
Submitted:
23 July 2024
Posted:
26 July 2024
You are already at the latest version
Abstract
The Axillary Seabream Pagellus acarne is of economic importance for recreational and commercial fisheries in the Atlantic and the Mediterranean, but a decline in total landings has been observed and raised some concern. Although biological information on the species has been provided for the Atlantic and Mediterranean, in the Portuguese continental slope data is restricted to the southern part of the country, which points to the relevance of the present work. A total of 520 individuals, between 7.8 cm and 39.9 cm TL, were monthly collected from September 2020 to August 2022 from commercial landings of fishing vessels operating off mainland Portugal, and used to study age and growth, the reproductive strategy and the annual sexual cycle. Age varied between 0 years (7.8 cm TL) and 15 years (34.2 cm TL) and the von Bertalanffy growth parameters were estimated from whole-otolith readings (L∞ = 35.2 cm, k = 0.133 yr–1 and t 0 = –2.78 yr). Pagellus acarne proved to be a protandric hermaphrodite species with males prevailing until 28.0 cm TL and females between 28.0 cm and 34.0 cm TL with a brief sex change phase. Females presented asynchronous oocyte development with a batch spawning pattern. The spawning season occurred from April to October with a spawning peak between June and August aas shown by histological analysis of the gonads and high levels of GSI during the period.
Keywords:
small scale fishery
; otolith
; von Bertalanffy
; histology
; reproductive strategy
; Portugal
1.Introduction
The Axillary Seabream Pagellus acarne (Risso, 1827) is a demersal species widely distributed along the European and African coasts, from Denmark to Senegal, and around the Madeira, Azores, Canary and Cape Verde Islands. It also occurs in the Mediterranean and the Black Sea [5]. This species typically occurs in seagrass beds and sand bottoms up to 500 m deep, usually between 40 and 100 m.
Pagellus acarne is caught by the demersal fleets operating along the eastern Atlantic coasts and the Mediterranean Sea [1], where it is mostly a by-catch of artisanal vessels and trawlers [2]. In Portugal, P. acarne is one of the main target species of small-scale commercial fisheries, namely in the Algarve [3], where it is mainly caught by trawl and gillnets [4]. Between 1995 and 2022, the national average landings were 820 tons per year [5]. However, since 2012, there has been an evident decline [5] in Portuguese total landings (reaching 293 tons in 2021), raising some concern. Currently, the only specific management measure applied for the species is fixing the minimum landing size at 18 cm TL. The biology of P. acarne has been studied in the Mediterranean [2,6,7,8,9,10,11] and the Atlantic [4,6,12,13,14]. Available information includes the validation of age assignment; the modelling of growth; the estimation of mortality and exploitation level; and the study of the spawning period, the sex ratio, the length at sexual inversion and first maturity, and the length-weight relationship. In Portugal, the only existing biological data for the species is restricted to the southern part of the country [4,13,14], with no biological information available for the western Atlantic coast. To date, only one study of the species’ reproductive cycle has included a histological analysis of the gonads [7].
The western continental Portuguese coast presents unique and dynamic geographic characteristics. Characterized by its position on the Iberian Peninsula, facing the Atlantic Ocean, the region has significant geographic and environmental diversity [15]. The coast is a biogeographic transition zone, with cooler northern waters and warmer southern waters, contributing to its rich marine and coastal habitats [16]. Along the western continental Portuguese coast, there is a complex and dynamic wind regime. North and northwest winds dominate in the summer, but the north wind is often deflected to blow from the west, southwest, or south over the Troia Peninsula [17]. Also, the impact of river drainage on coastal fisheries is complex and may be masked by other factors such as the upwelling regime [18].
This study aims to improve the available biological information on the age, growth, and reproduction of P. acarne off the western Atlantic coast of Portugal, extending the knowledge of the species geographically, and providing decision-makers with reliable information for the sustainable management of this resource.
2. Material and Methods
2.1. Sampling
The present study was based on a sample of 520 specimens between 7.8 cm and 39.9 cm TL, collected monthly from September 2020 to August 2022. The individuals were acquired from commercial landings of fishing vessels operating off mainland Portugal (Peniche, western Atlantic, 39°03′20.4′′N, 9°39′54.2′′W to 38°00′32.7′′N, 8°55′30.7′′W; Figure 1), using bottom trawl nets. Pagellus acarne individuals were identified as sparid fish with tall bodies without stripes but with a black spot at the pectoral fin base (Figure 2).
Each fish sampled was measured (total length, TL with 1 mm precision) and weighed (total and gutted weights, TW and GW, respectively, with 0.1 g precision). The gonad and liver weights (gonW and livW respectively, with 0.01 g precision) were also recorded, and sex was determined. Gonads were fixed in 4% buffered formalin for posterior histological analysis and sex confirmation. The otoliths sagitta were removed, rinsed with water, air-dried, and stored in labelled plastic tubes for future analysis.
2.2. Age and Growth
Individual age was estimated using right otoliths immersed in tap water, with the sulcus acusticus down. They were observed with a binocular stereomicroscope (magnification: 18x) Nikon SMZ745T, Tokyo, Japan and photographed with a coupled digital camera (Leica DFC290, Heerbrugg, Switzerland). The growth increment formation was considered annual, as previously validated [6,12,13,14]. Annual growth increments were observed using reflected light against a dark background, to enhance the differentiation between opaque and translucent increments. One opaque and one translucent increment corresponded to one year. Translucent increments were counted from the otolith nucleus towards the posterior edge to estimate age.
To evaluate the precision between readers, a subsample of 96 otoliths (a maximum of 10 for each 1 cm length class) was read by two independent readers. The average percentage error (APE) [19], the coefficient of variation (CV) [20], and the index of precision (D) [20] were determined. Bias was evaluated based on age-bias plots [21], which help identify departures from the 1:1 equivalence line. Systematic differences in assigned ages between readers were assessed with the Bowker-type test for symmetry [22] using the cran.r-project.org [23]. The remaining otoliths were read twice by the first author. Estimates that did not differ by more than 1 year were accepted and used to assign an average age, assuming the 1st of January as the birthdate [24]. If estimates differed by more than 1 year, the otolith was discarded.
To model species growth, three different growth models were fitted to the length-at-age data, including the typical three parameter von Bertalanffy growth model and two biphasic growth models (Hyper L∞ and Hyper k), described in Table 1. Both biphasic models were based on Soriano et al. [25] and correspond to variants of the von Bertalanffy model that allow changes in the growth parameter at a specific moment of the lifespan. By including a hyperbolic function that relies on a factor, designated as At (Table 1), these models modify either L∞ or k as age increases. To compare each model’s performance, the Akaike Information Criterion (AIC) was calculated using the stats package. A smaller value of AIC corresponded to a better fit. All growth models were implemented in R Statistical Software (v 4.3.0) [26], using the FSA [23] and nlstools packages [27].
2.3. Reproduction
From the total sample, 372 gonads (TL range between 19.8 cm and 34.2 cm) were analysed histologically. The gonads of all the females sampled between May and October were processed for histological analysis. Concerning the gonads sampled in the remaining months, a subsample of five individuals representing each macroscopic maturation stage was histologically analysed to ensure that the maturation stage was correctly assigned. The specimens classified as spawning capable and actively spawning were all analysed histologically, as well as all the gonads potentially with both ovarian and testicular tissues.
Fragments of the gonad mid-region preserved in formal were dehydrated with ethanol, embedded in methacrylate, sectioned at 3-5µm, stained with toluidine blue, and examined on a Leica DM 2500 LED microscope with a Leica MC 190 HD camera.
Gonads were histologically classified based on the most advanced cellular type of each developmental stage. Maturity phases were assigned according to the standardised terminology proposed by Brown-Peterson et al. [28]: immature (I), developing (D), spawning capable (SC) and its actively spawning (AS) subphase in females, regressing (RG), and regenerating (RE). Hermaphrodite specimens in the transition phase were defined as those with both visible ovarian and testicular tissues, with clear degeneration of the former and proliferation of the latter [29].
All stages of oocytes and cells of spermatogenic lineage were measured to characterise each stage of development. Each cell was measured for maximum and minimum diameter and averaged afterwards (to minimise the error from the loss of spherical shape caused by the histological processing). In females, only oocytes with a visible nucleus were measured. A total of 829 oocytes and 666 cells of spermatogenic lineage were analysed, in 27 females and 15 males in each maturity stage. These measurements were made using the software package ImageJ (http://imagej.nih.gov/ij/).
After characterizing each maturity phase, their monthly frequency was investigated to determine the spawning season, defined by the occurrence of the first and last actively spawning females during the sampling period. The monthly evolution of the gonadosomatic index (GSI=100×gonW/GW) was estimated to further corroborate the spawning season period. The fish’s condition was evaluated with two bioenergetic indices, the hepatosomatic index (HSI=100×livW/GW) and Fulton’s condition factor (K=100×GW/TL3). Monthly variation of the three indices was tested using the Kruskal-Wallis rank sum test and post hoc Dunn’s test for pairwise comparisons [30].
3. Results
3.1. Age and Growth
3.1.1. Length-Weight Relationship
The overall sex ratio (M : F) did not differ significantly from 1 : 1 (1 : 0.89; χ2 = 1.83, d.f. = 1, p > 0.05).
Pagellus acarne showed a strong relationship between TL and GW (r2 = 0.94) expressed by the equation: GW = 0.01 x TL 3.01 p > 0.05. This species shows isometric growth as b was not significantly different from 3 (t = 0.005, df = 489, p > 0.05).
3.1.2. Ageing Methodology, Precision, and Bias
Otoliths of P. acarne showed the typical deposition pattern for teleosts, with alternate opaque and translucent concentric increments around an opaque nucleus (Figure 2). As validated and assumed by other authors [2,4,20], the growth increments are formed annually with the opaque and rapid-growth regions deposited during spring and summer, and the translucent and slow-growth during autumn and winter. Only 2.8% of otoliths were considered unreadable.
The precision indices used for comparing readers were 7.4% and 5.2% for CV and APE, respectively. The percentage of total agreement was 37.5% rising to 87.5% at a tolerance of ±1 year. No significant differences between readers were found (U, p = 0.64) and there was no evidence of systematic disagreement between readers (Bowker’s symmetry test, p = 0.38). The age bias plot (Figure 3) shows the level of agreement between ages assigned by the two readers.
3.1.3. Growth Model
A total of 456 individuals (5 immature, 54 hermaphrodites, 213 females, and 183 males) were used for modelling growth. The smallest individual was 7.8 cm TL and was aged 0 years, and the largest was 34.2 cm TL and was aged 15 years. The age-length key obtained from direct otolith reading is shown in Table 2.
All models applied to the present dataset are represented in Figure 4 and the respective parameters estimates are expressed in Table 3. Hyperbolic biphasic models provided the better fit to the present data, given the lower values of AIC. Both Hyper L∞ and Hyper k models provided very similar estimates for the growth parameters. These models indicated a change in growth pattern at around 3 years of age (Hyper L∞ th = 3.36 years and Hyper k th = 3.40 years), which corresponds to an approximate TL of 21.3 cm. Although the conventional 3-parameter von Bertalanffy growth model (3p VBGM) provided estimates for the growth parameters that were very close to other models, its AIC value was higher (AIC = 1275.36, ∆AIC = 86.36), indicating a poor fit to the dataset.
3.2. Reproduction
3.2.1. Sample Characterization
Of all the initially sampled individuals, 372 individuals (140 females ranging from 20.6 to 34.2 cm TL; 227 males ranging from 19.2 to 32.0 cm TL; 5 hermaphrodites ranging from 27.4 to 29.7 cm TL) were analysed for reproductive purposes (Figure 5). Although otoliths from immature individuals were collected, the specimens were not used for reproductive studies. The smallest mature female measured 20.6 cm TL and the smallest mature male measured 19.8 cm TL. No females were observed at lengths < 20 cm TL and no males were sampled > 33 cm TL.
3.2.2. Structure of Ovaries and Testicles
A small portion of female tissue could be observed in most males. All gonads where tissues of both sexes were present, the ovarian and testicular components were completely separated by a thin layer of connective tissue (Figure 7a). In adult females, remaining male tissue was not observed.
All follicle and male germ cells were identified with histological examination of the gonads. Mean cell diameters are presented in Figure 6a and Figure 6b.
Pagellus acarne presented an asynchronous oocyte development, with a continuous oocyte diameter distribution and all oocyte stages occurring simultaneously in the mature phase (Figure 7d). Sex transition in this species appears to be brief since a small number of adult individuals (1.3%) presented both sex tissues in transition (Figure 7a).
The testicular tissue of P. acarne is structured into lobules, each containing numerous seminiferous tubules with cysts. These cysts are comprised of spermatogenic cells in the same developmental stage, enclosed by a slender layer of connective tissue (Figure 8a and Figure 8b).
Figure 7.
Transverse sections of female gonads of Pagellus acarne representing different maturation stages: a) hermaphrodite; b) developing (D); c) spawning capable (SC); d) actively spawning (AS); e) regressing (RG); and f) regenerating (RE). <Vtg3, early vitellogenic oocyte; Ac, cortical alveolar oocyte; Atr, atretic oocyte; Gvm, germinal vesicle migration oocyte; H, hydrated oocyte; O, ovary; Pg, primary growth oocyte; T, testicle; Vtg3, vitellogenic oocyte. Stain used: Toluidine blue. Magnification used: 4x (A); 10x (B, C, D, E, F).
Figure 7.
Transverse sections of female gonads of Pagellus acarne representing different maturation stages: a) hermaphrodite; b) developing (D); c) spawning capable (SC); d) actively spawning (AS); e) regressing (RG); and f) regenerating (RE). <Vtg3, early vitellogenic oocyte; Ac, cortical alveolar oocyte; Atr, atretic oocyte; Gvm, germinal vesicle migration oocyte; H, hydrated oocyte; O, ovary; Pg, primary growth oocyte; T, testicle; Vtg3, vitellogenic oocyte. Stain used: Toluidine blue. Magnification used: 4x (A); 10x (B, C, D, E, F).
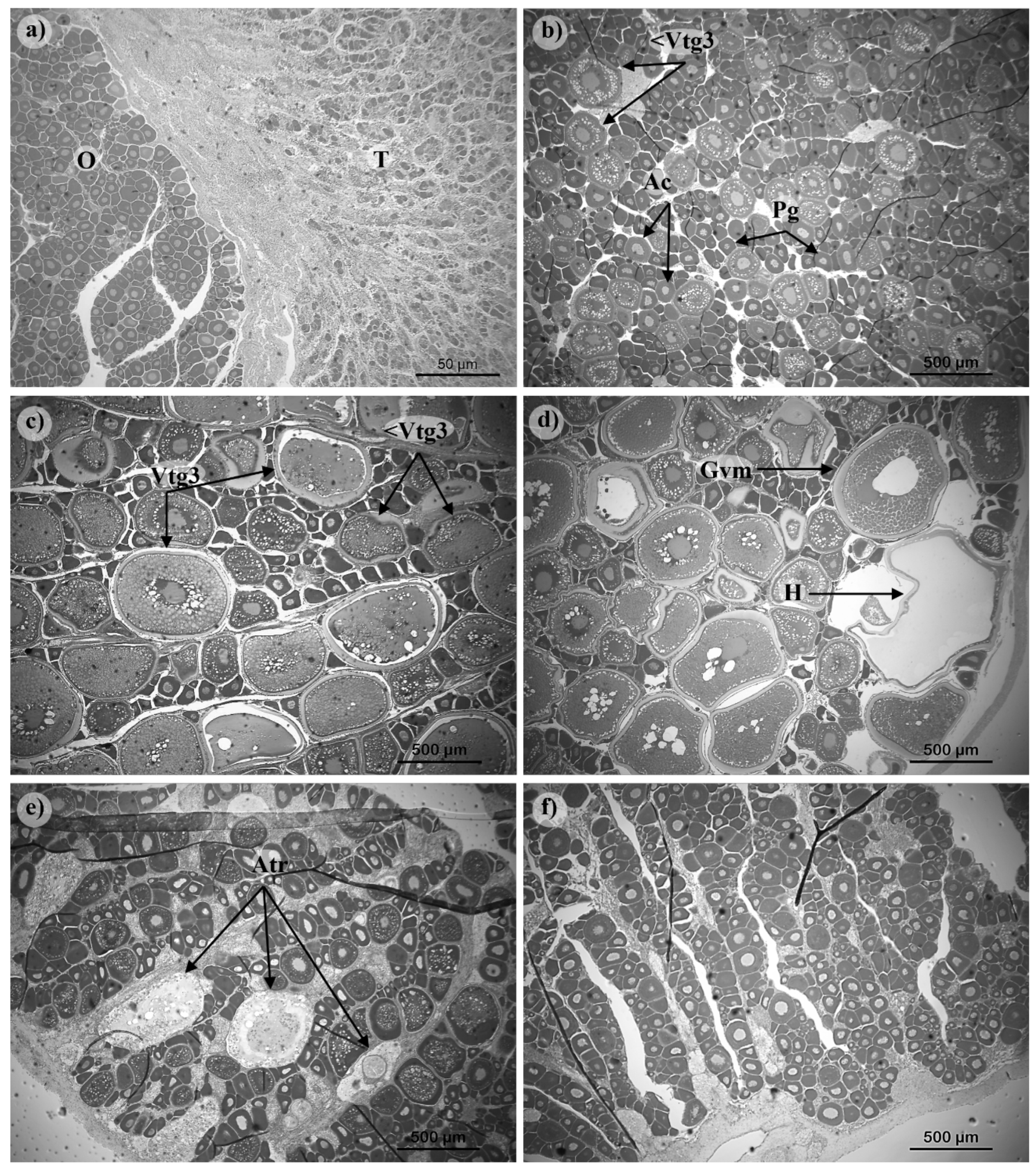
Figure 8.
Transverse section of male gonads of Pagellus acarne representing the different maturation stages: a) developing (D); b) spawning capable (SC); c) regressing (RG); and d) regenerating (RE). Sc, spermatocyte; Sg, spermatogonia; St, spermatocyst; Sz, spermatozoa. Stain used: Toluidine blue. Magnification used: 10x in all images.
Figure 8.
Transverse section of male gonads of Pagellus acarne representing the different maturation stages: a) developing (D); b) spawning capable (SC); c) regressing (RG); and d) regenerating (RE). Sc, spermatocyte; Sg, spermatogonia; St, spermatocyst; Sz, spermatozoa. Stain used: Toluidine blue. Magnification used: 10x in all images.

3.2.3. Sexual Cycle
Female and male sexual cycles proved to be synchronous with spawning capable/actively spawning gonads observed between April and October. Regressing and regenerating stages were registered between October and February and developing females in March and males in January, March and April (Figure 9).
Female and male P. acarne GSI presented the same annual developmental pattern, starting to increase in March, reaching its peak between June and August, and decreasing until December. There were significant differences in GSI among months and for both sexes, with the period from May to September showing significantly higher values than the rest of the year (H, d.f. females = 11 and d.f. males = 10, p < 0.05). The HSI also presented significant differences throughout the year for both sexes, with significantly higher values between April and October (H, d.f. = 11, p < 0.05) for females, and significantly lower values in December (H, d.f. = 11, p < 0.05) for males. The K condition factor showed an almost constant values throughout the year, with only significantly lower values in December compared with January and August in females, and in February compared to July to October in males (H, d.f. = 10, p < 0.05) (Figure 10a e 10b). The analysis of the evolution of GSI, HSI and K with the maturation phases of both sexes (Figure 10c e 10d) shows that the GSI is significantly higher in spawning capable and actively spawning individuals, and HSI and K are significantly lower in regressing and regenerating individuals (H, d.f. females = 4 and d.f. males = 3, p < 0.05).
4. Discussion
This study presents the first characterization of the age, growth, and reproduction of P. acarne in the western Atlantic coast and the second study using histological analyses of gonads to characterise the reproductive cycle of the species.
The species showed isometric growth, as already observed in the Aegean Sea [16], but contrary to the positive allometry found in most of the studies carried out in the Atlantic and Mediterranean (Table 4). This can be due to differences in habitat, environmental conditions, food availability and supply, temperature, salinity, and other ecological factors.
The precision between otolith readers was within the reference values (CV < 7.6% and APE < 5.5%) suggested by Campana [31], and a good percentage of agreement within ± 1 year (87.5%) was achieved, proving that sagitta otoliths are adequate structures for estimating age. This is also supported by the few discrepancies in age assignment of P. acarne across its distribution, with ages varying between 10 years and 13 years for individuals with 29 cm to 31 cm TL. In the present study, the maximum age estimated was 15 years for a female with 34 cm TL. Only works by Coelho et al. [13] and Abecasis et al. [14], off the Portuguese south coast, registered higher ages (18 years) for individuals with 36 cm TL, which can be related to different ecological factors and environmental variables. This is not surprising as spatial differences in length-weigh relationships have been observed for multiple species in various regions, including the North-eastern Atlantic Ocean [32]. On the other hand, studies have shown that length-weight relationships can differ between locations for the same species, potentially indicating distinct populations [33]. These variations highlight the importance of considering spatial and temporal factors when using LWRs for fisheries management and stock assessment.
For P. acarne from the Portuguese west coast, the model that best described its growth (Hyper L∞) indicated that this species shows a change in its growth rate. This model suggests a faster growth rate in the initial years, with growth rate decreasing when individuals reach 3.4 years of age (21.3 cm TL). According to Wilson et al. [34], many factors can lead to a different allocation of energetic costs between somatic growth and other bioecological traits. Processes such as the investment in reproduction, changes in the habitat, food, or other stressors can motivate changes in the lifetime growth of a species, negatively impacting growth [35]. Therefore, describing a species’ growth with a single curve may conceal valuable ecological information. For P. acarne, the observed change in growth pattern could be related to the allocation of energy towards maturation and reproduction, which leads to a deceleration in growth. Although it was not possible to estimate length-at-first maturity in the present study, the smallest mature individual sampled had 19.8 cm TL and was 3 years old. This indicates that the inflection point observed in the hyperbolic growth model is indeed correlated with fish maturation. Additionally, when compared to other already published studies (Table 4), P. acarne individuals from the western Atlantic seem to grow slower than those from the south of Portugal, the Gulf of Cadiz and other regions of the Mediterranean, and the southwest Gran Canaria (Table 4).
In this study, histological techniques were used as they are the most accurate way to determine the individual phase of sexual maturation in fish. Histology provides more consistent results than a visual inspection of reproductive organs, and is also essential for the study of gametogenesis and characterization of reproductive strategies [36].
Pagellus acarne proved to be a protandric hermaphrodite species, as previously described in other studies [2,6,7,11,12,13]. Males prevailed until 28 cm TL and females between 28 and 34 cm TL. Transitional individuals occurred between 27 cm and 30 cm TL, but in a very low percentage, proving that the sex change phase is brief, as also concluded by Pajuelo and Lorenzo [12]. Nevertheless, in the present study, sexual inversion occurred in length classes higher than those observed in other studies [6,7,13], which may be a consequence of using histological procedures to identify the transitional individuals more precisely.
Histologically, ovaries presented asynchronous development with a batch spawning pattern. The simultaneous occurrence of previtellogenic, vitellogenic, and maturation oocytes in AS ovaries, together with a long spawning season and mass atresia at the end of the spawning period may suggest indeterminate fecundity type for P. acarne.
In what concerns the sexual cycle, the spawning season of P. acarne from the west Portuguese Atlantic coast was between April and October for both sexes, with a peak in the summer between June and August as suggested by the high GSI values and the frequency of maturation stages. This long spawning period was also observed in other studies [6,7,13]. Nevertheless, for the Canarian archipelago [12] and the Gulf of Tunis [2], the spawning season occurred between October and March and September and December, respectively, suggesting a delay in lower latitudes toward the autumn/winter months. Marine fish species can exhibit different spawning seasons across geographical areas due to various factors, including temperature, photoperiod, and rainfall [37]. In the case of P. acarne, the spawning season delays for autumn/winter months in the Canarian archipelago and the Gulf of Tunis. When analysing the sea surface temperature during the spawning season in the three locations it varies between 15ºC and 18ºC, 23ºC and 19ºC, and 27ºC and 17ºC, in the west Portuguese Atlantic coast, the Canarian, and Tunis, respectively [38]. This may indicate that the species has an optimal temperature interval that does not allow very high temperatures forcing the species to run from the values observed in the summer months in the Canarian archipelago and the Gulf of Tunis.
The values of HSI were generally higher during the spawning season, suggesting a correlation with higher values of GSI. Although K values were roughly constant throughout the year, higher values were observed in the summer (August in females and July to October in males). The absence of high HSI values before the spawning season suggests the use of both liver and muscle reserves during this period, which, together with the extension of this period and the asynchronous oocyte development, indicates a mixed income-capital breeder pattern [39]. As expected, significantly higher values of GSI were observed in spawning individuals (reflecting more developed gonads) and significantly lower values of HSI and K in regressing/regenerating individuals (as the spawning season is over and there is no need to continue to store energy in the liver and muscle).
It was not possible to estimate the mean length at first maturity (L50) for P. acarne in this study due to the lack of immature individuals in the samples. But, as it was possible to estimate the age of immature individuals (7-14 cm TL, 0-1 yr), confirm that the smallest mature male observed measured 19.8 cm TL, and conclude that a transition in growth occurs around 21 cm TL. Therefore, L50 must occur between 14 cm TL and 21 cm TL. The smallest individual caught for the present reproductive study is lower than the L50 estimated by Santos et al. [4] for the south coast of Portugal (19.7 cm TL) but higher than the L50 estimated in other works (Table 5).
5. Conclusions
This study provides valuable biological information for a geographic area where little knowledge is available for P. acarne: age varied between 0 years (7.8 cm TL) and 15 years (34.2 cm TL); the hyperbolic modification of the von Bertalanffy curve showed the best fit to the data, presenting a change in growth at 3 years, which may correspond to the average age for sex maturation in the species; the species proved to be a protandric hermaphrodite with males prevailing below 28.0 cm TL and females above this value, with a short transitional phase; females presented asynchronous oocyte development with a batch spawning pattern; and the spawning season occurred from April to October with a spawning peak between June and August.
On the Portuguese coast, the only existing management measure for P. acarne is based on a minimum legal landing size of 18 cm TL, which according to the present study fulfils its purpose. Nevertheless, the decrease in landing observed in the past 10 years cannot be ignored. Females are particularly vulnerable to exploitation since they are predominantly over 30 cm. The removal of larger and older females by fishing may contribute to a decrease in the species’ reproductive potential, as these females are potentially more fecund than younger females. The fact that the biological information available for P. acarne is one-off and geographically localised, with no long-term data available, compromises a deeper understanding of the species’ life history and the perception of the possible impacts of fishing or changes in P. acarne’s ecosystem. Other protection measures may include artificial reefs, closed seasons, and changes in fishing patterns, but more consistent studies and catch time series analyses are crucial for better fishery management.
Author Contributions
Conceptualization, I.M., S.M.F., V.S.; Methodology, I.M., M.I.S, V.S.; Formal Analysis, I.M., L.S.G., M.I.S, V.S.; Investigation, I.M., V.S.; Resources, I.M., A.N., C.A., V.S.; Data Curation, I.M., L.S.G., M.I.S, V.S.; Writing – Original Draft Preparation, I.M., V.S.; Writing – Review & Editing, I.M., L.S.G., A.N., A.R.V., M.I.S, S.A., J.R., S.M.F, V.S.; Visualization, I.M., M.I.S, V.S.; Supervision, V.S., J.R., S.M.F.; Project Administration, S.M.F., J.R., V.S.; Funding Acquisition, L.S.G., S.M.F., J.R., V.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação para a Ciência e a Tecnologia, through the project UIDP/04292/2020 (https://doi.org/10.54499/UIDB/04292/2020; https://doi.org/10.54499/UIDP/ 04292/2020) awarded to MARE, project LA/P/0069/2020 (https://doi.org/10.54499/LA/P/0069/2 020) granted to the Associate Laboratory ARNET, and through the contracts CEECIND/02705/2017 (https://doi.org/10.54499/CEECIND/02705/2017/CP1387/CT0042) and CEECIND/01528/2017 (https://doi.org/10.54499/CEECIND/01528/2017/CP1387/CT0040) assigned to V.S. and A.R.V, respectively.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the fact that the fish used in the present study came from commercial fishing and were bought in the auction.
Acknowledgments
The authors are grateful to “Mestre Ricardo”, who facilitated the acquisition of the samples used in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Russell, B.; Carpenter, K.E.; Pollard, D. Pagellus acarne. The IUCN Red List of Threatened Species, 2014, e.T170229A1297432. https://www.iucnredlist.org/species/170229/1297432.
- Di Maio, F.; Geraci, M.; Scannella, D.; Falsone, F.; Colloca, F.; Vitale, S.; Rizzo, P.; Fiorentino, F. Age structure of spawners of the axillary seabream, Pagellus Acarne (Risso, 1827), in the central Mediterranean Sea (strait of Sicily). Reg Stud Mar Sci 2020, 34, 101082. [Google Scholar] [CrossRef]
- Erzini, K.; Bentes, L.; Coelho, R.; Correia, C.; Lino, P.G.; Monteiro, P.; Ribeiro, J.; Gonçalves, J.M.S. Fisheries biology and assessment of demersal species (Sparidae) from the South of Portugal. Commission of the European Communities DG XIV/C/1, Final Report. 2001, Ref. 98/082, 263 pp.
- Santos, M.M.; Monteirom, C.; Erzinim, K. Aspects of the biology and gillnet selectivity of the axillary seabream (Pagellus acarne, Risso) and common pandora (Pagellus erythrinus, Linnaeus) from the Algarve (south Portugal). Fish Res 1995, 23, 223–236. [Google Scholar] [CrossRef]
- INE (2023). Estatísticas da Pesca 2022. Lisboa, Portugal: INE, Instituto Nacional de Estatística. Available online: https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_publicacoes&PUBLICACOESpub_boui=66322600&PUBLICACOESmodo=2 (accessed on 31 May 2023).
- Velasco, E.M.; Jiménez-Tenorio, N.; Del Árbol, J.; Bruzón, M.A.; Baro, J.; Sobrino, I. Age, growth and reproduction of the axillary seabream, Pagellus acarne, in the Atlantic and Mediterranean waters off southern Spain. JMBA 2011, 91, 1243–1253. [Google Scholar] [CrossRef]
- Bensahla Talet, L.; Gherram, M.; Dalouche, F.; Bensahla Talet, A.; Boutiba, Z. Reproductive biology of Pagellus acarne (Risso, 1927) (Teleostei: Sparidae) off western Algerian waters (Western Mediterranean). CBM 2017, 58, 443–452. [Google Scholar] [CrossRef]
- Soykan, O.; İlkyaz, A.; Metin, G.; Kınacıgil, H. Growth and reproduction of Boops boops, Dentex macrophthalmus, Diplodus vulgaris, and Pagellus acarne (Actinopterygii: Perciformes: Sparidae) from east-central Aegean Sea, Turkey. AleP 2015, 45, 39–55. [Google Scholar] [CrossRef]
- Ilhan, D. Age, growth, and diet of axillary seabream, Pagellus acarne (Actinopterygii: Perciformes: Sparidae), in the central Aegean Sea. AleP 2018, 48, 329–339. [Google Scholar] [CrossRef]
- Bentata-Keddar, I.; Abid-Kachour, S.; Bouderbala, M.; Mouffok, S. Reproduction and growth of Axillary seabream Pagellus acarne (Risso, 1827) (Perciformes Sparidae) from the Western Algerian coasts. Biodivers J 2020, 11, 351–358. [Google Scholar] [CrossRef]
- Ali-Basha, N.; Hamwi, N.; Saad, A. Age, growth, Mortality and Exploitation of the Axillary seabream, Pagellus acarne (Risso, 1826) (Sparidae) from the Syrian marine waters. Damascus Uni J Agric Sci 2023, 39, 161–172. [Google Scholar]
- Pajuelo, B.J.G.; Lorenzo, J.M. Reproduction, age, growth and mortality of axillary seabream, Pagellus acarne (Sparidae), from the Canarian archipelago. J Appl Ichthyol 2000, 16, 41–47. [Google Scholar] [CrossRef]
- Coelho, R.; Bentes, L.; Gonçalves, J.; Correia, C.; Lino, P.; Monteiro, P. Age, growth and reproduction of the axillary seabream, Pagellus acarne (Risso, 1827), from the South coast of Portugal. Thalassas 1999, 21, 77–84. [Google Scholar]
- Abecasis, D.; Bentes, L.; Coelho, R.; Correia, C.; Lino, P.G.; Monteiro, P.; Gonçalves, J.M.S.; Ribeiro, J.; Erzini, K. Ageing seabreams: A comparative study between scales and otoliths. Fish Res 2008, 89, 37–48. [Google Scholar] [CrossRef]
- de Figueiredo, T. Montesinho and the Mountains of Northern Portugal. In Natural Heritage from East to West; Evelpidou, N., Figueiredo, T., Mauro, F., Tecim, V., Vassilopoulos, A., Eds.; Springer: Berlin, Heidelberg, 2010. [Google Scholar] [CrossRef]
- Cardoso, P.G.; Dolbeth, M.; Sousa, R.; Relvas, P.; Santos, R.; Silva, A.; Quintino, V. Chapter 7 - The Portuguese Coast. In World Seas: an Environmental Evaluation (Second Edition); Sheppard, C., Ed.; Academic Press, p. 2019; pp. 189–208. [Google Scholar] [CrossRef]
- Neto, C.S. Circulação de ar na península de Tróia e costa da Galé. Finisterra 2000, 35. [Google Scholar] [CrossRef]
- Gamito, R.; Vinagre, C.; Teixeira, C.M.; Costa, M.J.; Cabral, H.N. Are Portuguese Coastal Fisheries Affected by River Drainage? Aquat. Living Resour. 2016, 29, 102. [Google Scholar] [CrossRef]
- Beamish, R.J.; Fournier, D.A. A method for comparing the precision of a set of age determinations. Can J Fish Aquat Sci 1981, 38, 982–983. [Google Scholar] [CrossRef]
- Chang, W.Y.B. A statistical method for evaluating the reproducibility of age determination. Can J Fish Aquat Sci 1982, 39, 1208–1210. [Google Scholar] [CrossRef]
- Campana, S.E.; Annand, M.C.; McMillan, J.I. Graphical and statistical methods for determining the consistency of age determinations. Trans Am Fish Soc 1995, 124, 131–138. [Google Scholar] [CrossRef]
- Hoenig, J.M.; Morgan, M.J.; Brown, C.A. Analysing differences between two age determination methods by tests of symmetry. Can J Fish Aquat Sci 1995, 52, 364–68. [Google Scholar] [CrossRef]
- Ogle, D.H.; Doll, J.C.; Wheeler, A.P.; Dinno, A. (2023). _FSA: Simple Fisheries Stock Assessment Methods_. R package version 0.9.4, <https://CRAN.R-project.org/package=FSA>.
- Williams, T.; Bedford, B.C. The use of otoliths for age determination. In The Ageing of Fish; Bagenal, T.B., Ed.; Gresham Press: Old Woking, England, 1974; pp. 114–123. [Google Scholar]
- Soriano, M.; Moreau, J.; Hoenig, J.M.; Pauly, D. New functions for the analysis of two-phase growth of juvenile and adult fishes, with application to Nile Perch. Transactions of the American Fisheries Society 1992, 121, 486–493. [Google Scholar] [CrossRef]
- R Core Team (2023). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
- Baty, F.; Ritz, C.; Charles, S.; Brutsche, M.; Flandrois, J.-P.; Delignette-Muller, M.-L. A Toolbox for Nonlinear Regression in R: The Package nlstools. Journal of Statistical Software 2015, 66, 1–21. [Google Scholar] [CrossRef]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A standardized terminology for describing reproductive development in fishes. Marine and Coastal Fisheries: Dynamics, Management, and Ecosystem Science 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Sadovy, Y.; Shapiro, D.Y. Criteria for the diagnosis of hermaphroditism in fishes. Copeia 1987, 136–156. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Campana, S.E. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Mahé, K.; Bellamy, E.; Delpech, J.P.; et al. Evidence of a relationship between weight and total length of marine fish in the North-eastern Atlantic Ocean: physiological, spatial and temporal variations. J Mar Biol Assoc UK 2018, 98, 617–625. [Google Scholar] [CrossRef]
- Biolé, F.G.; Valpedo, A.V.; Thompson, G.A. Length-weight and length-length relationship for three marine fish species of commercial importance from southwestern Atlantic Ocean coast. Lat Am J Aquat Res 2020, 48. [Google Scholar] [CrossRef]
- Wilson, K.L.; Honsey, A.E.; Moe, B.; Venturelli, P.; Reynolds, J. Growing the biphasic framework: Techniques and recommendations for fitting emerging growth models. Methods in Ecology and Evolution 2017, 9, 822–833. [Google Scholar] [CrossRef]
- Carbonara, P.; et al. Biphasic versus monophasic growth curve equation, an application to common sole (Solea solea, L.) in the northern and Central Adriatic Sea. Fisheries Research 2023, 263, 106694. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Brown-Peterson, N.J.; Murua, H.; Tomkiewicz, J.; Wyanski, D.M.; Saborido-Rey, F. Emerging issues and methodological advances in fisheries reproductive biology. Mar Coast Fish [dnlm] 2011, 3, 32–51. [Google Scholar] [CrossRef]
- Lin, H.R.; Peter, R.E. Hormones and Spawning in Fish. Asian Fish Sci 1996, 9, 21–23. [Google Scholar] [CrossRef]
- GLORYS12V1. E.U. Copernicus Marine Service Information (CMEMS). Marine Data Store (MDS). https://doi.org/10.48670/moi-00021 (Accessed on 10-07-2024). [CrossRef]
- Mcbride, R.S.; Somarakis, S.; Fitzhugh, G.R.; Albert, A.; Yaragina, N.A.; Wuenschel, M.J.; Alonso-Fernández, A.; Basilone, G. Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish Fish 2015, 16, 23–57. [Google Scholar] [CrossRef]
Figure 1.
Geographic location of the sampling area. Black rectangle marks the sampling area (Peniche, western Atlantic, 39°03′20.4′′N, 9°39′54.2′′W to 38°00′32.7′′N, 8°55′30.7′′W).
Figure 1.
Geographic location of the sampling area. Black rectangle marks the sampling area (Peniche, western Atlantic, 39°03′20.4′′N, 9°39′54.2′′W to 38°00′32.7′′N, 8°55′30.7′′W).
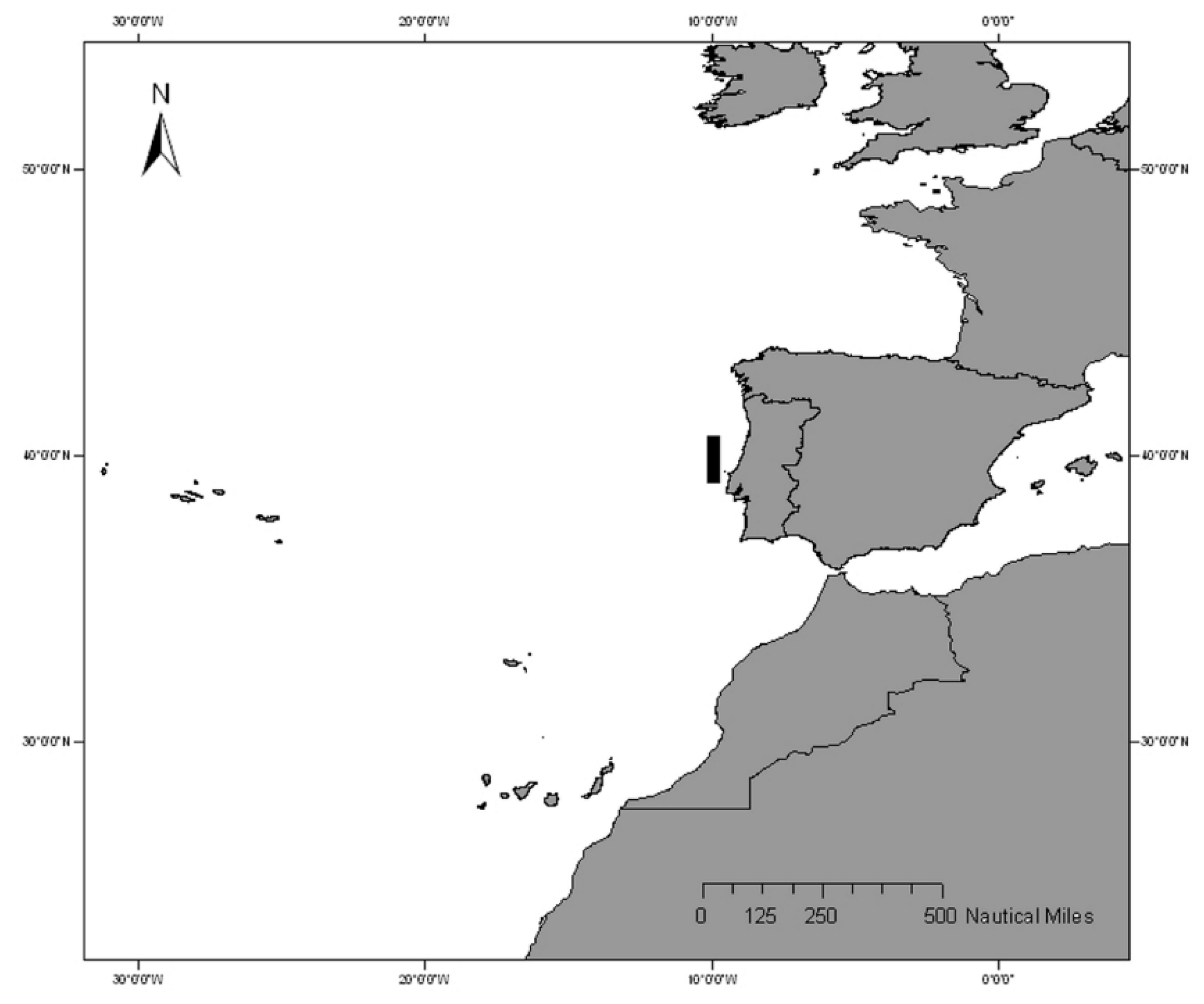
Figure 2.
Pagellus acarne image (© Scandinavian Fishing Year Book, in https://fish-commercial-names.ec.europa.eu/fish-names/species/pagellus-acarne_en).
Figure 2.
Pagellus acarne image (© Scandinavian Fishing Year Book, in https://fish-commercial-names.ec.europa.eu/fish-names/species/pagellus-acarne_en).

Figure 2.
Right sagittal otolith from 5 years old Pagellus acarne with 25.1 cm TL. Dots indicate the counted annual growth increments.
Figure 2.
Right sagittal otolith from 5 years old Pagellus acarne with 25.1 cm TL. Dots indicate the counted annual growth increments.

Figure 3.
Age-bias plots of age assignments from otoliths obtained by independent readers for Pagellus acarne from the Portuguese west coast. The 45° line represents 100% agreement.
Figure 3.
Age-bias plots of age assignments from otoliths obtained by independent readers for Pagellus acarne from the Portuguese west coast. The 45° line represents 100% agreement.
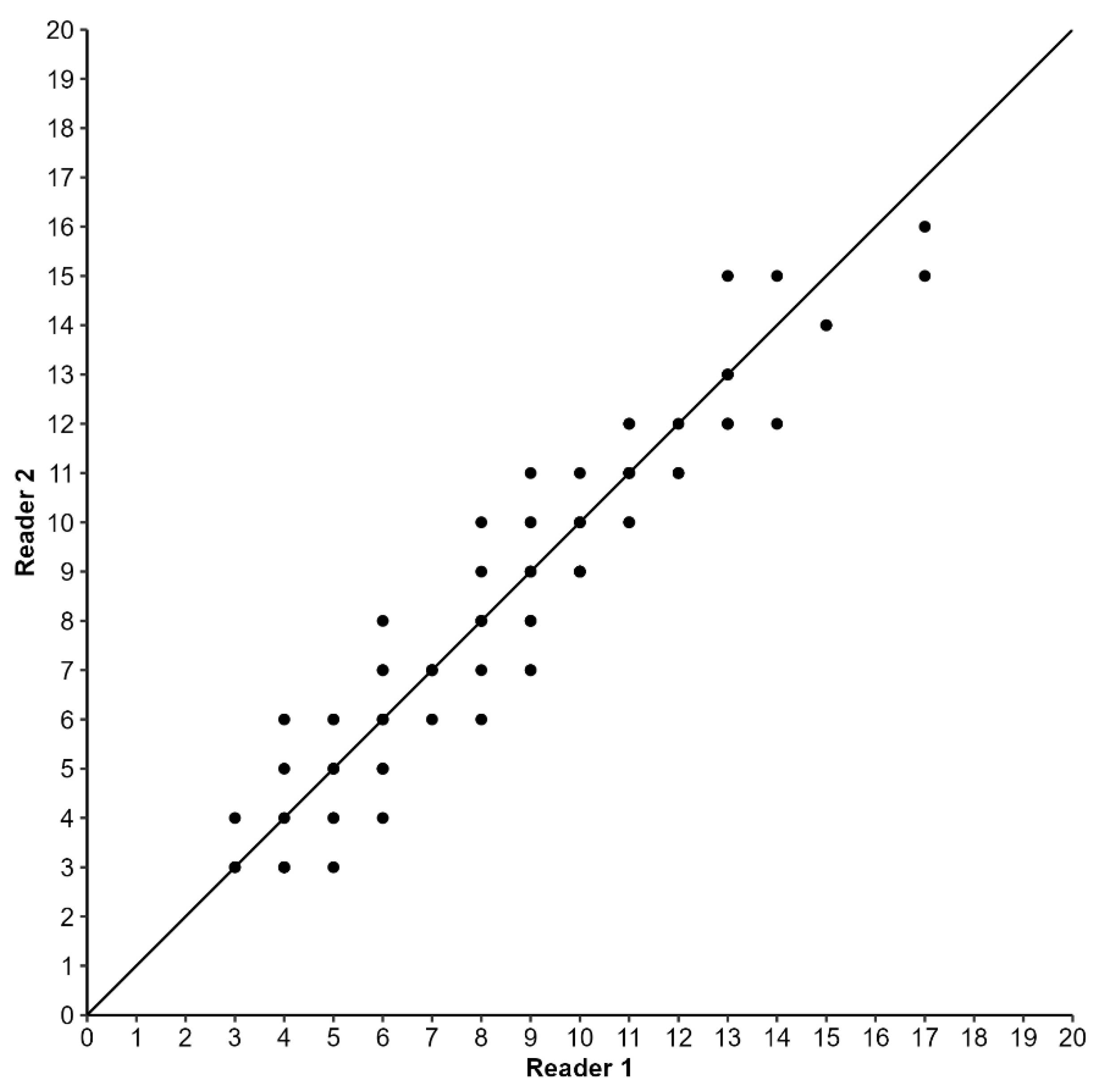
Figure 4.
The von Bertalanffy growth functions from the three approaches adjusted to age-at-length data of axillary seabream Pagellus acarne from the Portuguese west coast. Solid black line – von Bertalanffy growth model with 3 parameters; Solid dark grey line – L∞ hyperbolic biphasic model; Dashed light grey line – k hyperbolic biphasic model.
Figure 4.
The von Bertalanffy growth functions from the three approaches adjusted to age-at-length data of axillary seabream Pagellus acarne from the Portuguese west coast. Solid black line – von Bertalanffy growth model with 3 parameters; Solid dark grey line – L∞ hyperbolic biphasic model; Dashed light grey line – k hyperbolic biphasic model.
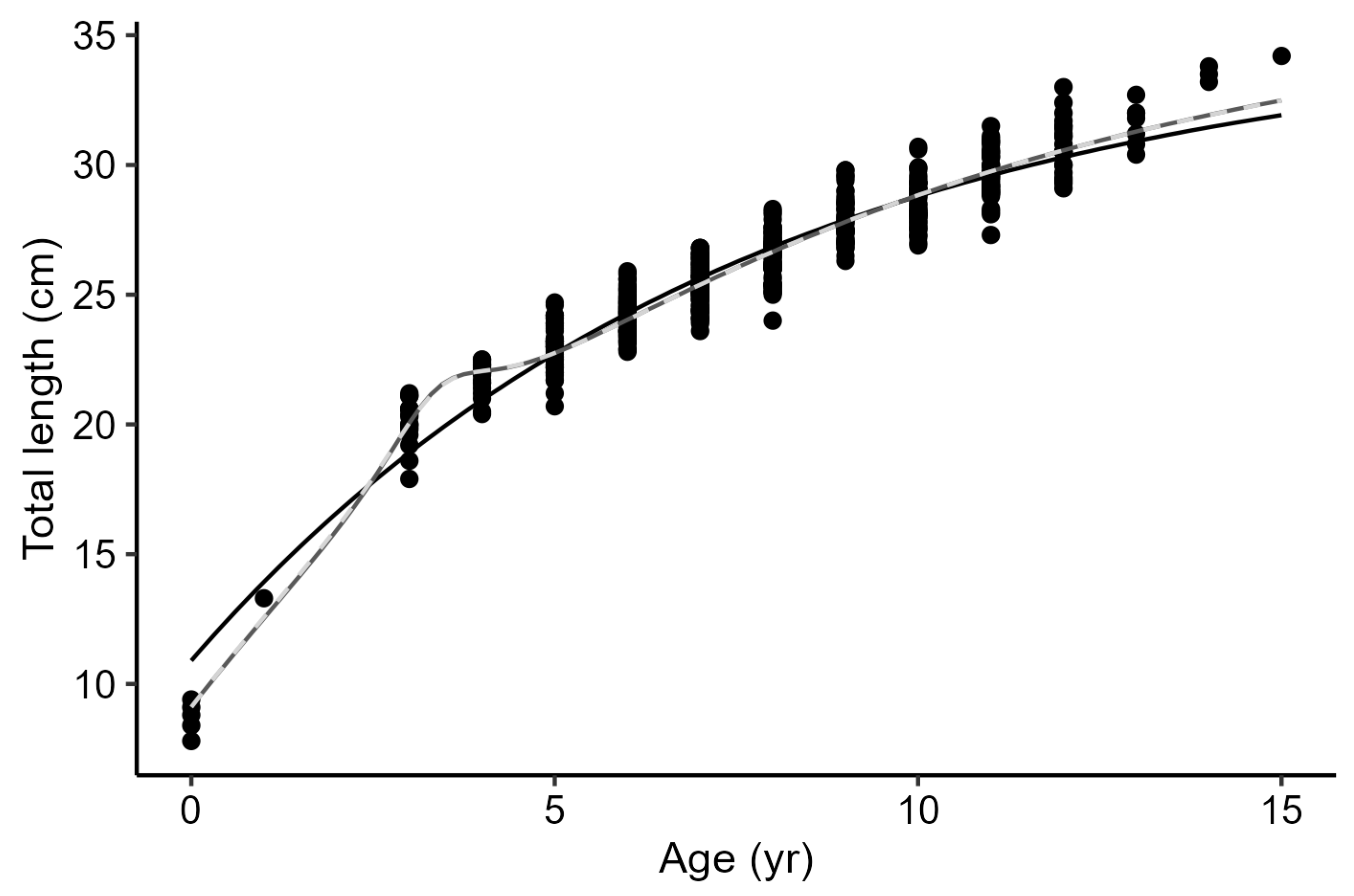
Figure 5.
Number of Pagellus acarne females (white), males (black) and hermaphrodites (grey) by 1 cm length class from the Portuguese west coast.
Figure 5.
Number of Pagellus acarne females (white), males (black) and hermaphrodites (grey) by 1 cm length class from the Portuguese west coast.
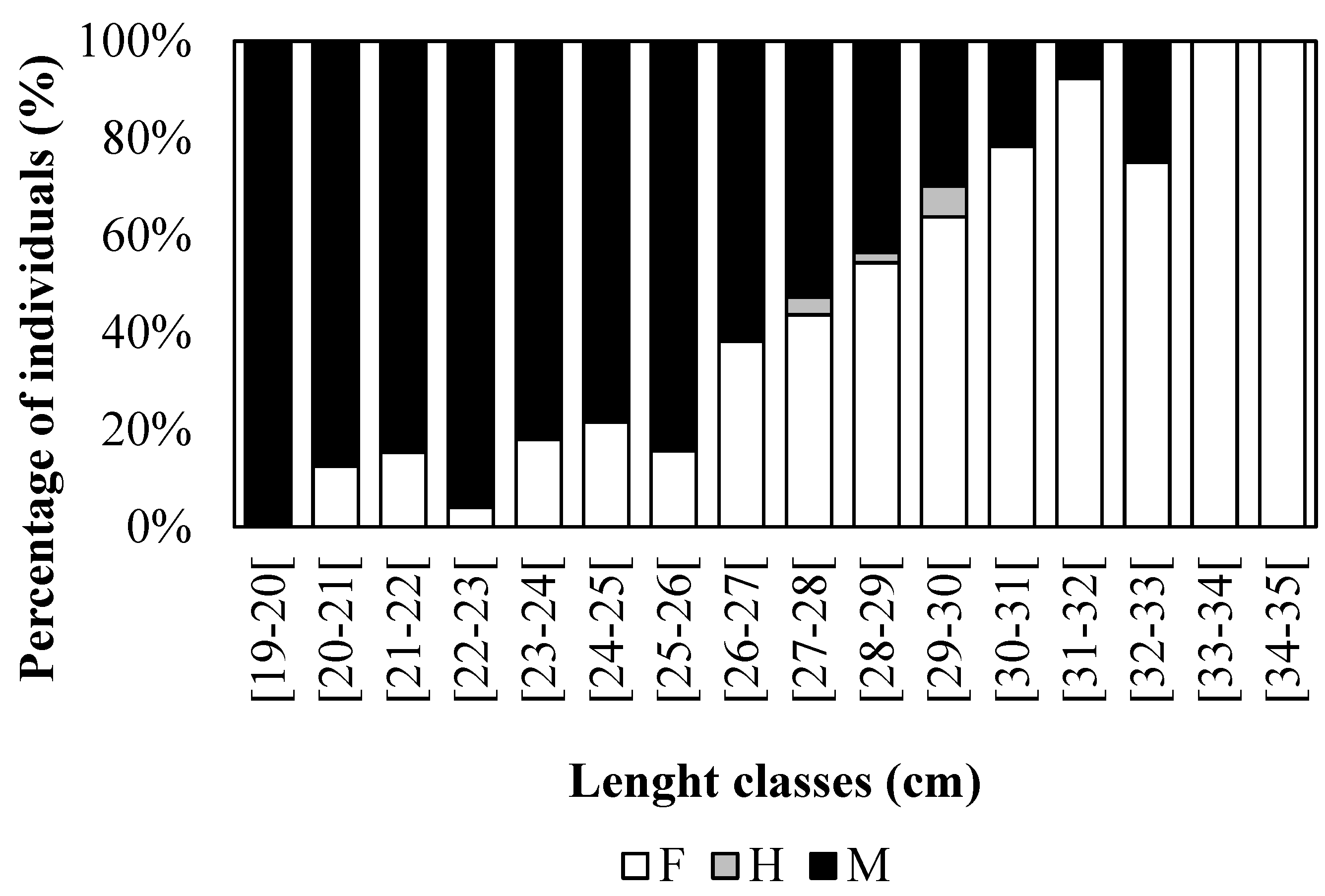
Figure 6.
Mean diameter and standard deviation of gametes from (a) females (triangles) and (b) males (squares) of Pagellus acarne. Pg, primary growth oocyte; Ac, cortical alveolar oocyte; <Vtg3, early vitellogenic oocyte; Vtg3, vitellogenic oocyte; Gvm, geminal vesicle migration oocyte; Yc, yolk coalescence oocyte; H, hydrated oocyte; Sg, spermatogonia; Sc, spermatocyte; St, spermatid; Sz, spermatozoa.
Figure 6.
Mean diameter and standard deviation of gametes from (a) females (triangles) and (b) males (squares) of Pagellus acarne. Pg, primary growth oocyte; Ac, cortical alveolar oocyte; <Vtg3, early vitellogenic oocyte; Vtg3, vitellogenic oocyte; Gvm, geminal vesicle migration oocyte; Yc, yolk coalescence oocyte; H, hydrated oocyte; Sg, spermatogonia; Sc, spermatocyte; St, spermatid; Sz, spermatozoa.
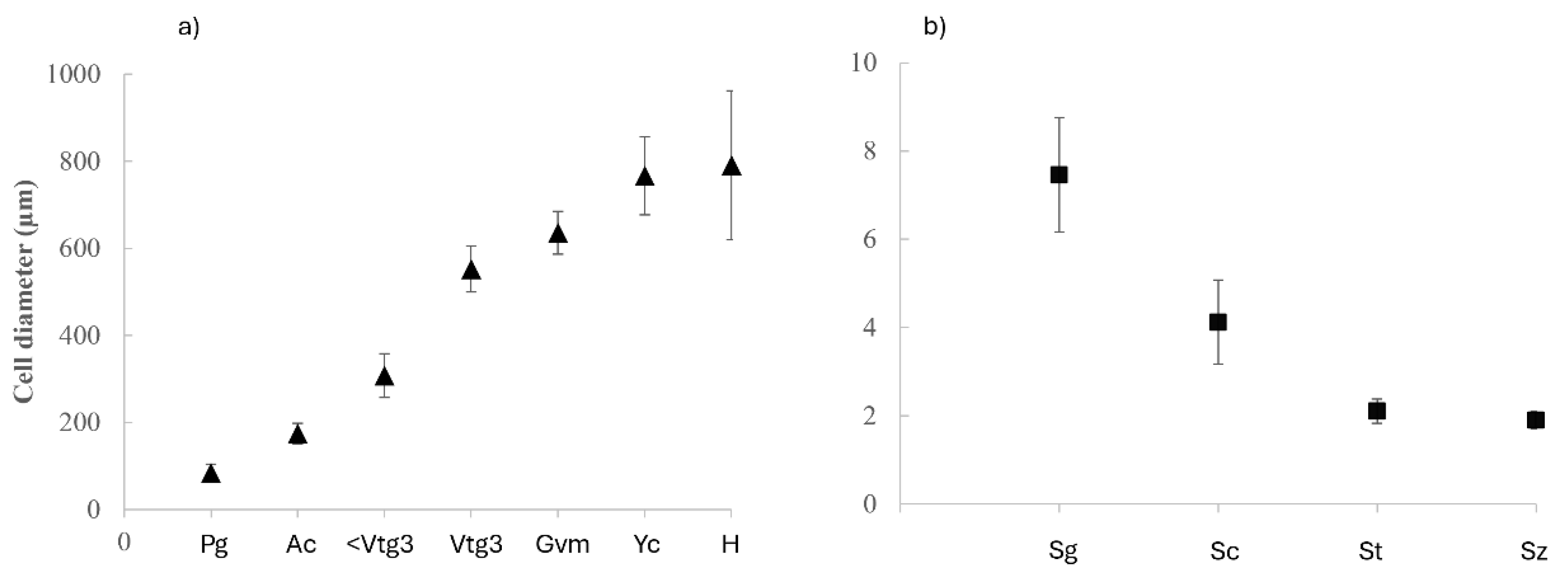
Figure 9.
Percentage of different maturation phases through the sampling period for females (a) and males (b) of Pagellus acarne from the Portuguese west coast. (D) developing; (SC) spawning capable; (AS) actively spawning; (RE) regressing; (RG) regenerating.
Figure 9.
Percentage of different maturation phases through the sampling period for females (a) and males (b) of Pagellus acarne from the Portuguese west coast. (D) developing; (SC) spawning capable; (AS) actively spawning; (RE) regressing; (RG) regenerating.
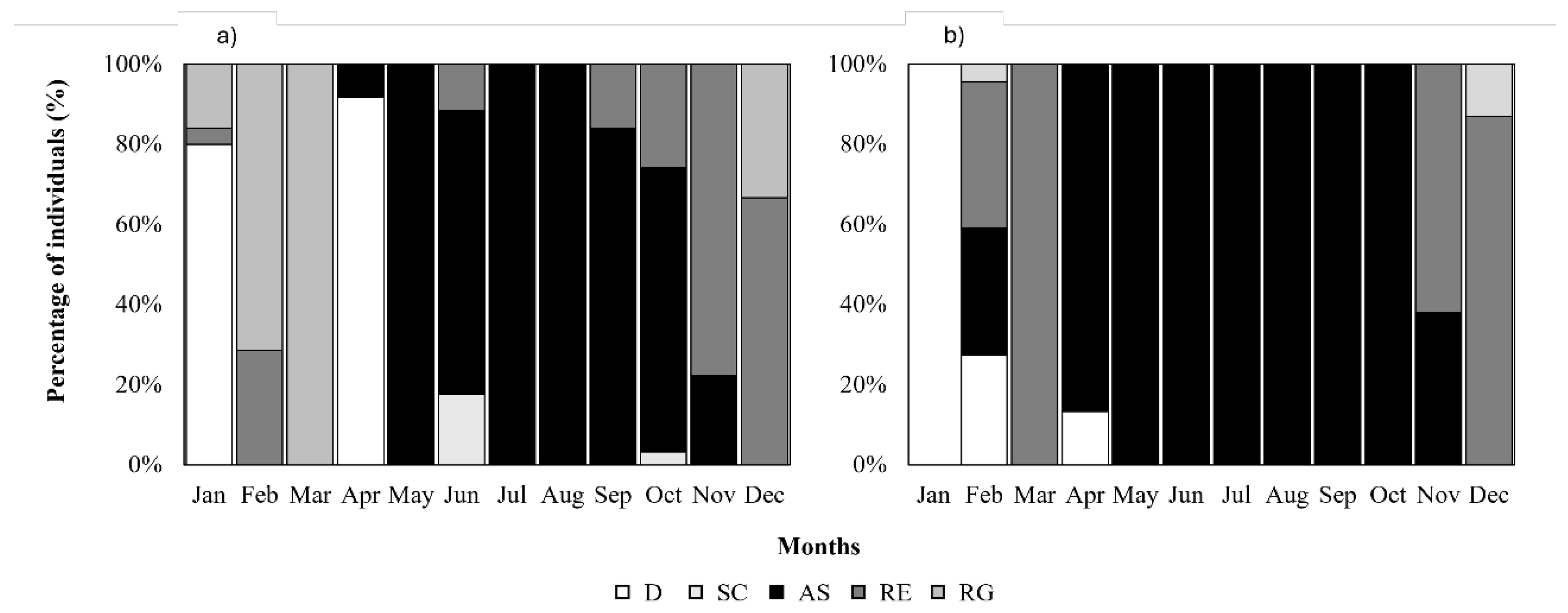
Figure 10.
Gonadosomatic index (GSI), hepatosomatic index (HSI) and Fulton’s condition factor (k) by month and maturation stage, for females (a and c) and males (b and d) of Pagellus acarne from the Portuguese west coast. Significant values are marked with (*). (D) developing; (SC) spawning capable; (AS) actively spawning; (RE) regressing; (RG) regenerating.
Figure 10.
Gonadosomatic index (GSI), hepatosomatic index (HSI) and Fulton’s condition factor (k) by month and maturation stage, for females (a and c) and males (b and d) of Pagellus acarne from the Portuguese west coast. Significant values are marked with (*). (D) developing; (SC) spawning capable; (AS) actively spawning; (RE) regressing; (RG) regenerating.
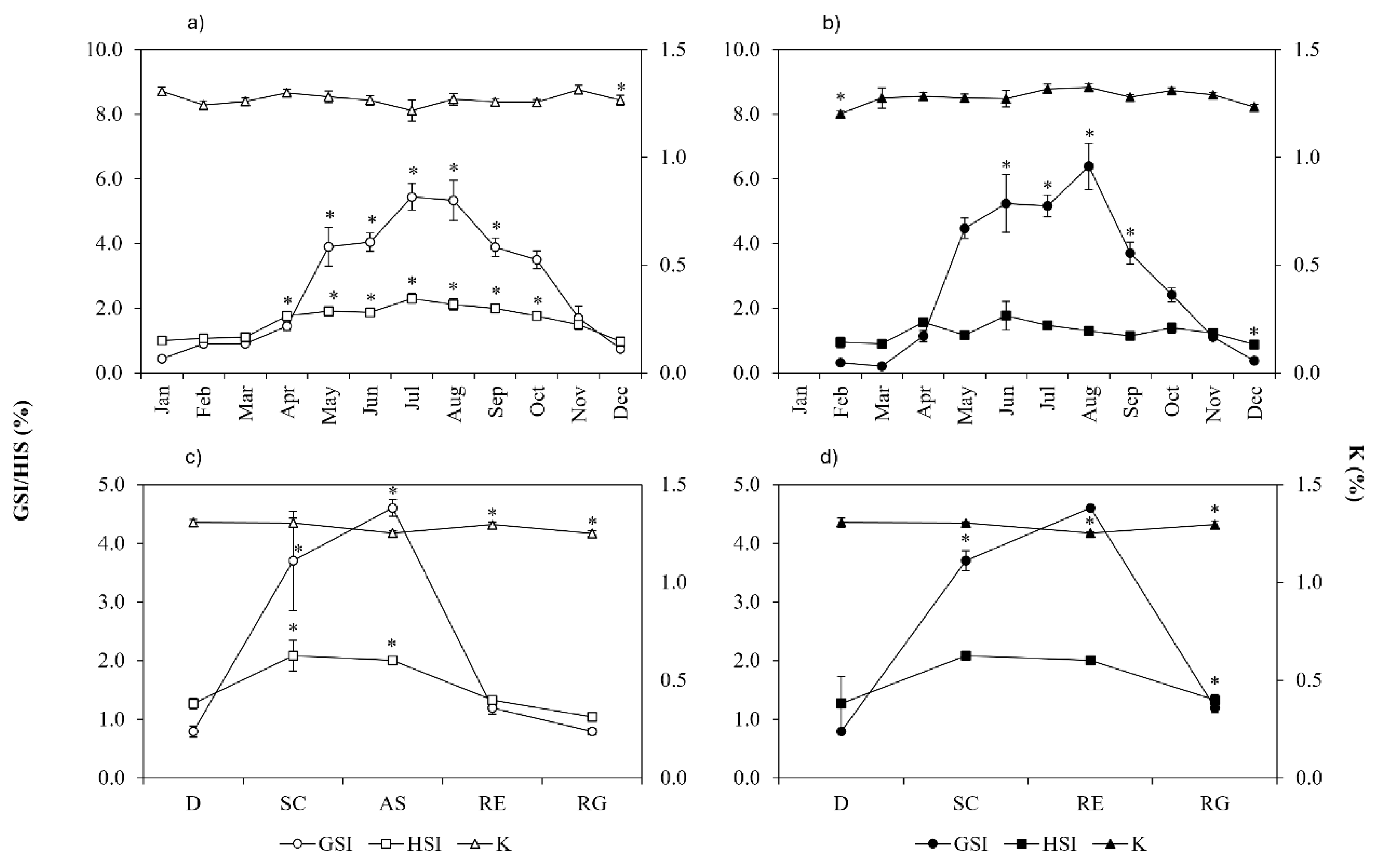

Table 1.
Model equations and number of parameters fitted to length-at-age data for axillary seabream Pagellus acarne.
Table 1.
Model equations and number of parameters fitted to length-at-age data for axillary seabream Pagellus acarne.
| Model | Number of parameters | Equation |
|---|---|---|
| 3p VBGM | 3 | |
| Hyper L∞ | 5 | |
| Hyper k | 5 |
Table 2.
Age-length key obtained from direct reading of otoliths of Pagellus acarne from the Portuguese west coast.
Table 2.
Age-length key obtained from direct reading of otoliths of Pagellus acarne from the Portuguese west coast.
| Length class (cm) | Age (yr) | ||||||||||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | Total | |
| 7-8 | 1 | 1 | |||||||||||||||
| 8-9 | 2 | 2 | |||||||||||||||
| 9-10 | 2 | 2 | |||||||||||||||
| 13-14 | 1 | 1 | |||||||||||||||
| 17-18 | 1 | 1 | |||||||||||||||
| 18-19 | 1 | 1 | |||||||||||||||
| 19-20 | 4 | 4 | |||||||||||||||
| 20-21 | 7 | 2 | 1 | 10 | |||||||||||||
| 21-22 | 3 | 7 | 4 | 14 | |||||||||||||
| 22-23 | 10 | 15 | 3 | 28 | |||||||||||||
| 23-24 | 19 | 16 | 2 | 37 | |||||||||||||
| 24-25 | 7 | 28 | 18 | 1 | 54 | ||||||||||||
| 25-26 | 10 | 34 | 15 | 59 | |||||||||||||
| 26-27 | 19 | 31 | 5 | 1 | 56 | ||||||||||||
| 27-28 | 17 | 21 | 12 | 1 | 51 | ||||||||||||
| 28-29 | 3 | 13 | 21 | 7 | 44 | ||||||||||||
| 29-30 | 8 | 15 | 19 | 5 | 47 | ||||||||||||
| 30-31 | 2 | 11 | 7 | 2 | 22 | ||||||||||||
| 31-32 | 3 | 7 | 2 | 12 | |||||||||||||
| 32-33 | 2 | 3 | 5 | ||||||||||||||
| 33-34 | 1 | 3 | 4 | ||||||||||||||
| 34-35 | 1 | 1 | |||||||||||||||
| N | 5 | 1 | - | 16 | 19 | 46 | 57 | 73 | 67 | 47 | 51 | 41 | 22 | 7 | 3 | 1 | 456 |
| TL mean (cm) | 8.7 | 13.3 | - | 20.1 | 21.8 | 23.0 | 24.2 | 25.4 | 26.4 | 28.0 | 28.5 | 29.6 | 30.8 | 31.6 | 33.5 | 34.2 | |
| TL SD (cm) | 0.6 | - | - | 0.9 | 0.6 | 0.9 | 0.8 | 0.8 | 0.9 | 0.9 | 0.9 | 0.9 | 1.0 | 0.8 | 0.3 | - | |
Table 3.
Summary of von Bertalanffy growth parameters resulting from the three models fitted to axillary seabream Pagellus acarne data.
Table 3.
Summary of von Bertalanffy growth parameters resulting from the three models fitted to axillary seabream Pagellus acarne data.
| Parameters | 3p VBGM | Hyper L∞ | Hyper k |
|---|---|---|---|
| L∞ (cm) | 35.22 | 36.79 | 36.70 |
| K (years-1) | 0.13 | 0.12 | 0.12 |
| t0 (years-1) | -2.78 | -2.26 | -2.24 |
| h | - | -0.16 | -0.24 |
| th (years) | - | 3.36 | 3.40 |
| AIC | 1275.36 | 1189.00 | 1189.45 |
| ∆AIC | 86.36 | 0.00 | 0.45 |
Table 4.
Comparison of age and growth data of Pagellus acarne collected by different authors in the Portuguese west coast. Amax, maximum age; C, combined sexes; F, female; k, growth coefficient; L∞, asymptotic length; LWR, length-weight relationship; M, male; t0, theoretical age when size is zero.
Table 4.
Comparison of age and growth data of Pagellus acarne collected by different authors in the Portuguese west coast. Amax, maximum age; C, combined sexes; F, female; k, growth coefficient; L∞, asymptotic length; LWR, length-weight relationship; M, male; t0, theoretical age when size is zero.
| Reference | Region | LWR | Amax (year) | L∞ (cm) | k (year-1) | t0 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | F | C | M | F | C | M | F | C | M | F | C | |||||||
| Santos et al. (1995) | Portugal South Coast (Atlantic) | Positive allometry | - | - | - | - | - | - | - | - | - | - | - | - | ||||
| Pajuelo & Lorenzo (2000) | Southwest Gran Canaria (Atlantic) | Positive allometry | - | - | 10 | 28.0 | 33.9 | 33.0 | 0.27 | 0.21 | 0.22 | -0.67 | -0.99 | -0.87 | ||||
| Coelho et al. (2005) | Portugal South Coast (Atlantic) | Positive allometry | 13 | 18 | - | 28.8 | 32.3 | 32.1 | 0.29 | 0.18 | 0.18 | -1.47 | -2.56 | -2.91 | ||||
| Abecasis et al. (2008) | Portugal South Coast (Atlantic) | - | - | - | 18 | - | - | 31.8 | - | - | 0.19 | - | - | -2.86 | ||||
| Velasco et al. (2011) | Gulf of Cadiz (Mediterranean) | Positive allometry | - | - | 12 | - | - | 31.7 | - | - | 0.21 | - | - | -1.76 | ||||
| Velasco et al. (2011) | Alboran Sea (Mediterranean) | Positive allometry | - | - | 12 | - | - | 32.1 | - | - | 0.17 | - | - | -2.69 | ||||
| Soykan et al. (2015) | Aegean Sea (Mediterranean) | Isometry | - | - | 6 | - | - | 27.7 | - | - | 0.32 | - | - | - | ||||
| Ílhan (2018) | Aegean Sea (Mediterranean) | Positive allometry | - | - | 4 | 22.5 | 27.8 | 25.6 | 0.34 | 0.2 | 0.25 | -1.55 | -2.35 | -1.94 | ||||
| Di Maio et al. (2020) | Strait of Sicily - Tunisian Coast (Central Mediterranean) | Positive allometry | 6 | 7 | - | - | - | - | - | - | - | - | - | - | ||||
| Bentata-Keddar et al. (2020) | Western Algerian coasts (Western Mediterranean) | - | - | - | - | 28.4 | 29.8 | 30.0 | 0.42 | 0.5 | 0.41 | -0.13 | -0.04 | -0.34 | ||||
| Ali-Basha et al. (2023) | Syrian waters | Positive allometry | - | - | 5 | 23.0 | 21.5 | 19.8 | 0.18 | 0.32 | 0.23 | -0.24 | -0.24 | -0.04 | ||||
Table 5.
Comparison of reproductive data collected by different authors on Pagellus acarne from the Portuguese west coast. F, female; L50, length at first maturity; LSI, length of sexual inversion; M, male.
Table 5.
Comparison of reproductive data collected by different authors on Pagellus acarne from the Portuguese west coast. F, female; L50, length at first maturity; LSI, length of sexual inversion; M, male.
| Reference | Region | Spawning season | L50 (cm) | LSI (cm) | |||
|---|---|---|---|---|---|---|---|
| M | F | ||||||
| Santos et al. (1995) | Portugal South Coast (Atlantic) | May to August | 19.7 | 20.9 | 15-23 | ||
| Pajuelo & Lorenzo (2000) | Southwest Gran Canaria (Atlantic) | October to March | 15.8 | 19.4 | 15-23 | ||
| Coelho et al. (2005) | Portugal South Coast (Atlantic) | May to November | 18.1 | 17.6 | 20-24 | ||
| Abecasis et al. (2008) | Portugal South Coast (Atlantic) | ||||||
| Velasco et al. (2011) | Gulf of Cadiz (Mediterranean) | Autumn, spring and beginning summer (peak April to June) | 18.0 | 21.7 | 23.5 | ||
| Velasco et al. (2011) | Alboran Sea (Mediterranean) | May to October | 17.7 | 20.1 | 21.5 | ||
| Soykan et al. (2015) | Aegean Sea (Mediterranean) | June to September | 13.9 | 14.5 | |||
| Bensahla Talet et al. (2017) | Western Algerian coasts (Western Mediterranean) | April to June andNovember to January | 15.9 | 12.8 | 17.5-26.3 | ||
| Ílhan (2018) | Aegean Sea (Mediterranean) | ||||||
| Di Maio et al. (2020) | Strait of Sicily - Tunisian Coast (Central Mediterranean) | October | 22 | ||||
| Bentata-Keddar et al. (2020) | Western Algerian coasts (Western Mediterranean) | Late spring and autumn | 16.9 | 18.6 | |||
| Ali-Basha et al. (2023) | Syrian waters | ||||||
| This study | Portugal West Coast (Atlantic) | Apr-Oct | 27-31 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



