
Submitted:
23 July 2024
Posted:
24 July 2024
You are already at the latest version
Abstract
The Mononuclear Phagocyte System includes monocytes, macrophages, some dendritic cells and multinuclear giant cells. These cell populations display marked heterogeneity depending on their differentiation from embryonic and bone marrow haematopoietic progenitors, tissue location and activation. They contribute to tissue homeostasis by interacting with local and systemic immune and non-immune cells, through trophic, clearance and cytocidal functions. During evolution they contributed to innate host defense before effector mechanisms of specific adaptive immunity emerged. Mouse macrophages appear at midgestation and are distributed throughout the embryo to facilitate organogenesis and clear cells undergoing programmed cell death. Yolk sac-, AGM- and foetal liver-derived tissue resident macrophages persist throughout postnatal and adult life, supplemented by bone marrow-derived blood monocytes, as required after injury and infection. Nobel awards to Elie Metchnikoff and Paul Ehrlich in 1908 drew attention to cellular phagocytic and humoral immunity, respectively. In 2011, prizes were awarded to Jules Hoffmann and Bruce Beutler for contributions to innate immunity and to Ralph Steinman for discovery of Dendritic Cells and their role in antigen presentation to T lymphocytes. We trace milestones in the history of mononuclear phagocyte research from the perspective of Nobel awards bearing directly and indirectly on their role in cellular immunity.
Keywords:
Mononuclear Phagocyte System
; Immunity
; Macrophages
; Dendritic Cells
; Multinucleated Giant Cells
; Nobel Prizes
; Phagocytosis
; Plasma Membrane Receptors
; Homeostasis
; History
Epigraph
Nothing in biology makes sense except in the context of evolution—Theodosius Dobzhansky
Nothing in macrophage biology makes sense except in the context of homeostasis—after Claude Bernard
1. Introduction
1870-1920. In keeping with the overall scope of the present review, we trace the growth of the subject during 50-year periods over the past 150 years in relation to selected Nobel awards, first awarded in 1901, in Physiology and Medicine, Chemistry and Physics (Figure 1A-D). Apart from the awards directly relevant to mononuclear phagocytes (MPs) in 1908 and 2011, we also note selected prizes in Immunology, Microbiology and Infectious diseases, as well as in Biochemistry/Metabolism, Genetics, Cell Biology and methodologic advances, which contributed substantially to their study (Figure 2).
During the mid-19th century Charles Darwin laid the groundwork for his theory of the origin of species and for natural selection as the driver of evolution [1]. Before the first Nobel awards, Rudolph Virchow had already put forward a cell-based theory of health and disease [2,3], soon followed by the stunning achievements of Louis Pasteur in chemistry, microbiology and vaccination [4]. Also in the later 19th century, Claude Bernard became a major influence in physiology and experimental medicine through his concept of the stable “milieu interieur” [5], later termed Hom(e)ostasis by Walter Cannon [6]. These and other investigators influenced the thinking of Metchnikoff, a zoologist interested in embryology, before his conversion to ”natural immunity” and comparative pathology [7]; Metchnikoff and Ehrlich shared Nobel awards in 1908 for their contributions to cellular and humoral immunity [8,9,10,11,12,13,14]. Other Nobel laureates in the early decades of the 20th century included (Table 1 and Figure 2):
- Emil von Behring, who received the first Nobel Prize for Physiology and Medicine in 1901 for introducing serum therapy as passive vaccination against diphtheria and tetanus [15]
- Robert Koch, famed for his studies on Tuberculosis and tuberculin-induced delayed hypersensitivity [16]
- Ramon Y. Cajal [17], who adapted Golgi’s silver staining method to map the intricate distribution of neurons in exquisite detail;
Metchnikoff (1845-1916) can be justly considered to be the grandfather of macrophage research. His life has been documented in several readable biographies [14,20]. Born in the Ukraine, he worked in Russia on vaccines for anthrax before joining the Pasteur Institute in Paris [21]. His putative Eureka moment with marine invertebrates at Messina [22] consolidated his interest in macrophages, large phagocytic cells, both sessile and migratory, which responded to chemotactic stimuli generated by microbial invasion. By the use of microscopy and cell staining, he identified, among other observations, intracellular acid-fast Mycobacteria of unusual appearance within multinucleated macrophage giant cells in tuberculous granulomata, emphasizing host-pathogen interactions. Anticipating later discovery of the microbiome [23], he postulated intoxication of the host by deleterious products derived from intestinal bacteria [14], in contrast with beneficial flora such as Lactobacilli which he advocated could promote human health in the form of yoghurt. Metchnikoff was prescient also in his fascination with aging, coining the term gerontology [24]. His remarkable insights into host-pathogen interactions extended to studies with his colleague Emile Roux on an experimental model of syphilis in primates [25]. He wrote several monographs on comparative pathology [7], expanding on lectures at the Pasteur Institute and had a memorable meeting with Tolstoy, an ardent anti-vivisectionist, who had written a moving short story, “The death of Ivan Illich”, based on Metchnikoff’s brother [26]. His collaborators and successors at the Pasteur Institute [21] included Emile Roux [25], Alexandre Besredka [27] and Waldemar Haffkine [28]. Jules Bordet [29] subsequently received a Nobel award (Figure 1A) for the discovery of complement and Clemens von Pirquet [30] contributed to acute hypersensitivity allergic reactions; Charles Richet, a French physician who coined the term anaphylaxis, was awarded a Nobel prize in 1913 [31].
In contrast with Metchnikoff’s citation for his Nobel award in cellular immunity, Paul Ehrlich (1854-1915) was cited for his work on humoral immunity [53] and his “side chain” cellular receptor theory of antibody induction [54]. He also used newly developed dyes [55] for differential staining of eosinophilic granulocytes, basophilic/mast cells and polymorphonuclear neutrophilic leukocytes, initially called “microphages” by Metchnikoff. Finally, Ehrlich also performed seminal studies on the role of complement in humoral defense under the direction of antibodies. If the Nobel judges were hoping to reconcile the two opposing immunology camps through a joint award, they did not succeed at the time. Ironically, we now know that both sides were correct in that cellular and humoral immunity as well as innate and acquired immunity are closely interrelated in function. The cellular faction (Metchnikoff and his colleagues from the Pasteur Institute) favored the innate “unspecific” character, whereas the humoral faction (Emil von Behring, Paul Ehrlich and colleagues) favored the humoral “specific” character as most critical. We now know that both the innate “unspecific” as well as the acquired “specific” immune response are composed of both cellular and humoral components and that successful immunity depends on a highly regulated interplay between innate and adaptive immune responses.
2. The Reticulo-Endothelial System (RES)
1920-1970. In a comprehensive review in 1924, Ludwig Aschoff drew together the findings of many investigators who studied the intravital clearance of carmine particles by tissue macrophages in liver (Kupffer cells), spleen, bone marrow, lymph nodes and the lungs [56]; often forgotten later, uptake by macrophages was also observed in endocrine organs, the adrenal and pituitary glands [56]. Walter Cannon [57] and Hans Selye [58] studied the importance of adrenaline in stress responses mediated by the sympathetic nervous system. Impressed by Metchnikoff’s work, Almroth Wright showed that agglutinins (antibodies) made particles and cells “tasty for eating”, coining the term opsonins/opsonization, enhancing phagocytosis in vivo and in vitro [59]. Much later, leukocyte receptors were shown to bind the Fc region of selected IgG antibodies, (originally defined by Rodney Porter and Gerald Edelman [60], who received Nobel awards for their structural studies); similar opsonic cell activation by distinct phagocyte complement receptors was observed for complement-derived C3bi, activated by IgG or IgM antibodies bound to cellular or microbial antigens [61], or via an alternative carbohydrate pathway [62]. IgE antibody bound to allergens can induce histamine release from mast cells via FcR, triggering anaphylaxis [63]. Almroth Wright’s attempts to” stimulate the phagocytes” to cure disease was spoofed by George Bernard Shaw in his play, The Doctor’s Dilemma [64]. We return to the characterisation and functions of these opsonic leukocyte receptors below.
Following on the belated appreciation of Gregor Mendel’s seminal discoveries on the genetic inheritance of discrete traits [65], Archibald Garrod coined the term “Inborn errors of metabolism” [66], revealing the great value of rare genetic diseases in deciphering pathophysiology in humans and other species. Later examples of discovery of dominant genes involved in myeloid phagocytes include Chronic Granulomatous Disease [67], Interferon receptor deficiencies [68], pyrinopathies [69] and lysosomal storage diseases [70]. An example of striking macrophage-related research is notable during this period; Peyton Rous, the discoverer of Rous sarcoma virus, for which he received a Nobel prize many decades later, used magnetic separation of Kupffer cells after uptake of magnetic beads [71]. Florence Sabin, the first female medical graduate at Johns Hopkins and then Member at the Rockefeller Institute, studied macrophage involvement in Tuberculosis [72], as did Arthur Dannenberg, later at Johns Hopkins [73]. Also at Rockefeller was Nobel laureate Karl Landsteiner, discoverer of the ABO blood group antigen polymorphism [74]. In a landmark study, Merrill Chase and Landsteiner definitively showed that delayed hypersensitivity depended on adoptive transfer of cells, rather than passive immunization with serum antibodies [75].
At the same time, Howard Florey, Ernst Chain and their colleagues at the Sir William Dunn School of Pathology in Oxford followed up two earlier discoveries by Alexander Fleming of the bacteriolytic enzyme, lysozyme [76,77], and of an uncharacterized Penicillium mold product with antibacterial activity [78]. This culminated in the clinically effective and safe antibiotic penicillin for which these three investigators shared a Nobel prize [79]. Having, to his mind, solved the problem of bacterial infection, Florey, who edited an influential textbook of General Pathology [80], turned in the 50’s to the host cellular response to which he assigned his students, George Mackaness [81], James Gowans [82] and Henry Harris [83] the task of defining the role of macrophages, lymphocytes and neutrophils, respectively. We shall pick up their subsequent contributions later. Another major figure in the history of immunological tolerance and organ transplantation, Peter Medawar, a zoology student at Oxford, built on his wartime studies of skin transplants in severely burnt patients in Glasgow, to investigate foetal tolerance to paternal antigens, culminating in a Nobel award with Leslie Brent and Rupert Billingham [84,85]; this was followed by a distinguished career, although handicapped by a premature stroke, as Director of the National Institute for Medical Research (NIMR) laboratory at Mill Hill in London; this became a major centre for immunologic research over three quarters of a century [86], before moving recently to the Crick Institute. From a macrophage perspective, we single out the long-standing research interest at Mill Hill in influenza viral infection, the discovery of interferon by Alick Isaacs and Jean Lindenmann in the 1950’s [87] and the groundbreaking studies by Philip D’Arcy Hart, who identified the role of Mycobacterial inhibition of phagosome fusion with lysosomes in the pathogenesis of tuberculosis [88]. John Humphrey and Deirdre Grennan made seminal contributions to the capture of capsulated bacteria such as pneumococci by spleen macrophages through polysaccharide recognition, important in T lymphocyte -independent initiation of immunity to infection [89]. The important contributions of Avrion Mitchison on B and T lymphocyte interactions [90] and by Brigitte Askonas and Emil Unanue on antigen presentation will be considered below.
3.1. The Mononuclear Phagocyte System (MPS)
1970-2020. We return to Rockefeller in the 60’s, by now a University, in the runup to the establishment of the laboratory of the influential Phagocyte research group by James G. Hirsch (neutrophils) [91], soon followed by Zanvil A. Cohn (monocytes and macrophages) [92], and later Ralph M. Steinman (Dendritic Cells) [93]. This Myeloid Leukocyte Biology group emerged from the earlier recruitment to the Rockefeller Institute of the microbiologist René Dubos whose benign presence persisted in the laboratory long after he turned his talents to Ecology [94]. A French agrarian-trained admirer and later biographer of Pasteur, he joined Selman Waksman as a student at Rutgers University, to investigate the interactions and antibacterial properties of soil organisms [94]. Waksman later won a Nobel award for streptomycin, which transformed the clinical treatment of Tuberculosis [95]. A graduate student in the laboratory of Waksman, Albert Schatz, had first isolated streptomycin and both were jointly granted a patent for this drug. Waksman alone received the Nobel Prize in 1952 [96].
Dubos himself developed earlier antibiotics Gramicidin and Tyrothricin which unfortunately proved too toxic for clinical use [94]. After early spells at Rockefeller with Oswald Avery of DNA fame and at Harvard, Dubos returned to Rockefeller to study M. tuberculosis and Bacillus Calmette-Guérin (BCG) interactions with the host. He pioneered studies of the anaerobic flora of the gut and produced germ-free mice, long before the microbiome became a general concern in immunity [23]. Dubos admired Metchnikoff whose portrait took pride of place in his office and inspired all members of the Phagocyte research group.
The range of research topics of this group has been documented in several books and reviews by Carol Moberg, secretary sequentially to Dubos [94], Hirsch [91], Cohn [92] and Steinman [93]. Hirsch was mainly interested in the study of neutrophil degranulation, phase contrast, electron microscopy of phagocytosis in a variety of species and the bactericidal properties of histones [97]. His group made important contributions to morphologic studies of intracellular infection of macrophages by a range of pathogens, including Legionnaire’s disease [98] and toxoplasmosis [99]. After early studies on isolation of human monocytes [100], Cohn turned to the isolation and characterization of mouse peritoneal macrophages in cell culture. Both Hirsch and Cohn were influenced by the cell biological studies of Nobel laureates George Palade [101] and Christian de Duve [102] on the pancreatic secretory pathway and lysosomal digestion, respectively. The Cohn laboratory studied a range of cellular and immune functions, including phagocytosis, endocytosis, membrane traffic and recycling, cell fusion, lysosomal digestion and permeability; the group established that macrophages were not only “professional” phagocytes [103], but also potent secretory cells of enzymes, reactive oxygen and nitrogen species and arachidonate metabolites [92]. Cohn’s interest turned to human macrophage infection by intracellular pathogens, such as tuberculosis, leprosy [104], Leishmaniasis [105] and HIV [106]. The Steinman -Cohn discovery of dendritic cells (DCs), culminating in the Nobel award to Steinman a few days after his death, will be dealt with below.
It is hard to overstate the international impact of this laboratory in macrophage research at the time, punctuated by a series of Leiden conferences organized by a Dutch collaborator, Ralph van Furth [107,108,109,110]. After discussion at one of these meetings, it was decided that the term RES was no longer appropriate and the family of cells was renamed the Mononuclear Phagocyte System [111].
3.2. Enter the Lymphocytes
Metchnikoff seems to have overlooked the lymphocyte in his observations and thought that after capture by phagocytes in organs such as spleen, the agglutinins, termed “antikorper” (antibodies) by Ehrlich, which were detected in blood after immunization, were also products of the phagocytes [112]. James Gowans and his group at the Dunn school demonstrated the role of lymphatic recirculation and the delivery of smaller lymphocytes, rather than macrophages, to blood through the thoracic duct [82,113]. Florey, Gowans and their students Alvin Volkman and Vincent Marchesi did, however trace non-recirculating bone marrow-derived blood monocytes and macrophages directly to peripheral organs [114,115]. It seems remarkable that the thymus had no identified function until relatively late when Jacques Miller published his first papers [116,117]; the existence and properties of B (bursa/bone marrow), and Thymus-derived lymphocytes and their heterogeneity (Th1, Th2 and even later, helper and cytotoxic, let alone memory, effector and regulatory T lymphocyte subpopulations as well as innate lymphoid cells), were only established from the 60’s. Their role in adaptive immunity dominated the awards of Nobel prizes until 2011 (Table 1). It is interesting to compare these awards with those discoveries selected in published reviews of T [118], B lymphocytes [119] and antibodies [120].
We emphasise here their essential collaboration with cells of the MPS in both the initiation and effector mechanisms of cell mediated immunity. Australian immunologists have made major contributions in this field. We include Macfarlane Burnet, remembered for clonal selection of antibody production, as well as self-nonself recognition and virology [121,122], and Peter Doherty who, with the Swiss Rolf Zinkernagel, established the role of the Major Histocompatibility complex in cell mediated immunity [123]. Donald Metcalf [124], also at the Walter and Eliza Hall Institute in Melbourne and later, Richard Stanley [125], Australia born, at Albert Einstein College of Medicine in New York, played a major role in identifying the lineage-determining growth factors (CSF-1/ M-CSF and GM-CSF) for macrophage and granulocyte differentiation. George Mackaness and Robert North, both Australians at the Trudeau Institute in upstate New York, performed pioneering studies with BCG and Listeria monocytogenes infection models in genetically defined mouse strains [126]. They established that cell mediated immunity to infection was antigen dependent during induction, but nonspecific during expression, involving both T lymphocyte priming and activation of macrophages. The soluble lymphokine factors, initially demonstrated independently by Barry Bloom and John David [127] and others, and their receptors were identified subsequently as appropriate methods became available. Charles Dinarello played an important role in characterizing interleukins/ cytokines [128]. Mosmann and Coffman discriminated between Th1 and Th2 lymphocyte cytokines [129]. Carl Nathan played an important role in characterizing Interferon gamma, the cytokine responsible for classical macrophage activation, [130] described further below, and Gray and Goeddel cloned its receptor [131].
3.3. Enter Dendritic Cells (DCs)
Prior to the discovery by Ralph Steinman and Zanvil Cohn that DCs are specialised, potent cells for antigen capture, processing and presentation (APCs) to naive B and T lymphocytes [93,132,133], it was assumed that the more abundant macrophages in adherent mononuclear phagocyte populations were responsible for the initiation of cell mediated immunity. Emil Unanue, in particular, had demonstrated in important studies with Brigitte Askonas at Mill Hill [134] that a fraction of protein antigen was not completely degraded after uptake by “macrophage” populations, as found in studies of endocytosis by Steinman, Silverstein and Cohn [135]. It was only later that Alain Townsend, Pamela Bjorkman and others identified the role of peptide binding by MHC molecules that protected antigenic epitopes for presentation to CD4/CD8 T cells [136,137]. In their classic paper [138], Steinman and Cohn distinguished the rare DCs from conventional macrophages in mouse splenic digests by phenotypic analysis and later demonstrated “immature “and “mature” functional states [93]. A subsequent paper with Michel Nussenzweig confirmed human DCs were also able to activate lymphocytes in allogeneic mixed lymphocyte cultures [139]. It took a decade before the unique ability of DCs to activate naïve lymphocytes became widely accepted [140]. Cultures of mouse bone marrow in GM-CSF and Interleukin 4, but not CSF1, give rise to DCs in vitro [141]. The haematopoietic origin of tissue and blood DCs has revealed considerable heterogeneity in DC subpopulations [142]. Studies of the “immune synapse” by which DCs induce B and T cell activation by MHC-peptide complexes, and effector recognition of target antigens by CD8 cytotoxic T cell receptors, continue to the present day [143]. DCs play a critical role in the decision to induce tolerance versus immune activation [144,145]. Artificial intelligence is being employed to identify tumour-specific peptides for personalised immunotherapy [146].
3.4. Enter Monoclonal Antibodies
The chemical and genetic elucidation of antibody structure and function was justly recognized by a series of Nobel awards in the 60’and 70’s (Figure 1). Notably, Susumu Tonegawa was awarded the Nobel Prize in 1987 for his work at the Basel Institute for Immunology in which he identified genetic rearrangement as basis for antibody diversity. At Rockefeller, Henry Kunkel characterized the properties of monoclonal immunoglobulins in sera and urine derived from patients with multiple myeloma [147]. The roles of MPs and DCs in induction and effector mechanisms of both humoral and cellular immunity are intimately associated with progress in research methods and our understanding of disease. Perhaps one of the most arresting advances in experimental and clinical immunology came from the application of somatic cell fusion to generate immortal hybrid cell lines producing monoclonal antibodies as tools to analyse phenotypic heterogeneity by flow and histochemical analysis. Henry Harris and John Watkins in Oxford started the ball rolling in their use of UV-irradiated Sendai virus for the artificial fusion of different species of somatic cells to form homokaryons and heterokaryons which could undergo subsequent unlimited proliferation as genetically labile, interspecific hybrids [148]. It soon became evident that differentiated cells extinguished their distinctive cell-specific properties unless fused with cells of a related lineage. Harris and Klein exploited cell fusion technology to demonstrate that malignancy was a recessive phenotypic trait and that fusion with normal diploid cells such as fibroblasts could transiently suppress this trait [149]. However, the tumour suppressor genes were readily lost in hybridomas because of chromosomal instability and selection for growth in vivo or in vitro. Kohler and Milstein fused primary antibody- producing B lymphocytes with immortal myeloma cell lines in vitro, to identify and isolate hybridoma clones by direct screening for monoclonal antibodies [150]. This method was optimized using polyethyleneglycol (PEG) instead of Sendai virus and could be adapted to screen in advance for a desired functional phenotype. In the MPS field it opened up the analysis of tissue heterogeneity in situ and provided biomarker antigens [151] for cell isolation by FACS and other methods of enrichment, leading to the discovery of novel plasma membrane receptor functions. An early mab, Mac-1, produced by Timothy Springer [152,153], directed at the CR3 receptor led to the discovery of beta -2 integrins as important in myeloid phagocyte recruitment and adhesion during inflammation. Monoclonal antibodies have also opened up therapeutic opportunities to manipulate the phenotype of immune cells, including mononuclear phagocytes [154]. In addition, somatic cell fusion is a natural property of mononuclear phagocytes in vivo, physiologically, in the generation of osteoclasts, and in immune and non-immune multinucleated giant cell (MNGC) formation, discussed further below.
3.5. Enter Monoclonal Antibodies
The F4/80 mouse antigen (Emr1), isolated by Austyn and Gordon 40 years ago [155], has become a widely used plasma membrane marker to identify macrophages in many mouse organs, from the embryo throughout adult life, normally and in a range of disease models [156,157]. It became the founding member of a family of adhesion G protein 7-transmembrane receptors [158]. The related human antigen (EMR2), which is absent in the mouse, has been implicated as a mechanoreceptor which undergoes a novel method of autoproteolytic activation [159]. Collaborative gene knockout studies with Hsi Hsien Lin and Joan Stein Streilein showed that the F4/80 molecule played a non-redundant role in peripheral tolerance in an anterior chamber-associated immune deviation (ACAID) model of delayed type hypersensitivity in the eye [160], extended to allografts and tumour implantation. Figure 4 illustrates its expression in selected mouse tissues. With this groundwork, the group repeated the approach to identify and/or characterize a range of macrophage plasma membrane lectin-like (CD206, Siglec-1, Dectin1) and Scavenger receptors (SRA, MARCO and CD36) involved in adhesion, phagocytosis, endocytosis, pathogen entry, macrophage fusion and cellular interactions [161]. By using a panel of surface markers, they were able to demonstrate the heterogeneous phenotypes of resident macrophages in a range of different organs (Figure 3).
Subsequent studies of temporal and spatial mRNA and protein gene expression in mouse and human tissue have considerably extended the concept that resident macrophages display distinct phenotypes within different tissue microenvironments [165,166]. The mechanisms which underlie their diversity include local interactions with neighbouring differentiated cell types and the extracellular matrix, exposure to endogenous systemic stimuli, including the microbiome, which, in sum, regulate epigenetic adaptation of macrophage gene expression [167,168].
3.6. Development and Distribution
Studies of haematopoietic origins and lineage tracing in the mouse have given rise to a major paradigm shift in our understanding of the development of the MPS [172]. Progenitors of F4/80+ macrophages arise sequentially in the yolk sac (d8), AGM (d10.5), foetal liver (d10-12) and bone marrow before birth (d18-22), with widespread distribution throughout developing organs such as the brain [165,170]. An early origin from aortic endothelial stem cells and other mesenchymal cells has been established [173] and colony stimulating factor1 (CSF-1)- responsive precursors of F4/80+ macrophages can be detected as early as d4 (SG, unpublished observations). Yolk sac- derived tissue-resident macrophages persist in many tissues such as the brain and epidermis throughout adult life, turning over locally and depending on different transcription factors such as Myb from those deriving from bone marrow [174,175], which populates organs of high turnover such as the gut. After birth, bone marrow derived blood monocytes are the main source of recruitment on demand such as inflammation, infection, tissue injury, metabolic needs and malignancy. Macrophage populations in adult organs are therefore chimaeras of varying embryonic and bone marrow origin. In several species, including humans, the CSF1R provides a pan monocytic lineage-determining surface marker which can be used to isolate subpopulations of monocytes and some DCs for further study, in conjunction with CD14 and CD16 antigens [176,177]. The mechanisms of circulation, tissue distribution and migration in the foetus are not clear. After birth, bone marrow derived monocytes use distinct adhesion molecules such as Beta2 integrins [178], L selectin and CD31 during constitutive and induced extravasation and re-entry [179]. Monocyte/macrophage reserves can be mobilized from the spleen into blood and, during inflammation from the peritoneal cavity to draining lymph nodes or injured liver [180,181]. The niche for adhesion and repopulation may involve ligands induced on endothelium, extracellular matrix or other cell types, e.g., hepatic stellate cells or epithelium [182]. Blood monocytes can be delivered directly to the brain from bone marrow in the skull or via a leaky blood brain barrier [183]. Monocytes recruited from blood contribute to the formation of osteoclast giant cells in bone and to MNGCs in granuloma formation during infection [184]. Unlike macrophages, DCs recirculate from the lymphatic circulation via the thoracic duct [185]. Recent studies have demonstrated that during stress such as fasting, blood monocytes can return to bone marrow for subsequent release into blood upon refeeding [179].
3.7. Distinct Properties of Elicited Monocyte-Derived Mononuclear Phagocytes
Depending on the nature of acute or chronic stimuli, metabolic, inflammatory, infectious, immune or malignant, circulating monocytes and newly recruited tissue macrophages display a range of altered phenotypes, reflecting their particular tissue environment. For example, sterile crystalline materials that are poorly degradable, accumulate in lysosomes and stimulate foreign body giant cell formation [186]; bacteria, viruses and helminths induce distinct cytokine responses and the formation of Th1 or Th2 types of granulomata; microbial products enhance a respiratory burst; uptake of apoptotic or necrotic cells inhibit or enhance inflammation, respectively; immune complexes and cytokines can enhance or suppress DC, T and B cell activities. The concomitant responses of activated granulocytes, NK, innate lymphoid cells, DCs, CD4 and CD8 T cells can exacerbate and may exceed homeostatic limits, contributing to altered monocyte/macrophage proliferation, viability, clearance, secretory, cytotoxic and repair functions. Below, we draw attention to some of the complex cellular aspects of these induced mononuclear phagocyte responses.
3.8. Re-Enter Complement
After earlier Nobel awards for complement and antibodies, research on humoral immunity centred almost exclusively on the characterisation of plasma proteins. The later discovery of an “Alternative Complement pathway” and lectin-like recognition actually predated “classical” antibody-dependent activation in evolution. Progress in defining opsonic Fc and Complement receptors on myeloid leukocytes provided an important link to cellular immunity, both innate and adaptive. Proteolytic cascades of complement, coagulation and kinin pathways regulated phagocyte migration, cell activation and target cell cytolysis via plasma membrane receptors and peptides. Analogous plasma proteins such as Mannose binding lectin (MBL) [187], calreticulin [188] and pentraxins [189], also serve as humoral Pattern Recognition receptors. The major source of many of these plasma proteins is the liver hepatocyte, but Harvey Colten and colleagues showed that macrophages were able to produce small amounts of all complement proteins during development, compatible with functions in local tissue environments [190]. Selected resident macrophages, for example microglia in the CNS, express C1Q which contributes to sculpting of synapses by poorly understood mechanisms [191]. Recent discoveries, reviewed by Wright and Kemper, have shown that complement also plays an unexpected role intracellularly, in innate and adaptive autoimmunity [192]. These involve T lymphocytes, as well as mononuclear phagocytes and DCs, and play a role in Toll-like, TNF-related and mitochondrial antiviral-signaling protein (MAVS) recognition and local responses. This discovery has rekindled interest in the role of locally produced complement as an integral part of cellular interactions during inflammasome activation, immunity and infection, as well as in metabolic homeostasis, development and malignancy.
4. Cellular Functions of Mononuclear Phagocytes
4.1. Recognition, Uptake and Degradation
Although many plasma membrane receptors and intracellular sensing mechanisms have been identified (Figure 5), the ability of MPs to discriminate healthy from altered self at the cell surface or within intracellular membrane -bound compartments and the cytosol, remain poorly defined.
These myeloid cells are more promiscuous than lymphoid cells and less dependent on MHC restricted selection of foreign or auto-antigens. Although the chemical composition of the walls of micro-organisms can be detected by a broad range of protein, carbohydrate, lipid and nucleic acid receptors, quality control of biosynthetic and intracellular transport products is less defined. We tend to fall back on abstractions such as “danger” [193] or “pattern recognition receptors” [194]; what can be stated with confidence is that there are few holes in the MP repertoire. The efficient clearance of particulates and apoptotic and necrotic cells by “professional” phagocytes is legendary [195], and “don’t eat me “ signals such as signal-regulatory protein α (Sirpα)-CD47 interactions have stimulated immunotherapeutic interest of oncologists [196]; apart from the elegant early studies of Silverstein [197] and the ultrastructural studies by Grinstein and their colleagues [198], we still have little insight into molecular aspects of the phagocytic synapse [199,200], with or without opsonization, and the regulation of cargo internalisation. Antibody- dependent enhancement of inflammation or infection via FcR and CR are important aspects of viral pathogenesis [201,202,203,204]. Also intriguing, is the ability of microglia to sculpt neuronal synapses and dendrites [205] and of stromal macrophages in bone marrow to ingest erythroid cell nuclei without compromising target cell viability [206]. Hidalgo and colleagues have demonstrated the uptake of effete mitochondria, by a heterophagic process, analogous to autophagy (Figure 6) [207]. Remarkably, intact mitochondria can be interchanged between adipocytes and macrophages without loss of integrity or function [208]. Various names have been coined to stress variants of phagocytosis such as efferocytosis, pyroptosis, phagoptosis, ferroptosis, neuronophagy and mitophagy. Apart from phago- and endolysosomal fusion, acidification, digestion and membrane recycling during intracellular invasion of macrophages are important aspects of pathogenesis [209] and determine immune cross presentation and peptide loading of MHC molecules by DCs [210] and monocytes [211].
The biochemical mechanisms that control protein degradation by ubiquitination and delivery to proteosomes have previously attracted Nobel awards to Hershko and Ciechanover [215]; new methods have been developed to degrade selected cellular proteins for experimental and possible therapeutic purposes [216]. Finally, we stress the importance of endocytosis in lipid metabolism, recognized by Nobel awards to Brown and Goldstein, and the recycling of iron and other essential nutrients [217,218].
4.2. Biosynthesis, Gene Expression, Metabolism, Cell Activation and Secretion
After terminal differentiation, MP shut down DNA synthesis, but continue to express a wide range of mRNAs and proteins becoming highly active secretory cells [219] as well as phagocytes. Upon activation by innate and immune stimuli they adapt to local and systemic environments to perform various homeostatic, inflammatory and immune trophic and effector functions (Figure 7).
Tissue resident cells are long lived whereas elicited cells turn over more rapidly and are primed to generate more proinflammatory and cytotoxic radicals after further stimulation. Their secretory products include lysozyme [223], a range of neutral proteinases [224] to activate plasma cascades of complement, coagulation and kinin generation, as well as arachidonate metabolites, leukotrienes, prostaglandins and their derivatives. In addition, they produce chemokines, pro- and anti-inflammatory cytokines and growth factors for lymphohaematopoietic and endothelial cells [225], as well as protease inhibitors such as TIMP, alpha1 anti trypsin and alpha2 macroglobulin. Activated macrophages also secrete collagenase and elastase, modify the extracellular matrix [226], and regulate fibroblast growth and repair. Their energy requirements of ATP are met by glycolysis, through the Krebs cycle and/or oxidative phosphorylation through mitochondrial respiration. The pioneering metabolic studies by Nobel laureates Otto Warburg [227], Hans Krebs and Fritz Lipmann [228] are currently undergoing a renaissance in immunology [229].
Macrophage phenotype signatures are associated with a spectrum of polarized functional states [221,230]. Markers of innate stimulation include enhanced expression of MARCO and CD200 [231]. Classical activation by interferon gamma, a product of CD4 Th1 lymphocytes and NK cells, induces a respiratory burst after local triggering, for example phagocytosis, to generate reactive oxygen and nitrogen radicals by activation of NADPH dehydrogenase [232] and iNOS [233], respectively. The vital roles of oxygen and nitrogen in metabolism have also been recognized by recent Nobel prizes.
An alternative activation pathway of macrophages [176,221,234,235] associated with allergy and nematode infestation and Th2 immunity, is mediated by IL-4/13 cytokines, which upregulate selected MHCII molecules, production of arginase and Chitinase-like proteins, and promote repair and fibrosis. Other macrophage functional states are associated with anti-inflammatory stimuli (IL10, TGF beta, corticosteroid, prostaglandin E2) or immunoregulatory immune complexes. Tumour-associated macrophages (TAM) promote malignancy and metastasis [236,237,238]; temporal and spatial analyses of RNA and protein expression reveal considerable heterogeneity in TAM subtypes, relevant to potential immunotherapy [239].
Several hundred rare genetic disorders have been discovered which result in familial auto-inflammatory and auto-immune syndromes [69]. IL-1 beta release and pyroptosis are associated with periodic fevers and a range of clinical manifestations, and can be treated with IL-1 Receptor Antagonist [240]. The discovery of inflammasome activation by the late Jürg Tschopp [241], and of intracellular Nod-like, RIG-like and DNA sensing pathways, has spawned enormous activity in this field [242,243]. Life threatening hyperinflammatory syndromes are associated with CD4 lymphocyte reconstitution during antiretroviral treatment of HIV/ AIDS and severe pulmonary infections with SARS-CoV-2 [244,245]. It is worth noting that the term “cytokine storm” is a misnomer since it encompasses many macrophage secretory products. Dexamethasone and several other repurposed drugs used for COVID-19 therapy act primarily on dysregulated macrophages, and the type1 interferon pathway has been implicated in genetic predisposition to critical outcome of SARS-CoV-2 infection [68,246,247]. The potential role of MPs in Long COVID sequelae should also be considered [248].
4.3. Cellular Immunity and Granuloma Formation
Mononuclear phagocytes play a prominent role in the formation and persistence of organized structures known as granulomas, which are not tumours, but assemblies of heterogeneous monocytes, macrophages and DCs, together with other myeloid and lymphoid cells, fibroblasts and blood vessels, embedded in an extracellular matrix [249,250,251]. Their morphology has been recognized since early descriptions by pathologists and they cover a range of immunologic and inflammatory host responses to infection, foreign or host-derived non-degradable materials and autoimmune disorders [252]. Although not malignant, per se, they have features in common with tumour-associated macrophages and can inflict considerable tissue injury upon their local environment, promote fibrosis and even progress to true tumour formation, as in Schistosomiasis [253]. A characteristic feature is macrophage multinucleation, the result of cell fusion induced by Th1 and Th2 lymphocytes and their secretory products [254]. Metchnikoff recognized MNGCs, known as Langhans giant cells, containing intracellular M. tuberculosis, with evident impairment of mycobacterial division [8,255]. There has been recent significant progress in studying the mechanism of fusion and its impact on the macrophage phenotype in a variety of cellular model systems, in vitro [256] and in autoimmune granulomatosis, in vivo [251]. This is a fascinating field for further study of cellular biology and immunology, relevant to the impact of environmental pollutant particles on human health [257].
5. Discussion
Our epigraph above [258] highlights the importance of homeostasis [259] and evolution in considering the roles of macrophages and closely related DCs and MNGCs in pathophysiology and natural selection. We have traced the growth of knowledge from the emergence of these cells as a Mononuclear Phagocyte System, during development and throughout life, and described selected cellular and molecular properties. These widely distributed phagocytes recognize and interact with every other cell type of the body in their micro-environment, including microbes and foreign particulates, through plasma membrane contact, internalisation and secretion (Figure 8). Reciprocal interactions between the MPS and neuroendocrine [260], cardiovascular [261] and gastrointestinal systems [262], for example, extend the role of mononuclear phagocytes beyond host defence, inflammation and immunity. Within the lymphohaematopoietic system it is important to recognize that macrophages are integral parts of all immunity, whether innate or adaptive, cellular or humoral, and participate in many disease processes as monocytes, tissue macrophages, DCs or MNGCs. These “non-immune” diseases include metabolic processes such as atherosclerosis [263] and fatty liver disease [264], neurodegeneration such as Alzheimer disease [265] and malignancy [237]. We have stressed the value of genetic, cellular and molecular analysis in deciphering and manipulating the MPS in situ and in vitro.
Although it is well recognized that macrophage-like cells are ancient, long preceding the emergence of lymphocytes, clonal selection and gene rearrangement, there has been much less investigation of myeloid cell evolution. Compelling studies by Max Cooper [266] and colleagues have brought insights into lymphocyte evolution; presumably the role of MHC molecules in peptide capture and antigen presentation by DCs should coincide with evolution of lymphocyte activation. Comparative genomics and DNA analysis are making it possible to trace the origin and migration of human populations and the role of pandemics in evolution. The recent Nobel award to Paabo [267] and the studies of Quintana-Murci [268] and Barreiro [269] suggest that the MPS contributed significantly to selection by infectious disease. The hypothesis by Lynn Margulis [270,271] that mitochondria and chloroplasts are endosymbionts of captured bacteria is now widely accepted and retention of retroviral sequences in the human genome [272] provides further evidence for the role of gene transfer and selection in evolution. Experiments to model the origin of eukaryotic cells [273], reveal cells which display macrophage-like features.
Apart from the specific Nobel prizes in the macrophage field in 1908 and 2011, we have shown how important many other awards in immunology and related subjects, including technological advances, have been in promoting and documenting progress of research in MPs. This acknowledgement attests to the wider legacy of the Nobel Awards in cellular, biomedical and chemical research.
6. Conclusion
We have outlined some of the evidence that the MPS is a unique dispersed organ consisting of a family of closely related cells, interacting with every other system in the body, consistent with its primary role in physiological homeostasis, cellular immunity and disease. Perturbation of its trophic cellular and molecular functions may contribute to the mechanisms of ageing and malignancy and provide opportunities for therapy, through genetic and immunologic manipulation. We have stressed the direct and indirect benefits of many Nobel awards to achievements in MPS research and noted some of the factors that promote scientific advances in this, as in other fields, where fundamental discoveries can lead to clinical applications. Our historic perspective has confirmed the international nature of scientific discovery and its dependence on institutional excellence, ready access to education, free movement and exchange of ideas. Finally, we suggest that the evolutionary origins of macrophage diversification need more research, to establish its rightful role in biology, beyond immunity.
Contributions: Conceptualization, SG and SHEK; writing—original draft preparation, SG, AR and SHEK; writing—review and editing, SG, AR and SHEK.; visualization, AR and SG. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Acknowledgements
SG would like to dedicate this review to the late Zanvil A. Cohn, mentor and friend. We thank all colleagues and collaborators, too many to cite, who have shared, in discussions and in print, over the past 50 years.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
ACAID: Anterior Chamber-Associated Immune Deviation, AGM: Aorta-gonad-mesonephros, APC: antigen-presenting cell, ATP: adenosine triphosphate, BCG: Bacillus Calmette-Guérin, CNS: central nervous system, CR: complement receptor, CSF: colony stimulating factor, DCs: dendritic cells, FACS: fluorescence-activated cell sorting, FcR: Fc receptor, HIV: human immunodeficiency virus, iNOS: inducible nitric oxide synthase, iPSCs: induced pluripotent stem cells, ITAM: immunoreceptor tyrosine-based activation motif, ITIM: immunoreceptor tyrosine-based inhibitory motif, MARCO: Macrophage receptor with collagenous structure, MAVS: Mitochondrial antiviral-signaling protein, MBL: mannose-binding lectin, MHC: major histocompatibility complex, MNGCs: multi-nucleated giant cells, MPs: mononuclear phagocytes, MPS: mononuclear phagocyte system, NADPH: Nicotinamide adenine dinucleotide phosphate, NK: natural killer cell, NO: nitric oxide, RES: reticulo-endothelial system, ROS: reactive oxygen species, SIRP: Signal regulatory protein, SR: scavenger receptor, TAM: tumour-associated macrophage, TIMP: tissue inhibitor of metalloproteinases, TLR: toll-like receptor.
References
- Darwin, C. and G. Beer, On the Origin of Species. 2008: OUP Oxford.
- Harris, H. , The Birth of the Cell. 2000: Yale University Press.
- Ackerknecht, E.H. , Rudolf Virchow: doctor, statesman, anthropologist. 1953.
- Brock, T.D., G. L. Geison, and R.J. Dubos, Pasteur and Modern Science. 2014: Springer Berlin Heidelberg.
- Bernard, C. and L.L.S. University College, An Introduction to the Study of Experimental Medicine. 1865: Schuman.
- Cannon, W.B. , The Wisdom of the Body. 1939: W.W. Norton, Incorporated.
- Metchnikoff, E. , Lectures on the Comparative Pathology of Inflammation: Delivered at the Pasteur Institute in 1891. 1968: Dover Publications.
- Kaufmann, S.H. , Immunology’s foundation: the 100-year anniversary of the Nobel Prize to Paul Ehrlich and Elie Metchnikoff. Nat Immunol, 2008. 9(7): p. 705-12.
- Ehrlich, P. Paul Ehrlich - Nobel Lecture. Partial Cell Functions 1908 [cited 2024 17.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1908/ehrlich/lecture/.
- Himmelweit, F. , The Collected Papers of Paul Ehrlich: in Four Volumes Including a Complete Bibliography. 2017: Elsevier Science.
- Ehrlich, P. , et al., The Collected Papers of Paul Ehrlich: Immunology and cancer research. 1957: Pergamon P.
- Ehrlich, P., T. Himmelweit, and M. Marquardt, The Collected Papers of Paul Ehrlich: Chemotherapy. 1960: Pergamon Press.
- Metchnikoff, E. Ilya Mechnikov - Nobel Lecture. On the Present State of the Question of Immunity in Infectious Diseases 1908 [cited 2024 17.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1908/mechnikov/lecture/.
- Metchnikoff, O. , Life of Elie Metchnikoff 1845-1916. 1921: Houghton Mifflin, New York.
- Behring, E.v. Emil von Behring - Nobel Lecture. Serum Therapy in Therapeutics and Medical Science 1901 [cited 2024 17.06.2024]; Available from: https://www.nobelprize.org/prizes/medicine/1901/behring/lecture/.
- Brock, T.D. , Robert Koch: A Life in Medicine and Bacteriology. 1999: ASM Press.
- Ehrlich, B. , The Brain in Search of Itself: Santiago Ramón Y Cajal and the Story of the Neuron. 2022: Farrar, Straus and Giroux.
- Dormandy, T. , The White Death: A History of Tuberculosis. 1999: Hambledon Press.
- Paolicelli, R.C. , et al., Microglia states and nomenclature: A field at its crossroads. Neuron, 2022. 110(21): p. 3458-3483.
- Vikhanski, L. , Immunity: How Elie Metchnikoff Changed the Course of Modern Medicine. 2016: Chicago Review Press Incorporated.
- Cavaillon, J.M. , The historical milestones in the understanding of leukocyte biology initiated by Elie Metchnikoff. J Leukoc Biol, 2011. 90(3): p. 413-24.
- Clarke, D.J.B. , et al., Gene set predictor for post-treatment Lyme disease. Cell Reports Medicine, 2022. 3(11).
- Rasmussen, N. , René Dubos, the Autochthonous Flora, and the Discovery of the Microbiome. J Hist Biol, 2022. 55(3): p. 537-558.
- Stambler, I.S. , Elie Metchnikoff- The founder of longevity science and a founder of modern medicine: In honor of the 170th anniversary Adv Gerontol, 2015. 28(2): p. 207-17.
- Maisonneuve, P., E. Metchnikoff, and P.P.E. Roux, The Experimental Prophylaxis Of Syphilis. 2015: Creative Media Partners, LLC.
- Tolstoy, L. , The Death of Ivan Ilyich. 2020: Open Road Media.
- Rietschel, E.T. and J.-M. Cavaillon, Richard Pfeiffer and Alexandre Besredka: creators of the concept of endotoxin and anti-endotoxin. Microbes and Infection, 2003. 5(15): p. 1407-1414.
- Schama, S. , Foreign Bodies: Pandemics, Vaccines and the Health of Nations. 2023: Simon & Schuster UK.
- Bordet, J. The Nobel Prize in Physiology or Medicine 1919. 1919 [cited 2024 18.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1919/summary/.
- Hansson, N. and D. Angetter-Pfeiffer, Remembering innovative paediatrician Clemens von Pirquet on the 150th anniversary of his birth. Acta Paediatrica. n/a(n/a).
- NobelPrize.org. Charles Richet - Nobel Lecture. [cited 2024 25.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1913/richet/lecture/.
- Nijweide, P.J., E. H. Burger, and J.H. Feyen, Cells of bone: proliferation, differentiation, and hormonal regulation. Physiological Reviews, 1986. 66(4): p. 855-886.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1945. [cited 2024 15.07.]; Available from: https://www.nobelprize.org/prizes/medicine/1945/summary/.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1999. [cited 2024 15.07.]; Available from: https://www.nobelprize.org/prizes/medicine/1999/summary/.
- Hamann, J. and A.G. Petrenko, Introduction: History of the Adhesion GPCR Field. Handb Exp Pharmacol, 2016. 234: p. 1-11.
- Brown, G.D. and S. Gordon, Immune recognition. A new receptor for beta-glucans. Nature, 2001. 413(6851): p. 36-7.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 2011. [cited 2024 15.07.]; Available from: https://www.nobelprize.org/prizes/medicine/2011/summary/.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1980. [cited 2024 15.07.]; Available from: https://www.nobelprize.org/prizes/medicine/1980/summary/.
- Dzierzak, E. and E. de Pater, Regulation of Blood Stem Cell Development. Curr Top Dev Biol, 2016. 118: p. 1-20.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 2019. [cited 2024 15.07.]; Available from: https://www.nobelprize.org/prizes/medicine/2019/summary/.
- Wake, K. , Karl Wilhelm Kupffer And His Contributions To Modern Hepatology. Comp Hepatol, 2004. 3 Suppl 1(Suppl 1): p. S2.
- Kodama, T. , et al., Type I macrophage scavenger receptor contains α-helical and collagen-like coiled coils. Nature, 1990. 343(6258): p. 531-535.
- Stanley, E.R. and V. Chitu, CSF-1 receptor signaling in myeloid cells. Cold Spring Harb Perspect Biol, 2014. 6(6).
- Pappenheim, A. and A. Ferrata, Über die verschiedenen lymphoiden Zellformen des normalen und pathologischen Blutes. Vol. 10. 1911: Klinkhardt.
- Beutler, B. and A. Cerami, The biology of cachectin/TNF--a primary mediator of the host response. Annu Rev Immunol, 1989. 7: p. 625-55.
- Henson, P.M. and D.L. Bratton, Recognition and removal of apoptotic cells, in Phagocyte-pathogen interactions: macrophages and the host response to infection, D.G. Russell and S. Gordon, Editors. 2009, ASM Press: Washington, D.C. p. 341-365.
- Crocker, P.R. and S. Gordon, Isolation and characterization of resident stromal macrophages and hematopoietic cell clusters from mouse bone marrow. J Exp Med, 1985. 162(3): p. 993-1014.
- Nagata, S. , Apoptosis and Clearance of Apoptotic Cells. Annu Rev Immunol, 2018. 36: p. 489-517.
- Rossi, F. , The O2- -forming NADPH oxidase of the phagocytes: nature, mechanisms of activation and function. Biochim Biophys Acta, 1986. 853(1): p. 65-89.
- Waddington, C.H. Epigenetics and evolution. in Symp Soc Exp Biol. 1953.
- Blackburn, D.G. and J.R. Stewart, Morphological research on amniote eggs and embryos: An introduction and historical retrospective. Journal of Morphology, 2021. 282(7): p. 1024-1046.
- Bessis, M. , L’ilot erythroblastique. Unite functionelle de la moelle osseuse. Rev Hematol, 1958. 13(1): p. 8-11.
- Kaufmann, S.H.E. , Immunology’s Coming of Age. Front Immunol, 2019. 10: p. 684.
- Silverstein, A.M. , Paul Ehrlich’s Receptor Immunology: The Magnificent Obsession. 2001: Elsevier Science.
- Barry Kay, A. , Paul Ehrlich and the Early History of Granulocytes, in Myeloid Cells in Health and Disease. 2017. p. 1-15.
- Aschoff, L. , Das reticulo-endotheliale system. Ergeb Inn Med Kinderheilk, 1924. 26: p. 1-118.
- Cannon, W.B. , The Way of an Investigator: A Scientist’s Experiences in Medical Research. 1984: W.W. Norton.
- Tan, S.Y. and A. Yip, Hans Selye (1907-1982): Founder of the stress theory. Singapore Med J, 2018. 59(4): p. 170-171.
- Dunnill, M.S. , The Plato of Praed Street: The Life and Times of Almroth Wright. 2000: Royal Society of Medicine Press.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1972. 1972 [cited 2024 18.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1972/summary/.
- Mastellos, D.C., G. Hajishengallis, and J.D. Lambris, A guide to complement biology, pathology and therapeutic opportunity. Nature Reviews Immunology, 2024. 24(2): p. 118-141.
- Janeway, C. , et al., Immunobiology: The Immune System in Health and Disease. 2005: Garland Science.
- MacPhail, T. , Allergic: How Our Immune System Reacts to a Changing World. 2023: Penguin Books Limited.
- Shaw, B. and D. Laurence, The Doctor’s Dilemma. 1987: Penguin Adult.
- Berry, A. and J. Browne, Mendel and Darwin. Proc Natl Acad Sci U S A, 2022. 119(30): p. e2122144119.
- Bearn, A.G. , Archibald Garrod and the Individuality of Man. 1993: Clarendon Press.
- Roos, D. and M. de Boer, Molecular diagnosis of chronic granulomatous disease. Clin Exp Immunol, 2014. 175(2): p. 139-49.
- Bastard, P. , et al., Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science, 2020. 370(6515).
- Savic, S., J. Coe, and P. Laws, Autoinflammation: Interferonopathies and Other Autoinflammatory Diseases. Journal of Investigative Dermatology, 2022. 142(3, Part B): p. 781-792.
- Grabowski, G.A. and D. Hughes, Emerging Therapies, in Lysosomal Storage Disorders. 2022. p. 287-294.
- Rous, P. and J.W. Beard, Selection with the magnet and cultivation of reticulo-endothelial cells (Kupffer cells). J Exp Med, 1934. 59(5): p. 577-91.
- Sabin, F.R. , Cunningham, Robert S., Doan, Charles A., Studies of the Blood in Experimental Tuberculosis: The Monocyte-Lymphocyte Ratio; The Anemia-Leucopenia Phase. National Tuberculosis Association, 1926(Transactions of the 22nd Annual Meeting of the National Tuberculosis Association).
- Dannenberg, A.M. , Cellular hypersensitivity and cellular immunity in the pathogensis of tuberculosis: specificity, systemic and local nature, and associated macrophage enzymes. Bacteriological reviews, 1968. 32(2): p. 85-102.
- NobelPrize.org. Karl Landsteiner - Nobel Lecture. On Individual Differences in Human Blood [cited 2024 25.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1930/landsteiner/lecture/.
- Landsteiner, K. and M.W. Chase, Experiments on Transfer of Cutaneous Sensitivity to Simple Compounds. Proceedings of the Society for Experimental Biology and Medicine, 1942. 49(4): p. 688-690.
- Fleming, A. and A.E. Wright, On a remarkable bacteriolytic element found in tissues and secretions. Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character, 1922. 93(653): p. 306-317.
- Leonard, C. , Alexander Fleming, 1881-1955. Biogr. Mems Fell. R. Soc.2117–127, 1956.
- Fleming, A. , On the antibacterial action of cultures of a penicillium, with special reference to their use in the isolation of B. influenzae. 1929. Bull World Health Organ, 2001. 79(8): p. 780-90.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1945. [cited 2024 25.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1945/summary/.
- Florey, H. , General Pathology. 1970: W. B. Saunders Company.
- Mackaness, G.B. , The action of drugs on intracellular tubercle bacilli. The Journal of Pathology and Bacteriology, 1952. 64(3): p. 429-446.
- Gowans, J.L., E. J. Knight, and H.W. Florey, The route of re-circulation of lymphocytes in the rat. Proceedings of the Royal Society of London. Series B. Biological Sciences, 1964. 159(975): p. 257-282.
- Harris, H. , Role of chemotaxis in inflammation. Physiol Rev, 1954. 34(3): p. 529-62.
- Billingham, R.E., L. Brent, and P.B. Medawar, ‘Actively Acquired Tolerance’ of Foreign Cells. Nature, 1953. 172(4379): p. 603-606.
- Medawar, P.B. , Immunological tolerance. Science, 1961. 133(3449): p. 303-6.
- Porterfield, J.S. , The National Institute for Medical Research, Mill Hill. Archives of Virology, 1995. 140(7): p. 1329-1336.
- Isaacs, A., J. Lindenmann, and C.H. Andrewes, Virus interference. I. The interferon. Proceedings of the Royal Society of London. Series B - Biological Sciences, 1957. 147(927): p. 258-267.
- Armstrong, J.A. and P.D. Hart, Response of cultured macrophages to Mycobacterium tuberculosis, with observations on fusion of lysosomes with phagosomes. J Exp Med, 1971. 134(3 Pt 1): p. 713-40.
- Humphrey, J.H. and D. Grennan, Different macrophage populations distinguished by means of fluorescent polysaccharides. Recognition and properties of marginal-zone macrophages. Eur J Immunol, 1981. 11(3): p. 221-8.
- Mitchison, T.J. and Beverly, C.L., Nicholas Avrion Mitchison. 5 May 1928 — 28 December 2022. Biogr. Mems Fell. R. Soc., 2024.
- Moberg, C.L. and R.M. Steinman, James Gerald Hirsch. Biogr Mem Natl Acad Sci, 2004. 84: p. 182-203.
- Steinman, R.M. and C.L. Moberg, Zanvil Alexander Cohn 1926-1993. J Exp Med, 1994. 179(1): p. 1-30.
- Moberg, C.L. , An appreciation of Ralph Marvin Steinman (1943-2011). J Exp Med, 2011. 208(12): p. 2337-42.
- Moberg, C.L. , René Dubos, Friend of the Good Earth: Microbiologist, Medical Scientist, Environmentalist. 2005: ASM Press.
- Woodruff, H.B. , Selman A. Waksman, winner of the 1952 Nobel Prize for physiology or medicine. Appl Environ Microbiol, 2014. 80(1): p. 2-8.
- Pringle, P. , Experiment Eleven: Dark Secrets Behind the Discovery of a Wonder Drug. 2012: Bloomsbury Publishing.
- Hirsch, J.G. , Bactericidal action of histone Journal of Experimental Medicine, 1958. 108(6): p. 925-944.
- Horwitz, M.A. , The Legionnaires’ disease bacterium (Legionella pneumophila) inhibits phagosome-lysosome fusion in human monocytes. J Exp Med, 1983. 158(6): p. 2108-26.
- Jones, T.C. and J.G. Hirsch, The interaction between Toxoplasma gondii and mammalian cells. II. The absence of lysosomal fusion with phagocytic vacuoles containing living parasites. J Exp Med, 1972. 136(5): p. 1173-94.
- Johnson, W.D., Jr., B. Mei, and Z.A. Cohn, The separation, long-term cultivation, and maturation of the human monocyte. J Exp Med, 1977. 146(6): p. 1613-26.
- NobelPrize.org. George E. Palade - Facts. [cited 2024 20.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1974/palade/facts/.
- De Duve, C. , Vital Dust: The Origin and Evolution of Life on Earth. 1995: Basic Books.
- Rabinovitch, M. , Professional and non-professional phagocytes: an introduction. Trends in Cell Biology, 1995. 5(3): p. 85-87.
- Kaplan, G. and Z.A. Cohn, The immunobiology of leprosy. Int Rev Exp Pathol, 1986. 28: p. 45-78.
- Kaplan, G. and Z.A. Cohn, Leprosy and cell-mediated immunity. Curr Opin Immunol, 1991. 3(1): p. 91-6.
- McElrath, M.J., R. M. Steinman, and Z.A. Cohn, Latent HIV-1 infection in enriched populations of blood monocytes and T cells from seropositive patients. J Clin Invest, 1991. 87(1): p. 27-30.
- Van Furth, R. and L. Conference on Mononuclear Phagocytes, Mononuclear Phagocytes. Edited by Ralph Van Furth. 1970: Blackwell Scientific Publications (C1970).
- Van Furth, R. , Mononuclear Phagocytes in Immunity, Infection and Pathology. 1975: Mosby, Incorporated.
- van Furth, R. , Mononuclear Phagocytes: Functional Aspects. 1980: Springer Netherlands.
- van Furth, R. , Mononuclear Phagocytes: Characteristics, Physiology and Function. 1985: Springer Netherlands.
- van Furth, R. , et al., The mononuclear phagocyte system: a new classification of macrophages, monocytes, and their precursor cells. Bull World Health Organ, 1972. 46(6): p. 845-52.
- Tauber, A.I. , The birth of immunology: III. The fate of the phagocytosis theory. Cellular Immunology, 1992. 139(2): p. 505-530.
- Gowans, J.L. , First Medawar prize lecture. The recirculating small lymphocyte. Transplant Proc, 1991. 23(1 Pt 1): p. 7-8.
- Volkman, A. and J.L. Gowans, The origin of macrophages from bone marrow in the rat. Br J Exp Pathol, 1965. 46(1): p. 62-70.
- Marchesi, V.T. and J.L. Gowans, The migration of lymphocytes through the endothelium of venules in lymph nodes: an electron microscope study. Proceedings of the Royal Society of London. Series B. Biological Sciences, 1964. 159: p. 283 - 290.
- Miller, J.F.A.P. , Immunological function of the thymus The Lancet, 1961. 278(7205): p. 748-749.
- Miller, J.F. , The discovery of thymus function and of thymus-derived lymphocytes. Immunol Rev, 2002. 185: p. 7-14.
- Nature Milestones: T cells. 2022 [cited 2024 20.06.]; Available from: https://www.nature.com/immersive/d42859-022-00032-7/index.html.
- Gitlin, A.D. and M.C. Nussenzweig, Immunology: Fifty years of B lymphocytes. Nature, 2015. 517(7533): p. 139-141.
- Zoltan Fehervari, Y.B. , Cosma Dellisanti, Nicholas Bernard & Jamie D. K. Wilson. Nature Milestones in Antibodies. 2016 [cited 2024 20.06.]; Available from: https://www.nature.com/articles/d42859-021-00068-1.
- Burnet, M. , Self and Not-Self: Cellular Immunology Book One. 1969: Cambridge University Press.
- Burnet, F.M. , Immunological recognition of self. Science, 1961. 133(3449): p. 307-11.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1996. [cited 2024 20.6.]; Available from: https://www.nobelprize.org/prizes/medicine/1996/summary/.
- Metcalf, D. , Growth and Differentiation Factors. Microbiol Spectr, 2016. 4(4).
- Pixley, F.J. and E.R. Stanley, CSF-1 regulation of the wandering macrophage: complexity in action. Trends Cell Biol, 2004. 14(11): p. 628-38.
- Mackaness, G.B. , The Immunological Basis of Acquired Cellular Resistance. J Exp Med, 1964. 120: p. 105-20.
- Sorg, C. and B.R. Bloom Products of activated lymphocytes: I. The use of radiolabeling techniques in the characterization and partial purification of the migration inhibitory factor of the guinea pig Journal of Experimental Medicine, 1973. 137(1): p. 148-170.
- Dinarello, C.A. , Historical insights into cytokines. Eur J Immunol, 2007. 37 Suppl 1(Suppl 1): p. S34-45.
- Mosmann, T.R. and R.L. Coffman, TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol, 1989. 7: p. 145-73.
- Nathan, C.F. , et al., Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. Journal of Experimental Medicine, 1983. 158(3): p. 670-689.
- Gray, P.W. and D.V. Goeddel, Structure of the human immune interferon gene. Nature, 1982. 298(5877): p. 859-863.
- Moberg, C.L. , The discovery of dendritic cells. Journal of Experimental Medicine, 2021. 218(6).
- Steinman, R. , Dendritic cells: linking innate to different forms of adaptive immunity in The Biomedical & Biomedical Life Sciences Collection. 2009, Henry Stewart Talks.
- Unanue, E.R. and B.A. Askonas, Persistence of immunogenicity of antigen after uptake by macrophages. J Exp Med, 1968. 127(5): p. 915-26.
- Silverstein, S.C., R. M. Steinman, and Z.A. Cohn, Endocytosis. Annu Rev Biochem, 1977. 46: p. 669-722.
- Elliott, T. , et al., Structural requirements for the peptide-induced conformational change of free major histocompatibility complex class I heavy chains. European Journal of Immunology, 1992. 22(8): p. 2085-2091.
- Bjorkman, P.J. , et al., Structure of the human class I histocompatibility antigen, HLA-A2. Nature, 1987. 329(6139): p. 506-512.
- Steinman, R.M. and Z.A. Cohn, Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med, 1973. 137(5): p. 1142-62.
- Nussenzweig, M.C. and R.M. Steinman, Contribution of dendritic cells to stimulation of the murine syngeneic mixed leukocyte reaction. J Exp Med, 1980. 151(5): p. 1196-212.
- Paul, W.E. , Dendritic cells bask in the limelight. Cell, 2007. 130(6): p. 967-70.
- Inaba, K. , et al., Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J Exp Med, 1992. 176(6): p. 1693-702.
- Merad, M. , et al., The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol, 2013. 31: p. 563-604.
- Bálint, Š. , et al., Supramolecular attack particles are autonomous killing entities released from cytotoxic T cells. Science, 2020. 368(6493): p. 897-901.
- Horton, C., K. Shanmugarajah, and P.J. Fairchild, Harnessing the properties of dendritic cells in the pursuit of immunological tolerance. Biomedical Journal, 2017. 40(2): p. 80-93.
- Qin, S. , et al., “Infectious” Transplantation Tolerance. Science, 1993. 259(5097): p. 974-977.
- Mellman, I. , et al., The cancer-immunity cycle: Indication, genotype, and immunotype. Immunity, 2023. 56(10): p. 2188-2205.
- Bearn, A.G., F. J. Dixon, and B. Benacerraf, Henry G. Kunkel 1916-1983. An appreciation of the man and his scientific contributions & a bibliography of his research papers. J Exp Med, 1985. 161(5): p. 869-95.
- Harris, H. , et al., Artificial heterokaryons of animal cells from different species. J Cell Sci, 1966. 1(1): p. 1-30.
- Klein, G., S. Friberg, Jr., and H. Harris, Two kinds of antigen suppression in tumor cells revealed by cell fusion. J Exp Med, 1972. 135(4): p. 839-49.
- Köhler, G. and C. Milstein, Continuous cultures of fused cells secreting antibody of predefined specificity. Nature, 1975. 256(5517): p. 495-7.
- Crumpton, M.J., Alan Frederick Williams 25 May 1945 - 9 April 1992. Biographical Memoirs of Fellows of the Royal Society, 2004. 50: p. 351-366.
- Springer, T. , et al., Mac-1: a macrophage differentiation antigen identified by monoclonal antibody. Eur J Immunol, 1979. 9(4): p. 301-6.
- Diamond, M.S. , et al., Binding of the integrin Mac-1 (CD11b/CD18) to the third immunoglobulin-like domain of ICAM-1 (CD54) and its regulation by glycosylation. Cell, 1991. 65(6): p. 961-971.
- Feldman, M. , et al., Anti-TNF alpha therapy is useful in rheumatoid arthritis and Crohn’s disease: analysis of the mechanism of action predicts utility in other diseases. Transplant Proc, 1998. 30(8): p. 4126-7.
- Austyn, J.M. and S. Gordon, F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur J Immunol, 1981. 11(10): p. 805-15.
- Gordon, S. and A. Pluddemann, Tissue macrophages: heterogeneity and functions. BMC Biol, 2017. 15(1): p. 53.
- Taylor, P.R. , et al., Macrophage receptors and immune recognition. Annu Rev Immunol, 2005. 23: p. 901-44.
- Lin, H.H. , et al., Ligands and Beyond: Mechanosensitive Adhesion GPCRs. Pharmaceuticals (Basel), 2022. 15(2).
- I, K.Y. , et al., Activation of Adhesion GPCR EMR2/ADGRE2 Induces Macrophage Differentiation and Inflammatory Responses via Gα(16)/Akt/MAPK/NF-κB Signaling Pathways. Front Immunol, 2017. 8: p. 373.
- Lin, H.H. , et al., The macrophage F4/80 receptor is required for the induction of antigen-specific efferent regulatory T cells in peripheral tolerance. J Exp Med, 2005. 201(10): p. 1615-25.
- Gordon, S., A. Plüddemann, and S. Mukhopadhyay, Plasma membrane receptors of tissue macrophages: functions and role in pathology. J Pathol, 2020. 250(5): p. 656-666.
- Zulu, M.Z. , et al., The Elusive Role of Placental Macrophages: The Hofbauer Cell. J Innate Immun, 2019: p. 1-10.
- Takahashi, K. and S. Yamanaka, Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell, 2006. 126(4): p. 663-76.
- Fejer, G. , et al., Nontransformed, GM-CSF-dependent macrophage lines are a unique model to study tissue macrophage functions. Proc Natl Acad Sci U S A, 2013. 110(24): p. E2191-8.
- Mass, E. , et al., Specification of tissue-resident macrophages during organogenesis. Science, 2016. 353(6304).
- Guilliams, M. and C.L. Scott, Liver macrophages in health and disease. Immunity, 2022. 55(9): p. 1515-1529.
- Amit, I., D. R. Winter, and S. Jung, The role of the local environment and epigenetics in shaping macrophage identity and their effect on tissue homeostasis. Nat Immunol, 2016. 17(1): p. 18-25.
- Ginhoux, F. , et al., Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science, 2010. 330(6005): p. 841-5.
- Hume, D.A., V. H. Perry, and S. Gordon, The mononuclear phagocyte system of the mouse defined by immunohistochemical localisation of antigen F4/80: macrophages associated with epithelia. Anat Rec, 1984. 210(3): p. 503-12.
- Lawson, L.J. , et al., Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience, 1990. 39(1): p. 151-70.
- Silverstein, S.C., J. Michl, and J.D. Loike, Studies of the Mechanism of Phagocytosis, in International Cell Biology 1980–1981: Papers Presented at the Second International Congress on Cell Biology Berlin (West), August 31 – September 5, 1980, H.G. Schweiger, Editor. 1981, Springer Berlin Heidelberg: Berlin, Heidelberg. p. 604-612.
- Yona, S. , et al., Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity, 2013. 38(1): p. 79-91.
- Mariani, S.A. , et al., Pro-inflammatory Aorta-Associated Macrophages Are Involved in Embryonic Development of Hematopoietic Stem Cells. Immunity, 2019. 50(6): p. 1439-1452.e5.
- Schulz, C. , et al., A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science, 2012. 336(6077): p. 86-90.
- Ginhoux, F. and M. Guilliams, Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity, 2016. 44(3): p. 439-449.
- Orsenigo, F. , et al., Unifying considerations and evidence of macrophage activation mosaicism through human CSF1R and M1/M2 genes. Cell Reports, 2024. 43(6).
- Ziegler-Heitbrock, L. , et al., Nomenclature of monocytes and dendritic cells in blood. Blood, 2010. 116(16): p. e74-80.
- Carlin, L. , et al., Functions and molecular mechanisms of patrolling monocytes. Vascular Pharmacology, 2012. 56: p. 328.
- Janssen, H. , et al., Monocytes re-enter the bone marrow during fasting and alter the host response to infection. Immunity, 2023. 56(4): p. 783-796.e7.
- Hossain, M. and P. Kubes, Innate immune cells orchestrate the repair of sterile injury in the liver and beyond. European Journal of Immunology, 2019. 49(6): p. 831-841.
- Bellingan, G.J. , et al., In vivo fate of the inflammatory macrophage during the resolution of inflammation: inflammatory macrophages do not die locally, but emigrate to the draining lymph nodes. J Immunol, 1996. 157(6): p. 2577-85.
- Bonnardel, J. , et al., Stellate Cells, Hepatocytes, and Endothelial Cells Imprint the Kupffer Cell Identity on Monocytes Colonizing the Liver Macrophage Niche. Immunity, 2019. 51(4): p. 638-654.e9.
- Cugurra, A. , et al., Skull and vertebral bone marrow are myeloid cell reservoirs for the meninges and CNS parenchyma. Science, 2021. 373(6553): p. eabf7844.
- Jacome-Galarza, C.E. , et al., Developmental origin, functional maintenance and genetic rescue of osteoclasts. Nature, 2019. 568(7753): p. 541-545.
- Alvarez, D., E. H. Vollmann, and U.H. von Andrian, Mechanisms and consequences of dendritic cell migration. Immunity, 2008. 29(3): p. 325-42.
- Towiwat, P., A. Chhana, and N. Dalbeth, The anatomical pathology of gout: a systematic literature review. BMC Musculoskeletal Disorders, 2019. 20(1): p. 140.
- Turner, M.W. , The role of mannose-binding lectin in health and disease. Mol Immunol, 2003. 40(7): p. 423-9.
- Sawaya, A.P. , et al., Calreticulin: a multifunctional protein with potential therapeutic applications for chronic wounds. Front Med (Lausanne), 2023. 10: p. 1207538.
- Mantovani, A. , et al., The long pentraxin PTX3: a paradigm for humoral pattern recognition molecules. Annals of the New York Academy of Sciences, 2013. 1285(1): p. 1-14.
- Colten, H.R. , Ontogeny of the human complement system: in vitro biosynthesis of individual complement components by fetal tissues. J Clin Invest, 1972. 51(4): p. 725-30.
- Wu, X. , et al., Complement C1q drives microglia-dependent synaptic loss and cognitive impairments in a mouse model of lipopolysaccharide-induced neuroinflammation. Neuropharmacology, 2023. 237: p. 109646.
- West, E.E. and C. Kemper, Complosome - the intracellular complement system. Nat Rev Nephrol, 2023. 19(7): p. 426-439.
- Matzinger, P. , Tolerance, Danger, and the Extended Family. Annual Review of Immunology, 1994. 12(Volume 12, 1994): p. 991-1045.
- Medzhitov, R. and C. Janeway, Jr., Innate immune recognition: mechanisms and pathways. Immunol Rev, 2000. 173: p. 89-97.
- Kerr, J.F., A. H. Wyllie, and A.R. Currie, Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer, 1972. 26(4): p. 239-57.
- Willingham, S.B. , et al., The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proceedings of the National Academy of Sciences, 2012. 109(17): p. 6662-6667.
- Griffin, F.M., Jr. Bianco, and S.C. Silverstein, Characterization of the macrophage receptro for complement and demonstration of its functional independence from the receptor for the Fc portion of immunoglobulin G. J Exp Med, 1975. 141(6): p. 1269-77.
- Botelho, R.J. , et al., Localized Biphasic Changes in Phosphatidylinositol-4,5-Bisphosphate at Sites of Phagocytosis. Journal of Cell Biology, 2000. 151(7): p. 1353-1368.
- Goodridge, H.S. , et al., Activation of the innate immune receptor Dectin-1 upon formation of a ‘phagocytic synapse’. Nature, 2011. 472(7344): p. 471-5.
- Davis, S.J. and P.A. Van Der Merwe, The kinetic-segregation model: TCR triggering and beyond. Nature immunology, 2006. 7(8): p. 803-809.
- Halstead, S.B. , Antibodies determine virulence in dengue. Ann N Y Acad Sci, 2009. 1171 Suppl 1: p. E48-56.
- Peiris, J.S. , et al., Monoclonal anti-Fc receptor IgG blocks antibody enhancement of viral replication in macrophages. Nature, 1981. 289(5794): p. 189-91.
- Cardosa, B.M.J., J. S. Porterfield, and S. Gordon, Complement receptor mediates enhanced flavivirus replication in macrophages. The Journal of Experimental Medicine, 1983. 158: p. 258 - 263.
- He, Y. , et al., In vitro enhancement of Zika virus infection by preexisting West Nile virus antibodies in human plasma-derived immunoglobulins revealed after P2 binding site-specific enrichment. Microbiology Spectrum, 2024. 12(6): p. e00758-24.
- Schafer, D.P. , et al., Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron, 2012. 74(4): p. 691-705.
- Menon, V. and S. Ghaffari, Erythroid enucleation: a gateway into a “bloody” world. Exp Hematol, 2021. 95: p. 13-22.
- Nicolás-Ávila, J.A. , et al., A Network of Macrophages Supports Mitochondrial Homeostasis in the Heart. Cell, 2020. 183(1): p. 94-109.e23.
- Brestoff, J.R. , et al., Intercellular Mitochondria Transfer to Macrophages Regulates White Adipose Tissue Homeostasis and Is Impaired in Obesity. Cell Metab, 2021. 33(2): p. 270-282.e8.
- Steinman, R.M. , et al., Endocytosis and the recycling of plasma membrane. J Cell Biol, 1983. 96(1): p. 1-27.
- Hilligan, K.L. and F. Ronchese, Antigen presentation by dendritic cells and their instruction of CD4+ T helper cell responses. Cellular & Molecular Immunology, 2020. 17(6): p. 587-599.
- Randolph, G.J., C. Jakubzick, and C. Qu, Antigen presentation by monocytes and monocyte-derived cells. Curr Opin Immunol, 2008. 20(1): p. 52-60.
- Puleston, D.J. and A.K. Simon, Autophagy in the immune system. Immunology, 2014. 141(1): p. 1-8.
- Russell, D.G. , Mycobacterium tuberculosis and the intimate discourse of a chronic infection. Immunological reviews, 2011. 240(1): p. 252-268.
- Gordon, S. , Phagocytosis: An Immunobiologic Process. Immunity, 2016. 44(3): p. 463-75.
- NobelPrize.org. The Nobel Prize in Chemistry 2004. [cited 2024 27.06.]; Available from: https://www.nobelprize.org/prizes/chemistry/2004/summary/.
- Hernández-Morán, B.A. , et al., Degron tagging for rapid protein degradation in mice. Disease Models & Mechanisms, 2024. 17(4).
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1985. [cited 2024 27.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1985/summary/.
- Nairz, M. , et al., “Pumping iron”-how macrophages handle iron at the systemic, microenvironmental, and cellular levels. Pflugers Arch, 2017. 469(3-4): p. 397-418.
- Nathan, C. , Secretory products of macrophages: twenty-five years on. J Clin Invest, 2012. 122(4): p. 1189-90.
- Liebold, I. , et al., Apoptotic cell identity induces distinct functional responses to IL-4 in efferocytic macrophages. Science, 2024. 384(6691): p. eabo7027.
- Martinez, F.O. and S. Gordon, The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep, 2014. 6: p. 13.
- Gordon, S. and F.O. Martinez, Alternative activation of macrophages: mechanism and functions. Immunity, 2010. 32(5): p. 593-604.
- Gordon, S., J. Todd, and Z.A. Cohn In vitro synthesis and secretion of lysozyme by mononuclear phagocytes Journal of Experimental Medicine, 1974. 139(5): p. 1228-1248.
- Gordon, S. and Z. Werb, Secretion of macrophage neutral proteinase is enhanced by colchicine. Proceedings of the National Academy of Sciences of the United States of America, 1976. 73(3): p. 872-876.
- Amersfoort, J., G. Eelen, and P. Carmeliet, Immunomodulation by endothelial cells — partnering up with the immune system? Nature Reviews Immunology, 2022. 22(9): p. 576-588.
- Winkler, J. , et al., Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nature Communications, 2020. 11(1): p. 5120.
- NobelPrize.org. The Nobel Prize in Physiology or Medicine 1931. [cited 2024 27.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1931/summary/.
- NobelPrize.org. [cited 2024 27.06.]; Available from: https://www.nobelprize.org/prizes/medicine/1953/summary/.
- Murphy, M.P. and L.A.J. O’Neill, A break in mitochondrial endosymbiosis as a basis for inflammatory diseases. Nature, 2024. 626(7998): p. 271-279.
- Mantovani, et al., Macrophage Polarization Comes of Age. Immunity, 2005. 23(4): p. 344-346.
- Gordon, S. , et al., Macrophage Class A Scavenger Receptors – A Functional Perspective. 2022.
- Pick, E. , In Memoriam: Filippo Rossi (1926–2022). Journal of Leukocyte Biology, 2023. 113.
- Nathan, C. , Role of iNOS in human host defense. Science, 2006. 312(5782): p. 1874-5; author reply 1874-5.
- Stein, M. , et al., Interleukin 4 potently enhances murine macrophage mannose receptor activity: a marker of alternative immunologic macrophage activation. J Exp Med, 1992. 176(1): p. 287-92.
- Mantovani, A. , et al., The chemokine system in diverse forms of macrophage activation and polarization. Trends in Immunology, 2004. 25(12): p. 677-686.
- Park, M.D. , et al., On the Biology and Therapeutic Modulation of Macrophages and Dendritic Cells in Cancer. Annual Review of Cancer Biology, 2023. 7(Volume 7, 2023): p. 291-311.
- Cassetta, L. and J.W. Pollard, Tumor-associated macrophages. Curr Biol, 2020. 30(6): p. R246-r248.
- Balkwill, F.R. and A. Mantovani, Cancer-related inflammation: common themes and therapeutic opportunities. Semin Cancer Biol, 2012. 22(1): p. 33-40.
- Blériot, C. , et al., A temporal perspective for tumor-associated macrophage identities and functions. Cancer Cell, 2024. 42(5): p. 747-758.
- Mantovani, A. , et al., Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity, 2019. 50(4): p. 778-795.
- Martinon, F., K. Burns, and J. Tschopp, The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell, 2002. 10(2): p. 417-26.
- Barnett, K.C. , et al., A 360° view of the inflammasome: Mechanisms of activation, cell death, and diseases. Cell, 2023. 186(11): p. 2288-2312.
- Hertzog, J. and J. Rehwinkel, Regulation and inhibition of the DNA sensor cGAS. EMBO Rep, 2020. 21(12): p. e51345.
- Lawn, S.D. and G. Meintjes, Pathogenesis and prevention of immune reconstitution disease during antiretroviral therapy. Expert Rev Anti Infect Ther, 2011. 9(4): p. 415-30.
- Webb, B.J. , Defining COVID-19-associated hyperinflammatory syndrome in specific populations. The Lancet Rheumatology, 2021. 3(9): p. e609-e611.
- Dexamethasone in Hospitalized Patients with Covid-19. New England Journal of Medicine, 2021. 384(8): p. 693-704.
- Roberti, A., L. E. Chaffey, and D.R. Greaves, NF-κB Signaling and Inflammation-Drug Repurposing to Treat Inflammatory Disorders? Biology (Basel), 2022. 11(3).
- Altmann, D.M. , et al., The immunology of long COVID. Nature Reviews Immunology, 2023.
- Mariano, M. and W.G. Spector, The formation and properties of macrophage polykaryons (inflammatory giant cells). J Pathol, 1974. 113(1): p. 1-19.
- Hughes, E.J. and D.M. Tobin, Decoding the tuberculous granuloma. Immunity, 2022. 55(5): p. 819-821.
- Henderson, S.R. , et al., Proteinase 3 promotes formation of multinucleated giant cells and granuloma-like structures in patients with granulomatosis with polyangiitis. Annals of the Rheumatic Diseases, 2023. 82(6): p. 848-856.
- Spector, W.G. , Immunologic components of granuloma formation. Epithelioid cells, giant cells, and sarcoidosis. Ann N Y Acad Sci, 1976. 278: p. 3-6.
- Schwartz, C. and P.G. Fallon, Schistosoma “Eggs-Iting” the Host: Granuloma Formation and Egg Excretion. Frontiers in Immunology, 2018. 9.
- Ahmadzadeh, K. , et al., Multinucleation resets human macrophages for specialized functions at the expense of their identity. EMBO Rep, 2023: p. e56310.
- Brooks, P.J., M. Glogauer, and C.A. McCulloch, An Overview of the Derivation and Function of Multinucleated Giant Cells and Their Role in Pathologic Processes. The American Journal of Pathology, 2019. 189(6): p. 1145-1158.
- Pereira, M. , et al., Common signalling pathways in macrophage and osteoclast multinucleation. J Cell Sci, 2018. 131(11).
- Chung, K.F., D. Togbe, and B. Ryffel, Editorial: Ozone as a Driver of Lung Inflammation and Innate Immunity and as a Model for Lung Disease. Frontiers in Immunology, 2021. 12.
- Dobzhansky, T. , Nothing in Biology Makes Sense except in the Light of Evolution. The American Biology Teacher, 1973. 35(3): p. 125-129.
- Cooper, S.J. , From Claude Bernard to Walter Cannon. Emergence of the concept of homeostasis. Appetite, 2008. 51(3): p. 419-427.
- Pirzgalska, R.M. , et al., Sympathetic neuron-associated macrophages contribute to obesity by importing and metabolizing norepinephrine. Nat Med, 2017. 23(11): p. 1309-1318.
- Hulsmans, M. , et al., Macrophages Facilitate Electrical Conduction in the Heart. Cell, 2017. 169(3): p. 510-522 e20.
- Schulthess, J. , et al., The Short Chain Fatty Acid Butyrate Imprints an Antimicrobial Program in Macrophages. Immunity, 2019. 50(2): p. 432-445.e7.
- Tabas, I. , 2016 Russell Ross Memorial Lecture in Vascular Biology. Arteriosclerosis, Thrombosis, and Vascular Biology, 2017. 37(2): p. 183-189.
- Huby, T. and E.L. Gautier, Immune cell-mediated features of non-alcoholic steatohepatitis. Nature Reviews Immunology, 2022. 22(7): p. 429-443.
- Mildner, A. , et al., Distinct and non-redundant roles of microglia and myeloid subsets in mouse models of Alzheimer’s disease. J Neurosci, 2011. 31(31): p. 11159-71.
- Cooper, M. , Which came first, T cells or B cells? Nature Reviews Immunology, 2021. 21(10): p. 616-617.
- NobelPrize.org. Svante Pääbo - Nobel Lecture. [cited 2024 01.07.]; Available from: https://www.nobelprize.org/prizes/medicine/2022/paabo/lecture/.
- Quintana-Murci, L. and A.G. Clark, Population genetic tools for dissecting innate immunity in humans. Nature Reviews Immunology, 2013. 13(4): p. 280-293.
- Barreiro, L.B. and L. Quintana-Murci, From evolutionary genetics to human immunology: how selection shapes host defence genes. Nature Reviews Genetics, 2010. 11(1): p. 17-30.
- Margulis, L. and U.M.A.M.L. Margulis, Symbiosis in Cell Evolution: Life and Its Environment on the Early Earth. 1981: W. H. Freeman.
- Margulis, L. and D. Sagan, Lynn Margulis: The Life and Legacy of a Scientific Rebel. 2012: Chelsea Green Publishing.
- Dopkins, N. and D.F. Nixon, Activation of human endogenous retroviruses and its physiological consequences. Nat Rev Mol Cell Biol, 2024. 25(3): p. 212-222.
- Xu, C. , et al., Living material assembly of bacteriogenic protocells. Nature, 2022. 609(7929): p. 1029-1037.
Figure 1.
Selected historical figures who contributed to understanding the role of Mononuclear Phagocytes in Cellular Immunity. (A) 1870-1920, (B) and (C) 1920-1970 (D) 1970-2020. See text and attachments for details. In this manuscript, we use the French spelling Elie Metchnikoff instead of the Russian version Ilya Metchnikov.
Figure 1.
Selected historical figures who contributed to understanding the role of Mononuclear Phagocytes in Cellular Immunity. (A) 1870-1920, (B) and (C) 1920-1970 (D) 1970-2020. See text and attachments for details. In this manuscript, we use the French spelling Elie Metchnikoff instead of the Russian version Ilya Metchnikov.


Figure 2.
Selected Milestones of Mononuclear Phagocyte Research and Cellular Immunity. Note indirect contributions of numerous Nobel awards. Asterisks indicate Nobel Prize laureates. Abbreviations: RES: Reticulo-endothelial system, MPS: Mononuclear Phagocyte system, AdGPCRs: Adhesion G-protein coupled receptors [32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52].
Figure 2.
Selected Milestones of Mononuclear Phagocyte Research and Cellular Immunity. Note indirect contributions of numerous Nobel awards. Asterisks indicate Nobel Prize laureates. Abbreviations: RES: Reticulo-endothelial system, MPS: Mononuclear Phagocyte system, AdGPCRs: Adhesion G-protein coupled receptors [32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52].
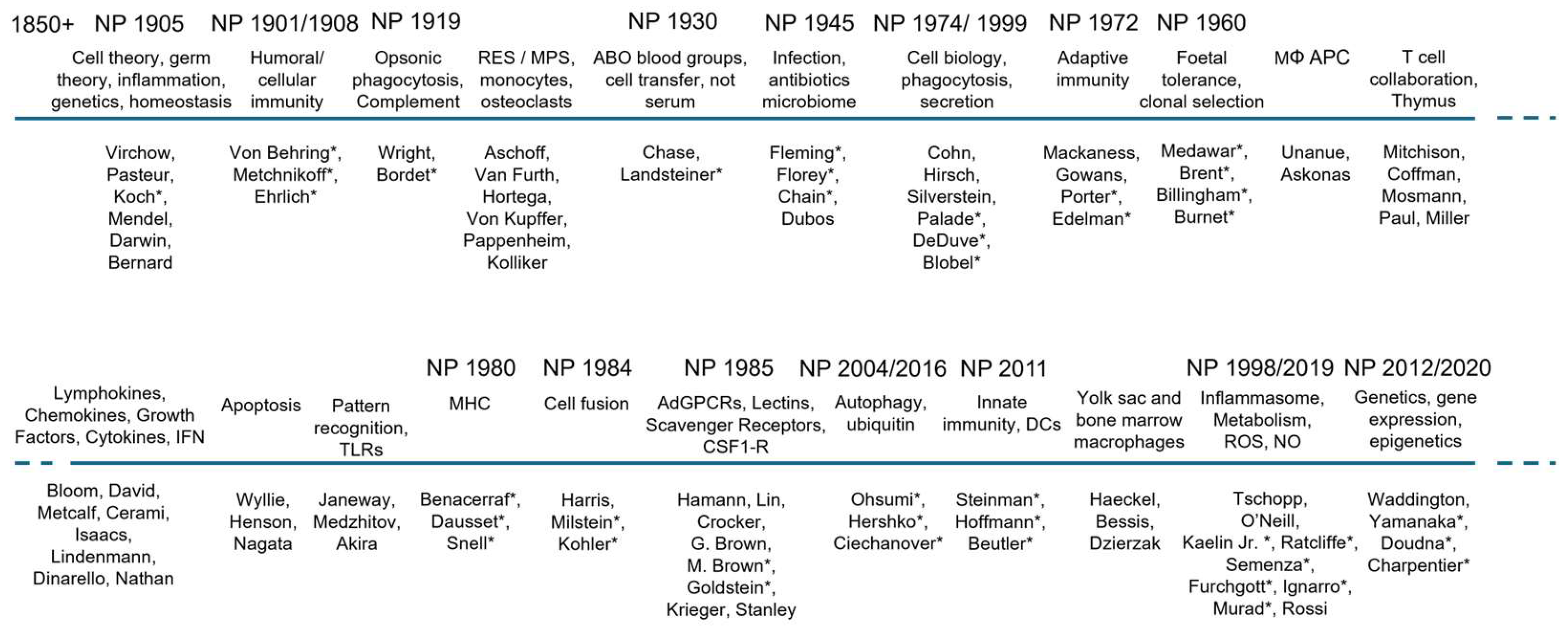
Figure 3.
The Mononuclear Phagocyte System. The differentiation and distribution of monocytes, macrophages, DCs and multinucleated giant cells (MNGCs) in vivo and in vitro, based mainly on F4/80 antigen expression in the mouse [156], on macrophage phenotypes in human placenta [162] and induced pluripotent stem cells (iPSC) [163]. Fejer, Galanos, Freudenberg et al. have developed primary macrophage cell lines from mouse foetal liver which are superior to tumour-derived cell lines (MPI cells) in studying macrophage differentiation in vitro [164]. Abbreviations: AGM= Aorta-gonad-mesonephros, HSC= Haematopoietic stem cell, MDSC= Myeloid-derived suppressor cell, GM-CSF= Granulocyte-macrophage colony stimulating factor, MPI= Max Planck Institute.
Figure 3.
The Mononuclear Phagocyte System. The differentiation and distribution of monocytes, macrophages, DCs and multinucleated giant cells (MNGCs) in vivo and in vitro, based mainly on F4/80 antigen expression in the mouse [156], on macrophage phenotypes in human placenta [162] and induced pluripotent stem cells (iPSC) [163]. Fejer, Galanos, Freudenberg et al. have developed primary macrophage cell lines from mouse foetal liver which are superior to tumour-derived cell lines (MPI cells) in studying macrophage differentiation in vitro [164]. Abbreviations: AGM= Aorta-gonad-mesonephros, HSC= Haematopoietic stem cell, MDSC= Myeloid-derived suppressor cell, GM-CSF= Granulocyte-macrophage colony stimulating factor, MPI= Max Planck Institute.
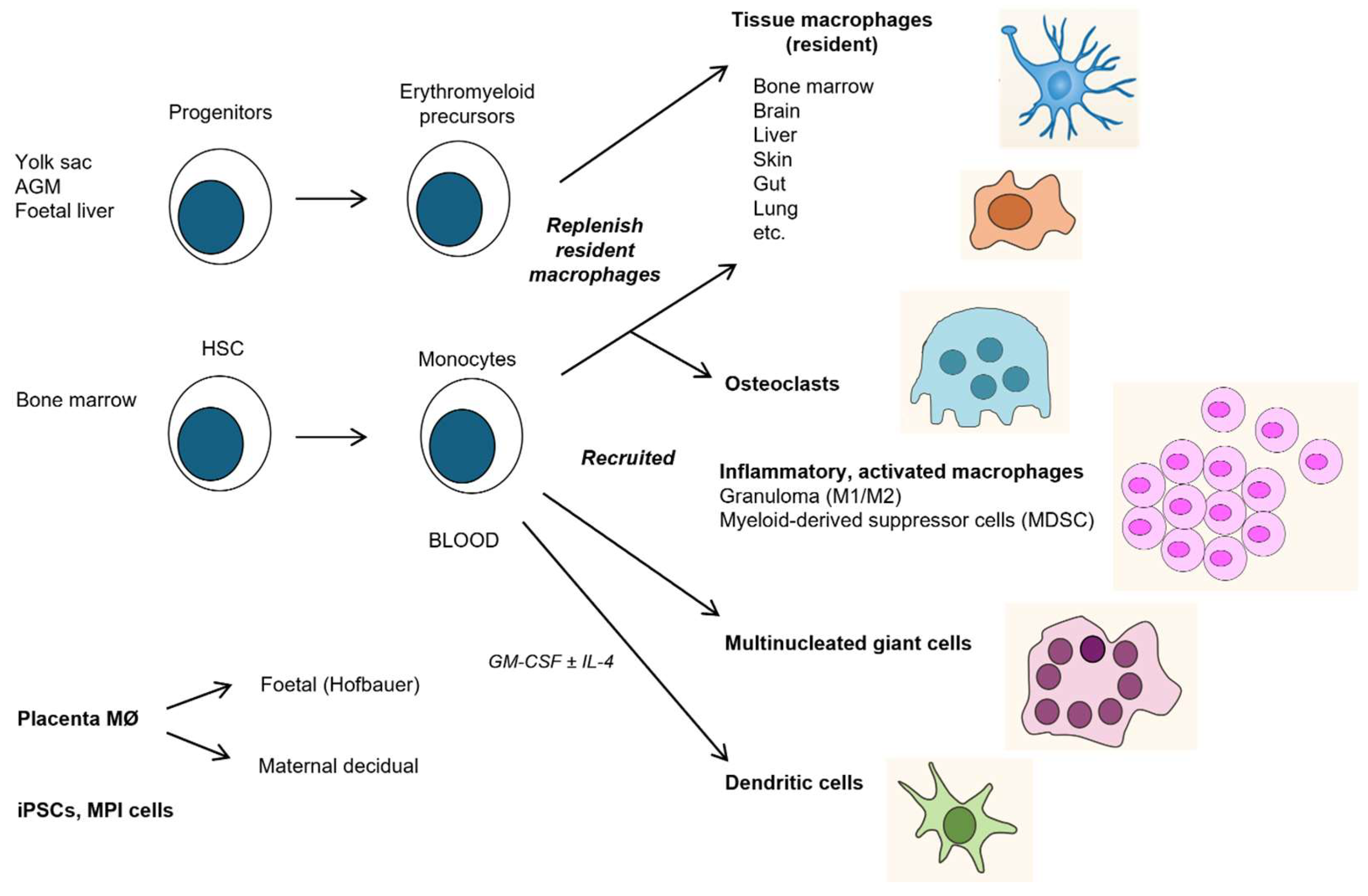
Figure 4.
Morphology of Mononuclear Phagocytes. (A) Dendritic cell isolated from mouse spleen. Phase contrast microscopy reveals characteristic nuclear structure, cytoplasmic organelles and dendritic processes [138]. (B) Mouse peritoneal macrophage engulfing antibody coated sheep erythrocytes, note plasma membrane ruffling revealed by scanning electron microscopy. (C) F4/80+ Langerhans cells in the mouse epidermis [169]. (D, E) Mouse F4/80+ microglia in situ and montage. Note ramified processes of individual cells. (E) Striking regional morphologic heterogeneity in grey and white matter [170,171].
Figure 4.
Morphology of Mononuclear Phagocytes. (A) Dendritic cell isolated from mouse spleen. Phase contrast microscopy reveals characteristic nuclear structure, cytoplasmic organelles and dendritic processes [138]. (B) Mouse peritoneal macrophage engulfing antibody coated sheep erythrocytes, note plasma membrane ruffling revealed by scanning electron microscopy. (C) F4/80+ Langerhans cells in the mouse epidermis [169]. (D, E) Mouse F4/80+ microglia in situ and montage. Note ramified processes of individual cells. (E) Striking regional morphologic heterogeneity in grey and white matter [170,171].
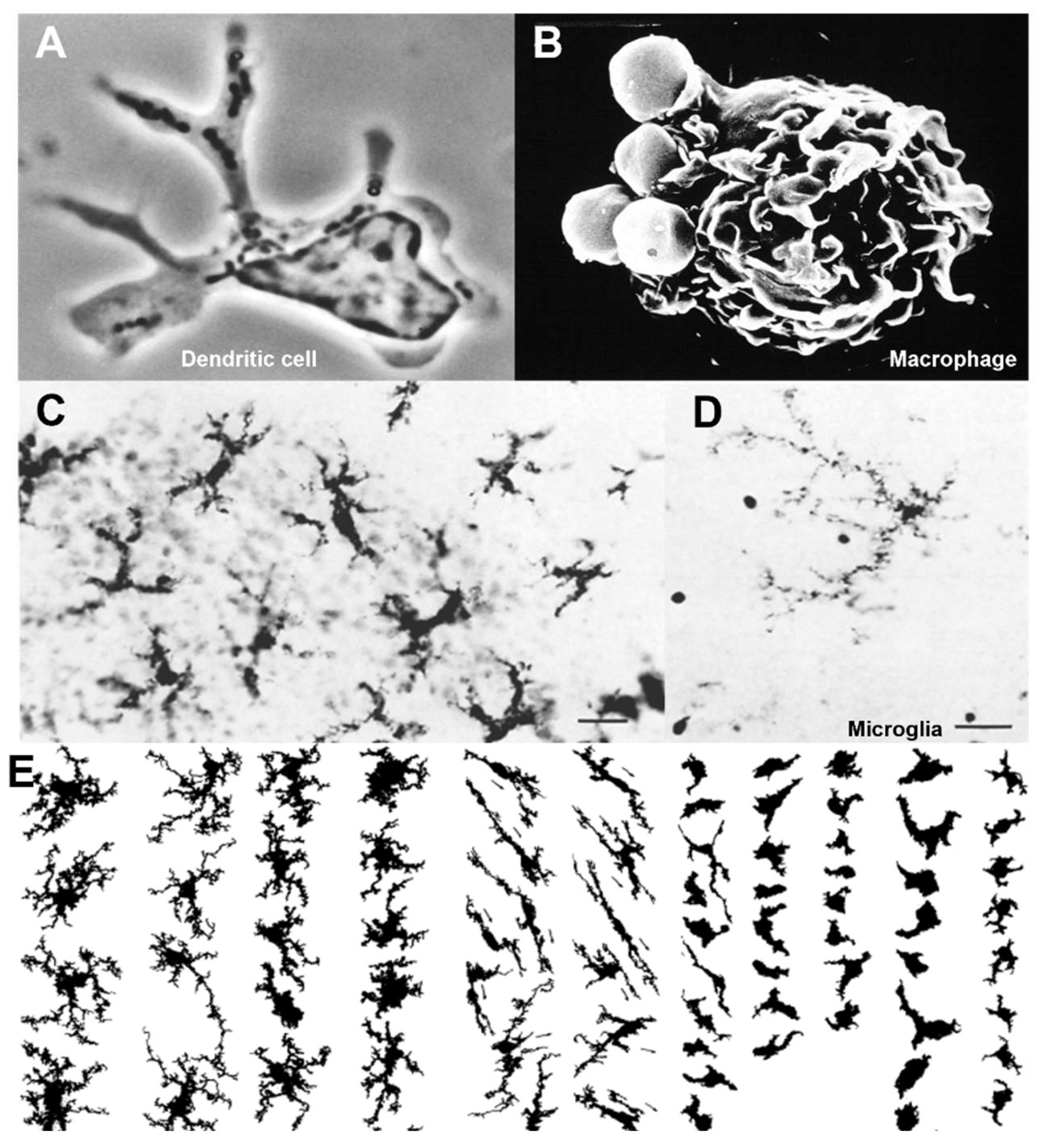
Figure 5.
Sensing the environment and Mononuclear Phagocyte Responses. Schematic representation of plasma membrane opsonic Fc and Complement receptors, Toll-like receptors (TLRs) and non-opsonic Lectin-like and Scavenger receptors. Signalling pathways of opsonic receptors depend on ITAM and ITIM cytoplasmic domains for activatory and inhibitory responses. Cytoplasmic sensors include mitochondrial antiviral signaling (MAVS) proteins, RIG-like helicases, NOD-like receptors, and components of the DNA sensing CGAS pathway. Inflammasome activation depends on ASC and Caspase-1. Abbreviations: ITAM= immunoreceptor tyrosine-based activation, ITIM= immunoreceptor tyrosine-based inhibitory motif, NF-κB= Nuclear factor kappa-light-chain-enhancer of activated B cells, IRF= interferon regulatory factors, RIG= Retinoic acid-inducible gene/protein; NOD= nucleotide binding and oligomerisation domain; cGAS= cyclic GMP-AMP synthase. Adapted from A Pluddemann.
Figure 5.
Sensing the environment and Mononuclear Phagocyte Responses. Schematic representation of plasma membrane opsonic Fc and Complement receptors, Toll-like receptors (TLRs) and non-opsonic Lectin-like and Scavenger receptors. Signalling pathways of opsonic receptors depend on ITAM and ITIM cytoplasmic domains for activatory and inhibitory responses. Cytoplasmic sensors include mitochondrial antiviral signaling (MAVS) proteins, RIG-like helicases, NOD-like receptors, and components of the DNA sensing CGAS pathway. Inflammasome activation depends on ASC and Caspase-1. Abbreviations: ITAM= immunoreceptor tyrosine-based activation, ITIM= immunoreceptor tyrosine-based inhibitory motif, NF-κB= Nuclear factor kappa-light-chain-enhancer of activated B cells, IRF= interferon regulatory factors, RIG= Retinoic acid-inducible gene/protein; NOD= nucleotide binding and oligomerisation domain; cGAS= cyclic GMP-AMP synthase. Adapted from A Pluddemann.
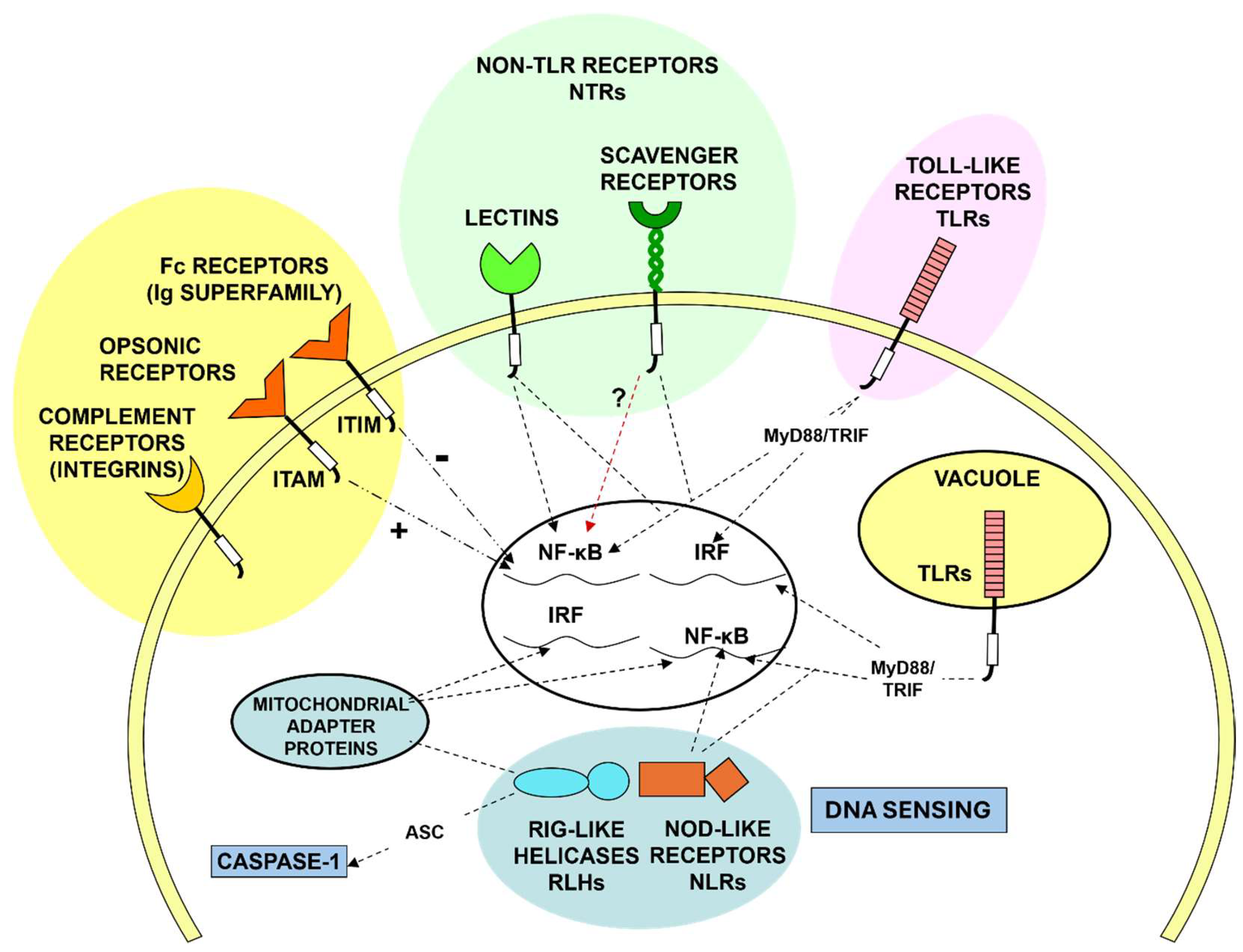
Figure 6.
Heterophagy and Autophagy: (A) Heterophagy is a defining property of Mononuclear Phagocytes. The schema shows key stages in the process of recognition, membrane and vesicular fusion and fission, and their recycling during cargo internalization and product secretion. Two important outcomes of phagocytosis are acidification/ digestion and the generation of oxygen and nitrogen radicals in host defense, critical determinants of intracellular infection. (B) The autophagy pathway employs analogous mechanisms to recognize, isolate and degrade damaged intracellular constituents. The genetic and molecular aspects of autophagy have attracted considerable attention recently [212]. Adapted from [212,213,214]. Abbreviations: Atg= autophagy-related gene, LC3= Microtubule-associated proteins 1A/1B-light chain 3.
Figure 6.
Heterophagy and Autophagy: (A) Heterophagy is a defining property of Mononuclear Phagocytes. The schema shows key stages in the process of recognition, membrane and vesicular fusion and fission, and their recycling during cargo internalization and product secretion. Two important outcomes of phagocytosis are acidification/ digestion and the generation of oxygen and nitrogen radicals in host defense, critical determinants of intracellular infection. (B) The autophagy pathway employs analogous mechanisms to recognize, isolate and degrade damaged intracellular constituents. The genetic and molecular aspects of autophagy have attracted considerable attention recently [212]. Adapted from [212,213,214]. Abbreviations: Atg= autophagy-related gene, LC3= Microtubule-associated proteins 1A/1B-light chain 3.
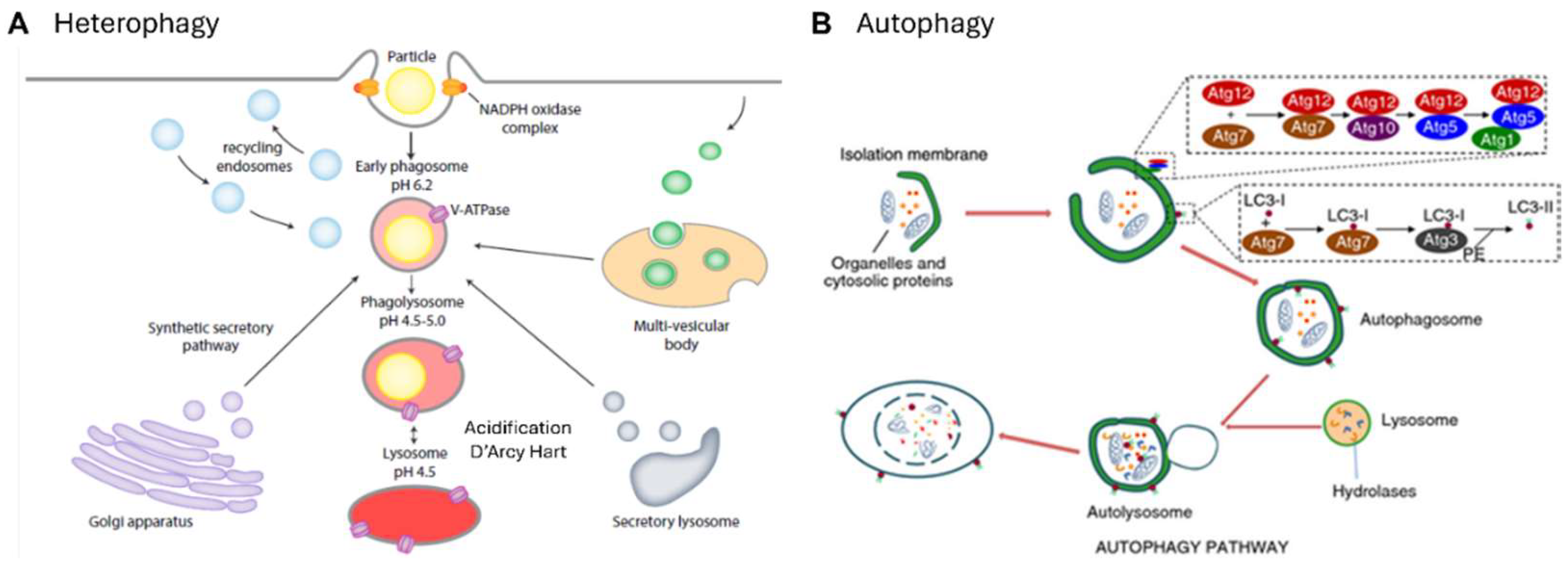
Figure 7.
Paradigm of Macrophage Polarisation. Macrophage gene expression is regulated differentially after priming by the Th1/2 cytokines, Interferon gamma versus IL4/13, followed by a local innate phagocytic stimulus. The spectrum of activation extends from a signature cluster of genes characteristic of Classical or Alternative activation, to deactivation by IL-10, tgf beta, glucocorticoid steroids [176], or by the uptake of apoptotic cells [46,220]. Polarisation of macrophages mirrors that of T lymphocytes [221]. Adapted from [222].
Figure 7.
Paradigm of Macrophage Polarisation. Macrophage gene expression is regulated differentially after priming by the Th1/2 cytokines, Interferon gamma versus IL4/13, followed by a local innate phagocytic stimulus. The spectrum of activation extends from a signature cluster of genes characteristic of Classical or Alternative activation, to deactivation by IL-10, tgf beta, glucocorticoid steroids [176], or by the uptake of apoptotic cells [46,220]. Polarisation of macrophages mirrors that of T lymphocytes [221]. Adapted from [222].
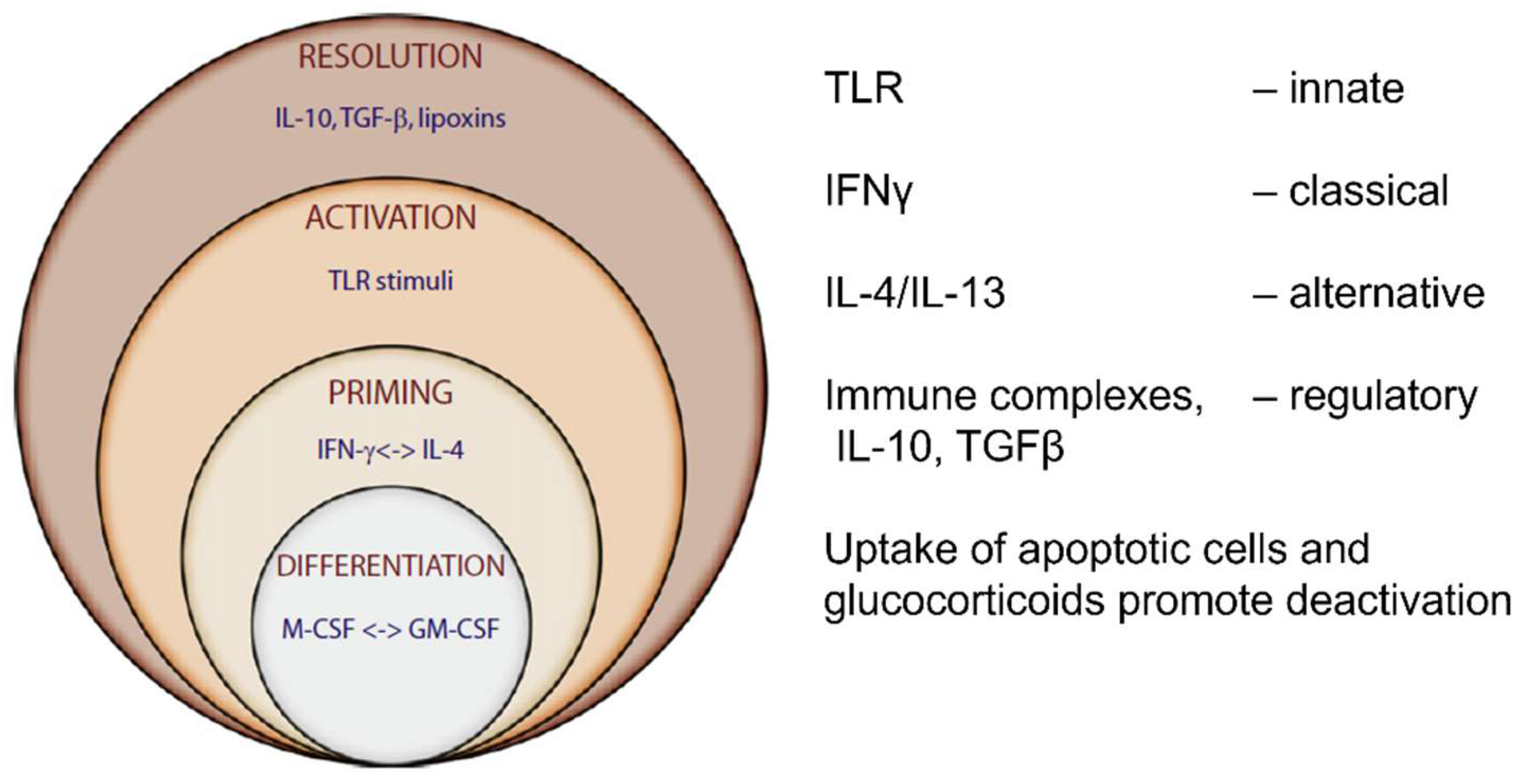
Figure 8.
Interactions of the MPS with other systems. We place the cells of the MPS at the centre of the homeostatic networks which interact reciprocally with all other cellular systems of the body. Illustrative examples of such systemic tissue interactions are given in the text.
Figure 8.
Interactions of the MPS with other systems. We place the cells of the MPS at the centre of the homeostatic networks which interact reciprocally with all other cellular systems of the body. Illustrative examples of such systemic tissue interactions are given in the text.
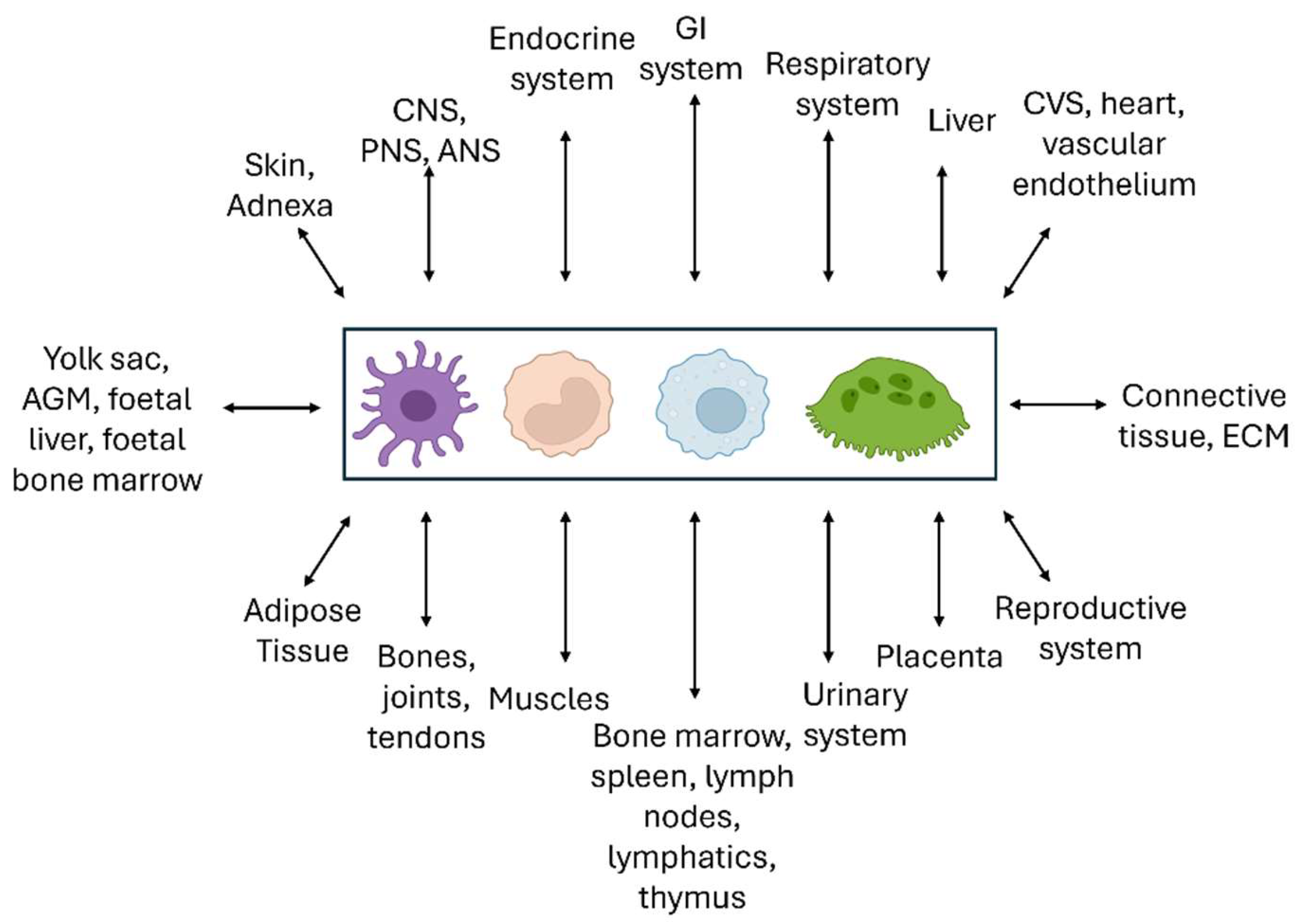

Table 1.
Selection of Nobel Awards in Physiology or Medicine, Chemistry and Physics which bear indirectly or directly on Mononuclear Phagocytes.
Table 1.
Selection of Nobel Awards in Physiology or Medicine, Chemistry and Physics which bear indirectly or directly on Mononuclear Phagocytes.
| Year | Names | Topic | Comment |
| 1901 | Von Behring | Diphtheria | Antibodies |
| 1905 | Koch | TB | Cell-mediated |
| 1908 | Metchnikoff, Ehrlich | Phagocytosis, Receptors | Cell-mediated and antibodies |
| 1912 | Carrel | Work on vascular suture and the transplantation of blood vessels and organs | Cell culture |
| 1913 | Richet | Anaphylaxis | Cell-mediated and antibodies |
| 1919 | Bordet | Complement | Humoral |
| 1930 | Landsteiner | Blood groups | Antibodies |
| 1931 | Warburg | Respiration | Metabolism |
| 1945 | Fleming, Chain, Florey | Penicillin | Antibiotics |
| 1950 | Kendall, Reichstein, Hench | Corticosteroids | Anti-inflammatory |
| 1951 | Theiler | Yellow fever vaccine | Adaptive |
| 1952 | Waksman | Streptomycin | Antibiotic TB |
| 1953 | Krebs, Lipmann | Citric acid cycle, coenzyme A | Metabolism |
| 1954 | Enders, Weller, Robbins | Polio virus | Isolation |
| 1960 | Burnet, Medawar | Self and non-self, tolerance | Cell-mediated, transplantation |
| 1966 | Rous, Huggins | Viral malignancy | Magnetic beads macrophage isolation |
| 1972 | Edelman, Porter | Antibodies | Structure |
| 1974 | Claude, de Duve, Palade | Cell structure | Cell biology, transmission EM |
| 1975 | Baltimore, Dulbecco, Temin | Viral replication | NF-κB pathway |
| 1976 | Blumberg, Gajdusek | Hepatitis, Prions | Infection |
| 1980 | Benacerraf, Dausset, Snell | Histocompatibility antigens | Genetics |
| 1982 | Bergstrom, Samuelsson, Vane | Prostaglandins | Regulatory |
| 1984 | Jerne, Kohler, Milstein | Monoclonal antibodies | Specific targeting |
| 1985 | Brown, Goldstein | Cholesterol | Scavenger receptor |
| 1987 | Tonegawa | Genetic control of antibodies | Lymphocytes |
| 1990 | Murray, Thomas | Transplantation | Cell-mediated |
| 1994 | Gilman, Rodbell | G-proteins | Signalling |
| 1996 | Doherty, Zinkernagel | MHC peptide recognition | Cell-mediated and lymphocytes |
| 1998 | Furchgott, Ignarro, Murad | NO | Effector metabolite |
| 1999 | Blobel | Protein signal peptide | Membrane transport |
| 2001 | Hartwell, Hunt, Nurse | Regulators of cell cycle | Cell cycle |
| 2002 | Brenner, Horvitz, Sulston | Genetics of organ development, programmed cell death | Evolution, C. elegans |
| 2005 | Marshall, Warren | Helicobacter pylori infection | Peptic ulcers |
| 2008 | Zur Hausen, Barre-Sinoussi, Montagnier | HPV and HIV | CD4 depletion, opportunistic infection |
| 2011 | Hoffmann, Beutler, Steinman | Insect, TNF, TLR, DCs | Innate immunity and antigen presentation |
| 2012 | Gurdon, Yamanaka | Reprogramming differentiation | iPSC |
| 2013 | Rothman, Schekman, Sudhof | Vesicular trafficking | Cell biology |
| 2016 | Ohsumi | Autophagy | Macrophages |
| 2018 | Allison, Honjo | Cancer immunotherapy | Checkpoint inhibitors |
| 2019 | Ratcliffe, Kaelin, Semenza | Oxygen sensing | Respiratory burst |
| 2020 | Houghton, Alter, Rice | Hepatitis C virus | Virus |
| 2021 | Patapoutian, Julius | Receptors for temperature and touch | Receptor biology |
| 2022 | Paabo | Genomes of extinct hominins and human evolution | Evolution and genetics |
| Chemistry and Physics | |||
| Year | Names | Topic | Comment |
| 1902 | Fischer | Sugar and purine synthesis | Biochemistry |
| 1948 | Tiselius | Electrophoresis | Method |
| Pauling | Protein bonds | Structural Biology/Chemistry | |
| 1958 | Sanger | Proteins | Sequence |
| 1962 | Perutz, Kendrew | Protein Structure | Analytic |
| 1972 | Anfinsen | Ribonuclease | Protein strucure |
| 1980 | Berg, Gilbert, Sanger | DNA | Sequencing |
| 1982 | Klug | Crystallography | Protein structure |
| 1993 | Mullis, Smith | PCR and mutagenesis | Nucleic acid |
| 2004 | Ciechanover, Hershko, Rose | Protein ubiquitination | Degradation |
| 2009 | Ramakrishnan, Steitz, Yonath | Ribosome | Structural biology |
| 2008 | Shimomura, Chalfie, Tsien | Green fluorescent protein | Detection, microscopy |
| 2012 | Lefkowitz, Kobilka | GPCRs | Signalling |
| 2018 | Arnold, Smith, Winter | Enzymes, phage display of peptides and antibodies | Directed evolution |
| 2020 | Charpentier, Doudna | Genome editing | CRISPR |
| 2022 | Bertozzi, Meldal, Sharpless | Click chemistry and biorthogonal chemistry | Protein and therapeutics |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



