Submitted:
26 July 2024
Posted:
29 July 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Long COVID (Post-Acute Sequelae of COVID-19, PASC) is a consequence of infection by SARS-CoV-2 that continues to disrupt the well-being of millions of affected individuals for many months beyond their first infection. While the exact mechanisms underlying PASC remain to be defined, hypotheses regarding the pathogenesis of Long COVID are varied and include (but are not limited to) dysregulated local or systemic inflammatory responses, autoimmune mechanisms, viral-induced hormonal imbalances and long-term persistence of virus and/or fragments of viral RNA or proteins. This review article is based on a comprehensive review of the wide range of symptoms most often observed in long COVID and an attempt to integrate that information into a plausible hypothesis for the pathogenesis of PASC. In particular, it is proposed that longterm dysregulation of the gut in response to viral persistence could lead to the myriad of symptoms observed in PASC.
Keywords:
Long COVID
; PASC
; viral persistence
; gut
; SARS-CoV-2
1. Introduction
In the four years since the first appearance of Covid-19, there have been more than 774 million cases globally and a reported death rate of almost 1% from the disease (1). These are likely underestimates given the inadequacy of detection and reporting methodologies across the world. While vaccinations and new treatment methods have helped to bring the disease under control to some degree, post-COVID-19 sequelae, a syndrome termed Long Covid, has blighted the lives of many individuals. This review highlights the pathology of the chronic disease and the wide range of reported symptoms, and proposes underlying mechanisms which drive disease progression and viral persistence. The key hypothesis proposed here is that Long Covid (Post-Acute Sequelae of COVID-19, PASC) is primarily a disease driven by the gut that is perturbed by underlying inflammation.
Long Covid was first documented in the months after the Covid-19 pandemic began and has been associated with over 200 symptoms, with occurrence and severity varying among affected individuals. Long Covid can best be described as a multi-layered pathology that causes a variety of symptoms which are dependent on the baseline characteristics of the individual patient. The multi-faceted nature of this disease makes it inherently difficult to diagnose and treat. Some of the most common symptoms include fatigue, headache, myalgia, concentration deficit and sleep abnormalities (2).
The approach taken by the authors to clarifying this complex disease is, however, not to focus on symptoms, but to identify the primary pathophysiology and explain how symptoms can be so varied. Long COVID has been described as more likely among patients infected during early pandemic waves and who developed severe disease (3); the study of Sudre et al. (4) found that long COVID was more likely to occur in patients who reported five or more symptoms. It is important to differentiate Long Covid – which may occur after either a mild or severe infection – from the impact of heart, lung or kidney damage caused by a severe cytokine immune response to Covid-19. Patients with organ damage are more likely to have primarily specific symptoms like shortness of breath, and not the wide array of symptoms typically seen in Long Covid (5,6,7,8). People who have had severe COVID-19 are not necessarily more prone to Long Covid than those who had a milder case, but the resulting damage to vital organs could lead to a longer term impact from Long Covid. The present review includes studies of patients who had relatively mild to moderate COVID-19 and who did not need hospitalisation (9).
2. The Cellular Receptor for COVID Infection: Angiotensin Converting Enzyme-2 (ACE-2)
Following the original SARS coronavirus outbreak in South Asia in 2003, the cell surface receptor for the virus was rapidly identified as ACE-2, a protein discovered only three years earlier as a homologue of ACE, a key enzyme of the renin-angiotensin system (RAS) (10,11). Similarly, ACE-2 is the primary receptor for SARS-CoV2 (12) and also for the coronavirus CoV-NL63 which causes the common cold. ACE-2, like ACE, is a cell-surface zinc metallopeptidase; it acts as a counter-regulatory protein to ACE in the cardiovascular system (13). The removal of ACE-2 from the plasma membrane following Covid infection therefore contributes to the cardiovascular complications seen in the disease. However, ACE-2 is a multifunctional protein also facilitating intestinal amino acid transport, particularly tryptophan, through its association with the B0AT1 amino acid transporter protein, similar to the role of the protein collectrin in the kidney, which is homologous to the C-terminal domain of ACE-2 (14).
3. Viral Infection Routes, ACE-2 and Autoimmunity in Severe COVID-19
Severe COVID-19 with respiratory compromise has been described as a viral-mediated autoimmune response triggered by the combination of viral spike protein and serum ACE-2. It has previously been demonstrated that serum ACE-2, shed from the cell membrane, could bind tightly to the viral spike protein and potentially become caught up in the immune presentation of the virus by antigen-presenting cells (15,16). The immune system could then produce antibodies to ACE-2 (17) as well as to the virus. The ACE-2 autoantibodies could cause endothelial inflammation in the blood vessels of the lungs, heart and kidneys, leading to the development of microthrombi and respiratory compromise. ACE-2 is primarily located in the lungs, intestine, heart and kidneys, testes, the endothelial lining of blood vessels and especially the upper airways (18). It is also located in the brain, playing a key role in central cardiovascular regulation (19). Interestingly, a study of host genetic polymorphisms (20) showed that the ACE-2 rs2285666 T allele, among several other gene polymorphisms, was associated with an increased risk of long COVID; at present, however, the mechanisms underlying these associations are unknown.
The SARS-CoV2 virus infects the nasopharynx through binding to ACE-2 primarily on sustentacular cells (21), which express a high level of cell-surface ACE-2. Once infected, the virus employs a number of strategies to evade the interferon response (22) allowing further infection and spread asymptomatically to the lungs. Viral replication allows further infection of the lower airways, finally involving alveolar macrophages (23). Local spread to endothelial cells of lung blood vessels (24) is the final stage prior to blood-borne dissemination of the virus which leads to infection of intestinal cells, causing a positive stool result within the first four days of infection, and positive infection of intestinal enterocytes can also be observed (25).
4. Viral Persistence in COVID-19
Viral persistence is central to the pathology of Long Covid as revealed by long-term circulating levels of viral proteins in the serum of patients with ongoing symptoms (26). It is important to note that attempted culture of these viral remnants did not reveal viable viruses, that is, there are pieces of viral protein circulating rather than a persistent viral infection. A key question is which cells are host to these viral remnants?
There is a close association between the occurrence of Long Covid and inflammatory bowel disease (IBD) – of the ~10% of IBD patients who have contracted COVID, up to 40% of those develop Long Covid with symptoms of IBD (27) – which suggests that sites of intestinal pathology may be a primary location of viral persistence. Intestinal biopsies demonstrated concentration of viral protein, including nucleocapsid and spike proteins, in specific regions of the intestine (28). Identifying the concentration of viral remnants in specific intestinal cells facilitates an understanding of whether a specific type of intestinal cell is the primary target.
A recent tissue study in Long Covid was based on 46 patients with a background of IBD and recent SARS-CoV2 infection. An endoscopy study was conducted with biopsies from the small and large intestines. The biopsy specimens were collected in formalin and tissue samples were further processed using SARS-CoV2 PCR, immunofluorescence and viral culture to analyse for antigen persistence (29).
The presence of viral RNA was detected in 31% of biopsy specimens from the duodenum, ileum and colon, but there was no evidence of ability to culture the virus. Specific immunofluorescence demonstrated involvement of epithelial intestinal cells and CD8 T lymphocytes. It is unclear which type of epithelial cell is primarily involved. The intestinal lining is primarily composed of absorptive enterocytes covering the lamina propria and muscularis layers. An understanding of primary cell involvement can provide clues about the associated pathophysiology in Long Covid patients. Potential cell candidates are considered in Figure 1 below.
Figure 1 illustrates the various cell types of which enterocytes are the most common in the intestinal lining and have the primary function of nutrient absorption. The biopsy review (28), however, revealed that the concentration of viral particles was localised to specific cells in the small intestine and colon as opposed to the diffuse pattern that would be expected if enterocytes were involved. Additionally, enterocytes are typically present for only about five days before being replaced by new ones, which makes them a less likely site for long-term concentration of viral remnants (29). A more probable candidate is an intestinal cell with the ability to survive for longer periods of time.
Stem cells, which are the progenitor cells for most of the intestinal cells, are typically located in the intestinal crypts and help to maintain normal gut function. The location of these cells appears not to fit the observed pattern of localisation from the intestinal samples. Additionally, if the stem cells played a key role, the viral remnants would likely be in multiple types of progenitor cells, which has not been demonstrated.
The microfold or M cells, which sample intestinal fluid to identify pathogens, are a possible source of viral persistence. However, they are typically localised to Peyer’s patches, which are immune-sensing regions in the intestine, rather than being diffusely spread through the intestinal lining (30,31,32,33). Paneth cells modulate the microbiome and mediate the inflammatory response (34,35). They are longer lived than enterocytes, with a lifespan of up to 60 days (36), and represent a potential source of SARS-CoV2 viral remnants.
Similar to M cells are the cup cells which are another unique group of intestinal cells, but they are not localised on Peyer’s patches. Critically, these cells express vimentin G protein receptors which are normally involved in sensing, and vimentin is reported to be a co-receptor facilitating SARS-CoV2 cell entry. Extracellular vimentin could, therefore, be a target against viral invasion (37,38). The occurrence of persistent G protein receptor antibodies in Long Covid suggests that cup cells could be relevant to viral persistence.
Enterochromaffin cells (EC) are neuroendocrine cells within the intestinal lining. They are a diverse collection of cells comprising less than 1% of intestinal cells and are primarily secretors of gut peptides that influence the intestine and distant organs (39). They are involved in the control of gut inflammation and motility.
Finally, Tuft cells are secretory epithelial cells in the intestine with the ability to sense the surrounding environment and produce biogenic amines and peptides such as serotonin and calcitonin -gene-related peptide (CGRP).
The involvement of T-cells in all biopsy specimens has been noted, specifically CD8 T-cells in regions of active intestinal inflammation. The persistence of viral antigen has been demonstrated in CD8 pharyngeal cells (40) which could account for the persistence seen within intestinal CD8 cells. If T lymphocytes are found to be central to Long Covid, then the primary pathophysiology should point to a mechanism that is consistent across all patterns of the disease.
Autopsy studies have also shed some light on the issue of viral persistence in Long COVID. Stein et al. (41) looked at 44 autopsy specimens with a focus on the burden of infection, both acutely and over seven months following symptom onset. The data showed a wide distribution of viral RNA in multiple pulmonary and extrapulmonary tissues for up to 230 days following symptom onset, but with little inflammation or evidence of direct tissue or organ system injury, nor any apparent direct viral cytopathic effect outside of the lungs. These results were taken to indicate that the virus can survive and thrive in the body without generating a direct inflammatory response and suggested that the immune response is less capable outside of the lungs. The authors detected sgRNA (subgenomic RNA) in at least one tissue in over half of cases (14/27) beyond day 14, suggesting that prolonged viral replication may occur in extra pulmonary tissues as late as day 99. Outside the lungs, histological changes were mainly related to complications of therapy or preexisting co-morbidities, mainly obesity, diabetes, and hypertension. In the 44 cases RNA was detected in 79 of 85 anatomic locations with at least 100 N (nucleocapsid) gene copies/ng of RNA from every tissue. The authors also recovered replication-competent virus from the thalamus. In the hypothalamus, neurons were stained with anti-S and anti-N antibodies. Thus, non-intestinal and non-epithelial cells also serve as reservoirs of replicating viruses.
5. The Gut-Brain Axis: Potential Role in COVID-Related Cognitive Dysfunction
One of the symptoms of long COVID has been described as “brain fog”, difficulty in concentrating or cognitive decline. In the light of growing literature describing the now well-established concept of the “Gut-Brain Axis” (42), it can be hypothesized that gut dysfunction due to SARS-Co-V2 could be responsible for at least some of the cognitive dysfunction seen in long COVID. Indeed, Chen et al. (43) documented an association between gut microbiota dysbiosis and poor functional outcomes in acute ischemic stroke patients with COVID-19 infection. The potential crosstalk between COVID-19 and the gut-brain axis has been discussed critically (44), and Plummer et al. (45) have proposed a hypothetical scenario in gut-brain axis pathogenesis might invoke mechanisms leading to neurocognitive decline, such as decreased intestinal and brain barrier functions that promote immune-mediated systemic and neural inflammation. On the other hand, Wong et al. (46) showed that serotonin is reduced in long COVID, and reasoned that serotonin reduction might impede the activity of the vagus and thereby impair hippocampal function and memory. Regardless, in the light of this growing literature, it seems reasonable to speculate that gut dysfunction caused by SARS-CoV-2 persistence in the gut could contribute to the neurocognitive decline in PASC.
6. Gut Autoimmunity
How is the intestine predisposed to persistent viral replication and is it the main viral reservoir in Long Covid? There are higher concentrations of EC in coeliac disease and IBS (47). If these cells are the primary route to the intestinal infection, higher numbers of EC with basolateral B0AT1 transporters could increase the risk of this infection occurring. Basolateral B0AT1 transporters with ACE-2 have not been identified, but there are high numbers on the apical side to allow probable infection (48). Additionally, faecal microbiome changes point strongly to SARS-CoV2 infection as inducing an immune response in the gut (49).
Immune responses related to food intolerance in the gut are tightly regulated, primarily through T-cell activation as demonstrated in coeliac disease (50). Intestinal infection with SARS-CoV2 can dysregulate the balance between regulatory and effector T-cells and potentially increase intestinal inflammation (51). The association between SARS-CoV2 infection and lymphopenia suggests an immune response in the gut (52) where cell-mediated immunity is a pre-existing issue.
7. Viral Persistence in Specific Cell Types
This paper is focused on intestinal EC, tuft and cup cells as potential primary locations for viral remnants related to SARS-CoV2 and as possible drivers for persistent immune responses in Long Covid.
EC cells have serotonin as the primary biogenic amine and account for over 95% of total body serotonin content. EC cell uptake of its precursor, tryptophan, from the intestinal lumen is likely to be facilitated by the B0AT1 amino acid transporter with associated ACE-2 acting as a chaperone protein, like its homologue, collectrin (53).
SARS-CoV2 serum viremia could target the intestinal EC cells for infection. There is evidence that SARS-CoV2 can broadly infect intestinal cells with ongoing diffuse replication, suggesting luminal enterocyte infection (54). In the context of serum viremia, it is probable that EC cells are one of the primary routes to gain access to the intestinal lumen. Once in intestinal secretions and faecal matter, viral particles can continue to replicate in the latter (55) and subsequently infect enterocytes which have ACE-2 concentrated on the luminal portion of the cell. Prolonged faecal shedding of viral RNA for up to 210 days post infection points to the intestine as the source of ongoing viremia (56).
Tuft cells are potentially associated with Long Covid symptoms. It has already been established that they have the characteristic for viral persistence with Norovirus (57). These tuft cells, once infected, are able to evade clearance from the immune system.
Additionally, tuft cells have the unique ability to be chemosensing with regards to sweet and bitter (58). Chemosensing takes place through a G-protein receptor. It is possible that interaction with bacterial toxins through persistent infection (59) could stimulate an immune response to these G-protein receptors. Persistently elevated autoantibodies to G-protein receptors could also explain many of the diverse changes in Long Covid (60,61,62). The association of G-protein receptors with chemosensing intestinal cells could also be linked to taste and smell abnormalities which are some of the most common symptoms in Long Covid (63).
Viral antigens were definitively located in CD8+T cells within the intestinal lining. What was not clear was the exact morphology of these CD8+ cells. One group of CD8+ cells are the Mucosal-Associated Invariate T (MAIT) cells that have unique characteristics with the ability to bridge innate and adaptive immunity (64). Interaction with intestinal microbes and concentration in the intestinal tract could constitute a trigger mechanism for inflammation (65). These cells circulate throughout the body and thus could be relevant to the broad spectrum of symptoms associated with long COVID (66).
8. Mosaic Theory of Long COVID
The spectrum of disease manifestations in Long Covid is broad. The immune response to persistence of SARS-CoV2 viral remnants in specific intestinal cells is likewise expected to vary and will depend on the number and type of cells involved as well as the predisposition of the individual. However, patterns are likely to evolve which are relatively consistent. This understanding has led to the formulation of a mosaic theory of Long Covid. The mosaic theory has been applied elsewhere in medical research, for example, in hypertension, it was proposed that interaction of many factors led to elevated blood pressure and end organ damage (67).
Smell and taste abnormalities in the disease suggest involvement of tuft cells as a main driver for some Covid-19 symptoms; a study by Santin et al. (68) suggested the possible involvement of genetic variants of the extra-oral bitter (TAS2Rs) and sweet (TAS1Rs) taste receptors. Cup cells with associated vimentin could be related to G protein receptor autoantibodies, potentially leading to low cortisol in Long Covid (69) and associated fatigue through potential impact on the G protein cortisol feedback receptors in the pituitary. Infected EC cell involvement might cause postural orthostatic tachycardia syndrome (POTS)-type symptoms and depression through serotonin interaction, since POTS is a multisystem disorder frequently initiated by viral infection (70). Up to 60% of Covid-19 patients can develop POTS-like symptoms (71).
Symptoms of long Covid fatigue are very similar to post viral fatigue syndrome (72). On the other hand, some subjective yet specific symptoms as chronic fatigue, myalgia, concentration deficit, insomnia should be screened for false positives, especially in patients developing mild-to moderate disease (3). It has been noted that reactivation of Epstein-Barr virus (EBV) is associated with Covid infection (73) and could be a contributor to some of the patterns in the disease. The association with EBV gut persistence and gut immunity (74) is especially important in the context of SARS-COV2 viral persistence in the gut immune system.
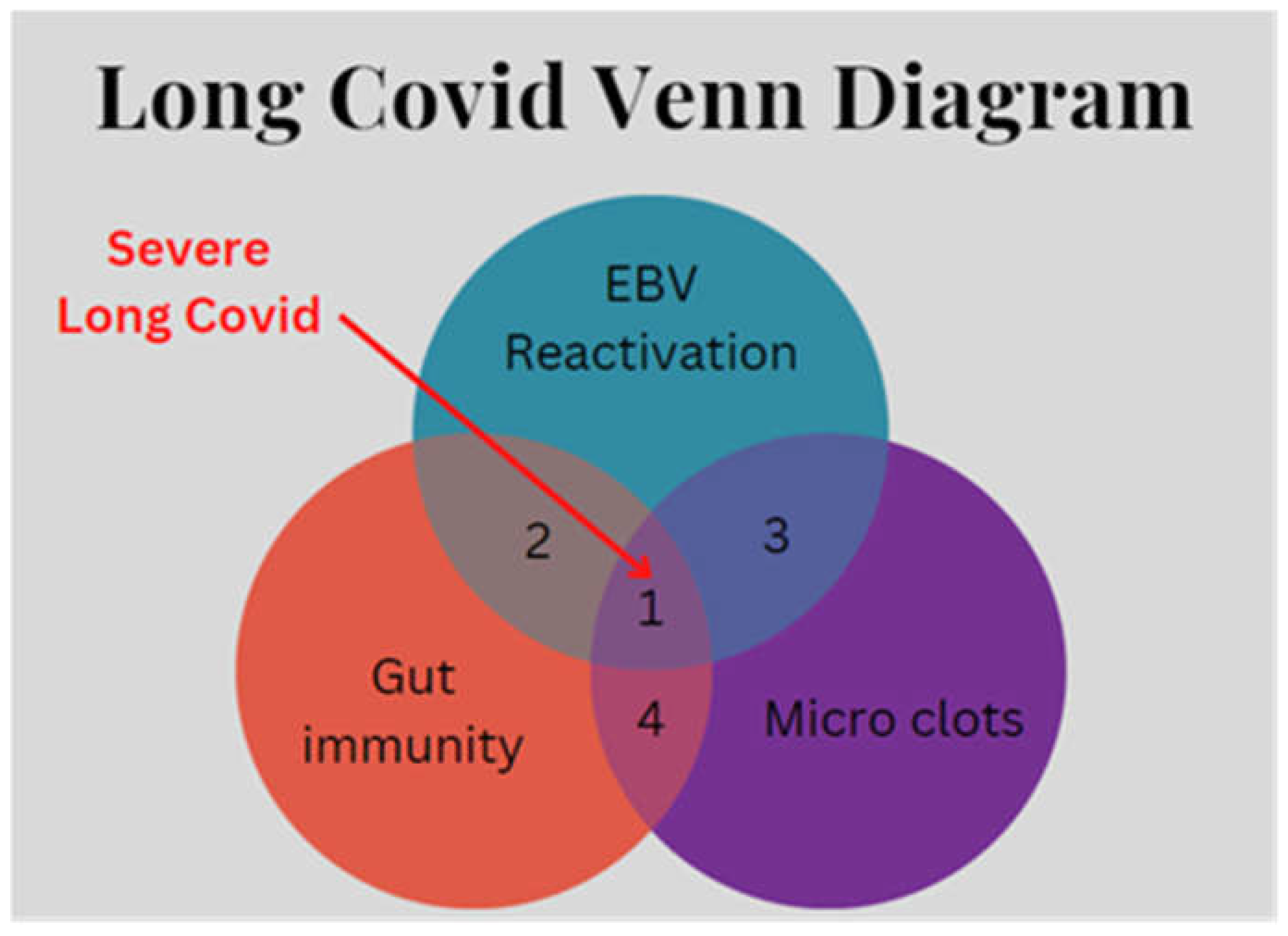
Coagulation activation and microclots are a typical feature of Long Covid and highlight an additional facet of the disease. Elevated levels of von Willebrand Factor (VWF) in Long Covid along with low levels of the metalloprotease ADAMTS13 could point to an intestinal toxin as an associated factor (75). In shigellosis, the exotoxin from the bacterial infection can lead to excessive clotting in the form of thrombotic thrombocytopenic purpura (76). The parallel with toxin-related microclots in Long Covid is striking. In consideration of all these findings, it is proposed (see Figure 2) that at least three key factors can interact to promote the occurrence of severe Long Covid: (EBV) reactivation, gut immunity and the occurrence of microclots.
9. Discussion
Long Covid appears to be a viral-mediated inflammatory disease with dysregulated intestinal immune responses as the primary driver of symptoms. The predisposition for Long Covid appears to be prior chronic intestinal inflammation. The inflamed areas have the potential for increases in specific cell types, such as EC, tuft and cup cells, and could be the location of viral remnants which cause ongoing symptoms.
Long Covid has a heterogeneous pattern which depends on the characteristics of prior gut inflammation and whether it is T-cell or humoral mediated. Overall, the SARS-CoV2 viral infection may cause dysregulated T-cell control in inflamed areas of the intestine, exacerbating the chronic inflammation. The heterogeneity of Long Covid indicates that multiple factors could be at play. With a more detailed understanding of the symptoms, specific cell types may be targeted to reduce the probability of viral persistence. Broad symptoms suggest there are a number of overlapping factors at play which make it difficult to identify primary pathology. Bacterial toxins could be a contributing factor in Long Covid; they are known to be a part of other diseases such as chronic fatigue syndrome and fibromyalgia and could be related to immune responses to bacterial exotoxins.
An important but poorly understood issue is the ability of COVID vaccinations to prevent the longterm symptoms often observed in PASC. In a staggered cohort study of COVID patients in the UK, Spain and Estonia, Catala et al. (77) found that both mRNA-based and adenoviral-based COVID vaccines were effective at reducing the risk of long COVID symptoms as defined by WHO. The mechanism(s) by which vaccination reduces the risk of long COVID symptoms are at present unknown, but deserve further attention, particularly in the light of prior vaccine hesitancy due to safety concerns and the lower risk of severe COVID in young people (ibid).
On the other hand, a decreased risk of long-covid-19 symptoms was reported not only following vaccination, but also after SARS-Cov-2 reinfections, suggesting the role of pre-existing immunity acquired by either natural infection, vaccination or both (hybrid) to prevent post-COVID-19 sequelae (3). LC dropped in fact dramatically after reinfections, which almost entirely surged during Omicron (3,78), although rare reinfections observed during the first 1-2 pandemic years were reportedly more severe than the respective primary COVID-19 events.
Clarification of the characteristics of the immune response could direct the appropriate immune-suppressive approach to manage Long Covid. The evidence strongly points to Long Covid being a complex, multisystemic viral-mediated autoimmune disease. Developing comprehensive answers to the questions raised above will undoubtedly take considerable time and further research to resolve the origins and potential therapies for Long Covid.
Author Contributions
The primary conceptualization, organization and writing of the text was performed by P.M. In subsequent drafts, A.T. and B.U. performed restructuring and literature searching to expand the text. The manuscript drafts were edited, revised and formatted by A.T. and B.U. All authors approved the final version of the submitted manuscript.
Acknowledgments
The authors acknowledge the expert editorial and preparatory work of Jacqueline Elaine Allison, Ph.D.
Conflicts of Interest
No Conflicts of Interest exist.
References
- World Health Organization COVID-19 Dashboard, March 26, 2024.
- Frontera JA, Guekht A, Allegri RF, Ashraf M, Baykan B, Crivelli L, Easton A, Garcia-Azorin D, Helbok R, Joshi J, Koehn J, Koralnik I, Netravathi M, Michael B, Nilo A, Özge A, Padda K, Pellitteri G, Prasad K, Romozzi M, Saylor D, Seed A, Thakur K, Uluduz D, Vogrig A, Welte TM, Westenberg E, Zhuravlev D, Zinchuk M, Winkler AS. Evaluation and treatment approaches for neurological post-acute sequelae of COVID-19: A consensus statement and scoping review from the global COVID-19 neuro research coalition. J Neurol Sci. 2023. Nov 15;454:120827. Epub 2023 Oct 13. PMID: 37856998. [CrossRef]
- Cegolon L, Mauro M, Sansone D, Tassinari A, Gobba FM, Modenese A, Casolari L, Liviero F, Pavanello S, Scapellato ML, Taus F, Carta A, Spiteri G, Monaco MGL, Porru S, Larese Filon F. A Multi-Center Study Investigating Long COVID-19 in Healthcare Workers from North-Eastern Italy: Prevalence, Risk Factors and the Impact of Pre-Existing Humoral Immunity-ORCHESTRA Project. Vaccines (Basel). 2023 Nov 27;11(12):1769. PMID: 38140174; PMCID: PMC10747427. [CrossRef]
- Sudre CH, Murray B, Varsavsky T, Graham MS, Penfold RS, Bowyer RC, Pujol JC, Klaser K, Antonelli M, Canas LS, Molteni E, Modat M, Jorge Cardoso M, May A, Ganesh S, Davies R, Nguyen LH, Drew DA, Astley CM, Joshi AD, Merino J, Tsereteli N, Fall T, Gomez MF, Duncan EL, Menni C, Williams FMK, Franks PW, Chan AT, Wolf J, Ourselin S, Spector T, Steves CJ. Attributes and predictors of long COVID. Nat Med 2021, 27:626-631. [CrossRef]
- Raveendran AV, Jayadevan R, Sashidharan S. Long COVID: An overview. Diabetes Metab Syndr 2021, 15:869-875. [CrossRef]
- Crook H, Raza S, Nowell J, Young M, Edison P.. Long covid-mechanisms, risk factors, and management. BMJ 2021, 374:n1648. [CrossRef]
- Sandler CX, Wyller VBB, Moss-Morris R, Buchwald D, Crawley E, Hautvast J, Katz BZ, Knoop H, Little P, Taylor R, Wensaas KA, Lloyd AR. Long COVID and Post-infective Fatigue Syndrome: A Review. Open Forum Infect Dis. 2021 Sep 9;8(10):ofab440. PMID: 34631916; PMCID: PMC8496765. [CrossRef]
- Davis HE, Assaf GS, McCorkell L, Wei H, Low RJ, Re’em Y, Redfield S, Austin JP, Akrami. A. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. E Clinical Medicine 2021, 38:101019. [CrossRef]
- Lopez-Leon S, Wegman-Ostrosky T, Perelman C, Sepulveda R, Rebolledo PA, Cuapio A, Villapol S. More than 50 long-term effects of COVID-19: a systematic review and meta-analysis. Sci Rep 2021 11(1):16144. [CrossRef]
- Tipnis SR, Hooper NM, Hyde R, Karran E, Christie G, Turner AJ. A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J Biol Chem 2000, 275:33238-33243. [CrossRef]
- Donoghue M, Hsieh F, Baronas E, Godbout K, Gosselin M, Stagliano N, Donovan M, Woolf B, Robison K, Jeyaseelan R, Breitbart RE, Acton S. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1-9. Circ Res 2000 87:E1-9. [CrossRef]
- Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, Müller MA, Drosten C, Pöhlmann S. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181:271-280.e8. [CrossRef]
- Hooper NM, Lambert DW, Turner. Discovery and characterization of ACE2 - a 20-year journey of surprises from vasopeptidase to COVID-19. Clin Sci (Lond) 2020 134:2489-2501. [CrossRef]
- Stevens BR, Ellory JC, Preston RL. B0AT1 Amino Acid Transporter Complexed With SARS-CoV-2 Receptor ACE2 Forms a Heterodimer Functional Unit: In Situ Conformation Using Radiation Inactivation Analysis. Function (Oxf) 2021, May 13;2(4) 2:zqab027. [CrossRef]
- McMillan P, Dexhiemer T, Neubig RR, Uhal BD. COVID-19-A Theory of Autoimmunity Against ACE-2 Explained. Front Immunol 2021, 12:582166. [CrossRef]
- McMillan P, Uhal BD. COVID-19-A theory of autoimmunity to ACE-2. MOJ Immunol. 2020, 7:17-19.
- Arthur JM, Forrest JC, Boehme KW, Kennedy JL, Owens S, Herzog C, Liu J, Harville TO. Development of ACE2 autoantibodies after SARS-CoV-2 infection. PLoS One 2021, 16:e0257016. [CrossRef]
- Hou YJ, Okuda K, Edwards CE, Martinez DR, Asakura T, Dinnon KH 3rd, Kato T, Lee RE, Yount BL, Mascenik TM, Chen G, Olivier KN, Ghio A, Tse LV, Leist SR, Gralinski LE, Schäfer A, Dang H, Gilmore R, Nakano S, Sun L, Fulcher ML, Livraghi-Butrico A, Nicely NI, Cameron M, Cameron C, Kelvin DJ, de Silva A, Margolis DM, Markmann A, Bartelt L, Zumwalt R, Martinez FJ, Salvatore SP, Borczuk A, Tata PR, Sontake V, Kimple A, Jaspers I, O’Neal WK, Randell SH, Boucher RC, Baric RS. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell. 2020 Jul 23;182(2):429-446.e14. Epub 2020 May 27.PMID: 32526206. [CrossRef]
- Xia H, Lazartigues E. Angiotensin-converting enzyme 2 in the brain: properties and future directions. J Neurochem 2008, 107:1482-1494. [CrossRef]
- Udomsinprasert W, Nontawong N, Saengsiwaritt W, Panthan B, Jiaranai P, Thongchompoo N, Santon S, Runcharoen C, Sensorn I, Jittikoon J, Chaikledkaew U, Chantratita W. Host genetic polymorphisms involved in long-term symptoms of COVID-19. Emerg Microbes Infect. 2023 Dec;12(2):2239952. PMID: 37497655; PMCID: PMC10392286. [CrossRef]
- Bryche B, St Albin A, Murri S, Lacôte S, Pulido C, Ar Gouilh M, Lesellier S, Servat A, Wasniewski M, Picard-Meyer E, Monchatre-Leroy E, Volmer R, Rampin O, Le Goffic R, Marianneau P, Meunier N. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav Immun 2020, 89:579-586. [CrossRef]
- Xia H, Cao Z, Xie X, Zhang X, Chen JY, Wang H, Menachery VD, Rajsbaum R, Shi PY. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep 2020. 33:108234. [CrossRef]
- Dalskov L, Møhlenberg M, Thyrsted J, Blay-Cadanet J, Poulsen ET, Folkersen BH, Skaarup SH, Olagnier D, Reinert L, Enghild JJ, Hoffmann HJ, Holm CK, Hartmann R. SARS-CoV-2 evades immune detection in alveolar macrophages. EMBO Rep 2020, 21:e51252. [CrossRef]
- Liu F, Han K, Blair R, Kenst K, Qin Z, Upcin B, Wörsdörfer P, Midkiff CC, Mudd J, Belyaeva E, Milligan NS, Rorison TD, Wagner N, Bodem J, Dölken L, Aktas BH, Vander Heide RS, Yin XM, Kolls JK, Roy CJ, Rappaport J, Ergün S, Qin X. 2. SARS-CoV-2 Infects Endothelial Cells In Vivo and In Vitro. Front Cell Infect Microbiol 2021, 11:701278. [CrossRef]
- Moura IB, Buckley AM, Wilcox MH. Can SARS-CoV-2 be transmitted via faeces? Curr Opin Gastroenterol 2022, 38:26-29. [CrossRef]
- Swank Z, Senussi Y, Manickas-Hill Z, Yu XG, Li JZ, Alter G, Walt DR. Persistent Circulating Severe Acute Respiratory Syndrome Coronavirus 2 Spike Is Associated With Post-acute Coronavirus Disease 2019 Sequelae. Clin Infect Dis 2023, 76:e487-e490. [CrossRef]
- Salvatori S, Baldassarre F, Mossa M, Monteleone G. Long COVID in Inflammatory Bowel Diseases. J Clin Med 2021, 10:5575. [CrossRef]
- Zollner A, Koch R, Jukic A, Pfister A, Meyer M, Rössler A, Kimpel J, Adolph TE, Tilg H. Postacute COVID-19 is Characterized by Gut Viral Antigen Persistence in Inflammatory Bowel Diseases. Gastroenterology 2022, 163:495-506.e8. [CrossRef]
- Snoeck V, Goddeeris B, Cox E. The role of enterocytes in the intestinal barrier function and antigen uptake. Microbes Infect 2005, 7:997-1004. [CrossRef]
- Ohno H, Kanaya T, Williams IR. M cell differentiation: distinct lineage or phenotypic transition? Salmonella provides answers. Cell Host Microbe 2012, 12:607-609. [CrossRef]
- Neutra MR. Current concepts in mucosal immunity. V Role of M cells in transepithelial transport of antigens and pathogens to the mucosal immune system. Am J Physiol 1998, 274:G785-791. [CrossRef]
- Miller H, Zhang J, Kuolee R, Patel GB, Chen W. Intestinal M cells: the fallible sentinels? World J Gastroenterol 2007 13:1477-1486. [CrossRef]
- Neutra MR, Mantis NJ, Kraehenbuhl JP. Collaboration of epithelial cells with organized mucosal lymphoid tissues. Nat Immunol 2001, 2:1004-1009. [CrossRef]
- Lueschow SR, McElroy SJ. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front Immunol 2020, 11:587. [CrossRef]
- Clevers HC, Bevins CL. Paneth cells: maestros of the small intestinal crypts. Annu Rev Physiol 2013, 75:289-311. [CrossRef]
- Roth S, Franken P, Sacchetti A, Kremer A, Anderson K, Sansom O, Fodde R. Paneth cells in intestinal homeostasis and tissue injury. PLoS One 2012, 7:e38965. [CrossRef]
- Suprewicz Ł, Swoger M, Gupta S, Piktel E, Byfield FJ, Iwamoto DV, Germann D, Reszeć J, Marcińczyk N, Carroll RJ, Janmey PA, Schwarz JM, Bucki R, Patteson AE. Extracellular Vimentin as a Target Against SARS-CoV-2 Host Cell Invasion. Small 2022, 18:e2105640. [CrossRef]
- Amraei R, Xia C, Olejnik J, White MR, Napoleon MA, Lotfollahzadeh S, Hauser BM, Schmidt AG, Chitalia V, Mühlberger E, Costello CE, Rahimi N. Extracellular vimentin is an attachment factor that facilitates SARS-CoV-2 entry into human endothelial cells. Proc Natl Acad Sci U S A 2022, 119:e2113874119. [CrossRef]
- Diwakarla S, Fothergill LJ, Fakhry J, Callaghan B, Furness JB. Heterogeneity of enterochromaffin cells within the gastrointestinal tract. Neurogastroenterol Motil 2017, 29:10.1111/nmo.13101. [CrossRef]
- Vibholm LK, Nielsen SSF, Pahus MH, Frattari GS, Olesen R, Andersen R, Monrad I, Andersen AHF, Thomsen MM, Konrad CV, Andersen SD, Højen JF, Gunst JD, Østergaard L, Søgaard OS, Schleimann MH, Tolstrup M. SARS-CoV-2 persistence is associated with antigen-specific CD8 T-cell responses. EBioMedicine 2021, 64:103230. [CrossRef]
- Stein SR, Ramelli SC, Grazioli A, Chung JY, Singh M, Yinda CK, Winkler CW, Sun J, Dickey JM, Ylaya K, Ko SH, Platt AP, Burbelo PD, Quezado M, Pittaluga S, Purcell M, Munster VJ, Belinky F, Ramos-Benitez MJ, Boritz EA, Lach IA, Herr DL, Rabin J, Saharia KK, Madathil RJ, Tabatabai A, Soherwardi S, McCurdy MT; NIH COVID-19 Autopsy Consortium; Peterson KE, Cohen JI, de Wit E, Vannella KM, Hewitt SM, Kleiner DE, Chertow DS. SARS-CoV-2 infection and persistence in the human body and brain at autopsy. Nature 2022, 612:758-763. [CrossRef]
- Warren A, Nyavor Y, Beguelin A, Frame LA. Dangers of the chronic stress response in the context of the microbiota-gut-immune-brain axis and mental health: a narrative review. Front Immunol. 2024 May 2;15:1365871. PMID: 38756771; PMCID: PMC11096445. [CrossRef]
- Chen J, Gao X, Liang J, Wu Q, Shen L, Zheng Y, Ma Y, Peng Y, He Y, Yin J. Association between gut microbiota dysbiosis and poor functional outcomes in acute ischemic stroke patients with COVID-19 infection. mSystems. 2024 May 3:e0018524. Epub ahead of print. PMID: 38700338. [CrossRef]
- Kalam N, Balasubramaniam VRMT. Crosstalk between COVID-19 and the gut-brain axis: a gut feeling. Postgrad Med J. 2024 Mar 16:qgae030. Epub ahead of print. PMID: 38493312. [CrossRef]
- Plummer AM, Matos YL, Lin HC, Ryman SG, Birg A, Quinn DK, Parada AN, Vakhtin AA. Gut-brain pathogenesis of post-acute COVID-19 neurocognitive symptoms. Front Neurosci. 2023 28;17S:1232480. PMID: 37841680; PMCID: PMC10568482. [CrossRef]
- Wong AC, Devason AS, Umana IC, Cox TO, Dohnalová L, Litichevskiy L, Perla J, Lundgren P, Etwebi Z, Izzo LT, Kim J, Tetlak M, Descamps HC, Park SL, Wisser S, McKnight AD, Pardy RD, Kim J, Blank N, Patel S, Thum K, Mason S, Beltra JC, Michieletto MF, Ngiow SF, Miller BM, Liou MJ, Madhu B, Dmitrieva-Posocco O, Huber AS, Hewins P, Petucci C, Chu CP, Baraniecki-Zwil G, Giron LB, Baxter AE, Greenplate AR, Kearns C, Montone K, Litzky LA, Feldman M, Henao-Mejia J, Striepen B, Ramage H, Jurado KA, Wellen KE, O’Doherty U, Abdel-Mohsen M, Landay AL, Keshavarzian A, Henrich TJ, Deeks SG, Peluso MJ, Meyer NJ, Wherry EJ, Abramoff BA, Cherry S, Thaiss CA, Levy M. Serotonin reduction in post-acute sequelae of viral infection. Cell. 2023 Oct 26;186(22):4851-4867.e20. Epub 2023 Oct 16. PMID: 37848036; PMCID: PMC11227373. [CrossRef]
- Di Sabatino A, Giuffrida P, Vanoli A, Luinetti O, Manca R, Biancheri P, Bergamaschi G, Alvisi C, Pasini A, Salvatore C, Biagi F, Solcia E, Corazza GR. Increase in neuroendocrine cells in the duodenal mucosa of patients with refractory celiac disease. Am J Gastroenterol 2014, 109:258-69. [CrossRef]
- Zhang H, Shao B, Dang Q, Chen Z, Zhou Q, Luo H, Yuan W, Sun Z. Pathogenesis and Mechanism of Gastrointestinal Infection With COVID-19. Front Immunol 2021, 12:674074. [CrossRef]
- Zuo T, Zhan H, Zhang F, Liu Q, Tso EYK, Lui GCY, Chen N, Li A, Lu W, Chan FKL, Chan PKS, Ng SC. Alterations in fecal fungal microbiome of patients with COVID-19 during time of hospitalization until discharge. Gastroenterology 2020, 159:1302-1310.e5. [CrossRef]
- Christophersen A, Lund EG, Snir O, Solà E, Kanduri C, Dahal-Koirala S, Zühlke S, Molberg Ø, Utz PJ, Rohani-Pichavant M, Simard JF, Dekker CL, Lundin KEA, Sollid LM, Davis MM. Distinct phenotype of CD4+ T cells driving celiac disease identified in multiple autoimmune conditions. Nat Med 2019, 25:734-737. [CrossRef]
- Brunetti NS, Davanzo GG, de Moraes D, Ferrari AJR, Souza GF, Muraro SP, Knittel TL, Boldrini VO, Monteiro LB, Virgílio-da-Silva JV, Profeta GS, Wassano NS, Nunes Santos L, Carregari VC, Dias AHS, Veras FP, Tavares LA, Forato J, Castro IMS, Silva-Costa LC, Palma AC, Mansour E, Ulaf RG, Bernardes AF, Nunes TA, Ribeiro LC, Agrela MV, Moretti ML, Buscaratti LI, Crunfli F, Ludwig RG, Gerhardt JA, Munhoz-Alves N, Marques AM, Sesti-Costa R, Amorim MR, Toledo-Teixeira DA, Parise PL, Martini MC, Bispos-Dos-Santos K, Simeoni CL, Granja F, Silvestrini VC, de Oliveira EB, Faca VM, Carvalho M, Castelucci BG, Pereira AB, Coimbra LD, Dias MMG, Rodrigues PB, Gomes ABSP, Pereira FB, Santos LMB, Bloyet LM, Stumpf S, Pontelli MC, Whelan S, Sposito AC, Carvalho RF, Vieira AS, Vinolo MAR, Damasio A, Velloso L, Figueira ACM, da Silva LLP, Cunha TM, Nakaya HI, Marques-Souza H, Marques RE, Martins-de-Souza D, Skaf MS, Proenca-Modena JL, Moraes-Vieira PMM, Mori MA, Farias AS. SARS-CoV-2 uses CD4 to infect T helper lymphocytes. Elife 2023, 12:e84790. [CrossRef]
- Tavakolpour S, Rakhshandehroo T, Wei EX, Rashidian M. Lymphopenia during the COVID-19 infection: What it shows and what can be learned. Immunol Lett 2020, 225:31-32. [CrossRef]
- Stevens BR, Ellory JC, Preston RI. B0AT1 Amino Acid Transporter Complexed With SARS-CoV-2 Receptor ACE2 Forms a Heterodimer Functional Unit: In Situ Conformation Using Radiation Inactivation Analysis. Function (Oxf) 2021, 2(4):zqab027. [CrossRef]
- Qian Q, Fan L, Liu W, Li J, Yue J, Wang M, Ke X, Yin Y, Chen Q, Jiang C. Direct Evidence of Active SARS-CoV-2 Replication in the Intestine. Clin Infect Dis 2021, 73:361-366. [CrossRef]
- Petrillo M, Brogna C, Cristoni S, Querci M, Piazza O, Van den Eede G. Increase of SARS-CoV-2 RNA load in faecal samples prompts for rethinking of SARS-CoV-2 biology and COVID-19 epidemiology. F1000Res 2021, 10:370. [CrossRef]
- Natarajan A, Zlitni S, Brooks EF, Vance SE, Dahlen A, Hedlin H, Park RM, Han A, Schmidtke DT, Verma R, Jacobson KB, Parsonnet J, Bonilla HF, Singh U, Pinsky BA, Andrews JR, Jagannathan P, Bhatt AS. Gastrointestinal symptoms and fecal shedding of SARS-CoV-2 RNA suggest prolonged gastrointestinal infection. Med 2022, 3:371-387.e9. [CrossRef]
- Wilen CB, Lee S, Hsieh LL, Orchard RC, Desai C, Hykes BL Jr, McAllaster MR, Balce DR, Feehley T, Brestoff JR, Hickey CA, Yokoyama CC, Wang YT, MacDuff DA, Kreamalmayer D, Howitt MR, Neil JA, Cadwell K, Allen PM, Handley SA, van Lookeren Campagne M, Baldridge MT, Virgin HW. Tropism for tuft cells determines immune promotion of norovirus pathogenesis. Science. 2018, 360:204-208. [CrossRef]
- Ting HA, von Moltke J. The Immune Function of Tuft Cells at Gut Mucosal Surfaces and Beyond. J Immunol 2019, 202:1321-1329. [CrossRef]
- Petrillo M, Brogna C, Cristoni S, Querci M, Piazza O, Van den Eede G. Increase of SARS-CoV-2 RNA load in faecal samples prompts for rethinking of SARS-CoV-2 biology and COVID-19 epidemiology. F1000Res. 2021, 10:370. [CrossRef]
- Cabral-Marques O, Halpert G, Schimke LF, Ostrinski Y, Vojdani A, Baiocchi GC, Freire PP, Filgueiras IS, Zyskind I, Lattin MT, Tran F, Schreiber S, Marques AHC, Plaça DR, Fonseca DLM, Humrich JY, Müller A, Giil LM, Graßhoff H, Schumann A, Hackel A, Junker J, Meyer C, Ochs HD, Lavi YB, Scheibenbogen C, Dechend R, Jurisica I, Schulze-Forster K, Silverberg JI, Amital H, Zimmerman J, Heidecke H, Rosenberg AZ, Riemekasten G, Shoenfeld Y. Autoantibodies targeting GPCRs and RAS-related molecules associate with COVID-19 severity. Nat Commun 2022, 13:1220. [CrossRef]
- Szewczykowski C, Mardin C, Lucio M, Wallukat G, Hoffmanns J, Schröder T, Raith F, Rogge L, Heltmann F, Moritz M, Beitlich L, Schottenhamml J, Herrmann M, Harrer T, Ganslmayer M, Kruse FE, Kräter M, Guck J, Lämmer R, Zenkel M, Gießl A, Hohberger B. Long COVID: Association of Functional Autoantibodies against G-Protein-Coupled Receptors with an Impaired Retinal Microcirculation. Int J Mol Sci 2022, 23:7209. [CrossRef]
- Hohberger B, Harrer T, Mardin C, Kruse F, Hoffmanns J, Rogge L, Heltmann F, Moritz M, Szewczykowski C, Schottenhamml J, Kräter M, Bergua A, Zenkel M, Gießl A, Schlötzer-Schrehardt U, Lämmer R, Herrmann M, Haberland A, Göttel P, Müller J, Wallukat G. Case Report: Neutralization of Autoantibodies Targeting G-Protein-Coupled Receptors Improves Capillary Impairment and Fatigue Symptoms After COVID-19 Infection. Front Med (Lausanne). 2021, 8:754667. [CrossRef]
- Ballering AV, van Zon SKR, Olde Hartman TC, Rosmalen JGM; Lifelines Corona Research Initiative. Persistence of somatic symptoms after COVID-19 in the Netherlands: an observational cohort study. Lancet 2022, 400:452-461. [CrossRef]
- Toubal A, Nel I, Lotersztajn S, Lehuen A. Mucosal-associated invariant T cells and disease. Nat Rev Immunol 2019, 19:643-657. [CrossRef]
- Wang X, Liang M, Song P, Guan W, Shen X. Mucosal-associated invariant T cells in digestive tract: Local guardians or destroyers? Immunology 2023, 170:167-179. [CrossRef]
- Howson LJ, Salio M, Cerundolo V. MR1-Restricted Mucosal-Associated Invariant T Cells and Their Activation during Infectious Diseases. Front Immunol 2015, 6:303. [CrossRef]
- Harrison DG, Coffman TM, Wilcox CS. Pathophysiology of Hypertension: The Mosaic Theory and Beyond. Circ Res 2021, 128:847-863. [CrossRef]
- Santin, A. Spedicati, B. Pecori, A. Nardone, G.G. Concas, M.P. Piatti, G. Menini, A. Tirelli, G. Boscolo-Rizzo, P. Girotto, G. The Bittersweet Symphony of COVID-19: Associations between TAS1Rs and TAS2R38 Genetic Variations and COVID-19 Symptoms. Life 2024, 14, 219. [CrossRef]
- Klein J, Wood J, Jaycox JR, Dhodapkar RM, Lu P, Gehlhausen JR, Tabachnikova A, Greene K, Tabacof L, Malik AA, Silva Monteiro V, Silva J, Kamath K, Zhang M, Dhal A, Ott IM, Valle G, Peña-Hernández M, Mao T, Bhattacharjee B, Takahashi T, Lucas C, Song E, McCarthy D, Breyman E, Tosto-Mancuso J, Dai Y, Perotti E, Akduman K, Tzeng TJ, Xu L, Geraghty AC, Monje M, Yildirim I, Shon J, Medzhitov R, Lutchmansingh D, Possick JD, Kaminski N, Omer SB, Krumholz HM, Guan L, Dela Cruz CS, van Dijk D, Ring AM, Putrino D, Iwasaki A. Distinguishing features of long COVID identified through immune profiling. Nature 2023, 623:139-148. [CrossRef]
- Fedorowski A. Postural orthostatic tachycardia syndrome: clinical presentation, aetiology and management. J Intern Med 2019, 285:352-366. [CrossRef]
- Ormiston CK, Świątkiewicz I, Taub PR. Postural orthostatic tachycardia syndrome as a sequela of COVID-19. Heart Rhythm 2022, 19:1880-1889. [CrossRef]
- Lechner-Scott J, Levy M, Hawkes C, Yeh A, Giovannoni G. Long COVID or post COVID-19 syndrome. Mult Scler Relat Disord 2021, 55:103268. [CrossRef]
- Zubchenko S, Kril I, Nadizhko O, Matsyura O, Chopyak V. Herpesvirus infections and post-COVID-19 manifestations: a pilot observational study. Rheumatol Int 2022, 42:1523-1530. [CrossRef]
- Ryan JL, Shen YJ, Morgan DR, Thorne LB, Kenney SC, Dominguez RL, Gulley ML. Epstein-Barr virus infection is common in inflamed gastrointestinal mucosa. Dig Dis Sci 2012, 57:1887-98. [CrossRef]
- Fogarty H, Ward SE, Townsend L, Karampini E, Elliott S, Conlon N, Dunne J, Kiersey R, Naughton A, Gardiner M, Byrne M, Bergin C, O’Sullivan JM, Martin-Loeches I, Nadarajan P, Bannan C, Mallon PW, Curley GF, Preston RJS, Rehill AM, Baker RI, Cheallaigh CN, O’Donnell JS; Irish COVID-19 Vasculopathy Study (iCVS) Investigators. Sustained VWF-ADAMTS-13 axis imbalance and endotheliopathy in long COVID syndrome is related to immune dysfunction. J Thromb Haemost 2022, 20:2429-2438. [CrossRef]
- Motto DG, Chauhan AK, Zhu G, Homeister J, Lamb CB, Desch KC, Zhang W, Tsai HM, Wagner DD, Ginsburg G. Shigatoxin triggers thrombotic thrombocytopenic purpura in genetically susceptible ADAMTS13-deficient mice. J Clin Invest 2005, 115:2752-2761. [CrossRef]
- Català M, Mercadé-Besora N, Kolde R, Trinh NTH, Roel E, Burn E, Rathod-Mistry T, Kostka K, Man WY, Delmestri A, Nordeng HME, Uusküla A, Duarte-Salles T, Prieto-Alhambra D, Jödicke AM. The effectiveness of COVID-19 vaccines to prevent long COVID symptoms: staggered cohort study of data from the UK, Spain, and Estonia. Lancet Respir Med. 2024 Jan 11:S2213-2600(23)00414-9. Epub ahead of print. PMID: 38219763. [CrossRef]
- Cegolon L, Magnano G, Negro C, Larese Filon F; ORCHESTRA Working Group. SARS-CoV-2 Reinfections in Health-Care Workers, 1 March 2020-31 January 2023. Viruses. 2023 Jul 14;15(7):1551. PMID: 37515237; PMCID: PMC10384331. [CrossRef]
Figure 1.
Intestinal epithelial cell types. (reprinted with permission from Gerbe et al. 2012, ref. 71).
Figure 1.
Intestinal epithelial cell types. (reprinted with permission from Gerbe et al. 2012, ref. 71).

Figure 2.
Long Covid is a combination of multiple diseases.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



