Submitted:
24 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
Submergence stress challenges direct seeding in rice cultivation. In this study, we identified a heat shock protein, NAL11, with a DnaJ domain, which can regulate the length of rice coleoptiles under flooded conditions. Through bioinformatics analyses, we identified cis-regulatory elements in its promoter, making it responsive to abiotic stresses, such as hypoxia or anoxia. Expression of NAL11 was higher in the basal regions of shoots and coleoptiles during flooding. NAL11 knockout triggered the rapid accumulation of abscisic acid (ABA) and reduction of Gibberellin (GA), stimulating rice coleoptile elongation and contributes to flooding stress management. In addition, NAL11 mutants were found to be more sensitive to ABA treatments. Such knockout lines exhibited enhanced cell elongation for coleoptile extension. Quantitative RT-PCR analysis revealed that NAL11 mediated the gluconeogenic pathway, essential for the energy needed in cell expansion. Furthermore, NAL11 mutants reduced the accumulation of reactive oxygen species (ROS) and malondialdehyde under submerged stress, attributed to an improved antioxidant enzyme system compared to the wild-type. In conclusion, our findings underscore the pivotal role of NAL11 knockout in enhancing the tolerance of rice to submergence stress by elucidating its mechanisms. This insight offers a new strategy for improving resilience against flooding in rice cultivation.
Keywords:
Rice seed germination
; Coleoptile elongation
; Submergence
; NAL11
; Stress tolerance
1. Introduction
Rice (Oryza sativa L.) stands as one of the most pivotal crops cultivated globally, with over half of the world’s population relying on it as a primary staple food [1]. The practice of direct seeding plays a crucial role in rice cultivation and is widely employed in both rainfed and irrigated fields due to its substantial benefits, including reductions in labor, energy consumption, water usage, production costs, and mechanization [2, 3]. However, rice's vulnerability to prolonged flooding poses a significant challenge, leading to oxygen starvation and energy depletion in submerged plants [4]. Enhanced seedling vigor, characterized by the elongation of mesocotyls, coleoptiles, and shoots, is crucial for improving seedling emergence under such conditions [5, 6]. Rice employs an escape strategy to reduce submergence stress during seed germination [7], increasing coleoptile and/or mesocotyl length to improve survival under submerged conditions [8, 9].
Several genetic factors affect rice shoot growth, such as expansin genes, anaerobic metabolic pathways including glycolysis and fermentation, ROS scavenging and phytohormone signaling. Overexpression of the expansin gene OsEXP4 has been shown to promote mesocotyl and coleoptile elongation by cell wall stress relaxation and volumetric extension, a process that is repressed in OsEXP4-antisense plants [10]. In addition, up-regulation of EXPA7 and EXPB12 promotes the elongation of rice coleoptile under hypoxic conditions [11]. In rice, sugar availability has been considered one of the critical factors for tolerance to submergence [12]. Rice seeds can germinate and produce α-amylase enzymes required for starch degradation even without oxygen [13]. Under flooded conditions, anaerobic metabolic pathways, including glycolysis and fermentation, play a crucial role in coleoptile elongation [14]. Under sugar starvation, the transcription factor MYBS1 activates the Ramy3D gene, facilitating starch degradation to provide the necessary energy for subsequent leaf and root development [15]. During germination under submergence, rice gene CIPK15 (calcineurin B-like-interacting protein kinase 15) regulates coleoptile length through a sugar signaling pathway [16]. Alcohol dehydrogenase (ADH) activity in rice coleoptiles is correlated with a deceleration in coleoptile elongation under submergence conditions [17].
Abiotic stresses, like drought, salt, and temperature variations, enhance the production of ROS in plants [16]. However, excessive ROS can lead to oxidative damage to lipids, DNA, and proteins [17]. To mitigate ROS-induced damage, plants have developed an antioxidant system consisting of enzymes, such as catalase (CAT), ascorbate peroxidase (APX), glutathione peroxidase (GPX), glutathione reductase (GR), and glutathione sulfotransferase (GST) [18]. Abscisic acid (ABA) is a pivotal stress hormone that accumulates in response to stress and concurrently associated with a reduction in growth in stressed plants [19]. However, a growing body of evidence suggests that ABA plays a dual role in plant stress responses; while high concentrations inhibit growth, low concentrations can promote it [20, 21]. This balance is crucial in stressed plants, where ABA concentrations are finely tuned through a balance between ABA biosynthesis and catabolism processes [22]. Recent studies have shown that the crosstalk between ABA and ROS in the phytohormone network play important roles in many aspects of plant growth and development, including the response to adversity stresses [23, 24]. For example, it has been reported that alterations in ROS levels can affect ABA biosynthesis and signaling, as well as change ABA sensitivity [25], and ABA can also regulate the expression of ROS producing and scavenging genes [23]. For instance, overexpression of the rice ABA receptor 6 (OsPYL6) can improve drought tolerance by increasing ABA content and improving ROS detoxification, thereby stabilizing membrane [26]. Studies had suggested that ABA could improve oxidase activity and induce stomatal closure to reduce CO2 fixation, thereby inhibit the accumulation of ROS [27].
In addition to ABA, other phytohormones play crucial roles in regulating growth and developmental process and signaling networks involved in plant responses to environmental stresses, including flooding [28]. Gibberellin (GA) is considered essential in regulating the expression of α-amylase genes, which catalyse hydrolytic starch degradation during cereal seed germination in the air. However, under anoxic conditions, starch degradation through the gibberellin-induced α-amylase pathway fails to function properly because oxygen is also required for gibberellin biosynthesis, and rice become gibberellin insensitive under anoxic or hypoxic conditions [12]. Auxin is well known for promoting coleoptile elongation and rapid seedling growth during germination [29], but little is known about its role in rice germination and seedling establishment under submergence. A recent study has shown that auxin biosynthesis and the auxin influx carrier AUX1 regulated the final length of rice coleoptile under submergence [30].
Heat-shock proteins are proteins with molecular chaperone activity, responsible for protein folding, assembly, translocation and degradation in many normal cellular processes, stabilize proteins and membranes [31]. And HSPs may be newly synthesized or otherwise increase in abundance in vivo when plants are subjected to stress. Heat-shock proteins have been reported to play critical roles in stress resistance. For example, transgenic rice plants overexpressing sHSP17.7 showed increased survival under high-temperature conditions [32]. The overexpression of Hsp70 genes correlates positively with the acquisition of thermotolerance [25] and results in enhanced tolerance to salt, water and high-temperature stress in plants. HSP70s reduce heat tolerance, but under high-salt conditions, these proteins enhance seed germination and regulate the developmental transition from seeds to seedlings by suppressing seed-specific gene expression [33]. Expression of Hsp90 in Arabidopsis is developmentally regulated and responds to heat, cold, salt stress, heavy metals, phytohormones and light and dark transitions [34]. A correlation between HSPs and anaerobiosis has been observed in the hearts of turtles and mammals, where constitutive expression of certain HSP genes is associated with increased tolerance to anoxia [35]. Our previous study showed that the DnaJ domain-containing heat shock protein-encoding gene NAL11 regulates rice plant architecture and it is involved in GA metabolism [36]. Nevertheless, the explicit role of NAL11 in coping with abiotic stresses is poorly understood. Therefore, this study aims to Therefore, this study aims to investigate the effect of NAL11 on the elongation of rice coleoptile under flooded conditions. To elucidate the molecular mechanisms by which NAL11 regulates submergence tolerance in rice. To further explore the interactions between NAL11 and gibberellins, auxins and abscisic acid under submergence stress. Our findings unveil a novel mechanism by which HSPs contribute to flood tolerance in rice, which lays a foundation for further investigations into flooding tolerance in rice.
2. Results
2.1. Bioinformatics Characteristics
All amino acid and nucleotide sequences were retrieved and downloaded from the NCBI website. Using the NCBI blast online comparison, 28 homologous proteins were screened in Oryza sativa Japonica, Brachypodium distachyon, Sorghun bicolor, Zea mays, Glycine max, Nymphaea colorata, Arabidopsis, and wild rice (Oryza brachyantha, Oryza glaberrima). To comprehensively characterize NAL11 and its homologous proteins, a phylogenetic tree was constructed for the 28 homologous proteins and the NAL11 protein using MEGA 11.0 software. The constructed evolutionary tree was then merged with the phylogenetic tree of the conserved structural domain elements of the proteins using the TBtools software. Remarkably, in our phylogenetic analysis showed that NAL11 (XP_015645205.1) and XP_040381942.1 clustered together on the same branch (Figure 1). NP_001412649.1 was found to be orthologous to NAL11 in rice, suggesting that NAL11 has undergone a gene duplication event during evolution. Using the online software MEME was used to analyze the 28 homologous proteins, Motif 1 was identified in all 28 members, with conserved motifs 1, 2, and 3 in 22 encoding proteins (Figure 1). All homologues of NAL11, except for NP_001146964.1, contained the DnaJ structural domain (Figure S1). In addition to its conserved nature, NAL11 exhibited high homology with homologues in wild rice. These findings highlight NAL11 as a highly conserved gene throughout evolution. To further investigate potential regulatory mechanisms, we analyzed the 2.0-kb nucleotide sequences upstream of the start codon using Plant CARE. This revealed multiple cis-elements associated with stress response (Figure S2), including the ABRE-motif (ABA response element), the TGACG-motif (MeJA-responsiveness), the GGTCCAT-motif (Auxin-responsiveness), the TCTGTTG-motif (Gibberellin-responsiveness), and other elements critical for endosperm expression, anaerobic induction, anoxic specific regulation, and meristem expression. These results suggest that the NAL11 gene may respond to hypoxia or anoxia during rice growth and development.
2.2. The Expression of NAL11 Was Induced during Seed Germination under Submerged Conditions
Under aerobic conditions, NAL11 exhibited a consistently low level of expression, with a discernible decline at both 12 h and 72 h compared to the baseline at 0 h (Figure 2A). In contrast, during submergence, the gene was induced to express at high levels at 12 h, 24 h, 48 h, and 60 h, compared to the corresponding expression levels under submerged conditions. This finding highlights a substantial upregulation in the expression of NAL11 under submergence conditions compared to that under aerobic conditions. Peak expression occurred at 60 h under submergence, representing an approximately 3-fold increase compared to the expression levels under aerobic conditions. Despite a subsequent decrease in the expression at 72 h under submergence, the level remained significantly elevated compared to the expression level observed under aerobic conditions.
The expression pattern was assessed in transgenic plants carrying the pNAL11::GUS construct (GUS reporter gene driven by the promoter of the NAL11 gene) in the ZH11 background under both submerged and aerobic conditions (Figure 2B). The analysis revealed that NAL11 was predominantly expressed in the parenchyma tissues of the protruding embryo at 24 h and 48 h under aerobic conditions. Whereas at 72 h after seed germination, however, its expression was mainly observed in the growing bud, with minimal presence in the developing radicle. Conversely, under submergence conditions, NAL11 exhibited distinct expression patterns. At 24 h, expression was concentrated in the protruding embryo, and extended to both the embryo and the growing bud at 48 h. Under submerged conditions, the expression in the parenchyma tissues of the embryo was prominent. Due to the limited observation of radicles during seed germination under submergence, NAL11 expression was mainly observed in the elongating coleoptile, with a more pronounced signal closer to the base.
2.3. Knockout Lines Exhibited Longer Coleoptiles during Seed Germination under Submerged Conditions
Mature dry seeds of the WT (ZH11) and two knockout lines were subjected to germination under both air (aerobic) and waterlogged (anoxic) conditions. The coleoptile length of dry seeds on day 4 under submergence exhibited significant differences between the WT and the two knockout lines (Figure S3B). The average coleoptile length of the two knockout lines reached 4.64 cm and 4.55 cm, respectively, which was significantly greater than the 3.82 cm observed in the WT (Figure S3C). Moreover, the surface area of the coleoptile in the knockout lines was significantly greater than that observed in the WT (Figure S3D). However, no evident differences in coleoptile diameter were observed between the WT and knockout lines (Figure S3E). Remarkably, by day 4, both the WT and knockout lines achieved an impressive germination rate of approximately 100% on day 4 (Figure S3A). Additionally, when germinated under aerobic conditions on day 4, no evident differences in bud and root lengths were observed between the WT and knockout lines (Figure S3B). In conclusion, knockout of NAL11 significantly improved rice seed coleoptile growth under submerged conditions.
It has previously demonstrated that coleoptile elongation under anoxic conditions is attributed to cell expansion, and that expansins are likely to be key players in this physiological process [10, 37]. To further understand the role of expansins in this context, we compared the expression patterns of some expansin genes in ZH11 and knockout lines after 48 hours of flooding. Interestingly, only OsEXP7 showed downregulation in the coleoptiles of the knockout lines at 48 h. In contrast, the mRNA levels of OsEXP2, OsEXP4, OsEXP8, OsEXPB11, OsEXPB12, and OsEXPB15 were significantly higher in the coleoptiles of the knockout lines after submerged germination compared to those of the WT at the same time point (Figure 3), with OsEXP4 showing an impressive approximately 4-fold increase (Figure 3). The notable upregulation of the expression of these expansin genes in the knockout lines may provide a molecular basis for the accelerated growth of rice coleoptiles under submerged conditions compared to that in the WT. This suggests that expansins are likely to contribute to the elongation of coleoptiles in rice when exposed to anoxic conditions.
2.4. Knockout of NAL11 Affected Sugar and Energy Pathways under Submerged Conditions
As a crucial enzyme responsible for catalyzing the degradation of starch in cereal seeds, α-AMS plays a pivotal role in various key agronomic traits, including the germination rate [38] and resistance to hypoxia stress [39]. In our study, we aimed to identify the specific α-AMS family member directly involved in the response to endosperm starch degradation under submergence stress. To achieve this, we investigated the expression of rice α-AMS genes in seeds (containing coleoptiles) of ZH11 and knockout lines at 48 h after submergence treatment (Figure 4A). Our findings revealed that the expression levels of RAmy1A and RAmy2A in the knockout lines were significantly lower than those in ZH11. Conversely, the transcript levels of Ramy3A, RamyC, and RAmy3D were significantly higher in the knockout lines than ZH11 and that of RAmy3A in the knockout lines was approximately 5-fold higher in the knockout lines than in ZH11. Interestingly, no evident differences were observed in the expression levels of RAmy3C and RAmy3E between ZH11 and the knockout lines. Furthermore, the α-AMS activity in both knockout lines and WT showed a consistently significant increase from 12 h to 96 h during submerged germination. Compared to the WT, the knockout lines consistently showed higher α-AMS activity at different stages (Figure 4B).
Long-term submergence can lead to substantial carbohydrate consumption, resulting in energy deficiency [40]. Therefore, we investigated the expression patterns of glycolytic pathway genes in rice seeds (containing coleoptiles) under submerged conditions (Figure 4C). CDPK15, MYBS1, Adh1, and PDC1 exhibited distinct submergence-dependent expression. The expression of CDPK15 decreased in the knockout lines, while other key genes associated with energy pathways, including MYBS1, Adh1, and PDC1, showed a significant upregulation in the knockout lines at 48 h after submergence treatment. Concurrently, the enzyme activity of α-AMS exhibited a similar increasing trend mirroring the transcript levels of the energy-synthesizing genes. The observed expression patterns of these genes suggest that the knockout of NAL11 may have a substantial impact on the regulation of α-AMS genes and energy-synthesizing genes, potentially affecting the plant's ability to cope with submergence stress.
2.5. Knockout of NAL11 Affects ROS Levels and the Expression of Some Stress-Related Genes under Submerged Conditions
Given the anaerobic stress experienced by both the knockout lines and the WT during submergence, it is imperative to investigate into the potential physiological changes at different stages, aiming to elucidate he differences in coleoptile length. ROS are important signals that regulate the stress tolerance. Here, we compare the accumulation of hydrogen peroxide (H2O2) and MDA between seeds of the WT and knockout lines after submergence treatment. It shows that the seeds of the knockout lines accumulated more MDA and H2O2 than the WT (Figure 5A-B). Considering that OsRbohA and OsRbohE belong to the NOX family [41], which is key to the production of ROS. We determined the transcription level of these two genes. We found that both were reduced in the knockout lines compared to WT (Figure 5C-D), which was consistent with ROS levels.
In addition, we also determined the activities of ROS-scavenging enzymes (SOD, POD, CAT). The results showed that there was no significant difference between WT and knockout lines in the control group. CAT activity in the knockout lines was evidently lower than that in the WT at the beginning (0 h). Surprisingly, the knockout lines exhibited significantly higher CAT activity than the WT at 12 h, 72 h, and 96 h of seed germination under submergence, with no apparent differences at 24 h and 48 h (Figure S4A). Moreover, the knockout lines showed significantly increased SOD activity from 0 h to 96 h under submerged conditions compared to the WT (Figure S4B). However, no evident difference was observed at 48 h. In contrast, POD activity was increased in both knockout lines and WT after submergence compared to that at the baseline (0 h). From 12 h to 96 h under submergence, no evident differences in POD activity were observed between the knockout lines and WT (Figure S4C). The above results indicated that knockout of NAL11 can enhance rice tolerance to submergence stress by improving ROS scavenging ability.
To further investigate the possible molecular mechanisms of NAL11 in regulating submergence tolerance in plants, we also determined the transcript levels of some well-known stress-responsive genes. These included OsSUB1A and OsNAC9, encoding typical stress-related NAC-type transcription factors (TFs); OsSnRK1A and OsTPP7, encoding trehalose-6-phosphate(T6P) phosphatasegene (OsTPP7) proteins. After submergence treatment for 48 h, compared to WT, the mRNA levels of the above genes were both increased in knockout lines (Figure S5).
2.6. NAL11 Is Involved in the Phytohormone-Mediated Regulatory Pathway
Previous studies have revealed that Plant hormones are signaling compounds that regulate crucial aspects of growth, development and environmental stress responses [41]. The contents of several endogenous phytohormones, including ABA, IAA and GA, were measured in the WT and knockout lines to determine the regulatory pathway in which NAL11 is involved. There were no significant differences in endogenous ABA, GA and IAA concentrations between the WT and knockout lines at 24 h after the initiation of submergence treatment. However, as the submergence time increased, a significant increase in ABA and IAA concentrations was observed in the knockout lines, whereas no significant change was observed in the WT (Figure 6A). The knockout lines showed a significantly higher ABA and IAA levels than the WT at 48 h and 72 h after submergence treatment, while the opposite was true for GA (Figure 6B).
Therefore, we further analyzed the expression of genes related to ABA, GA and IAA synthesis and metabolism in the WT and knockout lines at 24 h, 48 h and 72 h after submergence. Consistent with the increase in the ABA and IAA levels, most of the genes related to ABA and IAA synthesis in the knockout lines were upregulated (Figure S6A-C, Figure S8). Moreover, most of the genes related to ABA metabolism, including several genes of the ABAox family genes, were downregulated simultaneously (Figure S6D-F). To further elucidate the effects of NAL11 knockout on the ABA signaling pathway, we examined the expression levels of key genes involved in ABA regulation and signaling. Since OsDET1, OsbZIP46, and OsbZIP72 have been identified as positive regulators of ABA signaling [42], our study specifically focused on analyzing the expression levels of OsDET1, two bZIP genes (OsbZIP46 and OsbZIP72) involved in ABA regulation, and several ABA receptor genes in both the WT and knockout lines under anaerobic conditions. At 48 h after submergence, the knockout lines exhibited a significant increase in the expression of OsbZIP72, whereas no significant differences were observed in the expression levels of OsDET1 and OsbZIP46 between the WT and knockout lines (Figure S6G). Moreover, OsPYL1, OsPYL2, OsPYL3, OsPYL8, and OsPYL10 were significantly upregulated compared to the WT, whereas OsPYL5 and OsPYL9 were downregulated in the knockout lines. In contrast, no significant changes were observed in the transcript levels of OsPYL4 and OsPYL7 in the WT and knockout lines (Figure S6H). At 72 h after submergence, most of the genes related to ABA signaling were upregulated (Figure S6I). This suggests that knockout of NAL11 has a significant effect on the expression of genes related to ABA biosynthesis, catabolism and signal transduction processes. Interestingly, the expression of most GA-related genes in the NAL11 knockout lines showed the opposite trend compared to ABA (Figure S7), indicating reduced GA activity. We therefore speculate that the longer coleoptiles of the NAL11 knockout lines under submergence could be attributed to the upregulated expression of ABA- and IAA-related genes and down-regulated expression of GA-related genes. Furthermore, we also analyzed the expression of genes related to auxin biosynthesis in one of the knockout lines. The expression of YUCCA2, YUCCA3 and YUCCA6, which are involved in auxin biosynthesis, was significantly increased in the knockout line compared to the WT under submerged conditions (Figure S8A-C). Besides, the expression pattern of TAA1, another phytoalexin biosynthesis gene, was also similar to that of these genes (Figure S8D). As a result, the endogenous IAA levels were significantly increased (Figure 6C). It suggests that under submerged conditions, NAL11 knockout may prolong the coleoptile by increasing the accumulation of auxin in rice seeds. Taken together, NAL11 is involved in the ABA, GA and auxin pathways to improve tolerance to submergence stress tolerance in rice. This intricate regulatory network contributes to the improved tolerance of the knockout lines to submergence stress during the germination stage of rice seeds.
2.7. Knockout Lines Are Sensitive to ABA
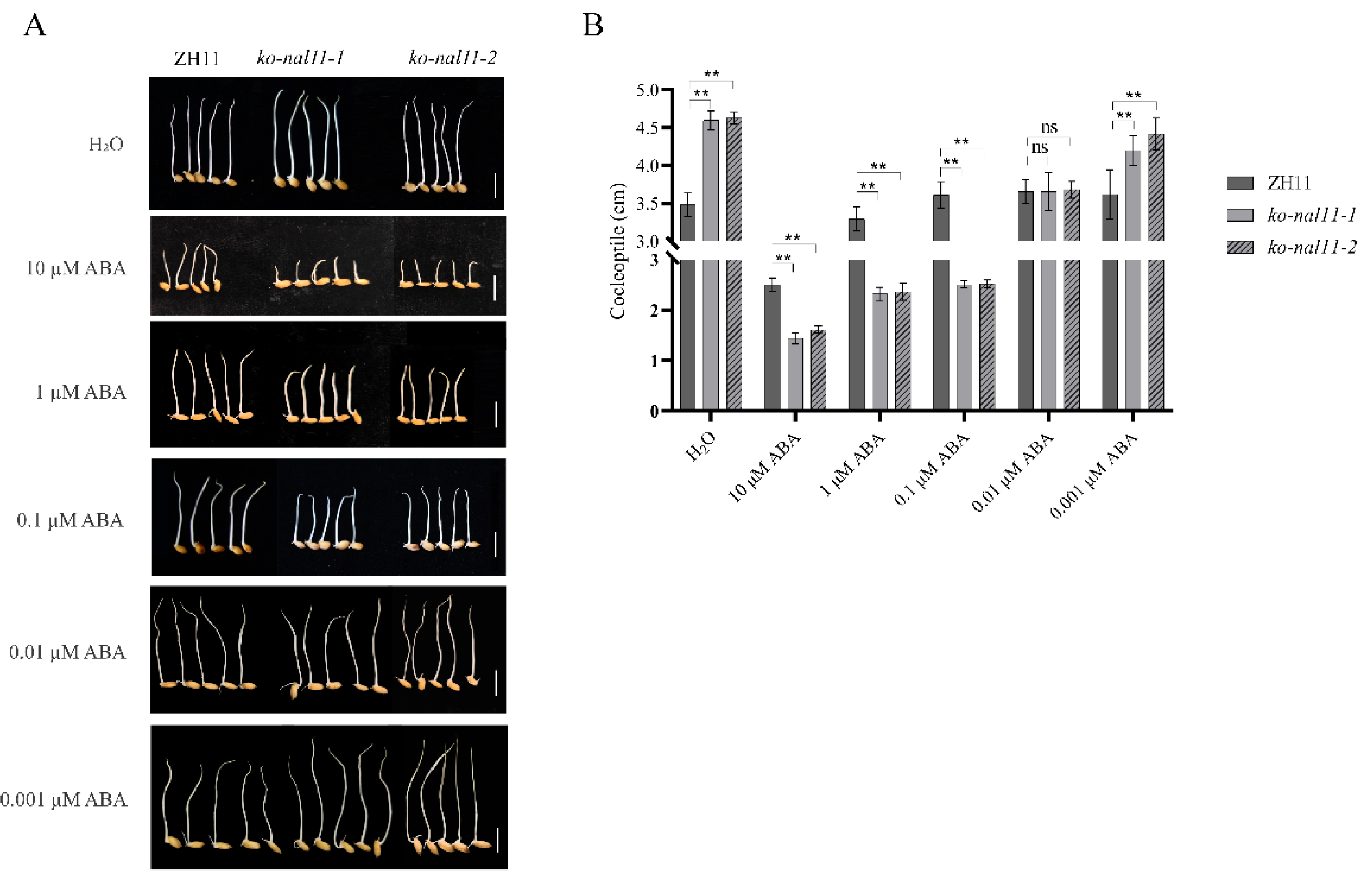
Crosstalk between ABA and ROS in the phytohormone network has been shown to be involved in the regulation of plant stress tolerance [27], and exogenous ABA treatment will decrease the level of ROS in rice seed germination embryos [43]. Based on these studies, we conducted the following experiment to assess the sensitivity of NAL11 to ABA. We conducted a statistical analysis of the growth status of the WT and knockout lines on day 4 after treatment with different concentrations of exogenous ABA (0 µM, 0.001 µM, 0.01 µM, 0.1 µM, 1 µM, and 10 µM). Under submerged conditions, the knockout lines exhibited longer coleoptiles compared to the WT in the absence of ABA treatment. However, the coleoptile length of the knockout lines was significantly reduced compared to that of the WT when treated with higher concentrations (10 µM, 1 µM, or 0.1 µM) of exogenous ABA (Figure 7A-B). Remarkably, at an ABA concentration of 0.01 µM, no significant difference in coleoptile length was observed between the WT and knockout lines. Interestingly, when treated with a lower concentration of 0.001 µM ABA, the coleoptile length of the knockout lines was significantly greater than that of the WT. These results suggest that high concentrations of exogenous ABA inhibit coleoptile elongation in the knockout lines, while low concentrations of exogenous ABA promote coleoptile elongation in the knockout lines. This suggests that the knockout of NAL11 may enhance sensitivity to ABA.
3. Discussion
The length and elongation rate of the coleoptile and/or mesocotyl are the crucial developmental traits that determines deterring the success of direct seeding in many cereal crops. Mesocotyl elongation responds to abiotic stresses, such as deep sowing drought, submergence, chilling, and salinity [9]. In the case of rice submerged seed germination, the long and rapidly elongating coleoptiles may promote submergence tolerance by providing oxygen when they reach to the water surface [10]. Knowledge on the genetic factors and molecular mechanisms of coleoptile development under submerged conditions remains limited. Here, we found that knockout of NAL11 plays a key role in determining rice coleoptile length under submergence. NAL11 integrates ABA signaling and is involved in coleoptile elongation by regulating the synthesis and metabolism of ABA, GA and IAA, the expression of expansin genes, and the catalysis of glycolysis. Thus, our results contribute to the understanding of the escape strategy of submergence tolerance in rice, which may be helpful in improving submergence tolerance in other cereal crops.
HSPs are known to respond to a variety of abiotic stresses, including deep-seeded drought, submergence, cold, and salinity, and they play a protective role in plants against these stresses [32]. As a member of this family, NAL11 was highly induced under submergence (Figure 2A). Based on the analysis of the tissue-specific gene expression pattern, the expression of NAL11 was primarily observed in the elongating coleoptile, with a more pronounced signal closer to the base (Figure 2B). In addition, the expression patterns of the expansin genes were significantly altered in the knockout lines. In fact, the overexpression of the expansin gene OsEXP4 has been shown to promote mesocotyl and coleoptile elongation through cell wall stress relaxation and volume expansion, a process that is repressed in OsEXP4-antisense plants [10]. Consistent with these results, the expression of the vast majority of expansin genes was significantly upregulated in the NAL11 knockout lines compared to the WT. In rice, coleoptile elongation during submergence is predominantly driven by cell elongation [15]. Thus, NAL11 may play a key role in coleoptile elongation activity by regulating expansins to maintain cell elongation and ultimately regulate submergence tolerance in rice at the seed germination stage. Several response-related cis-elements were identified in the promoter of the NAL11 gene (Figure S2), such as ABRE-motif (ABA response element), GGTCCAT-motif (Auxin response element) and TCTGTTG-motif (Gibberellin response element), suggesting that NAL11 may be directly or indirectly involved in the regulation of biosynthesis, metabolism, and signaling of ABA, GA, and Auxin, which was confirmed in our study (Figure 6, S6-8). Therefore, we first investigated and elucidated the regulatory role of NAL11, a DnaJ domain-containing HSP gene, in rice coleoptile elongation.
ROS function as key signal transduction molecules in plants. However, their excessive accumulation can lead to irreversible cell damage [44]. Previous studies have shown that plants adapt to abiotic stress by regulating ROS metabolism [45]. For example, overexpression of a MAPK kinase gene, DSM1, in rice improves drought tolerance in rice at the seedling stage by regulating ROS clearance [46]. Enhancing antioxidant activity to mitigate ROS overproduction is a critical strategy for improving stress tolerance in plants [47]. Recent studies have indicated that increased activities of POD, SOD, and CAT play a pivotal role in improving ROS clearance, thereby maintaining ROS homeostasis and consequently enhancing tolerance to environmental [48, 49]. Our result showed that knockout of NAL11 could reduce the over-accumulation of ROS under submergence stress conditions, which was further confirmed by the content of H2O2 and the transcription level of several genes. Moreover, NAL11 knockout enhanced the activity of ROS scavenging enzymes, namely CAT and SOD, in rice under submergence stress, although no significant enhancement in POD activity was observed (Figure S4). MDA, which is an indicator of the degree of ROS-induced cell membrane damage and lipid peroxidation. The result that the MDA content of the knockout lines was reduced compared to the WT, which was consistent with the phenotype under submergence, suggests that knocking out NAL11 could reduce lipid peroxidation in plants [50]. Therefore, we speculated that knockout of NAL11 could increase ROS-scavenging activity and lead to reduced ROS accumulation, thereby alleviating the damage caused by excessive ROS induced by submergence stress and further enhancing submergence tolerance in rice. In conclusion, the NAL11 knockout that confers submergence tolerance appears to be associated with enhanced antioxidant capacity.
The submergence tolerance of rice seedlings is intricately linked to the sustenance of energy supply, necessitating elevated levels of carbohydrates [51]. During germination under anoxic conditions, rice seeds can degrade starch into readily fermentable carbohydrates, facilitating the generation of energy essential for the growth of germinating embryos [40, 52]. The successful germination of rice seeds and the establishment of early seedlings under submergence conditions are contingent on the ability to induce α-AMS and glycolytic enzymes in low-oxygen environments. Specifically, in the present study, RAmy3A, RAmyC, and RAmy3D exhibited robust upregulation under submergence conditions, indicating an accelerated starch hydrolysis process. Additionally, knockout lines showed significantly higher amylase activity during submergence compared to WT (Figure 4B). The elevated amylase activity is consistent with the longer coleoptile in the knockout lines than in WT, a relationship further illustrated by strong positive correlations coleoptile surface area during submergence (Figure S3C-D). Under submergence conditions, RAmy1A expression was less pronounced in the knockout lines (Figure 4A), in agreement with previous findings by Hwang et al. [40]. Although the expression of RAmy1A and RAmy2A was reduced after submergence (Figure 4A), this did not affect the rate of band starch hydrolysis, as the isozymes they encode are not dominant during hypoxic sprouting [11, 53]. α-AMS 3 emerges as a major player, its mRNA accounting for approximately 60% of the total mRNA of amylase genes in glucose-starved rice cells [54]. In this study, high expression of α-AMS 3 accelerated the hydrolysis of starch (Figure 4A), thereby providing the energy required for the germination process and sustaining the subsequent alcoholic fermentation process. Previous studies have shown that submergence triggers sugar starvation and induces mRNA accumulation of calcineurin B-like (CBL) protein-interacting protein kinase 15 (CIPK15), thereby enhancing the accumulation of SnRK1A proteins. These two proteins interact and induce the MYBS1 transcription factor, subsequently activating the expression of starvation-induced α-amylase gene, αAmy3/RAmy3D [55]. As expected, the expression of CIPK15 and MYBS1 was significantly higher in the knockout lines compared to WT (Figure 4C).
Under anoxia/hypoxia, the metabolic shift in carbohydrate catabolism from aerobic to anaerobic pathways (alcoholic fermentation) results in the generation of ATP to support growth and maintenance processes [5]. Despite being 18-fold less efficient for ATP production than in aerobic respiration, alcoholic fermentation remains one of the most crucial mechanisms for alleviating the adverse effects of reduced ATP supply during oxygen deficiency [56, 57]. It has been previously demonstrated that the catalysis of the alcoholic fermentation is facilitated by enzymes such as pyruvate decarboxylase (PDC) and ADH, supporting glycolysis and ATP synthesis by recycling nicotinamide adenine dinucleotide (NAD). Our results showed that the expression of PDC and ADH in the knockout plants was significantly higher than that of the WT after 48 h of submergence (Figure 4C)., indicating a pivotal role for this metabolic pathway in coping with submergence conditions during germination. This observation is consistent with previous studies showing enhanced submergence tolerance in transgenic plants overexpressing PDC [58]. Moreover, it remains unclear whether sugar could affect the accumulation of endogenous free phytohormones to influence submergence tolerance and seedling establishment in rice.
Submergence stress causes hypoxia in plants and in this situation, the expression of genes is changed (be up- or down-regulated) and these genes encode a variety of proteins involved in physiological and biochemical processes and, in addition to the SnRK1A proteins, several regulatory factors, such as transcription factors, are also involved [59]. Under sugar starvation conditions, SnRK1A is an important mediator in the sugar signaling cascade response, acts upstream of MYBS1 and αAmy3 SRC interactions, and plays a key regulatory role in rice seed germination and seedling growth. Many transcription factors play critical roles in regulating the stress response in plants, including SUB1A and OsNAC9, which improve submergence tolerance when overexpressed [60, 61]. In our result, both were up-regulated in transcription level in the knockout lines compared to WT. Meanwhile, it has been shown that OsTPP7 is involved in T6P metabolism and catalyzes the conversion of T6P to trehalose, thereby allowing increased starch mobilization in the form of easily fermentable sugar, which ultimately enhances coleoptile elongation and embryo germination [62]. Similarly, in our research, better developed coleoptiles and higher levels of OsTPP7 expression were observed in the knockout lines (Figure S5). This suggests, therefore, that knockout of NAL11 could improve submergence tolerance in rice by affecting the transcript levels of these stress-responsive genes.
The phytohormone auxin has long been known to be important in stimulating coleoptile elongation and rapid seedling growth in the air [63], but little is known about its role in influencing t rice coleoptile elongation under water. However, the underlying mechanism of seed germination under anaerobic conditions is not fully understood. To understand the role of plant hormones in rice under air and hypoxic conditions, we carried out some analyses on ABA, GA and auxin and related genes under submerged conditions. ABA is a well-documented stress hormone that accumulates in response to stress [19]. Consequently, ABA levels in the NAL11 knockout lines gradually increased after 48 h of submergence treatment. We also found that ABA and GA levels were reversed at 48 h and 72 h after submergence treatment regardless of in both knockout lines and WT (Figure 6). It has previously been shown that the vivipary phenotype in maize kernels due to ABA deficiency can be reversed through inhibition of GA synthesis, demonstrating the role of GA in antagonizing the action of ABA. In this study, this phenomenon was well explained by the expression of genes related to the biosynthesis and metabolism of ABA and GA (Figure S6-7). These results suggest that NAL11 negatively regulates the antagonistic effects of ABA and GA by mediating the activities of a number of enzymes involved in ABA- and GA-related biosynthesis, which in turn come to regulate the elongation of the coleoptile under submergence. Thus, we can reasonably infer that the antagonistic regulation of GA and ABA metabolism mainly occurs by activating and repressing the opposing metabolic genes (NCED/GA2ox or ABA8ox/GA3ox family) to maintain a hormonal balance during plant growth and development and to respond to environmental cues. Auxin is well known for promoting coleoptile elongation and rapid seedling growth during germination [61], but little is known about its role in rice germination and seedling establishment under submergence. A recent report has also demonstrated that auxin is required for rice seed germination under submergence. The results indicate that auxin availability and transport play a critical role in determining the final coleoptile length in Japonica rice [64]. In submerged seeds, the knockout lines had higher levels of endogenous auxin than WT, which is consistent with the fact that the expression of four auxin biosynthesis genes, YUCCA2, YUCCA3, YUCCA6 and TAA1, was significantly increased in seedlings of hypoxic knockout lines in comparison to WT (Figure S8), consistent with the observed phenotype (Figure S3) and the previous studies. Consequently, it was postulated that the knockout of NAL11 could affect the transcription levels of related synthetic genes, including YUCCA2, YUCCA3, YUCCA6 and TAA1, which could subsequently alter auxin levels and further enhance submergence tolerance. Whether or not the auxin transport or distribution in hypoxic rice seedlings would be influenced by excessive accumulation of endogenous free IAA and affect the submergence tolerance remains to be investigated.
ABA is known for its dual function of inhibiting growth at high concentrations and promoting growth at low concentrations [23, 65]. In our study, ABA levels in the knockout lines significantly exceeded those in the WT at 48 h and 72 h under submergence (Figure 6A). This phenomenon was further confirmed by experiments using low concentrations (0.001 µM) of exogenous ABA treatment, which stimulated coleoptile growth. However, when exposed to higher concentrations of exogenous ABA, coleoptile growth was inhibited. When the optimal concentration for plant growth was exceeded, the addition of exogenous ABA led to the inhibition of coleoptile growth in both the knockout lines and the WT when treated with 0.1 µM and 1.0 µM ABA (Figure 7A), in agreement with which is consistent with the previous studies [66]. The increased sensitivity of the knockout lines to exogenous ABA (Figure 7A-B) implies an enhanced responsiveness of these lines to ABA. In the ABA signaling pathway, OsPYL/RCAR serves as a crucial functional ABA receptor that orchestrates ABA-dependent gene expression in rice [1]. Overexpression of bZIP72 was associated with augmented coleoptile elongation and increased ABA sensitivity in submerged seeds [67, 68]. In this study, the expression levels of OsPYL1, OsPYL2, OsPYL3, OsPYL8, OsPYL10, and OsbZIP72 were significantly upregulated in the knockout lines (Figure S6G-I). This strongly suggests that showing that NAL11 enhances rice tolerance to submergence stress by regulating the expression of ABA-mediated genes. These results suggest that NAL11 plays a crucial role in submergence stress via the ABA-mediated regulatory pathway.
4. Materials and methods
4.1. Bioinformatics Analysis of NAL11
The information and sequences of NAL11 homologues were retrieved from the NCBI database (http://www.ncbi.nlm.nih.gov). Utilizing ClustalW with default parameters, multiple sequence alignments were performed on protein sequences, followed by manual adjustments. Subsequently, a phylogenetic tree was constructed with aligned protein sequences using MEGA11 software, employing the neighbor-joining (NJ) method. Bootstrap values, derived from 1000 iterations, were calculated to assess the robustness of the tree [69]. For the identification of conserved motifs within protein sequences, online MEME (http://meme-suite.org/) was utilized with default parameters. Putative cis-acting elements were identified by analyzing the 2000-bp promoter region sequences of these paralogous genes, obtained from the NCBI database, using PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/). TBtools software facilitated the visualization of the phylogenetic tree, conserved motifs, gene structure, domains, and cis-acting elements in promoters [70].
4.2. Plant Materials and Growth Conditions
Zhonghua 11 (ZH11) is a japonica rice (Oryza sativa L.) variety used as the WT plant and the recipient for genetic transformation in this study. The T1 and T2 generation knockout lines were consecutively assayed for the target gene and the hygromycin resistance gene. Two stable T3-generation knockout lines (ko-nal11-1 and ko-nal11-2) without hygromycin were selected for follow-up studies. All plants were cultivated under natural conditions in the experimental field at South China Agricultural University (Guangzhou, Guangdong, China, 23.13°N,113.27°E). The WT and knockout lines without hygromycin were planted in a randomized block design. Each plot consisted of six rows with six plants per row at a planting interval of 20 cm × 20 cm. Field management was in accordance with normal agricultural practices.
4.3. Construction of Transgenic Plants
To achieve the knockout lines of NAL11, target sites were designed through the online tool Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-GE/targetDesign (http://skl.scau.edu.cn/) [71]. Genome-targeting constructs were then prepared using the pRGEB32 vector [72]. Pre-cultured seeds were immersed in the Agrobacterium suspension by gently inverting the tube for 1.5 min, then blotted dry with a sterilized filter paper to remove excess bacteria. These seeds were transferred onto a sterilized filter paper (9-cm diameter) that had been moistened with 0.5 mL of AAM medium placed on 2N6-AS medium solidified with 0.4% Gelrite. After 3 days of co-cultivation at 25°C in the dark, seeds were washed five times in sterile water and then washed once in sterile water containing 500 mg l−1 carbenicillin (Wako Pure Chemicals, Osaka, Japan) to remove Agrobacterium. The seeds were rapidly blotted dry on a sterilized filter paper and cultured on N6D medium containing 50 mg l−1 hygromycin and 400 mg l−1 carbenicillin under continuous light at 32°C for 2 weeks. Proliferating calli emerging from the scutellum were transferred to RE-III medium. Plantlets emerging from the calli were transferred to HF medium to induce roots. For validation, the target sites of T1 and T2 generation plants were sequenced and analyzed using CRISPR-GE/DSDecodeM [73].
To obtain the expression profile of NAL11, the promoter sequence of approximately 2,000 bp was identified from Ensemble Plants (LOC_Os07g09450) and amplified from rice genomic DNA. The pCAMBIA1305 vector was double digested with SacⅠ and BglⅡ endonucleases, followed by recombination to generate the pNAL11::GUS vector. The pNAL11::GUS vector was introduced into Agrobacterium tumefaciens strain EHA105, and the transformation procedure described above was used to generate transgenic rice lines. Transgenic plants carrying the pNAL11::GUS construct were screened and confirmed by PCR. All primers used in the construction process are detailed in Supplementary Table 1.
4.4. Evaluation of Germination Rate and Coleoptile Length
Seeds of each line were grown in the field, and their seeds were harvested 45 days after heading, air dried, and stored at 42 °C for 7 days to break dormancy. Three independent biological replicates of 30 seeds per replicate were then sterilized with 1.5% (v/v) sodium hypochlorite and subsequently incubated in a 9-cm diameter Petri dish. Seeds were considered germinated when the white embryo protrusion was visible, at which point the germination percentage was then calculated. For the anoxic experiments, five seeds were placed in a test tube (diameter: 2.7 cm; height: 11.7 cm) filled with distilled water to simulate anaerobic conditions [74]. All germination experiments were performed in a controlled environment at 28°C under a 12 h/12 h light/dark cycle. After 4 days, a WinRHIZO root image analysis system (Regent Instruments Inc., Québec, Canada) was used to measure the coleoptile length (CL), coleoptile surface area (CSA), and coleoptile diameter (CD). Three biological replicates were tested.
4.5. Germination Test by Exogenous Application of ABA
A total of 15 seeds of the WT and knockout line (nal11) were anaerobically incubated at 28°C with a gradient concentration of ABA solution as treatment and with pure water as control, respectively. The concentration of the ABA solution was adjusted to 0.001, 0.01, 0.1, 0, 1, and 10 µM, respectively. Photographs were taken and coleoptile lengths were measured after 4 d of incubation.
4.6. Measurement of Endogenous ABA, GA and IAA Levels
To measure the endogenous levels of ABA, GA and IAA, the seeds were prepared at 24, 48 and 72 HAI (hours after imbibition) under anaerobic stress. Liquid nitrogen-frozen germinated seeds (50 mg fresh weight) were ground to powder and extracted with the traction method (methanol/water/formic acid = 15:4:1, V/V/V). The extracts were vortexed and centrifuged at 4694 × g at 4°C for 10 min. The supernatants were dried by evaporation under the flow of nitrogen gas at room temperature, then dissolved in 200 μL of methanol. The sample extracts were analyzed using an LC-ESI-MS/MS system (HPLC, Shim-pack UFLC SHIMADZU CBM30A system; MS, Applied Biosystems 6500 Triple), and the data were analyzed by Zoonbio Biotechnology Co. Ltd. Nanjing, China. Three replicates of each assay were performed [75].
4.7. Analysis of Physiological Parameters Related to Submergence Stress
Ten seeds each from the WT and knockout lines were anaerobically incubated at 28°C, respectively. The treatment time was set at 0 h, 12 h, 24 h, 48 h, 72 h and 96 h, respectively, and the analysis of catalase (CAT), superoxide dismutase (SOD), peroxidase (POD) activity and content of malondialdehyde (MDA) and H2O2 content was performed as previous described [76] and adjusted. For all assays, the data for each time point represent the average of at least three biological replicates. Calculations were carried out according to the equations recommended by the manufacturer (Nanjing Jiancheng Bioengineering Institute).
4.8. RNA Extraction and Analysis of Gene Expression
Total RNA from germinated seeds was extracted using TRIzol reagent (R401-01, Vazyme). The first-strand cDNA was synthesized from 600 ng total RNA using a reverse transcription kit (R133-01, Vazyme). The qRT-PCR reaction was performed using ChamQ Universal SYBR qPCR Master Mix (Q711-03, Vazyme) on an ABI Step One Plus Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) [77]. Normalized transcript levels were calculated using the comparative 2-ΔΔCT method [78],with the OsActin serving as an internal control. Five biological replicates were performed. All primers used for qRT-PCR are listed in Table S1.
The developing seeds of pNAL11::GUS transgenic plants were collected and detected according to the previous method [79]. The developing seeds of pNAL11::GUS transgenic plants were collected and incubated in GUS staining buffer (750 μg·mL-1 X-gluc, 10 mM EDTA, 3 mM K3Fe(CN)6, 100 mM NaPO4 pH 7, and 0.1% Nonidet-P40) at 37 °C for 6 h. The samples were then transferred to 70% ethanol to remove chlorophyll. Finally, photographs were taken using a ZEISS stereomicroscope.
4.9. Statistical Analysis
Statistical analysis of the data was performed using the Prism 8.3.0 software package. Student’s t-tests were employed, with statistical significance set at P < 0.05. Significant differences between means are indicated by asterisks (*P < 0.05, **P < 0.01, ***P < 0.001). All data are presented as mean ± standard deviation (SD), with “n” representing the sample size.
5. Conclusion
Bioinformatic analysis reveals that the NAL11 gene belongs to the family of HSPs containing the DnaJ structural domain. Knockout of NAL11, which regulates the expression of genes involved in ABA, GA and auxin biosynthesis, catabolism and signaling pathways, increases the activities of many antioxidant defence enzymes to maintain ROS balance and improve tolerance to submergence stress in rice. Additionally, knockout of NAL11 significantly increased sugar metabolism and the expression of expansin genes in rice seeds, which promoted the elongation of rice coleoptiles. Taken together, these molecular and physiological changes resulted in improved tolerance to submergence stress in rice. These findings not only deepen our understanding of the function of NAL11, but also provide a solid foundation for future research aimed at improving crop tolerance to flooding, potentially leading to more resilient agricultural practices in flood-prone areas.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author information
Z.Z. and Y.X. have contributed equally to this work. W.X. and T.G. raised the project conception and designed the research. Z.Z. and Y.X. carried out most of the experiments and analyzed data. Z.Z. and Y.X. wrote, the manuscript. W.X. and T.G. provided laboratory support. Z.Z., M.T., C.C., J.Z. and J.L. analyzed the data. All authors have read and approved to the published version of the manuscript. Corresponding authors Correspondence to T.G. and W.X.
Funding
This work was supported by the Natural Science Foundation of China (Grant no. 31872885), the Guangdong Basic and Applied Basic Research Foundation (Grant no. 2021A1515010410), and the Seed Industry Revitalization Project of Guangdong Provincial Rural Revitalization Strategy Special Fund (2022-NPY-00-016).
Data Availability Statement
All of the datasets are included within the article and its additional files.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Long, Q.; Qiu, S.; Man, J.; Ren, D.; Xu, N.; Luo, R. OsAAI1 Increases Rice Yield and Drought Tolerance Dependent on ABA-Mediated Regulatory and ROS Scavenging Pathway. Rice 2023, 16, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, G.; Cui, Z.; Kong, X.; Yu, X.; Gui, R.; Han, Y.; Li, Z.; Lang, H.; Hua, Y.; et al. Regain flood adaptation in rice through a 14-3-3 protein OsGF14h. Nat. Commun. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Perata, P.; Alpi, A. Plant responses to anaerobiosis. Plant Sci. 1993, 93, 1–17. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L. Flooding Stress: Acclimations and Genetic Diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guo, Z.; Luo, L.; Gao, Q.; Xiao, W.; Wang, J.; Wang, H.; Chen, Z.; Guo, T. Identification of QTL and candidate genes involved in early seedling growth in rice via high-density genetic mapping and RNA-seq. Crop. J. 2020, 9, 360–371. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Lee, S.C.; Brinton, E. Waterproofing Crops: Effective Flooding Survival Strategies. Plant Physiol. 2012, 160, 1698–1709. [Google Scholar] [CrossRef] [PubMed]
- Pucciariello, C. Molecular Mechanisms Supporting Rice Germination and Coleoptile Elongation under Low Oxygen. Plants 2020, 9, 1037. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Lu, X.; Liu, H.; Zhao, Q.; Ye, G. Mesocotyl elongation, an essential trait for dry-seeded rice (Oryza sativa L.): a review of physiological and genetic basis. Planta 2019, 251, 27. [Google Scholar] [CrossRef]
- Choi, D.; Lee, Y.; Cho, H.-T.; Kende, H.; Ht, C. Regulation of Expansin Gene Expression Affects Growth and Development in Transgenic Rice Plants. Plant Cell 2003, 15, 1386–1398. [Google Scholar] [CrossRef]
- Lasanthi-Kudahettige, R.; Magneschi, L.; Loreti, E.; Gonzali, S.; Licausi, F.; Novi, G.; Beretta, O.; Vitulli, F.; Alpi, A.; Perata, P. Transcript Profiling of the Anoxic Rice Coleoptile. Plant Physiol. 2007, 144, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Loreti, E.; Yamaguchi, J.; Alpi, A.; Perata, P. Sugar modulation of α-amylase genes under anoxia. Annals of Botany 2003, 91, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Perata, P.; Geshi, N.; Yamaguchi, J.; Akazawa, T. Effet of anoxia on the induction of α-amylase in cereal seeds. Planta 1993, 191, 402–408. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kugimiya, T. Preferential Induction of Alcohol Dehydrogenase in Coleoptiles of Rice Seedlings Germinated in Submergence Condition. Biol. Plant. 2003, 46, 153–155. [Google Scholar] [CrossRef]
- Li, X.; Chen, L.; Forde, B.G.; Davies, W.J. The Biphasic Root Growth Response to Abscisic Acid in Arabidopsis Involves Interaction with Ethylene and Auxin Signalling Pathways. Front. Plant Sci. 2017, 8, 1493–1493. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated Responses to Oxygen and Sugar Deficiency Allow Rice Seedlings to Tolerate Flooding. Sci. Signal. 2009, 2, ra61–ra61. [Google Scholar] [CrossRef]
- Takahashi, H.; Saika, H.; Matsumura, H.; Nagamura, Y.; Tsutsumi, N.; Nishizawa, N.K.; Nakazono, M. Cell division and cell elongation in the coleoptile of rice alcohol dehydrogenase 1-deficient mutant are reduced under complete submergence. Ann. Bot. 2011, 108, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Xu, N.; Chu, Y.; Chen, H.; Li, X.; Wu, Q.; Jin, L.; Wang, G.; Huang, J. Rice transcription factor OsMADS25 modulates root growth and confers salinity tolerance via the ABA–mediated regulatory pathway and ROS scavenging. PLOS Genet. 2018, 14, e1007662. [Google Scholar] [CrossRef]
- Brookbank, B.P.; Patel, J.; Gazzarrini, S.; Nambara, E. Role of Basal ABA in Plant Growth and Development. Genes 2021, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cai, Z.; Wang, X. The primary signaling outputs of brassinosteroids are regulated by abscisic acid signaling. Proc. Natl. Acad. Sci. 2009, 106, 4543–4548. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Liu, J.; Weng, H.; Yuan, X.; Xiao, W.; Wang, H. A long noncoding RNA derived from lncRNA-mRNA networks modulates eeed vigor. Int. J. Mol. Sci. 2022, 23, 9472. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhu, Z.; Chern, M.; Yin, J.; Yang, C.; Ran, L.; Cheng, M.; He, M.; Wang, K.; Wang, J.; et al. A Natural Allele of a Transcription Factor in Rice Confers Broad-Spectrum Blast Resistance. Cell 2017, 170, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, W.; Zhang, Y.; Zhan, Q.; Liu, K.; Botella, J.R.; Bai, L.; Song, C. Abscisic acid-induced cytoplasmic translocation of constitutive photomorphogenic 1 enhances reactive oxygen species accumulation through the HY5-ABI5 pathway to modulate seed germination. Plant, Cell Environ. 2022, 45, 1474–1489. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; Sun, T.; Wang, D.; Wang, Z.; Zhang, C.; Que, Y.; Guo, J.; Xu, L.; Su, Y. Sugarcane ScDREB2B-1 l Nicotiana benthamiana by regulating the ABA signal, ROS level and stress-related gene expression. Int. J. Mol. Sci. 2022, 23, 9557. [Google Scholar] [CrossRef] [PubMed]
- Postiglione, A.E.; Muday, G.K. The Role of ROS Homeostasis in ABA-Induced Guard Cell Signaling. Front. Plant Sci. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.V.S.; Yadav, S.K.; Verma, R.K.; Shrivastava, S.; Ghimire, O.; Pushkar, S.; Rao, M.V.; Kumar, T.S.; Chinnusamy, V. The abscisic acid receptor OsPYL6 confers drought tolerance to indica rice through dehydration avoidance and tolerance mechanisms. J. Exp. Bot. 2020, 72, 1411–1431. [Google Scholar] [CrossRef]
- Kurup, S.; Jones, H.D.; Holdsworth, M.J. Interactions of the developmental regulator ABI3 with proteins identied from developing arabidopsis seeds. Plant J. 2000, 21, 143–155. [Google Scholar] [CrossRef]
- Khan, N.A.; Nazar, R.; Iqbal, N.; Anjum, N.A. Phytohormones and abiotic stress tolerance in plants; Anjum, N.A., Ed.; Springer Science & Business Media, 2012. [Google Scholar]
- Gallei, M.; Luschnig, C.; Friml, J. Auxin signaling in growth: schrödinger's cat out of the bag. Curr Opin Plant Biol. 2020, 53, 43–49. [Google Scholar] [CrossRef]
- Nghi, K.; Tagliani, A.; Mariotti, L.; Weits, D.; Perata, P.; Pucciariello, C. Auxin is required for the long coleoptile trait in japonica rice under submergence. New Phytol. 2021, 229, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Matsuba, S.; Funatsuki, H.; Kawaguchi, K.; Saruyama, H.; Tanida, M.; Sato, Y. Over-expression of a small heat shock protein, sHSP17.7, confers both heat tolerance and UV-B resistance to rice plants. Mol. Breed. 2004, 13, 165–175. [Google Scholar] [CrossRef]
- Zhao, H.; Jan, A.; Ohama, N.; Kidokoro, S.; Soma, F.; Koizumi, S.; Mogami, J.; Todaka, D.; Mizoi, J.; Shinozaki, K.; et al. Cytosolic HSC70s repress heat stress tolerance and enhance seed germination under salt stress conditions. Plant, Cell Environ. 2021, 44, 1788–1801. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.; Gloor, G. The Hsp90 family of proteins in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Knowlton, A.A.; Wasser, J.S.; Li, Q.; Lin, X.; Yang, X.; Weber, R.E.; Ostojic, H.; Fago, A.; Dewilde, S.; et al. Expression of heat shock proteins in turtle and mammal hearts: relationship to anoxia tolerance. Am. J. Physiol. Integr. Comp. Physiol. 2000, 278, R209–R214. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Xie, Y.; Yu, S.; Yang, J.; Chen, S.; Yuan, X.; Guo, T.; Wang, H.; Liu, Y.; Chen, C.; et al. The DnaJ domain-containing heat-shock protein NAL11 determines plant architecture by mediating gibberellin homeostasis in rice (Oryza sativa). New Phytol. 2023, 237, 2163–2179. [Google Scholar] [CrossRef] [PubMed]
- Magneschi, L.; Kudahettige, R.L.; Alpi, A.; Perata, P. Expansin gene expression and anoxic coleoptile elongation in rice cultivars. J. Plant Physiol. 2009, 166, 1576–1580. [Google Scholar] [CrossRef]
- Asatsuma, S.; Sawada, C.; Itoh, K.; Okito, M.; Kitajima, A.; Mitsui, T. Involvement of α-Amylase I-1 in Starch Degradation in Rice Chloroplasts. Plant Cell Physiol. 2005, 46, 858–869. [Google Scholar] [CrossRef]
- Hwang, Y.-S.; Thomas, B.; Rodriguez, R. Differential expression of rice α-amylase genes during seedling development under anoxia. Plant Mol. Biol. 1999, 40, 911–920. [Google Scholar] [CrossRef]
- Jackson, M.B.; Ram, P.C. Physiological and Molecular Basis of Susceptibility and Tolerance of Rice Plants to Complete Submergence. Ann. Bot. 2003, 91, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-H.; Wang, P.-Q.; Zhang, P.-P.; Nie, X.-M.; Li, B.-B.; Tai, L.; Liu, W.-T.; Li, W.-Q.; Chen, K.-M. NADPH Oxidases: The Vital Performers and Center Hubs during Plant Growth and Signaling. Cells 2020, 9, 437. [Google Scholar] [CrossRef]
- Hewage, K.A.H.; Yang, J.; Wang, D.; Hao, G.; Yang, G.; Zhu, J. Chemical Manipulation of Abscisic Acid Signaling: A New Approach to Abiotic and Biotic Stress Management in Agriculture. Adv. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Ye, N.; Zhu, G.; Liu, Y.; Zhang, A.; Li, Y.; Liu, R.; Shi, L.; Jia, L.; Zhang, J. Ascorbic acid and reactive oxygen species are involved in the inhibition of seed germination by abscisic acid in rice seeds. J. Exp. Bot. 2011, 63, 1809–1822. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: the new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Li, X.; Hicks, L.M.; Xiong, L. A Raf-Like MAPKKK Gene DSM1 Mediates Drought Resistance through Reactive Oxygen Species Scavenging in Rice. Plant Physiol. 2009, 152, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought Tolerance in Rice: Focus on Recent Mechanisms and Approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Aleem, M.; Aleem, S.; Sharif, I.; Wu, Z.; Aleem, M.; Tahir, A.; Atif, R.M.; Cheema, H.M.; Shakeel, A.; Lei, S.; Yu, D.; Wang, H.; Kaushik, P.; Alyemeni, M.N.; Bhat, J.A.; Ahmad, P. Characterization of SOD and GPX gene families in the Soybeans in response to drought and salinity stresses. Antioxidants 2022, 11, 460. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Miro, B.; Ismail, A.M. Tolerance of anaerobic conditions caused by flooding during germination and early growth in rice (Oryza sativa L.). Front. Plant Sci. 2013, 4, 269. [Google Scholar] [CrossRef] [PubMed]
- Perata, P.; Guglielminetti, L.; Alpi, A. Mobilization of Endosperm Reserves in Cereal Seeds under Anoxia. Ann. Bot. 1997, 79, 49–56. [Google Scholar] [CrossRef]
- Banti, V.; Loreti, E.; Novi, G.; Santaniello, A.; Alpi, A.; Perata, P. Heat acclimation and cross-tolerance against anoxia in Arabidopsis. Plant Cell Environ. 2008, 31, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-A.; Lim, E.-K.; Yu, S.-M. Sugar Response Sequence in the Promoter of a Rice α-Amylase Gene Serves as a Transcriptional Enhancer. J. Biol. Chem. 1998, 273, 10120–10131. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated Responses to Oxygen and Sugar Deficiency Allow Rice Seedlings to Tolerate Flooding. Sci. Signal. 2009, 2, ra61–ra61. [Google Scholar] [CrossRef]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa L.). Annu. Bot. 2008, 103, 197–209. [Google Scholar] [CrossRef]
- Gibbs, J.; Morrell, S.; Valdez, A.; Setter, T.; Greenway, H. Regulation of alcoholic fermentation in coleoptiles of two rice cultivars differing in tolerance to anoxia. J. Exp. Bot. 2000, 51, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Quimio, C.A.; Torrizo, L.B.; Setter, T.L.; Ellis, M.; Grover, A.; Abrigo, E.M.; Oliva, N.P.; Ella, E.S.; Carpena, A.L.; Ito, O.; et al. Enhancement of Submergence Tolerance in Transgenic Rice Overproducing Pyruvate Decarboxylase. J. Plant Physiol. 2000, 156, 516–521. [Google Scholar] [CrossRef]
- Lee, K.; Chen, J.J.W.; Wu, C.; Chang, H.; Chen, H.; Kuo, H.; Lee, Y.; Chang, Y.; Chang, H.; Shiue, S.; et al. Auxin plays a role in the adaptation of rice to anaerobic germination and seedling establishment. Plant, Cell Environ. 2022, 46, 1157–1175. [Google Scholar] [CrossRef]
- Singh, P.; Sinha, A.K. A Positive Feedback Loop Governed by SUB1A1 Interaction with MITOGEN-ACTIVATED PROTEIN KINASE3 Imparts Submergence Tolerance in Rice. Plant Cell 2016, 28, 1127–1143. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, X.; Jin, S.; Liu, X.; Zhu, L.; Nie, Y.; Zhang, X. Overexpression of Rice NAC Gene SNAC1 Improves Drought and Salt Tolerance by Enhancing Root Development and Reducing Transpiration Rate in Transgenic Cotton. PLOS ONE 2014, 9, e86895. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef] [PubMed]
- Gallei, M.; Luschnig, C.; Friml, J. Auxin signaling in growth: schrödinger's cat out of the bag. Current Opinion in Plant Biology 2020, 53, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Nghi, K.N.; Tagliani, A.; Mariotti, L.; Weits, D.A.; Perata, P.; Pucciariello, C. Auxin is required for the long coleoptile trait in japonica rice under submergence. New Phytol. 2020, 229, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.; Yuan, W.; Wang, Y.; Garcia-Maquilon, I.; Dang, X.; Li, Y.; Zhang, J.; Zhu, Y.; Rodriguez, P.L.; Xu, W. Low ABA concentration promotes root growth and hydrotropism through relief of ABA INSENSITIVE 1-mediated inhibition of plasma membrane H + -ATPase 2. Sci. Adv. 2021, 7, eabd4113. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhou, L.; Chen, W.; Ye, N.; Xia, J.; Zhuang, C. Overexpression of a microRNA-targeted NAC transcription factor improves drought and salt tolerance in Rice via ABA-mediated pathways. Rice 2019, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, Y.; Qiu, J.; Wang, H.; Wang, S.; Tang, L.; Tong, X.; Zhang, J. Abscisic acid promotes jasmonic acid biosynthesis via a ‘SAPK10-bZIP72-AOC’ pathway to synergistically inhibit seed germination in rice (Oryza sativa). New. Phytol. 2020, 228, 1336–1353. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, W.; He, Y.; Adegoke, T.V.; Ying, J.; Tong, X.; Li, Z.; Tang, L.; Wang, H.; Zhang, J.; et al. bZIP72 promotes submerged rice seed germination and coleoptile elongation by activating ADH1. Plant Physiol. Biochem. 2021, 169, 112–118. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; Xia, R. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant. 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Ma, X.; Zhu, Q.; Zeng, D.; Li, G.; Liu, Y.-G. CRISPR-GE: A Convenient Software Toolkit for CRISPR-Based Genome Editing. Mol. Plant 2017, 10, 1246–1249. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-X.; Yin, X.-X.; Li, S.; Peng, Y.-T.; Yan, X.-L.; Chen, C.; Hassan, B.; Zhou, S.-X.; Pu, M.; Zhao, J.-H.; et al. miR167d-ARFs Module Regulates Flower Opening and Stigma Size in Rice. Rice 2022, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, X.; Ma, X.; Li, J.; Chen, J.; Liu, Y.-G. DSDecode: A Web-Based Tool for Decoding of Sequencing Chromatograms for Genotyping of Targeted Mutations. Mol. Plant 2015, 8, 1431–1433. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Yang, J.; Li, D.; Peng, Z.; Xia, A.; Yang, M.; Luo, L.; Huang, C.; Wang, J.; Wang, H.; et al. Dynamic genome-wide association analysis and identification of candidate genes involved in anaerobic germination tolerance in rice. Rice 2021, 14, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Ma, J.; Zhai, H.; Xin, P.; Chu, J.; Qiao, Y.; Han, L. CHR729 Is a CHD3 Protein That Controls Seedling Development in Rice. PLOS ONE 2015, 10, e0138934. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Luo, L.; Cui, H.; Wang, H.; Guo, T.; Liu, Y.; Wang, J.; Huang, M.; Yang, G.; Chen, Z.; et al. Characterization and Fine Mapping of a Leaf Wilt Mutant, m3, Induced by Heavy Ion Irradiation of Rice. Crop. Sci. 2019, 59, 2679–2688. [Google Scholar] [CrossRef]
- Chen, T.; Luo, L.; Zhao, Z.; Wang, H.; Chen, C.; Liu, Y.; Li, X.; Guo, T.; Xiao, W. Fine mapping and candidate gene analysis of qGL10 affecting rice grain length. Crop. J. 2023, 11, 540–548. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Huang, L.; Hua, K.; Xu, R.; Zeng, D.; Wang, R.; Dong, G.; Zhang, G.; Lu, X.; Fang, N.; Wang, D.; Duan, P.; Zhang, B.; Liu, Z.; Li, N.; Luo, Y.; Qian, Q.; Yao, S.; Li, Y. The LARGE2-APO1/APO2 regulatory module controls panicle size and grain number in rice. Plant Cell. 2021, 33, 1212–1228. [Google Scholar] [CrossRef]
Figure 1.
Evolutionary tree of NAL11 homologs with Oryza sativa Japonica, Brachypodium distachyon, Sorghun bicolor, Zea mays, Glycine max, Nymphaea colorata, Arabidopsis, and wild rice (Oryza brachyantha, Oryza glaberrima). XP_015645205.1 is the accession number of NAL11 in NCBI, which is highlighted in the red box.
Figure 1.
Evolutionary tree of NAL11 homologs with Oryza sativa Japonica, Brachypodium distachyon, Sorghun bicolor, Zea mays, Glycine max, Nymphaea colorata, Arabidopsis, and wild rice (Oryza brachyantha, Oryza glaberrima). XP_015645205.1 is the accession number of NAL11 in NCBI, which is highlighted in the red box.

Figure 2.
Spatiotemporal expression analysis of NAL11. A, Transcription levels of NAL11 in germinating seeds of ZH11 under aerobic and submerged conditions using quantitative reverse transcription polymerase chain reaction (RT-PCR). Gene expression was normalized to that of OsActin, with relative expression levels represented as fold change relative to the expression level of NAL11 at 0 h. B, β-glucuronidase (GUS) staining of seeds after 24 h, 48 h, and 72 h of submergence stress. Scale Bar, 0.5 cm. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 2.
Spatiotemporal expression analysis of NAL11. A, Transcription levels of NAL11 in germinating seeds of ZH11 under aerobic and submerged conditions using quantitative reverse transcription polymerase chain reaction (RT-PCR). Gene expression was normalized to that of OsActin, with relative expression levels represented as fold change relative to the expression level of NAL11 at 0 h. B, β-glucuronidase (GUS) staining of seeds after 24 h, 48 h, and 72 h of submergence stress. Scale Bar, 0.5 cm. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
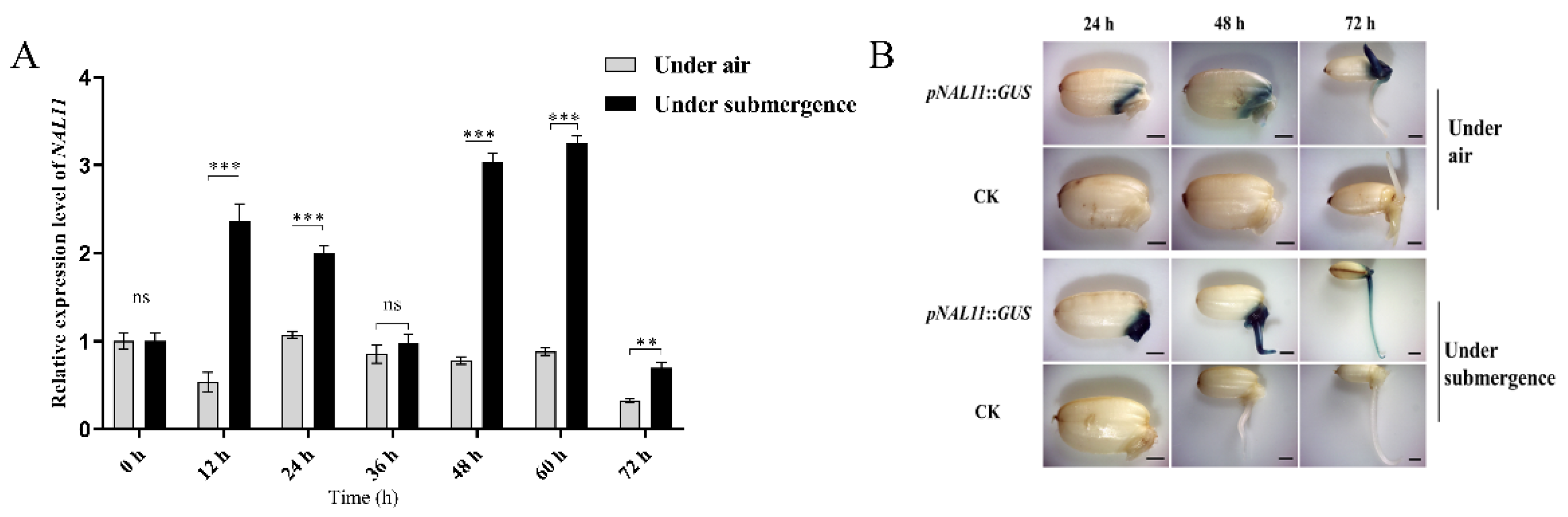
Figure 3.
Relative expression level of expansin genes in ZH11 and knockout lines at 48 h after submergence. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 3.
Relative expression level of expansin genes in ZH11 and knockout lines at 48 h after submergence. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
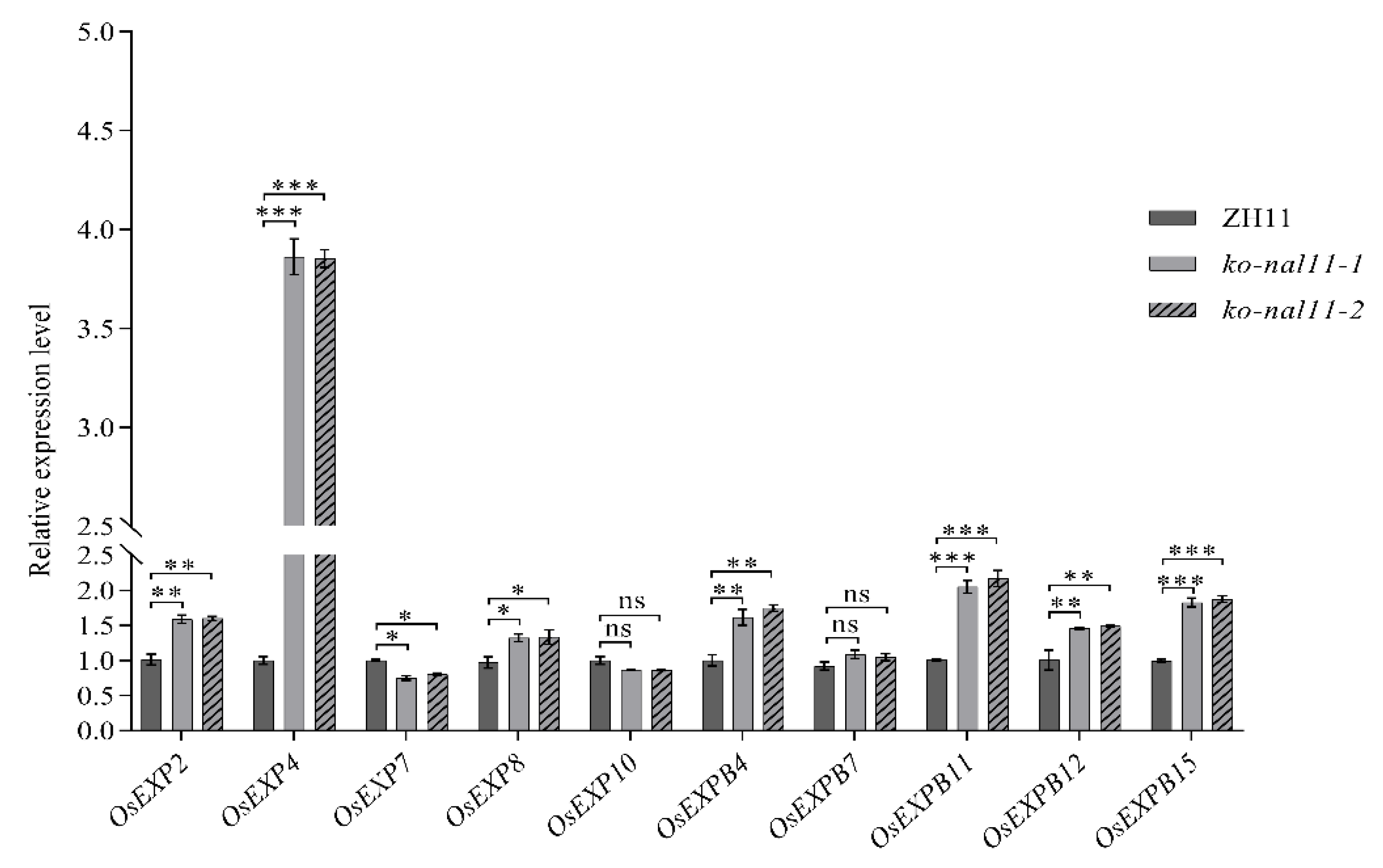
Figure 4.
NAL11 is involved in the sugar and energy pathway. A, Quantitative RT-PCR analysis of nine α-amylase family genes in seeds of ZH11 and knockout lines at 48 h after submergence treatment. B, α-AMS activity at different stages. C, RT-qPCR analysis of sugar and energy metabolism genes at 48 h after submergence treatment, respectively. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-ests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 4.
NAL11 is involved in the sugar and energy pathway. A, Quantitative RT-PCR analysis of nine α-amylase family genes in seeds of ZH11 and knockout lines at 48 h after submergence treatment. B, α-AMS activity at different stages. C, RT-qPCR analysis of sugar and energy metabolism genes at 48 h after submergence treatment, respectively. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-ests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
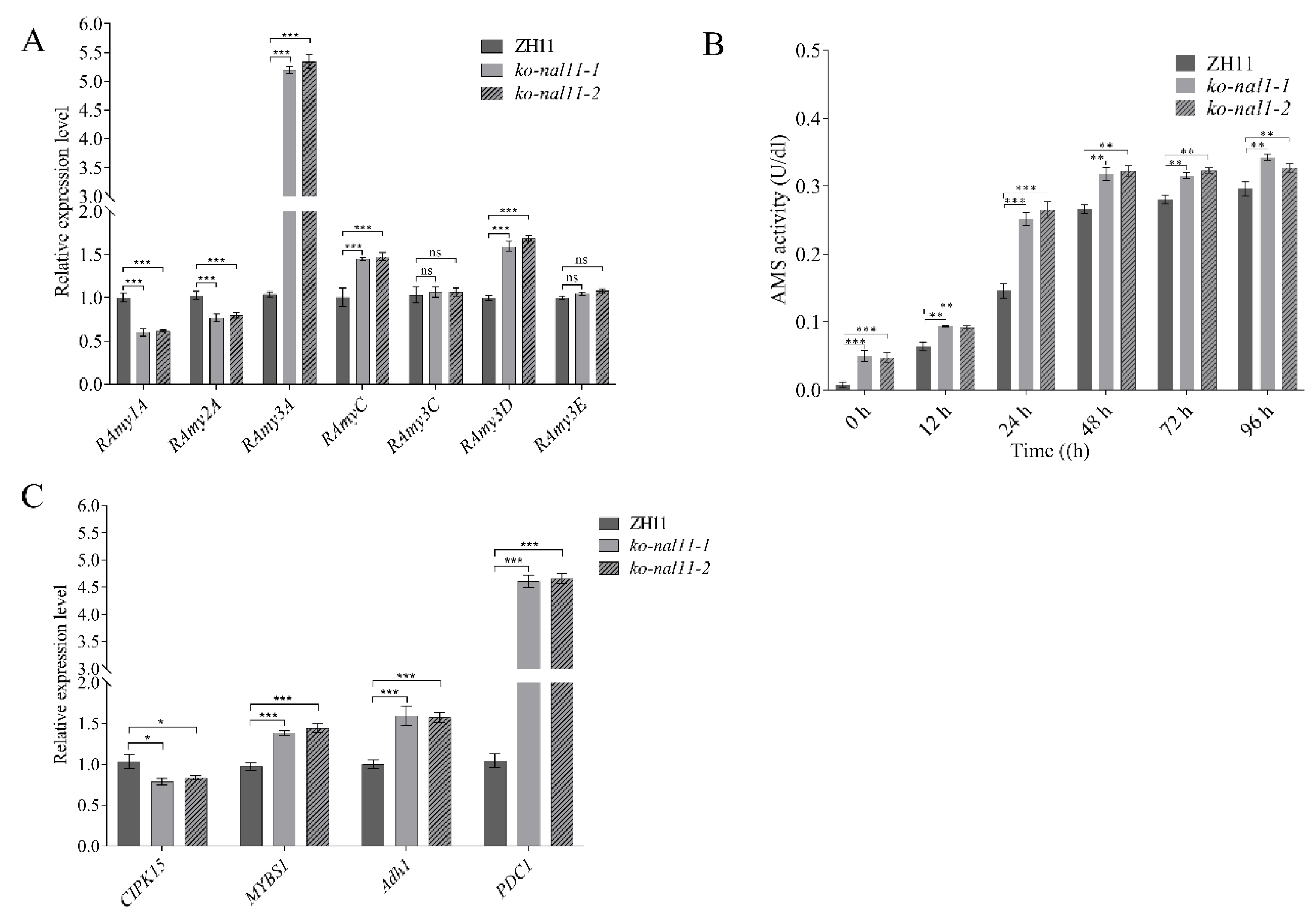
Figure 5.
Analysis of MDA and H2O2 content, transcript levels of ROS-production gene, OsRbohA and OsRbohE of WT and transgenic plants under normal and submerged conditions. A. MDA content. B. H2O2 content. C. OsRbohA. D. OsRbohE. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 5.
Analysis of MDA and H2O2 content, transcript levels of ROS-production gene, OsRbohA and OsRbohE of WT and transgenic plants under normal and submerged conditions. A. MDA content. B. H2O2 content. C. OsRbohA. D. OsRbohE. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 6.
Knockout of NAL11 affects the levels of ABA, GA and IAA. A, B and C. Content of endogenous ABA, GA and IAA at 24 h, 48 h, and 72 h after submergence. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 6.
Knockout of NAL11 affects the levels of ABA, GA and IAA. A, B and C. Content of endogenous ABA, GA and IAA at 24 h, 48 h, and 72 h after submergence. (Data are presented as mean ± SD, n = 5; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
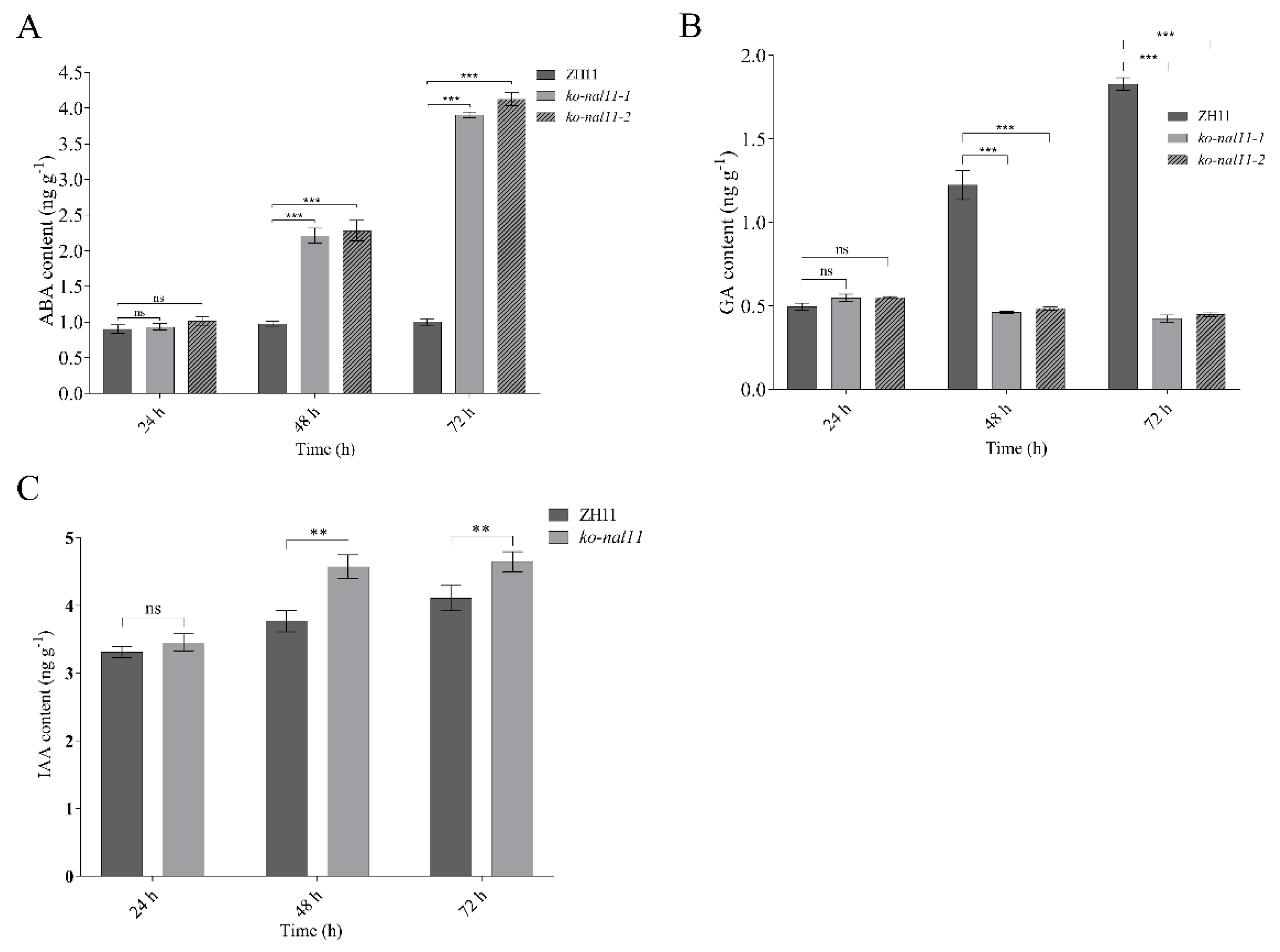
Figure 7.
Knockout of NAL11 shows sensitivity to ABA. A, B, Representative images of coleoptile length in response to different concentrations of ABA (0 µM, 0.001 µM, 0.01 µM, 0.1 µM, 1 µM, and 10 µM, respectively) after 4 d of submergence for both WT and knockout lines. Scale bars: 1 cm. B, Comparison of coleoptile lengths in WT and knockout lines in response to control (H2O) and ABA treatments after 4 d of submergence. (Data are presented as mean ± SD, n = 5 biologically independent samples; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Figure 7.
Knockout of NAL11 shows sensitivity to ABA. A, B, Representative images of coleoptile length in response to different concentrations of ABA (0 µM, 0.001 µM, 0.01 µM, 0.1 µM, 1 µM, and 10 µM, respectively) after 4 d of submergence for both WT and knockout lines. Scale bars: 1 cm. B, Comparison of coleoptile lengths in WT and knockout lines in response to control (H2O) and ABA treatments after 4 d of submergence. (Data are presented as mean ± SD, n = 5 biologically independent samples; significant differences were determined by two-tailed Student’s t-tests. *P < 0.05, **P < 0.01, ***P < 0.001, ns, no significance).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



