
Submitted:
26 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Cytokinesis, the process by which the cytoplasm divides to generate two daughter cells after mitosis, is a crucial stage of the cell cycle. Successful cytokinesis must be coordinated with chromosome segregation and requires the fine orchestration of several processes, such as constriction of the actomyosin ring, membrane reorganisation, and, in fungi, cell wall deposition. In Saccharomyces cerevisiae, commonly known as budding yeast, septins play a pivotal role in the control of cytokinesis by assisting the assembly of the cytokinetic machinery at the division site and controlling its activity. Yeast septins form a collar at the division site that undergoes major dynamic transitions during the cell cycle. This review will discuss the functions of septins in yeast cytokinesis, their regulation and the implications of their dynamic remodelling for cell division.
Keywords:
S. cerevisiae
; septins
; cytokinesis
Introduction
Septins form an evolutionarily conserved family of cytoskeletal proteins expressed in many eukaryotic organisms but absent in land plants [1,2]. Septins were initially discovered in budding yeast through mutagenesis screens aimed at identifying genes involved in the cell division cycle (CDC; [3]). Temperature-sensitive mutations affecting the genes CDC3, CDC10, CDC11, and CDC12 cause hyperpolarized growth and cytokinesis failure at high temperatures, suggesting a role in cell morphogenesis and cell division [4,5]. Electron and immunofluorescence microscopy studies revealed that the products of these genes localize at the bud neck, forming a filamentous collar [6,7,8]. Given their role in septation, these proteins were named "septins." Subsequently, three additional septin genes were identified in budding yeast based on sequence homology: SHS1, expressed like the aforementioned septins in vegetative cells, and SPR3 and SPR28, which are expressed exclusively during meiosis [9,10,11].
Yeast septins participate in a wide range of cellular functions, including cell polarity [12], cell cycle progression [13,14,15], spindle positioning [16], cytokinesis and septum formation [4,17,18,19]. Since their discovery in budding yeast, septin orthologs have been identified in animals, fungi and protists, where they are involved in numerous cellular processes, such as cytokinesis [20,21,22,23,24,25], exocytosis [26], phagocytosis [27], morphogenesis [28,29,30,31,32], ciliogenesis [33,34,35], cell motility [36,37], spermatogenesis [38], lipid metabolism [39,40], synaptic activity [41,42,43] and bacterial entrapment during cell infection [44,45].
Given their pivotal roles in cellular organisation and function, it is not surprising that septins are implicated in a range of pathological conditions. Indeed, septin dysfunction has been associated with cancer, neurodegenerative disorders, infectious diseases and infertility (reviewed in [46]).
Septins are GTP-binding proteins characterized by a conserved GTPase domain and variable N-terminal and C-terminal regions. A septin unique element is a distinctive feature of all septins, although its precise function remains unknown [47]. Septins invariably assemble into rod-shaped hetero-complexes composed of two copies of each septin monomer organised in a palindromic fashion (reviewed in [48]). Septin hetero-complexes then interact to form non-polar filaments and/or higher-order structures, such as bundles, rings or gauzes, as observed both in vitro and in vivo across various organisms [6,22,24,31,38,49,50,51,52,53,54,55,56,57,58,59,60].
Membranes play a crucial role in septin organisation. Septins associate with lipid membranes containing negatively charged phospholipids, particularly phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) [61,62,63,64,65]. In budding yeast, PI(4,5)P2 is enriched at the presumptive bud site in G1 [66], thus accounting for septin recruitment at this specific cellular location. Additionally, perturbing the amount of PI(4,5)P2 at the plasma membrane causes the septin collar to be disassembled at the bud neck and/or septin filaments to form aberrant structures cytoplasm, such as small rings and arcs [64,67], indicating that PI(4,5)P2 is crucial for septin organisation in vivo.
How septins bind to membranes is unclear. A protein motif rich in basic residues at the N-terminus of septins has been implicated in membrane binding via electrostatic interactions with the negative charges of phospholipids [63,68,69,70]. However, whether this polybasic motif is actually exposed on the surface of septins or buried, thus precluding a prominent role in membrane binding, is still debated [48].
Septin binding to membranes is also affected by membrane curvature, showing a strong preference for micrometric curvatures [71,72,73]. Consequently, septins are frequently located at curved membranes, such as the hyphal branch sites of filamentous fungi, the base of primary cilia, and the annulus of sperm tails [31,32,33,50,74,75].
Finally, multiple septin-associated proteins and post-translational modifications are likely involved in septin organisation in different physiological contexts, highlighting the complexity and dynamic nature of septin regulation.
Budding Yeast Septins and their Localisation
In vegetative cells, budding yeast septins form two hetero-octamers with a common core composed of Cdc12-Cdc3-Cdc10-Cdc10-Cdc3-Cdc12 and comprising either Cdc11 or Shs1 at both extremities [58,59,76]. Surprisingly, septin octamers have always symmetric termini, while asymmetric complexes capped by Cdc11 and Shs1 at each end cannot be formed, at least in vitro [76]. The linear septin complexes are often referred to as septin “rods” and measure 32-35 nm in length and 4-5 nm in diameter [58,77]. In vitro reconstitution assays using recombinant septins have shown a striking difference between Cdc11-capped and Shs1-capped yeast octamers, where the former can spontaneously polymerise end-to-end to form long paired filaments in solution, while the latter appear unable polymerise [50,58,59,78,79]. Shs1-capped octamers can nevertheless interact laterally and stagger on top of each other to form curved bundles, rings and spirals in vitro [77]. Additionally, in the presence of lipids or upon an SHS1 mutation mimicking constitutive phosphorylation of Ser259, Shs1-capped octamers form gauze-like structures in addition to rings, suggesting that the cellular context and post-translational modifications influence the supramolecular organisation of septins [60,77]. Despite being unable to polymerise into linear filaments, Shs1-capped rods can form heterotypic interactions with the termini of Cdc11-capped octamers [79], suggesting that they may be interspersed in septin filaments and higher-order structures. Consistently, the five mitotic septins display a completely superimposable localisation in vegetative cells throughout the cell cycle.
Formation of septin filament is essential for cell division, as shown by the lethality of mutants incapable of septin polymerisation [80]. This may explain why Cdc11 is required for viability in most yeast backgrounds, while Shs1 is dispensable [10,47,80]. During sporulation, the aforementioned septin hetero-octamers are replaced by the Spr28-Spr3-Cdc3-Cdc10-Cdc10-Cdc3-Spr3-Spr28 meiotic-specific complex, where Spr28 and Spr3 replace the terminal septin and Cdc12, respectively [81]. Unlike its mitotic Cdc11-capped counterpart, the Spr28-capped meiotic complex is unable to polymerise in solution but it readily forms filaments on PI(4,5)P2-containing lipid monolayers [81], suggesting that its polymerisation requires binding to the plasma membrane. Its function during meiosis is linked to proper biogenesis of the prospore membrane and cell wall assembly, thereby ensuring high sporulation efficiency [82].
The functional organisation of septins is based on their assembly into higher-ordered structures. In vegetative cells, septins undergo dynamic changes in their structural organisation during the cell cycle. First, in late G1 septins form a dynamic cortical ring at the presumptive bud site. Then, as the bud emerges the septin ring expands into an hourglass-shaped structure, referred to as septin collar, around the bud neck. Finally, at the onset of cytokinesis the septin collar splits into a double ring that sandwiches the AMR (Figure 1). This dramatic septin remodelling, which is essential for cell division (see below), is often referred to as septin ring splitting. Each split ring then persists in the mother and daughter cell, respectively, until the next cell cycle [7,83,84]. Florescence recovery after photobleaching (FRAP) experiments have shown that the newly forming septin ring is dynamic, as shown by its ability to exchange septins from an unassembled pool. In contrast, the septin collar is a stable and immobile structure. At cytokinesis, split septin rings become again relatively dynamic [85,86].
In cells exposed to mating pheromones, septins form a fuzzy band or a set of parallel bars at the base of the polarized projection (i.e. the shmoo) [7,84,87]. Upon cell fusion during mating, septins form an annulus at the midzone of the zygote that seems to affect the redistribution of supramolecular complexes and organelles [88].
During meiosis I, septins first form ring-like structures at the leading edge of membrane sacs, also known as prospore walls, that originate in close apposition to the cytoplasmic face of the spindle pole bodies and that will later extend to engulf each of the four haploid nuclei. At the end of meiosis II, septins seem to localize quite uniformly at the plasma membrane surrounding the developing spore [9,89].
Thus, budding yeast septins display a wide range of high order architectures depending on the physiological context, but they seem to be invariably associated with membranes, while in other eukaryotes septins clearly associate also with the actin and microtubule cytoskeleton (reviewed in [90]).
Septin Organisation and Cytokinesis in Budding Yeast
Our current knowledge of septin architecture at the bud neck of mitotic yeast cells comes mainly from ultrastructural studies. Freeze-fracture and platinum-replica electron microscopy on unroofed spheroplasts or cryo-electron tomography on intact cells have shown that the mature septin collar at the bud neck is made by a network of axial filaments lying parallel to the mother-bud axis and circumferential filaments that are orthogonally oriented, thus forming a gauze-like meshwork (Figure 2) [49,52,91]. Conversely, the septin double ring at cytokinesis is made exclusively of circumferential filaments [91], in line with polarized fluorescence microscopy studies showing that the average orientation of septin filaments in the split septin rings is perpendicular relative to the polarity axis [92,93,94]. Since fluorescently-labelled septins show a marked decrease in fluorescence at the bud neck at the moment of septin ring splitting [19,86,93,95], current models envision that at the onset of cytokinesis the axial septin filaments disassemble and possibly partially reassemble into two arrays of circumferential filaments, forming the double ring (reviewed in [96]).
Cytokinesis is a delicate process that must precisely partition an equal complement of the replicated genomes and organelles to each daughter cell. It relies on assembly and contraction of an actomyosin ring (AMR) and must be tightly controlled and coordinated with other cell cycle processes, such as chromosome segregation. Typically, yeast cytokinesis can be divided into four steps: selection of the division site, assembly of the AMR, cleavage furrow ingression and primary septum formation powered by the AMR, and localized membrane remodeling. In budding yeast, septins are involved in most of these cytokinetic steps by adopting different structural organisations.
The first steps of cytokinesis occur very early during cell cycle. The selection of the bud site, which is also the division site, is determined in late G1 by the cell polarization machinery controlled by the master polarity GTPase Cdc42 that also recruits septins (reviewed in [97]; see below). Bud site selection follows a very specific pattern depending on whether cells are haploid or diploid. In haploid cells the emergence of a new bud occurs proximally to the division site (axial budding pattern), while in diploid cells it occurs distally (bipolar budding pattern). Such a stereotypical pattern depends on polarity landmark proteins (reviewed in [97]). Despite their recruitment before bud emergence, septins are not required for bud emergence [4,98], but they contribute to the axial budding pattern by recruiting to the bud neck landmarks proteins, such as Bud3 and Bud4 [99,100,101,102]. In addition, they facilitate the formation of a narrow cluster of Cdc42 at the presumptive bud site by recruiting GTPase-activating proteins (GAPs) that in turn inhibit Cdc42 in the membrane area surrounding its activated cluster (Figure 2) [103].
Before bud emergence, Myo1, the sole myosin-II heavy chain in budding yeast, is recruited along with its regulatory light chain Mlc2 in a septin-dependent manner, forming a ring (Figure 1) [17,104,105,106]. At the time of the bud emergence, the septin ring expands around the bud neck forming an hourglass-shaped structure that remains stable during mitosis. AMR assembly proceeds until the end of anaphase (Figure 1), when the IQGAP Iqg1 appears at the bud neck to recruit actin [17,104,107,108]. Around the same time, the essential myosin light chain Mlc1 arrives at the bud neck to interact with Myo1 and to enhance Myo1 targeting to the neck that further increases its local levels [105,109]. Concomitantly, Mlc1 also promotes Iqg1 recruitment to the bud neck by binding to its IQ motifs [110]. Two formins, Bni1 and Bnr1, are redundantly essential for actin polymerisation at the AMR [111,112]. Furthermore, formins have been shown to contribute to the accumulation of Mlc1 at the bud neck during cytokinesis [113], suggesting an additional mechanism by which they could participate to AMR assembly. After mitotic exit, the chitin synthase Chs2 is recruited to the AMR to promote formation of the primary septum along with AMR constriction [114,115,116,117,118]. Additionally, a protein complex made by Hof1, Inn1 and Cyk3 also joins the AMR and partially constricts alongside it to boost Chs2 recruitment and activity [112,115,119,120,121,122,123,124,125,126,127]. Several of the aforementioned AMR components and regulators, namely Myo1 [128,129], Mlc1 [107,130], Bnr1 [131,132] and Hof1 [133], require septins for their bud neck localisation, suggesting that the septin collar orchestrates AMR assembly.
Constriction of the AMR marks the beginning of cytokinesis (Figure 1). AMR constriction may occur partly through sliding of bipolar myosin filaments along actin filaments that are tethered to the plasma membrane, similar to the way by which actomyosin generates force in the striated muscle [134,135]. However, actin depolymerisation by cofilin plays a predominant role in budding yeast AMR constriction [136]. Iqg1 degradation by the APC/C complex might also contribute to AMR disassembly [137,138].
AMR constriction drives membrane invagination inward and is also coupled to targeted deposition of post-Golgi vesicles to the division site to increase surface area and deliver the chitin synthase-II Chs2 to drive primary septum formation (PS) [117,139,140]. Thus, AMR constriction guides PS formation, that in turn stabilises the AMR during constriction [141,142,143].
At the onset of cytokinesis, the septin hourglass is split into two rings that sandwich the AMR (Figure 1) [83,95,144]. This septin reorganisation is an essential prerequisite for cytokinesis [95,145]. Indeed, lack of septin ring splitting prevents AMR constriction and PS formation. Thus, the septin collar has both a positive and a negative role in cytokinesis: during mitosis it organises the cytokinetic machinery by recruiting several cytokinetic proteins to the bud neck, but at cytokinesis it must be displaced from the bud neck (through septin ring splitting or clearance altogether) to allow AMR constriction [95,145]. Consistently, septins are not required for AMR constriction once the AMR has been assembled [95,146]. The septin double ring could nevertheless facilitate plasma membrane closure by acting as a diffusion barrier that concentrates membrane remodeling factors, such as the polarisome and the exocyst complex, to the cleavage site [146].
Once the primary septum has been laid down, a secondary septum (SS) is assembled by glucan synthases and the chitin synthase Chs3 on both the mother and daughter sides of the PS, thus leading to abscission (Figure 1) [139,147]. By recruiting Hof1 to the bud neck, which in turn inhibits precocious activation of Chs3, septins indirectly set the timing of SS assembly, so that it occurs only after the PS has been deposited [120,148].
The final step of yeast cytokinesis is cell separation (Figure 1), which involves PS digestion and SS remodeling by chitinase and endoglucanases, respectively (reviewed in [149]). If and how septins are involved in the control of cell separation is not known. Intriguingly, however, in fission yeast glucanases are localized in between the two split septin rings and are unable to form a proper ring at the cortex in septin mutants [150], suggesting that the septin double ring may form a diffusion barrier for the enzymes involved in cell separation.
The control of Septin Dynamics During the Cell Cycle
We can distinguish three main steps in the dynamics of septin architecture during the budding yeast cell cycle: 1. Septin recruitment and ring assembly; 2. Maturation into the septin collar; 3. Transition into the septin double ring.
Septin Recruitment and Ring Assembly
Septins accumulate at the presumptive bud site initially as a cloud or patch (Figure 2). This process depends on the Rho GTPase Cdc42 that is a conserved master regulator of cell polarity [83,85,151]. Indeed, Cdc42 depletion leads to an unbudded cell phenotype lacking cortical septin structures [152], and cdc42 mutants are defective in septin recruitment to the presumptive bud site or display septin mislocalization [99,151,153]. However, in some of these mutants Cdc42 cannot hydrolyse GTP and is locked in a GTP-bound state, while overexpression of Cdc42 GTPase-activating proteins (GAPs) can restore proper septin localization and function in septin mutants [151,152]. These observations have led to the proposal that Cdc42 must continuously cycle between a GTP- and a GDP-bound state in order to gather septins at the future bud neck [153]. Alternatively, active GTP-bound Cdc42 may be required to establish the first polarity cue necessary for septin recruitment (e.g. to define a membrane domain permissive for septin accumulation), while the active GTPase actually prevents septins from piling up.
Once at the plasma membrane, septins rapidly organise into a cortical ring (Figure 2). Polarized exocytosis and insertion of membrane vesicles by Cdc42 and the exocyst complex are critical to this process by inhibiting septin accumulation in the center of the septin cap at the incipient bud site, thus creating the ring [103]. Thus, it is possible that the septin mislocalization phenotype of some GTP-bound cdc42 mutants is accounted for by excessive or untargeted membrane vesicles tethering that disrupts septin ring assembly.
Several studies have attempted to link the function of Cdc42 in septin ring assembly to specific Cdc42 effectors (Figure 2). The p21-activated kinases (PAK) Cla4 and Ste20 are turned on by Cdc42 and Cla4 has been implicated in septin recruitment to the presumptive bud site through phosphorylation of several septins [98,154,155,156]. On the other hand, Cla4 binds the Cdc42 membrane scaffold Bem1 and recruits and phosphorylates Cdc24, the GEF (guanine-nucleotide exchange factor) for Cdc42, thus promoting its GTP-bound state [157,158,159]. Such a positive feedback loop suggests that the role of PAK kinases in septin recruitment may be partly mediated by a global enhancement in Cdc42 activity. In parallel, the Cdc24-Bem1 complex binds Cdc11, helping to bring septins to the bud site [160]. Two additional Cdc42 effectors, i.e. the paralogous membrane proteins Gic1 and Gic2, interact directly with septins and are involved in septin recruitment and organisation in vivo and in vitro [103,152,161,162,163]. Accordingly, septin deposition and budding mostly fail in gic1 gic2 double mutants at high temperatures [152]. Gic1 colocalizes with septins at the presumptive bud site at early stages of the cell cycle and at the bud neck later on [152], and it has been shown to bundle and crosslink septin filaments in vitro, thereby stabilising them [163]. However, similar to PAK kinases, Gic1 and Gic2 are also involved in a positive feedback loop for Cdc42 activation, and overexpression of CDC42 suppresses the lethality of cells lacking Gic1 and Gic2 at high temperatures [164,165], suggesting that septin recruitment to the future bud site may be empowered by Cdc42 itself or by effectors other than Gic proteins.
PAK kinases possess a pleckstrin-homology domain that binds preferentially to PI(4,5)P2 [166]. Similarly, Gic2 (and possibly Gic1) bears a cluster of basic residues that binds to PI(4,5)P2 [162]. Thus, the PI(4,5)P2 binding properties of these Cdc42 effectors might influence their interaction with and organisation of septin into a ring.
The formin Bni1 contributes to septin ring formation along with Cla4 (Figure 2), as bni1 cla4 double mutants form a cap of septins at the presumptive bud site that is not converted into a ring [155]. This could be related to the function of Cdc42 in polarized exocytosis along actin cables generated by Bni1 and in creating the hole in the middle of the septin ring [103].
Finally, Axl2 is a bud site landmark protein that also participates to septin recruitment (Figure 2) [167,168]. Recent two-hybrid and bimolecular fluorescence complementation (BiFC) assays indicate that Axl2 interacts with the septin Cdc10, as well as with Cdc42-GTP and Bud3, and contributes to the efficient gathering of septins to the cell division site [169].
In summary, while Cdc42 effectors, activators and cell polarity proteins have been implicated in septin organisation, the precise mechanistic processes underlying septin recruitment and septin ring formation need to be further elucidated.
Another Rho GTPase, Rho1, has been implicated in timely and efficient septin recruitment and septin collar stability through activation of one of its downstream effectors, the kinase Pkc1 that in turn phosphorylates the septin-interacting protein Syp1 and promotes its turnover at the bud neck [170,171]. Syp1 appears at the presumptive bud site concomitant with septins and, in vitro, is able to align laterally and bundle septin filaments, thus offering mechanical stability to the new-forming septin ring/collar [172].
Maturation Into the Septin Collar
As the bud emerges, the septin ring at the bud neck expands into a rigid hourglass-shaped septin collar. Whether this septin remodelling occurs suddenly at the G1/S transition or progressively throughout mitotic progression is not known. As mentioned above, the mature septin hourglass is a gauze-like, very stable septin assembly comprising axial and circumferential filaments (Figure 2) [49,85,86,91]. Likely, the transition from the initial septin ring, which is mainly composed by radial septin filaments converging towards a central hole [91], into the septin collar is controlled by the cell cycle machinery.
One of the first septin-interacting proteins that has been involved in septin organisation is the Nim-related kinase Gin4. Mutations in GIN4 are synthetically lethal with cdc12 and cause septins to form a fuzzy band or parallel axial “bars” at the bud neck instead of the hourglass [87]. A similar phenotype has been observed in cells lacking the Gin4-binding protein Nap1 [14] or the septin Shs1 [13] and in cells exposed to mating pheromones [7,84,87]. These observations suggest that Gin4 may be involved in stabilising the circumferential septin filaments, which in turn require Shs1 for the overall gauze-like organisation in a septin collar [91,93]. Consistently, Gin4 phosphorylates directly Shs1, thus contributing to the robustness of the hourglass [173,174]. Gin4 colocalises with septins from septin appearance throughout mitosis, but is displaced from the bud neck before septin ring splitting and is absent in septin structures of mating cells [87]. Its association with septins requires Nap1, Shs1, the PAK kinase Cla4 and cyclin-dependent kinases (CDKs) [173], thus partly explaining the links between septin collar organisation and cell cycle progression. Accordingly, Shs1 phosphorylation by G1 CDKs stabilises the interaction of septins with Gin4 [175]. Two additional Nim-related kinases, Hsl1 and Kcc4, associate with septins in yeast cells [176,177,178,179,180,181,182]. Although lack of Hsl1 or Kcc4 does not cause any obvious defect in septin organisation [14], these kinases may share with Gin4 partially overlapping functions in building up the architecture of the septin collar [179].
During bud emergence, Gin4 targets to the bud neck the LKB1-like kinase Elm1, which contributes to septin collar architecture and stability until cytokinesis (Figure 2) [183,184,185,186]. In turn, Elm1 phosphorylates and activates Cla4, Gin4 and Hsl1 [14,173,174,185,186,187]. Its efficient recruitment to the bud neck requires the Dma1 and Dma2 ubiquitin ligases, which in turn are involved in proper septin stability at the bud neck through an unknown molecular mechanism [188,189]. The role of Elm1 in septin organisation also involves phosphorylation and functionality of Bni5 [184,190], a septin-interacting protein that was identified as a dosage suppressor of septin mutants [191]. Bni5 can dimerize and in vitro is able to crosslinks septin filaments into networks by bridging pairs or multiple filaments [190,192]. Additionally, Bni5 interacts with the C-terminal extension of Cdc11 and Shs1 and recruits Myo1 to the septin ring and collar throughout most of the cell cycle except during cytokinesis [109,193]. The exact role of Myo1 at the septin collar is unclear, but myosin II filaments may assemble on the membrane-distal side of the septin hourglass perpendicular to the axial septin filaments and somehow contribute to the appropriate septin architecture [91].
Post-translational modifications (PTMs) likely play an important role in the organisation and stability of the septin hourglass. Besides the aforementioned septin phosphorylations by Cla4, Gin4 and CDKs, septins may be phosphorylated by other septin- and bud neck-associated kinases (e.g. Hsl1, Kcc4, polo kinase Cdc5, Mob1-Dbf2, etc.). Plenty of septin phosphorylation sites have been identified by phosphoproteomics and some them are regulated across the cell cycle [194,195,196,197,198,199,200,201,202,203], suggesting that they may play an important role in the control of septin dynamics. However, their elevated number has hampered so far their systematic characterization. Septin sumoylation occurs mainly during mitosis and could potentially promote the stability of the septin hourglass or septin interactions [204]. However, non-sumoylatable septin mutants do not show any obvious defects in septin architecture and experience only a mild delay in septin disassembly after cell division [204]. Septin acetylation may also impact the overall architecture of the septin collar, as septin mutants with reduced acetylation levels have defects in septin localization [205]. Finally, some septins have also been found to be ubiquitylated [196], but the functional significance of this PTM has not been explored so far.
Since the septin collar is necessary for the recruitment of several cytokinetic proteins to the bud neck (see above), it is often considered as a “scaffold” for cytokinesis. However, septins actually inhibit AMR constriction and are no longer required for cytokinesis after AMR assembly [95,146]. Thus, the term “scaffold” seems inappropriate. Why certain cytokinetic factors require to bind the septin collar in order to be incorporated into the AMR remains to be further investigated. Possibly, the septin collar congregates these proteins at the bud neck to reach threshold concentrations necessary for AMR assembly, thus acting as a molecular crowder. A non-mutually exclusive hypothesis is that binding to septins may induce specific protein conformations that are compatible with AMR interactions.
Transition Into the Septin Double Ring
During cytokinesis, the septin collar undergoes a sudden and dramatic reorganisation, resulting in its splitting into two distinct rings that sandwich the AMR (Figure 2) [129,144]. This septin displacement is crucial for AMR constriction and cytokinesis, indicating that building a force-generating AMR requires at least two septin-related steps: initially, the septin collar spurs the assembly of the cytokinetic machinery at the right place while preventing AMR-driven membrane ingression; subsequently, the confined eviction of septins from the division site during septin ring splitting initiates AMR constriction [95,145].
Formation and maintenance of the double septin ring depend on the anillin homologue Bud4 and the Rho-GEF Bud3 (Figure 2) [19,93,206,207,208], which had been previously implicated in the axial budding pattern of haploid yeast cells [101,207,209]. In the absence of Bud3 and Bud4 the septin collar disassembles completely at mitotic exit, without any obvious consequence on the kinetics of AMR constriction and cytokinesis [206], in line with the notion that septin clearance after AMR assembly is not detrimental for cytokinesis. Both Bud3 and Bud4 localise as a double ring at the edges of the septin collar during mitosis and then remain associated with the split septin rings, suggesting that these proteins act as spatial cues to pre-pattern the septin double ring [100,101,206,207,210,211].
Septin ring splitting requires the activity of the Mitotic Exit Network (MEN; Figure 2), a Hippo-like pathway triggered by a top GTPase (Tem1) that turns on two sequentially acting protein kinases (Cdc15 and Mob1-Dbf2) to ultimately promote the activation of the Cdc14 phosphatase (reviewed in [212]). In turn, Cdc14 dephosphorylates and activates its upstream kinases Cdc15 and Mob1-Dbf2 in a positive feedback loop [213,214]. Cdc14 is the main CDK-counteracting phosphatase in budding yeast and its function, together with that of its upstream MEN regulators, is essential for mitotic exit and cytokinesis [215,216,217,218,219,220]. Mitotic exit elicits several processes, such as spindle disassembly and licensing of replication origins, and is an essential prerequisite for cytokinesis through dephosphorylation of specific mitotic CDK substrates [221]. However, MEN promotes septin ring splitting also independently of its role in mitotic exit [95], suggesting that one or more MEN targets prompt septin remodelling at cytokinesis. We have recently identified the cytokinetic protein Hof1 as one of such septin ring splitting regulators (Figure 2) [222]. Hof1 is involved in cytokinesis and is an established target of MEN [112,127,223,224]. It plays a major role in the control of actin polymerisation and bundling, partly by modulating formin activity [225,226,227,228,229,230]. Consistently, it has been implicated in polarised growth [228,231]. However, the cytokinetic function of Hof1 was mainly linked to activation of the chitin synthase Chs2, and CHS2 overexpression or gain of function mutations can suppress the lethality of hof1Δ mutants at high temperatures [126,232]. Our recent data indicate that hof1 mutants also display defects in septin remodelling at cytokinesis [222]. During mitosis, Hof1 associates with septins, forming two closely spaced rings at the edges of the septin collar [133,222,223,233]. In vitro, it can induce the formation of intertwined septin bundles, suggesting that during this cell cycle stage it may contribute to the robustness of the septin hourglass [222,229]. However, shortly before septin ring splitting Hof1 is displaced from the septin collar and relocates to the AMR, where it partially constricts alongside it [133,222,223,233]. This rapid translocation from septins to the AMR is triggered by Hof1 phosphorylation, primarily by the MEN kinase Dbf2, which disengages Hof1 from the septins allowing it to join the AMR [222,223,224]. Phospho-mimicking HOF1 mutant alleles can bypass the septin reorganisation defects seen in MEN mutants by displacing Hof1 from septins and enhancing its translocation to the AMR. Importantly, septin remodelling by Hof1 depends on its membrane-binding F-BAR domain, suggesting that a local membrane remodeling could underlie septin disassembly from the cleavage site and remodelling into the double ring [222]. Since BAR domains can induce membrane curvature [234,235], Hof1 may bend the membrane at the division site to a radius that causes septin disassembly. Another possibility, stemming from the ability of BAR domains to cluster phosphoinositides [236,237], is that Hof1 may promote septin ring splitting by modifying the local composition of the plasma membrane. The non-essentiality of HOF1, especially in some strain background, suggests that additional, as yet unidentified proteins participate to septin ring splitting alongside Hof1.
Conclusions and Perspectives
Cytokinesis is a fundamental, yet vulnerable process that requires the orchestration of many players. Given the prominent role of septins in budding yeast cytokinesis, understanding how septin remodeling is controlled during the yeast cell cycle is key in order to gain insights into how cytokinesis is regulated in time and space. Unresolved questions that deserve further investigation regard the precise mechanism underlying septin-membrane interactions, the impact of septin-binding proteins and post-translational modifications on septin structural organisation, and the possible interplay between septins and other cytoskeletal elements. Unravelling how the yeast septin collar inhibits AMR constriction is another critical question for future research. As we continue to tease apart the complexity of septin biology, their integral role in cellular organisation and function becomes increasingly apparent, opening exciting avenues for our comprehension of septin functions in physiological and pathological conditions.
Acknowledgments
Work in Piatti’s lab is supported by the Agence Nationale pour la Recherche (ANR project CONSTRICT) and Fondation pour la Recherche Médicale (FRM project EQU202303016309) to S.P. M.V.S has been supported by Fondation pour la Recherche Médicale (FRM project FDT202106012881).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nishihama, R.; Onishi, M.; Pringle, J.R. New Insights into the Phylogenetic Distribution and Evolutionary Origins of the Septins. Biol Chem 2011, 392, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Shuman, B.; Momany, M. Septins From Protists to People. Frontiers in Cell and Developmental Biology 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H.; Culotti, J.; Reid, B. Genetic Control of the Cell-Division Cycle in Yeast, I. Detection of Mutants. Proc Natl Acad Sci U S A 1970, 66, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H. Genetic Control of the Cell Division Cycle in Yeast. IV. Genes Controlling Bud Emergence and Cytokinesis. Exp Cell Res 1971, 69, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H.; Mortimer, R.K.; Culotti, J.; Culotti, M. Genetic Control of the Cell Division Cycle in Yeast: V. Genetic Analysis of Cdc Mutants. Genetics 1973, 74, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Byers, B.; Goetsch, L. A Highly Ordered Ring of Membrane-Associated Filaments in Budding Yeast. The Journal of cell biology 1976, 69, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Ford, S.K.; Pringle, J.R. Cellular Morphogenesis in the Saccharomyces Cerevisiae Cell Cycle: Localization of the CDC11 Gene Product and the Timing of Events at the Budding Site. Dev Genet 1991, 12, 281–292. [Google Scholar] [CrossRef]
- Haarer, B.K.; Pringle, J.R. Immunofluorescence Localization of the Saccharomyces Cerevisiae CDC12 Gene Product to the Vicinity of the 10-Nm Filaments in the Mother-Bud Neck. Molecular and cellular biology 1987, 7, 3678–3687. [Google Scholar] [PubMed]
- De Virgilio, C.; DeMarini, D.J.; Pringle, J.R. SPR28, a Sixth Member of the Septin Gene Family in Saccharomyces Cerevisiae That Is Expressed Specifically in Sporulating Cells. Microbiology (Reading) 1996, 142 ( Pt 10) Pt 10, 2897–2905. [Google Scholar] [CrossRef]
- Mino, A.; Tanaka, K.; Kamei, T.; Umikawa, M.; Fujiwara, T.; Takai, Y. Shs1p: A Novel Member of Septin That Interacts with Spa2p, Involved in Polarized Growth inSaccharomyces Cerevisiae. Biochemical and Biophysical Research Communications 1998, 251, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Ozsarac, N.; Bhattacharyya, M.; Dawes, I.W.; Clancy, M.J. The SPR3 Gene Encodes a Sporulation-Specific Homologue of the Yeast CDC3/10/11/12 Family of Bud Neck Microfilaments and Is Regulated by ABFI. Gene 1995, 164, 157–162. [Google Scholar] [CrossRef]
- Barral, Y.; Mermall, V.; Mooseker, M.S.; Snyder, M. Compartmentalization of the Cell Cortex by Septins Is Required for Maintenance of Cell Polarity in Yeast. Molecular Cell 2000, 5, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Castillon, G.A.; Adames, N.R.; Rosello, C.H.; Seidel, H.S.; Longtine, M.S.; Cooper, J.A.; Heil-Chapdelaine, R.A. Septins Have a Dual Role in Controlling Mitotic Exit in Budding Yeast. Curr Biol 2003, 13, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; Theesfeld, C.L.; McMillan, J.N.; Weaver, E.; Pringle, J.R.; Lew, D.J. Septin-Dependent Assembly of a Cell Cycle-Regulatory Module in Saccharomyces Cerevisiae. Mol Cell Biol 2000, 20, 4049–4061. [Google Scholar] [CrossRef] [PubMed]
- Sakchaisri, K.; Asano, S.; Yu, L.-R.; Shulewitz, M.J.; Park, C.J.; Park, J.-E.; Cho, Y.-W.; Veenstra, T.D.; Thorner, J.; Lee, K.S. Coupling Morphogenesis to Mitotic Entry. Proceedings of the National Academy of Sciences 2004, 101, 4124–4129. [Google Scholar] [CrossRef]
- Kusch, J.; Meyer, A.; Snyder, M.P.; Barral, Y. Microtubule Capture by the Cleavage Apparatus Is Required for Proper Spindle Positioning in Yeast. Genes Dev 2002, 16, 1627–1639. [Google Scholar] [CrossRef] [PubMed]
- Bi, E.; Maddox, P.; Lew, D.J.; Salmon, E.D.; McMillan, J.N.; Yeh, E.; Pringle, J.R. Involvement of an Actomyosin Contractile Ring in Saccharomyces Cerevisiae Cytokinesis. J Cell Biol 1998, 142, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- DeMarini, D.J.; Adams, A.E.M.; Fares, H.; Virgilio, C.D.; Valle, G.; Chuang, J.S.; Pringle, J.R. A Septin-Based Hierarchy of Proteins Required for Localized Deposition of Chitin in the Saccharomyces Cerevisiae Cell Wall. The Journal of Cell Biology 1997, 139, 75. [Google Scholar] [CrossRef] [PubMed]
- Wloka, C.; Nishihama, R.; Onishi, M.; Oh, Y.; Hanna, J.; Pringle, J.R.; Krauss, M.; Bi, E. Evidence That a Septin Diffusion Barrier Is Dispensable for Cytokinesis in Budding Yeast. Biol Chem 2011, 392, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Estey, M.P.; Di Ciano-Oliveira, C.; Froese, C.D.; Bejide, M.T.; Trimble, W.S. Distinct Roles of Septins in Cytokinesis: SEPT9 Mediates Midbody Abscission. Journal of Cell Biology 2010, 191, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Joo, E.; Surka, M.C.; Trimble, W.S. Mammalian SEPT2 Is Required for Scaffolding Nonmuscle Myosin II and Its Kinases. Developmental Cell 2007, 13, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Karasmanis, E.P.; Hwang, D.; Nakos, K.; Bowen, J.R.; Angelis, D.; Spiliotis, E.T. A Septin Double Ring Controls the Spatiotemporal Organization of the ESCRT Machinery in Cytokinetic Abscission. Current Biology 2019, 29, 2174–2182. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Froese, C.D.; Estey, M.P.; Trimble, W.S. SEPT9 Occupies the Terminal Positions in Septin Octamers and Mediates Polymerization-Dependent Functions in Abscission. The Journal of cell biology 2011, 195, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, M.J.; Liu, J.; Lavoie, B.D.; Wilde, A. Anillin-Dependent Organization of Septin Filaments Promotes Intercellular Bridge Elongation and Chmp4B Targeting to the Abscission Site. Open Biol 2014, 4, 130190. [Google Scholar] [CrossRef] [PubMed]
- Surka, M.C.; Tsang, C.W.; Trimble, W.S. The Mammalian Septin MSF Localizes with Microtubules and Is Required for Completion of Cytokinesis. Mol Biol Cell 2002, 13, 3532–3545. [Google Scholar] [CrossRef] [PubMed]
- Tokhtaeva, E.; Capri, J.; Marcus, E.A.; Whitelegge, J.P.; Khuzakhmetova, V.; Bukharaeva, E.; Deiss-Yehiely, N.; Dada, L.A.; Sachs, G.; Fernandez-Salas, E.; et al. Septin Dynamics Are Essential for Exocytosis. J Biol Chem 2015, 290, 5280–5297. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-W.; Yan, M.; Collins, R.F.; DiCiccio, J.E.; Grinstein, S.; Trimble, W.S. Mammalian Septins Are Required for Phagosome Formation. Mol Biol Cell 2008, 19, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Bai, X.; Bowen, J.R.; Dolat, L.; Korobova, F.; Yu, W.; Baas, P.W.; Svitkina, T.; Gallo, G.; Spiliotis, E.T. Septin-Driven Coordination of Actin and Microtubule Remodeling Regulates the Collateral Branching of Axons. Curr Biol 2012, 22, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-R.; Lee, R.T.-H.; Wang, Y.-M.; Zheng, X.-D.; Wang, Y. Candida Albicans Hyphal Morphogenesis Occurs in Sec3p-Independent and Sec3p-Dependent Phases Separated by Septin Ring Formation. J Cell Sci 2007, 120, 1898–1907. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.A.; Field, C.M.; Groen, A.C.; Mitchison, T.J.; Loose, M. Using Supported Bilayers to Study the Spatiotemporal Organization of Membrane Bound Proteins. Methods Cell Biol 2015, 128, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Tada, T.; Simonetta, A.; Batterton, M.; Kinoshita, M.; Edbauer, D.; Sheng, M. Role of Septin Cytoskeleton in Spine Morphogenesis and Dendrite Development in Neurons. Curr Biol 2007, 17, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Vessey, J.P.; Konecna, A.; Dahm, R.; Macchi, P.; Kiebler, M.A. The GTP-Binding Protein Septin 7 Is Critical for Dendrite Branching and Dendritic-Spine Morphology. Curr Biol 2007, 17, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Milenkovic, L.; Jin, H.; Scott, M.P.; Nachury, M.V.; Spiliotis, E.T.; Nelson, W.J. A Septin Diffusion Barrier at the Base of the Primary Cilium Maintains Ciliary Membrane Protein Distribution. Science 2010, 329, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Shindo, A.; Park, T.J.; Oh, E.C.; Ghosh, S.; Gray, R.S.; Lewis, R.A.; Johnson, C.A.; Attie-Bittach, T.; Katsanis, N.; et al. Planar Cell Polarity Acts through Septins to Control Collective Cell Movement and Ciliogenesis. Science 2010, 329, 1337–1340. [Google Scholar] [CrossRef] [PubMed]
- Palander, O.; El-Zeiry, M.; Trimble, W.S. Uncovering the Roles of Septins in Cilia. Frontiers in Cell and Developmental Biology 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Tooley, A.J.; Gilden, J.; Jacobelli, J.; Beemiller, P.; Trimble, W.S.; Kinoshita, M.; Krummel, M.F. Amoeboid T Lymphocytes Require the Septin Cytoskeleton for Cortical Integrity and Persistent Motility. Nature cell biology 2009, 11, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Dolat, L.; Hunyara, J.L.; Bowen, J.R.; Karasmanis, E.P.; Elgawly, M.; Galkin, V.E.; Spiliotis, E.T. Septins Promote Stress Fiber–Mediated Maturation of Focal Adhesions and Renal Epithelial Motility. J Cell Biol 2014, 207, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Kinoshita, A.; Yamada, S.; Tanaka, H.; Tanigaki, A.; Kitano, A.; Goto, M.; Okubo, K.; Nishiyama, H.; Ogawa, O.; et al. Cortical Organization by the Septin Cytoskeleton Is Essential for Structural and Mechanical Integrity of Mammalian Spermatozoa. Dev Cell 2005, 8, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yan, B.; Ren, J.; Lyu, R.; Wu, Y.; Guo, Y.; Li, D.; Zhang, H.; Hu, J. FIT2 Organizes Lipid Droplet Biogenesis with ER Tubule-Forming Proteins and Septins. J Cell Biol 2021, 220, e201907183. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Castellanos, N.; Rodríguez, A.; Rabanal-Ruiz, Y.; Fernández-Vega, A.; López-Miranda, J.; Vázquez-Martínez, R.; Frühbeck, G.; Malagón, M.M. The Cytoskeletal Protein Septin 11 Is Associated with Human Obesity and Is Involved in Adipocyte Lipid Storage and Metabolism. Diabetologia 2017, 60, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Nurullin, L.F.; Khuzakhmetova, V.F.; Khaziev, E.F.; Samigullin, D.V.; Tsentsevitsky, A.N.; Skorinkin, A.I.; Bukharaeva, E.A.; Vagin, O. Reorganization of Septins Modulates Synaptic Transmission at Neuromuscular Junctions. Neuroscience 2019, 404, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.W.; Estey, M.P.; DiCiccio, J.E.; Xie, H.; Patterson, D.; Trimble, W.S. Characterization of Presynaptic Septin Complexes in Mammalian Hippocampal Neurons. Biol Chem 2011, 392, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-M.; Fedchyshyn, M.J.; Grande, G.; Aitoubah, J.; Tsang, C.W.; Xie, H.; Ackerley, C.A.; Trimble, W.S.; Wang, L.-Y. Septins Regulate Developmental Switching from Microdomain to Nanodomain Coupling of Ca2+ Influx to Neurotransmitter Release at a Central Synapse. Neuron 2010, 67, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Bonazzi, M.; Hamon, M.A.; Tham, T.N.; Mallet, A.; Lelek, M.; Gouin, E.; Demangel, C.; Brosch, R.; Zimmer, C.; et al. Entrapment of Intracytosolic Bacteria by Septin Cage-like Structures. Cell Host Microbe 2010, 8, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Krokowski, S.; Lobato-Márquez, D.; Chastanet, A.; Pereira, P.M.; Angelis, D.; Galea, D.; Larrouy-Maumus, G.; Henriques, R.; Spiliotis, E.T.; Carballido-López, R.; et al. Septins Recognize and Entrap Dividing Bacterial Cells for Delivery to Lysosomes. Cell Host Microbe 2018, 24, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Dolat, L.; Hu, Q.; Spiliotis, E.T. Septin Functions in Organ System Physiology and Pathology. Biol Chem 2014, 395, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Versele, M.; Gullbrand, B.; Shulewitz, M.J.; Cid, V.J.; Bahmanyar, S.; Chen, R.E.; Barth, P.; Alber, T.; Thorner, J. Protein-Protein Interactions Governing Septin Heteropentamer Assembly and Septin Filament Organization in Saccharomyces Cerevisiae. Mol Biol Cell 2004, 15, 4568–4583. [Google Scholar] [CrossRef] [PubMed]
- Cavini, I.A.; Leonardo, D.A.; Rosa, H.V.D.; Castro, D.K.S.V.; D’Muniz Pereira, H.; Valadares, N.F.; Araujo, A.P.U.; Garratt, R.C. The Structural Biology of Septins and Their Filaments: An Update. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Bertin, A.; McMurray, M.A.; Pierson, J.; Thai, L.; McDonald, K.L.; Zehr, E.A.; García, G.; Peters, P.; Thorner, J.; Nogales, E. Three-Dimensional Ultrastructure of the Septin Filament Network in Saccharomyces Cerevisiae. Mol. Biol. Cell 2012, 23, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Bridges, A.A.; Zhang, H.; Mehta, S.B.; Occhipinti, P.; Tani, T.; Gladfelter, A.S. Septin Assemblies Form by Diffusion-Driven Annealing on Membranes. Proc Natl Acad Sci U S A 2014, 111, 2146–2151. [Google Scholar] [CrossRef] [PubMed]
- John, C.M.; Hite, R.K.; Weirich, C.S.; Fitzgerald, D.J.; Jawhari, H.; Faty, M.; Schläpfer, D.; Kroschewski, R.; Winkler, F.K.; Walz, T.; et al. The Caenorhabditis Elegans Septin Complex Is Nonpolar. The EMBO Journal 2007, 26, 3296–3307. [Google Scholar] [CrossRef] [PubMed]
- Rodal, A.A.; Kozubowski, L.; Goode, B.L.; Drubin, D.G.; Hartwig, J.H. Actin and Septin Ultrastructures at the Budding Yeast Cell Cortex. MBoC 2005, 16, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Sirajuddin, M.; Farkasovsky, M.; Hauer, F.; Kühlmann, D.; Macara, I.G.; Weyand, M.; Stark, H.; Wittinghofer, A. Structural Insight into Filament Formation by Mammalian Septins. Nature 2007, 449, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Field, C.M.; al-Awar, O.; Rosenblatt, J.; Wong, M.L.; Alberts, B.; Mitchison, T.J. A Purified Drosophila Septin Complex Forms Filaments and Exhibits GTPase Activity. J Cell Biol 1996, 133, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Arbizzani, F.; Mavrakis, M.; Hoya, M.; Ribas, J.C.; Brasselet, S.; Paoletti, A.; Rincon, S.A. Septin Filament Compaction into Rings Requires the Anillin Mid2 and Contractile Ring Constriction. Cell Reports 2022, 39, 110722. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Morrell, J.L.; Jennings, J.L.; Link, A.J.; Gould, K.L. Requirements of Fission Yeast Septins for Complex Formation, Localization, and Function. Mol Biol Cell 2004, 15, 5551–5564. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Q.; Kuhn, J.R.; Kovar, D.R.; Pollard, T.D. Spatial and Temporal Pathway for Assembly and Constriction of the Contractile Ring in Fission Yeast Cytokinesis. Dev Cell 2003, 5, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Bertin, A.; McMurray, M.A.; Grob, P.; Park, S.S.; Garcia, G., 3rd; Patanwala, I.; Ng, H.L.; Alber, T.; Thorner, J.; Nogales, E. Saccharomyces Cerevisiae Septins: Supramolecular Organization of Heterooligomers and the Mechanism of Filament Assembly. Proc Natl Acad Sci U S A 2008, 105, 8274–8279. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G., 3rd; Bertin, A.; Li, Z.; Song, Y.; McMurray, M.A.; Thorner, J.; Nogales, E. Subunit-Dependent Modulation of Septin Assembly: Budding Yeast Septin Shs1 Promotes Ring and Gauze Formation. J Cell Biol 2011, 195, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Taveneau, C.; Blanc, R.; Péhau-Arnaudet, G.; Di Cicco, A.; Bertin, A. Synergistic Role of Nucleotides and Lipids for the Self-Assembly of Shs1 Septin Oligomers. Biochemical Journal 2020, 477, 2697–2714. [Google Scholar] [CrossRef] [PubMed]
- Beber, A.; Alqabandi, M.; Prévost, C.; Viars, F.; Lévy, D.; Bassereau, P.; Bertin, A.; Mangenot, S. Septin-Based Readout of PI(4,5)P2 Incorporation into Membranes of Giant Unilamellar Vesicles. Cytoskeleton 2019, 76, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Szuba, A.; Bano, F.; Castro-Linares, G.; Iv, F.; Mavrakis, M.; Richter, R.P.; Bertin, A.; Koenderink, G.H. Membrane Binding Controls Ordered Self-Assembly of Animal Septins. eLife 2021, 10, e63349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kong, C.; Xie, H.; McPherson, P.S.; Grinstein, S.; Trimble, W.S. Phosphatidylinositol Polyphosphate Binding to the Mammalian Septin H5 Is Modulated by GTP. Current Biology 1999, 9, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Bertin, A.; McMurray, M.A.; Thai, L.; Garcia, G.; Votin, V.; Grob, P.; Allyn, T.; Thorner, J.; Nogales, E. Phosphatidylinositol-4,5-Bisphosphate Promotes Budding Yeast Septin Filament Assembly and Organization. Journal of Molecular Biology 2010, 404, 711–731. [Google Scholar] [CrossRef] [PubMed]
- Tanaka-Takiguchi, Y.; Kinoshita, M.; Takiguchi, K. Septin-Mediated Uniform Bracing of Phospholipid Membranes. Current biology: CB 2009, 19, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Garrenton, L.S.; Stefan, C.J.; McMurray, M.A.; Emr, S.D.; Thorner, J. Pheromone-Induced Anisotropy in Yeast Plasma Membrane Phosphatidylinositol-4,5-Bisphosphate Distribution Is Required for MAPK Signaling. Proceedings of the National Academy of Sciences 2010, 107, 11805–11810. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Escudero, I.; Roelants, F.M.; Thorner, J.; Nombela, C.; Molina, M.; Cid, V.J. Reconstitution of the Mammalian PI3K/PTEN/Akt Pathway in Yeast. Biochem J 2005, 390, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Casamayor, A.; Snyder, M. Molecular Dissection of a Yeast Septin: Distinct Domains Are Required for Septin Interaction, Localization, and Function. Molecular and Cellular Biology 2003, 23, 2762–2777. [Google Scholar] [CrossRef] [PubMed]
- Omrane, M.; Camara, A.S.; Taveneau, C.; Benzoubir, N.; Tubiana, T.; Yu, J.; Guérois, R.; Samuel, D.; Goud, B.; Poüs, C.; et al. Septin 9 Has Two Polybasic Domains Critical to Septin Filament Assembly and Golgi Integrity. iScience 2019, 13, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Alaoui, F.E.; Al-Akiki, I.; Ibanes, S.; Lyonnais, S.; Sanchez-Fuentes, D.; Desgarceaux, R.; Cazevieille, C.; Blanchard, M.-P.; Parmeggiani, A.; Carretero-Genevrier, A.; et al. Septin Filament Assembly Assist the Lateral Organization of Membranes 2024, 2024.03.19.585775.
- Beber, A.; Taveneau, C.; Nania, M.; Tsai, F.-C.; Di Cicco, A.; Bassereau, P.; Lévy, D.; Cabral, J.T.; Isambert, H.; Mangenot, S.; et al. Membrane Reshaping by Micrometric Curvature Sensitive Septin Filaments. Nat Commun 2019, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Bridges, A.A.; Jentzsch, M.S.; Oakes, P.W.; Occhipinti, P.; Gladfelter, A.S. Micron-Scale Plasma Membrane Curvature Is Recognized by the Septin Cytoskeleton. Journal of Cell Biology 2016, 213, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Cannon, K.S.; Woods, B.L.; Crutchley, J.M.; Gladfelter, A.S. An Amphipathic Helix Enables Septins to Sense Micrometer-Scale Membrane Curvature. Journal of Cell Biology 2019, 218, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Ewers, H.; Tada, T.; Petersen, J.D.; Racz, B.; Sheng, M.; Choquet, D. A Septin-Dependent Diffusion Barrier at Dendritic Spine Necks. PLOS ONE 2014, 9, e113916. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Lin, Y.H.; Chen, H.I.; Wang, Y.Y.; Chiou, Y.W.; Lin, H.H.; Pan, H.A.; Wu, C.M.; Su, S.M.; Hsu, C.C.; et al. SEPT12 Mutations Cause Male Infertility with Defective Sperm Annulus. Hum Mutat 2012, 33, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Newby, J.; Gladfelter, A.S. Control of Septin Filament Flexibility and Bundling by Subunit Composition and Nucleotide Interactions. MBoC 2018, 29, 702–712. [Google Scholar] [CrossRef]
- Garcia, G., III; Bertin, A.; Li, Z.; Song, Y.; McMurray, M.A.; Thorner, J.; Nogales, E. Subunit-Dependent Modulation of Septin Assembly: Budding Yeast Septin Shs1 Promotes Ring and Gauze Formation. Journal of Cell Biology 2011, 195, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Frazier, J.A.; Wong, M.L.; Longtine, M.S.; Pringle, J.R.; Mann, M.; Mitchison, T.J.; Field, C. Polymerization of Purified Yeast Septins: Evidence That Organized Filament Arrays May Not Be Required for Septin Function. J Cell Biol 1998, 143, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Booth, E.A.; Vane, E.W.; Dovala, D.; Thorner, J. A Förster Resonance Energy Transfer (FRET)-Based System Provides Insight into the Ordered Assembly of Yeast Septin Hetero-Octamers *. Journal of Biological Chemistry 2015, 290, 28388–28401. [Google Scholar] [CrossRef] [PubMed]
- McMurray, M.A.; Bertin, A.; Garcia, G.; Lam, L.; Nogales, E.; Thorner, J. Septin Filament Formation Is Essential in Budding Yeast. Developmental Cell 2011, 20, 540–549. [Google Scholar] [CrossRef]
- Garcia, G.; Finnigan, G.C.; Heasley, L.R.; Sterling, S.M.; Aggarwal, A.; Pearson, C.G.; Nogales, E.; McMurray, M.A.; Thorner, J. Assembly, Molecular Organization, and Membrane-Binding Properties of Development-Specific Septins. J Cell Biol 2016, 212, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Heasley, L.R.; McMurray, M.A. Roles of Septins in Prospore Membrane Morphogenesis and Spore Wall Assembly in Saccharomyces Cerevisiae. MBoC 2016, 27, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Cid, V.J.; Adamikova, L.; Sanchez, M.; Molina, M.; Nombela, C. Cell Cycle Control of Septin Ring Dynamics in the Budding Yeast. Microbiology 2001, 147, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.B.; Haarer, B.K.; Pringle, J.R. Cellular Morphogenesis in the Saccharomycescerevisiae Cell Cycle: Localization of the CDC3 Gene Product and the Timing of Events at the Budding Site.
- Caviston, J.P.; Longtine, M.; Pringle, J.R.; Bi, E. The Role of Cdc42p GTPase-Activating Proteins in Assembly of the Septin Ring in Yeast. Mol Biol Cell 2003, 14, 4051–4066. [Google Scholar] [CrossRef]
- Dobbelaere, J.; Gentry, M.S.; Hallberg, R.L.; Barral, Y. Phosphorylation-Dependent Regulation of Septin Dynamics during the Cell Cycle. Dev Cell 2003, 4, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; Fares, H.; Pringle, J.R. Role of the Yeast Gin4p Protein Kinase in Septin Assembly and the Relationship between Septin Assembly and Septin Function. J Cell Biol 1998, 143, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Tartakoff, A.M.; Aylyarov, I.; Jaiswal, P. Septin-Containing Barriers Control the Differential Inheritance of Cytoplasmic Elements. Cell Reports 2013, 3, 223–236. [Google Scholar] [CrossRef]
- Fares, H.; Goetsch, L.; Pringle, J.R. Identification of a Developmentally Regulated Septin and Involvement of the Septins in Spore Formation in Saccharomyces Cerevisiae. J Cell Biol 1996, 132, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Spiliotis, E.T.; Nakos, K. Cellular Functions of Actin- and Microtubule-Associated Septins. Current Biology 2021, 31, R651–R666. [Google Scholar] [CrossRef]
- Ong, K.; Wloka, C.; Okada, S.; Svitkina, T.; Bi, E. Architecture and Dynamic Remodelling of the Septin Cytoskeleton during the Cell Cycle. Nat Commun 2014, 5, 5698. [Google Scholar] [CrossRef] [PubMed]
- Vrabioiu, A.M.; Mitchison, T.J. Structural Insights into Yeast Septin Organization from Polarized Fluorescence Microscopy. Nature 2006, 443, 466–469. [Google Scholar] [CrossRef] [PubMed]
- McQuilken, M.; Jentzsch, M.S.; Verma, A.; Mehta, S.B.; Oldenbourg, R.; Gladfelter, A.S. Analysis of Septin Reorganization at Cytokinesis Using Polarized Fluorescence Microscopy. Front Cell Dev Biol 2017, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Demay, B.S.; Bai, X.; Howard, L.; Occhipinti, P.; Meseroll, R.A.; Spiliotis, E.T.; Oldenbourg, R.; Gladfelter, A.S. Septin Filaments Exhibit a Dynamic, Paired Organization That Is Conserved from Yeast to Mammals. J Cell Biol 2011, 193, 1065–1081. [Google Scholar] [CrossRef] [PubMed]
- Tamborrini, D.; Juanes, M.A.; Ibanes, S.; Rancati, G.; Piatti, S. Recruitment of the Mitotic Exit Network to Yeast Centrosomes Couples Septin Displacement to Actomyosin Constriction. Nat Commun 2018, 9, 4308. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, J.; Chen, X.; Bi, E. Architecture, Remodeling, and Functions of the Septin Cytoskeleton. Cytoskeleton 2019, 76, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Bi, E.; Park, H.O. Cell Polarization and Cytokinesis in Budding Yeast. Genetics 2012, 191, 347–387. [Google Scholar] [CrossRef] [PubMed]
- Cvrckova, F.; De Virgilio, C.; Manser, E.; Pringle, J.R.; Nasmyth, K. Ste20-like Protein Kinases Are Required for Normal Localization of Cell Growth and for Cytokinesis in Budding Yeast. Genes Dev 1995, 9, 1817–1830. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.J.; Mullner, R.; Lian, K.; Park, H.-O. Cdc42 Couples Septin Recruitment to the Axial Landmark Assembly via Axl2 in Budding Yeast. Journal of Cell Science 2023, 137, jcs261080. [Google Scholar] [CrossRef] [PubMed]
- Chant, J.; Mischke, M.; Mitchell, E.; Herskowitz, I.; Pringle, J.R. Role of Bud3p in Producing the Axial Budding Pattern of Yeast. J Cell Biol 1995, 129, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Sanders, S.L.; Herskowitz, I. The BUD4 Protein of Yeast, Required for Axial Budding, Is Localized to the Mother/BUD Neck in a Cell Cycle-Dependent Manner. J Cell Biol 1996, 134, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.J.; Angerman, E.; Jung, C.H.; Park, H.O. Bud4 Mediates the Cell-Type-Specific Assembly of the Axial Landmark in Budding Yeast. Journal of cell science 2012, 125, 3840–3849. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Leda, M.; Hanna, J.; Savage, N.S.; Bi, E.; Goryachev, A.B. Daughter Cell Identity Emerges from the Interplay of Cdc42, Septins, and Exocytosis. Developmental Cell 2013, 26, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Lippincott, J.; Li, R. Sequential Assembly of Myosin II, an IQGAP-like Protein, and Filamentous Actin to a Ring Structure Involved in Budding Yeast Cytokinesis. J Cell Biol 1998, 140, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Vallen, E.A.; Dravis, C.; Tcheperegine, S.E.; Drees, B.; Bi, E. Identification and Functional Analysis of the Essential and Regulatory Light Chains of the Only Type II Myosin Myo1p in Saccharomyces Cerevisiae. The Journal of cell biology 2004, 165, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Watts, F.Z.; Shiels, G.; Orr, E. The Yeast MYO1 Gene Encoding a Myosin-like Protein Required for Cell Division. The EMBO journal 1987, 6, 3499–3505. [Google Scholar] [CrossRef] [PubMed]
- Boyne, J.R.; Yosuf, H.M.; Bieganowski, P.; Brenner, C.; Price, C. Yeast Myosin Light Chain, Mlc1p, Interacts with Both IQGAP and Class II Myosin to Effect Cytokinesis. Journal of cell science 2000, 113 Pt 24, 4533–4543. [Google Scholar] [CrossRef] [PubMed]
- Epp, J.A.; Chant, J. An IQGAP-Related Protein Controls Actin-Ring Formation and Cytokinesis in Yeast. Current Biology 1997, 7, 921–929. [Google Scholar] [CrossRef]
- Fang, X.; Luo, J.; Nishihama, R.; Wloka, C.; Dravis, C.; Travaglia, M.; Iwase, M.; Vallen, E.A.; Bi, E. Biphasic Targeting and Cleavage Furrow Ingression Directed by the Tail of a Myosin II. J Cell Biol 2010, 191, 1333–1350. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.B.; Li, R. A Myosin Light Chain Mediates the Localization of the Budding Yeast IQGAP-like Protein during Contractile Ring Formation. Curr Biol 2000, 10, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Tolliday, N.; VerPlank, L.; Li, R. Rho1 Directs Formin-Mediated Actin Ring Assembly during Budding Yeast Cytokinesis. Curr Biol 2002, 12, 1864–1870. [Google Scholar] [CrossRef] [PubMed]
- Vallen, E.A.; Caviston, J.; Bi, E. Roles of Hof1p, Bni1p, Bnr1p, and Myo1p in Cytokinesis in Saccharomyces Cerevisiae. Mol Biol Cell 2000, 11, 593–611. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Okada, S.; Cai, G.; Zhou, B.; Bi, E. MyosinII Heavy Chain and Formin Mediate the Targeting of Myosin Essential Light Chain to the Division Site before and during Cytokinesis. Molecular biology of the cell 2015, 26, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Chuang, J.S.; Schekman, R.W. Differential Trafficking and Timed Localization of Two Chitin Synthase Proteins, Chs2p and Chs3p. The Journal of cell biology 1996, 135, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Meitinger, F.; Petrova, B.; Lombardi, I.M.; Bertazzi, D.T.; Hub, B.; Zentgraf, H.; Pereira, G. Targeted Localization of Inn1, Cyk3 and Chs2 by the Mitotic-Exit Network Regulates Cytokinesis in Budding Yeast. J Cell Sci 2010, 123, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Foltman, M.; Filali-Mouncef, Y.; Crespo, D.; Sanchez-Diaz, A. Cell Polarity Protein Spa2 Coordinates Chs2 Incorporation at the Division Site in Budding Yeast. PLOS Genetics 2018, 14, e1007299. [Google Scholar] [CrossRef]
- VerPlank, L.; Li, R. Cell Cycle-Regulated Trafficking of Chs2 Controls Actomyosin Ring Stability during Cytokinesis. Mol Biol Cell 2005, 16, 2529–2543. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.F.; Bennett, A.M.; Ma, W.K.; Hall, M.C.; Yeong, F.M. Dependence of Chs2 ER Export on Dephosphorylation by Cytoplasmic Cdc14 Ensures That Septum Formation Follows Mitosis. Molecular biology of the cell 2012, 23, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Nishihama, R.; Schreiter, J.H.; Onishi, M.; Vallen, E.A.; Hanna, J.; Moravcevic, K.; Lippincott, M.F.; Han, H.; Lemmon, M.A.; Pringle, J.R.; et al. Role of Inn1 and Its Interactions with Hof1 and Cyk3 in Promoting Cleavage Furrow and Septum Formation in S. Cerevisiae. J Cell Biol 2009, 185, 995–1012. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Schreiter, J.H.; Okada, H.; Wloka, C.; Okada, S.; Yan, D.; Duan, X.; Bi, E. Hof1 and Chs4 Interact via F-BAR Domain and Sel1-like Repeats to Control Extracellular Matrix Deposition during Cytokinesis. Current Biology 2017, 27, 2878–2886. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Nishihama, R.; Onishi, M.; Pringle, J.R. Role of the Hof1–Cyk3 Interaction in Cleavage-Furrow Ingression and Primary-Septum Formation during Yeast Cytokinesis. Molecular Biology of the Cell 2018, 29, 597–609. [Google Scholar] [CrossRef]
- Devrekanli, A.; Foltman, M.; Roncero, C.; Sanchez-Diaz, A.; Labib, K. Inn1 and Cyk3 Regulate Chitin Synthase during Cytokinesis in Budding Yeasts. Journal of cell science 2012, 125, 5453–5466. [Google Scholar] [CrossRef] [PubMed]
- Jendretzki, A.; Ciklic, I.; Rodicio, R.; Schmitz, H.P.; Heinisch, J.J. Cyk3 Acts in Actomyosin Ring Independent Cytokinesis by Recruiting Inn1 to the Yeast Bud Neck. Molecular genetics and genomics: MGG 2009, 282, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Ko, N.; Nishihama, R.; Pringle, J.R. Distinct Roles of Rho1, Cdc42, and Cyk3 in Septum Formation and Abscission during Yeast Cytokinesis. J Cell Biol 2013, 202, 311–329. [Google Scholar] [CrossRef]
- Sanchez-Diaz, A.; Marchesi, V.; Murray, S.; Jones, R.; Pereira, G.; Edmondson, R.; Allen, T.; Labib, K. Inn1 Couples Contraction of the Actomyosin Ring to Membrane Ingression during Cytokinesis in Budding Yeast. Nature cell biology 2008, 10, 395–406. [Google Scholar] [CrossRef]
- Foltman, M.; Molist, I.; Arcones, I.; Sacristan, C.; Filali-Mouncef, Y.; Roncero, C.; Sanchez-Diaz, A. Ingression Progression Complexes Control Extracellular Matrix Remodelling during Cytokinesis in Budding Yeast. PLOS Genetics 2016, 12, e1005864. [Google Scholar] [CrossRef] [PubMed]
- Lippincott, J.; Li, R. Dual Function of Cyk2, a Cdc15/PSTPIP Family Protein, in Regulating Actomyosin Ring Dynamics and Septin Distribution. J Cell Biol 1998, 143, 1947–1960. [Google Scholar] [CrossRef] [PubMed]
- Bi, E.; Maddox, P.; Lew, D.J.; Salmon, E.D.; McMillan, J.N.; Yeh, E.; Pringle, J.R. Involvement of an Actomyosin Contractile Ring in Saccharomyces Cerevisiae Cytokinesis. J Cell Biol 1998, 142, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Lippincott, J.; Li, R. Sequential Assembly of Myosin II, an IQGAP-like Protein, and Filamentous Actin to a Ring Structure Involved in Budding Yeast Cytokinesis. Journal of Cell Biology 1998, 140, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Vallen, E.A.; Dravis, C.; Tcheperegine, S.E.; Drees, B.; Bi, E. Identification and Functional Analysis of the Essential and Regulatory Light Chains of the Only Type II Myosin Myo1p in Saccharomyces Cerevisiae. Journal of Cell Biology 2004, 165, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Liu, W.; Bretscher, A. The Yeast Formin Bnr1p Has Two Localization Regions That Show Spatially and Temporally Distinct Association with Septin Structures. Mol Biol Cell 2010, 21, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Pruyne, D.; Gao, L.; Bi, E.; Bretscher, A. Stable and Dynamic Axes of Polarity Use Distinct Formin Isoforms in Budding Yeast. Molecular biology of the cell 2004, 15, 4971–4989. [Google Scholar] [CrossRef] [PubMed]
- Lippincott, J.; Li, R. Dual Function of Cyk2, a Cdc15/PSTPIP Family Protein, in Regulating Actomyosin Ring Dynamics and Septin Distribution. Journal of Cell Biology 1998, 143, 1947–1960. [Google Scholar] [CrossRef] [PubMed]
- Huxley, H.E. The Mechanism of Muscular Contraction. Science 1969, 164, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Huxley, H.; Hanson, J. Changes in the Cross-Striations of Muscle during Contraction and Stretch and Their Structural Interpretation. Nature 1954, 173, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Mendes Pinto, I.; Rubinstein, B.; Kucharavy, A.; Unruh, J.R.; Li, R. Actin Depolymerization Drives Actomyosin Ring Contraction during Budding Yeast Cytokinesis. Developmental Cell 2012, 22, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Ko, N.; Nishihama, R.; Tully, G.H.; Ostapenko, D.; Solomon, M.J.; Morgan, D.O.; Pringle, J.R. Identification of Yeast IQGAP (Iqg1p) as an Anaphase-Promoting-Complex Substrate and Its Role in Actomyosin-Ring-Independent Cytokinesis. MBoC 2007, 18, 5139–5153. [Google Scholar] [CrossRef] [PubMed]
- Tully, G.H.; Nishihama, R.; Pringle, J.R.; Morgan, D.O. The Anaphase-Promoting Complex Promotes Actomyosin-Ring Disassembly during Cytokinesis in Yeast. MBoC 2009, 20, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E. The Septation Apparatus, a Chitin-Requiring Machine in Budding Yeast. Archives of biochemistry and biophysics 2004, 426, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Kashimshetty, R.; Ng, K.E.; Tan, H.B.; Yeong, F.M. Exit from Mitosis Triggers Chs2p Transport from the Endoplasmic Reticulum to Mother-Daughter Neck via the Secretory Pathway in Budding Yeast. The Journal of cell biology 2006, 174, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Bi, E. Cytokinesis in Budding Yeast: The Relationship between Actomyosin Ring Function and Septum Formation. Cell Struct Funct 2001, 26, 529–537. [Google Scholar] [CrossRef]
- Schmidt, M.; Bowers, B.; Varma, A.; Roh, D.H.; Cabib, E. In Budding Yeast, Contraction of the Actomyosin Ring and Formation of the Primary Septum at Cytokinesis Depend on Each Other. J Cell Sci 2002, 115, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Roh, D.-H.; Bowers, B.; Schmidt, M.; Cabib, E. The Septation Apparatus, an Autonomous System in Budding Yeast. Mol Biol Cell 2002, 13, 2747–2759. [Google Scholar] [CrossRef] [PubMed]
- Lippincott, J.; Shannon, K.B.; Shou, W.; Deshaies, R.J.; Li, R. The Tem1 Small GTPase Controls Actomyosin and Septin Dynamics during Cytokinesis. J Cell Sci 2001, 114, 1379–86. [Google Scholar] [CrossRef] [PubMed]
- Tamborrini, D.; Piatti, S. Septin Clearance from the Division Site Triggers Cytokinesis in Budding Yeast. Microbial Cell 2019, 6, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Dobbelaere, J.; Barral, Y. Spatial Coordination of Cytokinetic Events by Compartmentalization of the Cell Cortex. Science 2004, 305, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Ko, N.; Nishihama, R.; Pringle, J.R. Distinct Roles of Rho1, Cdc42, and Cyk3 in Septum Formation and Abscission during Yeast Cytokinesis. Journal of Cell Biology 2013, 202, 311–329. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; MacTaggart, B.; Ohya, Y.; Bi, E. The Kinetic Landscape and Interplay of Protein Networks in Cytokinesis. iScience 2021, 24, 101917. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.L. Mitotic Exit and Separation of Mother and Daughter Cells. Genetics 2012, 192, 1165–1202. [Google Scholar] [CrossRef] [PubMed]
- Martin-Cuadrado, A.B.; Morrell, J.L.; Konomi, M.; An, H.; Petit, C.; Osumi, M.; Balasubramanian, M.; Gould, K.L.; Del Rey, F.; de Aldana, C.R. Role of Septins and the Exocyst Complex in the Function of Hydrolytic Enzymes Responsible for Fission Yeast Cell Separation. Mol Biol Cell 2005, 16, 4867–4881. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, A.S.; Moskow, J.J.; Zyla, T.R.; Lew, D.J. Isolation and Characterization of Effector-Loop Mutants of CDC42 in Yeast. Mol Biol Cell 2001, 12, 1239–1255. [Google Scholar] [CrossRef]
- Iwase, M.; Luo, J.; Nagaraj, S.; Longtine, M.; Kim, H.B.; Haarer, B.K.; Caruso, C.; Tong, Z.; Pringle, J.R.; Bi, E. Role of a Cdc42p Effector Pathway in Recruitment of the Yeast Septins to the Presumptive Bud Site. Mol Biol Cell 2006, 17, 1110–1125. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, A.S.; Bose, I.; Zyla, T.R.; Bardes, E.S.; Lew, D.J. Septin Ring Assembly Involves Cycles of GTP Loading and Hydrolysis by Cdc42p. The Journal of cell biology 2002, 156, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Benton, B.K.; Tinkelenberg, A.; Gonzalez, I.; Cross, F.R. Cla4p, a Saccharomyces Cerevisiae Cdc42p-Activated Kinase Involved in Cytokinesis, Is Activated at Mitosis. Mol Cell Biol 1997, 17, 5067–76. [Google Scholar] [CrossRef] [PubMed]
- Kadota, J.; Yamamoto, T.; Yoshiuchi, S.; Bi, E.; Tanaka, K. Septin Ring Assembly Requires Concerted Action of Polarisome Components, a PAK Kinase Cla4p, and the Actin Cytoskeleton in Saccharomyces Cerevisiae. Molecular biology of the cell 2004, 15, 5329–5345. [Google Scholar] [CrossRef] [PubMed]
- Versele, M.; Thorner, J. Septin Collar Formation in Budding Yeast Requires GTP Binding and Direct Phosphorylation by the PAK, Cla4. J Cell Biol 2004, 164, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Bose, I.; Irazoqui, J.E.; Moskow, J.J.; Bardes, E.S.; Zyla, T.R.; Lew, D.J. Assembly of Scaffold-Mediated Complexes Containing Cdc42p, the Exchange Factor Cdc24p, and the Effector Cla4p Required for Cell Cycle-Regulated Phosphorylation of Cdc24p. The Journal of biological chemistry 2001, 276, 7176–7186. [Google Scholar] [CrossRef] [PubMed]
- Gulli, M.-P.; Jaquenoud, M.; Shimada, Y.; Niederhäuser, G.; Wiget, P.; Peter, M. Phosphorylation of the Cdc42 Exchange Factor Cdc24 by the PAK-like Kinase Cla4 May Regulate Polarized Growth in Yeast. Molecular Cell 2000, 6, 1155–1167. [Google Scholar] [CrossRef] [PubMed]
- Kozubowski, L.; Saito, K.; Johnson, J.M.; Howell, A.S.; Zyla, T.R.; Lew, D.J. Symmetry-Breaking Polarization Driven by a Cdc42p GEF-PAK Complex. Current biology: CB 2008, 18, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Chollet, J.; Dünkler, A.; Bäuerle, A.; Vivero-Pol, L.; Mulaw, M.A.; Gronemeyer, T.; Johnsson, N. Cdc24 Interacts with the Septins to Create a Positive Feedback during Bud Site Assembly in Yeast. Journal of Cell Science, 2402. [Google Scholar] [CrossRef]
- Brown, J.L.; Jaquenoud, M.; Gulli, M.P.; Chant, J.; Peter, M. Novel Cdc42-Binding Proteins Gic1 and Gic2 Control Cell Polarity in Yeast. Genes Dev 1997, 11, 2972–2982. [Google Scholar] [CrossRef] [PubMed]
- Orlando, K.; Sun, X.; Zhang, J.; Lu, T.; Yokomizo, L.; Wang, P.; Guo, W. Exo-Endocytic Trafficking and the Septin-Based Diffusion Barrier Are Required for the Maintenance of Cdc42p Polarization during Budding Yeast Asymmetric Growth. Mol Biol Cell 2011, 22, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Sadian, Y.; Gatsogiannis, C.; Patasi, C.; Hofnagel, O.; Goody, R.S.; Farkasovsky, M.; Raunser, S. The Role of Cdc42 and Gic1 in the Regulation of Septin Filament Formation and Dissociation. Elife 2013, 2, e01085. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.J.; Miller, K.E.; Guegueniat, J.; Beven, L.; Park, H.-O. The Shared Role of the Rsr1 GTPase and Gic1/Gic2 in Cdc42 Polarization. MBoC 2018, 29, 2359–2369. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.N.; Zyla, T.R.; Lew, D.J. A Role for Gic1 and Gic2 in Cdc42 Polarization at Elevated Temperature. PLoS ONE 2018, 13, e0200863. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Ferguson, K.M.; O’Brien, R.; Sigler, P.B.; Schlessinger, J. Specific and High-Affinity Binding of Inositol Phosphates to an Isolated Pleckstrin Homology Domain. Proceedings of the National Academy of Sciences of the United States of America 1995, 92, 10472–10476. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.-D.; Sperber, L.M.; Kane, S.A.; Tong, Z.; Tong, A.H.Y.; Boone, C.; Bi, E. Sequential and Distinct Roles of the Cadherin Domain-Containing Protein Axl2p in Cell Polarization in Yeast Cell Cycle. MBoC 2007, 18, 2542–2560. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.; Chiou, J.-G.; Zhurikhina, A.; Zyla, T.R.; Tsygankov, D.; Lew, D.J. Temporal Regulation of Morphogenetic Events in Saccharomyces Cerevisiae. Mol Biol Cell 2018, 29, 2069–2083. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.J.; Mullner, R.; Lian, K.; Park, H.-O. Cdc42 Couples Septin Recruitment to the Axial Landmark Assembly via Axl2 in Budding Yeast. Journal of Cell Science 2024, 137, jcs261080. [Google Scholar] [CrossRef]
- Merlini, L.; Bolognesi, A.; Juanes, M.A.; Vandermoere, F.; Courtellemont, T.; Pascolutti, R.; Seveno, M.; Barral, Y.; Piatti, S. Rho1- and Pkc1-Dependent Phosphorylation of the F-BAR Protein Syp1 Contributes to Septin Ring Assembly. Molecular biology of the cell 2015, 26, 3245–3262. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Neo, S.P.; Yu, X.; Cai, M. A Novel Septin-Associated Protein, Syp1p, Is Required for Normal Cell Cycle-Dependent Septin Cytoskeleton Dynamics in Yeast. Genetics 2008, 180, 1445–1457. [Google Scholar] [CrossRef] [PubMed]
- Ibanes, S.; El-Alaoui, F.; Lai-Kee-Him, J.; Cazevieille, C.; Hoh, F.; Lyonnais, S.; Bron, P.; Cipelletti, L.; Picas, L.; Piatti, S. The Syp1/FCHo2 Protein Induces Septin Filament Bundling through Its Intrinsically Disordered Domain. Cell Rep 2022, 41, 111765. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, E.M.; McDonald, H.; Yates, J., 3rd; Kellogg, D.R. Cell Cycle-Dependent Assembly of a Gin4-Septin Complex. Mol Biol Cell 2002, 13, 2091–2105. [Google Scholar] [CrossRef] [PubMed]
- Asano, S.; Park, J.E.; Yu, L.R.; Zhou, M.; Sakchaisri, K.; Park, C.J.; Kang, Y.H.; Thorner, J.; Veenstra, T.D.; Lee, K.S. Direct Phosphorylation and Activation of a Nim1-Related Kinase Gin4 by Elm1 in Budding Yeast. J Biol Chem 2006, 281, 27090–27098. [Google Scholar] [CrossRef] [PubMed]
- Egelhofer, T.A.; Villen, J.; McCusker, D.; Gygi, S.P.; Kellogg, D.R. The Septins Function in G1 Pathways That Influence the Pattern of Cell Growth in Budding Yeast. PloS one 2008, 3, e2022. [Google Scholar] [CrossRef] [PubMed]
- Shulewitz, M.J.; Inouye, C.J.; Thorner, J. Hsl7 Localizes to a Septin Ring and Serves as an Adapter in a Regulatory Pathway That Relieves Tyrosine Phosphorylation of Cdc28 Protein Kinase in Saccharomyces Cerevisiae. Mol Cell Biol 1999, 19, 7123–7137. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.L.; Blacketer, M.J.; Edgington, N.P.; Myers, A.M. Assembly Interdependence among the S. Cerevisiae Bud Neck Ring Proteins Elm1p, Hsl1p and Cdc12p. Yeast 2003, 20, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, G.C.; Duvalyan, A.; Liao, E.N.; Sargsyan, A.; Thorner, J. Detection of Protein-Protein Interactions at the Septin Collar in Saccharomyces Cerevisiae Using a Tripartite Split-GFP System. Molecular biology of the cell 2016, 27, 2708–2725. [Google Scholar] [CrossRef] [PubMed]
- Barral, Y.; Parra, M.; Bidlingmaier, S.; Snyder, M. Nim1-Related Kinases Coordinate Cell Cycle Progression with the Organization of the Peripheral Cytoskeleton in Yeast. Genes Dev 1999, 13, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Breitkreutz, A.; Choi, H.; Sharom, J.R.; Boucher, L.; Neduva, V.; Larsen, B.; Lin, Z.Y.; Breitkreutz, B.J.; Stark, C.; Liu, G.; et al. A Global Protein Kinase and Phosphatase Interaction Network in Yeast. Science 2010, 328, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, G.C.; Sterling, S.M.; Duvalyan, A.; Liao, E.N.; Sargsyan, A.; Garcia, G.; Nogales, E.; Thorner, J. Coordinate Action of Distinct Sequence Elements Localizes Checkpoint Kinase Hsl1 to the Septin Collar at the Bud Neck in Saccharomyces Cerevisiae. Mol Biol Cell 2016, 27, 2213–2233. [Google Scholar] [CrossRef]
- Okuzaki, D.; Nojima, H. Kcc4 Associates with Septin Proteins of Saccharomyces Cerevisiae. FEBS Letters 2001, 489, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Bouquin, N.; Barral, Y.; Courbeyrette, R.; Blondel, M.; Snyder, M.; Mann, C. Regulation of Cytokinesis by the Elm1 Protein Kinase in Saccharomyces Cerevisiae. J Cell Sci 2000, 113 ( Pt 8) Pt 8, 1435–1445. [Google Scholar] [CrossRef]
- Marquardt, J.; Yao, L.-L.; Okada, H.; Svitkina, T.; Bi, E. The LKB1-like Kinase Elm1 Controls Septin Hourglass Assembly and Stability by Regulating Filament Pairing. Current Biology 2020, 30, 2386–2394. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, A.; Kellogg, D. The Elm1 Kinase Functions in a Mitotic Signaling Network in Budding Yeast. Mol Cell Biol 1999, 19, 7983–7994. [Google Scholar] [CrossRef]
- Marquardt, J.; Chen, X.; Bi, E. Reciprocal Regulation by Elm1 and Gin4 Controls Septin Hourglass Assembly and Remodeling. Journal of Cell Biology 2024, 223, e202308143. [Google Scholar] [CrossRef] [PubMed]
- Szkotnicki, L.; Crutchley, J.M.; Zyla, T.R.; Bardes, E.S.; Lew, D.J. The Checkpoint Kinase Hsl1p Is Activated by Elm1p-Dependent Phosphorylation. Mol Biol Cell 2008, 19, 4675–4686. [Google Scholar] [CrossRef] [PubMed]
- Fraschini, R.; Bilotta, D.; Lucchini, G.; Piatti, S. Functional Characterization of Dma1 and Dma2, the Budding Yeast Homologues of Schizosaccharomyces Pombe Dma1 and Human Chfr. Mol Biol Cell 2004, 15, 3796–3810. [Google Scholar] [CrossRef] [PubMed]
- Merlini, L.; Fraschini, R.; Boettcher, B.; Barral, Y.; Lucchini, G.; Piatti, S. Budding Yeast Dma Proteins Control Septin Dynamics and the Spindle Position Checkpoint by Promoting the Recruitment of the Elm1 Kinase to the Bud Neck. PLoS Genet 2012, 8, e1002670. [Google Scholar] [CrossRef] [PubMed]
- Patasi, C.; Godocikova, J.; Michlikova, S.; Nie, Y.; Kacerikova, R.; Kvalova, K.; Raunser, S.; Farkasovsky, M. The Role of Bni5 in the Regulation of Septin Higher-Order Structure Formation. Biological chemistry 2015, 396, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.R.; Song, S.; Ro, H.S.; Park, C.J.; Lippincott, J.; Li, R.; Pringle, J.R.; De Virgilio, C.; Longtine, M.S.; Lee, K.S. Bni5p, a Septin-Interacting Protein, Is Required for Normal Septin Function and Cytokinesis in Saccharomyces Cerevisiae. Mol Cell Biol 2002, 22, 6906–6920. [Google Scholar] [CrossRef]
- Booth, E.A.; Sterling, S.M.; Dovala, D.; Nogales, E.; Thorner, J. Effects of Bni5 Binding on Septin Filament Organization. Journal of Molecular Biology 2016, 428, 4962–4980. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, G.C.; Booth, E.A.; Duvalyan, A.; Liao, E.N.; Thorner, J. The Carboxy-Terminal Tails of Septins Cdc11 and Shs1 Recruit Myosin-II Binding Factor Bni5 to the Bud Neck in Saccharomyces Cerevisiae. Genetics 2015, 200, 843–862. [Google Scholar] [CrossRef] [PubMed]
- Touati, S.A.; Kataria, M.; Jones, A.W.; Snijders, A.P.; Uhlmann, F. Phosphoproteome Dynamics during Mitotic Exit in Budding Yeast. EMBO J 2018, 37. [Google Scholar] [CrossRef] [PubMed]
- Renz, C.; Oeljeklaus, S.; Grinhagens, S.; Warscheid, B.; Johnsson, N.; Gronemeyer, T. Identification of Cell Cycle Dependent Interaction Partners of the Septins by Quantitative Mass Spectrometry. PLOS ONE 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Swaney, D.L.; Beltrao, P.; Starita, L.; Guo, A.; Rush, J.; Fields, S.; Krogan, N.J.; Villen, J. Global Analysis of Phosphorylation and Ubiquitylation Cross-Talk in Protein Degradation. Nature methods 2013, 10, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, C.P.; Smolka, M.B.; Payne, S.H.; Bafna, V.; Eng, J.; Zhou, H. A Multidimensional Chromatography Technology for In-Depth Phosphoproteome Analysis. Molecular & cellular proteomics: MCP 2008, 7, 1389–1396. [Google Scholar] [CrossRef]
- Holt, L.J.; Tuch, B.B.; Villen, J.; Johnson, A.D.; Gygi, S.P.; Morgan, D.O. Global Analysis of Cdk1 Substrate Phosphorylation Sites Provides Insights into Evolution. Science 2009, 325, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Smolka, M.B.; Albuquerque, C.P.; Chen, S. -h.; Zhou, H. Proteome-Wide Identification of in Vivo Targets of DNA Damage Checkpoint Kinases. Proceedings of the National Academy of Sciences 2007, 104, 10364–10369. [Google Scholar] [CrossRef] [PubMed]
- Bodenmiller, B.; Wanka, S.; Kraft, C.; Urban, J.; Campbell, D.; Pedrioli, P.G.; Gerrits, B.; Picotti, P.; Lam, H.; Vitek, O.; et al. Phosphoproteomic Analysis Reveals Interconnected System-Wide Responses to Perturbations of Kinases and Phosphatases in Yeast. Science signaling 2010, 3, rs4. [Google Scholar] [CrossRef] [PubMed]
- Soufi, B.; Kelstrup, C.D.; Stoehr, G.; Frohlich, F.; Walther, T.C.; Olsen, J.V. Global Analysis of the Yeast Osmotic Stress Response by Quantitative Proteomics. Molecular bioSystems 2009, 5, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Paulo, J.A.; Nusinow, D.P.; Huttlin, E.L.; Gygi, S.P. Investigation of Proteomic and Phosphoproteomic Responses to Signaling Network Perturbations Reveals Functional Pathway Organizations in Yeast. Cell Reports 2019, 29, 2092–2104. [Google Scholar] [CrossRef] [PubMed]
- Lanz, M.C.; Yugandhar, K.; Gupta, S.; Sanford, E.J.; Faça, V.M.; Vega, S.; Joiner, A.M.N.; Fromme, J.C.; Yu, H.; Smolka, M.B. In-Depth and 3-Dimensional Exploration of the Budding Yeast Phosphoproteome. EMBO reports 2021, 22, e51121. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.S.; Blobel, G. Cell Cycle-Regulated Attachment of the Ubiquitin-Related Protein SUMO to the Yeast Septins. J Cell Biol 1999, 147, 981–994. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, L.; Lau, A.; Lambert, J.P.; Zhou, H.; Fong, Y.; Couture, J.F.; Figeys, D.; Baetz, K. Regulation of Septin Dynamics by the Saccharomyces Cerevisiae Lysine Acetyltransferase NuA4. PloS one 2011, 6, e25336. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, K.; Svitkina, T.; Bi, E. Critical Roles of a RhoGEF-Anillin Module in Septin Architectural Remodeling during Cytokinesis. Current Biology 2020, 30, 1477–1490. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.J.; Hood-DeGrenier, J.K.; Park, H.O. Coupling of Septins to the Axial Landmark by Bud4 in Budding Yeast. Journal of cell science 2013, 126, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Guo, J.; Zhou, Y.T.; Gao, X.D. The Anillin-Related Region of Bud4 Is the Major Functional Determinant for Bud4’s Function in Septin Organization during Bud Growth and Axial Bud Site Selection in Budding Yeast. Eukaryotic cell 2015, 14, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Chant, J.; Herskowitz, I. Genetic Control of Bud Site Selection in Yeast by a Set of Gene Products That Constitute a Morphogenetic Pathway. Cell 1991, 65, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Gong, T.; Gao, X.-D. Identification of an Amphipathic Helix Important for the Formation of Ectopic Septin Spirals and Axial Budding in Yeast Axial Landmark Protein Bud3p. PLoS ONE 2011, 6, e16744. [Google Scholar] [CrossRef] [PubMed]
- Schrock, M.N.; Yan, Y.; Goeckel, M.E.; Basgall, E.M.; Lewis, I.C.; Leonard, K.G.; Halloran, M.; Finnigan, G.C. Characterization of Bud3 Domains Sufficient for Bud Neck Targeting in S. Cerevisiae. Access Microbiol 2022, 4, 000341. [Google Scholar] [CrossRef] [PubMed]
- Stegmeier, F.; Amon, A. Closing Mitosis: The Functions of the Cdc14 Phosphatase and Its Regulation. Annu Rev Genet 2004, 38, 203–232. [Google Scholar] [CrossRef] [PubMed]
- Jaspersen, S.L.; Morgan, D.O. Cdc14 Activates Cdc15 to Promote Mitotic Exit in Budding Yeast. Curr Biol 2000, 10, 615–618. [Google Scholar] [CrossRef] [PubMed]
- König, C.; Maekawa, H.; Schiebel, E. Mutual Regulation of Cyclin-Dependent Kinase and the Mitotic Exit Network. J Cell Biol 2010, 188, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Visintin, R.; Craig, K.; Hwang, E.S.; Prinz, S.; Tyers, M.; Amon, A. The Phosphatase Cdc14 Triggers Mitotic Exit by Reversal of Cdk- Dependent Phosphorylation. Mol Cell 1998, 2, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.H.; Good, V.M.; Tonks, N.K.; Barford, D. The Structure of the Cell Cycle Protein Cdc14 Reveals a Proline-Directed Protein Phosphatase. Embo J 2003, 22, 3524–3535. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.P.; Hall, H.; Chaparian, R.; Mara, M.; Mueller, A.; Hall, M.C.; Shannon, K.B. Dephosphorylation of Iqg1 by Cdc14 Regulates Cytokinesis in Budding Yeast. Molecular biology of the cell 2015, 26, 2913–2926. [Google Scholar] [CrossRef] [PubMed]
- Bloom, J.; Cristea, I.M.; Procko, A.L.; Lubkov, V.; Chait, B.T.; Snyder, M.; Cross, F.R. Global Analysis of Cdc14 Phosphatase Reveals Diverse Roles in Mitotic Processes *. Journal of Biological Chemistry 2011, 286, 5434–5445. [Google Scholar] [CrossRef]
- Palani, S.; Meitinger, F.; Boehm, M.E.; Lehmann, W.D.; Pereira, G. Cdc14-Dependent Dephosphorylation of Inn1 Contributes to Inn1–Cyk3 Complex Formation. Journal of Cell Science 2012, 125, 3091–3096. [Google Scholar] [CrossRef] [PubMed]
- Shou, W.; Seol, J.H.; Shevchenko, A.; Baskerville, C.; Moazed, D.; Chen, Z.W.; Jang, J.; Charbonneau, H.; Deshaies, R.J. Exit from Mitosis Is Triggered by Tem1-Dependent Release of the Protein Phosphatase Cdc14 from Nucleolar RENT Complex. Cell 1999, 97, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Kuilman, T.; Maiolica, A.; Godfrey, M.; Scheidel, N.; Aebersold, R.; Uhlmann, F. Identification of Cdk Targets That Control Cytokinesis. EMBO J 2015, 34, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Varela Salgado, M.; Adriaans, I.E.; Touati, S.A.; Ibanes, S.; Lai-Kee-Him, J.; Ancelin, A.; Cipelletti, L.; Picas, L.; Piatti, S. Phosphorylation of the F-BAR Protein Hof1 Drives Septin Ring Splitting in Budding Yeast. Nat Commun 2024, 15, 1–17. [Google Scholar] [CrossRef]
- Meitinger, F.; Boehm, M.E.; Hofmann, A.; Hub, B.; Zentgraf, H.; Lehmann, W.D.; Pereira, G. Phosphorylation-Dependent Regulation of the F-BAR Protein Hof1 during Cytokinesis. Genes Dev 2011, 25, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Meitinger, F.; Palani, S.; Hub, B.; Pereira, G. Dual Function of the NDR-Kinase Dbf2 in the Regulation of the F-BAR Protein Hof1 during Cytokinesis. Mol Biol Cell 2013, 24, 1290–1304. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, C.; Reymond, A.; Cerutti, L.; Utzig, S.; Hofmann, K.; Simanis, V. The S. Pombe Cdc15 Gene Is a Key Element in the Reorganization of F-Actin at Mitosis. Cell 1995, 82, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Willet, A.H.; McDonald, N.A.; Bohnert, K.A.; Baird, M.A.; Allen, J.R.; Davidson, M.W.; Gould, K.L. The F-BAR Cdc15 Promotes Contractile Ring Formation through the Direct Recruitment of the Formin Cdc12. The Journal of cell biology 2015, 208, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Carnahan, R.H.; Gould, K.L. The PCH Family Protein, Cdc15p, Recruits Two F-Actin Nucleation Pathways to Coordinate Cytokinetic Actin Ring Formation in Schizosaccharomyces Pombe. The Journal of cell biology 2003, 162, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Graziano, B.R.; Yu, H.Y.; Alioto, S.L.; Eskin, J.A.; Ydenberg, C.A.; Waterman, D.P.; Garabedian, M.; Goode, B.L. The F-BAR Protein Hof1 Tunes Formin Activity to Sculpt Actin Cables during Polarized Growth. Molecular biology of the cell 2014, 25, 1730–1743. [Google Scholar] [CrossRef] [PubMed]
- Garabedian, M.V.; Wirshing, A.; Vakhrusheva, A.; Turegun, B.; Sokolova, O.S.; Goode, B.L. A Septin-Hof1 Scaffold at the Yeast Bud Neck Binds and Organizes Actin Cables. Molecular Biology of the Cell 2020, 31, 1988–2001. [Google Scholar] [CrossRef] [PubMed]
- Garabedian, M.V.; Stanishneva-Konovalova, T.; Lou, C.; Rands, T.J.; Pollard, L.W.; Sokolova, O.S.; Goode, B.L. Integrated Control of Formin-Mediated Actin Assembly by a Stationary Inhibitor and a Mobile Activator. Journal of Cell Biology 2018, 217, 3512–3530. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, K.A.; Gould, K.L. Cytokinesis-Based Constraints on Polarized Cell Growth in Fission Yeast. PLOS Genetics 2012, 8, e1003004. [Google Scholar] [CrossRef] [PubMed]
- Vallen, E.A.; Caviston, J.; Bi, E. Roles of Hof1p, Bni1p, Bnr1p, and Myo1p in Cytokinesis in Saccharomyces Cerevisiae. Mol Biol Cell 2000, 11, 593–611. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Schreiter, J.; Nishihama, R.; Wloka, C.; Bi, E. Targeting and Functional Mechanisms of the Cytokinesis-related F-BAR Protein Hof1 during the Cell Cycle. Molecular Biology of the Cell 2013, 24, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Frost, A.; Unger, V.M.; De Camilli, P. The BAR Domain Superfamily: Membrane-Molding Macromolecules. Cell 2009, 137, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, S.; Toyooka, K.; Senju, Y. Subcellular Membrane Curvature Mediated by the BAR Domain Superfamily Proteins. Seminars in cell & developmental biology 2010, 21, 340–349. [Google Scholar] [CrossRef]
- Zhao, H.; Michelot, A.; Koskela, E.V.; Tkach, V.; Stamou, D.; Drubin, D.G.; Lappalainen, P. Membrane-Sculpting BAR Domains Generate Stable Lipid Microdomains. Cell reports 2013, 4, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Picas, L.; Viaud, J.; Schauer, K.; Vanni, S.; Hnia, K.; Fraisier, V.; Roux, A.; Bassereau, P.; Gaits-Iacovoni, F.; Payrastre, B.; et al. BIN1/M-Amphiphysin2 Induces Clustering of Phosphoinositides to Recruit Its Downstream Partner Dynamin. Nat Commun 2014, 5, 5647. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Main cell cycle events relevant for budding yeast cytokinesis and cell division. See text for details.
Figure 1.
Main cell cycle events relevant for budding yeast cytokinesis and cell division. See text for details.
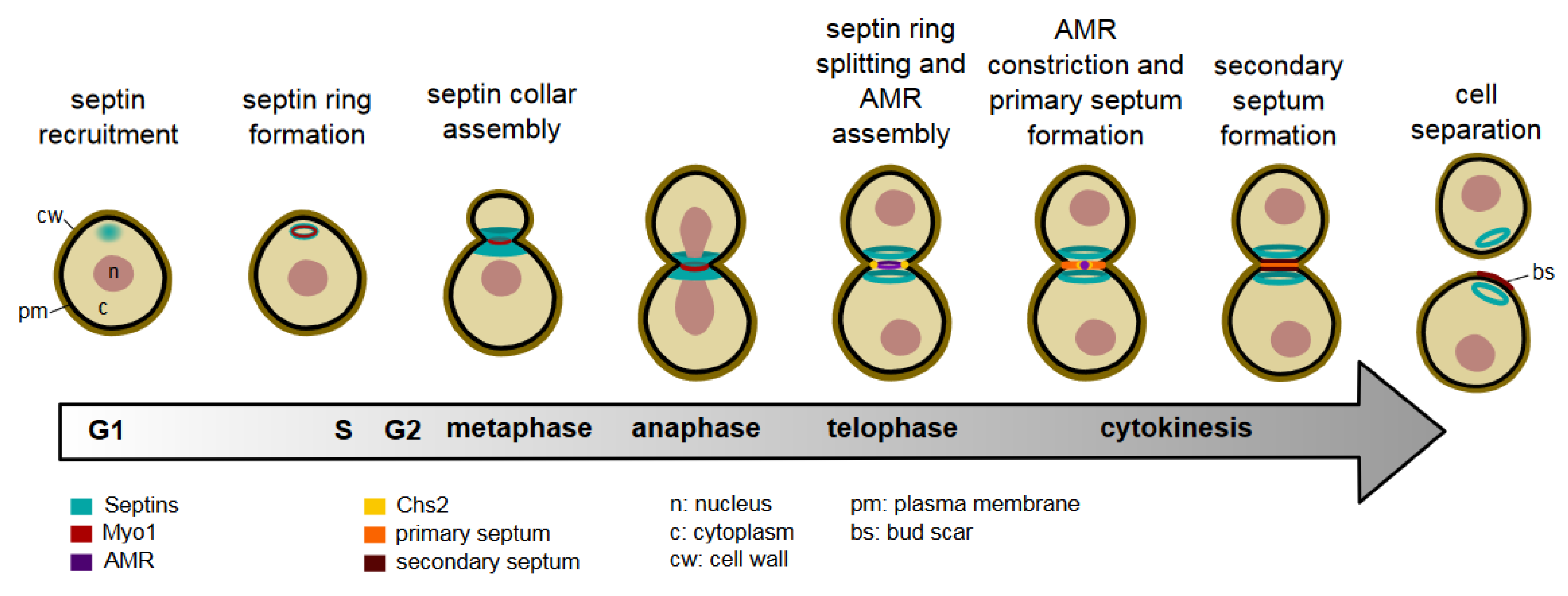
Figure 2.
Septin architecture during different cell cycle phases. Septins are first recruited in G1 to the presumptive bud side as an unorganised patch that is rapidly converted into a cortical ring formed by radial septin filaments. Septin recruitment and/or ring formation involve the Cdc42 GTPase, in turn activated by the scaffold Bem1 and the GEF Cdc24, the Cdc42 effectors Cla4 and Gic1-2, the formin Bni1 and the bud site selection protein Axl2. At S phase entry, the septin ring expands into a septin collar that spans the bud neck and then matures into an hourglass-shaped structure formed by axial and circumferential septin filaments. Septin collar assembly involves the kinases Cla4, Gin4 and Elm1, CDKs, the septin-interacting protein Bni5 and the ubiquitin ligases Dma1 and Dma2. At the onset of cytokinesis, the axial septin filaments are depolymerised and possibly repolymerised into two arrays of circumferential filaments that make a double ring sandwiching the constricting AMR. Septin reorganisation requires the MEN (Mitotic Exit Network), its phosphorylation target Hof1 and other unidentified effectors. The septin double ring is patterned and stabilised by the Bud3 protein and the anillin-like Bud4. Solid arrows indicate established regulatory relationships; dashed arrows indicate hypothetical regulatory interactions. See text for further details.
Figure 2.
Septin architecture during different cell cycle phases. Septins are first recruited in G1 to the presumptive bud side as an unorganised patch that is rapidly converted into a cortical ring formed by radial septin filaments. Septin recruitment and/or ring formation involve the Cdc42 GTPase, in turn activated by the scaffold Bem1 and the GEF Cdc24, the Cdc42 effectors Cla4 and Gic1-2, the formin Bni1 and the bud site selection protein Axl2. At S phase entry, the septin ring expands into a septin collar that spans the bud neck and then matures into an hourglass-shaped structure formed by axial and circumferential septin filaments. Septin collar assembly involves the kinases Cla4, Gin4 and Elm1, CDKs, the septin-interacting protein Bni5 and the ubiquitin ligases Dma1 and Dma2. At the onset of cytokinesis, the axial septin filaments are depolymerised and possibly repolymerised into two arrays of circumferential filaments that make a double ring sandwiching the constricting AMR. Septin reorganisation requires the MEN (Mitotic Exit Network), its phosphorylation target Hof1 and other unidentified effectors. The septin double ring is patterned and stabilised by the Bud3 protein and the anillin-like Bud4. Solid arrows indicate established regulatory relationships; dashed arrows indicate hypothetical regulatory interactions. See text for further details.
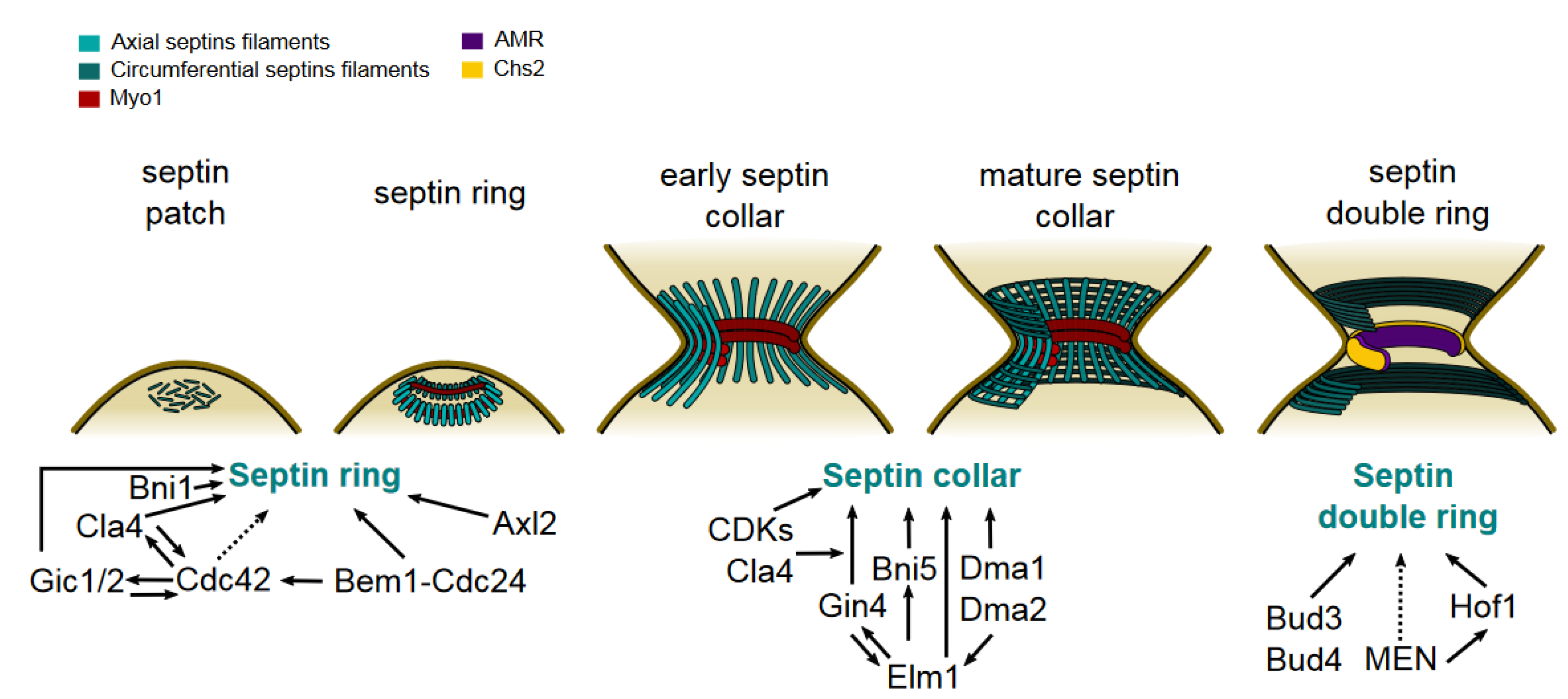
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



