
Submitted:
23 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
Shrimp is the most important aquaculture industry. Therefore, we determined the effect of nitrifying-probiotic bacteria on water quality, growth, survival, immune response and intestinal microbiota of Litopenaeus vannamei cultured without water exchange. In vitro, only B. licheniformis used TAN, nitrites and nitrates since nitrogen bubbles were produced. TAN decreased significantly in the treatments with bacilli and LAB, but no differences were observed in nitrites. Nitrates were significantly higher in the treatments with bacteria. The final weight and survival of shrimp in the bacterial treatments increased significantly and superoxide anion increased significantly only in LAB treatment. The activity of phenoloxidase decreased significantly in the treatment with LAB and with the mixture of bacilli and LAB. Shrimp treated with bacilli in the water showed lower species richness. The gut bacterial community after treatments was significantly different from that of the control. Linoleic acid metabolism was positively correlated with final weight and superoxide anion, whereas quorum sensing was correlated with survival. Thus, bacilli and LAB in the water of hyperintensive culture systems act as nitrifers, modulate the intestinal microbiota and immune response, and improve the growth and survival of shrimp. This is the first report on Pediococcus pentosaceus and Leuconostoc mesenteroides as nitrifying bacteria.
Keywords:
nitrifying bacteria
; Litopenaeus vannamei
; intestinal microbiota
; water quality
1. Introduction
From the economic and nutritional point of view, shrimp is the most important aquaculture market, generating 4 million tons annually [1]. Shrimp hyperintensive culture is a culture system carried out in open-air ponds or ponds covered with a plastic membrane ranging from 0.5 to 1 ha and is characterized by stocking densities exceeding 100 organisms m-3. Therefore, it is necessary to add oxygen to the water through aeration equipment, allowing to improve culture conditions and optimize feeding [2]. Due to the high stocking density, excess nitrogenous waste (ammonium and nitrites) is generated, which deteriorates the water quality and makes it toxic to shrimp. The main ammonium sources are shrimp excretions and sediment derived from mineralization of organic matter and molecular diffusion of reduced sediment [3, 4].
Nitrification is the general biochemical process of oxidation of ammonium (NH4+) to nitrite (NO2-) and, finally, to nitrate (NO3-) [5]. Bacterial nitrification is one of the commonly used methods for ammonium removal from aquaculture systems without water exchange [6]. The oxidation of ammonium to nitrite is generally carried out by bacteria that possess the ammonia-oxygenase enzyme such as Nitrosomonas, Nitrosococcus, Nitrosospira, Nitrosolobus, and Nitrosovibrio; whereas oxidation of nitrite to nitrate can be produced by the genera Nitrospina, Nitrococcus and Nitrobacter, using the nitrite-oxidase enzyme. However, these genera of anaerobic bacteria are more susceptible to changes in the environment and have a slower metabolism [7, 8]. Therefore, there are other bacterial groups used for bioremediation in aquaculture, which include species such as Bacillus licheniformis, Bacillus subtilis, Bacillus cereus, Pseudomonas, and Paracoccus [9,10,11]. These bacteria are also recognized as probiotics in aquaculture because they provide protection against bacterial and viral invasions in shrimp by stimulating the immune response (cellular and humoral reactions) [12, 13].
The invertebrate immune system is made up of cellular (hemocytes) and humoral effectors. Hemocytes, which are the first line of defense, participate in phagocytosis, capsule and nodule formation, cell adhesion, cytotoxicity, and coagulation [14]. Humoral effectors found in plasma or bound to hemocytes include antimicrobial peptides, lysozyme, lysosomal hydrolytic enzymes, lectins, the prophenoloxidase system, α2-macroglobulin, and transglutaminase [15,16,17,18].
Some bacterial species are culturable in the bacteriological media used in aquaculture; however, they could not offer an accurate picture as other species of bacteria could be present but hardly or even not culturable (Intriago et al. 2018). Therefore, metagenomics can be used to know the structure and function of the microorganisms of a specific sample [19]). The modulation of the intestinal microbiota in cultured shrimp has been carried out with the addition of probiotic bacteria [20]. The study of the microbial communities in the digestive tract of aquatic animals aims to highlight the benefit provided by the microbe-host relationship, which influences the health of the organism and its protection against pathogens [21].
Therefore, in this work, the effect of nitrifying-probiotic bacteria on water quality, growth, intestinal microbiota, immune response, and survival of Litopenaeus vannamei cultured with zero water exchange was determined.
2. Materials and Methods
2.1. Bacillus Licheniformis BCR 4-3 and Lactic Acid Bacteria (LAB) Culture
Bacillus licheniformis BCR 4-3, isolated and characterized by [22], was grown in trypticase soy broth (BD Bioxon®, Mexico) with 2.5% NaCl at 32 °C for 24 h, and centrifuged at 5000 g for 20 min. Lactic acid bacteria (LAB), such as Pediococcus pentosaceus, isolated and characterized by [23], and Leuconostoc mesenteroides, isolated and characterized by [22], were grown in Man Rogosa and Sharpe (MRS, Difco®, Mexico) broth with 2.5% NaCl at 32 °C for 48 h, and centrifuged at 5000 g for 20 min.
The bacterial pellets were resuspended in a 2.5% NaCl solution and brought to an absorbance (580 nm) of 1.0 in a Thermo Spectronic Genesys 2® spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA) before being inoculated into the shrimp culture water according to [22].
2.2. In Vitro Utilization of Ammonium, Nitrites and Nitrates by Bacilli and LAB
The medium used for the study of nitrification and denitrification consists of 20 g of peptone, 2 g of (NH4)2SO4, 1.5 g of KNO3, 1.5 g of MgSO4, 1.5 g of KH2PO4, and 20 g of NaCl per 1000 mL of distilled water. The medium was distributed, in triplicate, in test tubes with 15 mL each, to which a Durham hood (Smooth culture tube) was placed and inoculated with the isolated strains, leaving three tubes uninoculated as a control. They were incubated at 32 °C and positive or negative gas (nitrogen) formation was observed in the Durham hoods for one week.
2.3. Experimental Design
2.3.1. Origin and Acclimatization of Experimental Shrimp
Experimental shrimp (0.74 ± 0.07 g) were obtained from the hatchery systems of the Acuícola Cuate Machado farm (Guasave, Sinaloa, Mexico). Animals were transported to the Aquaculture Laboratory at CIIDIR-Sinaloa in 250-L plastic containers with 200 L of seawater from the culture tanks and constant aeration. Hypersaline water (90-100‰) previously treated with chlorine for one day was used. After chlorination (1.5%), water was aerated with diffuser stones to eliminate the chlorine by volatilization. The animals were acclimatized for 15 d at 30 PSU salinity in 1000-L plastic tank with 300 L of seawater, kept at room temperature and constant aeration [24] and fed with commercial feed (35% Purina®, Mexico) at 08:00, 13:00, and 17:00 h.
2.3.2. Bioassay: Effect of Bacilli and LAB on Water Quality, Growth, Survival, Immune System, and Intestinal Microbiota of Shrimp
The bioassay lasted 35 d and began with shrimp weighing 0.74 ± 0.07 g. Shrimp were cultured in plastic tanks (30 L) with 20 L of filtered seawater (20 µm), 30 PSU salinity, and constant aeration. Twelve animals were placed per tank and fed 3 times a day (08:00, 13:00, and 16:00 00 h) with commercial feed (35% Purina®, Mexico), adjusting the amount of feed according to the shrimp biomass (weight). The bioassay consisted of 4 treatments: I) control (without bacteria in water); II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Bacteria (3 × 106 CFU·L-1) were placed in the water every 7 d. There was no cleaning of the tanks or water exchange. The determination of physicochemical parameters (dissolved oxygen, temperature, pH, salinity) and survival were determined daily. Total ammonia nitrogen (TAN= NH3 and NH4+), nitrites, and nitrates were determined at the beginning and every 7 d.
The photoperiod was a 12:12 h light: dark cycle. Values of temperature (T), pH (HI 98127 pHep, Hanna Instruments, Woonsocket, RI, USA), salinity (S, Refractometer W/ATC 300011, Sper Scientific, Scottsdale, AZ, USA) and dissolved oxygen (YSI model 55 oximeter, Yellow Spring Instruments, Yellow Springs, OH, USA) were determined daily. During the bioassay, the physicochemical parameters behaved as shown in Table 1, indicating an optimal range [25].
On the 35th day, the shrimp were weighed, and five shrimp were taken from the three tanks of each treatment (2:2:1) to obtain intestine samples. The intestine of each shrimp was dissected, placed in a 1.5-mL Eppendorf tube with 1 mL of 96% (v/v) ethanol, and stored at -80 ºC. The samples (five per treatment) were sent to the Research Center for Food and Development (CIAD, Mazatlán, Sinaloa, Mexico).
Specific growth rate (SGR, %d-1) was calculated as follows [26]:
where W1 and W2 are the weights of the shrimp at times t1 and t2
2.3.3. Hemolymph Collection
Nine shrimp per treatment were obtained for the extraction of hemolymph (3: 3:3). Hemolymph was withdrawn from intermolt shrimp in the ventral sinus (second pair of pleopods) using a 1-mL tuberculin syringe with a 25-gauge needle. The syringe was previously loaded with precooled anticoagulant (450 mM NaCl, 10 mM KCl, 10 mM Hepes + 10 mM EDTA-Na2, pH 7.3) [27] in a ratio of 3:1 (3 volumes of anticoagulant for each volume of hemolymph).
2.3.4. Hemocyte Count
Fifty microliters of anticoagulant-hemolymph from the nine shrimp were diluted in 150 µL of formaldehyde (6%) and then 10 µL were placed in a hemocytometer (Neubauer chamber) to count hemocytes (THC) using a compound microscope.
2.3.5. Superoxide Anion
The free radical superoxide anion was quantified according to [28]. Samples (100 μL) of anticoagulant–hemolymph from three shrimp per tank (nine per treatment) were centrifuged at 800 g for 10 min at 4 °C and the plasma was discarded. Hemocyte pellet was washed three times with SIC-EDTA buffer and stained with 100 μL of nitro blue tetrazolium (NBT) solution (0.3%) for 30 min at 37°C. The superoxide anion reduces NBT to a formazan. The reaction was completed through the elimination of the NBT solution by centrifuging and adding 100 μL of absolute methanol to the cell pellet. After three washings with 70% methanol, hemocytes were air dried for 30 min, and 140 μL of DMSO and 120 μL KOH (2 M) were added to dissolve the cytoplasmic formazan. The optical density of the dissolved formazan was read at 630 nm in a Thermo Spectronic Genesys 2 Spectrophotometer 2.10 (Thermo Fisher Scientific).
2.3.6. Phenoloxidase Activity (PO) in Hemolymph
Hemolymph from three shrimp per tank (nine shrimp per treatment) was subjected to freeze thaw (-20°C). Finally, the samples were centrifuged at 14, 000 g for 10 min at 4°C to obtain the hemolymph supernatant. As the extraction buffer containing EDTA, which chelates calcium in plasma that is necessary for activating the prophenoloxidase activating enzyme, prophenoloxidase was activated with trypsin. Fifty microliters of sample were incubated with 50 μL of trypsin (0.1 mg mL-1) and incubated for 30 min at 37°C. Next, the sample was incubated with 50 μL of L-DOPA (3 mg mL-1 of distilled water) for 10 min at 37°C. Phenoloxidase activity was measured spectrophotometrically by recording the formation of dopachrome produced from L-DOPA at 492 nm in a microplate reader spectrophotometer from Awarness Technology (Palm City, FL, USA). Enzyme activity was expressed as absorbance at 492 nm.
2.4. Water Sampling, TAN, Nitrites, and Nitrates Determination
Samples of 1000 mL of culture water were taken from each tank and placed in plastic bottles with screw caps (1000 mL). The samples were shaken and the solids allowed to settle down for 3 min. Subsequently, samples were filtered by gravity with 696-grade fiberglass filters of 4.7 cm diameter (VWR International, Ltd., United Kingdom). The concentration of TAN, nitrites, and nitrates was determined following the Practical Manual of Seawater Analysis [29].
2.5. Metagenomic Analysis
2.5.1. Extraction of Bacterial DNA, Library Preparation, and Sequencing in Illumina MiniSeq
Microbial DNA was extracted from intestine samples using the cetyltrimethylammonium bromide (CTAB) method [30]. The variable region V3 of the bacterial 16S rRNA gene was amplified by PCR with the primers 338F (ACT CCT ACG GGAGGC AGC AG) and 533R (TTA CCG CGG CTG CTG GCAC) [31]. DNA amplification was carried out with the KAPA kit (2x KAPA HiFi HotStart ReadyMix) from Roche (Basel, Switzerland) in a 25-μL reaction volume. PCR was performed in a thermal cycler using the following program [32]: one cycle of 30 s at 95 °C; followed by 25 cycles, each one of 30 s at 95 °C, 55 °C for 30 s, 72 °C for 15 s, and a final extension at 72 °C for 7 min. AMPure XP magnetic beads were used to clean up amplicons from free primers and primer dimers. For sequencing, purified amplicons were associated with dual indices and Illumina sequencing adapters, using the Nextera XT index kit (Illumina, San Diego, CA, USA). Illumina MiniSeq platform was used under standard conditions (300 cycles, 2 × 150 pair-end) to perform sequencing. Before their quantification, the libraries were purified with AMPure XP magnetic beads. Raw reads from Illumina MiniSeq sequencing were deposited in the NCBI Sequence Read Archive (SRA) under accession number PRJNA1044443.
2.5.2. Gut Bacterial Taxonomy, Abundance, and Diversity Analysis
The raw sequences were cleaned with pair-end cleaner v. 1.0.3 and then analyzed with the web-based Shaman [33] and MicrobiomeAnalyst [34, 35], platforms for microbial taxonomy, abundance, and diversity. The analysis of read quality control, dereplication, removing singletons, removing chimera sequences, and grouping was carried out on the Shaman platform to construct operational taxonomic units (OTU). The OTUs shared by the three groups were determined using the Venn diagram analysis (http: //jvenn.toulouse.inra.fr/app/example.html) [36]. On the Shaman platform, the reads obtained from the V3 hypervariable region of the bacterial 16S rRNA gene were annotated against the SILVA (version 138.1, https://bioweb.pasteur.fr/data?search=silva) ribosomal RNA database with a confidence threshold of 0.8. The analyses of the alpha (Shannon, Simpson, Chao 1, ACE) and beta [Non-metric multi-dimensional scaling (NMDS)] indices were performed in the MicrobiomeAnalyst platform to explore the effects of bacilli and LAB in water on the bacterial community composition of cultured shrimp intestines.
Bacterial functional profile of the white shrimp intestine was performed on the multimodular web platform iVikodak. Using the Global Mapper module (independent contribution algorithm), functional profiles were inferred based on the KEGG database pathways [37].
2.6. Statistical Analysis
Most data are shown as mean ± SD. One-way analysis of variance (ANOVA) was applied to determine the differences in concentration of ammonium, nitrites, and nitrates, SGR, survival, total hemocytes, FO activity, and superoxide anion. Survival data in percentage were arcsine-transformed according to [38]. If significant differences were found in the ANOVA, a Tukey's HSD test was used to identify these differences at P < 0.05. For alpha diversity (Shannon, Simpson, Chao 1, ACE), the Kruskal-Wallis test was used (P < 0.05). For beta diversity analysis, the ANOSIM test (P < 0.05) was performed in the MicrobiomeAnalyst web-based platform. Spearman correlation analysis was performed to correlate some functional profiles with immune and productive variables (P < 0.05). Correlation plots were made online with SRplot (https://www.bioinformatics.com.cn/srplot).
3. Results
3.1. In Vitro Utilization of Ammonium, Nitrites and Nitrates by Bacilli and LAB
Results showed that in vitro test, bacilli oxidize TAN and nitrites (nitrification process) and reduce nitrates (denitrification process) since nitrogen bubbles were produced. Each LAB grew in the culture medium but did not produce nitrogen bubbles.
3.2. Effect of Bacilli and LAB on Culture Water Quality
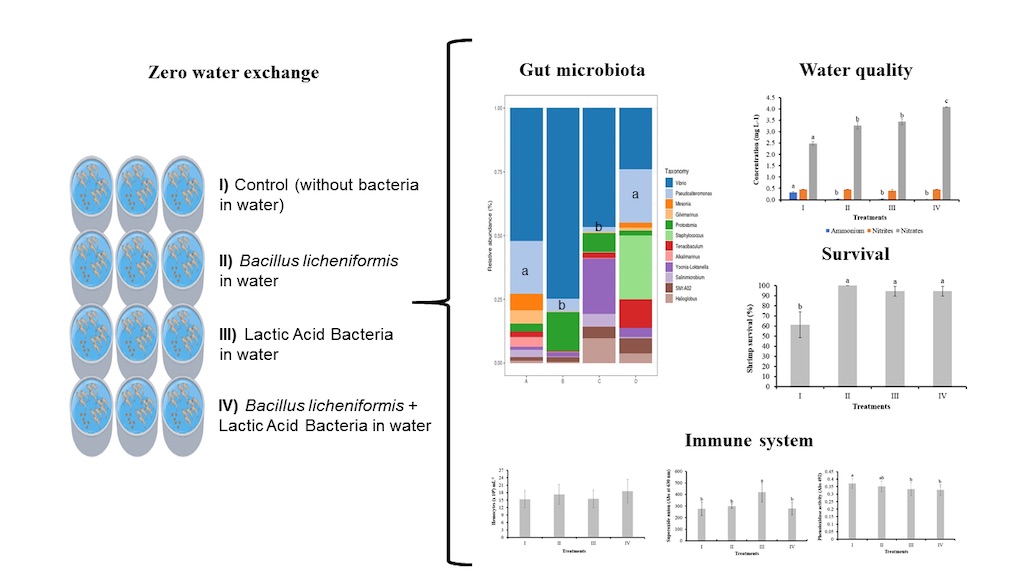
The TAN concentration of water in the control (I), bacilli in the water (II), LAB in the water (III), and bacilli and LAB in the water (IV) was 0.3 ± 0.029, 0.030 ± 0.001, 0.034 ± 0.004, and 0.015 ± 0.001 mg L-1, respectively. A significant decrease (P < 0.05) was observed in treatments II, III, and IV with respect to the control (I). No significant differences in the concentration of nitrites (0.40-0.46 mg L-1) were observed among treatments (P > 0.05). The nitrate concentration of water in the control (I, and treatment groups, bacilli (II), LAB (III), and bacilli and LAB (IV) was 2.47 ± 0.08, 3.27 ± 0.16, 3.44 ± 0.13, and 4.10 ± 0.02 mg L-1, respectively. An increase (P < 0.05) was observed in treatments II, III, and IV with respect to the control (I). Treatment IV presented a significant increase (P < 0.05) compared to treatments II and III (Figure 1).
3.2.1. Shrimp Growth
The initial average weight was between 0.70 g (control) and 0.73-0.79 g (treatments), no significant differences were observed between the treatments with bacteria in the water and the control (P > 0.05). The average final weight was between 3.09 g (control) and 3.87-4.18 g (treatments with bacteria in the water). Significant differences were observed between the control and bacilli in the water (II), and between the control and bacilli and LAB in the water (IV) (P < 0.05). The SGR was between 4.16 ± 0.25% d-1 and 4.78 ± 0.21% d-1. No significant differences were observed between the treatments and the control (P > 0.05) (Table 2).
3.2.2. Shrimp Survival
On the 34th day, dead shrimp were found in the tanks. The survival of shrimp in the control group (I), bacilli in the water (II), LAB (III), and bacilli and LAB in the water (IV) on the 35th day was 60 ± 12.72%, 100 ± 0.0%, 95 ± 4.81%, and 95 ± 4.81%, respectively. Significant differences were found between control and treatments (P < 0.05) (Figure 2).
3.3. Immune System
3.3.1. Total Hemocyte Count
Figure 3 shows the results of the number of shrimp hemocytes in the control and treatment groups with bacteria in water (II, III, IV). The number of hemocytes ranged between 15.38 (× 106) and 18.60 (× 106) per milliliter. No significant differences were observed (P > 0.05).
3.3.2. Superoxide Anion
Figure 4 shows the results of superoxide anion concentration (absorbance at 630 nm) in shrimp hemocytes. A significant increase (P < 0.05) was observed in treatment III (LAB, 418.44 ± 82.39) compared to I (control, 275.94 ± 57.65), II (bacilli in the water, 299.44 ± 19.25), and IV (bacilli and LAB in the water, 277.44 ± 54.01).
3.3.3. Phenoloxidase Activity in Shrimp Hemolymph
Phenoloxidase activity (absorbance at 492 nm) was detected in shrimp hemolymph (Figure 5). The phenoloxidase activities in the hemolymph of the control group (I), bacilli in the water (II), LAB in the water (III), and bacilli and LAB in the water (IV) were 0.37 ± 0.03, 0.35 ± 0.03, 0.33 ± 0.04, and 0.32 ± 0.03 mg L-1, respectively. No significant differences (P > 0.05) were observed between the control (I) and the treatment with bacilli in the water (II); however, a significant decrease (P < 0.05) of the enzyme activity was observed in treatments III and IV compared to I.
3.4. Gut Microbiota Analysis
In total, 428 OTUs were annotated by the Silva ribosomal RNA database. The control had the highest number (391) of OTUs and the treatment with bacilli in the water had the lowest (278). According to the Venn analysis, the four groups shared 189 OTUs. Regarding the unique OTUs, the group with the most OTUs was the control (26) and the one with the least was the mixture of bacilli and LAB in the water (6) (Figure 6).
3.4.1. Relative Bacterial Abundance in the Shrimp Gut
The relative abundance of the most relevant taxa at the phylum and genus levels in the intestines of shrimp treated with bacteria (bacilli and LAB) in the water and control was determined. The most abundant taxa (Figure 7) in control and treatments were Proteobacteria and Bacteroidota, and no significant differences were observed among the treatment groups with bacteri in the water (P > 0.05). Patescibacteria, the third phylum in relative abundance, was significantly enriched (P < 0.05) in treatment II (bacilli in the water) compared to I (control), III (LAB in the water), and IV (bacilli and LAB in the water).
Results show the most abundant genera composed of Vibrio, Pseudoalteromonas, Mesonia, Gilvimarinus, Staphylococcus, Tenacibaculum, Alkalimarinus, Yoonia-Loktanella, Salinimicrobium, and Halioglobus. Among these 10 genera, Vibrio was the most abundant, followed by Pseudoalteromonas. Only, Pseudoalteromonas decreased significantly (P < 0.05) in treatments II (bacilli in the water) and III (LAB in the water) as compared to control and treatment IV (bacilli and LAB in the water) (Figure 8).
3.4.2. Alpha Diversity Indices
Good's estimated sample coverage in the three groups was 99.93-99.99%, which means adequate data sampling and good quality sequences. Rarefaction analysis showed that samples reached the plateau, which means that the sequencing depth was sufficient to capture all the OTUs in the samples. No significant differences were found in Shannon and Simpson indices; however, they showed a tendency to be lower in the treatment groups with bacteria in the water. In the Chao index, the values were between 93.95 ± 25.25 and 149.74 ± 27.71; significant differences were observed between treatment II and treatment III, and between IV and the control (P < 0.05). In the ACE, the values were between 88.95 ± 28.97 and 146.34 ± 27.87, there were significant differences between the control and treatment II (P < 0.05) (Table 3).
3.4.3. Beta Diversity Index
Jaccard similarity matrices were visualized using non-metric multi-dimensional scaling (NMDS). At genus level, samples clustered according to treatment (NMDS stress = 0.13419). Control samples grouped separately from the experimental groups, II, III, IV. The ANOSIM analysis showed significant differences, although R values were low (R = 0.18067; P < 0.008) (Figure 9).
3.4.4. Bacterial Functional Profile Based on KEGG Pathway Analysis
In the shrimp gut samples, six functional categories were found, that is, metabolism (60.03-61.05%), genetic information processing (10.88-11.01%), human diseases (11.43-11.94%), environmental information processing (1.39-1.59%), cellular processes (6.90-738%), and organismal systems (8.10-8.27%). No significant differences were found in the treatments (bacteria in the water) compared to the control (P < 0.05) (Table 4).
3.5. Correlation between Functional Categories and Immune and Productive Variables
Only metabolism of linoleic acid and quorum sensing significantly correlate with productive variables in the KEGG pathway analysis at level 3. Figure 10 shows the positive correlation between the metabolism of linoleic acid and superoxide anion (P = 0.045), as well as the final weight (P < 0.05). Figure 11 shows the positive correlation between quorum sensing and shrimp survival (P < 0.05).
4. Discussion
The high animal densities that are managed in aquaculture systems deteriorate the water quality, which generates stress and susceptibility to diseases [39, 40]. Ammonia and nitrites at high concentrations are toxic to aquaculture animals and their toxicity depends on temperature, salinity, pH, or the developmental stage of the organism [41,42,43,44]. Furthermore, aquatic nitrogenous wastes, such as nitrites, can directly affect the nitrite content in shrimp intestine as this organ is exposed to the water environment [45]. Therefore, reducing nitrogenous waste in culture systems is very important for producing cultured animals and preventing deterioration of the surrounding environment [46].
The bacterial consortia, composed of several probiotic strains, usually have synergy leading to a bioremediation effect to improve water quality [47]. Some bacterial strains contain enzymes that oxidize ammonia and nitrites, contributing to the nitrification process of nitrogenous waste in culture systems [48]. In this work, only bacilli oxidize TAN and nitrites and reduce nitrates in vitro since nitrogen bubbles were produced, indicating a nitrification and denitrification process. The addition of bacilli and LAB to the water decreased TAN and nitrite concentrations, showing a nitrifying effect, so a denitrifying effect was not observed as occurred in vitro with bacilli. Also, it is important to mention that in the control there was also a nitrification process as TAN and nitrites were not high (0.3 and 0.4 mg L-1) and nitrates were 2.47 mg L-1, which could be caused by the natural bacteria community from the shrimp and initial filtered water. The nitrification process was reported in the culture of L. vannamei [49, 50], when Bacillus subtilis L10 and G1 were inoculated into the water. Denitrification process was observed in Bacillus sp. SC16 in an intensive fishery aquaculture pond [51]. Finally, there are no reports indicating Pediococcus and Leuconostoc as nitrifying bacteria.
In the aquatic environment, bacterial components in the sediment and water affect the bacterial communities in the shrimp intestine [52,53,54] mention that 35.32% of bacteria in the intestine of the white shrimp come from water and 54.58% from sediments. The intestinal microbiota of organisms affects their digestion, absorption, growth, and immune response [55]. Regarding the effect of bacteria and nitrogenous waste on growth, the best shrimp growth was observed in the treatments with bacilli or the mixture of bacilli and LAB in the water. It is known that bacterial species, such as Bacillus, inoculated into the water, improve the growth of L. vannamei [49, 56, 57] thanks to the increase in the production of enzymes that digest feed nutrients [58] and short-chain fatty acids [59]. On the other hand, elevated nitrites affected the growth of L. vannamei [60, 61] and reduced food consumption in Farfantepenaeus brasiliensis [62]. Similarly, [63] mention that ammonia and nitrites reduce the growth in weight and length of L. vannamei cultured in the laboratory.
The optimal concentration of nitrogen in water in the form of ammonium (NH4+), ammonia (NH3), nitrites (NO2-), and nitrates (NO3-) is 0.2-2.0, 0.09-0.11, < 0.23, and 0.2-10 mg·L-1, respectively [64,65,66] mention that crustaceans excrete ammonia and cannot convert it to the less toxic compounds, so the high concentration of these compounds is toxic to crustaceans through the gill absorption. In the present work, TAN’s concentration was below the toxic concentration. However, the stress caused by ammonia reduces the survival of L. vannamei grown in the laboratory [63, 65]. Nitrites are toxic to crustaceans because they convert hemocyanin into meta-hemocyanin, which is incapable of transporting oxygen [68]. In our work, nitrites were in the range of 0.40 and 0.46 mg·L-1, above the optimal concentration in all treatments. Therefore, it is possible that mortality was caused, in part, due to the stress induced by nitrites. The nitrification driven by bacteria (bacilli and LAB) could thus protect L. vannamei against the negative effect of nitrites. In the work of [60], a high mortality of L. vannamei was observed due to nitrite poisoning (5.0-40.0 mg L-1) and they mention that during molting there is a high consumption of oxygen so nitrites cause hypoxia or metabolic anoxia. In contrast, [61] did not observe mortalities in L. vannamei exposed to high concentrations of nitrites (2.0, 6.67, and 20 mg L-1).
Invertebrates possess a natural, non-specific immune response, which plays an important role in resistance to microbial diseases [68] as they do not produce antibodies like vertebrates [17]. In our work, no significant change was observed in the number of hemocytes in shrimp treated with bacteria in the water and the accumulation of nitrogenous waste. [69] found that L. vannamei treated with 400 and 600 mg·L-1 of Gracilaria tenuistipitata extract for 3 h and then exposed to 5 mg·L-1 of ammonia, increased hemocytes compared to the control group. Regarding the superoxide anion, it increased in the hemocytes of the organisms treated with LAB in the water compared to the control and the other treatments. In this treatment, nitrites were above the optimal range (0.09-0.11 mg L-1). [70] found that the concentration of nitrites in the hemolymph of L. vannamei was similar to that of the culture water and that this exposure increased the generation of ROS (measured indirectly by the high activity of catalase in the hepatopancreas). Similaly, [69] found that in the hemolymph of shrimp treated with G. tenuistipitata extract and then exposed to ammonia, the activity of the enzyme superoxide dismutase increased, which has the superoxide anion radical as a substrate. In contrast, [61] found no changes in SOD activity in the hepatopancreas of L. vannamei exposed to high nitrite concentrations, and [71] found similar results in L. vannamei pretreated with dietary inositol and exposed to ammonia during 24 h. Regarding phenoloxidase, its activity decreased in the treatments with LAB alone and the mixture of bacteria in the water. The decrease in phenoloxidase activity could indicate an optimal physiological state with less biotic stress. Converseley, data reported by [69] showed an increase in the activity of the enzyme in shrimp treated with G. tenuistipitata and ammonia.
In the shrimp intestine, the microbiota is complex and variable, mainly affected by diet, developmental phase, immune response, metabolism, and the environment surrounding the animals [72, 73]. At the phylum level, Proteobacteria and Bacteroidota predominated; however, no significant differences were observed in bacterial treatments. In the work of [61], at the concentration of 2 mg L-1 of nitrites, the highest relative abundance corresponded to Bacteroidota, followed by Proteobacteria and Actinobacteria. At the highest nitrite concentration (20 mg L-1), the highest abundance corresponded to Proteobacteria followed by Bacteroidota and Actinobacteria. All the above demonstrated that Proteobacteria and Bacteroidota are the most important components in shrimp intestine under this condition. Proteobacteria is a core member of shrimp gut microbiota [74]. According to [75] and [76], the abundance of this phylum indicates efficient colonization of the shrimp intestinal epithelium, and possible degradation of agar and cellulose and nitrogen fixation in the shrimp rectum [77]. Regarding the phylum Bacteroidota, it increases when the amount of dietary protein and fat increases [78, 79] and has a very important role due to its ability to utilize nitrogenous waste, biotransform steroids, and ferment carbohydrates [71, 72, 79, 80].
Within the native microbiota that predominates in marine species, the genus Vibrio constitutes the greatest abundance [81]. In our study, the highest relative abundance was presented by the Vibrio genus, with the highest abundance in the bacilli treatment followed by the treatment with LAB in the water. [82] and [83] reported the Vibrio genus as abundant in white shrimp intestines in treatments supplementing Bacillus species to the culture water, which coincides with the report of [84], where the mentioned genus predominated in most treatments supplemented with different Bacillus mixtures. The control of Vibrio in shrimp culture is very important as it could affect the shrimp health [85]. However, some Vibrio strains are beneficial to shrimp health such as and V. hepatarius and V. diabolicus that protect Penaeus vannamei larvae against V. parahaemolyticus [86] or V. parahaemolyticus, V. diazotrophicus, V. natriegens, and V. campbellii strains that utilize several organic carbon sources (unused organic matter) and can fix nitrogen [87, 88]. The second most abundant genera was Pseudoalteromonas, which is a probiotic bacteria that is antagonistic against V. parahaemolyticus, V. harveyi, and V. nigripulchritudo [89,90,91]. Bioencapsulated Pseualteromonas in Artemia sp. increases the immune response of shrimp and their resistance to infections caused by V. harveyi [92, 93].
Functional redundancy provided by high microbial diversity allows an ecosystem to be more stable and resistant to stress [94, 95]. The total bacterial species richness in a sample can be determined with the Chao1 and ACE alpha indices [96, 97]. On the other hand, Shannon and Simpson alpha indices consider the richness of the microbial community and the evenness (relative abundance of different species) [100,101,102,103]. The treatment with bacilli in the water showed lower species richness (alpha diversity) compared to the control and the other treatments. [54] found that the intestine of L. vannamei cultured in a nutrient-rich environment (shrimp feces and organic waste) showed higher bacterial diversity. It is possible that the treatment with bacilli in the water had a lower nutrient load and, although it presented a lower richness of bacteria, it did show greater survival. Beta diversity is the degree of change or replacement in species composition between different communities [104]. Regarding the analysis of diversity among communities (NMDS), this showed a grouping in each treatment; however, the bacterial community of the control was significantly different from the community of the treatments with bacteria in the water, where it was similar. In contrast, [105] found that the bacterial community in intestines of shrimp cultured in an indoor-culture system showed no variations. Similarly, [106] found that the intestine of L. vannamei fed with synbiotics and postbiotics of bacilli and vibrio showed similar bacterial communities.
The potential function of shrimp gut bacterial microbiota can be predicted using the KEGG database [107]. The functional profile has a fundamental role in the ecological balance of intestinal microbiota; however, it is important to mention that the predominant function is metabolism [106, 107]. [108] mentioned that overrepresented bacterial metabolism may be related to energy consumption to satisfy the physiological activities of the host (shrimp). In this work, the bacterial functional profile did not change significantly by the treatments. However, linoleic acid metabolism (fatty acid) was positively correlated with final weight. Fatty acids play a fundamental role in cell structure and cell homeostasis [109] and are a source of energy stored in triacylglycerols [110]. It is known that fatty acids modulate the immune response thanks to their influence on the structure, function, metabolism, surface proteins, and intracellular receptors of cells [109]. Linoleic acid stimulates the production of ROS by activating the NADPH oxidase enzyme in rat fibroblasts [111], which agrees with the positive correlation of the immune response as revealed by the detection of superoxide anion in the present study.
Quorum sensing (QS) is a cell-to-cell communication system that regulates biofilm formation and the expression of virulence genes [112, 113]. In this work, QS was not significantly different among treatments. However, a positive correlation with shrimp survival was observed, which suggests a benefit to its health. Similarly, QS had a health benefit on farmed L. vannamei shrimp [113] and the farmed turbot (Scophthalmus maximus) [112].
5. Conclusions
In hyperintensive culture systems, high stocking density produces excess nitrogenous wastes, which deteriorate water quality, affecting the physiology and gut microbiota of shrimp. The results of this study showed that bacilli and LAB in the water of hyper-intensive culture systems act as nitrifiers, modulate the intestinal microbiota and immune response, and improve the growth and survival of shrimp. To our knowledge, this is the first report on Pediococcus pentosaceus and Leuconostoc mesenteroides as nitrifying bacteria.
Author Contributions
Conceptualization, A.L.G. and R.E.M.; methodology, A.S.V.C., R.E.M., J.A.F.C., G.D.P., and A.L.G.; software, A.S.V.C. and A.L.G.; validation, A.S.V.C., R.E.M. and A.L.G.; formal analysis, A.S.V.C., R.E.M., and A.L.G.; investigation, A.S.V.C.; resources, A.L.G. and R.E.M.; data curation, A.S.V.C., R.E.M., J.A.F.C., G.D.P., and A.L.G.; writing—original draft preparation, A.S.V.C. and A.L.G; writing—review and editing, A.L.G, R.E.M, X.G. and C.G.G.; visualization, A.S.V.C. and R.E.M.; supervision, A.L.G and R.E.M.; project administration, A.L.G. and R.E.M.; funding acquisition, A.L.G. and R.E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants (20212061, 20221104) from SIP-IPN.
Data Availability Statement
The sequences were submitted to the NCBI (PRJNA1044443). The rest of the data is available upon request.
Acknowledgments
Ana S. Vega-Carranza acknowledges BEIFI from IPN and the Consejo Nacional de Humanidades, Ciencias y Tecnología (CONAHCyT, Mexico) for Ph.D. fellowships.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO. Estado Mundial de la Pesca y la Acuicultura 2020. Departamento de Pesca Acuicultura de la FAO. 2020.
- Villarreal, H.; Juárez, L. Super-intensive shrimp culture: Analysis and future challenges. J. World Aqua. Soc. 2012, 53, 928–932. [Google Scholar] [CrossRef]
- Thompson, F.L.; Abreu, P.C.; Wasielesky, W. Importance of biofilm for water quality and nourishment in intensive shrimp. Aquaculture 2002, 203, 263–278. [Google Scholar] [CrossRef]
- Dioses-Imán, D.S. Consorcios bacterianos nitrificantes inmovilizados en filtros de desechos agrícolas, como biorremediadores para cultivos de Litopenaeus vannamei, en agua dulce. Tesis. Universidad Nacional de Piura. Facultad de Ciencias. Escuela de Ciencias Biológicas Piura Perú, 2017, 50 pp.
- Hoang, P.H.; Nguyen, H.T.; Tran, T.T.; Do, L.P.; Le, T.N.C. Isolation and selection of nitrifying bacteria with high biofilm formation for treatment of ammonium polluted aquaculture water. J Vietn. Environ. 2016, 8, 33–40. [Google Scholar] [CrossRef]
- Ayyappan, S.; Mishra, S. Bioamelioration in aquaculture with a special reference to nitrifying bacteria. In: I.S.B. Singh, S.S. Pai, R. Philip and A. Mohandas (eds.) Aquaculture Medicine. CFDDM, CUSAT, India, 2003, pp 89-107.
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia-nitrogen in aquaculture systems. Aquaculture. 2006, 257, 346–358. [Google Scholar] [CrossRef]
- Claros-Bedoya, J.A. Graduate Thesis. Estudio del proceso de nitrificación y desnitrificación vía nitrito para el tratamiento biológico de corrientes de agua residual con alta carga de nitrógeno amoniacal. Universidad Politécnica de Valencia. 2012, 254 pp.
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Molec. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [CrossRef]
- Yang, X.P.; Wang, S.M.; Zhang, D.W.; Zhou, L.X. Isolation and nitrogen removal characteristics of an aerobic heterotrophic nitrifying–denitrifying bacterium, Bacillus subtilis A1. Bioresour. Technol. 2011, 102, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Noorak, S.; Rakkhiaw, S.; Limjirakhajornt, K.; Uppabullung, A.; Keawtawee, T.; Sangnoi, Y. Nitrite oxidizing bacteria for water treatment in coastal aquaculture system. IOP Conference Series: Environ. Earth Sci. 2018, 137, 012005. [Google Scholar] [CrossRef]
- Doñate, C.; Balasch, J.C.; Callol, A.; Bobe, J.; Tort, L.; MacKenzie, S. The effects of immunostimulation through dietary manipulation in the rainbow trout; evaluation of mucosal immunity. Mar. Biotechnol. 2010, 12, 88–99. [Google Scholar] [CrossRef]
- Luna-González, A.; Moreno-Herrera, J.T.; Campa-Córdova, Á.I.; González-Ocampo, H.A.; Fierro-Coronado, J.A.; Álvarez-Ruíz, P.; Bueno-Ibarra, M.A. Respuesta inmune y expresión de genes en el camarón blanco (Litopenaeus vannamei) inducida por inmunoestimulantes microbianos. Latin. Am. J. Aquatic. Res. 2013, 41, 898–907. [Google Scholar] [CrossRef]
- Lv, S.; Xu, J.; Zhao, J. Classification and phagocytosis of circulating haemocytes in Chinese mitten crab (Eriocheir sinensis) and the effect of extrinsic stimulation on circulating haemocytes in vivo. Fish Shellfish Immunol. 2014, 39, 415–422. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Shao, M.K.; Kang, H. Classification of haematopoietic cells and haemocytes in Chinese prawn Fenneropenaeus chinensis. Fish Shellfish Immunol. 2006, 21, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Fagutao, F.F.; Maningas, M.B.; Kondo, H.; Aoki, T.; Hirono, I. Transglutaminase regulates immune-related genes in shrimp. Fish Shellfish Immunol. 2012, 32, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Wang, J.X. Pattern recognition receptors acting in innate immune system of shrimp against pathogen infections. Fish Shellfish Immunol. 2013, 34, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Ponprateep, S.; Vatanavicharn, T.; Lo, C.F. Alpha-2-macroglobulin is a modulator of prophenoloxidase system in pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 62, 68–74. [Google Scholar] [CrossRef]
- Ortiz-Estrada, A.M.; Gollas-Galvan, T.; Martinez-Cordova, L.R.; Martinez-Porchas, M. Predictive functional profiles using metagenomic 16S rRNA data: a novel approach to understanding the microbial ecology of aquaculture systems. Rev. Aquacul. 2019, 11, 234–245. [Google Scholar] [CrossRef]
- Sha, Y.; Liu, M.; Wang, B.; Jiang, K.; Qi, C.; Wang, L. Bacterial population in intestines of Litopenaeus vannamei fed different probiotics or probiotic supernatant. J. Microbiol. Biotechnol. 2016, 26, 1736–1745. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Escamilla-Montes, R.; Luna-González, A.; Flores-Miranda, M.C.; Álvarez- Ruiz, P.; Fierro-Coronado, J.A. Isolation and characterization of potential probiotic bacteria suitable for mollusk larvae cultures. Thai J. Vet. Med. 2015, 45, 11. [Google Scholar] [CrossRef]
- Leyva-Madrigal, K.Y.; Luna-González, A.; Escobedo-Bonilla, C.M.; Fierro-Coronado, J.A.; Maldonado-Mendoza, I.E. Screening for potential probiotic bacteria to reduce prevalence of WSSV and IHHNV in whiteleg shrimp (Litopenaeus vannamei) under experimental conditions. Aquaculture 2011, 322-323, 16–22. [Google Scholar] [CrossRef]
- Jafari, S.M.; He, Y.; Bhandari, B. “Encapsulation of nanoparticles of dlimonene by spray drying: role of emulsifiers and emulsifying techniques, ” Dry. Technol. 2007, 25, 1079–1089. [Google Scholar]
- Lara-Espinoza, C.; Espinosa-Plasencia, A.; Rivera-Domínguez, M.; Astorga-Cienfuegos, K.; Acedo-Félix, E.; Bermúdez-Almada, M. Desarrollo de camarón Litopenaeus vannamei en un sistema de cultivo intensivo con biofloc y nulo recambio de agua. AquaTIC 2015, 43, 1–13. [Google Scholar]
- Ricker, W.E. Growth rates and models. In: W. S. Hoar, D. J. Randall & J. R. Brett (Eds.). Fish physiology, Volume VIII, Bioenergetics and growth. Academic Press. New York, United States. 1979, pp. 599–675.
- Vargas-Albores, F.; Guzmán, M.A.; Ochoa, J.L. An anticoagulant solution for haemolymph collection and prophenoloxidase studies of penaeid shrimp (Penaeus californiensis). Comp. Biochem. Physiol. Part A: Physiol. 1993, 106, 299–303. [Google Scholar] [CrossRef]
- Song, Y.L.; Hsieh, Y.T. Immunostimulation of tiger shrimp (Penaeus monodon) hemocytes for generation of microbicidalsubstances: Analysis of reactive oxygen species. Dev. Comp. Immunol. 1994, 18, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Strickland, J.D.H.; Parsons, T.R. A Parctical Handbook of Seawater Analysis. Bulletin 167. Second edition. 1972, .323 pp.
- Azmat, M.A.; Khan, I.A.; Cheema, H.M.; Rajwana, I.A.; Khan, A.S.; Khan, A.A. Extraction of DNA suitable for PCR applications from mature leaves of Mangifera indica L. J. Zhejiang Univ. Sci. 2012, 13, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring Microbial Diversity and Taxonomy Using SSU rRNA Hypervariable Tag Sequencing. PLOS Gen. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y. Design and experimental application of a novel non-degenerate universal primer set that amplifies prokaryotic 16S rRNA genes with a low possibility to amplify eukaryotic rRNA genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Volant, S.; Lechat, P.; Woringer, P.; Motref, L.; Campagne, P. SHAMAN: a user-friendly website for metataxonomic analysis from raw reads to statistical analysis. BMC Bioinforma. 2020, 21, 345. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, L.I.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: a web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 1–9. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nature Protocols 2020, 15, 1–23. [Google Scholar] [CrossRef]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero, N.; Laporte, P.; Balzergue, S. Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev. Cell. 2014, 30, 166–176. [Google Scholar] [CrossRef]
- Nagpal, S.; Haque, M.M.; Mande, S.S. Vikodak - A Modular Framework for Inferring Functional Potential of Microbial Communities from 16S Metagenomic Datasets. PLOS ONE 2016, 11, e0148347. [Google Scholar] [CrossRef] [PubMed]
- Daniel, P. Introducción a la Estadistica. 1997. MacGraw-Hill, Interamericana 428 p.
- Zokaeifar, H.; Babaei, N.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Balcazar, J.L. Administration of Bacillus subtilis strains in the rearing water enhances the water quality, growth performance, immune response, and resistance against Vibrio harveyi infection in juvenile white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2014, 36, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Lieke, T.; Meinelt, T.; Hoseinifar, S.H.; Pan, B.; Straus, D.L.; Steinberg, C.E. Sustainable aquaculture requires environmental-friendly treatment strategies for fish diseases. Rev. Aquacu. 2019, 12, 943–965. [Google Scholar] [CrossRef]
- Lin, Y.C.; Chen, J.C. Acute toxicity of ammonia on Litopenaeus vannamei Boone juveniles at different salinity levels. J. Exp. Mar. Biol. Ecol. 2001, 259, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chen, J.C. Acute toxicity of nitrite on Litopenaeus vannamei (Boone) juveniles at different salinity levels. Aquaculture. 2003, 224, 193–201. [Google Scholar] [CrossRef]
- Romano, N.; Zeng, C. Toxic Effects of Ammonia, Nitrite, and Nitrate to Decapod Crustaceans: A Review on Factors Influencing their Toxicity, Physiological Consequences, and Coping Mechanisms. Rev. Fish. Sci. Aquac. 2013, 21, 1–21. [Google Scholar] [CrossRef]
- Barbieri, E.; Bondioli, A.C.V.; Melo, C.; Henriques, M.B. Nitrite toxicity to Litopenaeus schmitti (Burkenroad, 1936, Crustacea) at different salinity levels. Aquac. Res. 2014, 1–9. [Google Scholar] [CrossRef]
- Suo, Y.; Erchao, L.; Tongyu, L.; Yongyi, J.; Jian, G.; Qin, Z.G.; Liqiao, C. Response of gut health and microbiota to sulfide exposure in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 63, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Anwar, K.; Safitri, R.; Fajriani, N.; Gifari, Z.A.; Wariata, I.W.; Rosyidi, A. In vitro screening of ammonia and nitrite-degrading bacteria isolated from broiler chicken (Gallus gallus domesticus) intestines and pond sediment of nile tilapia (Oreochromis niloticus): a preliminary study. IOP Conf. Ser. Earth Environ. Sci. 2021, 913, 012072. [Google Scholar] [CrossRef]
- Lalloo, R.; Ramchuran, S.; Ramduth, D.; Görgens, J.; Gardiner, N. Isolation and selection of Bacillus spp. as potential biological agents for enhancement of water quality in cultura of ornamental fish. J. Appl. Microbiol. 2007, 103, 1471–1479. [Google Scholar]
- Daims, H.; Lücker, L.; Wagner, M.A. New Perspective on Microbes Formerly Known as Nitrite-Oxidizing Bacteria. Trends Microbiol. 2016, 24, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Zokaeifar, H.; Balcázar, J.L.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Arshad, A.; Njat, N. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2012, 33, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Lezama, C.; Paniagua, J.; Zamora, J. Bioremediation of effluents ones of the culture of Litopenaeus vannamei (Boone, 1931) using microbial mats in a recirculating system. Latin Am. J. Aquatic Res. 2010, 38, 129–142. [Google Scholar] [CrossRef]
- Sang, C. G.; Fu, Y.-W.; Guo, S.-Q.; Luo, J.-J.; Zhang, Q.-Z. Isolation and Characterization of an Aerobic Denitrifier Bacillus sp. SC16 from an Intensive Aquaculture Pond. Water. 2020, 12, 3559. [Google Scholar]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Verreth, J.; Verdegem, M. The Colonization Dynamics of the Gut Microbiota in Tilapia Larvae. PLoS ONE 2014, 9, e103641. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-Gempeler, C.; Leibold, M.A. Multiple colonist pools shape fiddler crab-associated bacterial communities. The ISME J. 2018, 12, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yayu, Q.; Jian, G.; Qin, L.C.; Fenglu, H.; Erchao, L. Deep insight into bacterial community characterization and relationship in the pond water, sediment and the gut of shrimp (Penaeus japonicus). Aquaculture 2021, 539, 736658. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nat. Cell Biol. 2016, 535, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Nimrat, S.; Suksawat, S.; Boonthai, T.; Vuthiphandchai, V. Potential Bacillus probiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeus vannamei). Vet. Microbiol. 2012, 159, 443–450. [Google Scholar] [CrossRef]
- Madani, N.S.H.; Adorian, T.; Ghafari, H.; Farsani, J.; Hoseinifar, S.H. The effects of dietary probiotic Bacilli (Bacillus subtilis and Bacillus licheniformis) on growth performance, feed efficiency, body composition and immune parameters of whiteleg shrimp (Litopenaeus vannamei) postlarvae. ” Aquac. Res. 2018, 49, 1926–1933. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Dadar, M.; Ringø, E. Modulation of nutrient digestibility and digestive enzyme activities in aquatic animals: the functional feed additives scenario. Aquac. Res. 2017, 48, 3987–4000. [Google Scholar] [CrossRef]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: lights and shadows. Fron. Cell. Infec. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.; Plínio, S.A.; Valenzuela, J.; Rodriguez-Fuentes, G.; Campos, B. R.; Wasielesky Jr, W.; Gaxiola, G.G. Chronic effect of nitrite on the rearing of the white shrimp Litopenaeus vannamei in two salinities. Mar. Freshwater Behav. Physiol. 2016, 49, 201–211. [Google Scholar] [CrossRef]
- Huang, M.; Jia, X.; Qiuran, Y.; Chang, X.; Li, Z.; Jian, G.Q. Toxic effect of chronic nitrite exposure on growth and health in Pacific white shrimp Litopenaeus vannamei. Aquaculture. 2020, 529, 735664. [Google Scholar] [CrossRef]
- Campos, B.R.; de Furtado, P.S.; D'Incao, F.; Wasielesky, W.; Poersch, L.H. Compostos nitrogenados sobre o consumo alimentar de camarão-rosa Farfantepenaeus brasiliensis. Ciên Rural 2013, 43, 2202–2207. [Google Scholar] [CrossRef]
- Han, S.; Wang, B.; Wang, M.; Liu, Q.; Zhao, W.; Wang, L. Effects of ammonia and nitrite accumulation on the survival and growth performance of white shrimp Litopenaeus vannamei. Inv. Surv. J. 2017, 14, 221–232. [Google Scholar]
- Lee, D.O.C.; Wickins, J.F. Crustacean farming: ranching and culture. First Edition. Blackwell Sci Publ Oxford U.K., 1992, 392 p.
- Boyd, C.E. Consideraciones sobre la calidad del agua y del suelo en cultivos de camarón. In: Haws, M. C., Boyd, C.E. (eds.). Métodos para Mejorar la Camaronicultura en Centroamérica. Editorial-Imprenta UCA, Managua, Nicaragua, 2002, pp 24-25.
- Boardman, G.D.; Starbuck, S.M.; Hudgins, D.B.; Li, X.Y.; Kuhn, D.D. Toxicity of ammonia to three marine fish and three marine invertebrates. Environ. Toxicol. 2004, 19, 34–142. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, J.; Huang, J.; Shigui, J. Effects of Dietary Clostridium butyricum on the Growth, Digestive Enzyme Activity, Antioxidant Capacity, and Resistance to Nitrite Stress of Penaeus monodon. Prob. Antim. Prot. 2019, 11, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gai, C.; Ye, G. Aeromonas hydrophila, an emerging causative agent of freshwaterfarmed whiteleg shrimp Litopenaeus vannamei. Microorganisms 2019, 7, 1–20. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chen, J.C.; Lin, Y.C.; Yeh, S.T.; Huang, C.L. White Shrimp Litopenaeus vannamei That Have Received Gracilaria tenuistipitata Extract Show Early Recovery of Immune Parameters after Ammonia Stressing. Mar. Drugs. 2015, 13, 3606–3624. [Google Scholar] [CrossRef]
- Liao, S.; Li, Q.; Wang, A.; Xian, J.; Chen, X.; Gou, N. Effect of nitrite on immunity of the white shrimp Litopenaeus vannamei at low temperature and low salinity. Ecotoxicology 2012, 21, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.C.; Yeh, S.P.; Hu, S.Y.; Lin, H.L; Liu, C.H. Intestinal microbiota of white shrimp, Litopenaeus vannamei, fed diets containing Bacillus subtilis E20-fermented soybean meal (FSBM) or an antimicrobial peptide derived from B. subtilis E20- FSBM. Aquac. Res. 2019, 51, 41–50. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, X.; Xiong, J.; Zhu, J.; Wang, Y.; Zhao, Q.; Chen, H.; Guo, A.; Wu, J.; Dai, H. Bacterioplankton assemblages as biological indicators of shrimp health status. Ecol. Indic. 2014, 38, 218–224. [Google Scholar] [CrossRef]
- Cornejo-Granados, F.; Gallardo-Becerra, L.; Leonardo-Reza, M.; Ochoa-Romo, J.P.; Ochoa-Leyva, A. A meta-analysis reveals the environmental and host factors shaping the structure and function of the shrimp microbiota. Peer J. 2018, 6, e5382. [Google Scholar] [CrossRef] [PubMed]
- Li, E.C.; Xu, C.; Wang, X.D.; Wang, S.F.; Zhao, Q.; Zhang, M.L.; Qin, J.G.; Chen, L.Q. Gut microbiota and its modulation for healthy farming of Pacific white shrimp Litopenaeus vannamei. Rev. Fish Sci. Aquacult. 2018, 26, 381–399. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, K.; Wu, J.; Qiuqian, L.; Yang, K.; Qian, Y.; Demin, Z. Changes in intestinal bacterial communities are closely associated with shrimp disease severity. Appl. Microbiol. Biotechnol. 2015, 99, 6911–6919. [Google Scholar] [CrossRef] [PubMed]
- Rungrassamee, W.; Klanchui, A.; Maibun, S.; Karoonuthaisiri, N. Bacterial dynamics in intestines of the black tiger shrimp and the Pacific white shrimp during Vibrio harveyi exposure. J. Invertebr. Pathol. 2015, 133, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wen, M.; Xiang, L.; Shen, H.; Jiang, G.; Cheng, J.; Hu, Y.; Qian, J. Segmental variations in intestinal microbiota composition and functional capacity along the digestive tract of Litopenaeus vannamei. Aquac. Rep. 2024, 34, 101922. [Google Scholar] [CrossRef]
- Daniel, H.; Gholami, A.M.; Berry, D.; Desmarchelier, C.; Hahne, H.; Loh, G.; Mondot, S.; Lepage, P.; Rothballer, M.; Walker, A.; et al. High-fat diet alters gut microbiota physiology in mice. ISME J. 2014, 8, 295–308. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M.H. Gut Bacteroides species in health and disease. Gut Microbes. 2021, 13, 1848158. [Google Scholar] [CrossRef]
- Larsbrink, J.; Zhu, Y.; Kharade, S.S.; Kwiatkowski, K.J.; Eijsink, V.G.H.; Koropatkin, N. M.; McBride, M.J.; Pope, P.B. A polysaccharide utilization locus from Flavobacterium johnsoniae enables conversion of recalcitrant chitin. Biotechnol. Biofuels 2016, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Otta, S.; Karunasagar, I.K.I. Bacterial fora associated with shrimp culture ponds growing Penaeus monodon. J. Aquac. Trop. 1999, 14, 309–318. [Google Scholar]
- Intriago, J.; Quimi, J.; Risco, J.; López, J.; Yalta, J.; Bermudez, M.; Mialhe, E. Metagenómica de la microbiota de juveniles de Litopenaeus vannamei inoculados con bacterias probióticas y patógenas. AquaTIC 2018, 51, 16–29. [Google Scholar]
- Vega-Carranza, A.S. Efecto de plantas medicinales, probióticos, prebióticos, paraprobióticos y postbióticos en la microbiota intestinal, crecimiento, supervivencia y respuesta inmune del camarón blanco (Penaeus vannamei) retado con Vibrio parahaemolyticus. MS Dissertion. 2020. CIIDIR-IPN. 100 p.
- Zheng, Y.; Yu, M.; Liu, Y.; Su, Y.; Xu, T.; Yu, M.; Zhang, X.H. Comparison of cultivable bacterial communities associated with Pacific white shrimp (Litopenaeus vannamei) larvae at different health statuses and growth stages. Aquaculture. 2016, 451, 163–169. [Google Scholar] [CrossRef]
- Gao, S.; Pan, L.; Huang, F.; Song, M.; Tian, C.; Zhang, M. Metagenomic insights into the structure and function of intestinal microbiota of the farmed Pacific white shrimp (Litopenaeus vannamei). Aquaculture. 2019, 499, 109–118. [Google Scholar] [CrossRef]
- Ramirez, M.; Domínguez-Borbor, C.; Salazar, L.; Debut, A.; Vizuete, K.; Sonnenholzner, S.; Alexis, F.; Rodríguez, J. The probiotics Vibrio diabolicus (Ili), Vibrio hepatarius (P62), and Bacillus cereus sensu stricto (P64) colonize internal and external surfaces of Penaeus vannamei shrimp larvae and protect it against Vibrio parahaemolyticus. Aquaculture 2022, 549, 737826. [Google Scholar] [CrossRef]
- Criminger, J.D.; Hazen, T.H.; Sobecky, P.A.; Lovell, C.R. Nitrogen Fixation by Vibrio parahaemolyticus and Its Implications for a New Ecological Niche. Appl. Environ. Microbiol. 2007, 73. [Google Scholar] [CrossRef] [PubMed]
- Huang, W. Characterization and classification of novel marine nitrogen-fixing bacterial isolates from the lagoon sediment of Dongsha Island. 2021. [Google Scholar] [CrossRef]
- Widanarni, W.; Tepu, I.; Sekenda, S.; Setiawati, M. Seleksi bakteri probiotik untuk biokontrol vibriosis pada larva udang windu (Penaues monodon) menggunakan cara kultur bersama. J. Riset Akuakultur 2009, 4, 95–105. [Google Scholar] [CrossRef]
- Sorieul, N.; Wabete, D.; Ansquer, J.R.; Mailliez, M.; Pallud, C.; Zhang, M.; Lindivat, V.; Buolo, D. Pham. Survival improvement conferred by the Pseudoalteromonas sp. NC201 probiotic in Litopenaeus stylirostris exposed to Vibrio nigripulchritudo infection and salinity stress. Aquaculture 2018, 495, 888–898. [Google Scholar] [CrossRef]
- Wang, W.L.; Xu, S.Y.; Ren, Z.G.; Tao, L.; Jiang, J.W.; Zheng, S.S. Application of metagenomics in the human gut microbiome. World J. Gastroenterol. 2015, 21, 803–814. [Google Scholar] [CrossRef]
- Hamsah, H.; Widanarni, W.; Alimuddin, A.; Yuhana, M.; Junior, M.Z.; Hidayatullah, D. Immune response and resistance of Pacific white shrimp larvae administered probiotic, prebiotic, and synbiotic through the bio-encapsulation of Artemia sp. Aquac. Intern. 2019, 27, 567–580. [Google Scholar] [CrossRef]
- Ramadhani, D.E.; Widanarni, W.; Sukenda, S. Microencapsulation of probiotics and its applications with prebiotic in Pacific white shrimp larvae through Artemia sp. Jurnal Akuakultur Indonesia 2019, 18, 130–140. [Google Scholar] [CrossRef]
- Le Chatelier, E. T.; Nielsen, J.; Qin, E.; Prifti, F.; Hildebrand, G. Richness of human gut microbiome correlates with metabolic markers. Nature. 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-Induced Obesity Is Linked to Marked but Reversible Alterations in the Mouse Distal Gut Microbiome. Cell Host Microb. 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Bellwood, D.R.; Folke, C.; Steneck, R.; Wilson, J. New paradigms for supporting the resilience of marine ecosystems. Trends Ecol. Evol. 2005, 20, 380–386. [Google Scholar] [CrossRef]
- Chao, A.; Robin, L.; Chazdon, R.; Colwell, K.; Tsung-Jen, S. Abundance-based similarity indices and their estimation when there are unseen species in samples. Biometrics. 2016, 62, 361–371. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scandinavian J. Statistics. 1984, 11, 265–270. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R. M.; Hartmann, E.; Hollister, B. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Handelsman, J. Introducing SONS, a tool for operational taxonomic unit-based comparisons of microbial community memberships and structures. Appl. Environ. Microbiol. 2006, 72, 6773–6779. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar] [CrossRef]
- Landsman, A.; St-Pierre, B.; Rosales-Leija, M.; Brown, M.; Gibbons, W. Investigation of the potential effects of host genetics and probiotic treatment on the gut bacterial community composition of aquaculture- raised Pacific whiteleg shrimp Litopenaeus vannamei. Microorganisms. 2019, 7, 217. [Google Scholar] [CrossRef]
- Vega-Carranza, A.S.; Escamilla-Montes, R.; Luna-González, A.; Fierro-Coronado, J.A.; Diarte-Plata, G.; García-Gutiérrez, C. Survival, immune response, and gut microbiota in Litopenaeus vannamei fed with synbiotics and postbiotics and challenged with Vibrio parahaemolyticus. Aquac. Intern. 2024, 32, 361–381. [Google Scholar] [CrossRef]
- Cheng, Y.; Ge, C.; Li, W.; Yao, H. The intestinal bacterial community and functional potential of Litopenaeus vannamei in the coastal areas of China. Microorganisms 2021, 9, 1793. [Google Scholar] [CrossRef] [PubMed]
- Dopson, M.; Ossandon, F.J.; Lövgren, L.; Holmes, D.S. Metal resistance or tolerance? Acidophiles confront high metal loads via both abiotic and biotic mechanisms. Frontiers in Microbiology 2014, 5, 157. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Yun, W.; Xian, D.; Dalin, X.; Jiasong, Z. Response of intestine microbiota, digestion, and immunity in Pacific white shrimp Litopenaeus vannamei to dietary succinate. Aquaculture 2020, 517, 734–762. [Google Scholar] [CrossRef]
- Wang, W.L.; Xu, S.Y.; Ren, Z.G.; Tao, L.; Jiang, J.W.; Zheng, S.S. Application of metagenomics in the human gut microbiome. World J. Gastroenterol. 2015, 21, 803–814. [Google Scholar] [CrossRef]
- Brown, H.A.; Marnett, L.J. Introduction to lipid biochemistry, metabolism, and signaling. Chemical Reviews 2011, 111, 5817–5820. [Google Scholar] [CrossRef] [PubMed]
- Radzikowska, U.; Rinaldi, A.; Çelebi, O.; Sözener, Z.; Karaguzel, D.; Wojcik, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, E.; Levada-Pires, A.C.; Pithon-Curi, T.C.; Curi, R. Systematic study on ROS production induced by oleic, linoleic, and gamma-linolenic acids in human and rat neutrophils. Free Rad. Biol. Med. 2006, 41, 1124–1132. [Google Scholar] [CrossRef]
- Xing, M.; Zhanhu, H.; Jianbo, Y.; Yuan, L.; Yanmei, Q.; Bin, L. Taxonomic and functional metagenomic profiling of gastrointestinal tract microbiome of the farmed adult turbot (Scophthalmus maximus). FEMS Microbiol. Ecol. 2013, 86, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Luqing, P.; Fei, H.; Shuo, G.; Chen, S.; Mingzhu, Z.; Ziyan, H. Metagenomic analysis of composition, function and cycling processes of microbial community in water, sediment and effluent of Litopenaeus vannamei farming environments under different culture modes. Aquaculture 2019, 506, 280–293. [Google Scholar] [CrossRef]
Figure 1.
Concentration of TAN, nitrites, and nitrates in the shrimp culture system without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
Figure 1.
Concentration of TAN, nitrites, and nitrates in the shrimp culture system without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
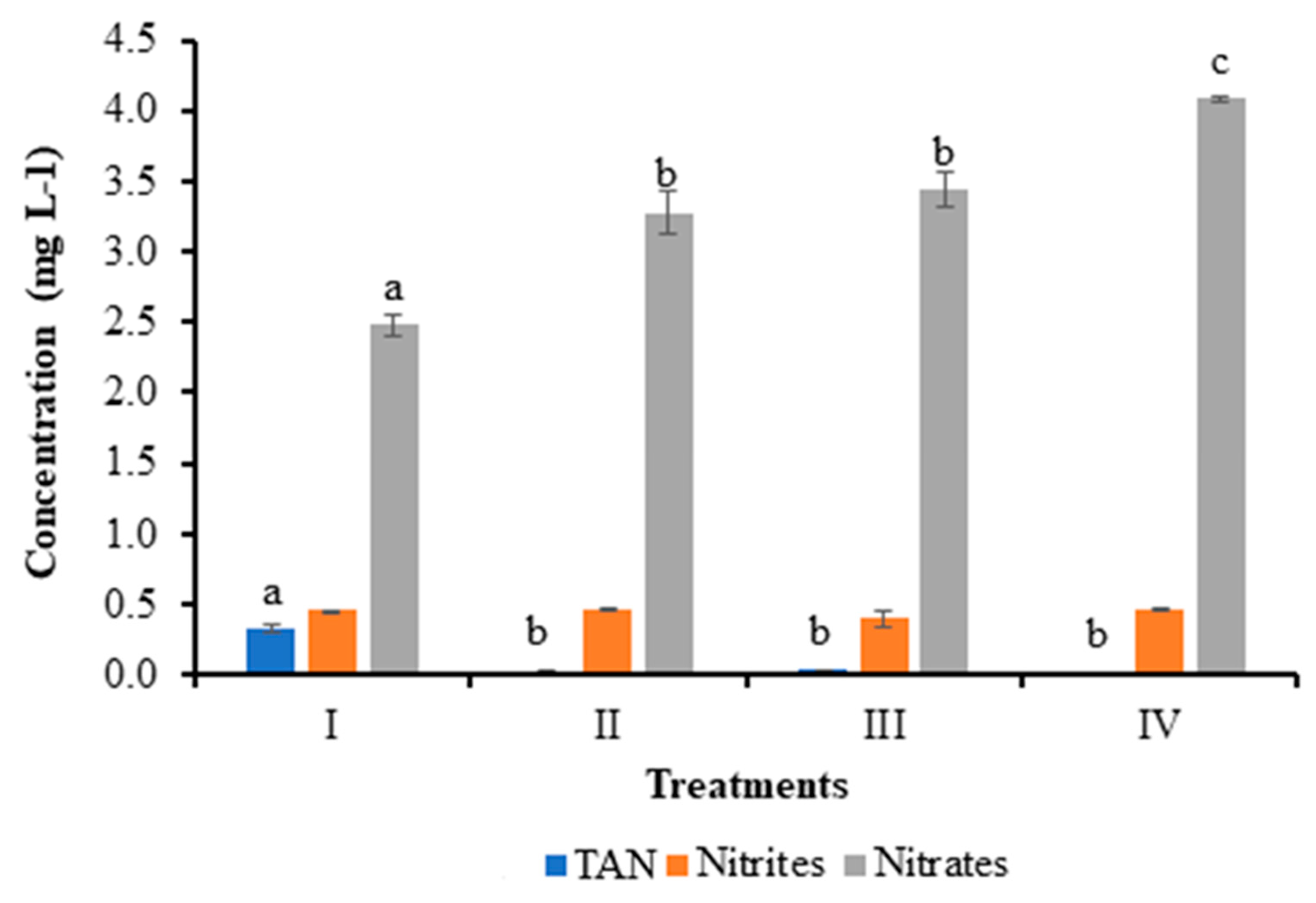
Figure 2.
Survival of shrimp cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
Figure 2.
Survival of shrimp cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
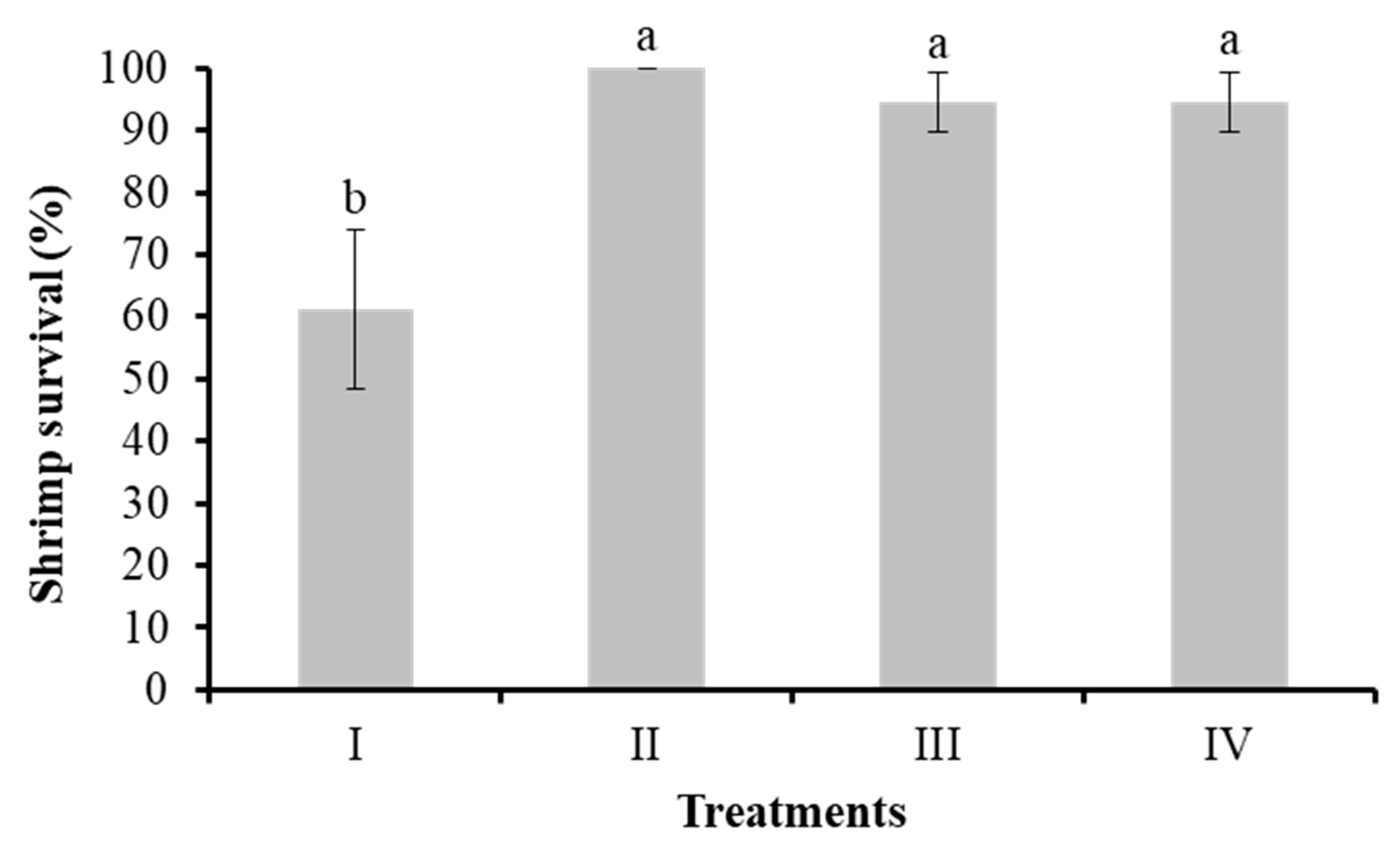
Figure 3.
Total hemocyte count in L. vannamei cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD.
Figure 3.
Total hemocyte count in L. vannamei cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD.
Figure 4.
Superoxide anion in hemolymph of L. vannamei cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
Figure 4.
Superoxide anion in hemolymph of L. vannamei cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
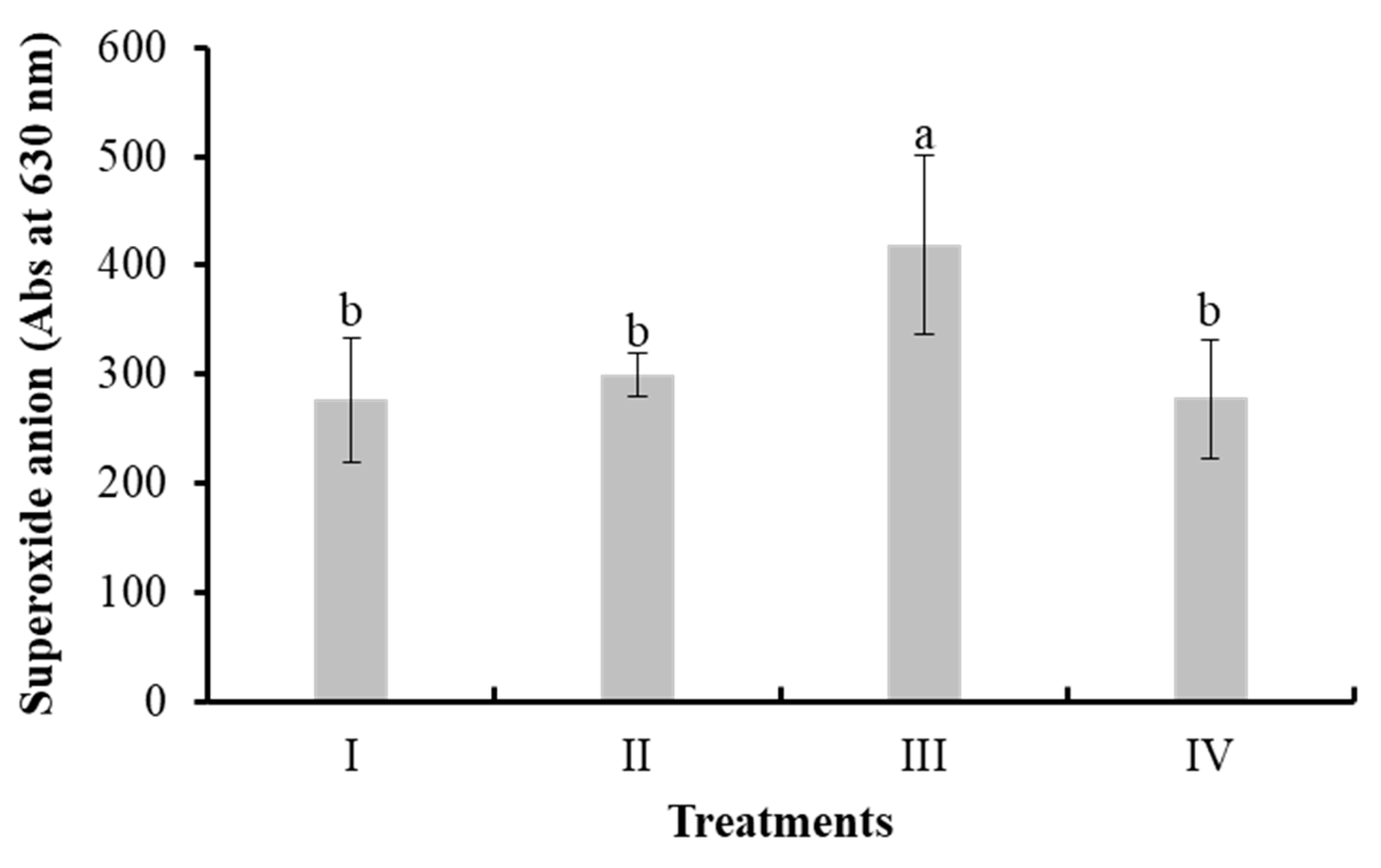
Figure 5.
Phenoloxidase activity in hemolymph of L. vannamei cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
Figure 5.
Phenoloxidase activity in hemolymph of L. vannamei cultured without water exchange and treated with bacteria. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Data are mean ± SD. Different letters indicate significant differences.
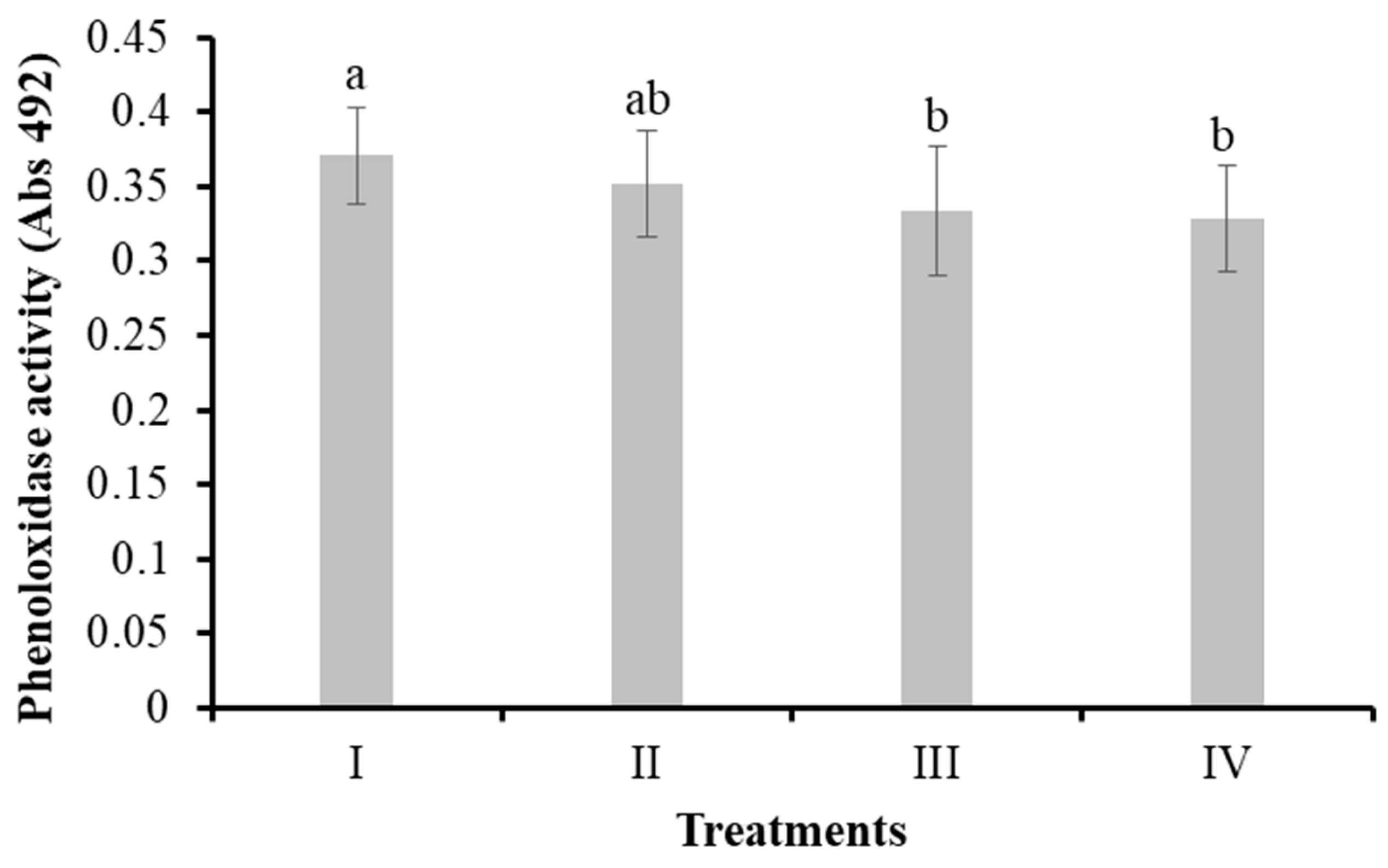
Figure 6.
Venn analysis of the bacterial communities in the intestine of shrimp at the OTUs level. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water.
Figure 6.
Venn analysis of the bacterial communities in the intestine of shrimp at the OTUs level. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water.
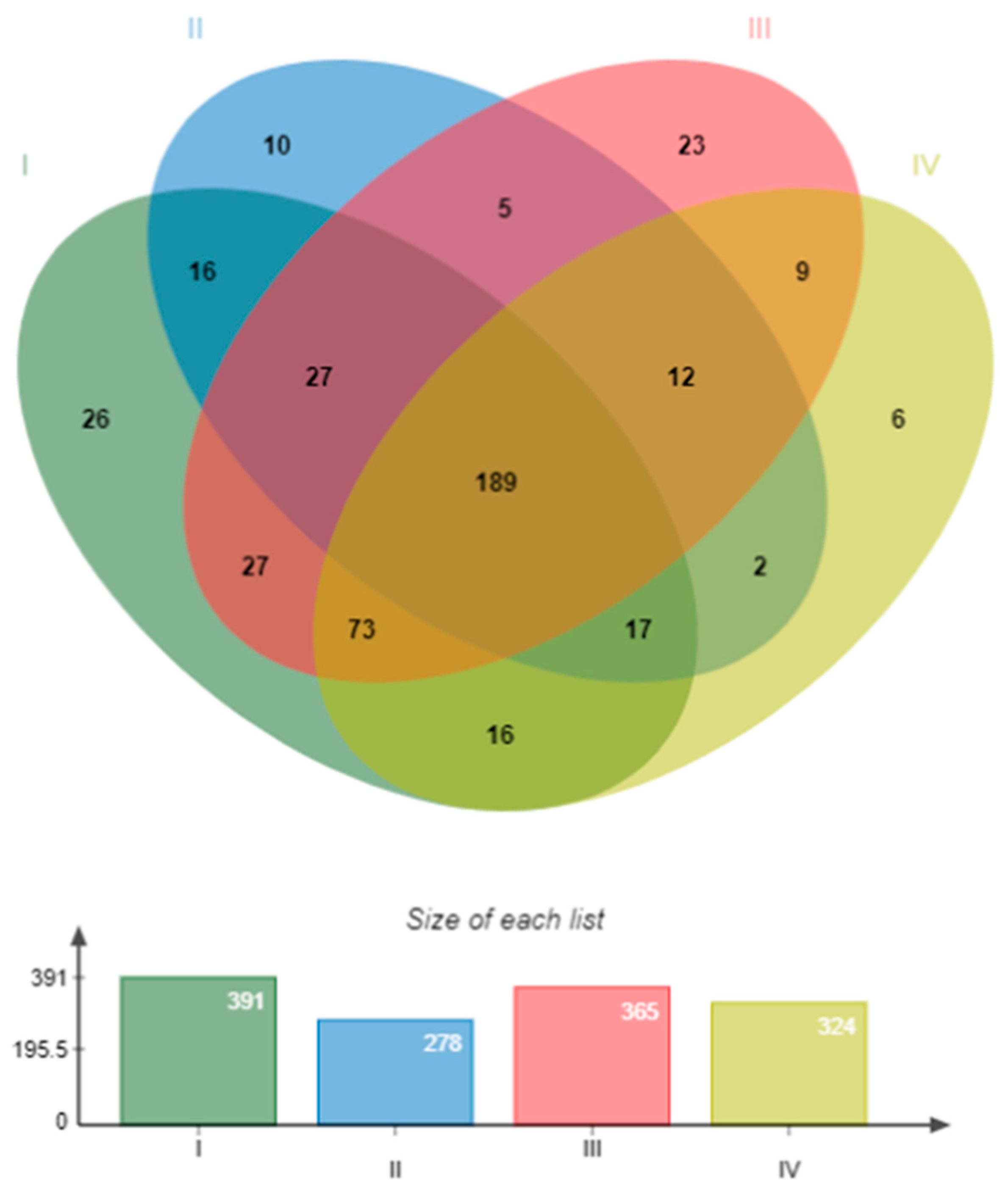
Figure 7.
Most abundant bacterial phyla (%) in the intestines of shrimp. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Patescibacteria phylum (*no significant differences [P > 0.05], **significant differences [P < 0.05]). The analysis was done with Shaman.
Figure 7.
Most abundant bacterial phyla (%) in the intestines of shrimp. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Patescibacteria phylum (*no significant differences [P > 0.05], **significant differences [P < 0.05]). The analysis was done with Shaman.
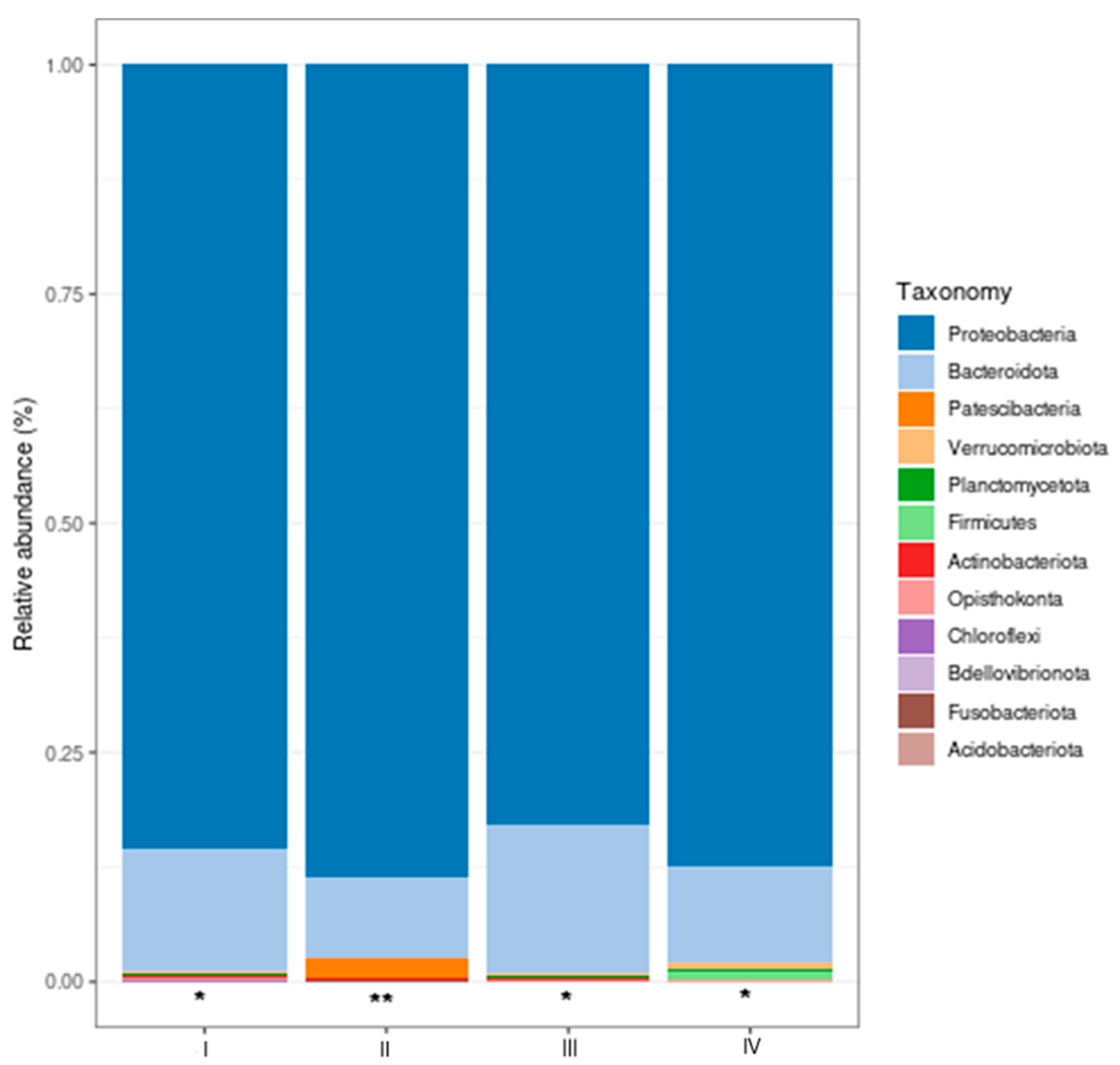
Figure 8.
Most abundant bacterial genera (%) in the intestines of shrimp. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Different letters indicate significant differences (P < 0.05). The analysis was done with Shaman.
Figure 8.
Most abundant bacterial genera (%) in the intestines of shrimp. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. Different letters indicate significant differences (P < 0.05). The analysis was done with Shaman.
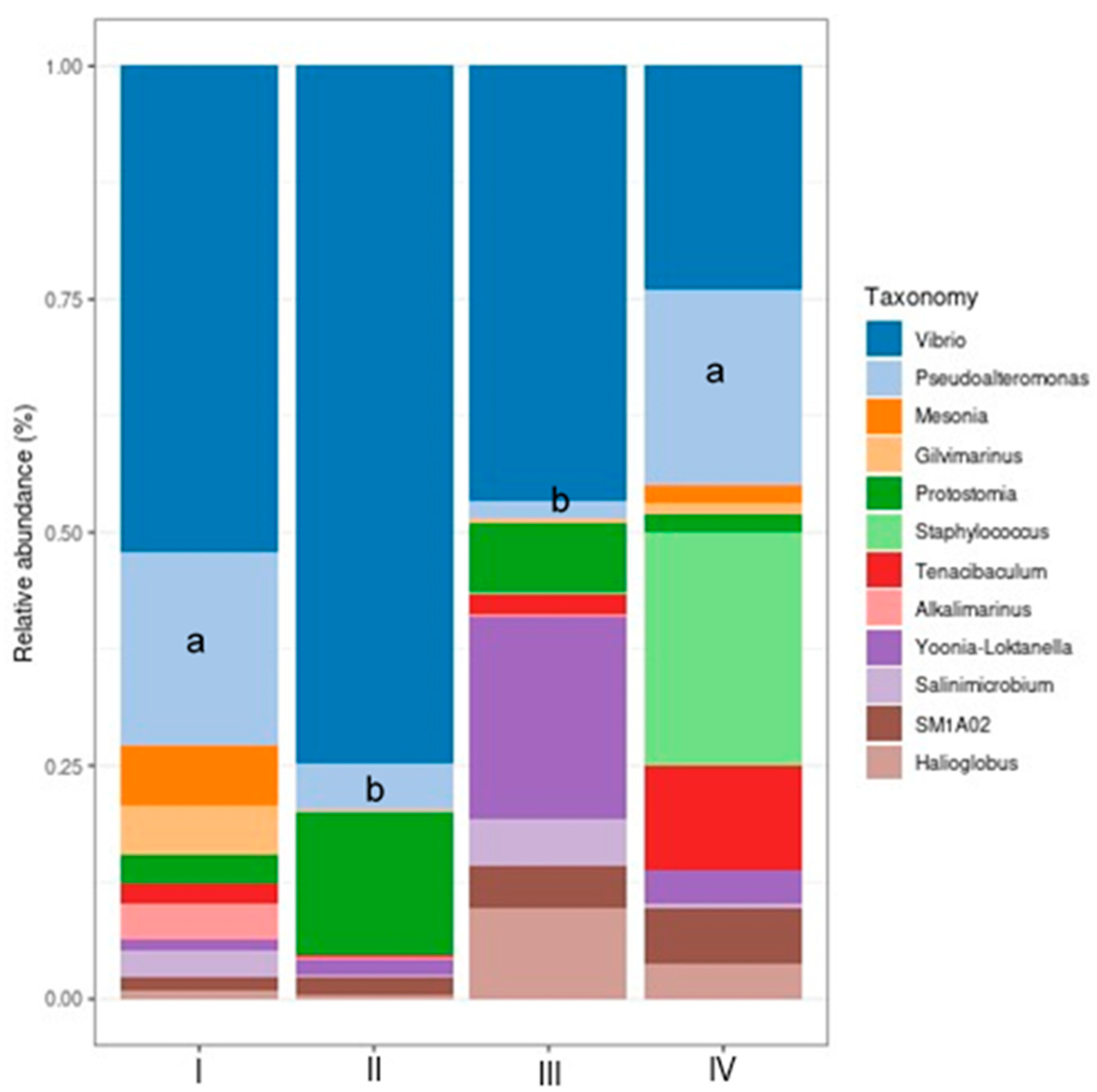
Figure 9.
Beta diversity of intestinal microbiota of L. vannamei at the genus level using non-metric multi-dimensional scaling based on Jaccard distances in MicrobiomeAnalyst. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. ANOSIM test, P < 0.008.
Figure 9.
Beta diversity of intestinal microbiota of L. vannamei at the genus level using non-metric multi-dimensional scaling based on Jaccard distances in MicrobiomeAnalyst. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. ANOSIM test, P < 0.008.
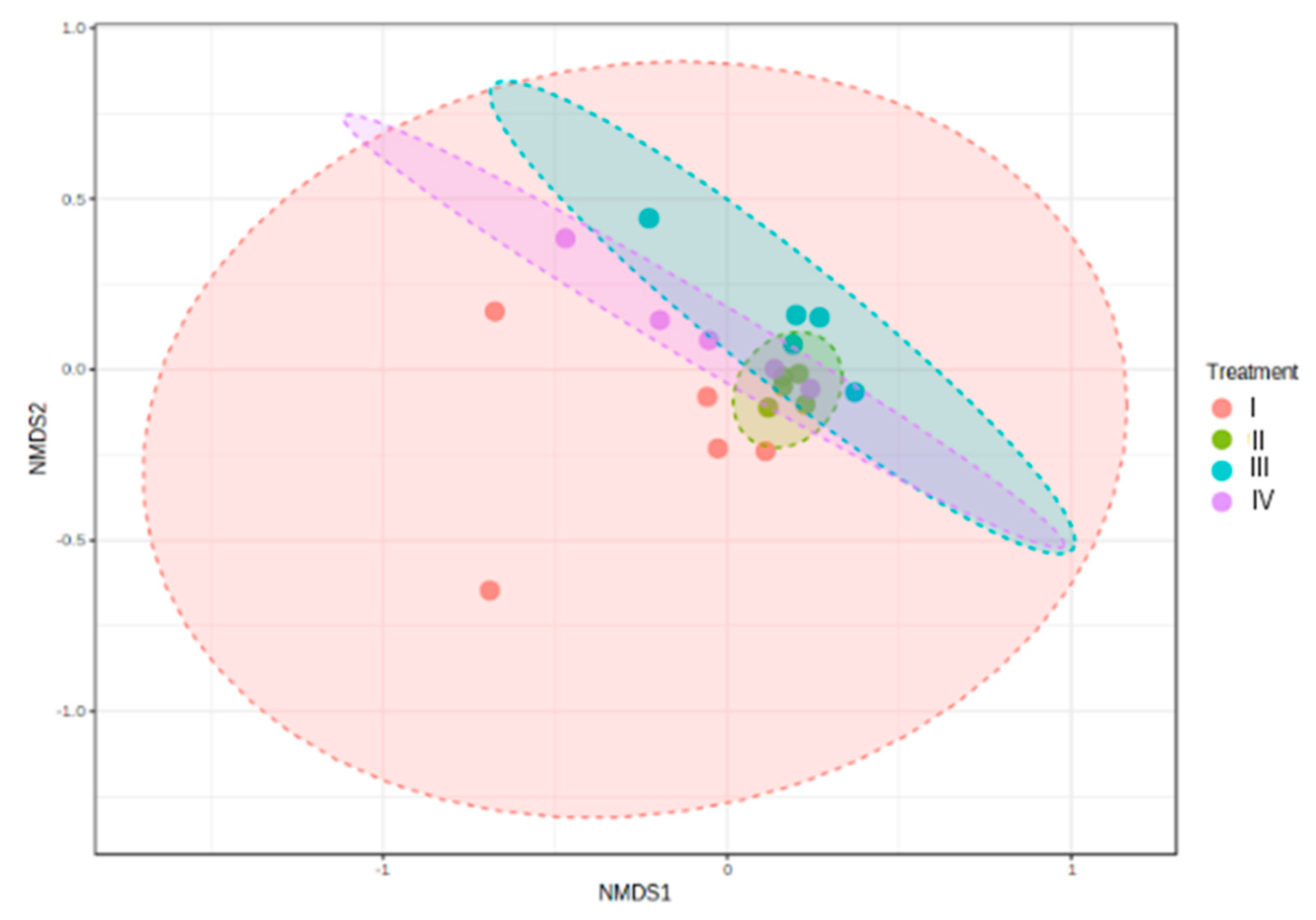
Figure 10.
Correlation among linoleic acid metabolism of intestinal bacteria of L. vannamei and immune and productive variables. Spearman correlation analysis.
Figure 10.
Correlation among linoleic acid metabolism of intestinal bacteria of L. vannamei and immune and productive variables. Spearman correlation analysis.
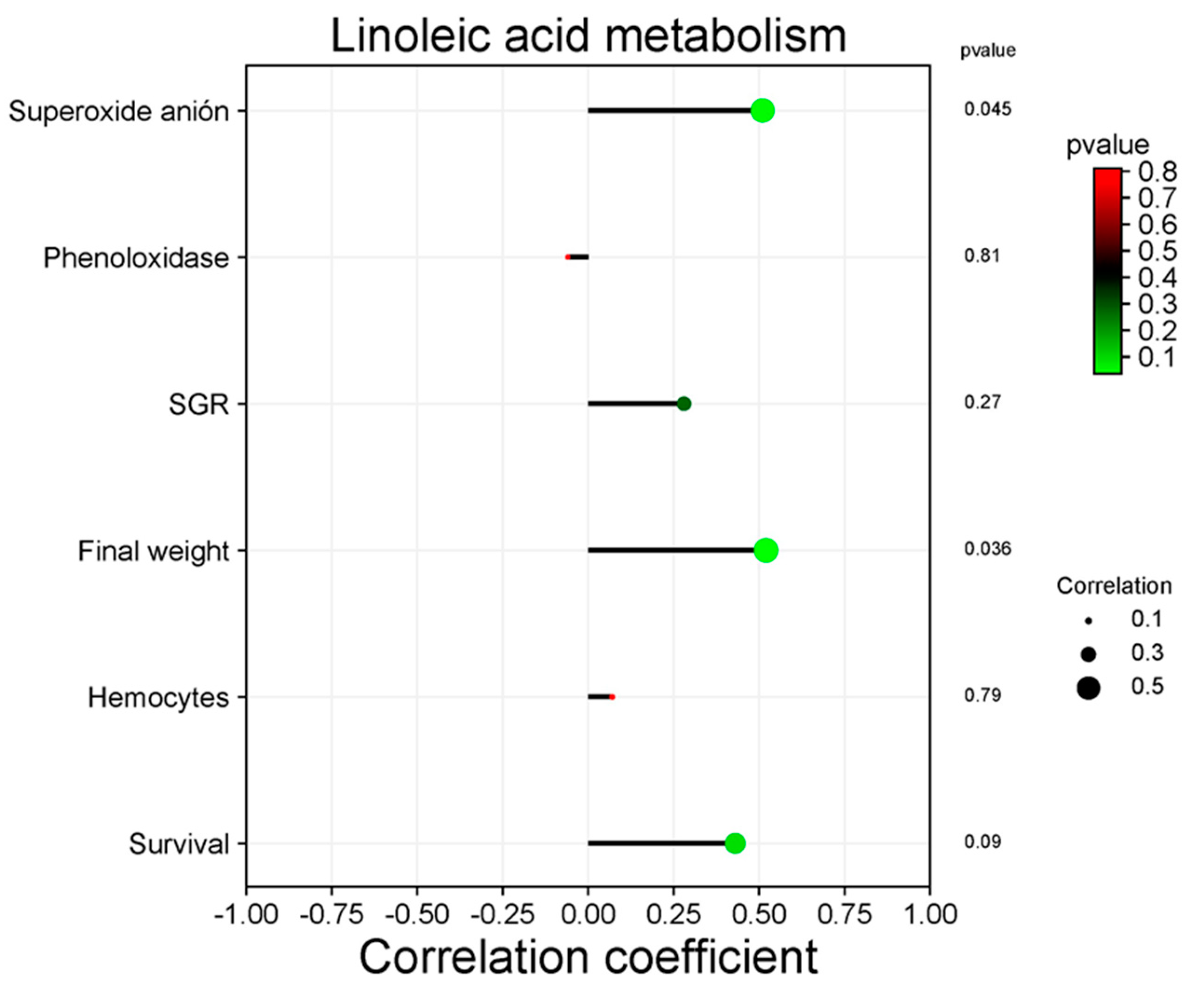
Figure 11.
Correlation among quorum sensing of intestinal bacteria of L. vannamei and immune and productive variables. Spearman correlation analysis.
Figure 11.
Correlation among quorum sensing of intestinal bacteria of L. vannamei and immune and productive variables. Spearman correlation analysis.
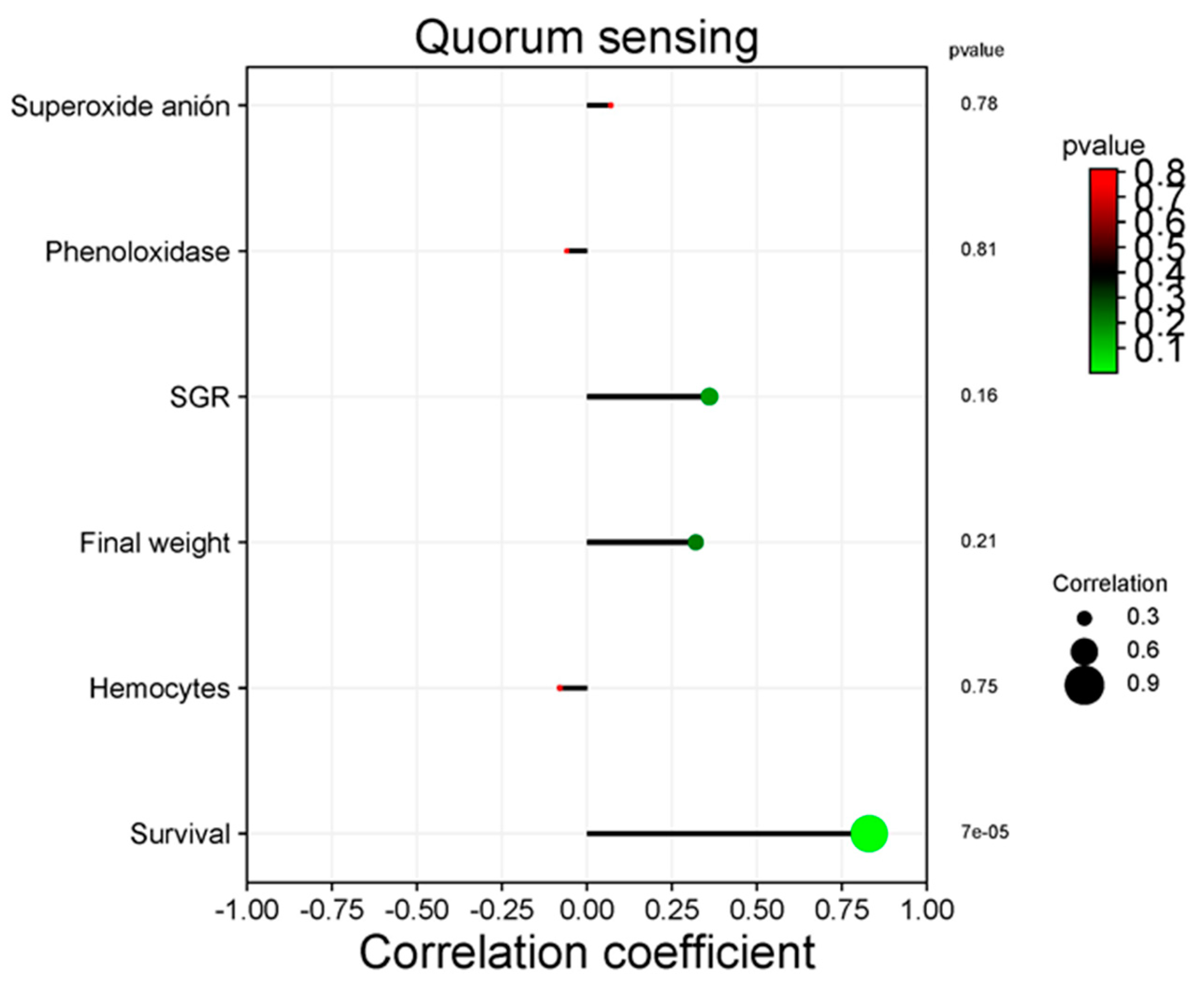

Table 1.
Physicochemical parameters in shrimp culture system without water exchange and treated with nitrifying bacteria. Treatments: I) Control; II) Bacilli in the water); III) LAB in the water; IV) Bacilli + LAB in the water. Values = mean ± SD.
Table 1.
Physicochemical parameters in shrimp culture system without water exchange and treated with nitrifying bacteria. Treatments: I) Control; II) Bacilli in the water); III) LAB in the water; IV) Bacilli + LAB in the water. Values = mean ± SD.
| Treatment | DO (mg mL-1) | pH | T (°C) | S (PSU) |
|---|---|---|---|---|
| Control (I) | 5.2±0.05 | 8.2±0.05 | 29.8±0.3 | 30±0.03 |
| II | 5.2±0.01 | 8.2±0.02 | 30.0±0.4 | 30±0.02 |
| III | 5.2±0.04 | 8.2±0.04 | 29.8±0.5 | 30±0.04 |
| IV | 5.3±0.08 | 8.2±0.06 | 30.0±0.3 | 30±0.03 |
| Optimal range | 4 to 10 | 8.1 to 9 | 23 to 30 | 15 to 35 |
Table 2.
Growth of shrimp cultured without water exchange and treated with bacteria. Control and with bacteria in the water: I) Control without bacteria; II) bacilli); III) LAB; IV) bacilli + LAB. Data are mean ± SD. Different letters indicate significant differences.
Table 2.
Growth of shrimp cultured without water exchange and treated with bacteria. Control and with bacteria in the water: I) Control without bacteria; II) bacilli); III) LAB; IV) bacilli + LAB. Data are mean ± SD. Different letters indicate significant differences.
| Shrimp growth | I | II | III | IV |
|---|---|---|---|---|
| Initial weigth (g) | 0.7±0.05 | 0.79±0.06 | 0.73±0.05 | 0.77 0.07 |
| Final weigth (g) | 3.09±0.27b | 4.18±0.53a | 3.87±0.23ab | 4.10±0.09a |
| SGR (%d-1) | 4.16±0.32 0.3271b | 4.75±0.25 25.2522 | 4.76±0.33 | 4.78±0.35 |
Table 3.
Alpha diversity indices in the treatment group and the control derived from MicrobiomeAnalyst. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. The mean ± SD are indicated. Different letters indicate significant differences.
Table 3.
Alpha diversity indices in the treatment group and the control derived from MicrobiomeAnalyst. Treatments: I) Control; II) Bacilli in the water; III) LAB in the water; IV) Bacilli + LAB in the water. The mean ± SD are indicated. Different letters indicate significant differences.
| Indices | I | II | III | IV |
|---|---|---|---|---|
| Shannon | 1.71±0.62 | 1.22±0.18 | 1.19±0.53 | 1.47±0.35 |
| Simpson | 0.60±0.16 | 0.53±0.13 | 0.40±0.21 | 0.47±0.13 |
| Chao1 | 149.74±27.71a | 93.95±25.25b 25.2522 | 137.03±10.27a | 137.10±14.54a |
| ACE | 146.34±27.87a 27.87.64 | 88.95±28.97b 2282828.9728.9705 | 132.22±15.70ab | 115.50±45.48ab |
Table 4.
The KEGG functional categories at level 1 (iVikodak) of microbiota found in the shrimp gut.
Table 4.
The KEGG functional categories at level 1 (iVikodak) of microbiota found in the shrimp gut.
| Treatment | Metabolism (%) | GIP (%) | HD (%) | EIP (%) | CP (%) | OS (%) |
|---|---|---|---|---|---|---|
| Control | 61.05±1.22 | 11.01±0.15 | 11.43±0.63 | 1.39±0.23 | 6.90±0.58 | 8.19±0.11 |
| Bacilli in water | 60.03±0.14 | 10.92±0.05 | 11.94±0.04 | 1.59±0.01 | 7.38±0.04 | 8.10±0.01 |
| LAB in water | 60.19±0.44 | 10.88±0.13 | 11.87±0.17 | 1.55±0.10 | 7.22±0.35 | 8.27±0.32 |
| Bacilli and LAB in water | 60.16±0.39 | 11.00±0.05 | 11.85±0.17 | 1.49±0.14 | 7.21±0.22 | 8.27±0.22 |
GIP = Genetic information processing; HD = Human diseases; EIP = Environmental information processing; CP = Cellular processes; OS = Organismal systems.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



