the rapid conversion of superoxide to hydrogen peroxide inside the cells, which has a longer life cycle and permeates cell environments better, being capable of causing persistent cellular changes, such as lipid and membrane oxidation and protein oxidation [60,63,64].
In this context, the current study investigated the effect of capsinoids treatment on biomarkers of cardiac oxidative stress. Our results indicate the inefficiency of the capsinoids treatment in reducing carbonyl proteins and advanced oxidation protein products caused by obesity. Some reasons for these effects may be related for these findings, including the dosage, exposure to treatment and and vehicle. Furthermore, it may be that the antioxidant portion of the capsinoid molecule (suggested to be the phenol portion) is not targeted or does not have easy access to ROS binding sites in the myocardium [16]. An important finding, different from what was expected, was that treatment with capsinoids caused greater lipid peroxidation in the hearts of animals with obesity, observed by the increase in MDA. This result suggests that compensatory mechanisms related to chronic antioxidant exposure in ROS-producing systems [16] may be involved. We also suggest that somehow the treatment, or even the activation of the TRPV1 receptor in the heart, has induced an increase in enzymatic lipid peroxidation, in which enzymes such as lipoxygenases (LOX), cyclooxygenases (COX) and cytochrome P450 (CYPs) synthesize molecules capable of trigger peroxidation [64].
Furthermore, our organism has antioxidant systems that work to maintain the balance of these molecules and reactions [16,60]. For example, reactive species are neutralized by intracellular antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT) [60]. SOD constitutes a group of metalloenzymes that dismutate superoxide into hydrogen peroxide [64,65], being important for maintaining an adequate flow of hydrogen peroxide [65]. While CAT constitutes a group of enzymes responsible for the decomposition of peroxide [16,64].
Our results showed that the antioxidant capacity carried out by FRAP was elevated in obesity, without differences observed with CAP treatment. This finding indicates that oxidative alterations induced by obesity somehow provoked the antioxidant response to counterbalance the damage. Furthermore, the increase in biomarkers in the obesity was not accompanied by an elevation in activity of antioxidant enzymes, which was already expected. However, it is worth highlighting that treatment with capsinoids promoted an improvement in antioxidant activity visualized by the increased in SOD and CAT activities, respectively. One possible explanation is that this result may not necessarily be an isolated protective effect of capsinoids on heart, but may be a consequence of increased lipid peroxidation. In this sense, the literature reports that lipid peroxidation products are capable of inducing the transcription factor Nrf2, important for regulating the expression of antioxidant enzymes (NIEMANN et al., 2017), which could promote protective feedback against oxidative damage itself.
5. Conclusions
The treatment with capsinoids does not prevent the process of cardiac remodeling and does not lead to positive modulation of oxidative stress induced by obesity. However, treatment with capsinoids causes a reduction in troponin I and induces antioxidant activity in cardiac tissue.
Author Contributions
KCCS, DSB, APLL and ASL conceived the design of the study. KKCS, LFD, FMN, LMS, ERC, FMF and CRC performed the experiments. KCCS, APLL and ASL interpreted, discussed the data, wrote the manuscript and statistical analysis. All authors revised the manuscript and the final version of the manuscript.
Funding
This study was supported by Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (grant numbers: 2023-MHRH6 and 2022-13H7D).
Institutional Review Board Statement
This study was approved by the Animal Use Ethics Committee of the Federal University of Espírito Santo under protocol 08/2022; approved on November 4, 2022.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We are grateful to Raissa Corrêa de Andrade, to RedOx e Inflammation Laboratory at UNESP-Botucatu (Brazil), and the LECTA, LHT and LHMI laboratories at the Federal University of Espírito Santo (Brazil) for technical assistance. To Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (FAPES, Brazil) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil) for assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Čolak, E.; Pap, D. The role of oxidative stress in the development of obesity and obesity-related metabolic disorders. J. Med. Biochem. .40. [CrossRef]
- Hawkes, C.; Fanzo, J.; Udomkesmalee, E. Nourishing the SDGs: Global Nutrition Report 2017. Bristol: Development Initiatives Poverty Research Ltd. 2017.
- Mosqueda-Solís, A.; Sánchez, J.; Reynés, B.; Palou, M.; Portillo, M.P.; Palou, A.; Picó, C. Hesperidin and capsaicin, but not the combination, prevent hepatic steatosis and other metabolic syndrome-related alterations in western diet-fed rats. Sci. Rep. 1510; .8. [Google Scholar] [CrossRef]
- Zheng, J.; Zheng, S.; Feng, Q.; Zhang, Q.; Xiao, X. Dietary capsaicin and its anti-obesity potency: from mechanism to clinical implications. Biosci. Rep. 2017; .37. [Google Scholar] [CrossRef]
- World Obesity Federation (WOF). World obesity atlas 2024. Disponível em: https://data.worldobesity.org/publications/?cat=22. (Accessed in March, 2024).
- World Health Organization (WHO). Obesity and overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed in May, 2023).
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, Oxidative Stress, and Obesity. Int. J. Mol. Sci. 3117; .12. [Google Scholar] [CrossRef]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Cell Physiol. .3. [CrossRef]
- Koenen, M.; Hill, M.A.; Cohen, P.; Sowers, J.R. Obesity, adipose tissue and vascular dysfunction. Circ. Res. 7. [CrossRef]
- Salehi, B.; Martorell, M.; Arbiser, J.L.; Sureda, A.; Martins, N.; Maurya, P.K.; Sharifi-Rad, M.; Kumar, P.; Sharifi-Rad, J. Antioxidants: positive or negative actors? Biomolecules. .8. [CrossRef]
- Martelli, F.; Nunes, F.M.F. Radicais livres: em busca do equilíbrio. Ci. e Cult, .66. [CrossRef]
- Ferreira, L.G.; Lunz, W. Tópicos em fisiologia e bioquímica com ênfase no exercício e treinamento físico, 1st ed.; Editora da Universidade Federal do Espírito Santo: Vitória, Brazil, 2022; pp. 53–67. [Google Scholar]
- Prasad, S.; Srivastava, S.K. Oxidative Stress and Cancer: chemopreventive and therapeutic role of triphala. Antioxidants, .9. [CrossRef]
- Jones, D.P. Redefining Oxidative Stress. Antioxid. Redox Signal. 1865; .8. [Google Scholar] [CrossRef]
- Niemann, B.; Rohrbach, S.; Miller, M.R.; Newby, D.E.; Fuster, V.; Kovacic, J.C. Oxidative Stress and Cardiovascular Risk: obesity, diabetes, smoking, and pollution: Part 3 of a 3-Part Series. J. Am. Coll. Cardiol. .70. [CrossRef]
- Münzel, T.; Camici, G.G.; Maack, C.; Bonetti, N.R.; Fuster, V.; Kovacic, J.C. Impact of Oxidative Stress on the Heart and Vasculature: Part 2 of a 3-Part Series. J. Am. Coll. Cardiol. 70. [CrossRef]
- Cote, B.; Elbarbry, F.; Bui, F.; Su, J.W.; Seo, K.; Nguyen, A.; Lee, M.; Rao, D.A. Mechanistic Basis for the Role of Phytochemicals in Inflammation-Associated Chronic Diseases. Molecules, .27. [CrossRef]
- Tran, N.; Pham, B.; LE, L. Bioactive Compounds in Anti-Diabetic Plants: from herbal medicine to modern drug discovery. Biology, .9. [CrossRef]
- Cansian, A.C.C. Efeitos da ingestão de capsinóides sobre a adiposidade corporal em ratos Wistar. Dissertação (Mestrado em Clínica Médica) - Programa de Pós-Graduação em Clínica Médica, Universidade de São Paulo, Ribeirão Preto, 2016.
- Gupta, R.; Kapoor, B.; Gulati, M.; Kumar, B.; Gupta, M.; Singh, S.K.; Awasthi, A. Sweet pepper and its principle constituent capsiate: functional properties and health benefits. Crit. Rev. Food Sci. Nutr, 7370; .62. [Google Scholar] [CrossRef]
- Snitker, S.; Fujishima, Y.; Shen, H.; Ott, S.; Pi-Sunyer, X.; Furuhata, Y.; Sato, H.; Takahashi, M. Effects of novel capsinoid treatment of fatness and energy metabolism in humans: possible pharmacogenetic implications. Am. J. Clin. Nutr, 89. [CrossRef]
- Galgani, J.E.; Ryan, D.H.; Ravussin, E. Effect of capsinoids on energy metabolism in humans. Br. J. Nutr. .1. [CrossRef]
- Sasahara, I.; Furuhata, Y.; Iwasaki, Y.; Inue, N.; Sato, H.; Watanabe, T.; Takahashi, M. Assessment of the biological similarity of three capsaicin analogs (Capsinoids) found in non-pungent Chilli Pepper (CH-Sweet) fruits. Biosc. Biotechnol. Biochem, .74. [CrossRef]
- Koch, C.E.; Lowe, C.; Pretz, D.; Steger, J.; Williamns, L.M.; Tups, A. High-fat diet induces leptin resistance in leptin-deficient mice. J. Neuroendocrinol, .26. [CrossRef]
- Irving, B.A.; Weltman, J.Y.; Patrie, J.T.; Davis, C.K.; Brock, D.W.; Swift, D.; Barret, E.J.; Gaesser, G.A.; Weltman, A. Effects of exercise training intensity on nocturnal growth hormone secretion in obese adults with the metabolic syndrome. J. Clin. Endocrinol. Metab. 1979; .94. [Google Scholar] [CrossRef]
- Kim, C.H.; Youn, J.H.; Park, J.Y.; Hong, S.K.; Park, K.S.; Park, S.W.; Suh, K.I.; Lee, K.U. Effects of high-fat diet and exercise training on intracellular glucose metabolism in rats. Am. J. Physiol. Endocrinol. Metab. 6. [CrossRef]
- Kobi, J.B.B.S.; Matias, A.M.; Gasparini, P.V.F.; Torezani-Sales, S.T.; Madureira, A.R.; Silva, D.S.; Corrêa, C.R.; Garcia, J.L.; Haese, D.; Nogueira, B.V.; Assis, A.L.E.M.; Lima-Leopoldo, A.P. ; Leopoldo A,S. High-fat, high-sucrose, and combined high-fat/high-sucrose diets effects in oxidative stress and inflammation in male rats under presence or absence of obesity. Physiol. Rep. e: v.11, n.7, 1563; .11. [Google Scholar] [CrossRef]
- Matias, A.M.; Coelho, P.M.; Marques, V.B.; Santos, L.; Assis, A.L.E.M.; Nogueira, B.V.; Lima-Leopoldo, A.P.; Leopoldo, A.S. Hypercaloric diet models do not develop heart failure, but the excess sucrose promotes contractility dysfunction. Plos One, 0228; .15. [Google Scholar] [CrossRef]
- Universidade Federal de São Paulo (UNIFESP). Guia de eutanásia para animais de ensino e pesquisa, 2019.
- Rolls, B.L.; Shide, D.J. The influence of dietary fat on food intake and body weight. Nutr. Rev. 50. [CrossRef]
- Taylor, B.A.; Phillips, S.J. Detection of obesity QTLs on mouse chromosomes 1 and 7 by selective DNA pooling. Genomics, .34. [CrossRef]
- Mendes, B.F.; Costa-Pereira, L.V.; Andrade, J.A.; Magalhães, C.O.D.; Pereira, R.R.S.; Esteves, E.A.; Cassilhas, R.C.; Andrade, E.F.; Gripp, F.; Magalhães, F.C.; Sampaio, K.H.; Amorim, F.T.; Dias-Peixoto, M.F. Superior cardiometabolic and cellular adaptive responses to multiple versus single daily sessions of high-intensity interval training in Wistar rats. Sci. Rep. 2118; 12. [Google Scholar] [CrossRef]
- Sociedade Brasileira de Diabetes (SBD). Diretrizes da Sociedade Brasileira de Diabetes 2019-2020. Clannad Editora Científica, 2019.
- Yin, F.C.; Spurgeon, H.A.; Rakusan, K.; Weisfeldt, M.L.; Lakatta, E.G. Use of tibial length to quantify cardiac hypertrophy: application in the aging rat. Am. J. Physiol. Heart Circ. Physiol, 6. [CrossRef]
- Doulberis, M.; Papaefthymiou, A.; Polyzos, S.A.; Katsinelos, P.; Grigoriadis, N.; Srivastava, D.S.; Kountouras, J. Rodent models of obesity. Minerva Endocrinol, 2020; .45. [Google Scholar] [CrossRef]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev.. 2010; v. 23, n.2, p. 270-299. [CrossRef]
- Moura e Dias, M.; Reis, S.A.; Conceição, L.L.; Sediyama, C.M.N.O.; Pereira, S.S.; Oliveira, L.L.; Peluzio, M.C.G.; Martinez, J.A.; Milagro, F.I. Diet-induced obesity in animal models: points to consider and influence on metabolic markers. Diabetol. Metab. Syndr. .13. [CrossRef]
- Feriani, A.; Bizzarri, M.; Tir, M.; Aldawood, N.; Alobaid, H.; Allagui, M.S.; Dahmash, W.; Tlili, N.; Alwasel, S.; Harrath, A.H. High-fat diet-induced aggravation of cardiovascular impairment in permethrin-treated Wistar rats. Exotoxicol. Environ. Saf. 1124. [Google Scholar] [CrossRef]
- Michicoti-Meneses, M.M.; Thompson-Bonilla, M.R.; Reyes-López, C.A.; García-Pérez, B.E.; López-Tenorio, I.I.; Ordaz-Pichardo, C.; Jaramillo-Flores, M.E. Inflammation Markers in Adipose Tissue and Cardiovascular Risk Reduction by Pomegranate Juice in Obesity Induced by a Hypercaloric Diet in Wistar Rats. Nutrients, 2577; 13. [Google Scholar] [CrossRef]
- Chapa-Oliver, A.; Mejía-Teniente, L. Capsaicin: from plants to a cancer-suppressing agent. Molecules, 21. [CrossRef]
- Hong, Q.; Xia, C.; Xiangying, H.; Quan, Y. Capsinoids supress fat accumulation via lipid metabolism. Mol. Med. Rep. 1669; 11. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, S.; Li, D.; Zhang, Y.; Tang, B.; Qiu, C.; Yang, Y.; Yang, D. Dietary Capsaicin Ameliorates Pressure Overload-Induced Cardiac Hypertrophy and Fibrosis Through the Transient Receptor Potential Vanilloid Type 1. Am. J. Hypertens. 1521; .27. [Google Scholar] [CrossRef]
- Ohnuki, K.; Haramizu, S.; Oki, K.; Watanabe, T.; Yazawa, S.; Fushiki, T. Administration of Capsiate, a Non-Pungent Capsaicin Analog, Promotes Energy Metabolism and Suppresses Body Fat Accumulation in Mice. Biosci. Biothecnol. Biochem. 2735; .65. [Google Scholar] [CrossRef]
- Vilariño-García, T.; Polonio-González, M.L.; Pérez-Pérez, A.; Ribalta, J.; Arrieta, F.; Aguilar, M.; Obaya, J.C.; Gimeno-Orna, J.A.; Iglesias, P.; Navarro, J.; Durán, S.; Pedro-Botet, J.; Sánchez-Margalet, V. Role of Leptin in Obesity, Cardiovascular Disease, and Type 2 Diabetes. Int. J. Mol. Sci, 2338; .25. [Google Scholar] [CrossRef]
- Bays, H.E.; Kirkpatrick, C.F.; Maki, K.C.; Toth, P.P.; Morgan, R.T.; Tondt, J.; Christensen, S.M.; Dixon, D.L.; Jacobson, T.A. Obesity, Dyslipidemia, and Cardiovascular Disease: A joint expert review from the Obesity Medicine Association and the National Lipid Association 2024. J. Clin. Lipidol, .18. [CrossRef]
- Tani, Y.; Fujioka, T.; Sumioka, M.; Furuichi, Y.; Hamada, H.; Watanabe, T. Effects of capsinoid on serum and liver lipids in hyperlipidemic rats. J. Nutr. Sci. Vitaminol, .50. [CrossRef]
- Zhang, L.; Fang, G.; Zheng, L.; Chen, Z.; Liu, X. The hypocholesterolemic effect of capsaicinoids in ovariectomized rats fed with a cholesterol-free diet was mediated by inhibition of hepatic cholesterol synthesis. Food Funct. 4. [CrossRef]
- Pereira-Lancha, L.O.; Campos-Ferraz, P.L.; Lancha Junior, A.H. Obesity: considerations about etiology, metabolism, and the use of experimental models. Diabetes Metab. Syndr. Obes. .5. [CrossRef]
- Han, Y.; Sun, Q.; Chen, W.; Gao, Y.; Ye, J.; Chen, Y.; Wang, T.; Gao, L.; Liu, Y.; Yang, Y. New advances of adiponectin in regulating obesity and related metabolic syndromes. J. Pharm. Anal, 1009; .14. [Google Scholar] [CrossRef]
- Cavalera, M.; Wang, J.; Frangogiannis, N.G. Obesity, metabolic dysfunction and cardiac fibrosis: pathophysiologic pathways, molecular mechanisms and therapeutic opportunities. Transl. Res. .4. [CrossRef]
- Powell-Wiley, T.M.; Poirier, P.; Burke, L.E.; Després, J.P.; Gordon-Larsen, P.; Lavie, C.J.; Lear, S.A.; Ndumele, C.E.; Neeland, I.J.; Sanders, P.; St-Onge, M.P. ; American Heart Association Council on Lifestyle and Cardiometabolic Health; Council on Cardiovascular and Stroke Nursing; Council on Clinical Cardiology; Council on Epidemiology and Prevention; Stroke Council. Obesity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation. 1010; .21. [Google Scholar] [CrossRef]
- Ilkun, O.; Boudina, S. Cardiac dysfunction and oxidative stress in the metabolic syndrome: an update on antioxidant therapies. Curr. Pharm. Des, 4806; .19. [Google Scholar] [CrossRef]
- Gasparini, P.V.F.; Matias, A.M.; Torezani-Sales, S.; Kobi, J.B.B.S.; Siqueira, J.S.; Corrêa, C.R.; Leopoldo, A.P.L.; Leopoldo, A.S. High-Fat and Combined High-Fat and Sucrose Diets Promote Cardiac Oxidative Stress Independent of Nox2 Redox Regulation and Obesity in Rats. Cell. Physiol. Biochem, .55. [CrossRef]
- Wang, Q.; Ma, S.; Li, D.; Zhang, Y.; Tang, B.; Qiu, C.; Yang, Y.; Yang, D. Dietary Capsaicin Ameliorates Pressure Overload-Induced Cardiac Hypertrophy and Fibrosis Through the Transient Receptor Potential Vanilloid Type 1. Am. J. Hypertens. 1521; .27. [Google Scholar] [CrossRef]
- Gao, F.; Liang, Y.; Lu, Z.; Li, L.; Zhu, S.; Liu, D.; Yan, Z.; Zhu, Z. TRPV1 Activation Attenuates High-Salt Diet-Induced Cardiac Hypertrophy and Fibrosis through PPAR-δ Upregulation. PPAR Res, 2014. [Google Scholar] [CrossRef]
- Ohyama, K.; Suzuky, K. Dihydrocapsiate improved age-associated impairments in mice by increasing energy expenditure. Am. J. Physiol. Endocrinol. Metab, .5. [CrossRef]
- Bhardwaj, A.; Baran, D.A. Biomarkers in HF: How Does the “Weight” Weigh in? J. Card. Fail. 1132; .29. [Google Scholar] [CrossRef]
- Horiuchi, Y.; Wettersten, N.; Vanveldhuisen, D.J.; Mueller, C.; Nowak, R.; Hogan, C.; Contas, M.C.; Cannon, C.M.; Birkhahn, R.; Vilke, G.M.; Mahon, N.; Nuñez, J.; Briguori, C.; Duff, S.; Murray, P.T.; Maisel, A. The Influence of Body Mass Index on Clinical Interpretation of Established and Novel Biomarkers in Acute Heart Failure. J. Card. Fail, 1121; .29. [Google Scholar] [CrossRef]
- Huang, W.; Rubistein, J.; Prieto, A.R.; Thang, L.V.; Wang, D.H. TRPV1 gene deletion exacerbates inflammation and atypical cardiac remodeling after myocardial infarction. Hypertension, .53. [CrossRef]
- Panth, N.; Paudel, K.R.; Parajuli, K. Reactive Oxygen Species: a key hallmark of cardiovascular disease. Advances In Medicine, 2016. [Google Scholar] [CrossRef]
- Rosa, A.; Deiana, M.; Corona, G.; Atzeri, A.; Incani, A.; Appendino, G.; Dessi, M.A. Protective effect of capsinoid on lipid peroxidation in rat tissues induced by Fe-NTA. Free Radic. Res. 1155; 39. [Google Scholar] [CrossRef]
- McCarty, M.F.; Dinicolantonio, J.J.; O'keefe, J.H. Capsaicin may have important potential for promoting vascular and metabolic health. Open Heart, 0002; .2. [Google Scholar] [CrossRef]
- Näpänkangas, J.P.; Liimatta, E.V.; Joensuu, P.; Bergmann, U.; Ylitalo, K.; Hassinen, I.E. Superoxide production during ischemia–reperfusion in the perfused rat heart: A comparison of two methods of measurement. J. Mol. Cell. Cardiol. 53. [CrossRef]
- Demirci-Çekiç, S.; Özkan, G.; Avan, C.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 1144. [Google Scholar] [CrossRef]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of oxidative stress in metabolic syndrome and associated diseases. Oxid. Med. Cell. Longev, 2019. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of experimental protocol (27 weeks). SD: standard diet (n = 17); HFD: high-fat diet (n = 27); C: control (n = 11) ; CCap: standard diet with capsinoids (n = 6); Ob: obese (n = 13); ObCap: obese with capsinoids (n = 14).
Figure 1.
Schematic representation of experimental protocol (27 weeks). SD: standard diet (n = 17); HFD: high-fat diet (n = 27); C: control (n = 11) ; CCap: standard diet with capsinoids (n = 6); Ob: obese (n = 13); ObCap: obese with capsinoids (n = 14).
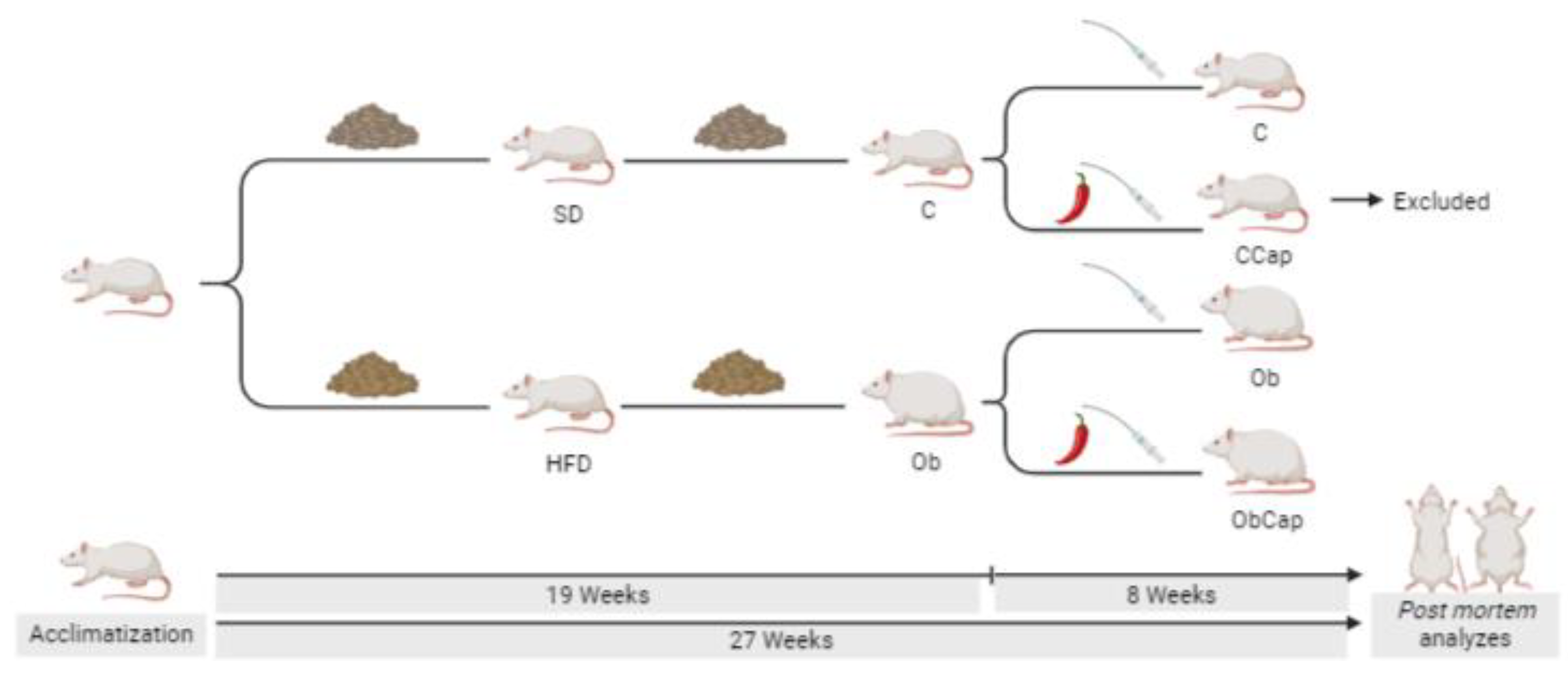
Figure 2.
Evolution of body mass during the period of exposure to experimental diets. Groups SD: standard diet (n = 17); HFD: high-fat diet (n = 27); After the onset and characterization of obesity, the groups were renamed: C: control (n = 17), Ob: obese (n = 27). Values expressed as mean ± standard deviation. Two-way ANOVA for independent samples complemented with Bonferroni post hoc test - * p <0.05 Ob vs. C.
Figure 2.
Evolution of body mass during the period of exposure to experimental diets. Groups SD: standard diet (n = 17); HFD: high-fat diet (n = 27); After the onset and characterization of obesity, the groups were renamed: C: control (n = 17), Ob: obese (n = 27). Values expressed as mean ± standard deviation. Two-way ANOVA for independent samples complemented with Bonferroni post hoc test - * p <0.05 Ob vs. C.
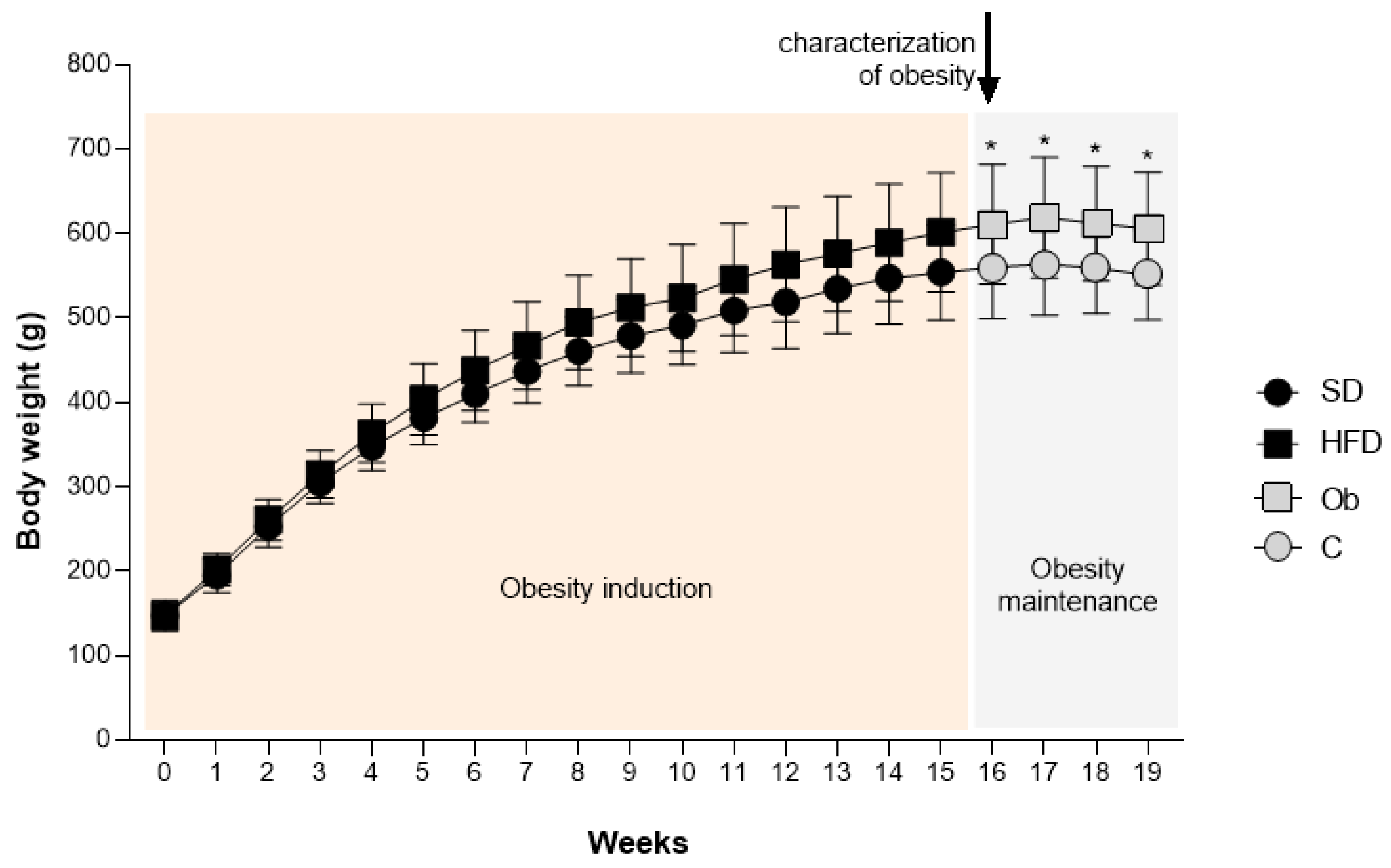
Figure 3.
Body weight and adiposity profile after treatment with capsinoids. AI: adiposity index. C: control (n = 11); Ob: obese (n = 13); ObCap: obese treated with capsinoids (n = 14). Values expressed as mean ± standard deviation. Student t test for independent samples - * p <0.05 Ob vs. C, # ObCap vs. Ob.
Figure 3.
Body weight and adiposity profile after treatment with capsinoids. AI: adiposity index. C: control (n = 11); Ob: obese (n = 13); ObCap: obese treated with capsinoids (n = 14). Values expressed as mean ± standard deviation. Student t test for independent samples - * p <0.05 Ob vs. C, # ObCap vs. Ob.
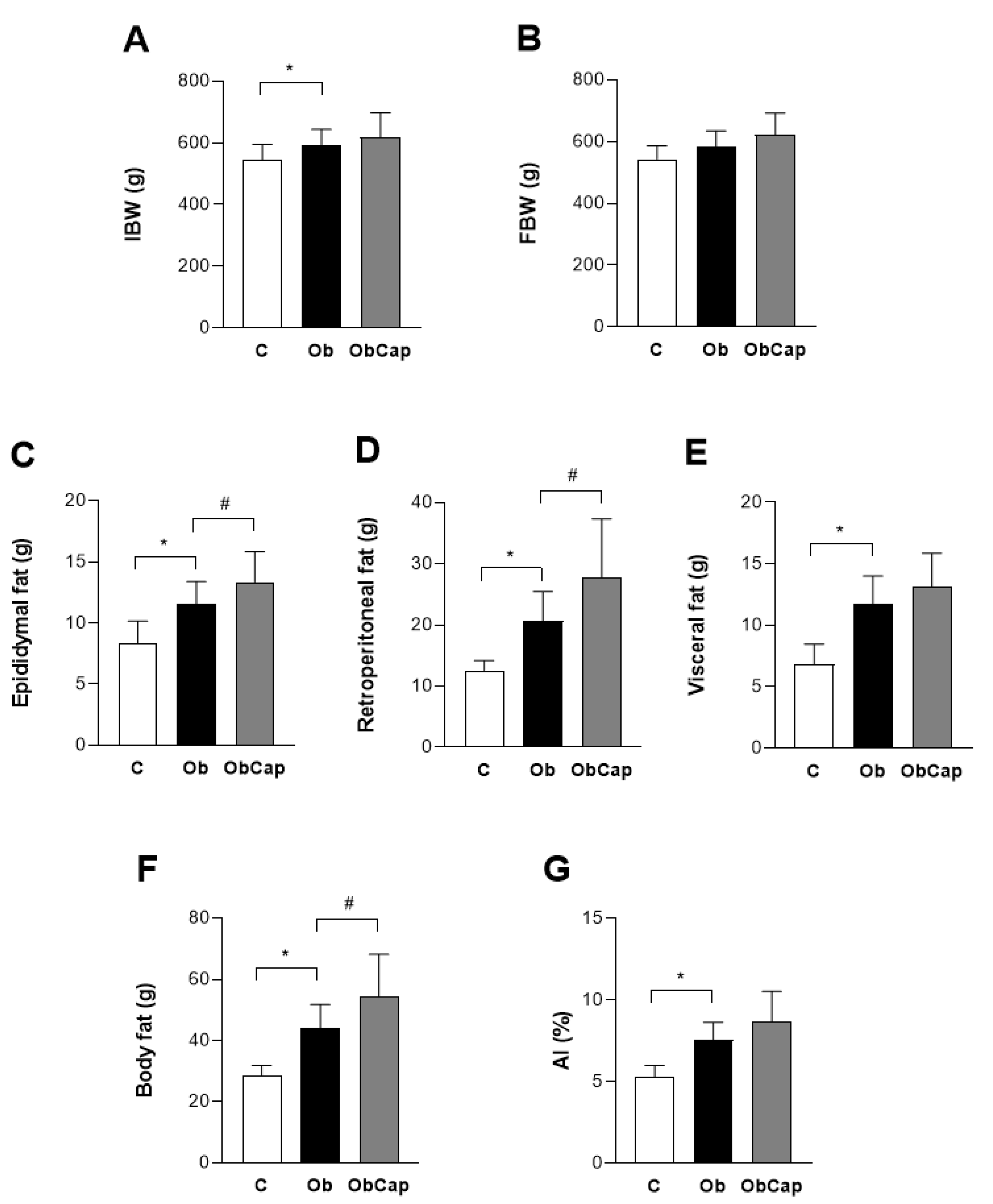
Figure 4.
Adipocytes of visceral adipose tissue. (A) Adipocyte area of visceral fat pad. (B) Representative histological images of visceral fat pad of groups stained with hematoxylin and eosin. The scale bar is 50 μm. C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7).Values expressed as mean ± standard deviation. Student t test for independent samples.
Figure 4.
Adipocytes of visceral adipose tissue. (A) Adipocyte area of visceral fat pad. (B) Representative histological images of visceral fat pad of groups stained with hematoxylin and eosin. The scale bar is 50 μm. C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7).Values expressed as mean ± standard deviation. Student t test for independent samples.
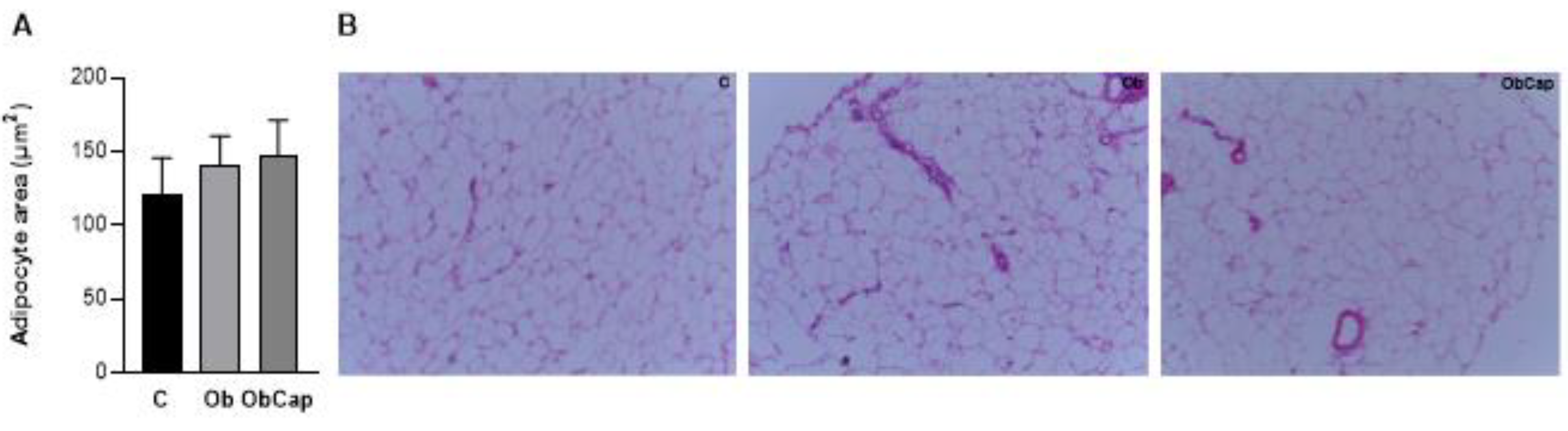
Figure 5.
Analysis of superoxide formation in sections of cardiac tissue by the dihydroetidium fluorescence (DHE). (A) DHE Fluorescence. (B) Representative images of LV with DHE fluorescence. The scale bar is 100 μm. C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7).Values expressed as mean ± standard deviation. Student t test for independent samples.
Figure 5.
Analysis of superoxide formation in sections of cardiac tissue by the dihydroetidium fluorescence (DHE). (A) DHE Fluorescence. (B) Representative images of LV with DHE fluorescence. The scale bar is 100 μm. C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7).Values expressed as mean ± standard deviation. Student t test for independent samples.
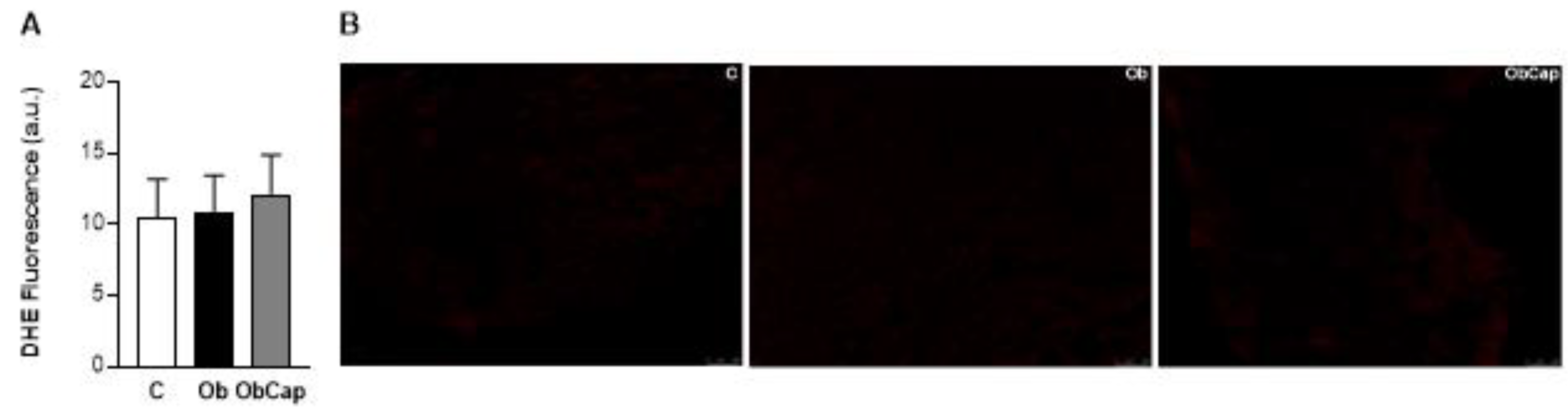
Figure 6.
Effect of capsinoid treatment on cardiac biomarkers of oxidative stress and on antioxidant enzymes. MDA: malondialdehyde. CBO: carbonyl proteins. AOPP: advanced oxidation protein products. FRAP: iron-reducing antioxidant capacity. SOD: superoxide dismutase. CAT: catalase. C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7). Values expressed as mean ± standard deviation. Student t test for independent samples - * p < 0,05 Ob vs. C. # ObCap vs. Ob.
Figure 6.
Effect of capsinoid treatment on cardiac biomarkers of oxidative stress and on antioxidant enzymes. MDA: malondialdehyde. CBO: carbonyl proteins. AOPP: advanced oxidation protein products. FRAP: iron-reducing antioxidant capacity. SOD: superoxide dismutase. CAT: catalase. C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7). Values expressed as mean ± standard deviation. Student t test for independent samples - * p < 0,05 Ob vs. C. # ObCap vs. Ob.
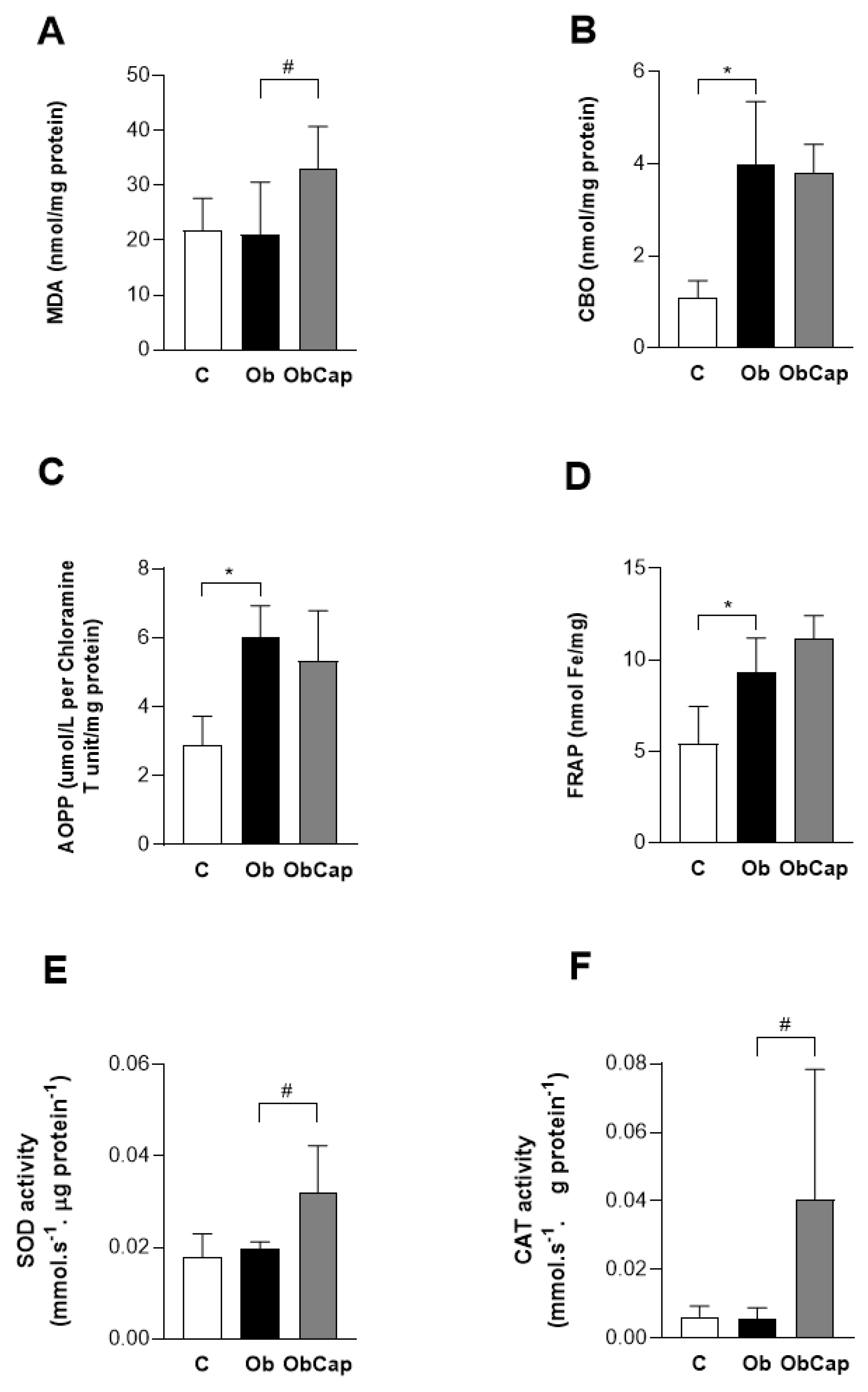

Table 1.
Lipid, glycemic and hormonal profiles after treatment with capsinoids.
| Variable | Groups | ||
|---|---|---|---|
| C | Ob | ObCap | |
| Cholesterol (mg/dL) | 41.8 ± 9.0 | 70.8 ± 6.8* | 57.5 ± 18.2# |
| Triglycerides (mg/dL) | 25.5 ± 9.2 | 32.1 ± 17,.1 | 19.5 ± 6.4 |
| HDL (mg/dL) | 15.9 ± 3.3 | 18.2 ± 3.3 | 17.5 ± 5.3 |
| LDL (mg/dL) | 14.9 ± 5.4 | 13.1 ± 2.7 | 12.9 ± 3.3 |
| Glucose (mg/dL) | 90.2 ± 12,2 | 102 ± 8 | 118 ± 7# |
| AUC for glucose (mg/dL.min) | 936 ± 144 | 943 ± 106 | 1071 ± 178 |
| Insulin (pg/mL) | 35.4 ± 9.0 | 80.8 ± 19.9* | 73.1 ± 11.3 |
| HOMA-IR | 0.23 ± 0.07 | 0.57 ± 0.14* | 0.62 ± 0.09 |
| Leptin (ng/mL) | 1.81 ± 0.60 | 2.90 ± 0.30* | 3.03 ± 0,38 |
| Glucagon (ng/mL) | 0.14 ± 0.02 | 0.12 ± 0.04 | 0.11 ± 0,01 |
| Adiponectin (ng/mL) | 37.7 ± 11.5 | 43.5 ± 10.1 | 45.5 ± 5.5 |
HDL: high-density lipoprotein. LDL: low-density lipoprotein. AUC: area under the curve for glucose. HOMA-IR: homeostatic model assessment for insulin resistance. Groups C: control; Ob: obese; ObCap: obese treated with capsinoids. Leptin (C: n = 7, Ob: n = 7, ObCap: n = 7); Glucagon and Adiponectin (C: n = 7, Ob: n = 6, ObCap: n = 7); Cholesterol, triglycerides, HDL e LDL (C: n = 9, Ob: n = 9, ObCap: n = 9); Blood glucose (C: n = 6, Ob: n = 8, ObCap: n = 10); Insulin e HOMA-IR (C: n = 5, Ob: n = 7, ObCap: n = 5). Values expressed as mean ± standard deviation. Student t test for independent samples - * p <0.05 - Ob vs. C; # p <0.05 ObCap vs. Ob.
Table 2.
Cardiac injury and remodeling process after treatment with capsinoids.
| Variable | Groups | ||
|---|---|---|---|
| C | Ob | ObCap | |
| Heart (g) | 1.28 ± 0.13 | 1.51 ± 0.17* | 1.47 ± 1.10 |
| Hearth/tibia length (g/cm) | 0.30 ± 0.03 | 0.37 ± 0.03 | 0.33 ± 0.02 |
| LV (g) | 0.72 ± 0.12 | 0.99 ± 0.22* | 1.06 ± 0.07 |
| LV/tibia length (g/cm) | 0.17 ± 0.03 | 0.22 ± 0.05 | 0.24 ± 0.01 |
| Troponin I (pg/mL) | 15.32 ± 3.24 | 18.46 ± 3.28 | 12.37 ± 2.79# |
LV: left ventricle. Groups C: control (n = 5); Ob: obese (n = 6); ObCap: obese treated with capsinoids (n = 7). Values expressed as mean ± standard deviation. Student t Test for independent samples - * p <0.05 Ob vs. C; # ObCap vs. Ob.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



