Submitted:
24 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
(1) Background: Impaired colonic motility is common in patients with ulcerative colitis. The patients often complain the symptoms related to colonic motility dysfunction such as loose stools or diarrhea and these symptoms make therapy more challenging; (2) Methods: This study aimed to examine the movement of the colon in mice with ulcerative colitis and determine if the glial-derived neurotrophic factor (GDNF) may enhance aberrant colonic motility through Connexin 43 (Cx43).A model of ulcerative colitis was created using 32 male C57BL/6 mice. Colonic transit time was measured in all mice, and colon tissues were examined using hematoxylin-eosin staining (HE) and transmission electron microscopy (TEM). The levels of GDNF and Cx43 were determined via immunohistochemistry, immunofluorescence, and RT-PCR techniques. The levels of C-reactive protein (CRP),TNF-α, IL-1β, and IL-6 were quantified using the enzyme-linked immunosorbent assay (ELISA); (3) Results: 1) The UC models were effectively established by administering dextran sodium sulfate (DSS) water. Mice with ulcerative colitis exhibited a delay in colonic transit time. 2) GDNF can improve the delayed movement of the colon in mice with UC by activating the Cx43 protein; (4) Conclusions: GDNF can ameliorate impaired colonic motility in mice with ulcerative colitis, and its mechanism is related to the activity of Cx43. GDNF could potentially be utilized as a therapeutic option for addressing aberrant colonic motility in patients.
Keywords:
GDNF
; ulcerative colitis
; colonic motility
; Cx43
1. Introduction
Ulcerative colitis (UC) is a chronic, frequently relapsing, and non-specific inflammatory disorder affecting the intestinal system that usually starts in the rectum [1]. The incidence rate of UC has dramatically increased in China over the last ten years, according to newly available clinical epidemiological data, as a result of dietary and lifestyle changes [2,3]. The clinical symptoms of UC are usually various, such as bloody diarrhea, weight loss, rectal pain, and abdominal cramps, which significantly lower the quality of life of patients and place a heavy socioeconomic load on healthcare systems [4,5,6,7]. For many years, it has been understood that individuals with UC experience colonic dysfunction [8,9,10]. Reduced post-prandial colonic contraction amplitude and varied transit have been observed in gastrointestinal motility assessments of UC patients [11]. According to reports, the entire gastrointestinal tract is covered by the enteric nervous system (ENS), which regulates bowel motility. [12,13]. The ENS is impaired in colonic tissue from UC patients [14,15,16]. People with ulcerative colitis (UC) often have diarrhea because their colons don't move properly and their intestinal epithelial barrier (IEB) isn't working right. These problems may affect each other. Changes in the ENS and subsequently colonic motility are caused by the accumulation of histological disturbance and gut wall structural damage. This leads to clinical symptoms such as loose stools or diarrhea, abdominal pain, or unpleasantness, and makes therapy more challenging [17,18,19]. However, the exact mechanisms of abnormal colonic motility in UC patients still remain unknown and confusing.
Gap junctions are cylindrical channels between animal cells that let ions and tiny molecules pass from the inside of one cell to the inside of the next [20]. Gap junctions regulate cell proliferation, migration, and differentiation [21]. They have channel clusters made of the connexin protein family, which create hemichannels and allow ions and signaling chemicals to be transferred between cells to facilitate intercellular communication [22,23]. By facilitating the transfer of electrical or second messenger impulses from one cell to another, they serve crucial roles in coordinating cellular function in numerous organs, including the heart, kidney, liver, and gastrointestinal system [24]. Humans have at least twenty-one distinct connexin subtypes, each of which can create heteromeric hemichannels with distinct connexin subtypes or homomeric channels with the same connexin subtype [25].
Connexin proteins are the basic units of hemichannels. Named for its molecular weight of 43 kDa, connexin 43 (Cx43) is the most prevalent and well-studied connexin [26,27]. Studies have shown that their key role is in regulating cell growth and apoptosis. Impaired motility, gastrointestinal infections, and IBD are linked to changes in Cx43 expression in the gastrointestinal tract [28]. Furthermore, studies have demonstrated a significant decrease in Cx43 expression in the colon of kids with Hirschsprung's illness. Such abnormalities cause disturbances in normal colonic motility [29]. That means Cx43 is involved in gastrointestinal motility regulation. The complex structure of the intestinal wall's enteric nervous system (ENS) organizes gastrointestinal functions [30]. Enteric glial cells (EGCs) and neurones are the two primary cell populations, the former being much more abundant than neurons. Previous studies had shown that abnormal EGCs were one of the important factors causing gastrointestinal motility disorders [31,32,33]. In the gastrointestinal tract, the Cx43 protein plays a crucial role in facilitating coordinated contraction of interstitial cells of Cajal and smooth muscle cells by forming gap junctions [34]. Initially, it was thought that EGCs were the primary source of GDNF secretion, and it has been found that intestinal tissue samples from UC patients had decreased levels of GDNF [35]. According to Gang Chen et al. [36], individuals with Parkinson's disease who experienced constipation also had reduced blood levels of GDNF, and low GDNF levels may be a risk factor for constipation in patients with Parkinson's disease. GDNF can trigger ENS regeneration in mouse models of Hirschsprung disease [37], and our previous animal investigations confirmed that aging-induced delayed stomach emptying might be linked to the inactivation of EGCs [32].GDNF plays a crucial role in the viability and maturation of the nervous system. Research has demonstrated that gap junction channels are controlled by a chemical gating mechanism that is responsive to the concentration of cytosolic Ca2+. The active EGCs have the ability to produce ATP, which plays a role in facilitating and transmitting the Ca2+ waves that spread from the EGCs to neighboring cells through gap junctions within the EGC networks [38].
Hence, the objectives of this investigation were to elucidate the alterations of GDNF and Cx43 in the colon tissues of mice with UC. Meanwhile, through Gap19, a selective inhibitor toward Cx43 hemichannels [39,40], we further investigate whether the effect of GDNF on impairment of colonic motility plays a role through Cx43. This evidence may contribute to the development of a possible novel drug or therapy for people with UC with impaired colonic motility.
2. Results
2.1. GDNF Alleviates Colitis Symptoms and Attenuates Serum Inflammatory Markers in DSS-induced UC Mice
DSS-induced colitis is characterized by pronounced diarrhea and the presence of severe bloody feces. The mice in this group showed significantly drier and duller hair over time. In comparison to the CN group, the mice had a significantly decreased appetite, body weight, and increased DAI score. The length of the colon in mice with UC caused by DSS was significantly shortened (Figure 1:a–c). The contents of inflammatory response indexes (CRP, TNF-α, IL-1β, and IL-6) were all significantly elevated in the mice with UC (Figure 2:a–d). In comparison to the UC group, GDNF reduced the symptoms (including more physical activity, reduced hair loss, and more alertness) and body weight loss, attenuated the increased DAI score, and shortened colon length during the progression of ulcerative colitis in mice in the UC+GDNF group. Moreover, TNF-α, IL-1β, and IL-6 levels, as well as CRP concentrations, were all reduced by GDNF. To further confirm whether the therapeutic effect of GDNF on UC was achieved by Cx43, the Cx43 inhibitor Gap19 was initially administered in mice to inhibit the pathway before the administration of GDNF. We discovered that the treatment effects of GDNF, which included the reduction of the DAI score and the inhibition of intestinal mucosal injury, were blocked. In the interim, no remarkable improvements in either the DAI or the symptoms and inflammatory response indexes were observed in UC+Gap19+GDNF group mice. The findings suggest that GDNF can mitigate the inflammatory reaction of colon tissue in UC mice through the Cx43 signaling pathway.
3.2. GDNF Improves Colonic Transit Time in DSS-Induced UC Mice through Cx43
To evaluate colonic transit time in all of the mice, we used the bead explusion test to determine colonic transit time. The colonic transit time in mice with UC was 13.13 ± 2.09 min, which was substantially delayed compared to that of the CN group (5.77 ± 0.58 min, P <0.05). However, the slowed colonic transit time in the UC group improved after the treatment of exogenous GDNF (7.84 ± 0.98 min). Interestingly, the differences in time to evacuate a glass bead from the rectums of UC and UC+Gap19+GDNF mice were not significant (13.13 ± 2.09 min versus 12.32 ± 1.35 min, P > 0.05). That means the therapeutic efficacy of GDNF in UC mice was inhibited after administration of Gap19. The effects of GDNF could restore colonic transit of UC mice by Cx43 (Figure 3).
2.3. GDNF Improves Colonic Histopathological Changes in DSS-Induced UC Mice through Cx43
Histopathological examinations were conducted to observe the pathology alterations in the intestinal tissue of each group. The colon tissue in the CN group had typical crypt architecture, the presence of goblet cells, and intact colonic epithelial cells, without ulceration or thickening of the mucosa. Conversely, the colons of the UC group showed impaired integrity of the colonic mucosa and significant pathological alterations such as multiple superficial ulcers, degeneration of mucosal epithelial cells, loss of goblet cells, necrosis, epithelial shedding, cryptitis, crypt abscesses, and extensive inflammatory cell infiltration, which led to symptoms of UC, such as abdominal cramping, rectal pain, weight loss, and bloody diarrhea. The pathological alterations seen in the intestinal tissue of the UC+Gap19+GDNF group are comparable to those seen in the UC group. However, the colons of the UC+GDNF group revealed less structural damage and less inflammatory cell infiltration (Figure 1d), indicating that GDNF prevents histopathological change in UC mice induced by DSS through Cx43.
3.4. GDNF and Cx43 Expression in the Colon Are Decreased in Mice with DSS-Induced UC
The primary gap junction protein, Cx43, was analyzed using quantitative Real-Time PCR in mice from various groups. Significantly, the mRNA levels of GDNF and Cx43 in the colon exhibited a distinct reduction in UC mice. Interestingly, in comparison to the UC group mice, the mRNA expression of Cx43 was significantly elevated in the GDNF+UC group (Figure 4). Further, to clarify the localization of Cx43 and GDNF and investigate whether impaired colonic motility in UC group mice is related to the defect of Cx43 protein, immunohistochemistry and immunofluorescent staining techniques were used in our study. We conducted double-labelling immunofluorescence with Cx43 and GDNF. Interestingly, immunohistochemistry revealed that Cx43 and GDNF were not only localized in the smooth muscle portion of the intestine but also highly concentrated in colonic epithelial cells. The Cx43 immunopositive product (green) was shown as plaques, and Cx43-immunoreactive plaques colocalized with the GDNF immunopositive product (red) and were distributed across colon tissue specimens. Cx43-puncta per field in the UC group was markedly sparse and disarrayed compared with the CN group; however, it was partially attenuated by exogenous GDNF treatment. In addition, the expression of Cx43 was significantly decreased after the administration of Gap19. The treatment effects of exogenous GDNF disappeared after Cx43 was blocked by Gap19 (Figure 5 and Figure 6).
2.5. Ultrastructure of Colon Tissue in DSS-Induced UC Mice
In order to more accurately evaluate the morphological alterations of colon tissue sections from each group. Electron microscopy revealed that the ultrastructure of colon tissue in the CN group exhibited normal morphology (such as absorptive columnar cells with clearly identified microvilli, regular oval euchromatic nuclei, numerous microvilli, and plentiful mitochondria). Mice that received DSS treatment exhibited a typical scorch phase, which was characterized by the presence of unusually expanded cells, rupture of the cell membranes, uneven microvilli, and shriveled mucin granules. The collagen fibers were looser, and the arrangement was irregular. Damaged cells appeared with dark, small nuclei, degenerated organelles, and rarified cytoplasm. The shape of the nucleus was irregular and edematous. In the DSS+GDNF group, although some cells displayed pyroptosis morphology, the overall number of damaged cells was lower. Additionally, there were fewer irregular microvilli, and the diameters of the injured cells were smaller compared to the DSS group. In addition, the microvilli of colonic epithelial cells in GDNF-treated mice arranged neatly without shedding, suggesting that GDNF mitigated the impact of DSS-induced pyroptosis. However, in the DSS+Gap19+GDNF group mice, the ultrastructure of colon tissues in mice exhibited similarities to those in the DSS group. The therapeutic effect of GDNF on UC mice was inhibited (Figure 7).
3. Discussion
The model of UC in mice was successfully produced by administering DSS aqueous solution in this current study. The model is widely regarded for its ability to accurately replicate human clinical symptoms of ulcerative colitis, such as weight loss and bloody diarrhea [41,42]. It was found in the experiments that colonic transit time in UC mice was significantly delayed than that in normal mice. Moreover, the pathological damage appeared in the colorectal tissue of UC mice. These indicators of the UC model mice confirmed that the UC mice mode was successfully established. In addition, our study revealed the beneficial impacts of GDNF on experimental UC in mice. The treatment with GDNF alleviated the morphological changes of the colon, improved the DAI scores, ameliorated delayed colonic transit time, and reduced the weight loss associated with colitis. Therefore, these data indicate that GDNF could be a promising target for treating the aberrant movement of the colon observed in mice with colitis. Nevertheless, the therapeutic impact of GDNF in UC mice was impeded by Gap19, a specific inhibitor of Cx43 hemichannels.
In the UC mice model, CRP, TNF-α, IL-1β, and IL-6 have been recognized as important proinflammatory cytokines in the development of UC in mice. The DAI score is commonly employed to indicate the level of inflammatory activity in animal experiments. The levels of CRP, TNF-α, IL-1β, and IL-6 in the blood were quantified using the ELISA. As anticipated, there was a notable rise in the levels of CRP, TNF-α, IL-1β, and IL-6, as measured by ELISA, in the mice with UC (P<0.05). The DAI score showed a considerable increase in the UC group. A study demonstrated that the rats with UC exhibited an increased level of P2X7R expression in the tissues of their colon [43]. Prior research has demonstrated the involvement of Cx43 in the extracellular release of ATP. This ATP, in turn, plays a role in excitatory neurotransmission inside the ENS through the activation of P2X and P2Y receptors [44,45]. ATP-activated receptors have a well-established role in the inflammatory response since they have been demonstrated to facilitate the release of proinflammatory cytokines from immune cells [46]. Furthermore, our research findings revealed that GDNF effectively inhibited the excessive release of these proinflammatory cytokines in mice with UC, indicating that GDNF reduced aberrant inflammatory responses.
Increasing data indicates that individuals with ulcerative colitis experience a considerable delay in colonic transit time and a notable decrease in the contractile force of the smooth muscle, despite an increased frequency of bowel movements. The symptoms of diarrhea in people with ulcerative colitis (UC) may not be attributed to an increased speed of movement in the colon but rather to a hypersensitive and inflamed distal colon and rectum that is not compliant, leading to frequent and urgent defecation [47]. Similar research results were observed in our study. However, the exact signaling mechanism remains unclear [48,49,50]. Impaired intestinal motility can exacerbate diarrhea in people with ulcerative colitis, lead to symptoms that are indistinguishable from those directly associated with the inflammatory process, and have an adverse effect on quality of life.
Connexin proteins are the basic unit of hemichannels, and Cx43 is the primary gap junction protein in astrocytes [51]. Nevertheless, the morphology and function of fully developed EGC closely resemble those of astrocytes in the CNS. EGCs, which are the primary constituents of the ENS, have been shown to possess a highly intricate and sophisticated nature in their involvement in the regulation of gastrointestinal tract movement and the preservation of intestinal balance [52]. Neural crest cells, which originate from the ectoderm, are widely recognized as multi-potent cells that migrate to several organs, including the gastrointestinal tract, where they contribute to the formation of the ENS. Peter Bannerman and his colleagues discovered that gap junctions are essential for the survival of neural crest cells. They also found that migrating rat neural crest cells express the gap junction component Cx43 [53]. By using of messenger molecules carried by Cx43, the interstitial cells of Cajal found in the deep muscular plexus area of the rat small intestine promote communication between themselves and smooth muscle cells [54]. Recent studies have demonstrated that the expression of Cx43 is modified in the colon of children diagnosed with Hirschsprung's disease. Consequently, this anomaly leads to improper communication between cells and a disruption in the movement of the colon [55].Based on these studies, we hypothesized that Cx43 is involved in gastrointestinal motility regulation. Cx43 is the primary gap junction protein involved in the development of the gastrointestinal motility transmission system. Phosphorylation is the main form in which Cx43 exerts biological effects in electrical coupling in the gastrointestinal tract [56]. However, there is limited knowledge regarding the precise mechanism of phosphorylation. In our current study, we discovered a decrease in the expression of Cx43 in the colon tissues of mice with UC. This suggests that Cx43 may have a protective role in the development of UC.
Our study confirmed that GDNF decreased the concentrations of CRP, TNF-α, IL-1β, and IL-6 and relieved the morphological changes in UC mice. GDNF, a neurotrophic factor, is mainly produced by EGCs in the gastrointestinal tract [57]. Our findings suggest that GDNF upregulates Cx43 expression in colonic tissue, including EGCs. GDNF can protect EGCs from apoptosis, and in mice deficient in GDNF, ENS entirely fails to mature [58,59]. In DSS-induced rat UC, Li et al. showed that GDNF significantly decreased colonic inflammation, partially reversed the loss of enteric neurons, and enhanced delayed intestinal transit [60]. The enhanced function of the intestinal epithelial barrier may have contributed to the anti-inflammatory impact of GDNF in our investigation. The underlying mechanisms are still unclear, though.
Cx43 is crucial in the transmission of regular action potentials. Reducing the expression of Cx43 can diminish the intercellular communication between myocytes [61]. We further investigated the relationship between GDNF and Cx43 and the effect of specific inhibition of Cx43 hemichannels named Gap19 on experimental UC in mice, an important mechanism about impaired colonic motility in UC. Previous research has demonstrated that Gap19 effectively and selectively inhibits the opening of Cx43 hemichannels in astrocytes [62]. Our analysis verified that the level of Cx43 expression was dramatically reduced. We hypothesized that the decrease in the expression of Cx43 would weaken the ability of EGCs to communicate and transmit signals, resulting in aberrant colonic motility. What's more, the therapeutic effect of GDNF on UC mice was inhibited after administration of Gap19. The presence of Cx43 hemichannels is necessary for the modulation of colonic motility in the ENS. Cx43 serves as the primary gap protein that facilitates the connection and transmission of signals across gastrointestinal neurons, smooth muscle networks, and the interstitial cells of Cajal in the ENS [63].Tingyi Sun et al, confirmed that the expression of Cx43 in the gastrointestinal tract of mice decreases with age, and this decline occurs prior to the loss of interstitial cells of Cajal and neurons. The results of the research team's study suggest that Cx43 might be the upstream regulation target for the gastrointestinal motility disorder sufferers due to the growth of age [63]. Furthermore, there were studies indicating that the interstitial cells of Cajal in the deep muscular plexus region of the rat small intestine establish communication among themselves and with smooth muscle cells by means of messenger molecules passing through Cx43 [64]. Our findings indicated that the expression quantity of Cx43 was decreased in UC mice.
The research from McClain J. et al. showed that Cx43 is distributed throughout the EGCs of the colon myenteric nerve plexus [65]. However, we found that Cx43 is distributed not only in the EGCs in the myenteric nerve plexus of the mouse colon but aslo enriched in epithelial cells, circular muscle layer, and muscularis mucosa. It was clearly confirmed that disturbance of Ca2+ channel function has been identified as a contributing factor to decreased movement in the colon [66]. McClain J. et al. also revealed that Ca2+ responses in EGCs are facilitated by Cx43 hemichannels. The absence of Cx43, particularly its channel activity, reduces the spread of intercellular Ca2+ waves (ICWs) [67]. Meanwhile, Cx43 has previously been shown to be involved in extracellular release of ATP; the activation of EGCs were accompanied by the release of ATP via Cx43 [44,68,69], following the administration of Cx43 inhibitor in mice, The capacity of Ca2+ to undergo reactive diffusion in EGCs is restricted, and Cx43 has a direct role in mediating Ca2+ responses and an indirect role in contributing to gastrointestinal transit [65]. In future work, the upstream regulatory mechanism of GDNF will be further investigated to explore more information on the pathological process of UC and promising candidate for future medication development.
4. Materials and Methods
4.1. Animals
We used thirty-two male C57BL/6-specific pathogen-free (SPF) mice (25 g) aged six to eight weeks in this experiment. These mice were acquired from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijiing, China). Prior to any surgical or experimental operations, all mice were provided with adapted feeding for a duration of 5 days while being kept in SPF circumstances. Subsequently, they were placed in an identical setting with a temperature range of 22–23 °C, humidity levels of 40–50%, and a light-dark cycle of 12 hours each. Cages were thoroughly cleaned, and mice were monitored every day. Prior to the current study, we obtained authorization from the Institutional Animal Care and Use Committee for all of our experimental procedures (approval no. 2022175). We also ensured the minimum suffering of the mice as much as possible during the whole experimental procedure.
4.2. Induction of Ulcerative Colitis
The mice were randomly allocated into following four groups: normal control group (CN), UC model group (UC), UC+GDNF treatment group (UC+GDNF), and UC+Gap19+GDNF treatment group (UC+Gap19+GDNF), with 8 mice in each group. The UC model was constructed according to the description provided in the literature [70]. The dextran sodium sulfate (DSS) was dissolved in distilled water to create a 3.5% DSS solution, and the mice of the UC model group received 3.5% DSS dissolved in drinking water for 10 days to induce UC, and the CN group received drinking the same amount of water alone. In the UC+GDNF group, after administering DSS aqueous solution, the mice were treated with 5μL/kg body weight of GDNF in 100μl 0.9% sodium chloride intraperitoneally, and in the UC+Gap19+GDNF group, mice were injected with Gap19 intraperitoneally at a dose of 25mg/kg body weight after the administration of GDNF. Every day, we measured body weight, diarrhea, and rectal bleeding in each group.
4.3. Surgical Procedure
Following an overnight fast, all mice were administered pentobarbital sodium
(75 mg/kg) intraperitoneally for euthanasia and sacrificed on day 11. The animals were handled and sacrificed in a humane manner. The mice were monitored every day using the disease activity index (DAI). The colon lengths were measured, and the tissues from the colon were divided and kept in liquid nitrogen in preparation for further tests. The DAI was determined as follow Table1 [71]. Higher values of DAI reflect the increasing severity of UC.
4.4. Analysis of Colonic Transit Time
The bead expulsion test was used to quantify colonic transit time, as previously reported [72,73]. In our investigation, we slightly altered this approach. An overnight fasting mouse's colon (3 cm from the anus) was punctured with a single 2-mm glass bead using a plastic rod that had been gently greased with lubricating jelly. The mice were then put in their own cages once the bead was inserted. Colonic transit time was evaluated by measuring the time between bead placement and bead expulsion.
4.5. Histopathological Examinations
The colon samples were removed from the mice on day eleven, preserved for an overnight period in 10% neutral-buffered formalin, and then sliced into thin strips for conventional processing and paraffin embedding. The colon samples were sectioned into 4μm-thick sections and stained with picrosirius red staining, periodic acid-Schiff (PAS), and hematoxylin and eosin (HE). The stained sections were examined by two of the investigators and photographed using a microscope for pathological changes (Eclipse E100 Nikon, Japan).
4.6. Transmission Electron Microscopy
The colon tissues of each group of mice were collected and preserved in 2.0% glutaraldehyde. They were then washed with 0.1 M phosphate buffer and subsequently fixed with 1% osmic acid. Following a sequence of dehydration, embedding, sectioning, polymerization, and staining with uranium acetate, transmission electron microscopy (TEM) was conducted, as outlined in prior research [74]. The ultrastructure of the colon tissues was evaluated with TEM (Hitachi HT7700, Japan). Each sample was seen by three specialists separately.
4.7. Immunohistochemistry Staining
The colonic tissues were dissected, separated, and rinsed with cold PBS. Subsequently, the samples were treated with a 4% paraformaldehyde solution to stabilize them. The expression of GDNF and Cx43 was analyzed in 4μm-thickness sections of paraffin-embedded colonic tissue. We followed the manufacturer's protocol, using anti-GDNF antibody (dilution 1:200, Servicebio, Wuhan, China) and anti-Cx43 antibody (dilution 1:200, Servicebio, Wuhan, China) at a temperature of 4 °C overnight. Thereafter, the slides were immersed in PBS and then exposed to a secondary antibody (goat anti-rabbit IgG) for a duration of 50 minutes at room temperature. The sections were washed three times with PBS for a duration of 5 minutes each time. Finally, slides were visualized with diaminobenzidine (DAB) and counterstained with hematoxylin for microscopic examination (Nikon E100, Japan). We used pathological image analysis software (Image Pro Plus 6.0) to quantify the immunohistochemistry staining results.
4.8. Immunofluorescent studies of Cx43 and GDNF
Astrocytes in the central nervous system (CNS) mostly express Cx43 hemichannels [75,76], while the morphology of mature EGCs in the gastrointestinal tract is similar to that of astrocytes in the CNS. Immunofluorescence staining was conducted to examine the precise location and distribution of Cx43 and GDNF in the colon tissues. After dewaxing, the antigen was restored using Tris-EDTA buffer (pH 8.0) and then treated with 3% BSA for 30 minutes to prevent non-specific binding. Then, the colon tissue sections were exposed to a mixture of primary antibodies: mouse anti-Cx43 (dilution 1:200, Servicebio, Wuhan, China) and rabbit anti-GDNF (dilution 1:200, Servicebio, Wuhan, China) antibodies. This was followed by incubation with corresponding secondary antibodies: Alexa Fluor-488-labeled goat anti-mouse IgG (dilution 1:200, Servicebio, Wuhan, China) and CY3-labeled goat anti-rabbit IgG (dilution 1:200, Servicebio, Wuhan, China) for 50 minutes at room temperature. The sections were stained with 4’,6-diamidino-2-phenylindole (DAPI) and then sealed using an anti-fluorescence quenching sealing solvent. The specimens were analyzed using a fluorescent microscope (Nikon Eclipse C1, Japan).
4.9. Relative quantitative Real-Time PCR Analysis
The relative expression levels of GDNF and Cx43 in colon samples at the mRNA level were measured using real-time polymerase chain reaction (RT-PCR). Colon samples were subjected to RNA extraction using Trizol reagent (Invi-Trogen, USA) following the manufacturer's procedure. The set of primers for mouse GDNF, Cx43, and GAPDH were prepared as shown in Table 2. The ABI-StepOne Real-Time System (Applied Biosystems, USA) was used for all reactions. The PCR was carried out under the following cycling conditions: incubation at 25°C for 5 minutes, followed by incubation at 42°C for 60 minutes, and finally incubation at 70°C for 5 minutes. The dissociation curve analysis provided confirmation of the amplification of products unique to the primers used. The 2-△△Ct technique was employed to calculate the relative change in gene expression [77].
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
The serum was acquired from whole blood samples using the process of centrifugation at a temperature of 4 °C, with a speed of 3000 revolutions per minute for a duration of 15 minutes. The serum was collected and kept at a temperature of -80 °C. The levels of c-reactive protein (CRP), tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) in the blood were determined using ELISA kits specific to each sample, following the instructions provided by the manufacturer. The optical density (OD) was quantified at a wavelength of 450 nm using a microplate reader (BioTeK, Winooski, USA), and the concentration was determined using a standard curve.
5. Conclusions
In summary, the data demonstrate a novel function of GDNF in controlling the movement of the colon in mice with UC. Mechanistically, we believed that GDNF improved impaired colonic motility in mice with UC by Cx43. Our ongoing findings suggest that GDNF has the potential to offer therapeutic advantages that could improve abnormal colonic motility in UC patients, and Cx43 represents a new potential target for UC therapy. Understanding the cell and molecular drivers of these processes is critical to investigating new therapeutic approaches to treat patients with UC. There are many gaps in our knowledge of the pathogenesis of UC and many opportunities for future research before we develop a more complete understanding of cells and molecules.
Author Contributions
Conceptualization, F.Xu.; investigation, R.Liu and W.Yang.; resources, W.Yang. and F.Xu.; writing—original draft preparation, W.Yang.; writing—review and editing, W.Yang. and F.Xu., supervision, R.Liu. All authors have read and agreed to the published final version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The present study was approved by the Animal Care and Use Committee of Wuhan Servicebio Technology Co., Ltd. (approval no. 2022175) .
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the conclusions of this investigation are accessible upon request from the corresponding author.
Acknowledgments
My deepest gratitude goes first and foremost to Professor F.Xu, my supervisor, for his constant encouragement and guidance. Second, I would like to thank Dr.R.Liu for her technical assistance during the experiment. Last but not least, I would also like to thank my families for their constant support and encouragement throughout this research.
Conflicts of Interest
No potential conflict of interest was reported by the authors..
References
- Gros,B.; Kaplan,G.G. Ulcerative Colitis in Adults: A Review. JAMA. 2023;330(10):951-965.
- Cui,G.; Li,J.; Liu,H.; Laugsand, J.B.; Liu,Z. Differences in inflammatory bowel diseases between East and West: a Chinese perspective. J Public Health (Berl.).2021; 29:19–26. [CrossRef]
- Ng, S. C.; Shi, H. Y.; Hamidi, N.; Underwood, F. E.; Tang, W.; Benchimol, E. I.; Panaccione, R.; Ghosh, S.; Wu, J. C. Y.; Chan, F. K. L.; Sung, J. J. Y.; Kaplan, G. G. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies [published correction appears in Lancet. 2020 Oct 3;396(10256):e56]. Lancet.2017;390(10114):2769-2778.
- Walsh, A.J; Bryant, R.V; Travis, S.P. Current best practice for disease activity assessment in IBD. Nat Rev Gastroenterol Hepatol. 2016;13(10):567-579. [CrossRef]
- Khan, S.; Sebastian, S. A.; Parmar, M. P.; Ghadge, N.; Padda, I.; Keshta, A. S.; Minhaz, N.; Patel, A. Factors influencing the quality of life in inflammatory bowel disease: A comprehensive review. Dis Mon. 2024;70(1S):101672. [CrossRef]
- Kaplan, G.G.; Ng,S.C. Understanding and Preventing the Global Increase of Inflammatory Bowel Disease. Gastroenterology. 2017;152:313-321 e2. [CrossRef]
- Salem, M. B.; El-Lakkany, N. M.; Seif El-Din, S. H.; Hammam, O. A.; Samir, S. Diosmin alleviates ulcerative colitis in mice by increasing Akkermansiamuciniphila abundance, improving intestinal barrier function, and modulating the NF-κB and Nrf2 pathways. Heliyon. 2024;10(6):e27527.
- Kishi,K.; Kaji,N.; Tsuru,Y.; Hori,M. A Novel Noninvasive Method for Quantitative Detection of Colonic Dysmotility Using Real-Time Ultrasonography. Digestion. 2021;102(5):731-741.
- da Silva Watanabe, P.; Cavichioli, A. M.; D'Arc de Lima Mendes, J.; Aktar, R.; Peiris, M.; Blackshaw, L. A.; de Almeida Araújo, E. J. Colonic motility adjustments in acute and chronic DSS-induced colitis. Life Sci. 2023;321:121642.
- KERN, F.; Jr, ALMY, T. P.; ABBOT, F. K.; BOGDONOFF, M. D. The motility of the distal colon in nonspecific ulcerative colitis. Gastroenterology. 1951;19:492–503. [CrossRef]
- Reddy, S. N.; Bazzocchi, G.; Chan, S.; Akashi, K.; Villanueva-Meyer, J.; Yanni, G.; Mena, I.; Snape, W. J. JrColonic motilityand transit in health and ulcerative colitis. Gastroenterology. 1991; 101: 1289–97.
- Gershon, M.D.; Margolis, K.G. The gut, its microbiome, and the brain: connections and communications. J Clin Invest. 2021;131(18):e143768. [CrossRef]
- Choi, E.L.; Taheri, N.; Zhang,Y.; Matsumoto, K.; Hayashi Y. The critical role of muscularis macrophages in modulating the enteric nervous system function and gastrointestinal motility. J Smooth Muscle Res. 2024;60:1-9.
- Li, H.; Fan, C.; Lu, H.; Feng, C.; He, P.; Yang, X.; Xiang, C.; Zuo, J.; Tang, W.Protective role of berberine on ulcerative colitis through modulating enteric glial cells-intestinal epithelial cells-immune cells interactions. Acta Pharm Sin B. 2020;10(3):447-461. [CrossRef]
- Tavakoli, P.; Vollmer-Conna, U.; Hadzi-Pavlovic, D.; Grimm, M.C. A Review of Inflammatory Bowel Disease: A Model of Microbial, Immune and Neuropsychological Integration. Public Health Rev. 2021;42:1603990.
- Le Berre, C.; Naveilhan, P.; Rolli-Derkinderen, M. Enteric glia at center stage of inflammatory bowel disease. Neurosci Lett. 2023;809:137315. [CrossRef]
- Xiao, H.; Li, H.; Wen, Y.; Jiang, D.; Zhu, S.; He, X.; Xiong, Q.; Gao, J.; Hou, S.; Huang, S.; He, L.; Liang, J. Tremella fuciformis polysaccharides ameliorated ulcerative colitis via inhibiting inflammation and enhancing intestinal epithelial barrier function. Int J Biol Macromol. 2021;180:633-642. [CrossRef]
- Spalinger, M. R.; Sayoc-Becerra, A.; Ordookhanian, C.; Canale, V.; Santos, A. N.; King, S. J.; Krishnan, M.; Nair, M. G.; Scharl, M.; McCole, D. F. The JAK Inhibitor Tofacitinib Rescues Intestinal Barrier Defects Caused by Disrupted Epithelial-macrophage Interactions. J Crohns Colitis. 2021;15(3):471-484. [CrossRef]
- Schlegel, N.; Boerner, K.; Waschke, J. Targeting desmosomal adhesion and signalling for intestinal barrier stabilization in inflammatory bowel diseases-Lessons from experimental models and patients. Acta Physiol (Oxf). 2021;231(1):e13492.
- Peracchia, C. Gap Junction Channelopathies and Calmodulinopathies. Do Disease-Causing Calmodulin Mutants Affect Direct Cell-Cell Communication?. Int J Mol Sci. 2021;22(17):9169.
- Zuo, T.; Liu, Y.; Duan, M.; Pu, X.; Huang, M.; Zhang, D.; Xie, J. Platelet-derived growth factor PDGF-AA upregulates connexin 43 expression and promotes gap junction formations in osteoblast cells through p-Akt signaling. BiochemBiophys Rep. 2023;34:10146.
- Totland, M.Z.; Rasmussen, N.L.; Knudsen, L.M.; Leithe E. Regulation of gap junction intercellular communication by connexin ubiquitination: physiological and pathophysiological implications. Cell Mol Life Sci. 2020;77(4):573-591.
- Schumacher, S.; Tahiri, H.; Ezan, P.; Rouach, N.; Witschas, K.; Leybaert L. Inhibiting astrocyte connexin-43 hemichannels blocks radiation-induced vesicular VEGF-A release and blood-brain barrier dysfunction. Glia. 2024;72(1):34-50.
- Katturajan, R.; Evan Prince, S. A role of connexin 43 on the drug-induced liver, kidney, and gastrointestinal tract toxicity with associated signaling pathways. Life Sci. 2021;280:119629.
- Meşe, G.; Richard, G.; White, T.W. Gap junctions: basic structure and function. J Invest Dermatol. 2007;127(11):2516-2524.
- Jindal, S.; Chockalingam, S.; Ghosh, S.S.; Packirisamy, G. Connexin and gap junctions: perspectives from biology to nanotechnology based therapeutics. Transl Res. 2021;235:144-167.
- Goodenough, D.A.; Paul, D.L.Gapjunctions.Cold Spring HarbPerspect Biol. 2009;1(1):2576.
- Wong, J.; Chopra, J.; Chiang, L. L. W.; Liu, T.; Ho, J.; Wu, W. K. K.; Tse, G.; Wong, S. H. The Role of Connexins in GastrointestinalDiseases. J Mol Biol. 2019; 431: 643-652.
- Coyle. D.; Doyle, B.; Murphy, J.M.; O'Donnell, A.M.; Gillick J, Puri.P. Expression of connexin 26 and connexin 43 is reduced in Hirschsprung's disease. J Surg Res. 2016;206(1):242-251.
- DiCello, J. J.; Carbone, S. E.; Saito, A.; Pham, V.; Szymaszkiewicz, A.; Gondin, A. B.; Alvi, S.; Marique, K.; Shenoy, P.; Veldhuis, N. A.; Fichna, J.; Canals, M.; Christopoulos, A.; Valant, C.; & Poole, D. P. Positive allosteric modulation of endogenous delta opioid receptor signaling in the enteric nervous system is a potential treatment for gastrointestinal motility disorders. Am J PhysiolGastrointest Liver Physiol. 2022;322(1):G66-G78. [CrossRef]
- Broadhead, M. J.; Bayguinov, P. O.; Okamoto, T.; Heredia, D. J.; & Smith, T. K. Ca2+ transients in myenteric glial cells during the colonic migrating motor complex in the isolated murine large intestine. J Physiol. 2012 Jan 15;590(2): 335-350. [CrossRef]
- Yang, W.; Wang, N.; Shi, X.; Chen, J. Synchronized dual pulse gastric electrical stimulation induces activation of enteric glial cells in rats with diabetic gastroparesis. Gastroenterol Res Pract. 2014;2014:964071. [CrossRef]
- Howlett, P.J.; Ward, A.S.; Duthie, H.L. Gastric emptying after vagotomy. Proc RSoc Med. 1974;67: 836-838.
- Maes, M.; Cogliati, B.; Crespo, Yanguas S.; Willebrords, J.; Vinken, M. Roles of connexins and pannexins in digestive homeostasis. Cell Mol Life Sci. 2015;72:2809–21.
- Meir, M.; Burkard, N.; Ungewiß, H.; Diefenbacher, M.; Flemming, S.; Kannapin, F.; Germer, C. T.; Schweinlin, M.; Metzger, M.; Waschke, J.; Schlegel, N. Neurotrophic factor GDNF regulates intestinal barrier function in inflammatory bowel disease. J Clin Invest. 2019;129(7):2824-2840. [CrossRef]
- Chen, G.; Du, Y.; Li, X.; Kambey, P. A.; Wang, L.; Xia, Y.; Tang, C.; Shi, M., Zai-Li, L.; Zai-E, X.; Xiao-Ling, Q.; Dian-Shuai, G. Lower GDNF Serum Level Is a Possible Risk Factor for Constipation in Patients With Parkinson Disease: A Case-Control Study. Front Neurol. 2022;12:777591. [CrossRef]
- Soret, R.; Schneider, S.; Bernas, G.; Christophers, B.; Souchkova, O.; Charrier, B.; Righini-Grunder, F.; Aspirot, A.; Landry, M.; Kembel, S. W.; Faure, C.; Heuckeroth, R. O.; Pilon, N. Glial Cell-Derived Neurotrophic Factor Induces Enteric Neurogenesis and Improves Colon Structure and Function in Mouse Models of Hirschsprung Disease. Gastroenterology. 2020;159(5):1824-1838.e17. [CrossRef]
- Weiss, S.; Clamon, L.C.; Manoim, J.E.; Ormerod, K.G.; Parnas, M.; Littleton, J.T. Glial ER and GAP junction mediated Ca2+ waves are crucial to maintain normal brain excitability. Glia. 2022;70(1):123-144. [CrossRef]
- Ponsaerts, R.; Wang, N.; Himpens, B.; Leybaert, L.; Bultynck, G. The contractile system as a negative regulator of the connexin 43 hemichannel. Biol Cell. 2012;104(7):367-377.
- Abudara, V.; Bechberger, J.; Freitas-Andrade, M.; De Bock, M.; Wang, N.; Bultynck, G.; Naus, C. C.; Leybaert, L.; Giaume, C. The connexin43 mimetic peptide Gap19 inhibits hemichannels without altering gap junctional communication in astrocytes. Front Cell Neurosci. 2014;8:306. [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar M. Dextran sulfate sodium (DSS)-induced colitis in mice. CurrProtoc Immunol. 2014;104:15.25.1-15.25.14. [CrossRef]
- Fan, X.; Ding, X.; Zhang, Q.Y. Hepatic and intestinal biotransformation gene expression and drug disposition in a dextran sulfate sodium-induced colitis mouse model. Acta Pharm Sin B. 2020;10(1):123-135. [CrossRef]
- Saber, S.; Youssef, M. E.; Sharaf, H.; Amin, N. A.; El-Shedody, R.; Aboutouk, F. H.; El-Galeel, Y. A.; El-Hefnawy, A.; Shabaka, D.; Khalifa, A.; Saleh, R. A.; Osama, D.; El-Zoghby, G.; Gobba, N. A. BBG enhances OLT1177-induced NLRP3 inflammasome inactivation by targeting P2X7R/NLRP3 and MyD88/NF-κB signaling in DSS-induced colitis in rats. Life Sci. 2021;270:119123.
- Brown, I.A.; McClain, J.L.; Watson, R.E.; Patel, B.A.; Gulbransen, B.D. Enteric glia mediate neuron death in colitis through purinergic pathways that require connexin-43 and nitric oxide. Cell Mol Gastroenterol Hepatol. 2016;2(1):77-91. [CrossRef]
- Burnstock, G.; Williams, M. P2 purinergic receptors: modulation of cell function and therapeutic potential. J Pharmacol Exp Ther. 2000;295(3):862-869.
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. Nature. 2014;509(7500):310-317. [CrossRef]
- Drewes, A.M.; Frøkjaer, J.B.; Larsen, E.; Reddy, H.; Arendt-Nielsen, L.; Gregersen, H. Pain and mechanical properties of the rectum in patients with active ulcerative colitis. Inflamm Bowel Dis. 2006;12(4):294-303. [CrossRef]
- Fornai, M.; Colucci, R.; Antonioli, L.; Ghisu, N.; Tuccori, M.; Gori, G.; Blandizzi, C.; Del Tacca, M. Effects of a bicarbonate-alkaline mineral water on digestive motility in experimental models of functional and inflammatory gastrointestinal disorders. Methods Find Exp Clin Pharmacol. 2008;30(4):261-269. [CrossRef]
- Reddy, S. N.; Bazzocchi, G.; Chan, S.; Akashi, K.; Villanueva-Meyer, J.; Yanni, G.; Mena, I.; Snape, W. J. Colonic motility and transit in health and ulcerative colitis. Gastroenterology. 1991;101(5):1289-1297. [CrossRef]
- Haase, A. M.; Gregersen, T.; Christensen, L. A.; Agnholt, J.; Dahlerup, J. F.; Schlageter, V.; Krogh, K. Regional gastrointestinal transit times in severe ulcerative colitis. NeurogastroenterolMotil. 2016;28(2):217-224. [CrossRef]
- Schumacher, S.; Tahiri, H.; Ezan, P.; Rouach, N.; Witschas, K.; Leybaert L. Inhibiting astrocyte connexin-43 hemichannels blocks radiation-induced vesicular VEGF-A release and blood-brain barrier dysfunction. Glia. 2024;72(1):34-50.
- Montalbán-Rodríguez, A.; Abalo, R.; López-Gómez, L. From the Gut to the Brain: The Role of Enteric Glial Cells and Their Involvement in the Pathogenesis of Parkinson's Disease. Int J Mol Sci. 2024;25(2):1294.
- Bannerman, P.; Nichols, W.; Puhalla, S.; Oliver, T.; Berman, M.; Pleasure, D. Early migratory rat neural crest cells express functional gap junctions: Evidence that neural crest cell survival requires gap junction function. Journal of Neuroscience Research. 2000, 61(6):605-615.
- Seki, K.; KomuroT. Immunocytochemical demonstration of the gap junction proteins connexin 43 and connexin 45 in the musculature of the rat small intestine. Cell Tissue Res. 2001 Dec; 306(3):417-22.
- Coyle, D.; Doyle, B.; Murphy, J. M.; O'Donnell, A. M.; Gillick, J.; Puri, P.Expression of connexin 26 and connexin 43 is reduced in Hirschsprung's disease. J Surg Res. 2016 Nov; 206(1):242-251.
- Katturajan, R.; Evan Prince, S. A role of connexin 43 on the drug-induced liver, kidney, and gastrointestinal tract toxicity with associated signaling pathways. Life Sci. 2021;280:119629.
- Bar, K.J.; Facer, P.A.; Williams, N.S.; Tam, P.K.; Anand, P.R. Glial-derived neurotrophic factor in human adult and fetal intestine and in Hirschsprung's disease. Gastroenterology 4: 1381-1385, 1997.
- Sánchez, M.P.; Silos-Santiago, I.; Frisén, J.; He, B.; Lira SA.; Barbacid M. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 1996;382(6586):70-73. [CrossRef]
- Steinkamp, M.; Gundel, H.; Schulte, N.; Spaniol, U.; Pflueger, C.; Zizer, E.; von Boyen, G. B. GDNF protects enteric glia from apoptosis: evidence for an autocrine loop. BMC Gastroenterol. 2012;12:6. [CrossRef]
- Li, H.;Fan, C.; Lu, H.; Feng, C.; He, P.; Yang, X.; Xiang, C.; Zuo, J.; Tang, W.Protective role of berberine on ulcerative colitis through modulating enteric glial cells-intestinal epithelial cells-immune cells interactions. Acta Pharm Sin B. 2020;10(3):447-461. [CrossRef]
- Delmar, M.; Makita, N. Cardiac connexins, mutations and arrhythmias. CurrOpinCardiol. 2012;27(3):236-241. [CrossRef]
- Abudara, V.; Bechberger, J.; Freitas-Andrade, M.; De Bock, M.; Wang, N.; Bultynck, G.; Naus, C. C.; Leybaert, L.; Giaume, C. The connexin43 mimetic peptide Gap19 inhibits hemichannels without altering gap junctional communication in astrocytes. Front Cell Neurosci. 2014;8:306.
- Sun, T.; Li, D.; Hu, S.; Huang, L.; Sun, H.; Yang, S.; Wu, B.; Ji, F.; Zhou, D. Aging-dependent decrease in the numbers of enteric neurons, interstitial cells of Cajal and expression of connexin43 in various regions of gastrointestinal tract. Aging (Albany NY). 2018 Dec 11;10(12):3851-3865. [CrossRef]
- Seki, K.; Komuro, T. Immunocytochemical demonstration of the gap junction proteins connexin 43 and connexin 45 in the musculature of the rat small intestine. Cell Tissue Res. 2001;306(3):417-422.
- McClain, J.; Grubišić, V.; Fried, D.; Gomez-Suarez, R. A.; Leinninger, G. M.; Sévigny, J.; Parpura, V.; Gulbransen, B. D. Ca2+ Responses in Enteric Glia Are Mediated by Connexin-43 Hemichannels and Modulate Colonic Transit in Mice. Gastroenterology. 2014 Feb;146(2):497-507. [CrossRef]
- Akbarali, H.I.; Pothoulakis, C.; Castagliuolo, I. Altered ion channel activity in murine colonic smooth muscle myocytes in an experimental colitis model. BiochemBiophys Res Commun. 2000;275(2):637-642.
- Zhang, J.; Chandrasekaran, G.; Li, W.; Kim, D. Y.; Jeong, I. Y.; Lee, S. H.; Liang, T.; Bae, J. Y.; Choi, I.; Kang, H.; Maeng, J. S.; Kim, M. K.; Lee, T.; Park, S. W.; Kim, M. J.; Kim, H. S.; Ro, H.; Bae, Y. C.; Park, H. C.; Choi, E. Y., et al. Wnt-PLC-IP3-Connexin-Ca2+ axis maintains ependymal motile cilia in zebrafish spinal cord. Nat Commun. 2020;11(1):1860.
- Schneider, R.; Leven, P.; Glowka, T.; Kuzmanov, I.; Lysson, M.; Schneiker, B.; Miesen, A.; Baqi, Y.; Spanier, C.; Grants, I.; Mazzotta, E.; Villalobos-Hernandez, E.; Kalff, J. C.; Müller, C. E.; Christofi, F. L.; Wehner, S. A novel P2X2-dependent purinergic mechanism of enteric gliosis in intestinal inflammation. EMBO Mol Med. 2021;13(1):e12724. [CrossRef]
- Shen, F.; Huang, W.L.; Xing, B.P.; Fang, X.; Feng, M.; Jiang, C.M. Genistein Improves the Major Depression through Suppressing the Expression of miR-221/222 by Targeting Connexin 43. Psychiatry Investig. 2018;15(10):919-925.
- Yao, H.; Yan, J.; Yin, L.; Chen, W. Picroside II alleviates DSS-induced ulcerative colitis by suppressing the production of NLRP3 inflammasomes through NF-κB signaling pathway. ImmunopharmacolImmunotoxicol. 2022;44(3):437-446.
- Jeon, Y.D.; Lee, J.H.; Lee, Y.M.; Kim, D.K. Puerarin inhibits inflammation and oxidative stress in dextran sulfate sodium-induced colitis mice model. Biomed Pharmacother. 2020;124:109847.
- Jin, B.; Ha, S. E.; Wei, L.; Singh, R.; Zogg, H.; Clemmensen, B.; Heredia, D. J.; Gould, T. W.; Sanders, K. M.; Ro, S. Colonic Motility Is Improved by the Activation of 5-HT2B Receptors on Interstitial Cells of Cajal in Diabetic Mice. Gastroenterology. 2022 Jan;162(1):352.
- Camilleri, M.; Linden, D.R. Measurement ofgastrointestinal and colonic motor functionsin humans and animals. Cell Mol Gastroenterol Hepatol. 2016 Jul;2(4):412–28.
- Tizro, P.; Choi, C.; Khanlou, N. Sample preparation fortransmission electron microscopy. Methods Mol Biol.2019;1897:417–424.
- Cheung, G.; Chever, O.; Rollenhagen, A.; Quenech'du, N.; Ezan, P.; Lübke, J. H. R.; Rouach, N. Astroglial Connexin 43 Regulates Synaptic Vesicle Release at Hippocampal Synapses. Cells. 2023;12(8):1133.
- Orellana, J. A.; Froger, N.; Ezan, P.; Jiang, J. X.; Bennett, M. V.; Naus, C. C.; Giaume, C.; Sáez, J. C. ATP and glutamatereleased via astroglial connexin 43 hemichannelsmediate neuronal death through activation of pannexin 1hemichannels. J Neurochem. 2011;118:826–840.
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402-408.
Figure 1.
Effect of GDNF and Gap19 on UC group mice. (a) The mice’s body weight was monitored from 1 to 10 days. (b) The DAI score was monitored from 1 to 10 days. (c) The length of the colon was measured in different mice groups. (d) Histopathological staining in each colon in different mice groups (scale bar: 100 μm). CN: normal control group mice;UC: mice treated with 3.5% DSS alone;UC+GDNF: mice treated with DSS plus GDNF;UC+Gap19+GDNF: mice treated with DSS plus Gap19 and GDNF. Notes: All date are expressed as mean ± SD. (n = 8 for each group).** p < 0.01, * p < 0.05.
Figure 1.
Effect of GDNF and Gap19 on UC group mice. (a) The mice’s body weight was monitored from 1 to 10 days. (b) The DAI score was monitored from 1 to 10 days. (c) The length of the colon was measured in different mice groups. (d) Histopathological staining in each colon in different mice groups (scale bar: 100 μm). CN: normal control group mice;UC: mice treated with 3.5% DSS alone;UC+GDNF: mice treated with DSS plus GDNF;UC+Gap19+GDNF: mice treated with DSS plus Gap19 and GDNF. Notes: All date are expressed as mean ± SD. (n = 8 for each group).** p < 0.01, * p < 0.05.
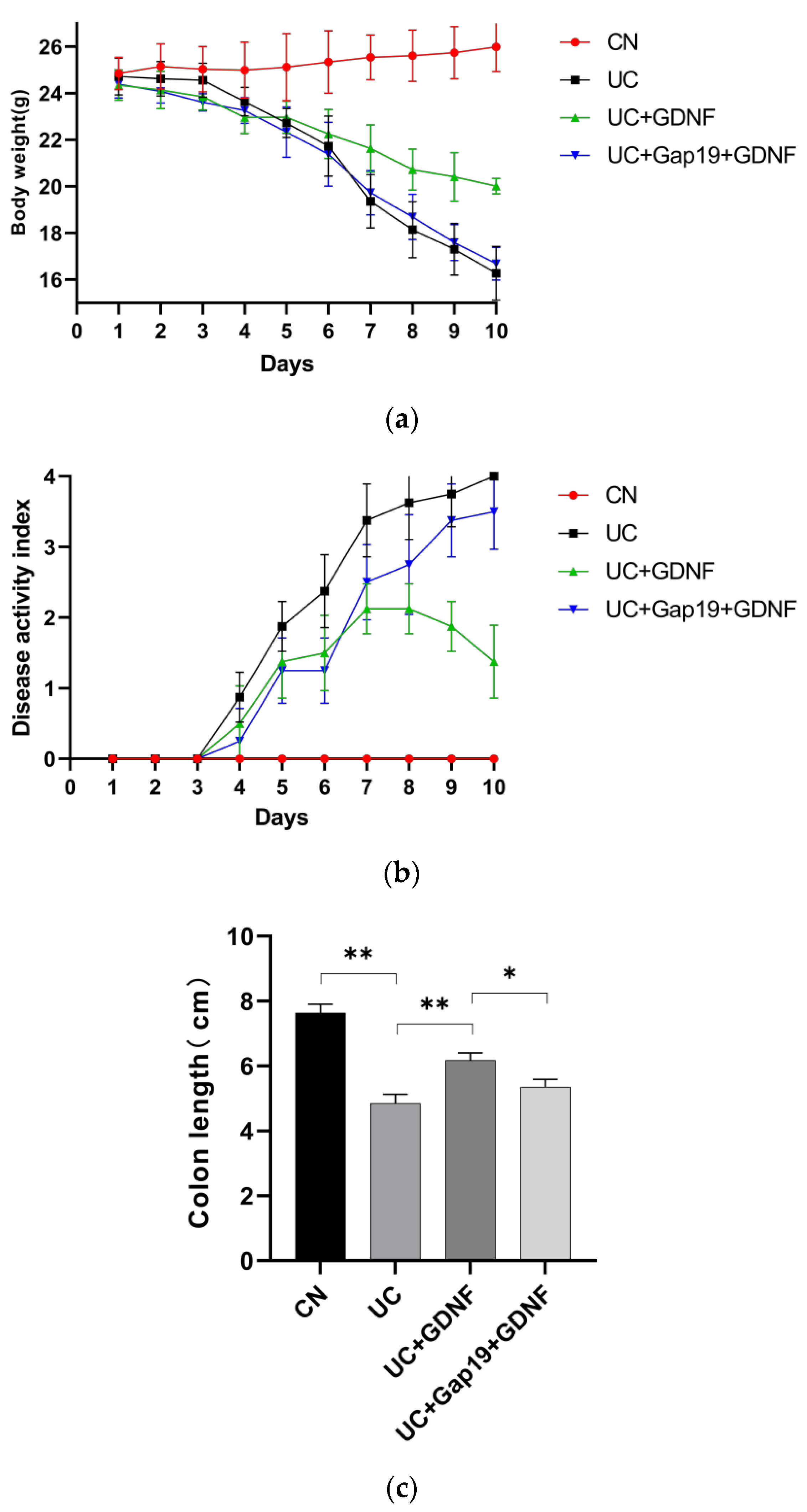
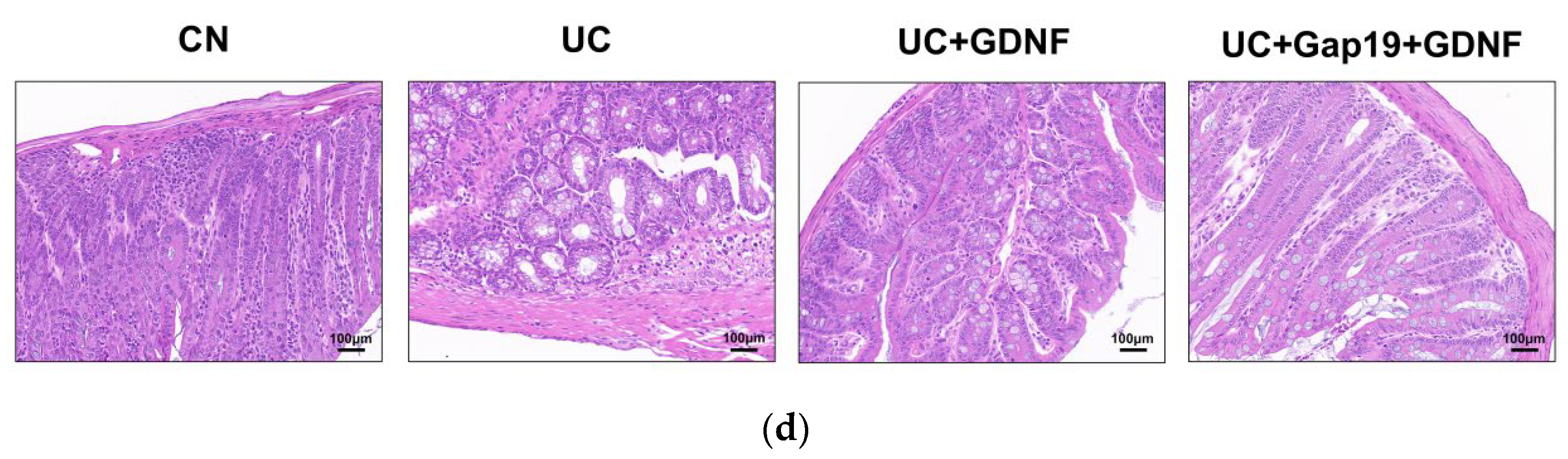
Figure 2.
Inflammatory response indexes in different mice groups (n = 8 for each group). (a) TNF-α, (b) IL-1β, (c) IL-6, and (d) CRP levels were measured by ELISA kits. Statistical analysis was performed using Student’s t-test, and the data were represented as mean ± SD. * P < 0.05, ** P < 0.01.
Figure 2.
Inflammatory response indexes in different mice groups (n = 8 for each group). (a) TNF-α, (b) IL-1β, (c) IL-6, and (d) CRP levels were measured by ELISA kits. Statistical analysis was performed using Student’s t-test, and the data were represented as mean ± SD. * P < 0.05, ** P < 0.01.
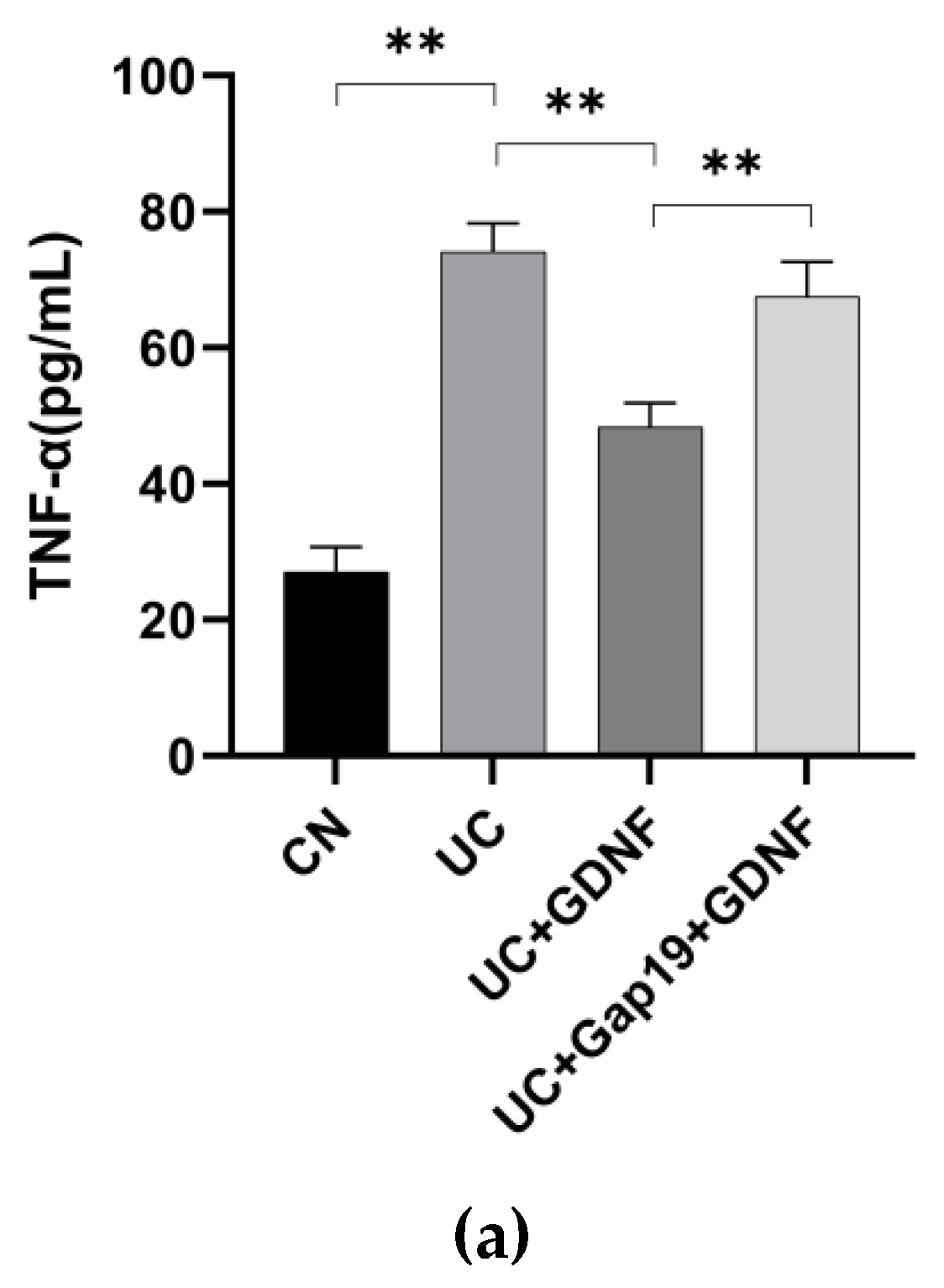
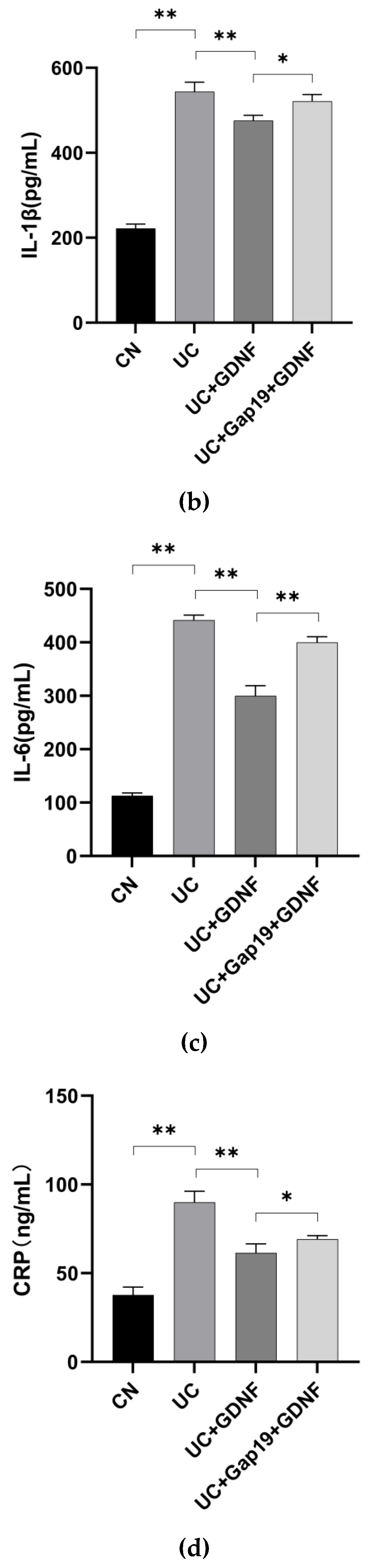
Figure 3.
The colonic transit time in different mice groups (n = 8 for each group). CN: normal control group mice; UC: mice treated with 3.5% DSS alone; UC+GDNF: mice treated with DSS plus GDNF; UC+Gap19 +GDNF: mice treated with DSS plus Gap19 and GDNF. Notes: All date are expressed as mean ± SD. ** p < 0.01, # p > 0.05.
Figure 3.
The colonic transit time in different mice groups (n = 8 for each group). CN: normal control group mice; UC: mice treated with 3.5% DSS alone; UC+GDNF: mice treated with DSS plus GDNF; UC+Gap19 +GDNF: mice treated with DSS plus Gap19 and GDNF. Notes: All date are expressed as mean ± SD. ** p < 0.01, # p > 0.05.
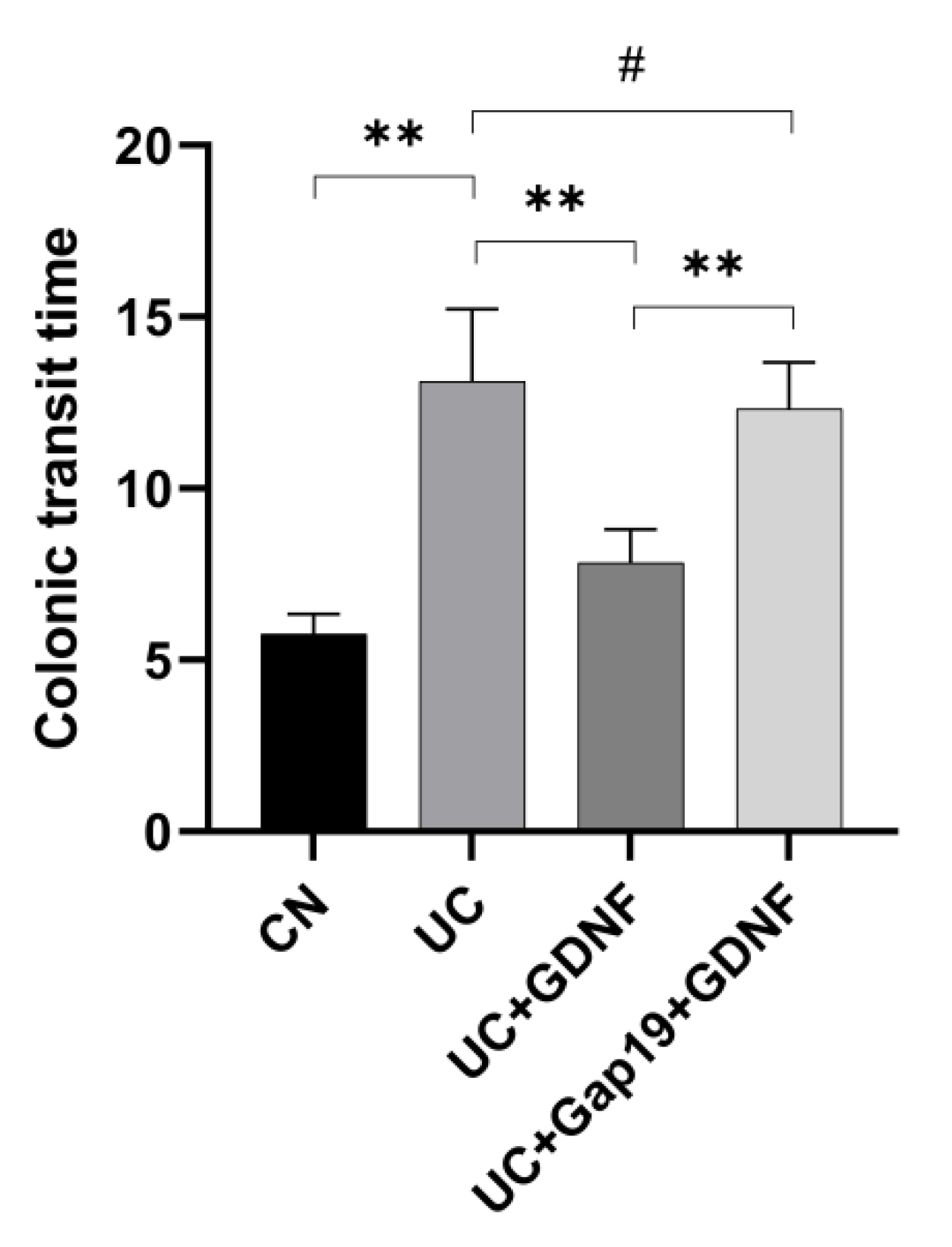
Figure 4.
The mRNA expression of Cx43 (a) and GDNF (b) were detected by using quantitative Real-Time PCR assay. Notes: All date are expressed as mean ± SD. (n = 8 for each group). ** p < 0.01, # p > 0.05.
Figure 4.
The mRNA expression of Cx43 (a) and GDNF (b) were detected by using quantitative Real-Time PCR assay. Notes: All date are expressed as mean ± SD. (n = 8 for each group). ** p < 0.01, # p > 0.05.
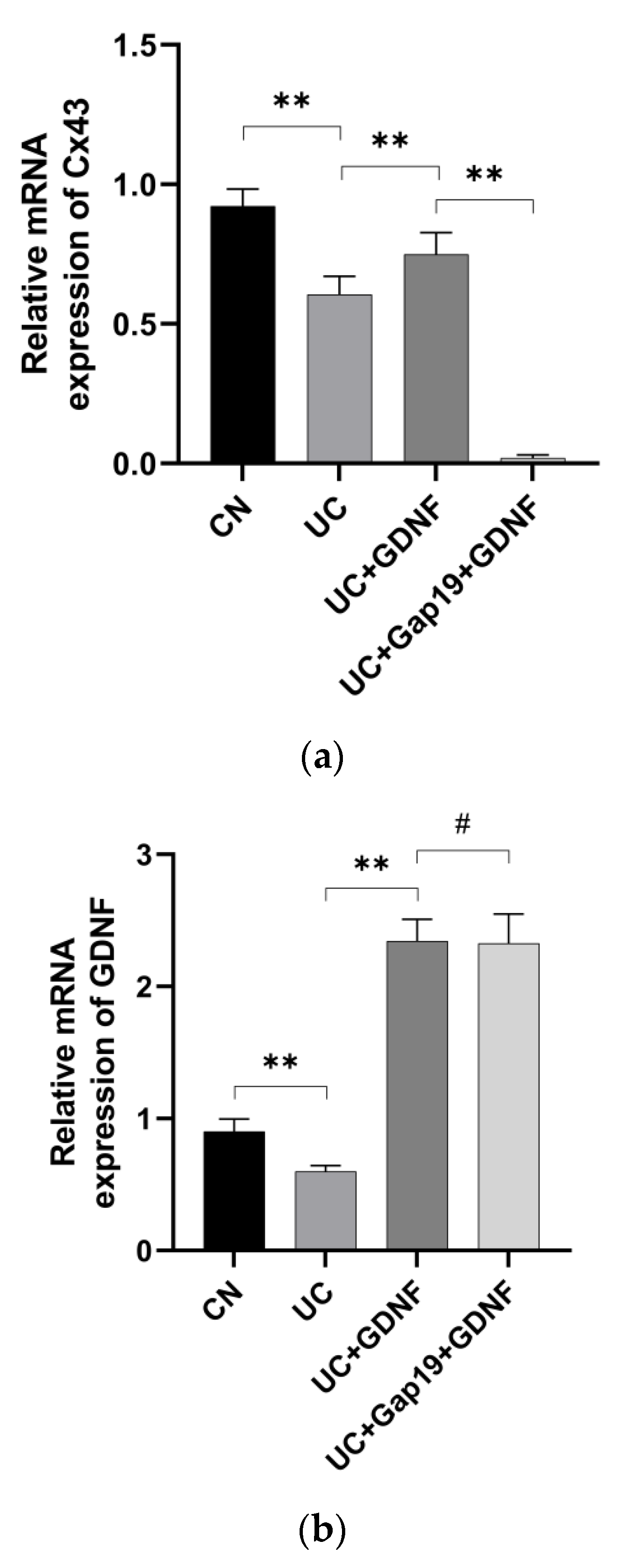
Figure 5.
Immunohistochemical staining (a, scale bar: 50 μm) of Cx43 (b) and GDNF (c) in colonic tissues. The relative immunohistochemical staining index was quantified. Notes: All date are expressed as mean ± SD. (n = 8 for each group). ** p < 0.01, # p > 0.05.
Figure 5.
Immunohistochemical staining (a, scale bar: 50 μm) of Cx43 (b) and GDNF (c) in colonic tissues. The relative immunohistochemical staining index was quantified. Notes: All date are expressed as mean ± SD. (n = 8 for each group). ** p < 0.01, # p > 0.05.
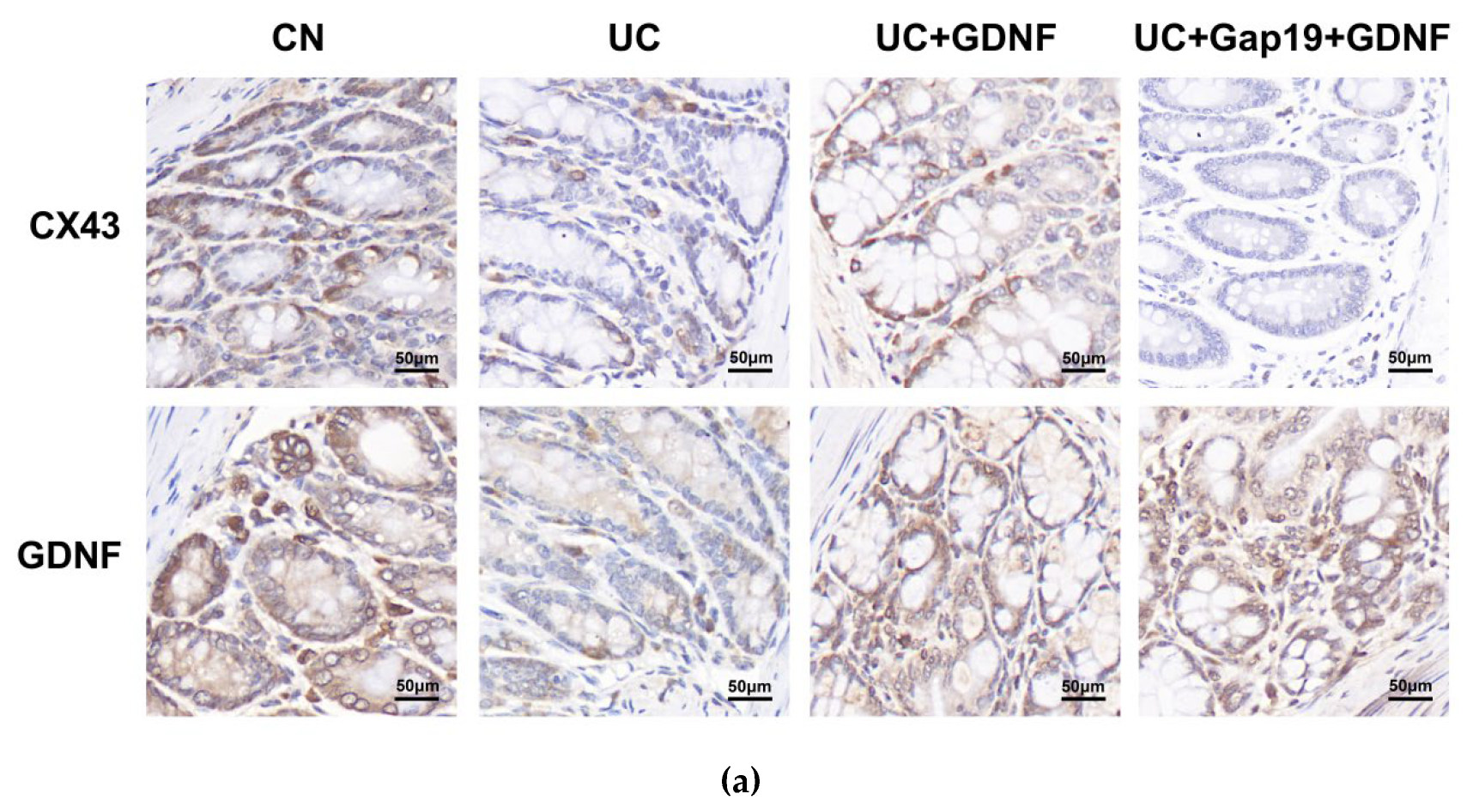
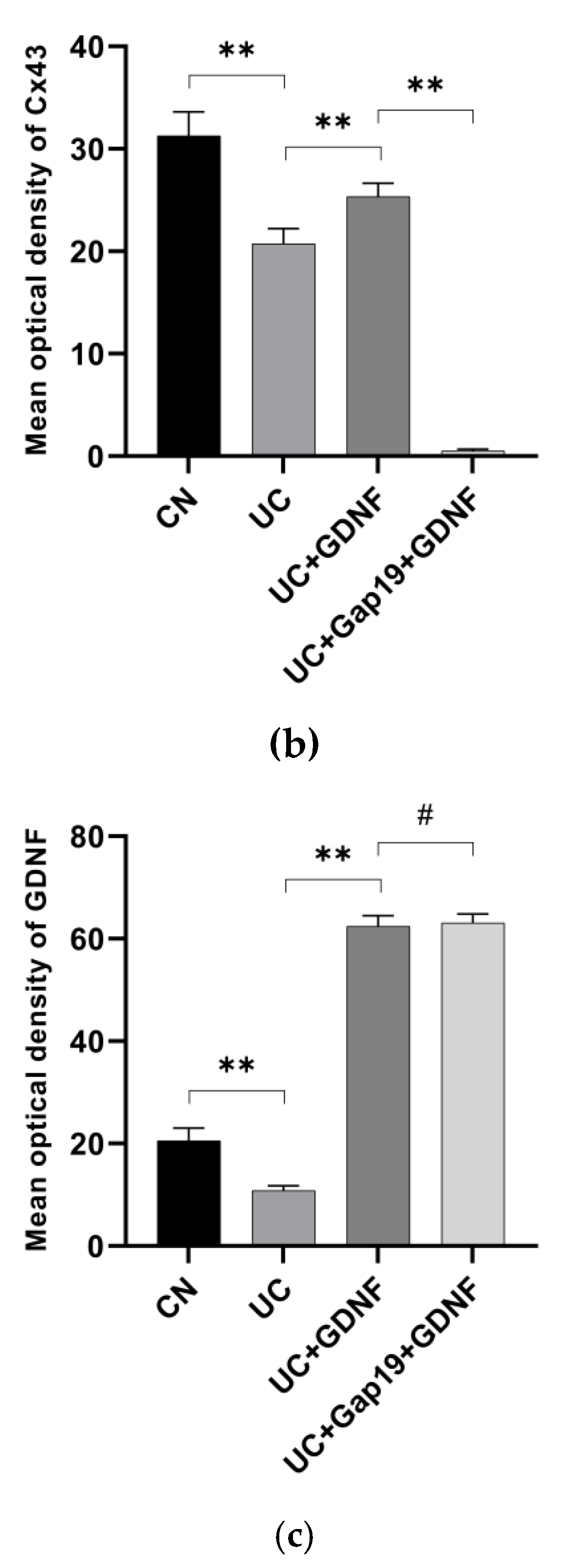
Figure 6.
Double-labelling immunofluorescence analysis of GDNF (red) and Cx43 (green) in colonic tissues in different mice groups. Same field showing DAPI (blue).Merged image showing co-localization of GDNF and Cx43. Scale bar: 50 μm.
Figure 6.
Double-labelling immunofluorescence analysis of GDNF (red) and Cx43 (green) in colonic tissues in different mice groups. Same field showing DAPI (blue).Merged image showing co-localization of GDNF and Cx43. Scale bar: 50 μm.
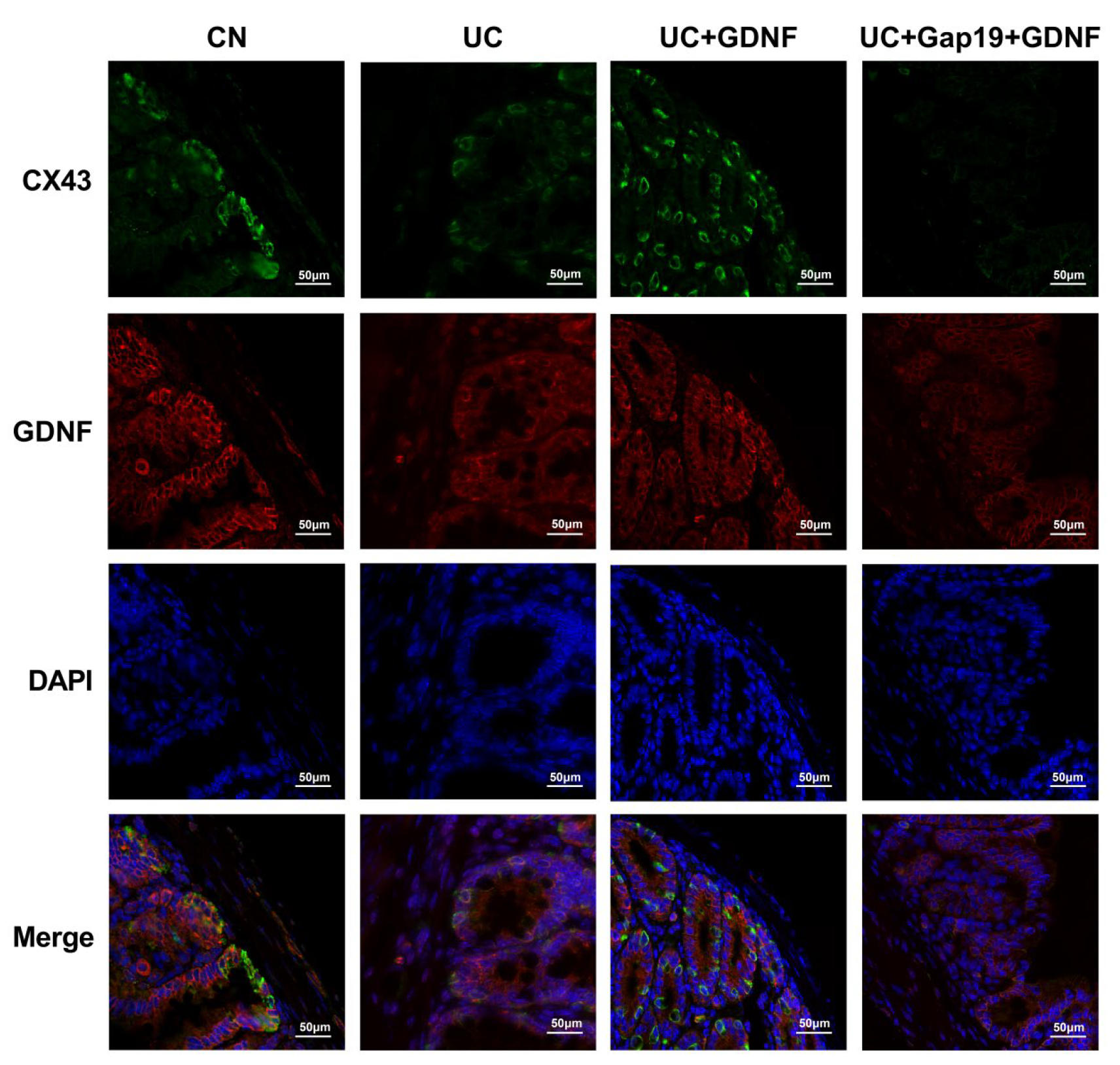
Figure 7.
The ultrastructure of the colon was observed by transmission electron microscopy in each group. The original amplification was 5000×, scale bar: 2.0 μm.
Figure 7.
The ultrastructure of the colon was observed by transmission electron microscopy in each group. The original amplification was 5000×, scale bar: 2.0 μm.
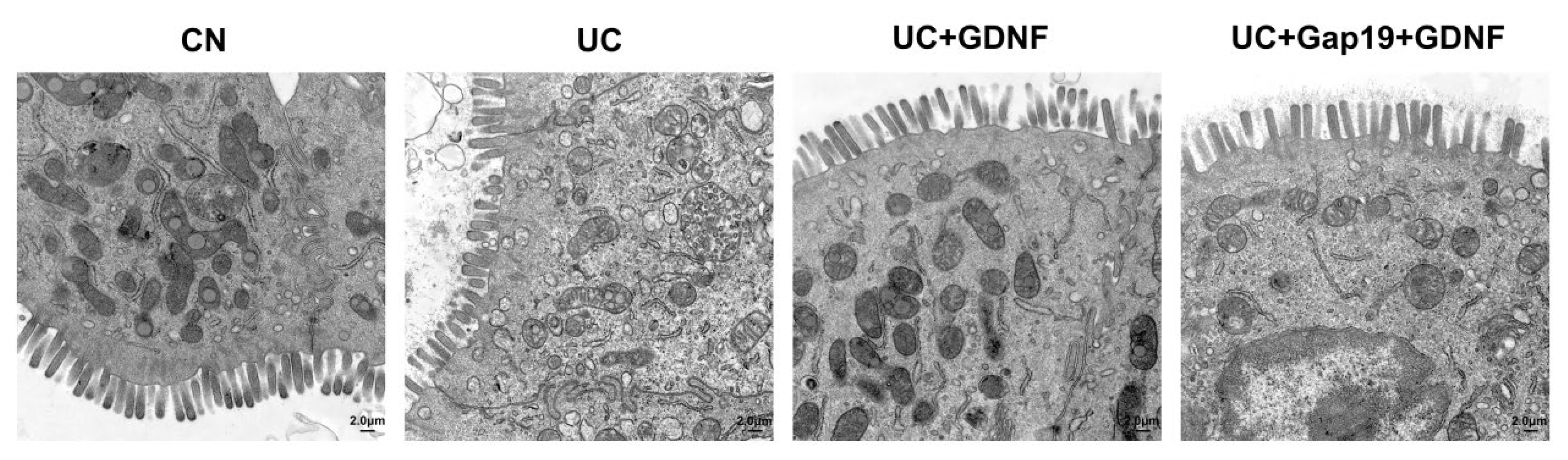

Table 1.
DAI scoring system.
| DAI score | Weight loss(%) | Stool condition | Gross bleeding |
| 0 | None | Normal | None |
| 1 | 1-5 | ||
| 2 | 5-10 | Loose stools | Hemoccult positive |
| 3 | 10-20 | ||
| 4 | >20 | Diarrhea | Severe bleeding |
Table 2.
Primers for quantitative real-time PCR analysis.
| GDNF | Forward | 5'-GTTAATGTCCAACTGGGGGTCTA-3' |
| GDNF | Reverse | 5'-ACAGCCACGACATCCCATAACT-3' |
| Cx43 | Forward | 5'-GGGTGATGAACAGTCTGCCTTT-3' |
| Cx43 | Reverse | 5'-AGCTTCTCTTCCTTTCTCATCACAT-3' |
| GAPDH | Forward | 5'-CCTCGTCCCGTAGACAAAATG-3' |
| GAPDH | Reverse | 5'-TGAGGTCAATGAAGGGGTCGT-3' |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



