
Submitted:
23 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
Sialyllactose (SL) is a functional human milk oligosaccharide essential for immune support,
brain development, intestinal maturation, and antiviral defense. However, despite its
established health benefits, the effect of SL on exercise performance and muscle mass in mice
remains unknown. Here, we aimed to investigate, for the first time, the effects of 6′-SL on
muscle functions. Seven-week-old male C57BL/6J mice were administered 100 mg/kg 6′-SL
for 12 weeks, after which exhaustive treadmill performance, muscle strength, and muscle
phenotype were examined. The administration of 6′-SL significantly improved exhaustive
treadmill performance metrics, including distance and exhaustion time. Grip strength was also
increased by 6′-SL administration. Additionally, 6′-SL increased muscle mass in both the
gastrocnemius (GAS) and soleus. 6′-SL administration led to an increase in the minimum
Feret’s diameter and the protein expression of total myosin heavy chain in the GAS muscle. In
conclusion, 6′-SL administration in vivo led to increased running distance and time by
increasing muscle mass and strength. These findings collectively indicate that 6′-SL is a
potential agent for improving muscle health and exercise performance.
Keywords:
6′-sialyllactose
; exercise performance
; muscle strength
; muscle mass
1. Introduction
Exercise performance is influenced by various factors, including muscle fiber composition, metabolic efficiency, muscle mass, muscle strength, and neuromuscular coordination [1,2,3]. Many studies corroborate that increased muscle strength and mass facilitate the execution of common sports skills such as jumping, sprinting, and changing direction [4,5]. Strength training and dietary supplements are established strategies for increasing muscle mass, strength, and overall health [6]. Athletes and fitness enthusiasts commonly use various dietary supplements [7,8], such as creatine, protein powders, dehydroepiandrosterone (DHEA), and leucine, all known to enhance muscle mass and strength [9,10]. However, long-term or high-dose usage of these supplements may lead to potential side effects, such as gastrointestinal disturbances and drug interactions [11]. Although creatine supplementation is particularly popular for muscle growth, its effectiveness and safety remain debated [12]. Therefore, safe and accessible alternatives to address these concerns are imperative [4].
Human milk oligosaccharides (HMOs), abundant in breast milk, play a pivotal role in several bioactive functions and significantly contribute to the nutritional value of breast milk [13]. Over 100 different HMOs, including sialylated and fucosylated oligosaccharides, have been identified to date [13]. Sialyllactose (SL) is the most abundant sialylated oligosaccharide, characterized by the linkage of N-acetylneuraminic acid to the galactosyl subunit of lactose. SL confers various health benefits and plays crucial roles in several physiological processes, such as gastrointestinal microbiota development, gut maturation, brain and cognitive development, and the enhancement of innate immunity as a decoy receptor for viruses, potential pathogens, and bacteria [14]. The predominant forms of SL are 3′-SL and 6′-SL, among which 6′-SL exerts several therapeutic effects on diseases such as necrotizing enterocolitis, neuritogenesis, and benign prostatic hyperplasia [15,16,17]. In relation to muscle health, 6′-SL has also been demonstrated to ameliorate myopathic phenotypes, such as muscle weight and locomotor activity, in symptomatic bifunctional UDP-N-acetylglucosamine 2-epimerase/N-acetylmannosamine kinase (GNE) myopathy [18]. A recent study further demonstrated that 6′-SL could increase limb muscle power in patients with GNE myopathy [19]. Although the ameliorative effects of 6′-SL on myopathy have been reported, the efficacy of 6′-SL on muscle function under normal conditions has not been evaluated. Hence, in the present study, we aimed to investigate the effects of 6′-SL on exercise performance capacity and muscle phenotype in normal young mouse.
2. Materials and Methods
2.1. Animal and Treatments
Four-week-old male C57BL/6J mice were obtained from Daehan Biolink (Seoul, South Korea) and housed in a controlled environment (22–23 ℃, 12/12-h light/dark cycle), with ad libitum access to water and a normal diet. This study was conducted in accordance with the guidelines of the Pohang Technopark Animal Ethics Committee (ABCC 2022009; Pohang, South Korea). After a 3-week acclimatization period, the mice were randomly divided into two groups. Group 1 (n = 10) was orally administered with water, whereas Group 2 was administered 100 mg/kg 6′-SL in water. Both groups were treated once a day, 5 days per week, for 12 weeks. The 6′-SL used in this study was produced via enzyme synthesis by GeneChem, Inc. (Daejeon, South Korea). All experimental procedures were approved by the Institutional Ethics Committee for the Care and Use of Animals.
2.2. Sample Collection
Following humane euthanasia, blood samples and gastrocnemius (GAS) and soleus (SOL) muscles were collected from the mice. Serum was obtained by centrifugation and stored at −80 °C until analysis. GAS and SOL muscle tissues were weighed, and GAS muscle tissues were either frozen in liquid nitrogen for protein extraction or fixed in 10% formalin for histological staining.
2.3. Exercise Function Measurement
After 10 weeks of 6′-SL administration, an exhaustive treadmill exercise test was conducted. The test began at a speed of 10 m/min on a flat surface (0% slope) for 3 min. Subsequently, the speed was gradually increased to 20 m/min and maintained until the mice reached exhaustion, which was defined as the inability to continue running for 10 s [20]. A forelimb grip strength test was conducted at 3- and 10-weeks post-6′-SL administration using a maximal voluntary force testing system (BIO-G53; BIOSEB, Pinellas Park, FL, USA) [21].
2.4. Dual-Energy X-ray Absorptiometry Measurement
Fat mass, bone mineral content (BMC), bone mineral density (BMD), bone area, and bone volume were measured using dual-energy X-ray absorptiometry (DEXA; InAlyzer; MEDIKORS, Seongnam-si, South Korea).
2.5. Histological Tissue Staining
Formalin-fixed GAS muscles were embedded in paraffin and cut into 4-μm thick slices, stained with hematoxylin and eosin (H&E), and imaged using a Digital Fluorescence Slide Scanner (Axio Scan Z1, Carl Zeiss Microscopy GmbH, Jena, Germany). The minimum Feret’s diameter of muscle fibers and their percentage distribution were measured using ImageJ 1.53t [22].
2.6. Western Blotting
Total protein was isolated from GAS muscles using the T-PER™ Tissue Protein Extraction Reagent (78510; Thermo Fisher, Rockford, IL, USA). Protein concentration was determined using the Quick Start™ Bradford Protein Assay (5000202; Bio-Rad Laboratories, Hercules, CA, USA). Equal amounts of protein were loaded onto 4–20% Mini-PROTEAN® TGX™ Precast Protein Gels (4561096; Bio-Rad Laboratories, Hercules, CA, USA). Subsequently, the resolved proteins were transferred onto a polyvinylidene difluoride membrane using the Trans-Blot Turbo Transfer System (1704156; Bio-Rad Laboratories, Hercules, CA, USA) and subjected to immunoblotting using anti-total myosin heavy chain (MHC) (sc-376157; Santa Cruz Biotechnology (SCBT)) and anti-α-tubulin (sc-5286; Santa Cruz Biotechnology (SCBT)) overnight at 4 ℃. Next, the membranes were incubated with the secondary antibodies (LF-SA8001; Abfrontier, Seoul, South Korea) at room temperature for 1 h. After three washes with phosphate-buffered saline containing 0.1% Tween, protein bands were detected using an enhanced chemiluminescence reagent (1705061; Bio-Rad Laboratories, Hercules, CA, USA).
2.7. Statistical Analysis
Data are presented as mean ± standard deviation. All statistical analyses were performed using GraphPad Prism 10.1.0 (GraphPad Software, Inc., San Diego, CA, USA). Significant differences were determined using a two-tailed unpaired t-test or one-way analysis of variance followed by Dunnett’s multiple comparison test. Results with p < 0.05 were considered significant.
3. Results
3.1. 6′-SL Enhances Muscle Function
Exercise performance and grip strength were assessed to evaluate the effects of 6′-SL on muscle functions in mice. For exercise performance, mice were orally administered 100 mg/kg of 6′-SL for 10 weeks and subjected to an exhaustive treadmill exercise test, where the maximum running distance, time to fatigue, and work performed were measured. Administration of 6′-SL led to significant increases in distance (control: 559.2 ± 63.5 m, 6′-SL: 674 ± 72.8 m), time (control: 33.5 ± 2.3 min, 6′-SL: 37.1 ± 2.2 min), and work (control: 185 18,502.2 ± 1,844.9 m × kg, 6′-SL: 23,140.7 ± 2,251.7 m × kg) compared with those observed in the control group (Figure 1A).
Grip strength was measured at weeks 3 and 10 of 6'-SL administration, and the rate of increase in grip strength between weeks 3 and 10 was calculated. The administration of 6'-SL led to a greater increase in grip strength than that observed in the control group (control: 96.3% ± 6.2%, 6′-SL: 108.6% ± 12%) (Figure 1B).
3.2. 6′-SL Increases the Volume and Size of GAS Muscles
Muscle weights and fiber diameter were measured to examine the effects of 6′-SL on muscle phenotype. The administration of 6′-SL increased the absolute weights of the GAS and SOL muscles (Figure 2A), which are key muscles of the calf used for walking, running, and jumping. To support the observed increase in muscle weights, we measured body weight, fat mass, and various bone mass parameters (BMC, BMD, bone area, and bone volume) (Table 1). An increase in body weight without corresponding fat and bone mass gain was observed, partly suggesting an increase in muscle mass. Additionally, 6′-SL administration increased the protein expression of total MHC, a late-stage differentiation marker (Figure 2B) [23]. H&E staining of the GAS muscle revealed larger muscle fiber diameters in the 6′-SL administration group than in the control group. This finding was confirmed by quantifying fiber diameter size using a percentage distribution curve, which showed a trend toward larger sizes, and by measuring the average minimum Ferret diameter, which was significantly increased in the 6′-SL administration group (Figure 2C–E).
4. Discussion
This study demonstrated that 6′-SL, an HMO, affected exercise performance by altering muscle mass, fiber size, and MHC protein expression in C57BL/6J mice. Exercise performance is influenced by intrinsic factors, such as genetic constitution and athletic ability, as well as extrinsic factors, such as exercise training and proper nutrition [24,25].
Muscle mass plays a key role in exercise performance, influencing many aspects of strength and athletic ability [26]. Increased muscle mass improves endurance by enabling greater force exertion and improved storage of glycogen, the main fuel required for prolonged exercise [27]. Various nutritional supplements have been reported to increase muscle mass, strength, and exercise performance [28]. For example, DHEA, a precursor of sex steroid hormones, promotes protein synthesis and anabolism, resulting in increased muscle mass and strength [29]. Beta-methyl-hydroxy-beta-methylbutyrate has demonstrated efficacy in preserving muscle mass and strength in older individuals and promoting skeletal muscle hypertrophy in bodybuilders and strength/power athletes [30,31]. Creatine has also been reported to positively affect several aspects of exercise performance, including muscle mass and strength, glycogen synthesis, and aerobic capacity [32]. Additionally, amino acids, including branched-chain amino acids, glutamine, aspartates, and arginine, as well as protein supplements, such as whey protein and colostrum, have demonstrated efficacy in improving physical performance [33]. Despite the availability of numerous dietary supplements, many uncertainties persist regarding their safety and effectiveness.
MHC is crucial for muscle function by influencing muscle contraction, energy consumption, and structure [34]. Additionally, MHC is a marker protein commonly used to assess muscle fiber lengths, muscle mass, and strength [35]. According to previous studies, various materials such as α- lipoic acid, creatine, dihydromyricetin, ursolic acid and leucine have been identified to increase MHC expression and improve muscle function. α-lipoic acid, a potent biological antioxidant has been reported to promote MHC gene expression and maintain muscle mass in Otsuka Long-Evans Tokushima Fatty rats, and reduce muscle degradation, promote muscle regeneration, and consequently maintain muscle mass in type 2 diabetes mellitus rats [35]. Creatine supplementation increased muscle strength and size through increased MHC mRNA and protein levels [36]. Dihydromyricetin has been shown to upregulate MHC I expression through the AMPK signaling pathway, enhancing muscle performance [37]. Moreover, a previous study showed that ursolic acid and leucine significantly induced MHC protein expression and promoted C2C12 muscle cell differentiation [38].
In this study, 6′-SL increased muscle weight (GAS and SOL), GAS muscle fiber size, and MHC protein expression in GAS muscle. These results suggest that 6′-SL could enhance endurance exercise performance, including exhaustion time, total distance, and work output, by increasing muscle mass and strength in C57BL/6J mice.
Additionally, the safety of 6′-SL has been previously reported in piglets, healthy adults, and infant formulas [39,40,41].
Proposed mechanisms for improving muscle health include enhanced anti-inflammatory and antioxidant properties, increased expression of the mammalian target of rapamycin (mTOR) signaling pathway, reduced protein degradation, and improved mitochondrial function [42,43]. Whey protein has been reported to induce the mTOR pathway in resistance-exercising young men [44,45]. Ginseng, which has antioxidant and anti-inflammatory properties, improves muscle regeneration post-exercise in healthy adults [46]. Branched-chain amino acids and vitamin D supplementation improved mitochondrial function and enhanced strength and performance in atrophic muscle [47]. Notably, the precise mechanism by which 6′-SL enhances exercise performance was not determined in this study. However, 6′-SL has been demonstrated to inhibit lipopolysaccharide-induced inflammatory symptoms in intestinally inflamed suckling mice [48]. Furthermore, in Caenorhabditis elegans, 6'-SL improved endurance exercise performance by increasing glycogenolysis and affecting mitochondrial function [49]. Based on these previous reports, we intend to conduct more comprehensive studies to elucidate the mechanisms by which 6′-SL affects muscle phenotype and functions.
In conclusion, this study demonstrated that 6′-SL increased muscle mass and strength, thereby enhancing exercise performance in young C57BL/6J mice. These findings suggest that 6′-SL is both beneficial and safe for enhancing exercise performance.
Author Contributions
Conceptualization and methodology, E.J.P, L.K., Y-A.K.; investigation, H.G., Y-A.K.; writing - original draft preparation, H.G.; writing - review & editing, E.J.P, J-O. L., H.L.; formal analysis, H.G.; visualization and validation, H.G., Y-A.K.; project administration, H.G.; supervision, Y-A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Bio-industry Core Technology Development Business of the Ministry of Trade, Industry and Energy (20014827), the Technology development Program of the Ministry of SMEs and Startups (Korea) (S3122086) and the National Research Foundation of Korea (NRF) (NRF-2020M3H1A107531414).
Institutional Review Board Statement
The experimental procedures were conducted following the guidelines of the NIH for the Care and Use of Laboratory Animals and were approved by the Pohang Technopark Animal Ethics Committee (ABCC 2022009; Pohang, South Korea).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Conflicts of Interest
The authors from POSTECH declare no conflicts of interest. Hiroe Go, Eun Jung Park, and Lila Kim were employed by GeneChem Inc. Lila Kim holds stock in GeneChem Inc.
Acknowledgments
We would like to thank Editage (www.editage.co.kr) for English language editing.
References
- Zierath, J. R.; Hawley, J. A. Skeletal muscle fiber type: influence on contractile and metabolic properties. PLoS Biol 2004, 2, e348. [Google Scholar] [CrossRef] [PubMed]
- Bogdanis, G. C. Effects of physical activity and inactivity on muscle fatigue. Front Physiol 2012, 3, 142. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S.; Soh, K. G.; Jazaily Mohd Nasiruddin, N.; Bashir, M.; Cao, S.; Soh, K. L. Effects of neuromuscular training on athletes physical fitness in sports: A systematic review. Front Physiol 2022, 13, 939042. [Google Scholar] [CrossRef] [PubMed]
- Suchomel, T. J.; Nimphius, S.; Stone, M. H. The Importance of Muscular Strength in Athletic Performance. Sports Med 2016, 46, 1419–1449. [Google Scholar] [CrossRef] [PubMed]
- Nakai, Y.; Usumoto, Y.; Takeshita, Y. The Effects of Regional Muscle Strength and Mass on Standing Long Jump Performance. Muscles 2024, 3, 60–70. [Google Scholar] [CrossRef]
- Samadi, M.; Khosravy, T.; Azadbakht, L.; Rezaei, M.; Mosafaghadir, M.; Kamari, N.; Bagheri, A.; Pasdar, Y.; Najafi, F.; Hamze, B.; Soleimani, D. Major dietary patterns in relation to muscle strength status among middle-aged people: A cross-sectional study within the RaNCD cohort. Food Sci Nutri 2021, 9, 6672–6682. [Google Scholar] [CrossRef] [PubMed]
- Maughan, R.; Depiesse, F.; Geyer, H. The use of dietary supplements by athletes. J Sports Sci 2007, 25 Suppl 1, S103–113. [Google Scholar] [CrossRef]
- Rawson, E. S.; Miles, M. P.; Larson-Meyer, D. E. Dietary Supplements for Health, Adaptation, and Recovery in Athletes. Int J Sport Nutr Exerc Metab 2018, 28, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Kreider, R. B. Dietary supplements and the promotion of muscle growth with resistance exercise. Sports Med 1999, 27, 97–110. [Google Scholar] [CrossRef]
- Beaudart, C.; Rabenda, V.; Simmons, M.; Geerinck, A.; Araujo de Carvalho, I.; Reginster, J. Y.; Amuthavalli Thiyagarajan, J.; Bruyère, O. Effects of Protein, Essential Amino Acids, B-Hydroxy B-Methylbutyrate, Creatine, Dehydroepiandrosterone and Fatty Acid Supplementation on Muscle Mass, Muscle Strength and Physical Performance in Older People Aged 60 Years and Over. A Systematic Review of the Literature. J Nutr Health Aging 2018, 22, 117–130. [Google Scholar]
- Beck, K. L.; Thomson, J. S.; Swift, R. J.; von Hurst, P. R. Role of nutrition in performance enhancement and postexercise recovery. Open Access J Sports Med 2015, 6, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Antonio, J.; Candow, D. G.; Forbes, S. C.; Gualano, B.; Jagim, A. R.; Kreider, R. B.; Rawson, E. S.; Smith-Ryan, A. E.; VanDusseldorp, T. A.; Willoughby, D. S.; et al. Common questions and misconceptions about creatine supplementation: what does the scientific evidence really show? J Int Soc Sports Nutr 2021, 18, 13. [Google Scholar] [CrossRef] [PubMed]
- ten Bruggencate, S. J.; Bovee-Oudenhoven, I. M.; Feitsma, A. L.; van Hoffen, E.; Schoterman, M. H. Functional role and mechanisms of sialyllactose and other sialylated milk oligosaccharides. Nutr Rev 2014, 72, 377–389. [Google Scholar] [CrossRef]
- Guo, L.; Chen, X.; Xu, L.; Xiao, M.; Lu, L. Enzymatic Synthesis of 6'-Sialyllactose, a Dominant Sialylated Human Milk Oligosaccharide, by a Novel exo-alpha-Sialidase from Bacteroides fragilis NCTC9343. Appl Environ Microbiol 2018, 84, e00071–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, E. Y.; Jin, B. R.; Chung, T. W.; Bae, S. J.; Park, H.; Ryu, D.; Jin, L.; An, H. J.; Ha, K. T. 6-sialyllactose ameliorates dihydrotestosterone-induced benign prostatic hyperplasia through suppressing VEGF-mediated angiogenesis. BMB Rep 2019, 52, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. X.; He, J. H.; Mi, Y. J.; Shen, H. F.; Schachner, M.; Zhao, W. J. A mimetic peptide of alpha2,6-sialyllactose promotes neuritogenesis. Neural Regen Res 2020, 15, 1058–1065. [Google Scholar] [PubMed]
- Sodhi, C. P.; Wipf, O.; Yamaguchi, Y.; Fulton, W. B. ; K., M.; Niño, D. F.; Zhou, Q.; Banfield, E.; Werts, A. D.; Ladd, M. R.; Buck, R. H.; Goehring, K. C.; Prindle Jr, T.; Wang, S.; Jia, H.; Lu, P.; Hackam, D. J. The human milk oligosaccharides 2'-fucosyllactose and 6'-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr Res, 2021; 89, 91–101. [Google Scholar]
- Yonekawa, T.; Malicdan, M. C.; Cho, A.; Hayashi, Y. K.; Nonaka, I.; Mine, T.; Yamamoto, T.; Nishino, I.; Noguchi, S. Sialyllactose ameliorates myopathic phenotypes in symptomatic GNE myopathy model mice. Brain 2014, 137, 2670–2679. [Google Scholar] [CrossRef] [PubMed]
- Park, Y. E.; Park, E.; Choi, J.; Go, H.; Park, D. B.; Kim, M. Y.; Sung, N. J.; Kim, L.; Shin, J. H. Pharmacokinetics and clinical efficacy of 6'-sialyllactose in patients with GNE myopathy: Randomized pilot trial. Biomed Pharmacother 2023, 168, 115689. [Google Scholar] [CrossRef]
- Kim, Y. A.; Oh, S. H.; Lee, G. H.; Hoa, P. T.; Jin, S. W.; Chung, Y. C.; Lee, Y.C.; Jeong, H. G. Platycodon grandiflorum-derived saponin attenuates the eccentric exercise-induced muscle damage. Food Chem Toxicol 2018, 112, 150–156. [Google Scholar] [CrossRef]
- Menalled, L. B.; Patry, M.; Ragland, N.; Lowden, P. A.; Goodman, J.; Minnich, J.; Zahasky, B.; Park, L.; Leeds, J.; Howland, D.; Signer, E.; Tobin, A. J.; Brunner, D. Comprehensive behavioral testing in the R6/2 mouse model of Huntington's disease shows no benefit from CoQ10 or minocycline. PLoS One 2010, 5, e9793. [Google Scholar] [CrossRef]
- Valenzuela, N.; Soibam, B.; Li, L.; Wang, J.; Byers, L. A.; Liu, Y.; Schwartz, R. J.; Stewart, M. D. HIRA deficiency in muscle fibers causes hypertrophy and susceptibility to oxidative stress. J Cell Sci 2017, 130, 2551–2563. [Google Scholar] [CrossRef] [PubMed]
- Murru, C.; Duvert, L.; Magdinier, F.; Casanova, A.; Alloncle, A.-P.; Testa, S.; Al-Kattan, A. Assessment of laser-synthesized Si nanoparticle effects on myoblast motility, proliferation and differentiation: towards potential tissue engineering applications. Nanoscale Adv 2024, 6, 2104–2112. [Google Scholar] [CrossRef] [PubMed]
- Kuok Ho, D. T. A Review of the Association between Environmental Factors and Athletic Performance. Sport Sci 2021, 1, 21–30. [Google Scholar]
- Spagnolo, A.; Klug, S.; Schenkl, C.; Schwarzer, M. Links between Exercise Capacity, Exercise Training, and Metabolism. Compr Physiol 2023, 13, 5115–5155. [Google Scholar] [PubMed]
- Pourreza, S.; Shahinfar, H.; Bazshahi, E.; Gholami, F.; Djafarian, K.; Shab-Bidar, S. Association of the Mediterranean Dietary Quality Index with handgrip strength and muscle endurance: A cross-sectional study. Food Sci Nutr 2022, 10, 2749–2759. [Google Scholar] [CrossRef] [PubMed]
- Schytz, C. T.; Ørtenblad, N.; Birkholm, T. A.; Plomgaard, P.; Nybo, L.; Kolnes, K. J.; Andersen, O. E.; Lundby, C.; Nielsen, J.; Gejl, K. D. Lowered muscle glycogen reduces body mass with no effect on short-term exercise performance in men. Scand J Med Sci Sports 2023, 33, 1054–1071. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, P. M.; Rawson, E. S. Nutritional supplements to increase muscle mass. Crit Rev Food Sci Nutr 1999, 39, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Iemitsu, M. The Role of Dehydroepiandrosterone (DHEA) in Skeletal Muscle. Vitam Horm 2018, 108, 205–221. [Google Scholar] [PubMed]
- Wilson, G. J.; Wilson, J. M.; Manninen, A. H. Effects of beta-hydroxy-beta-methylbutyrate (HMB) on exercise performance and body composition across varying levels of age, sex, and training experience: A review. Nutr Metab (Lond) 2008, 5, 1. [Google Scholar] [CrossRef]
- Oktaviana, J.; Zanker, J.; Vogrin, S.; Duque, G. The Effect of β-hydroxy-β-methylbutyrate (HMB) on Sarcopenia and Functional Frailty in Older Persons: A Systematic Review. J Nutr Health Aging 2019, 23, 145–150. [Google Scholar] [CrossRef]
- Wax, B.; Kerksick, C. M.; Jagim, A. R.; Mayo, J. J.; Lyons, B. C.; Kreider, R. B. Creatine for Exercise and Sports Performance, with Recovery Considerations for Healthy Populations. Nutrients 2021, 13, 1915. [Google Scholar] [CrossRef] [PubMed]
- Williams, M. Dietary supplements and sports performance: amino acids. J Int Soc Sports Nutr 2005, 2, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C. T. A.; Ochala, J. Myosin Heavy Chain as a Novel Key Modulator of Striated Muscle Resting State. Physiology (Bethesda) 2023, 38, 0. [Google Scholar] [CrossRef] [PubMed]
- Ko, C. Y.; Wu, C. H.; Huang, W. J.; Lo, Y. M.; Lin, S. X.; Wu, J. S.; Huang, W. C.; Shen, S. C. Alleviative effects of α-lipoic acid on muscle atrophy via the modulation of TNF-α/JNK and PI3K/AKT pathways in high-fat diet and streptozotocin-induced type 2 diabetic rats. Food Sci Nutr 2023, 11, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, D. S.; Rosene, J. Effects of oral creatine and resistance training on myosin heavy chain expression. Med Sci Sports Exerc 2001, 33, 1674–1681. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Chen, X.; Chen, D.; Yu, B.; He, J.; Zheng, P.; Luo, Y.; Chen, H.; Yan, H.; Huang, Z. Dihydromyricetin alters myosin heavy chain expression via AMPK signaling pathway in porcine myotubes. Food & Function 2022, 13, 10525–10534. [Google Scholar]
- Kim, M.; Sung, B.; Kang, Y. J.; Kim, D. H.; Lee, Y.; Hwang, S. Y.; Yoon, J. H.; Yoo, M. A.; Kim, C. M.; Chung, H. Y.; Kim, N. D. The combination of ursolic acid and leucine potentiates the differentiation of C2C12 murine myoblasts through the mTOR signaling pathway. Int J Mol Med 2015, 35, 755–762. [Google Scholar] [CrossRef]
- Phipps, K. R.; Baldwin, N. J.; Lynch, B.; Stannard, D. R.; Šoltésová, A.; Gilby, B.; Mikš, M. H.; Röhrig, C. H. Toxicological safety evaluation of the human-identical milk oligosaccharide 6'-sialyllactose sodium salt. J Appl Toxicol 2019, 39, 1444–1461. [Google Scholar] [CrossRef]
- Kim, J. H.; Yong, S. Y.; Kim, S. H.; Baek, A.; Go, T. H.; Kang, D. R. Randomized, triple-blind, placebo-controlled study to evaluate the safety of 6'-Sialyllactose in healthy adults. Regul Toxicol Pharmacol 2022, 129, 105110. [Google Scholar] [CrossRef]
- Golden, R.; Sutkus, L.; Bauer, L.; Donovan, S.; Dilger, R. Determining the safety and efficacy of dietary supplementation with 3ˊ-sialyllactose or 6ˊ-sialyllactose on growth, tolerance, and brain sialic acid concentrations. Front Nutr 2023, 10. [Google Scholar] [CrossRef]
- Buonocore, D.; Negro, M.; Arcelli, E.; Marzatico, F. Anti-inflammatory Dietary Interventions and Supplements to Improve Performance during Athletic Training. J Am Coll Nutr 2015, 34 Suppl 1, 62–67. [Google Scholar] [CrossRef]
- Therdyothin, A.; Phiphopthatsanee, N.; Isanejad, M. The Effect of Omega-3 Fatty Acids on Sarcopenia: Mechanism of Action and Potential Efficacy. Mar Drugs 2023, 21, 399. [Google Scholar] [CrossRef] [PubMed]
- Farnfield, M. M.; Carey, K. A.; Gran, P.; Trenerry, M. K.; Cameron-Smith, D. Whey protein ingestion activates mTOR-dependent signalling after resistance exercise in young men: a double-blinded randomized controlled trial. Nutrients 2009, 1, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Yoshida, N.; Maekawa, D.; Kitakaze, T.; Kobayashi, Y.; Kitano, T.; Fujita, T.; Okuwa-Hayashi, H.; Harada, N.; Nakano, Y.; Yamaji, R. 5-Hydroxy-7-methoxyflavone derivatives from Kaempferia parviflora induce skeletal muscle hypertrophy. Food Sci Nutr 2018, 7, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Castellanos, B.; Martínez-López, P.; Bailón-Moreno, R.; Esquius, L. Effect of Ginseng Intake on Muscle Damage Induced by Exercise in Healthy Adults. Nutrients 2024, 16, 90. [Google Scholar] [CrossRef] [PubMed]
- Cochet, C.; Belloni, G.; Buondonno, I.; Chiara, F.; D'Amelio, P. The Role of Nutrition in the Treatment of Sarcopenia in Old Patients: From Restoration of Mitochondrial Activity to Improvement of Muscle Performance, a Systematic Review. Nutrients 2023, 15, 3730. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. Y.; Lee, S.; Kim, G.; Shin, H. J.; Lee, E. J.; Lee, C. S.; Yoon, S.; Lee, E.; Lim, A.; Kim S., H. Ameliorating effect of 2'-Fucosyllactose and 6'-Sialyllactose on lipopolysaccharide-induced intestinal inflammation. J Dairy Sci 2024, 107, 4147–4160. [Google Scholar] [CrossRef]
- Arellano Spadaro, J.; Hishida, Y.; Matsunaga, Y.; van Es-Remers, M.; Korthout, H.; Kim, H. K.; Poppelaars, E.; Keizer, H.; Iliopoulou, E.; van Duijn, B.; Wildwater, M.; van Rijnberk, L. 3'sialyllactose and 6'sialyllactose enhance performance in endurance-type exercise through metabolic adaptation. Food Sci Nutr 2023, 11, 6199–6212. [Google Scholar] [CrossRef]
Figure 1.
Effect of 6′-SL on exercise performance and muscle strength. (A) Total running distance, exhaustion time, and work during exhaustive exercise were measured at 10 weeks. Work was calculated as running distance (m) × body weight (g). (B) The rate of increase in grip strength/body weight between weeks 3 and 10. Data are presented as mean ± standard deviation values for each group (n = 8). A two-tailed unpaired t-test was used: *P < 0.05, ** P < 0.01 vs. control. Abbreviations: SL, sialyllactose; BW, body weight.
Figure 1.
Effect of 6′-SL on exercise performance and muscle strength. (A) Total running distance, exhaustion time, and work during exhaustive exercise were measured at 10 weeks. Work was calculated as running distance (m) × body weight (g). (B) The rate of increase in grip strength/body weight between weeks 3 and 10. Data are presented as mean ± standard deviation values for each group (n = 8). A two-tailed unpaired t-test was used: *P < 0.05, ** P < 0.01 vs. control. Abbreviations: SL, sialyllactose; BW, body weight.
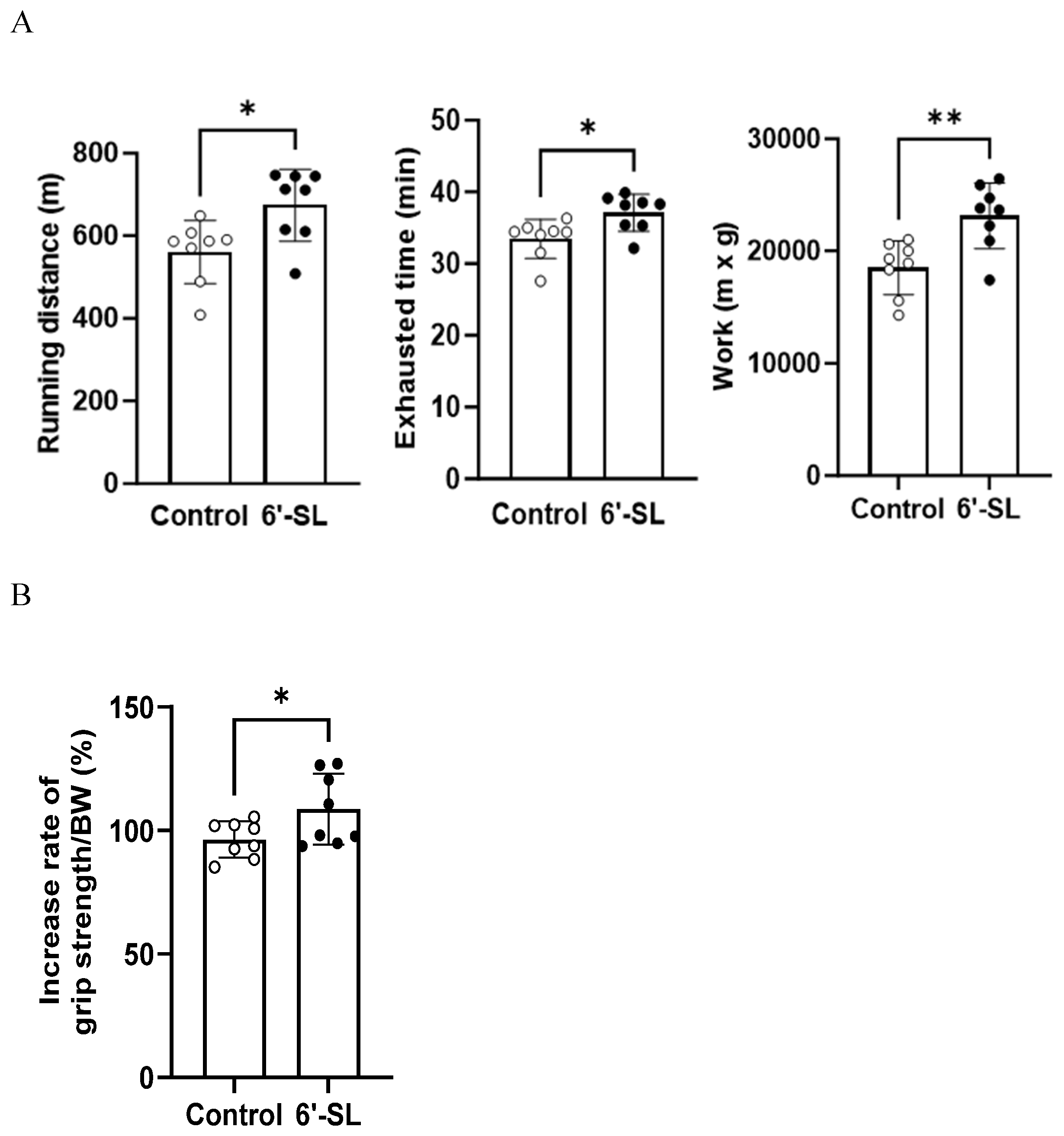
Figure 2.
Effect of 6′-SL on muscle mass and fiber formation in GAS and SOL muscles. (A) Absolute tissue weight (gastrocnemius (GAS) and soleus muscles) was measured at sacrifice. (B) Protein expression levels of the total myosin heavy chain of GAS muscle were measured using western blotting. The relative band intensities of each protein were normalized to those of α-tubulin, and the quantified data are shown in the bar graphs. (C) Images of hematoxylin and eosin-stained cross sections of the GAS muscle; Scale bar, 50 μm. (D) Percentage muscle fiber distribution based on Min Feret’s diameter of muscle fiber size of the GAS muscle. The numbers of muscle fibers measured in the control and 6′-SL groups were 1,535 and 1,697, respectively. (E) Mean of Min Feret’s diameter muscle fiber size of the GAS muscle. The numbers of muscle fibers measured in the control and 6′-SL groups were 591 and 633, respectively. Data are presented as mean ± standard deviation values for each group (n = 8 for (A), n = 3 for (B)-(E)). A two-tailed unpaired t-test was used: *P < 0.05, ** P < 0.01, **** P < 0.0001 vs. control. Abbreviations: SL, sialyllactose; GAS, gastrocnemius; SOL, soleus; MHC, myosin heavy chain.
Figure 2.
Effect of 6′-SL on muscle mass and fiber formation in GAS and SOL muscles. (A) Absolute tissue weight (gastrocnemius (GAS) and soleus muscles) was measured at sacrifice. (B) Protein expression levels of the total myosin heavy chain of GAS muscle were measured using western blotting. The relative band intensities of each protein were normalized to those of α-tubulin, and the quantified data are shown in the bar graphs. (C) Images of hematoxylin and eosin-stained cross sections of the GAS muscle; Scale bar, 50 μm. (D) Percentage muscle fiber distribution based on Min Feret’s diameter of muscle fiber size of the GAS muscle. The numbers of muscle fibers measured in the control and 6′-SL groups were 1,535 and 1,697, respectively. (E) Mean of Min Feret’s diameter muscle fiber size of the GAS muscle. The numbers of muscle fibers measured in the control and 6′-SL groups were 591 and 633, respectively. Data are presented as mean ± standard deviation values for each group (n = 8 for (A), n = 3 for (B)-(E)). A two-tailed unpaired t-test was used: *P < 0.05, ** P < 0.01, **** P < 0.0001 vs. control. Abbreviations: SL, sialyllactose; GAS, gastrocnemius; SOL, soleus; MHC, myosin heavy chain.
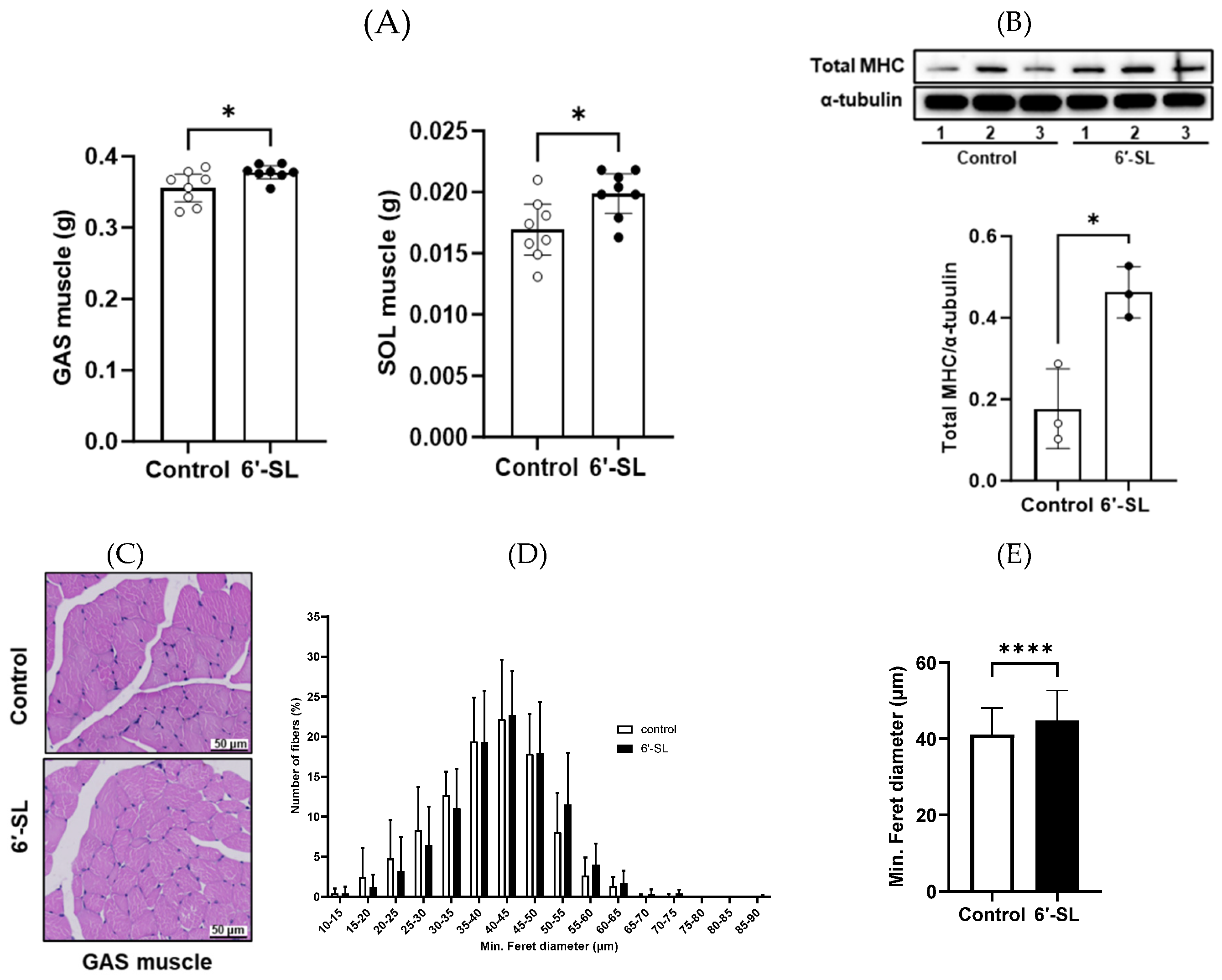

Table 1.
Data of Dual-energy X-ray absorptiometry measurements.
| . | Control (n = 8) |
6′-SL (n = 8) |
P-value |
|---|---|---|---|
|
Body weight (g) Fat mass (g) |
34.15 ± 1.40 6.67 ± 1.13 |
35.92 ± 1.00* 6.83 ± 1.10 |
0.0164 0.7987 |
| Fat mass (%) | 19.97 ± 3.24 | 19.43 ± 3.08 | 0.7542 |
| BMC (g) | 0.75 ± 0.05 | 0.78 ±0.08 | 0.4361 |
| BMD (g/cm2) | 0.07 ± 0.00 | 0.07 ± 0.00 | 0.9153 |
| Bone area (cm2) | 11.33 ± 0.40 | 11.72 ± 0.71 | 0.2328 |
| Bone volume (cm2) | 0.45 ± 0.03 | 0.47 ± 0.05 | 0.4358 |
The levels of body weight, fat mass, bone mineral content, bone mineral density, bone area, and bone volume were evaluated. Data are presented as mean ± standard deviation values for each group (n = 8). A two-tailed unpaired t-test was used: *P < 0.05 vs. control. Abbreviations: SL, sialyllactose; BMC, bone mineral content; BMD, bone mineral density.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



