Submitted:
29 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
The Ylistrum japonicum is a commercially valuable scallop known for its long-distance swimming abilities. Despite its economic importance, genetic and genomic research on this species is limited. This study presents the first complete mitochondrial genome of Y. japonicum. The mitochondrial genome is 19,475 bp long and encompasses 13 protein-coding genes, 3 ribosomal RNA genes, and 23 transfer RNA genes. By selecting 15 Pectinidae species and additional outgroup taxa, we have constructed phylogenetic trees to elucidate the evolutionary placement of Ylistrum within the family Pectinidae. Our analysis reveals that Ylistrum is a basal lineage to the Pectininae clade, distinct from its previously assigned tribe, Amusiini. This study offers critical insights into the genetic makeup and evolutionary history of Y. japonicum, enhancing our knowledge of this economically vital species.
Keywords:
saucer scallop
; Ylistrum japonicum
; mitochondrial genome
; taxonomy
1. Introduction
The genus Ylistrum, a member of the phylum Mollusca, class Bivalvia, order Pectinoida, family Pectinidae, established by Mynhardt et al. [1] in 2014, derived from the Greek word “ylistro”, meaning “glide”, reflecting the gliding habits of this group, and dividing the original genus Amusium into two genera, Ylistrum and Amusium. Both Ylistrum and Amusium primarily found in the Indian-Pacific region and exhibit similar lifestyles and morphological characters [1,2,3]. Despite this similarities, the evolutionary relationships and placement of Ylistrum within the family Pectinidae are not conclusively determined. Research using molecular phylogenetic studies has indicated that the Ylistrum is genetically distinct from its close relatives in the genus Amusium studies by Alejandrino et al. [4], Mynhardt et al. [1], and Sherratt et al. [5] support this genetic separation. However, the precise phylogenetic placement of Ylistrum within the family remains a topic of debate. Serb [6] proposed that the Ylistrum might belong to the tribe Decatopectinini, but this suggestion has not been strongly supportted by the abaliable evidence.
Ylistrum species are known for their long-distance swimming or gliding abilities, and their distinctive features include colorful left and right shells and radiating ribs on the inner shell [1,3,7]. There are two extant species within the Ylistrum genus that are found across the globe: Ylistrum japonicum (Gmelin,1791) was originally discovered in Japan [8], and Ylistrum balloti (Bernardi, 1861), which predominantly encountered in Australia’s western, eastern, and southern regions [3,9], as well as in New Caledonia [10,11]. There is also a known fossil species from Morgan Limestone [12], Ylistrum morganense (Beu and Darragh, 2001). Y. balloti is pivotal to the commercial trawl fisheries in Australia, warranting extensive research. In contrast, studies on Y. japonicum are scarcer, with most research originating from Japan and South Korea. Okada [13] have delved into the species’ ecology and morphology, while Kanmizutaru and Anraku [14] have investigated the effects of MgCl2 injection into the adductor muscle for shell opening, and in 2005, Kanmizutaru et al. [15] have also assessed the pallial eyes’ light perception through electroretinogram tests. In South Korea, research have illuminated including its reproductive cycle [16], the development of its gonads, the age at first sexual maturity, and the sex ratio [17], as well as the correlation between age and growth [18]. In China, there is limited research available, focusing primarily ecology and the possibilities of artificial breeding [19,20].
The taxonomic classification of Y. japonicum is still unclear due to a lack of molecular data. In a recent taxonomic investigation, Dijkstra and Beu [3] have provisionally maintained Ylistrum within the Amusiini tribe, awaiting a conclusive molecular phylogenetic analysis of the Pectinidae family.
2. Results
2.1. Mitochondrial Genome Composition
The mitochondrial genome structure of of Y. japonicum is depicted Figure 1 and detailed in Table 1. The complete mitochondrial genome sequence has been submitted to NCBI GenBank database (Accession number: PP571649). The structure is a typical circular closed double-stranded molecule with a total length of 19475 bp, containing 39 genes including 13 protein-coding genes (PCGs), 3 ribosomal RNAs, and 23 transfer RNAs. The base composition is as follows: (A) constitutes 21.9%, thymine (T) 36%, guanine (G) 29%, and cytosine (C) 13.1%. The mitochondrial genome exhibits an A+T content of 57.9% and a G+C content of 42.1%.
2.2. Protein-Coding Genes
There are a total of 13 protein-coding genes, 12 out of the 13 Protein-coding genes (PCGs) of Y. japonicum commonly found across most pectinid species [21,22]. Notebly, gene atp8 which is typically abesent in most mitogenomes of most bivalve [23,24] is present in Y. japonicaum. The total length of PCGs was 11884 bp, comprising approximately 61% of the complete genome. Among the genes only four (atp6, cox1, cox2, nad4L) utilize the standard start codon ATG. And the remaining nine had alternative start codon, six genes (atp8, cox3, nad1, nad2, nad4, nad6) had GTG, two TTG (cytb, nad5) and nad3 had ATC. Seven genes had the TAG stop codon, five had TAA, and cytb was terminated by T.
2.3. rRNA and tRNA Genes
The rrnS (rrn12) gene spans 957 bp (from position 15459 to 16415) while rrnL (rrn16) gene has two copies with length 1492 bp and 1486 bp (18-1509, 16453-17939 respectively). The mitochondrial genome of Y. japonicum contains 23 tRNA genes ranging in length from 65 to 72 nucleotides. Three tRNA genes are present in two copies, all of three tRNA genes was found with distict anticodons. Two trnS (rRNA-ser) have UCU or UGA, two trnL (tRNA-Leu) had UAA or UAG, and two trnM (rRNA-Met) had UAU or CAU. The occurrence multiple trnM genes in the mitochondiral genomes of bivalves is common [21], and the presence of two copies of trnS also frequently observed in most mitochondiral genomes of most animals [25]. None of the tRNA genes overlapp with any protein-coding genes (PCGs).
2.4. Gene Order
The occurrence of mitogenomes rearrangement is prevalent among mollusks [26], arrangement observed in mitogenome of Y. japonicum is also a novel configuration for the family Pectinidae, with no matching gene junctions found in other Pectinidae species (Figure 2). Species with higher gene order similarity were selected for comparison and newly annotated atp8 genes according to Malkócs et al. [25]. Due to the lacks of annotation of rRNA sequence, Mizuhopecten yessoensis (FJ595959) was excluded from the gene order analysis, and based on the high similarity of gene order between three Argopecten species [27], only one species was selected as a representative.
Comparing gene arrangements of four selected species, there is one gene cluster “nad6-trnL-cytb” were shared. When excluding the tRNA genes, four scallop species shared another gene clusters “nad1-rrnL-cox1”; Gene cluster “nad4L-cox3” present in Y. japonicum, Argopecten irradians irradians, Amusium pleuronctes and gene cluster “cox3-nad2-nad3” were shared by Y. japonicum, A. pleuronctes and Chlamys farreri. The “nad5-atp6” cluster in Y. japonicum is split by the insertion of cox2, make it different from the other two Pectininae species. The gene cluster “nad5-atp6-rrnS” was also shared between A. irradians irradians and A. pleuronctes when those variable tRNA were excluded, this indicates the close phylogenetic affinity between two species as well.
2.5. Gene Collinearity
Gene collinearity analysis using the progressiveMauve algorithm in Mauve, delineated 7 locally collinear blocks (LCBs) across complete mitochondrial genome of 5 Pectininae species (Figure 3). These LCBs are conserved across all mitogenomes analyzed, although variations in the sequence order are evident among the different species. The order of LCBs demonstrated a high degree of similarity among the three Argopecten species, indicating their close evolutionary relationship. In contrast, Y. japonicum exhibited a significantly distict order of LBCs arrangement when compared to both three Argopecten species and A. pleuronectes.
2.6. Phylogenetic Analysis
To delve deeper into the phylogenetic position of Y. japonicum and the taxonomic status of family Pectinidae, a phylogenetic tree (Figure 4) was constructed based on complete or nearly complete mitochondrial genome data of various Pectinidae species and outgroup taxa. The results of the phylogenetic analysis were found to be comparable with previous studies by Smedley et al. [28], Yao et al. [29], and Malkócs et al. [25], and were mostly consistent with Waller’s classification [30,31]. Phylogenies based on two methods (Maximum Likelihood and Bayesian inference) of the concatenated protein sequences showed almost complete agreement, with high bootstrap values or posterior probabilities supporting all nodes. The systematic arrangement, as proposed by Waller [31], subdivides the family Pectindae into four subfamilies: Pectininae, Chlamydinae, Pallioline, and Camptonectinae. The outgroup Mytilinae and Crassotreinae were found to be consistent with the phylogenetic position proposed by Xu et al. [21], where the clade Mytilinae forms a sister group with the clade Osteridae+Pectinidae.
Our study has confirtmed the previously hypothesized monophylicity of Pectinidae, as concluded by Waller [32]. However, due to the absence of complete mitochondrial genome sequences of the Camptonectinae, no representatives from this subfamily were included in the phylogenetic analysis. The species within Pectinidae were effectively categorized into three subfamilies Palliolinae, Chlamydinae, and Pectininae. Placopecten magellanicus, serving as the representative of the Palliolinae, was positioned at the basal position of the branch Palliolinae+Chlamydinae. The clade Palliolinae+Chlamydinae was well supported as the sister group to the Pectininae clade [33,34]. Within the subfamily Chlamydinae, M. yessonesis and C. farreri were found to have the closest relationship, forming a sister taxon with Mimachlamys, consistent with the results obtained by Xu et al. [21]. Ylistrum was identified as a lineage basal to the clade Pectininae, separated from its considered tribe Amusiini, a conclusion supported by Alejandrino et al. [4], Sherratt et al. [5], and Serb [6]. Argopecten were clustered on the same branch, forming a sister group with the clade Amusium+Pecten. The close relationship between A. pleuronectes and Pecten maximus + Pecten albicans was also noted, consistent with studies of Barucca et al. [35], Alejandrino et al. [4], and Feng et al. [34].
Another ML tree (Figure 5) was constructed based on 16S rRNA sequences, three specimens of Y. japonicum from China (PP571649) and Japan (HM622702, KF982785) [4,36] were selected. The result was comparable with Mynhardt et al. [1]. Genus Amusium and Pecten forming a sister group again. Antillipecten antillarum as a lineage basal, forming a sister group with the clade Anguipecten+Ylistrum. Two Ylistrum taxa formed a sister clade and well separated with Amusium. Specimens HM622702 and KF982785 were in same branch, sharing an ancestor with the Chinese individual. All Y. japonicum finally converged into the same branch, this provided evidence that it was a monophyletic clade, and also shown the closely genetic distance between its individual.
2.7. Systematic Descriptions
Order Pectinida Gray, 1854
Superfamily Pectinoidea Rafnesque, 1815
Family Pectinidae Rafnesque, 1815
Subfamily Pectininae Rafnesque, 1815
Tribe Decatopectinini Waller, 1986
Genus Ylistrum Mynhardt and Alejandrino, 2014
Ylistrum japonicum (Gmelin, 1791)
Type locality: Japan.
Distribution: Japan (South of central Honshu Island), Korean (Jeju Island), China (Taiwan Province, Guangdong Province, Guangxi Province and Hainan Province.).
Morphological description: Shell large, round, smooth, and glossy. The left valve is dark red to reddish-brown, covered with concentrically arranged dark brown fine lines and spots, The color is slightly lighter at the umbo, with small light-colored spots. The right valve is slightly flat, pale yellow to light tan, white near the umbo, also concentrically arranged brown spots on surface. Two small auricles and slightly different in size, The color of auricles on left valve is darker The inner surface of the shell is white, with the left valve having a yellow to light brown edge, sometimes the inner edge of left valve is pale brown. Interior radial ribbing on both valves, the specimen collected from the Hailing Island with 33-43 ribs on the left valve and 42-49 ribs on the right valve .
Remarks: In original description, the species group from China were record as a subspecies Amusium japonicum taiwanicum Habe, 1992 [37], and now its a synonymised name of Y. japonicum. Unlike individuals from Japan, the color of specimens from China are not bright, and the concentrically arranged brown spots are presence on the right valve, these are the morphological difference between the individuals from above two producing areas. Despite there are morphological differences, their molecular biological evidence indicates that they are the same species. The counting of internal rib by different author is not always the same (e.g. Zhang et al. [38], Wang [7], Mynhardt et al. [1]). In addition, as counts had completely overlapping rangesand it also could not be used to differentiate between the two Ylistrum species [1]. Overall, the most significant difference between Y. japonicum and Y. balloti are the color of their auricles on right valve and the spots in a concentric pattern on both two valves, to the former species, the auricles on right valve are generally darker, and spots always appear along with theirs valves repair marks.
When mixed with the long-ribbed scallop in trawl nets, 15 out of 52 scallops were Y. japonicum in one trawl, with sandy bottom sediment and others are A. pleuronectes. The average shell length of Y. japonicum, in the population was 84.27±10.63mm (n=15), the average shell height was 82.97±9.48mm, and the average shell width was 16.70±2.23mm.
3. Discussion
The family Pectinidae, as a clade within Bivalvia, exhibits a significant range of morphological and behavioral variations, rendering it of great importance in ecology, evolution, and commercial activities. Nevertheless the taxonomy of Pectinidae has long been a source of debate within the scientific community.
Waller [30] hypothesis for the classification and evolution of Pectinidae based on morphology, particularly focusing on pre-radial stage shell microsculpture, and incorporating fossil data and geological evidence. In 2006, Waller revised his scallopidae phylogenetic hypothesis in conjunction with previous molecular genetic studies, leading to the successful establishment of a stable classification method. Much of the contemporary taxonomic research on Pectinidae is grounded in the taxonomy framework developed by Waller, as evidenced by the work of Serb [6] and Smedley et al. [28]. Despite the enhancements to Waller‘s hypothesis, which have been fueled by advances in molecular techniques and the expansion of fossil evidence, controversies over the correct classification of Pectinidae remain In molecular phylogeny, discrepancies in findings can stem from a variety of factors, such as the choice of single genetic sequences [39] or the concatenation of multiple sequences [40,41]. Additionally, the accuracy and limitations of phylogenetic tree construction methods [6,42,43], the number and distribution of sampled species, the selection of outgroups, and the processing of sequences can all influence the results obtained. A case in point is the phylogenetic placement of the Palliolinae, a monophyletic subgroup within the Pectinidae. Different studies have placed the Palliolinae within various clades, either with the Pectininae or the Chlamyinae, highlighting the ongoing inconsistencies in phylogenetic resolution within this family (e.g. Alejandrino et al. [4], Sherratt et al. [5], Xu et al. [21], Lin et al. [22], Malkócs et al. [25], Li et al. [27], Smedley et al. [28], Saavedra and Peña [33], Feng et al. [34] and Malkowsky and Klussmann-Kolb [40]). In contrast, Waller‘s hypothesis placed Palliolinae with Pectininae as a sister group [31]. On the mito-phylogenomics level, differences in sequence selection and methodologies can lead to varying results, as evidenced by studies by Lin et al. [22], Malkócs et al. [25] and Li et al. [27]. The divergence time estimation analysis conducted by Lin et al. [22] based on concatenated mitochondrial PCG gene sequences yielded a similar topology to our studies, suggesting that similarities and differences in results may be related to sample selection. In summary, differences in these results can be attributed to variations in sequence selection, analytical approaches, and species sampled.
Although the robustness of a phylogenetic tree can be affected by a variety of factors, it is noteworthy that Ylistrum and Amusium have consistently been distinguished in past molecular phylogenetic investigations. The placement of Ylistrum within the subfamily Pectininae is well-supported, as evidenced by the study of Matsumoto and Hayami [39] and subsequent research. Nevertheless, due to the significant morphological and distributional similarities between Y. japonicum and A. pleuronectes, Ylistrum has historically been grouped with the Amusiini, even though molecular genetic studies have consistently pointed to its distinctiveness from Amusium. Our phylogenetic study based on complete mitochondrial genome indicate that Ylistrum has ancient origins, but its preciseplacement within the subfamily Pectininae remains ambiguous due to insufficient sample data. Alejandrino et al. [4] analyzed the phylogeny of 81 extant taxa from the Pectinidae, based on the nuclear Histone H3, 12S rRNA, 16S rRNA data and 28S rRNA data, the result shows that Ylistrum was placed among the species of the tribe Decatopectinini, and Aumsium were nest in different clade (Pectinini). Subsequently, Smedley et al. [28] expanded the dataset to 62 Pectinidae species on the basis of Alejandrino et al. [4] into a new phylogenetic analysis, clade Ylistrum was once again placed in the Decatopectinini tribe. Interestingly, it forms a sister clade with two Annachlamys species belongs to the tribe Pectinini, but the phylogenetic location of Annachlamys was still debated [3]. Mynhardt et al. [1] focused on the phylogenetic analysis of Aumsium and Ylistrum, restored their respective monophyletic clades and described Ylistrum as a new genus. In this study, Ylistrum also form a sister groups with a Decatopectinini species (A. antillarum). Our phylogenetic analysis based on 16S rRNA indicates that Y. japonicum from Japan and China are monophyletic, and same as former studies, two Ylistrum taxa were still nest in the tribe Decatopectinini. In light of these findings and previous studies, we adhere to Serb‘s classification [6], positioning Ylistrum within the Decatopectinini tribe.
The complete mitochondrial genome sequence data for Ylistrum remains inadequate. The available data for Y. balloti (accession number ON041136) is in fact, based on an erroneous identification. When analyzed using NCBI-BLAST (with an alignment length greater than 1200), this sequence exhibits a high degree of similarity (greater than 98.99%) to Y. japonicum, but less than 94.10% similarity to Y. balloti. This strongly suggests that ON041136 is more closely related to Y. japonicum. Additionally, the collection location of ON041136 is Beihai, Guangxi, China, which is problematic since there are no documented distributions of Y. balloti within China. The mention of Y. balloti in the Chinese Zoology book [7] is considered a misidentification, highlighting the need for a critical revision of this information.
4. Materials and Methods
4.1. Sample Collection
A total of 50 specimens were collected during 2022 to 2023 from Hailing Island, Yangjiang City, Guangdong Province, China (21.61N, 111.93E) (Figure 6). Morphometric measurements were performed using an electronic vernier caliper (0.1mm), body measurement traits (shell length, shell height, shell width, shell weighet, etc.) were recorded for further investigation. All collected samples are intended for commercial purposes, and there are no concerns regarding animal ethics. The morphological characteristics of these specimens were categorized and compared in accordance with Zhang et al. [38] and Zhang [44]. From this collection, one specimen had its adductor muscles extracted (5g), which were then preserved in a -80℃ freezer for subsequent analytical procedures.
4.2. DNA Extraction, Library Preparation and Next Generation Sequence
gDNA was extracted by MagPure Bacterial DNA Kit (Magen, Guangzhou, China) following pre-grind in liquid nitrogen. Qubit dsDNA HS assay kit (Sangon, Shanghai, China) was used to test concentration and 1% Agarose gel electrophoresis to confirm intergrity. The library preparation and next generation sequence was finished by Sangon Biotech (Shanghai) Co., Ltd.. First, 500ng quantified DNA was randomly fragmented by Covaris (Woburn, USA). Next, Hieff NGS®MaxUp II DNA Library Prep Kit for Illumina® (YEASEN, Shanghai, China) was used for next steps. Briefly, Endprep enzyme was added to repair end and 3’.end A tail ligation. Then adaptor was ligated by enhancer and Fast T4 DNA ligase. Index primer was added by PCR and the amplified product about 400bp was selected by DNA selection beads. The library concertration and size was confirmed by Qubit 4.0 (Thermo, Waltham, USA) and 2% agarose gel electrophoresis respectively.
Then the libraries were pooled and loaded on Novaseq 6000 (Illumina. San Diego, USA)/DNBseq-T7 (BGI, Shenzhen, China) sequencer by 2×150 bp paired end sequence kit according to the manufacture´s instructions.
4.3. Sequence Assembly and Annotation
Rawbases yielded at least 6 GB was used for downstream analysis. First, all of the raw reads were trimmed by Fastqc v0.11.2 [45]. The software SPAdes v3.15 [46] was used to assemble the raw sequence reads into contigs. tBLASTn and GeneWise were used to obtain the CDS gene boundary by reverse alignment with the near-source reference database, the tRNA sequence annotation was obtained by MiTFi, Rfam used cmsearch alignment identifies non-coding rRNA and the final summary into complete annotation results. The circular gene maps of the specie Y. japonicum was drawn by Circos.
4.4. Gene Collinearity
Gene collinearity among complete mitochondrial sequences of five Pectininae species was explored to assess their phylogenetic relationship with Y. japonicum, using the progressiveMauve algorithm and default parameters (including default seed weight, determine locally collinear blocks, and full alignment) in the Mauve v2.4.0 [47].
4.5. Phylogenetic Analysis with Mitochondrial Genome
Two phylogenetic analyses were conducted based on the complete mitochondrial sequences of Y. japonicum in this study. Following the meyhodologies established in previous studies by Malkowsky and Klussmann-Kolb [39], Xu et al. [21] and Malkócs et al. [25], a total of 15 mitochondrial sequences of Pectinidae species and outgroup taxa were selected to construct phylogenetic trees. This selection includes 11 Pectinidae species across three subfamilies: Pectininae, Palliolinae, and Chlamydinae. The available mitochondrial genome sequences were obtained from GeneBank, incorporating 13 complete mitochondrial genomes and 2 incomplete sequences (KP900974, KP900975), each over 16,000 base pairs in length. Two Ostreidae species Magallana bilineata, Magallana gigas and two Mytilidae species Mytilus galloprovincialis, Mytilus trossulus were used as outgroups.
PhyloSuite v1.2.3 [48] was utilized to extract the protein-coding genes (PCGs) from each sequence. All Sequences were aligned in batches with MAFFT v7.505 [49]. The alignments were refined using the codon-aware program MACSE v2.06 [50] which preserves reading frame and allows incorporation of sequencing errors or sequences with frameshifts. Ambiguously aligned fragments of the alignments were removed in batches using Gblocks 0.91b [51]. ModelFinder v2.2.0 [52] was used to select the best-fit partition model. The phylogenetic tree was subsequently constructed using both the Maximum-likelihood (ML) method in IQ-TREE v2.2.0 [53,54] and Bayesian inference (BI) in MrBayes v3.2.7a [55]. Branch support was determined with 5000 bootstrap iterations for best-scoring ML tree. Markov Chain Monte Carlo (MCMC) analyses were run for 1,000,000 generations (sampling every 1000 generations), in which initial 50% of sampled data were discarded as burn-in. The result was beautified with FigTree v1.4.4.
To explore in more detail the monophyletic development of Y. japonicum and its taxonomic position in Pectininae, another ML tree was constructed using 16S rRNA by PhyloSuite v1.2.3 [48], 3 Y. japonicum specimens from different regions (China and Japan) and the other 8 specimens of subfamliy Pectininae were selected. All Sequences were aligned in batches with MAFFT v7.505 [49] and pruned by Gblocks 0.91b [51]. Branch support was determined with 5000 bootstrap iterations for best-scoring ML tree. List of specimens included in molecular studies shown in Table 2.
Author Contributions
Conceptualization, Y.T.; Methodology, Y.T.; Investigation, Y.H., X.Y. and Y.T.; Resources,Y.T.; Funding acquisition, Y.T. and Z.H.; Writing—original draft, Y.H. and X.Y; Writing—review & editing, Y.T., J.M. and X.W.; Supervision,Y.C.
Funding
This research was supported by the National Key Research and Development Program of China (2022YFD2400302); the Central Government Subsidy Project for Liaoning Fisheries (2023); Science and Technology Foundation of Dalian (2021JB11SN035).
Institutional Review Board Statement
The experimental protocol was designed in accordance with the recommendations of the Regulations of the Laboratory Animal Guideline for Ethical Review of Animal Welfare (National Standards of P. R. China, GB/T 35823—2018) and reviewed and approved by the animal care and use committee of Dalian Ocean University (DLOU-2023009).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mynhardt, G.; Alejandrino, A.; Puslednik, L.; Corrales, J.; Serb, J.M. Shell shape convergence masks biological diversity in gliding scallops: Description of Ylistrum n. gen. (Pectinidae) from the Indo-Pacifc Ocean. J. Mollus. Stud. 2014, 80, 400–411. [Google Scholar] [CrossRef]
- Dijkstra, H.H. Pectinoidea (Bivalvia: Propeamussiidae and Pectinidae) from the Panglao region, Philippine Islands. Vita Malacologica. 2013, 10, 1–108. [Google Scholar]
- Dijkstra, H.H.; Beu, A.G. Living scallops of Australia and adjacent waters (Mollusca: Bivalvia: Pectinoidea: Propeamussiidae, Cyclochlamydidae and Pectinidae). Rec. Aust. Mus. 2018, 70, 113–330. [Google Scholar] [CrossRef]
- Alejandrino, A.; Puslednik, L.; Serb, J.M. Convergent and parallel evolution in life habit of the scallops (Bivalvia: Pectinidae). BMC Evol Biol. 2011, 11, 1–9. [Google Scholar] [CrossRef]
- Sherratt, E.; Alejandrino, A.; Kraemer, A.C.; Serb, J.M.; Adams, D.C. Trends in the sand: directional evolution in the shell shape of recessing scallops (Bivalvia: Pectinidae). Evolution, 2016, 70, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Serb, J.M. Reconciling morphological and molecular approaches in developing a phylogeny for the Pectinidae (Mollusca: Bivalvia). In Scallops Biology, Ecology, Aquaculture, and Fisheries, 3rd ed.; Shumway, S.E., Parsons, G.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–29. [Google Scholar]
- Wang, Z.R. Fauna Sinica Invertebrate Vol. 31 Mollusca Bivalvia. Science Press: Beijing, BJ, CN, 2002; pp.1-374.
- Gmelin, J.F. In Gmelin J.F. (Ed.) Caroli a Linnaei Systema Naturae per Regna Tria Naturae, 13 ed, Tom. 1, Pars 6. Lipsiae: Georg. Emanuel Deer, 1791; pp. 3021-3910.
- Bernardi, M. Description d‘espéces nouvellas. Jour. de conchyl, 1861, 9, 46–49. [Google Scholar]
- Abbott, R.T.; Dance, S.P. Compendium of seashells: A colour guide to more than 4,200 of the world‘s marine shells. EP Dutton Inc.: New York, NY, USA, 1982; pp.1-411.
- Carpenter, K.E.; Niem, V.H. The living marine resources of the Western Central Pacifc. Volume 2: Cephalopods, crustaceans, holothurians and sharks. Food and Agriculture Organization of the United Nations, Rome, 1998; pp. 1–686.
- Beu, A.G.; Darragh, T.A. Revision of southern Australian Cenozoic fossil Pectinidae (Mollusca: Bivalvia). Proceedings of the Royal Society of Victoria, 2001, 113, 1–205. [Google Scholar]
- Okada, Y.; Uchida, S.; Uchida, T. New Illustrated Encyclopediaof the Fauna of Japan. Ⅱ. Hokuryukan, Tokyo, TKY, JPN, 1998; pp. 1-803.
- Kanmizutaru, T.; Anraku, K. Inducement of Shell Opening of Amusium japonicum by MgCl2 Injection into the Adductor Muscle. Nippon Suisan Gakk. 1999, 65, 856–859. [Google Scholar] [CrossRef]
- Kanmizutaru, T.; Anraku, K.; Toyoda, S. Light perception capability of pallial eyes in Japanese moon scallop Amusium japonicum as determined by electroretinogram. Nippon Suisan Gakk. 2005, 71, 928–934. [Google Scholar] [CrossRef]
- Son, P.W.; Chung, E.Y. 1994. Annual Reproductive Cycle and Size at First Sexual Maturity of the Sun and Moon Scallop Amusium Japonlcum Japonicum (Gmelin, 1791) (Bivalvia: Pectinidae) in the Coastal Waters of Jejudo, Korea. Malacologia. 1994, 51, 119–129. [Google Scholar] [CrossRef]
- Son, P.W.; Chung, E.Y. Gonadal Development, First Sexual Maturity and Sex Ratio of the Sun and Moon Scallop Amusium japonicum japonicum on the Coastal Waters of Jejudo, Korea. Developmant & Reproduction. 2005, 9, 95–103. [Google Scholar]
- Son, P.W.; Ha, D.S.; Rho, S.; Chang, D.S. Studies on the Age and Growth fo Sun and Moon Scallop, Amusium japonicum japonicum (GMELIN). Journal of Aquaculture. 1996, 9, 409–417 [In Korean]. [Google Scholar]
- Ye, W.J.; Liang, G.Y. A proliminary study on artificial breeding of Amussium japonica (Gmelin). Transactions of Oceanology and Limnology. 1989, 4, 86–93 [In Chinese]. [Google Scholar]
- Ye, W.J.; Liang, G.Y. A preliminary observation on the ecology of Amussium japonica (Gmelin). Chinese Journal of Zoology. 1990, 25, 5–7 [In Chinese]. [Google Scholar]
- Xu, K.; Kanno, M.; Yu, H.; Li, Q.; Kijima, A. Complete mitochondrial DNA sequence and phylogenetic analysis of Zhikong scallop Chlamys farreri (Bivalvia: Pectinidae). Mol Biol. Rep. 2011, 38, 3067–3074. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Li, Y.X.; Sun, Y.; Tao, J.; Qiu, J.W. A new species of the genus Catillopecten (Bivalvia: Pectinoidea: Propeamussiidae): morphology, mitochondrial genome, and phylogenetic relationship. Front. Mar. Sci. 2023, 10, 1168991. [Google Scholar] [CrossRef]
- Boore, J.L; Medina, M.; Rosenberg, L.A. Complete sequences of the highly rearranged molluscan mitochondrial genomes of the scaphopod Graptacme eborea and the bivalve Mytilus edulis. Mol. Biol. Evol. 2004, 21, 1492–1503. [Google Scholar] [CrossRef]
- Smith, D.R.; Snyder, M. Complete mitochondrial DNA sequence of the scallop Placopecten magellanicus: evidence of transposition leading to an uncharacteristically large mitochondrial genome. J. Mol. Evol. 2007, 65, 380–391. [Google Scholar] [CrossRef]
- Malkócs, T.; Viricel, A.; Becquet, V.; Evin, L.; Dubillot, E.; Pante, E. Complex mitogenomic rearrangements within the Pectinidae (Mollusca: Bivalvia). BMC Ecol Evol. 2022, 22, 29. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–80. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Y.; Zhong, T.; Heng, X.; Ao, T.; Gu, Z.; Wang, A.; Liu, C.; Yang, Y. The Complete Mitochondrial Genomes of Two Rock Scallops (Bivalvia: Spondylidae) Indicate Extensive Gene Rearrangements and Adaptive Evolution Compared with Pectinidae. Int. J. Mol. Sci. 2023, 24, 13844. [Google Scholar] [CrossRef] [PubMed]
- Smedley, G.D.; Audino, J.A.; Grula, C.; Porath-Krause, A.; Pairett, A.N.; Alejandrino, A.; Serb, J.M. Molecular phylogeny of the Pectinoidea (Bivalvia) indicates Propeamussiidae to be a non-monophyletic family with one clade sister to the scallops (Pectinidae). Mol. Phylogenet. Evol. 2019, 137, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.J.; Yu, H.W.; Liu, Y.R.; Zhang, Y.H.; Li, Y.l. The complete mitochondrial genome and phylogenetic analysis of Amusium pleuronectes. Mitochondrial DNA B. 2020, 5, 2318–2319. [Google Scholar] [CrossRef] [PubMed]
- Waller, T.R. Evolutionary relationships among commercial scallops (Mollusca: Bivalvia: Pectinidae). In Scallops: Biology, Ecology and Aquaculture; Shumway, S.E., Ed.; Elsevier: New York, NY, USA, 1991; pp. 1–73. [Google Scholar]
- Waller, T.R. New phylogenies of the Pectinidae (Mollusca: Bivalvia): Reconciling morphological and molecular approaches. In Scallops: Biology, Ecology and Aquaculture. Shumway S.E., Parsons G.J., Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2006; pp. 1–44. [Google Scholar]
- Waller, T.R. The ctenolium of scallop shells: functional morphology and evolution of a key family-level character in the Pectiniacea (Mollusca: Bivalvia). Malacologia. 1984, 25, 20. [Google Scholar]
- Saavedra, C.; Peña, J.B. Phylogenetics of American scallops (Bivalvia: Pectinidae) based on partial 16S and 12S ribosomal RNA gene sequences. Mar Biol. 2006, 150, 111–119. [Google Scholar] [CrossRef]
- Feng, Y.W.; Li, Q.; Kong, L.F.; Zheng, X.D. DNA barcoding and phylogenetic analysis of Pectinidae (Mollusca: Bivalvia) based on mitochondrial COI and 16S rRNA genes. Mol Biol. Rep. 2011, 38, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Barucca, M.; Olmo, E.; Schiaparelli, S.; Canapa, A. Molecular phylogeny of the family Pectinidae (Mollusca: Bivalvia) based on mitochondrial 16S and 12S rRNA genes. Mol. Phylogenet. Evol. 2004, 31, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Marin, A.; Fujimoto, T.; Arai, K. The variable 5′end of the 16S rRNA gene as a novel barcoding tool for scallops (Bivalvia, Pectinidae). Fisheries sci. 2015, 81, 73–81. [Google Scholar] [CrossRef]
- Habe, T. New name for Amusium japonicum formosum. Venus. 1992, 50, 235. [Google Scholar]
- Zhang, X.; Qi, Z.Y.; Li, J.M.; Ma, X.T.; Wang, Z.R.; Huang, X.M.; Zhuang, Q.Q. Bivalve mollusca of The South China sea. Science Press: Beijing, BJ, CHN. 1960; pp.1-274.
- Matsumoto, M.; Hayami, I. Phylogenetic analysis of the family Pectinidae (Bivalvia) based on mitochondrial cytochrome C oxidase subunit I. J. Mollus. Stud. 2000, 66, 477–488. [Google Scholar] [CrossRef]
- Malkowsky, Y.; Klussmann-Kolb, A. Phylogeny and spatio-temporal distribution of European Pectinidae (Mollusca: Bivalvia). Syst. Biodivers. 2012, 10, 233–242. [Google Scholar] [CrossRef]
- Puslednik, L.; Serb, J.M. Molecular phylogenetics of the Pectinidae (Mollusca: Bivalvia) and the effect of outgroup selection and increased taxon sampling on tree topology. Mol. Phylogenet. Evol. 2008, 48, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.-X.; Li, Y.; Hittinger, C.T.; Chen, X.; Rokas, A. An investigation of irreproducibility in maximum likelihood phylogenetic inference. Nat. Commun. 2020, 11, 6096. [Google Scholar] [CrossRef]
- Haag, J.; Höhler, D.; Bettisworth, B.; Stamatakis, A. From Easy to Hopeless-Predicting the Difficulty of Phylogenetic Analyses. Mol. Biol. Evol. 2022, 39, msac254. [Google Scholar] [CrossRef]
- Zhang, S.P. Atlas of Marine Mollusks of China. China Ocean Press:Beijing, BJ, CHN. 2008, pp.1-383.
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics, 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A. S.; Lesin, V. M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. http://wwwgenomeorg/cgi/doi/101101/gr2289704. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Xiang, C.; Gao, F.; Jakovlić, I.; Lei, H.; Hu, Y.; Zhang, H.; Zou, H.; Wang, G.; Zhang, D. Using PhyloSuite for Molecular Phylogeny and Tree-based Analyses. iMeta. 2023, 2, e87. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. MACSE v2: toolkit for the alignment of coding sequences accounting for frameshifts and stop codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods. 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
Figure 1.
Mitochondrial genome map of Ylistrum japonicum in this study.
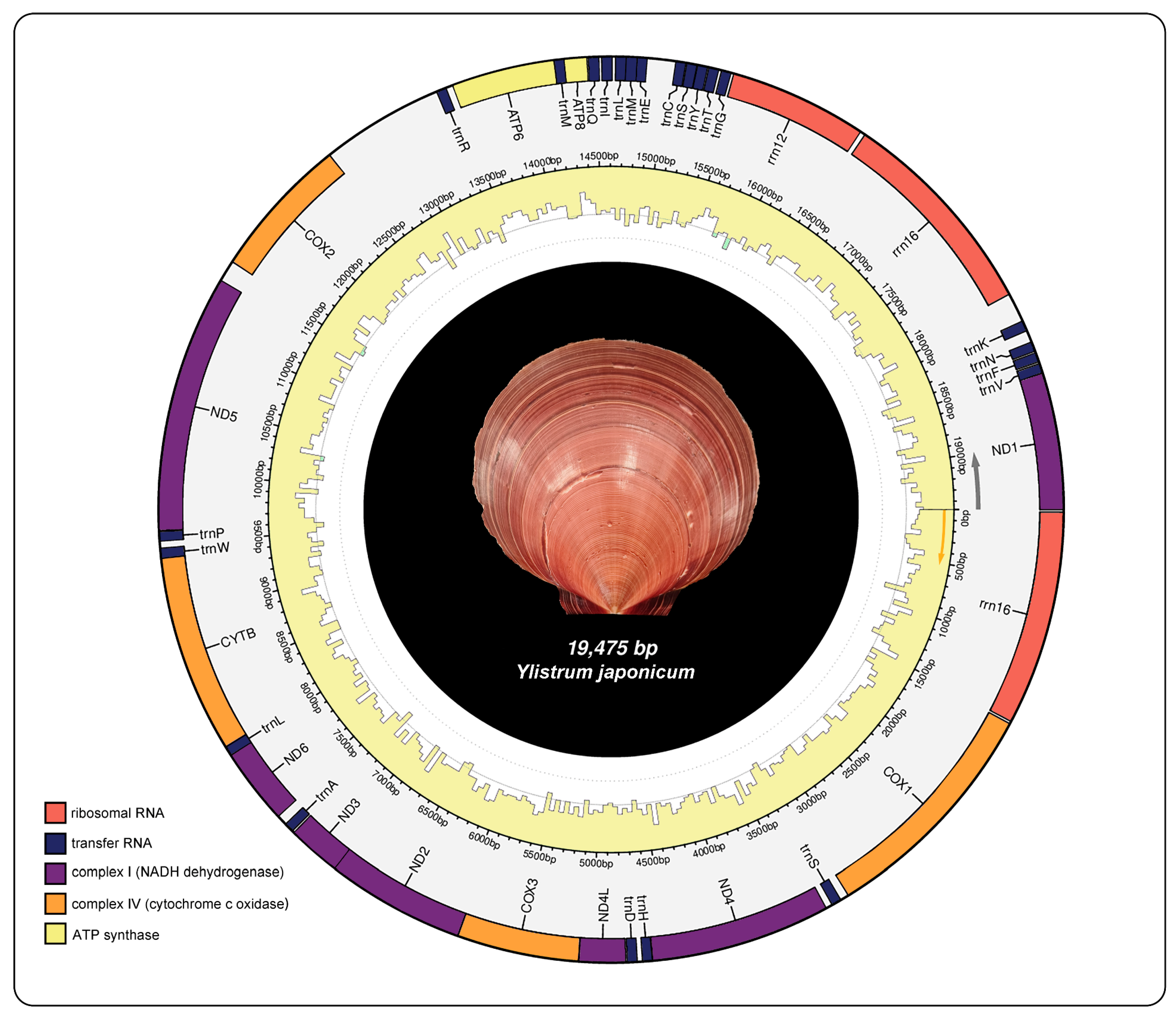
Figure 2.
Gene orders of Ylistrum japonicum, Argopecten irradians irradians, Amusium pleuronctes and Chlamys farreri, with newly annotated atp8 genes. The same color indicate identical gene junctions (excluding the tRNA genes).
Figure 2.
Gene orders of Ylistrum japonicum, Argopecten irradians irradians, Amusium pleuronctes and Chlamys farreri, with newly annotated atp8 genes. The same color indicate identical gene junctions (excluding the tRNA genes).
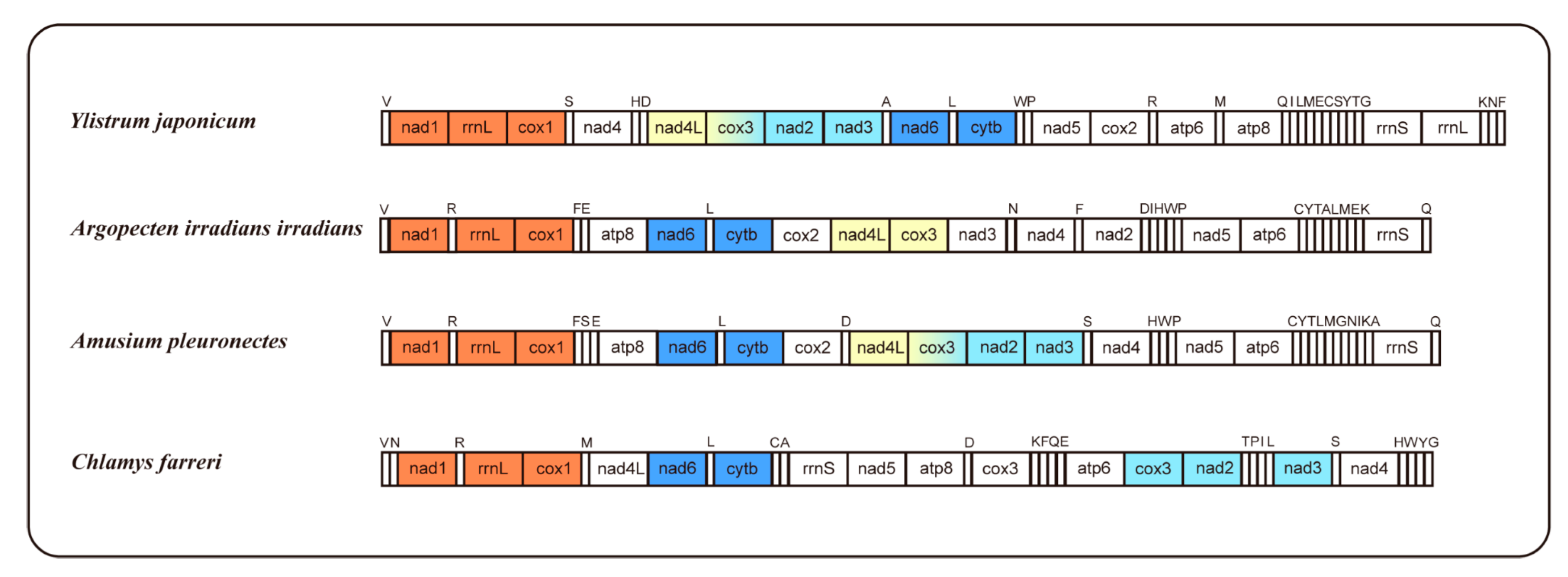
Figure 3.
Gene collinearity analysis of 5 Pectininae species. The level of similarity at each position is shown in the blocks. The white, red and green boxes represent protein coding, rRNA, tRNA genes.
Figure 3.
Gene collinearity analysis of 5 Pectininae species. The level of similarity at each position is shown in the blocks. The white, red and green boxes represent protein coding, rRNA, tRNA genes.
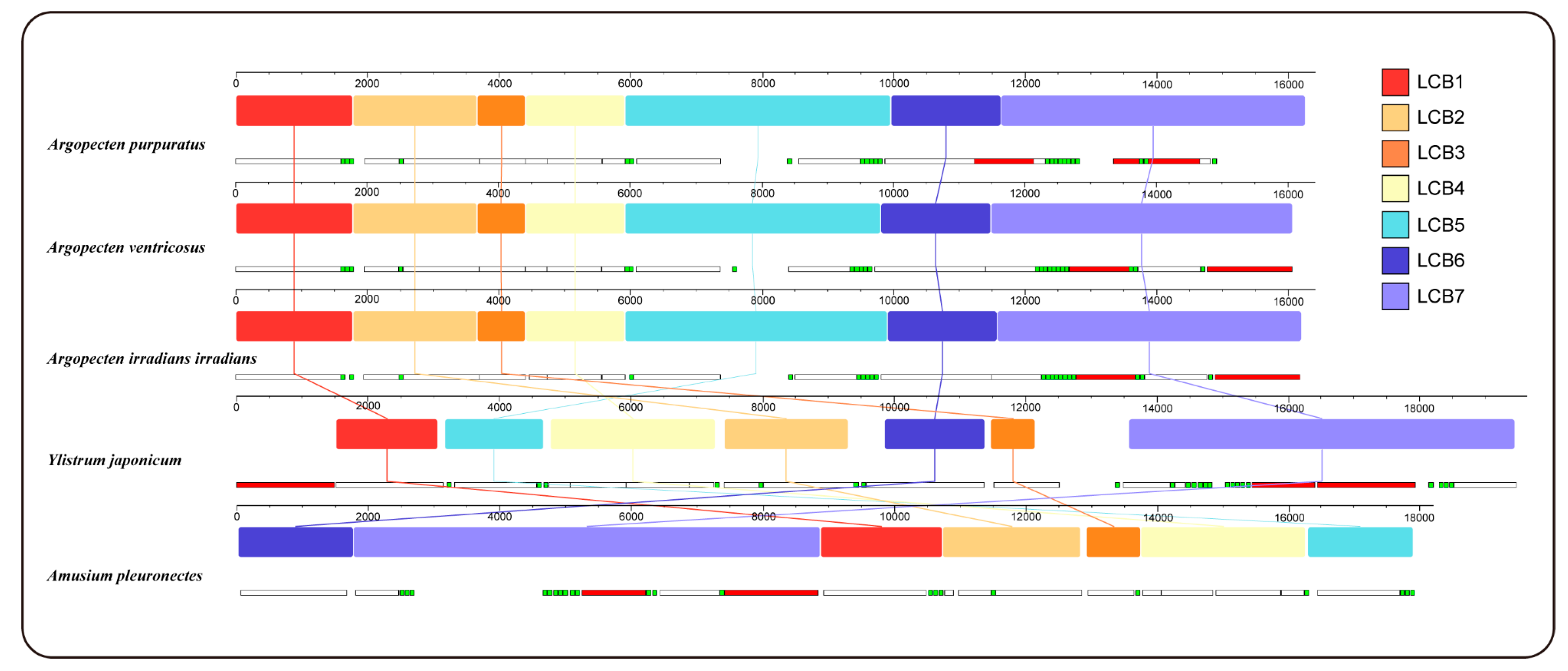
Figure 4.
Phylogenetic tree derived from Maximum likelihood (ML) and Bayesian inference (BI) based on the sequences of mitochondrial protein-coding genes (PCGs). Numbers above the branches indicate bootstrap support; numbers below branches are Bayesian posterior probability. A dash indicates no support for that node.
Figure 4.
Phylogenetic tree derived from Maximum likelihood (ML) and Bayesian inference (BI) based on the sequences of mitochondrial protein-coding genes (PCGs). Numbers above the branches indicate bootstrap support; numbers below branches are Bayesian posterior probability. A dash indicates no support for that node.
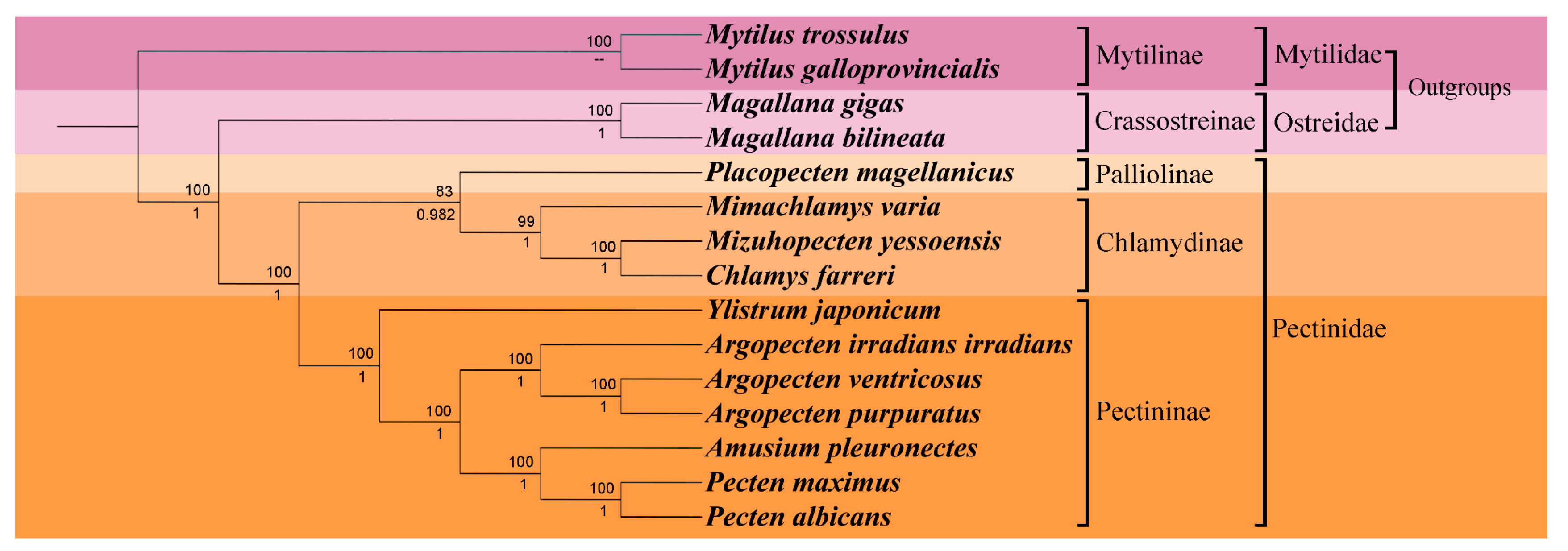
Figure 5.
Phylogenetic tree of genus Ylistrum and some species from three tribe of Pectininae inferred by Maximum likelihood (ML) of 16S rRNA sequences. Numbers indicate bootstrap support.
Figure 5.
Phylogenetic tree of genus Ylistrum and some species from three tribe of Pectininae inferred by Maximum likelihood (ML) of 16S rRNA sequences. Numbers indicate bootstrap support.
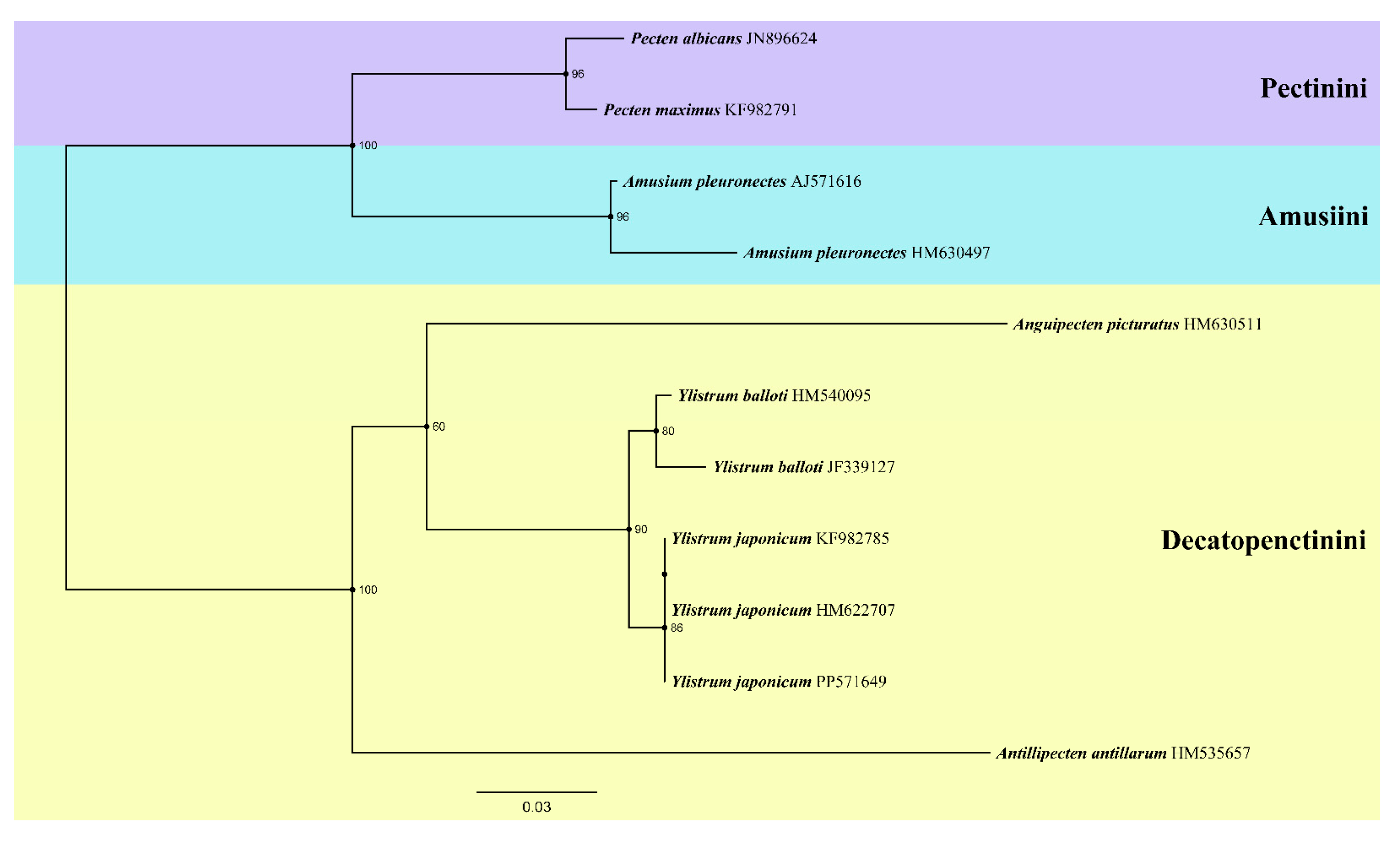
Figure 6.
Sampling location of Ylistrum japonicum (modified from d-maps: https://d-maps.com).
Figure 6.
Sampling location of Ylistrum japonicum (modified from d-maps: https://d-maps.com).
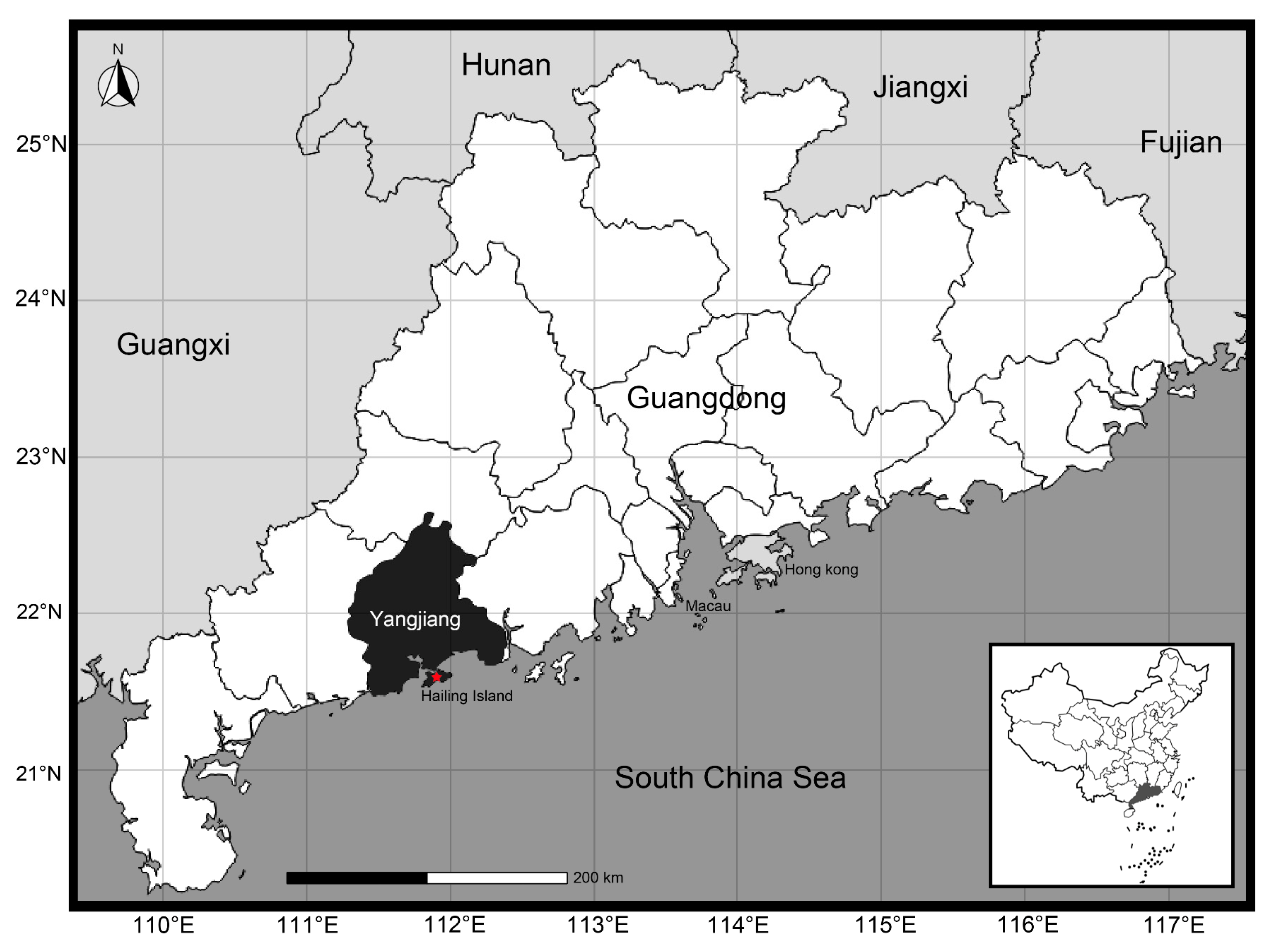

Table 2.
Structural features of the mitochondrial genome of Ylistrum japonicum.
| Gene | Sequence location | Size (bp) | Start codon | Stop codon | Intergenic nucleotide (bp) |
|---|---|---|---|---|---|
| rrnL | 18-1509 | 1492 | 17 | ||
| cox1 | 1527-3158 | 1632 | ATG | TAA | 17 |
| trnS | 3219-3286 | 68 | 60 | ||
| nad4 | 3334-4581 | 1248 | GTG | TAG | 47 |
| trnH | 4588-4652 | 65 | 6 | ||
| trnD | 4690-4757 | 68 | 37 | ||
| nad4L | 4769-5089 | 321 | ATG | TAA | 11 |
| cox3 | 5094-5939 | 846 | GTG | TAG | 4 |
| nad2 | 5942-6904 | 963 | GTG | TAA | 2 |
| nad3 | 6904-7275 | 372 | ATC | TAA | -1 |
| trnA | 7291-7356 | 66 | 15 | ||
| nad6 | 7431-7958 | 528 | GTG | TAA | 74 |
| trnL | 7960-8024 | 65 | 1 | ||
| cytb | 8028-9402 | 1375 | TTG | T | 3 |
| trnW | 9403-9471 | 69 | 0 | ||
| trnP | 9522-9586 | 65 | 50 | ||
| nad5 | 9594-11381 | 1788 | TTG | TAG | 7 |
| cox2 | 11532-12524 | 993 | ATG | TAG | 150 |
| trnR | 13380-13447 | 68 | 855 | ||
| ato6 | 13498-14211 | 714 | ATG | TAG | 50 |
| trnM | 14216-14280 | 65 | 4 | ||
| atp8 | 14284-14439 | 156 | GTG | TAG | 3 |
| trnQ | 14451-14521 | 71 | 11 | ||
| trnI | 14542-14612 | 71 | 20 | ||
| trnL | 14638-14706 | 69 | 25 | ||
| trnM | 14713-14784 | 72 | 6 | ||
| trnE | 14788-14853 | 66 | 3 | ||
| trnC | 15053-15118 | 66 | 199 | ||
| trnS | 15129-15195 | 67 | 10 | ||
| trnY | 15205-15271 | 68 | 9 | ||
| trnT | 15283-15350 | 68 | 11 | ||
| trnG | 15372-15437 | 66 | 21 | ||
| rrnS | 15459-16415 | 957 | 21 | ||
| rrnL | 16453-17939 | 1486 | 37 | ||
| trnK | 18145-18216 | 72 | 205 | ||
| trnN | 18301-18367 | 67 | 84 | ||
| trnF | 18381-18445 | 65 | 13 | ||
| trnV | 18459-18525 | 67 | 13 | ||
| nad1 | 18528-19475 | 948 | GTG | TAG | 2 |
Table 2.
List of specimens included in the molecular studies.
| Subfamily | Tribe in this research | Previous Tribe | Species | Genbank accession numbers | Genetic compartments |
| Ingroup | |||||
| Aequipectinini | Aequipectinini | Argopecten irradians irradians | DQ665851 | mitogenome | |
| Aequipectinini | Aequipectinini | Argopecten purpuratus | KT161260 | mitogenome | |
| Aequipectinini | Aequipectinini | Argopecten ventricosus | KT161261 | mitogenome | |
| Pectininae | Amusiini | Amusiini | Amusium pleuronectes | MT419374 | mitogenome |
| Amusiini | Amusiini | Amusium pleuronectes | AJ571616 | 16S rRNA | |
| Amusiini | Amusiini | Amusium pleuronectes | HM630497 | 16S rRNA | |
| Decatopectinini | Amusiini | Ylistrum japonicum | PP571649 | mitogenome | |
| Decatopectinini | Amusiini | Ylistrum japonicum | KF982785 | 16S rRNA | |
| Decatopectinini | Amusiini | Ylistrum japonicum | HM622707 | 16S rRNA | |
| Decatopectinini | Amusiini | Ylistrum balloti | HM540095 | 16S rRNA | |
| Decatopectinini | Amusiini | Ylistrum balloti | JF339127 | 16S rRNA | |
| Decatopectinini | Decatopectinini | Anguipecten picturatus | HM630511 | 16S rRNA | |
| Decatopectinini | Decatopectinini | Antillipecten antillarum | HMS35657 | 16S rRNA | |
| Pectinini | Pectinini | Pecten maximus | KP900975 | mitogenome | |
| Pectinini | Pectinini | Pecten maximus | KF982791 | 16S rRNA | |
| Pectinini | Pectinini | Pecten albicans | KP900974 | mitogenome | |
| Pectinini | Pectinini | Pecten maximus | JN896624 | 16S rRNA | |
| Palliolinae | Palliolini | Palliolini | Placopecten magellanicus | DQ088274 | mitogenome |
| Chlamydini | Chlamydini | Chlamy farreri | EF473269 | mitogenome | |
| Chlamydinae | Fortipectinini | Fortipectinini | Mizuhopecten yessoensis | FJ595959 | mitogenome |
| Mimachlamydini | Mimachlamydini | Mimachlamys varia | MZ520326 | mitogenome | |
| Outgroup | |||||
| Crassostreinae | Magallana bilineata | MT985154 | mitogenome | ||
| Magallana gigas | MZ497416 | mitogenome | |||
| Mytilinae | Mytilus galloprovincialis | DQ399833 | mitogenome | ||
| Mytilus trossulus | AY823625 | mitogenome |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



