
Submitted:
27 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Bicarbonate and CO2 are essential substrates for carboxylation reactions in bacterial central metabolism. In Staphylococcus aureus, the bicarbonate transporter, MpsABC (membrane potential-generating system) is the only carbon concentrating system. A mpsABC deletion mutant can hardly grow in ambient air. In this study, we investigated the changes that occur in S. aureus when it suffers from CO2/bicarbonate deficiency. Electron microscopy revealed that mpsABC has a twofold thicker cell wall thickness compared to the parent strain. The mutant was also substantially inert to cell lysis induced by lysostaphin and the non-ionic surfactant Triton X-100. Mass spectrometry analysis of muropeptides revealed the incorporation of alanine into the pentaglycine interpeptide bridge, which explains the mutant’s lysostaphin resistance. Flow cytometry analysis of wall teichoic acid (WTA) glycosylation patterns revealed a significantly lower -glycosylated and higher ß-glycosylated WTA, explaining the mutant’s increased resistance towards Triton X-100. Comparative transcriptome analysis showed altered gene expression profiles. Autolysin-encoding genes such as sceD, a lytic transglycosylase encoding gene, were upregulated, while genes related to cell wall-anchored proteins, secreted proteins, transporters, and toxins were downregulated. Overall, we demonstrate that bicarbonate deficiency is a stress response that causes changes in cell wall composition and global gene expression resulting in increased resilience to cell wall lytic enzymes and detergents.
Keywords:
Staphylococcus aureus
; bicarbonate transporter
; MpsABC
; cell wall
; peptidoglycan
; transcriptome
1. Introduction
Bicarbonate and CO2 are two major forms of dissolved inorganic carbon (DIC) that serve as essential substrates for many metabolic processes in bacteria. Unlike autotrophic bacteria which can directly fix CO2, nonautotrophic bacteria rely on external sources of bicarbonate/CO2 for anaplerotic reactions. Bicarbonate plays a crucial role in the central metabolism of these bacteria because it is a substrate for biotin carboxylase which is part of the multi enzyme complex, acetyl-CoA carboxylase, that catalyzes the first step in fatty acid synthesis [1,2]. Other carboxylases such as phosphoenolpyruvate carboxylase, pyruvate carboxylase, acetyl-CoA carboxylase among others play a vital role in central metabolism such as fueling the TCA cycle or biosynthesis of amino acids or nucleic bases like uracil and adenine [3,4]. Given the importance of CO2/bicarbonate, bacteria have evolved specialized mechanisms to concentrate these molecules to ensure a constant supply to cells. In numerous organisms, the enzyme carbonic anhydrase (CA) can rapidly convert CO2 to bicarbonate (HCO3-), thereby capturing it in the cytoplasm. In contrast to CO2, bicarbonate cannot freely diffuse in and out of the cell, and thus requires dedicated transporters to facilitate its movement across the cell membrane [5]. In cyanobacteria, three families of bicarbonate transporters have been identified: SbtA, a high-affinity sodium-dependent symporter; BicA, a low-affinity, high-flux sodium-dependent symporter; and BCT1, a high-affinity four-subunit transporter belonging to the ATP-binding cassette family [6,7]. Recently, a novel two-component transporter that facilitates DIC uptake was described in the chemolithoautotroph Hydrogenovibrio crunogenus, and surprisingly, these homologs are also found in nonautotrophic pathogen Staphylococcus aureus [8,9]. In S. aureus, the homologous bicarbonate transporter is encoded by the mpsABC operon, which was initially reported for its functionality as a membrane potential-generating system [10]. Radiolabeled NaH14CO3 uptake profiling showed that MpsABC represents a bicarbonate concentrating system, the first example of such transporter in the phylum Firmicutes [9]. As the only DIC supply system in S. aureus, MpsABC plays an important role in concentrating bicarbonate for key anaplerotic pathways, because CA is not present [11]. Deletion of mpsABC causes severe growth delay under ambient air conditions, which is reversible only by CO2 or bicarbonate supplementation. Of note, MpsABC is highly conserved and widespread in both autotrophic and nonautotrophic bacteria, suggesting its essential role in growth and survival [9,11,12]. In addition, MpsABC has an advantage in species or strains where CO2 diffusion is hampered; for example, in mucus and biofilm-forming bacteria [12]. Recent studies of clinical Methicillin-Resistant S. aureus (MRSA) isolates revealed a novel phenotype, termed ‘NaHCO3 responsiveness’, in which a substantial proportion of MRSA strains exhibit enhanced susceptibility to β-lactams such as cefazolin and oxacillin in the presence of NaHCO3 [13]. It turned out that the NaHCO3 transporter MpsABC is a key contributor to the NaHCO3–β-lactam responsiveness phenotype in MRSA [14]. In this study, we aimed to investigate the significance of bicarbonate in methicillin-resistant S. aureus (MRSA) strain set JE2 in terms of phenotypic and transcriptomic changes by depleting the supply of bicarbonate to the cells. This was accomplished by deleting the mpsABC genes, which encode for the bicarbonate transporter., as it is the only source of bicarbonate entry into the cell. Here, we found by electron microscopy that the mpsABC deletion mutant (ΔmpsABC) has increased thickness in the cell wall (CW). Moreover, the mutant was largely inert to lysostaphin-induced cell lysis and non-ionic surfactant Triton X-100. Using liquid chromatography–tandem mass spectrometry (LC-MS/MS), we further analyzed the peptidoglycan (PG) fragments of the mutant digested with lysostaphin and cellosyl and detected the incorporation of alanine into the pentaglycine interpeptide bridge. We detected a lower content of α-glycosylated and higher content of ß-glycosylated wall teichoic acid (WTA) by flow cytometry. Comparative transcriptome analysis between the parent JE2 and mutant revealed altered gene expression profiles, particularly an upregulation of autolysins in the mutant.
2. Results
2.1. The mpsABC Deletion Mutant Has a Thicker Cell Wall
For our phenotypic studies, mpsABC was deleted in the background of a MRSA strain JE2, a well characterized and plasmid-free derivative of S. aureus USA300 LAC [15]. The deletion of mpsABC in JE2 (ΔmpsABC) (Figure 1a) resulted in an almost complete growth arrest under ambient air conditions (Figure 1b), as previously described for several other S. aureus strains [9,10]. After 24 h cultivation, the absorption at 578 nm (A578) of JE2ΔmpsABC was < 1, whereas the JE2 parent strain reached its stationary phase at an A578 of >10. This growth defect could be rescued by aeration with 5% CO2 (Figure S1). Attempts to isolate a suppressor mutant of JE2ΔmpsABC that thrives under ambient air conditions have failed; consistent with our earlier reports that S. aureus lacks an alternative CO2 - concentrating mechanism [9,11,12].
Although the growth of was severely affected under ambient aeration conditions, it did not completely halt, which enabled us to perform transmission electron microscopy (TEM). The cells were cultivated under ambient air conditions until an A578 of 0.5, which took 2.5 h for the parental strain and 4.5 h for the mutant. TEM image analysis of more than 20 cells showed that the CW of the mutant grown in ambient air was approximately twice as thick compared to the parent strain (Figure 2, Figure S2). In JE2, the average thickness was 15 nm, whereas in the mutant, it was 33 nm (Table 1). When the cells were cultured under 5% CO2, the CW diameter decreased from 15 to 12 nm in JE2 and from 33 to 15 nm in the mutant.
2.2. The ΔmpsABC Mutant Is Largely Inert to Cell Lysis Induced by Lysostaphin and Triton X-100
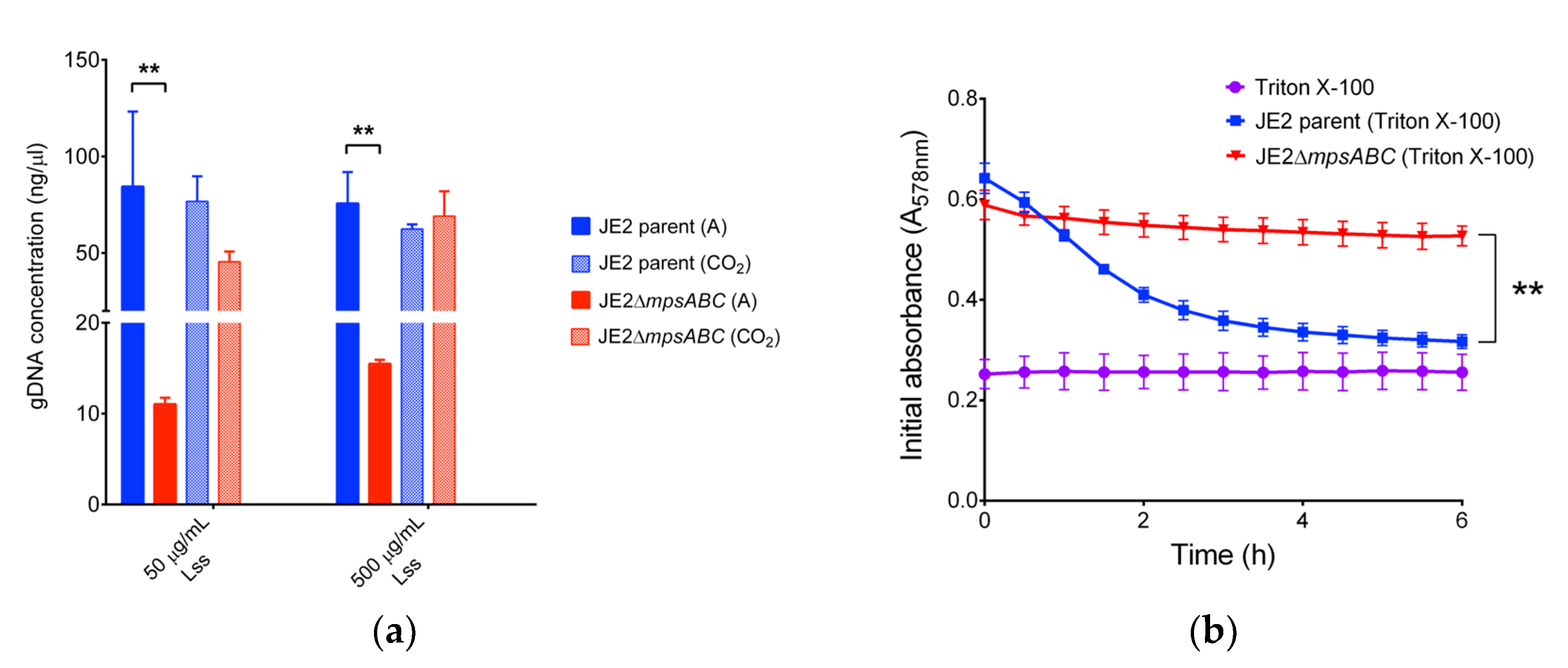
When cells were exposed to CW-degrading lysostaphin, the mutant showed almost a 10-fold lower release of genomic (g) DNA upon cell lysis compared to the parental strain (Figure 3a and Table S1). The mutant also exhibited increased resistance to the non-ionic surfactant Triton X-100, while the same treatment in the parental strain led to a continuous decrease in absorption from A578 0.65 to 0.3 over a 6 h period (Figure 3b).
2.3. The Peptidoglycan (PG) of the Mutant Contains Alanine in the Gly5-Bridge
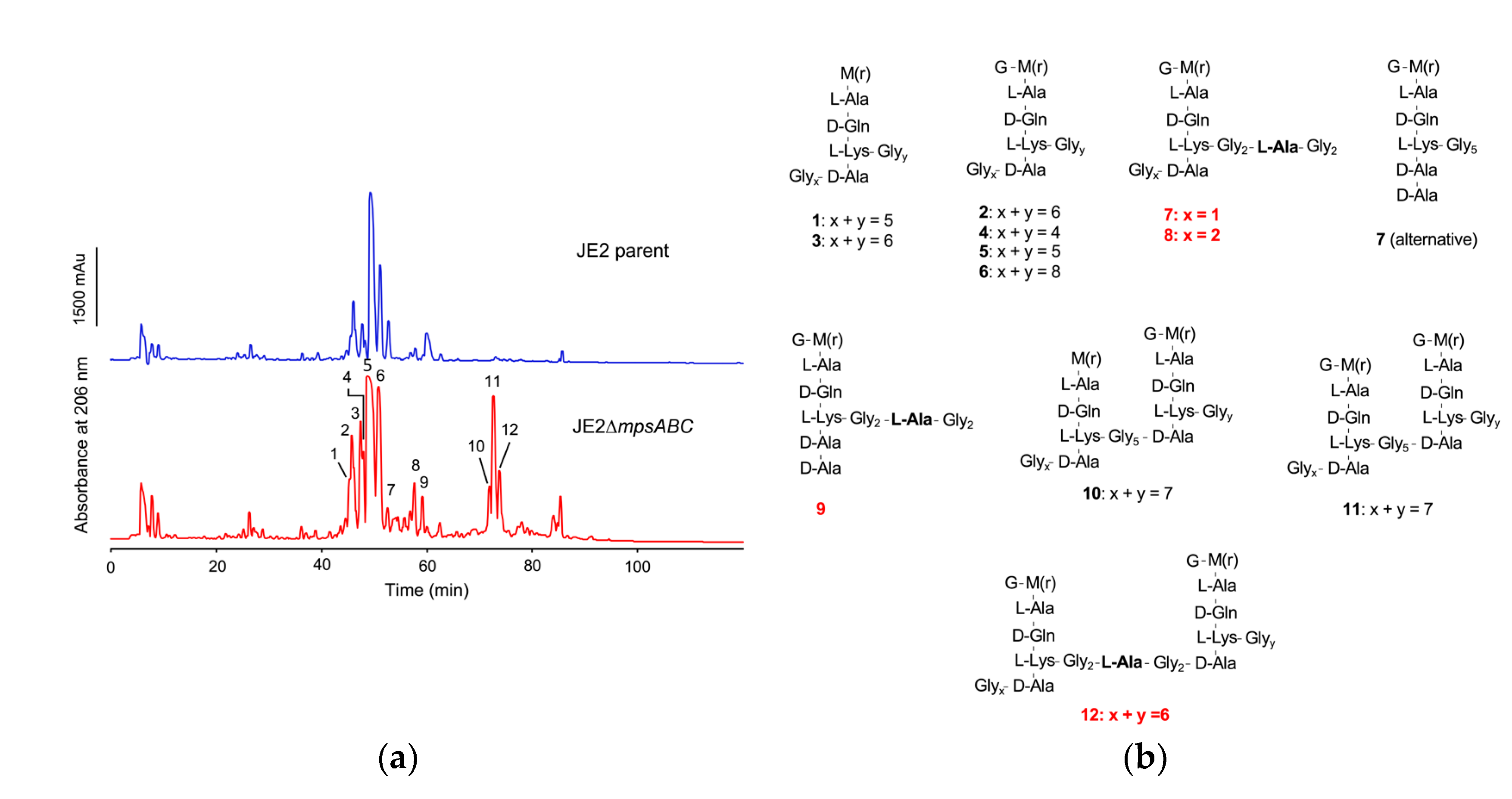
To determine the cause of the increased lysostaphin resistance in the mutant, we analyzed the composition of PG fragments by reversed-phase high-performance liquid chromatography (RP-HPLC) after PG digestion with cellosyl (muramidase from Streptomyces coelicolor), which yields cross-linked and non-cross-linked muropeptides [16]. PG digestion with cellosyl showed no significant differences in the composition between JE2 parent and mutant (Figure S3). When PG was double digested with lysostaphin and cellosyl, peaks at a retention time of ~75 min were detected in the mutant profile only (Figure 4a). The muropeptide peaks of the mutant were analyzed by LC-MS and LC-MS/MS (Figure S4). The calculated masses of peak 7, 8, 9 and 12 were consistent with the incorporation of alanine into the pentaglycine interpeptide bridge (Gly5-bridge). Furthermore, peaks 10, 11, and 12 represented dimers that most likely could not be cleaved owing to the restricted activity of lysostaphin (Figure 4b, Table S3). In summary, PG double digestion with lysostaphin and cellosyl detected the incorporation of alanine into the Gly5-bridge (-Glyx-ala-Glyy-), explaining the increased lysostaphin resistance of the mutant. For determination of the PG cross-linking degree, we compared the area distribution of the mono- and oligomeric muropeptide peaks after mutanolysin digestion to the total peak area. The percentage distribution of muropeptides in the mutant did not change with respect to crosslinking compared to the parent strain (Table S2).
2.4. Comparative Transcriptome Analysis Revealed Differentially Regulated Genes in ΔmpsABC under Ambient Air Condition
To uncover possible genetic factors responsible for the growth defect observed in the mutant strain in ambient air and its thickened cell wall, we performed comparative transcriptome analysis using RNA-sequencing (RNA-seq). The mRNA of the parent and mutant strain was isolated after 4 and 8 h of growth (Figure 1b). While the parent strain was in the mid- or end-exponential phase, no clear growth phase could be determined for the mutant due to its very slow growth. The mutant preculture was cultivated in the presence of 5% CO2, which allowed initial growth due to remaining CO2/bicarbonate. However, the growth stopped after 2-3 h (Figure 1b). During this period, the mutant was presumably affected by CO2 limitation as revealed by gene expression analysis. The complete list of differentially expressed genes is summarized in Excel files ; Table S5 (after 4 h), Table S6 (after 8 h) and Table S7 (5% CO2 condition). Table 2 comprises genes that were significantly up- or down-regulated. Based on their function, we classified them into the following groups: CW lytic enzymes, wall teichoic acid (WTA) biosynthesis, CW-anchored proteins, secreted enzymes, transporters, toxins, regulators, and prophage genes. To validate the RNA-seq data, we performed qRT-PCR analysis of selected genes (Figure S5). RNA-seq and qRT-PCR data showed a high level of correlation. The comparison between the mutant strain and its parental strain after 4 h of growth revealed that many genes (838) were downregulated and only 87 genes were upregulated by at least a factor of 3 (Figure S6a). If we consider the cluster of orthologous groups (COG) of protein categories of the mutant category U (intracellular trafficking and secretion) is 80%, category Q (secondary structure) is 70%, P (inorganic ion transport and metabolism) is 70%, E (amino acid metabolism and transport) and D (cell cycle control and mitosis) were about 70% and category H (coenzyme metabolism) is 100 downregulated. On the other hand, category O (posttranslational modification, protein turnover, chaperone function) was with 80% the highest expressed in the mutant. This might suggest that the mutant counteracts protein folding deficits with upregulation of chaperons and foldases (Figure S6b).
2.5. The ΔmpsABC Mutant’s Wall Teichoic Acids (WTA) Showed a Lower α- and Higher ß-Glycosylation
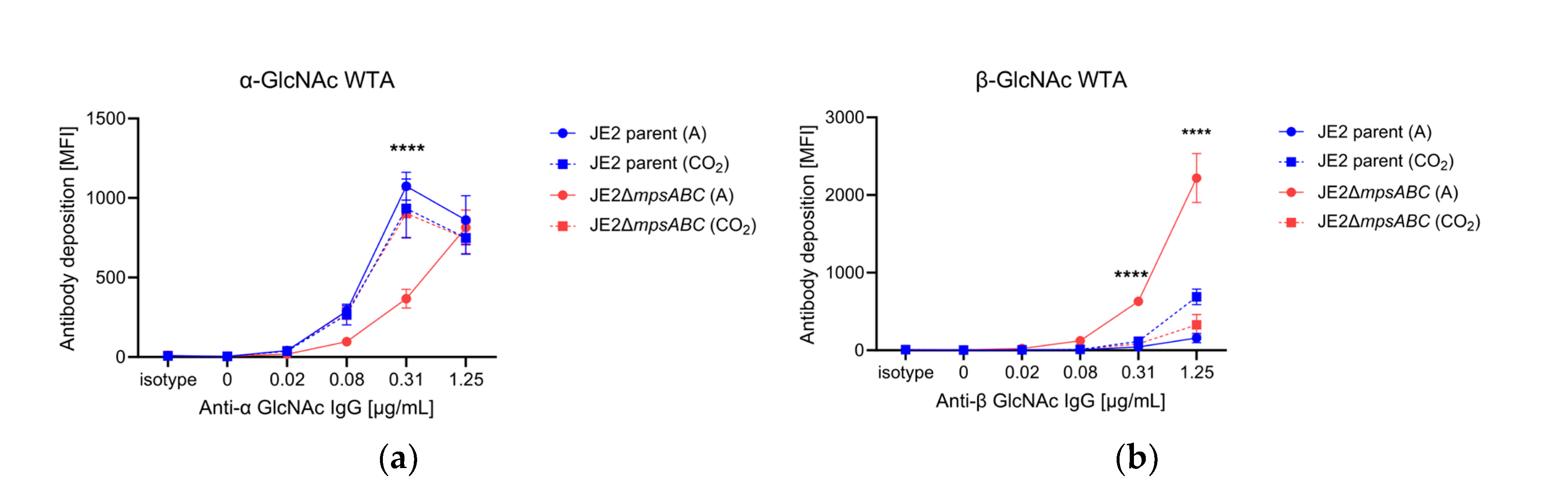
In S. aureus the WTA is glycosylated with GlcNAc either in the α- or β-configuration. TarM is the α-glycosyltransferase while TarS is a β-glycosyltransferase [17,18]. Transcriptome analysis revealed that tarM (α-1,4-glycosyltransferase) was approximately 40-fold downregulated in the ΔmpsABC mutant (Table 1). To see whether this is also associated with reduced α-glycosylation of the WTA, we analyzed the abundancy of α- and ß-glycosylated WTA by flow cytometry with Fab fragments targeting specific glycosylation patterns [19]. Indeed, we found less α-glycosylated WTA and significantly more ß-glycosylated WTA in the mutant strain compared to the parent strain or the mutant exposed to 5% CO2 during growth (Figure 5). This example shows that the genotype correlates well with the phenotype.
3. Discussion
In the present study, we observed significant phenotypic changes in the ΔmpsABC mutant following bicarbonate depletion. The most striking morphological difference between the mutant compared to the parent strain was the massively thickened CW (33 vs. 15 nm). Normally, the CW in S. aureus is approximately 20 nm thick [20,21]. However, CW thickness is not a fixed parameter but varies among strains, growth phase, medium and the presence of antibiotics [20,21]. Inhibition of protein synthesis by antibiotics, such as chloramphenicol, puromycin, or actinomycin, can induce cell wall thickening beyond 100 nm [22,23]. The antibiotic-induced thickening of the CW is probably since CW synthesis continues independently of the inhibition of protein biosynthesis. We assume that bicarbonate depletion with its negative effects on the TCA cycle, respiration and the synthesis of amino acids or other building blocks has a similar effect to the inhibition of translation by antibiotics. Berscheid et al. observed sceD upregulation, CW thickening, lysostaphin- and Triton X-100 resistance in the vancomycin-resistant laboratory mutant strain S. aureus VC40 [24], which was generated by serial passages of S. aureus RN4220ΔmutS in the presence of vancomycin [25]. Based on the RNA-seq of samples from two timepoints (4 and 8 h), alterations of gene expression profiles were observed after 4 h of bacterial growth in ambient air (Table 2). Genes encoding predicted CW hydrolases, such as SceD were highly upregulated (600-fold) and to a lesser extent, IsaA. These hydrolases show sequence identity and similarity, with E. coli soluble lytic transglycosylase (LT) Slt in their PFAM06737 domain and C-terminal part [26]. Other upregulated autolysins were SsaA (a prophage-encoded amidase), LytM (a glycine-glycine endopeptidase), and LysM. LysM has a PG-binding domain that is involved in bacterial PG synthesis and remodeling [27,28]. The mutant’s CW thickness (Figure 2), therefore, seems strongly related to sceD upregulation, which is further supported by the repeal of the mutant’s increased CW thickness as well as the upregulation of sceD when grown in 5% CO2 (Table S7). Like vancomycin-resistant S. aureus VC40 mutant, the ΔmpsABC mutant displayed increased resistance to lysostaphin and detergent Triton X-100 induced lysis. Both phenotypes suggested structural alterations in PG. An explanation could be that the mutant’s PG exhibits reduced cross-linking, thereby increasing its susceptibility to cleavage by cellosyl. During standard growth conditions in rich medium, S. aureus PG is highly cross-linked (80-90%) [29,30,31]. To investigate the extent of crosslinking, we analyzed the ratio of muropeptide abundance generated after mutanolysin digestion which indicates potential alterations in crosslinking. However, there were no significant differences in the distribution of mono- to hexameric muropeptides between the mutant and parent PG under ambient air or 5% CO2 (Table S2). The double digestion of mutant PG with lysostaphin and cellosyl revealed new dimeric muropeptides (peaks 10, 11, and 12) and alanine incorporation into the Gly5 – bridge (peaks 7, 8, 9 and 12) (Figure 4, Table S3). The detection of dimeric PG fragments suggests that they are not (or poorly) digested by lysostaphin, consistent with earlier observations of staphylococci with L-amino acids in peptide bridges [32,33]. In staphylococci, the pentaglycine bridge is sequentially built by FemABX using glycyl-charged tRNA [34]. Presumably, the mpsABC mutant incorporates alanine into the interpeptide bridges in a similar mechanism to the recently reported, FmhA-FmhC-mediated incorporation of L-serine in different S. aureus strains [35]. We explicitly looked for upregulation of genes in our RNAseq analysis encoding the Fem-like factors FmhA and FmhC, which incorporate Gly-Ser dipeptides into the pentaglycine bridges [35]; however, we observed no change in expression. The lysostaphin producer Staphylococcus simulans biovar staphylolyticus incorporates serine in the Gly5-bridge by the ‘lysostaphin immunity factor’ (Lif) to become resistant to lysostaphin [36,37]. FmhA and FmhC are homologs of Lif [38]. Therefore, we hypothesize that the incorporation of alanine into the interpeptide bridge causes lysostaphin resistance. The mechanisms of L-alanine incorporation are not clear. Since we could not observe a differential gene regulation of fmhA and fmhC, a possible explanation could be that the FmhA/C substrate alanyl-tRNA is more abundant than Ser- or Gly-tRNA. Another CW structural change was the altered glycosylation of WTA in the mutant. In S. aureus, approximately 60% of the CW is composed of WTA [39], a polymer covalently linked to MurNAc via phosphodiester bonds. WTA consists of chains of ribitol-phosphate units, esterified with D-alanine and glycosylated with N-acetylglucosamine (GlcNAc). This esterification process and the glycosylation pattern is crucial in regulating the activity of autolytic enzymes, it contributes to the resistance to cationic antimicrobial compounds and impacts the recognition by human immune receptors [19,39,40,41,42]. In the mutant tarM, which encodes the WTA α-1,4-glycosyltransferase [18], was significantly (>40-fold) downregulated. Indeed, we found that the α-1,4-glycosylation of WTA was significantly decreased in the mutant (Figure 5a) while the TarS mediated ß-1,4-glycosylation was increased compared to the parental strain (Figure 5b). In S. aureus tarS is highly conserved and most likely constantly expressed [39], and it was therefore not surprising that we did not observe much up- or downregulation of tarS in the mutant. We therefore assume that a depletion of the α-1,4-glycosylation in WTA is compensated by an increased ß-1,4-glycosylation by TarS. The increase of ß-1,4-glycosylation in VC40 has been described to be correlated with higher vancomycin and detergent resistance [24,43]. We therefore assume that the mutant’s increased WTA ß-glycosylation is responsible for its enhanced resistance to Triton X-100 induced autolysis. Apparently, the mutant’s CW structural response to CO2/bicarbonate depletion stress is comparable to vancomycin induced resistance CW adaptations in VC40. RNA-seq revealed the significant upregulation of ATP-dependent chaperone gene clpB. ClpB is a disaggregase and a key member of a multi-chaperone system that efficiently inhibits and reverses protein aggregation [44]. We hypothesize that in the mpsABC mutant, translation is affected due to substrate limitation, leading to incomplete and misfolded proteins and the expression of stress response factors such as ClpB. Consistent with this hypothesis, we identified a high number of other, differently regulated genes whose products are involved in protein turnover. In addition, we observed an induction of prophages since most phage genes were upregulated. Genes that encode secreted proteins, transporters, and toxins were found to be downregulated. The reduced expression of toxins correlated with the previously observed decrease in virulence of the ΔmpsABC mutant [9]. Many of these genes are activated by global regulators such as Agr and SarA, which were found to be downregulated in our study. This is consistent with the hypothesis that the mutant, when experiencing CO2/bicarbonate depletion, adjusts its metabolism to prioritize energy conservation and reduce the expression of genes that are not essential for survival. Therefore, we speculated that the lack of CO2/bicarbonate might be compensated by the overexpression of genes encoding enzymes involved in fatty acid biosynthesis and carboxylases. Contrary to our expectations, the mutant is unable to partially compensate the CO2/bicarbonate deficiency by overexpression of carboxylases. However, there was a downregulation of the pckA, PEP carboxylase, by 5-fold after 4 h and 100-fold after 8 h (Table 1). PckA was first described in S. aureus by Scovill et al. 1996 and they found that a mutant grew poorly in the absence of glucose [45]. PckA lies in the junction between glycolysis and TCA cycle. Reduced expression of pckA has far-reaching consequences: the TCA cycle is not sufficiently replenished with oxaloacetate which leads to its halt, the respiratory chain comes to a standstill and, consequently the membrane potential collapses. This aligns with our first phenotypic characterization of ΔmpsABC [10] in which we found decreased respiration and membrane potential.
4. Materials and Methods
4.1. Bacterial strains and Growth Conditions
4.2. Growth Studies
Overnight cultures were inoculated at a starting A578 of 0.1 in TSB with a flask-to-medium ratio of 1:10. Main cultures were grown at 37°C with continuous shaking under ambient air or 5% CO2 conditions. Aliquots were collected at 0 h, and subsequently every hour for up to 8 h, as well as after 24 h for A578 determination.
4.3. RNA Isolation, Library Construction, Sequencing, and Analysis
To obtain RNA, precultures of S. aureus JE2 and JE2ΔmpsABC were cultivated overnight under 5% CO2 conditions. The main cultures of each strain were inoculated at a starting A578 of 0.1 and grown under ambient air conditions. Aliquots of A578 2.0 were collected at 4 and 8 h. Qiagen RNAprotect (Qiagen, Germany) was added to the aliquots and washed once with Tris-EDTA buffer (TE, 10 mM Tris-HCL, 1 mM EDTA pH 8) followed by a 30 min digestion with lysostaphin (Sigma-Aldrich), mutanolysin (Sigma-Aldrich) and achromopeptidase (Sigma-Aldrich) at 37°C. Total RNA was extracted using RNeasy Mini kit (Qiagen, Germany) according to the manufacturer’s instructions, followed by an on-column DNase digestion (Qiagen, Germany). RNA quantification was performed using a Qubit fluorimeter (Thermo Fisher Scientific), and RNA integrity assessed using a BioAnalyzer (Agilent Technologies). The Stranded Total RNA Ribo-Zero Plus kit from Illumina was used for library preparation, with 500 ng of total RNA as input. Library molarity and quality were assessed using the Qubit and Tapestation (DNA High sensitivity chip, Agilent Technologies). Libraries were sequenced on a NovaSeq 6000 Illumina sequencer using oriented single-reads of 50 nt, yielding a minimum of 50 million mapped reads per sample. Raw reads were trimmed using Trimmomatic v.0.36 with the following options: SLIDINGWINDOW:10:30 MINLEN:25. The obtained clean-reads and the reference Staphylococcus aureus strain JE2 genome (GenBank accession: CP020619.1) were aligned using BWA v.0.7.17-r1188 with the BWA-MEM algorithm and the following options: -M –p. The samtools suite v.1.8 was used to obtain the final sorted bam files used to generate the reads count table. The reads count table was transformed into a RPKM table. Statistical analysis was performed using DSeq2 (https://yanli.shinyapps.io/DEApp/) with the Single-factor Experiment option. The lists of differentially regulated genes were exported and annotated as text files. Raw expression data for these samples were submitted to the European Nucleotide Archive (ENA) database under project accession number PRJEB65327. For qRT-PCR validation of selected genes bacteria were grown and total RNA was isolated as described above. Total RNA was treated with Turbo DNAse (Invitrogen, USA) and reverse-transcribed using random hexamers to generate a cDNA library with SuperScript IV (Invitrogen, USA). Following RNA-seq analyses, five genes were selected from the overall list of differentially expressed genes for qRT-PCR validation, representing the highest and lowest expressed genes from various categories including i) transporters (esxA), ii) CW-bound protein (sasD), iii) virulence (hla), iv) and CW hydrolase/modification (lytM and sceD). The primers used to amplify the selected genes to determine their relative expression are listed in Table S4. qRT-PCR was performed using StepOne thermocycler (Thermo Fisher, USA) and analyzed with StepOne Software. The gene gyrB was used as a housekeeping gene to normalize transcript quantification. Relative gene expression was calculated using the 2−ΔΔCT method from two independent biological replicates for each strain/condition and was performed in triplicate on at least two separate runs. Relative gene expression for the selected genes from strains grown under 5% CO2 conditions was normalized to its corresponding gene grown under ambient air, with the latter set equal to 1.0. Statistical significance of the relative expression profiles was determined by Student’s t-test, and a p value of < 0.05 is considered ‘significant’.
4.4. Triton X-100-Induced Autolysis Assay
The autolysis assay was performed as previously described [46]. Overnight cultures of S. aureus JE2 and JE2ΔmpsABC were adjusted to A578 of 0.05 and 0.1, respectively, and grown in BM (1% soy peptone, 0.5% yeast extract, 0.5% NaCl, 0.1% glucose, and 0.1% K2HPO4, pH 7.2) until mid-exponential phase to A578 0.5. Cells were centrifuged to remove the growth medium and then washed twice with phosphate buffered saline (PBS) (pH 7.2), followed by a final wash with ice-cold sterile MilliQ water. The cells were resuspended in PBS or PBS containing 0.05% Triton X-100, respectively, to an A578 of 1. Each cell suspension (500 μL) was added to a 48 well microplate (Greiner Bio-One, Germany) and incubated at 37°C with continuous shaking on a multiplate reader (Varioskan Lux, Thermo Scientific). Autolysis was monitored as a decrease in A578 after every 30 min for 6 h. PBS and PBS containing 0.05% Triton X-100 served as negative controls.
4.5. Lysostaphin Induced Lysis Assay
To evaluate the resistance of S. aureus JE2 parent and JE2ΔmpsABC towards the CW hydrolyzing enzyme lysostaphin, overnight cultures were adjusted to an A578 of 1. The cell suspensions were centrifuged for 5 min at 5000 × g to collect the cell pellets. The pellet was resuspended in 100 µL TSB containing lysostaphin at concentrations of 50, and 500 µg/mL and incubated at 37°C for 30 min. Subsequently, the gDNA was purified using Quick-DNATM Microprep Kit (Zymo Research, Germany). The gDNA concentration was measured using Nanodrop (NanoPhotometer® NP80, Implen GmbH, Germany).
4.6. Transmission Electron Microscopy
Overnight cultures of S. aureus JE2 and JE2ΔmpsABC (grown in CO2) were inoculated in TSB at a starting A578 of 0.05 and 0.1, respectively. All cultures were grown under their respective conditions with shaking at 37°C until they reached an A578 of 0.5. 1 mL aliquots of each culture were collected and pelleted by centrifugation at 1000 × g for 10 min before being washed twice with 1 mL of PBS (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.3). The cells were pre-fixed with fixing solution (4% formaldehyde, 2.5% glutaraldehyde in 0.1 M PO4 at pH 7.4) for 90 min at room temperature. Cells were high-pressure frozen (HPF Compact 03, Engineering Office M. Wohlwend GmbH) in capillaries and freeze substituted (AFS2, Leica Microsystems) with 2% OsO4 and 0.4% uranyl acetate in acetone as a substitution medium and embedded in Epon. Ultrathin sections were stained with uranyl acetate and lead citrate and analyzed with a Tecnai Spirit transmission electron microscope (Thermo Fisher Scientific) operated at 120 kV.
4.7. Muropeptide Analysis
PG was prepared as previously described [47,48] and incubated with 15 µg cellosyl in 20 mM sodium phosphate pH 4.8 for 16 h at 37°C with stirring. An additional 15 µg of cellosyl was added and the incubation was continued overnight. Enzymatic digestion was stopped by boiling the samples for 5 min at 100°C in a dry heat block and soluble material was collected after centrifugation at 13.000 × g for 10 min. The supernatant was reduced with sodium borohydride and the resulting muropeptides were separated by RP-HPLC using the conditions and gradients described previously [48]. PG was prepared as described previously [47,48] and incubated with lysostaphin (300 µg) in 20 mM NaPi, pH 6.0, for 24 h at 37°C with stirring. Subsequently, cellosyl (Höchst AG, Frankfurt, Germany) (15 µg) was added and the sample was incubated for 16 h at 37°C. An additional 15 µg of cellosyl was added and incubation was continued for 16 h. Enzymatic digestion was stopped by boiling the samples for 5 min at 100°C in a dry heat block and the soluble material was collected after centrifugation at 13.000 × g for 10 min. The supernatant was reduced with sodium borohydride and the resulting monomeric muropeptides were separated by RP-HPLC using a Prontosil (Bischoff, Leonberg, Germany) column (3 µm, particle size, 250 × 4.6 mm, 120 Å pore size) maintained at 52°C. A 140-minute linear gradient (0-100%) of 10 mM sodium phosphate pH 6.0 with 0.00065% NaN3 to 10 mM sodium phosphate pH 6.0 and 30% MeOH at a flow rate of 0.5 mL/min was used for separation of muropeptides [49].
4.8. Mass Spectrometry Analysis of Monomeric Muropeptides
For LC-MS/MS analysis [50], the collected fractions were reconstituted in 20 µL of 0.2% formic acid (aq.). Typically, 10 µL was injected onto a microbore RP-HPLC column (ACE 3 C18, 1.0 × 150 mm) flowing at 50 µL min-1 in a 1100 HPLC system (Agilent, UK) at 35°C, with the first 7 min of eluate diverted to waste due to high salt content. Buffer A was composed of water containing 0.1% (v/v) formic acid and buffer B was acetonitrile containing 0.1% (v/v) formic acid. The following elution gradient was used: starting at 0% buffer B, increasing to 4% B at 10 min, then on to 5% B at 30 min, rising to 10% B at 53 min, the gradient was ramped to 50% B at 58 min, then on to 85% B at 63 min, followed by a 2 min hold at 85% B, and finally 15 min re-equilibration at 0% B. The total run time was 80 min. The HPLC column eluate was directed to the mass spectrometer (LTQ Ion Trap MS, Thermo) via an IonMaxTM electrospray ion source (Thermo). The settings for the ion source were spray voltage and capillary temperature values of 4,200 V and 200°C, respectively, together with a sheath gas flow of 16 (arb) and a sweep gas flow of 1.0 (arb). MS data were acquired in positive ion mode over the range of 200 - 2,000 m/z in Triple Scan mode. The precursor scan (Enhanced scan rate) was immediately followed by an UltraZoom scan (lower = 3 m/z units, upper = 5 m/z units) and finally MS/MS acquisition was performed using a normal scan rate, with activation Q = 0.25 and activation time = 30 ms (with wide band activation turned on). The minimum signal threshold was set at 500 counts, MS/MS isolation width set at 2 m/z, preferred charge state range was set at +1 to +3 and undetermined charge states were excluded. The resulting mass spectral data were analyzed using QualBrowserTM software (Thermo).
4.9. Flow Cytometry Analysis with WTA Detecting Antibodies
S. aureus JE2 and its mpsABC deletion mutant were grown overnight at 37°C under ambient air and 5% CO2 condition. Cells were adjusted to an A578 of 0.4 in Tris-HCL buffer (pH 8) and treated with 200 μg/mL proteinase K for 1h at 37°C. Incubation was continued overnight at 4°C and stopped on the next day by inactivation at 95°C for 10 min followed by centrifugation at 5000 x g and buffer exchange with PBS containing 0.1% bovine serum albumin. 25 μL of the diluted bacteria were incubated with 25 μL serial dilutions of mAb (monoclonal antibody) 4461 or mAb 4497 Fab fragments in a 96-well plate for 30 min at 4°C [19,51,52]. The samples were subsequently washed, centrifuged, and incubated with fluorescein isothiocyanate-labeled goat anti-human IgG F(ab′)2 FITC conjugate (2 μg/mL) (Merck, AQ112F) for 20 min at 4°C in the dark. Labeled bacteria were washed, centrifuged, and fixed with 1% paraformaldehyde for 20 min at room temperature in the dark. The bacteria were centrifuged again and resuspended in PBS, and surface-bound IgG Fab was measured by flow cytometry using a BD FACSCalibur. Anti-HIV protein gp120 (b12-IgG) Fab fragment (5 μg/mL) was used as isotype control. The whole bacterial population was gated, and the mean FL-1 fluorescence was analyzed with FlowJo version 10.8.1. The WTA-specific mAb 4461, mAb 4497 and the B12 isotype control were kindly provided by Prof. N. van Sorge (Amsterdam UMC, The Netherlands) and described previously [19].
4.10. Statistical Analyses and Quantification of CW Thickness
All data are presented as sample mean ± SD, unless specified otherwise. All statistical analyses were performed using Student’s t test or two-way ANOVA for comparison between groups using GraphPad Prism 9 software. P values of < 0.05 were considered statistically significant. CW thickness was quantified using ImageJ [53] from a total of 20 cells for each strain under their respective growth conditions.
5. Conclusions
S. aureus serves as an exemplary model for mesophilic bacteria to study the stress response to bicarbonate depletion, as it relies solely on the MpsABC-type bicarbonate transporter for CO₂ concentration. Adequate bicarbonate supply is crucial, evidenced by the inability of a mpsABC mutant to grow in ambient air. This mutant exhibits characteristics of viable but non-culturable (VBNC) cells, making it a valuable model for investigating bicarbonate-depletion stress responses. Observations of the mpsABC mutant revealed differential gene expression and cell wall (CW) modifications, including increased thickness, enhancing resilience to lytic enzymes and detergents. A thickened CW has previously been reported in S. aureus treated with translation-inhibiting antibiotics and more recently in a vancomycin-resistant mutant. This raises the question: what parallels between antibiotic stress and bicarbonate deficiency lead to similar stress responses? Both conditions impair protein synthesis and growth. However, CW synthesis can proceed independently of protein synthesis, allowing CW thickening despite growth arrest. This might explain the similar stress response to antibiotic stress and bicarbonate depletion.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Growth in 5% CO2; Figure S2: Additional TEM images; Figure S3: Analysis of the PG composition from JE2 and JE2ΔmpsABC; Figure S4: LC- MS and LC-MS/MS spectra; Figure S5: qRT-PCR confirmation of selected genes; Figure S6: Comparative transcriptome analysis COG of proteins; Table S1: Concentrations of released genomic DNA (gDNA) upon cell lysis; Table S2: The percentage distribution of mutanolysin digested muropeptides; Table S3: Calculated masses by LC-MS/MS of JE2ΔmpsABC collected muropeptide fragments; Table S4: Oligonucleotides used in qRT-PCR; Table S5: Transcriptomic analysis 4h; Table S6: Transcriptomic analysis 8h; Table S7: Transcriptomic analysis in 5% CO2
Author Contributions
Conceptualization, F.G., E.L and SH.F; methodology, K.H, P.F, W.V, E.L, C.B, J.G and SH.F.; validation, F.G., P.F., K.H, J.G and W.V.; investigation, J.G, J.B, D.V, P.F, E.L, C.B, I.K, SH.F. and B.S; resources, F.G, P.F, W.V, A.P, A.B, J.G and K.H; data curation, P.F.; writing—original draft preparation, F.G. and E.L; writing—review and editing, C.B, W.V, K.H, P.F, J.G and SH.F; supervision, F.G, W.V, P.F, K.H, A.B, J.G and A.P; project administration, F.G; funding acquisition, F.G and W.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) Germany’s Excellence Strategy—EXC 2124—390838134 ‘Controlling Microbes to Fight Infections’ and the Biotechnology and Biological Sciences Research Council (UK) grant BB/W013630/1.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Raw expression data for these samples were submitted to the ENA database under project accession number PRJEB65327.
Acknowledgments
We would like to kindly thank Dr. Libera Lo Presti for her valuable comments and suggestions on scientific writing. We extend our sincere gratitude to Jeremiah Shuster, Dr. Stefan Fischer, and Prof. Dr. Andreas Kappler for their valuable collaboration in conducting Scanning Electron Microscopy. The authors would like to thank Eve-Julie Bonetti (GRL, University of Geneva) for technical assistance with RT-qPCR and RNA purification for RNA-seq analysis, as well as Mylène Docquier (iGe3 platform, University of Geneva) for library preparation and RNA sequencing. Lastly, we would like to thank Prof. N. van Sorge (Amsterdam UMC, The Netherlands) and Dr. A. Robin Temmig (Amsterdam UMC, the Netherlands) for producing and providing us with the WTA-specific and B12 control fragments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fujita Y, Matsuoka H, Hirooka K. Regulation of fatty acid metabolism in bacteria. Mol Microbiol. 2007, 66, 829–839. [Google Scholar] [CrossRef]
- Tong, L. Structure and function of biotin-dependent carboxylases. Cell Mol Life Sci. 2013, 70, 863–891. [Google Scholar] [CrossRef]
- Merlin C, Masters M, McAteer S, Coulson A. Why is carbonic anhydrase essential to Escherichia coli? J Bacteriol. 2003, 185, 6415–6424. [Google Scholar] [CrossRef]
- Langereis JD, Zomer A, Stunnenberg HG, Burghout P, Hermans PWM. Nontypeable Haemophilus influenzae Carbonic Anhydrase Is Important for Environmental and Intracellular Survival. J Bacteriol. 2013, 195, 2737–2746. [Google Scholar] [CrossRef]
- Casey, JR. Why bicarbonate? Biochem Cell Biol. 2006, 84, 930–939. [Google Scholar] [CrossRef]
- Price GD, Badger MR, Woodger FJ, Long BM. Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J Exp Bot. 2008, 59, 1441–1461. [Google Scholar] [CrossRef]
- Price, GD. Inorganic carbon transporters of the cyanobacterial CO2 concentrating mechanism. Photosynth Res. 2011, 109, 47–57. [Google Scholar] [CrossRef]
- Mangiapia M, Usf M, Brown TW, Chaput D, Haller E, Harmer TL, et al. Proteomic and Mutant Analysis of the CO2 Concentrating Mechanism of Hydrothermal Vent Chemolithoautotroph Thiomicrospira crunogena. J Bacteriol. [CrossRef]
- Fan SH, Ebner P, Reichert S, Hertlein T, Zabel S, Lankapalli AK, et al. MpsAB is important for Staphylococcus aureus virulence and growth at atmospheric CO2 levels. Nat Commun. 2019, 10, 3627. [Google Scholar] [CrossRef]
- Mayer S, Steffen W, Steuber J, Götz F. The Staphylococcus aureus NuoL-Like Protein MpsA Contributes to the Generation of Membrane Potential. J Bacteriol. 2015, 197, 794–806. [Google Scholar] [CrossRef]
- Fan SH, Liberini E, Götz F. Staphylococcus aureus Genomes Harbor Only MpsAB-Like Bicarbonate Transporter but Not Carbonic Anhydrase as Dissolved Inorganic Carbon Supply System. Microbiol Spectr. [CrossRef]
- Fan SH, Matsuo M, Huang L, Tribelli PM, Götz F. The MpsAB Bicarbonate Transporter Is Superior to Carbonic Anhydrase in Biofilm-Forming Bacteria with Limited CO2 Diffusion. Microbiol Spectr. [CrossRef]
- Ersoy SC, Manna AC, Proctor RA, Chambers HF, Harrison EM, Bayer AS, et al. The NaHCO(3)-Responsive Phenotype in Methicillin-Resistant Staphylococcus aureus (MRSA) Is Influenced by mecA Genotype. Antimicrob Agents Chemother. 2022, 66, e0025222. [Google Scholar] [CrossRef]
- Fan S-H, Proctor RA, Ersoy SC, Manna AC, Cheung AL, Götz F, et al. Role of the NaHCO(3) Transporter MpsABC in the NaHCO(3). 2013; ß-Lactam-Responsive Phenotype in Methicillin-Resistant Staphylococcus aureus. Microbiology Spectrum. 2023, 11, e00141–23. [Google Scholar] [CrossRef]
- Fey PD, Endres JL, Yajjala VK, Widhelm TJ, Boissy RJ, Bose JL, et al. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. mBio. 2013, 4, e00537–12. [Google Scholar] [CrossRef]
- Maya-Martinez R, Alexander JAN, Otten CF, Ayala I, Vollmer D, Gray J, et al. Recognition of Peptidoglycan Fragments by the Transpeptidase PBP4 From Staphylococcus aureus. Front Microbiol. 2018, 9, 3223. [Google Scholar] [CrossRef]
- Brown S, Xia G, Luhachack LG, Campbell J, Meredith TC, Chen C, et al. Methicillin resistance in Staphylococcus aureus requires glycosylated wall teichoic acids. Proceedings of the National Academy of Sciences. 2012, 109, 18909–18914. [Google Scholar] [CrossRef]
- Xia G, Maier L, Sanchez-Carballo P, Li M, Otto M, Holst O, et al. Glycosylation of wall teichoic acid in Staphylococcus aureus by TarM. J Biol Chem. 2010, 285, 13405–13415. [Google Scholar] [CrossRef]
- Hendriks A, van Dalen R, Ali S, Gerlach D, van der Marel GA, Fuchsberger FF, et al. Impact of Glycan Linkage to Staphylococcus aureus Wall Teichoic Acid on Langerin Recognition and Langerhans Cell Activation. ACS Infect Dis. 2021, 7, 624–635. [Google Scholar] [CrossRef]
- Beveridge, TJ. Ultrastructure, chemistry, and function of the bacterial wall. Int Rev Cytol. 1981, 72, 229–317. [Google Scholar] [CrossRef]
- Giesbrecht P, Kersten T, Maidhof H, Wecke J. Staphylococcal cell wall: morphogenesis and fatal variations in the presence of penicillin. Microbiol Mol Biol Rev. 1998, 62, 1371–1414. [Google Scholar] [CrossRef]
- Hancock R, Park JT. Cell-wall synthesis by Staphylococcus aureus in the presence of chloramphenicol. Nature. 1958, 181, 1050–1052. [Google Scholar] [CrossRef]
- Kim SJ, Chang J, Singh M. Peptidoglycan architecture of Gram-positive bacteria by solid-state NMR. Biochim Biophys Acta. [CrossRef]
- Berscheid A, François P, Strittmatter A, Gottschalk G, Schrenzel J, Sass P, et al. Generation of a vancomycin-intermediate Staphylococcus aureus (VISA) strain by two amino acid exchanges in VraS. J Antimicrob Chemother. 2014, 69, 3190–3198. [Google Scholar] [CrossRef]
- Sass P, Berscheid A, Jansen A, Oedenkoven M, Szekat C, Strittmatter A, et al. Genome sequence of Staphylococcus aureus VC40, a vancomycin- and daptomycin-resistant strain, to study the genetics of development of resistance to currently applied last-resort antibiotics. J Bacteriol. 2012, 194, 2107–2108. [Google Scholar] [CrossRef]
- Stapleton MR, Horsburgh MJ, Hayhurst EJ, Wright L, Jonsson IM, Tarkowski A, et al. Characterization of IsaA and SceD, two putative lytic transglycosylases of Staphylococcus aureus. J Bacteriol. 2007, 189, 7316–7325. [Google Scholar] [CrossRef]
- Buist G, Steen A, Kok J, Kuipers OR. LysM, a widely distributed protein motif for binding to (peptido)glycans. Molecular Microbiology. 2008, 68, 838–847. [Google Scholar] [CrossRef]
- Mesnage S, Dellarole M, Baxter NJ, Rouget JB, Dimitrov JD, Wang N, et al. Molecular basis for bacterial peptidoglycan recognition by LysM domains. Nat Commun. 2014, 5, 4269. [Google Scholar] [CrossRef]
- Gally D, Archibald AR. Cell wall assembly in Staphylococcus aureus: proposed absence of secondary crosslinking reactions. J Gen Microbiol. 1993, 139, 1907–1913. [Google Scholar] [CrossRef]
- Snowden MA, Perkins HR. Peptidoglycan cross-linking in Staphylococcus aureus. An apparent random polymerisation process. Eur J Biochem. 1990, 191, 373–377. [Google Scholar] [CrossRef]
- Vollmer W, Seligman SJ. Architecture of peptidoglycan: more data and more models. Trends Microbiol. 2010, 18, 59–66. [Google Scholar] [CrossRef]
- DeHart HP, Heath HE, Heath LS, LeBlanc PA, Sloan GL. The lysostaphin endopeptidase resistance gene (epr) specifies modification of peptidoglycan cross bridges in Staphylococcus simulans and Staphylococcus aureus. Appl Environ Microbiol. 1995, 61, 1475–1479. [Google Scholar] [CrossRef]
- Schindler CA, Schuhardt VT. LYSOSTAPHIN: A NEW BACTERIOLYTIC AGENT FOR THE STAPHYLOCOCCUS. Proc Natl Acad Sci U S A. 1964, 51, 414–421. [Google Scholar] [CrossRef]
- Monteiro JM, Covas G, Rausch D, Filipe SR, Schneider T, Sahl HG, et al. The pentaglycine bridges of Staphylococcus aureus peptidoglycan are essential for cell integrity. Sci Rep. 2019, 9, 5010. [Google Scholar] [CrossRef]
- Willing S, Dyer E, Schneewind O, Missiakas D. FmhA and FmhC of Staphylococcus aureus incorporate serine residues into peptidoglycan cross-bridges. J Biol Chem. 2020, 295, 13664–13676. [Google Scholar] [CrossRef]
- Strauss A, Thumm G, Götz F. Influence of Lif, the lysostaphin immunity factor, on acceptors of surface proteins and cell wall sorting efficiency in Staphylococcus carnosus. J Bacteriol. 1998, 180, 4960–4962. [Google Scholar] [CrossRef]
- Thumm G, Götz F. Studies on prolysostaphin processing and characterization of the lysostaphin immunity factor (Lif) of Staphylococcus simulans biovar staphylolyticus. Mol Microbiol. 1997, 23, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Rohrer S, Berger-Bachi B. FemABX peptidyl transferases: a link between branched-chain cell wall peptide formation and beta-lactam resistance in gram-positive cocci. Antimicrob Agents Chemother. 2003, 47, 837–846. [Google Scholar] [CrossRef] [PubMed]
- van Dalen R, Peschel A, van Sorge NM. Wall Teichoic Acid in Staphylococcus aureus Host Interaction. Trends in Microbiology. 2020, 28, 985–998. [Google Scholar] [CrossRef]
- Baddiley J, Buchanan JG, Martin RO, Rajbhandary UL. Teichoic acid from the walls of Staphylococcus aureus H. 2. Location of phosphate and alanine residues. Biochem J. 1962, 85, 49–56. [Google Scholar] [CrossRef]
- Fischer, W. Physiology of lipoteichoic acids in bacteria. Adv Microb Physiol. [CrossRef]
- Peschel A, Otto M. Phenol-soluble modulins and staphylococcal infection. Nat Rev Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Hort M, Bertsche U, Nozinovic S, Dietrich A, Schrötter AS, Mildenberger L, et al. The Role of β-Glycosylated Wall Teichoic Acids in the Reduction of Vancomycin Susceptibility in Vancomycin-Intermediate Staphylococcus aureus. Microbiol Spectr. 2021, 9, e0052821. [Google Scholar] [CrossRef]
- Alam A, Broms JE, Kumar R, Sjostedt A. The Role of ClpB in Bacterial Stress Responses and Virulence. Front Mol Biosci. 6689. [CrossRef]
- Scovill WH, Schreier HJ, Bayles KW. Identification and characterization of the pckA gene from Staphylococcus aureus. J Bacteriol. 1996, 178, 3362–3364. [Google Scholar] [CrossRef]
- Schlag M, Biswas R, Krismer B, Köhler T, Zoll S, Yu W, et al. Role of staphylococcal wall teichoic acid in targeting the major autolysin Atl. Mol Microbiol. 2010, 75, 864–873. [Google Scholar] [CrossRef]
- 10.1111/j.1365-2958.2009.07007.x.
- Bera A, Herbert S, Jakob A, Vollmer W, Götz F. Why are pathogenic staphylococci so lysozyme resistant? The peptidoglycan O-acetyltransferase OatA is the major determinant for lysozyme resistance of Staphylococcus aureus. Mol Microbiol. 2005, 55, 778–787. [Google Scholar] [CrossRef] [PubMed]
- de Jonge BL, Chang YS, Gage D, Tomasz A. Peptidoglycan composition of a highly methicillin-resistant Staphylococcus aureus strain. The role of penicillin binding protein 2A. J Biol Chem. 1992, 267, 11248–11254. [Google Scholar] [CrossRef]
- Figueiredo TA, Sobral RG, Ludovice AM, Almeida JM, Bui NK, Vollmer W, et al. Identification of genetic determinants and enzymes involved with the amidation of glutamic acid residues in the peptidoglycan of Staphylococcus aureus. PLoS Pathog. 2012, 8, e1002508. [Google Scholar] [CrossRef]
- Bui NK, Gray J, Schwarz H, Schumann P, Blanot D, Vollmer W. The peptidoglycan sacculus of Myxococcus xanthus has unusual structural features and is degraded during glycerol-induced myxospore development. J Bacteriol. 2009, 191, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Lehar SM, Pillow T, Xu M, Staben L, Kajihara KK, Vandlen R, et al. Novel antibody–antibiotic conjugate eliminates intracellular S. aureus. Nature. 2015, 527, 323–328. [CrossRef]
- van Dalen R, Molendijk MM, Ali S, van Kessel KPM, Aerts P, van Strijp JAG, et al. Do not discardsStaphylococcus aureus WTA as a vaccine antigen. Nature. 2019, 572, E1–E2. [Google Scholar] [CrossRef]
- Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012, 9, 676–682. [Google Scholar] [CrossRef]
Figure 1.
Deletion of mpsABC causes severe growth delay under ambient air. (a) Illustration of the construction of ΔmpsABC deletion mutant in the parent strain S. aureus JE2 (locus tag prefix: B7H15). For homologous recombination using allelic exchange, plasmid pBASE6-ΔmpsABC containing approximately 2 kb upstream and downstream DNA sequences of mpsABC was used. (b) Growth rates of ΔmpsABC were significantly lower than those of the parent JE2 strain (**p < 0.01 as determined by unpaired two-sided t-test). Arrows indicate the time points at 4 and 8 h when the samples were collected for transcriptomic analysis. Each time point in the graph represents the mean ± standard deviation (SD) from three independent biological replicates.
Figure 1.
Deletion of mpsABC causes severe growth delay under ambient air. (a) Illustration of the construction of ΔmpsABC deletion mutant in the parent strain S. aureus JE2 (locus tag prefix: B7H15). For homologous recombination using allelic exchange, plasmid pBASE6-ΔmpsABC containing approximately 2 kb upstream and downstream DNA sequences of mpsABC was used. (b) Growth rates of ΔmpsABC were significantly lower than those of the parent JE2 strain (**p < 0.01 as determined by unpaired two-sided t-test). Arrows indicate the time points at 4 and 8 h when the samples were collected for transcriptomic analysis. Each time point in the graph represents the mean ± standard deviation (SD) from three independent biological replicates.
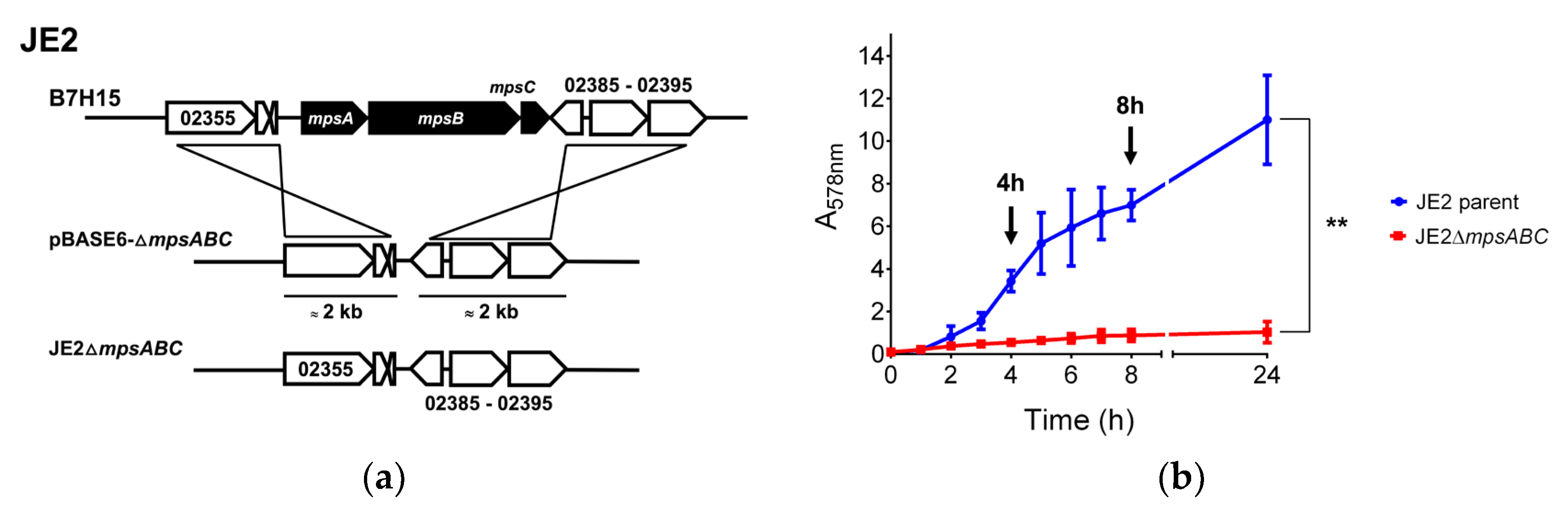
Figure 2.
Cell wall (CW) of the JE2ΔmpsABC grown in ambient air is thicker than that of the parent strain. Transmission electron microscopy (TEM) images of JE2 parent and JE2ΔmpsABC. Cells were grown under ambient air and 5% CO2 at a starting A578 of 0.1, harvested at A578 0.5 and washed with PBS pH 7.3 before fixation with 4% formaldehyde, 2.5% glutaraldehyde in 0.1 M phosphate buffer pH 7.4. The CW of JE2ΔmpsABC showed increased thickness. Scale bar is 200 nm.
Figure 2.
Cell wall (CW) of the JE2ΔmpsABC grown in ambient air is thicker than that of the parent strain. Transmission electron microscopy (TEM) images of JE2 parent and JE2ΔmpsABC. Cells were grown under ambient air and 5% CO2 at a starting A578 of 0.1, harvested at A578 0.5 and washed with PBS pH 7.3 before fixation with 4% formaldehyde, 2.5% glutaraldehyde in 0.1 M phosphate buffer pH 7.4. The CW of JE2ΔmpsABC showed increased thickness. Scale bar is 200 nm.
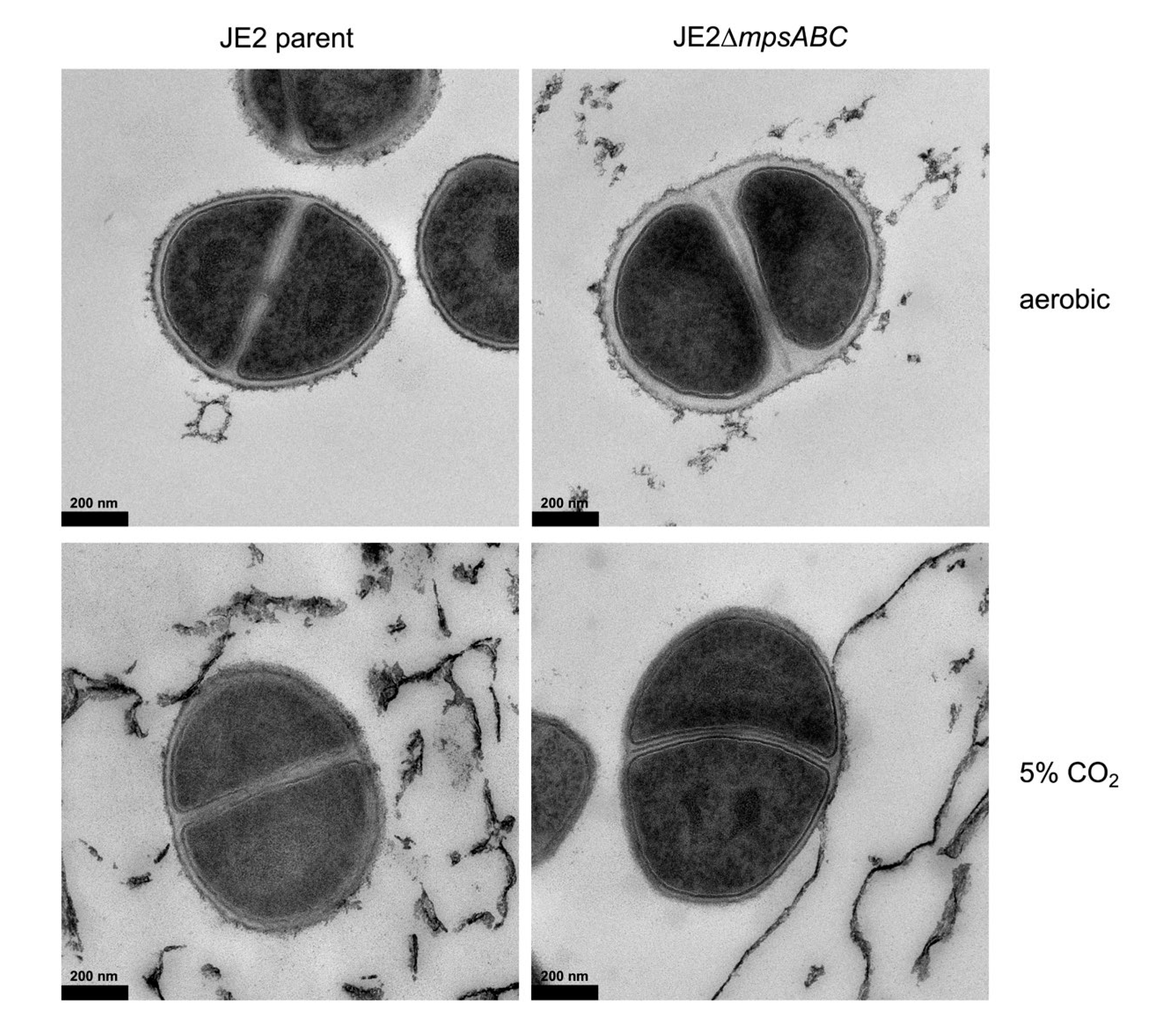
Figure 3.
JE2ΔmpsABC cells are more resistant to lysis. (a) Concentration of genomic DNA (gDNA) released upon cell lysis from JE2 parent and its ΔmpsABC grown under ambient air and 5% CO2 following treatment with lysostaphin (Lss). Genomic DNA was isolated using the commercial Quick- DNATM Microprep Kit (ZYMO Research, Germany) according to the manufacturer’s protocol. Significantly less gDNA was isolated from the mutant than from the parent strain. Each bar in the chart shows the mean ± standard deviations (SD) from three independent biological replicates. Significance was calculated by two-way ANOVA with (**p < 0.01). The concentration of released gDNA upon cell lysis is listed in Table S1. (b) Triton X-100 induced autolysis assay. Triton X-100 (0.05% in PBS) was added at 0 h to mid-exponential phase cells. Autolysis was monitored by measuring the decrease in absorption (A578) every 30 min for 6 h. (**p < 0.01 as determined by unpaired two-sided t-test). Triton X-100 alone served as negative control. Each value in the graph represents the mean ± standard deviations (SD) from three independent biological replicates.
Figure 3.
JE2ΔmpsABC cells are more resistant to lysis. (a) Concentration of genomic DNA (gDNA) released upon cell lysis from JE2 parent and its ΔmpsABC grown under ambient air and 5% CO2 following treatment with lysostaphin (Lss). Genomic DNA was isolated using the commercial Quick- DNATM Microprep Kit (ZYMO Research, Germany) according to the manufacturer’s protocol. Significantly less gDNA was isolated from the mutant than from the parent strain. Each bar in the chart shows the mean ± standard deviations (SD) from three independent biological replicates. Significance was calculated by two-way ANOVA with (**p < 0.01). The concentration of released gDNA upon cell lysis is listed in Table S1. (b) Triton X-100 induced autolysis assay. Triton X-100 (0.05% in PBS) was added at 0 h to mid-exponential phase cells. Autolysis was monitored by measuring the decrease in absorption (A578) every 30 min for 6 h. (**p < 0.01 as determined by unpaired two-sided t-test). Triton X-100 alone served as negative control. Each value in the graph represents the mean ± standard deviations (SD) from three independent biological replicates.
| Cell wall thickness in nm (mean ± SD) | ||
| Strain | Ambient air | 5% CO2 |
| JE2 parent | 15.37 ± 2.64 | 12.05 ± 2.44 |
| JE2ΔmpsABC | 33.43 ± 7.71 | 15.00 ± 2.78 |
Quantification of the CW thickness (nm) was determined using the program ImageJ (Fiji software). Each value represents the mean ± standard deviations (SD) from a total of 20 cells for each strain (JE2 parent and ΔmpsABC) in each condition (ambient air and 5% CO2).
Figure 4.
Muropeptides identified from lysostaphin-cellosyl PG digestion of JE2ΔmpsABC indicate the incorporation of L-alanine into the pentaglycine bridge. (a) Purified PG was digested with lysostaphin and cellosyl and the resulting profile of the JE2ΔmpsABC strain revealed distinct peaks at ~ 75-80 min retention time, which were absent in the PG digestion profile of the parent strain. (b) Collected peaks for lysostaphin-cellosyl digested PG of JE2ΔmpsABC via LC-MS and LC-MS/MS analysis show incorporation of L-alanine into the pentaglycine bridge in the red highlighted peaks. The calculated masses are summarized in Table S3.
Figure 4.
Muropeptides identified from lysostaphin-cellosyl PG digestion of JE2ΔmpsABC indicate the incorporation of L-alanine into the pentaglycine bridge. (a) Purified PG was digested with lysostaphin and cellosyl and the resulting profile of the JE2ΔmpsABC strain revealed distinct peaks at ~ 75-80 min retention time, which were absent in the PG digestion profile of the parent strain. (b) Collected peaks for lysostaphin-cellosyl digested PG of JE2ΔmpsABC via LC-MS and LC-MS/MS analysis show incorporation of L-alanine into the pentaglycine bridge in the red highlighted peaks. The calculated masses are summarized in Table S3.
| Cell wall thickness in nm (mean ± SD) | ||
| Strain | Ambient air | 5% CO2 |
| JE2 parent | 15.37 ± 2.64 | 12.05 ± 2.44 |
| JE2ΔmpsABC | 33.43 ± 7.71 | 15.00 ± 2.78 |
Figure 5.
JE2ΔmpsABC has less α- and more ß-glycosylated GlcNAc WTA. Flow cytometry analysis of GlcNAc WTA (N-acetylglucosamine wall teichoic acid). Cells were grown in ambient air (A) or 5% CO2 (CO2) overnight, adjusted to A578 0.4 and treated with 200 µg/mL proteinase K. Isotype control was anti-HIV protein gp120 (b12-IgG) Fab fragment (5 μg/mL). Secondary antibody was fluorescein isothiocyanate-labeled goat anti-human IgG F(ab′)2 FITC conjugate (2 μg/mL). (a) mAb (monoclonal antibody) 4461 Fab fragment against TarM dependent α-GlcNAc WTA. (b) mAb 4497 Fab fragment against TarS dependent ß-GlcNAc WTA. MFI: mean fluorescence intensity. Each value in the graph shows the mean ± standard error of the mean (SEM) from at least three independent biological replicates. Significance was calculated by two-way ANOVA with (****p < 0.05).
Figure 5.
JE2ΔmpsABC has less α- and more ß-glycosylated GlcNAc WTA. Flow cytometry analysis of GlcNAc WTA (N-acetylglucosamine wall teichoic acid). Cells were grown in ambient air (A) or 5% CO2 (CO2) overnight, adjusted to A578 0.4 and treated with 200 µg/mL proteinase K. Isotype control was anti-HIV protein gp120 (b12-IgG) Fab fragment (5 μg/mL). Secondary antibody was fluorescein isothiocyanate-labeled goat anti-human IgG F(ab′)2 FITC conjugate (2 μg/mL). (a) mAb (monoclonal antibody) 4461 Fab fragment against TarM dependent α-GlcNAc WTA. (b) mAb 4497 Fab fragment against TarS dependent ß-GlcNAc WTA. MFI: mean fluorescence intensity. Each value in the graph shows the mean ± standard error of the mean (SEM) from at least three independent biological replicates. Significance was calculated by two-way ANOVA with (****p < 0.05).
| Protein Number | Function | Fold change Mutant vs. Parent |
||
|---|---|---|---|---|
| 4h | 8h | |||
| Cell wall (CW) lytic enzymes | ||||
| B7H15_11645 | SceD, lytic transglycosylase | 592.3 | 517.1 | |
| B7H15_14290 | IsaA, lytic transglycosylase | 4.3 | 2.1 | |
| B7H15_12775 | SsaA, secretory antigen | 20 | 5.2 | |
| B7H15_10875 | Amidase, 251 aa, prophage-encoded | 16.8 | 9.9 | |
| B7H15_01500 | LytM, glycine-glycine endopeptidase | 14.8 | 8.4 | |
| B7H15_03720 | LysM, peptidoglycan-binding domain-containing protein, probably autolysin | 8 | 2 | |
| Wall teichoic acid (WTA) biosynthesis | ||||
| B7H15_05355 | TarM, poly(ribitol-phosphate) α-N-acetylglucosaminyltransferase | - 43.3 | - 61.5 | |
| CW anchored proteins | ||||
| B7H15_00750 | SasD, cell-wall-anchored protein | - 27.3 | - 3.2 | |
| B7H15_14775 | SasA, serine-rich repeat glycoprotein adhesin | - 23.4 | - 15 | |
| B7H15_13880 | SIRK signal domain/LPXTG anchor domain surface protein | - 22.2 | - 3.3 | |
| B7H15_00135 | AdsA, LPXTG-anchored adenosine synthase | - 11.5 | - 4.1 | |
| B7H15_00620 | Spa, staphylococcal protein A | - 6.4 | - 1.6 | |
| Secreted enzymes | ||||
| B7H15_01535 | Peptidase C51 domain-containing protein | - 64.6 | - 31 | |
| B7H15_14690 | Aureolysin, zinc metalloproteinase | - 44.6 | - 1.2 | |
| B7H15_00545 | Phospholipase C, phosphatidylinositol | - 23.1 | - 1.7 | |
| B7H15_01765 | Lip2(geh), triacylglycerol lipase | - 20.3 | - 200 | |
| Transporters | ||||
| B7H15_01540 | Type VII secretion effector EsxA (and all other type VII secretion protein genes) | - 56.5 | - 102 | |
| B7H15_01675 | Formate/nitrite transporter family protein | - 33.7 | - 1.7 | |
| B7H15_01120 | Peptide ABC transporter substrate-binding protein | - 15.3 | - 4.7 | |
| B7H15_01110 | ABC transporter permease | - 14.9 | - 6 | |
| B7H15_00420 | YdhK family protein, lipoprotein, putative | - 13.5 | - 29 | |
| B7H15_00415 | Copper-translocating P-type ATPase | - 11.2 | - 21.6 | |
| B7H15_03550 | Metal ABC transporter permease | - 11.2 | - 176 | |
| Toxins | ||||
| B7H15_02365 | PSM-α3, phenol-soluble modulin | - 479 | - 6360 | |
| B7H15_11280 | δ-lysin, phenol-soluble modulin | - 336 | - 11215 | |
| B7H15_06120 | PSM-ß, phenol-soluble modulin | - 235 | - 5200 | |
| B7H15_06050 | Hla, α-hemolysin | - 61 | - 2408 | |
| Regulators | ||||
| B7H15_14830 | IcaR, ica operon transcriptional repressor | - 20.7 | - 15.6 | |
| B7H15_11300 | AgrA, accessory gene regulator protein A | - 25 | - 91 | |
| B7H15_11285 | AgrB, accessory gene regulator protein B | - 24 | - 89 | |
| B7H15_11290 | AgrD, accessory gene regulator protein D | - 24 | - 69 | |
| B7H15_11295 | AgrC, accessory gene regulator protein C | - 28 | - 90 | |
| B7H15_03430 | SarA, staphylococcal accessory regulator | - 1.7 | - 3 | |
| B7H15_06535 | CodY, transcriptional repressor | - 1.2 | - 1.6 | |
| Prophage genes | ||||
| B7H15_10915 | Phage tail family protein (and most phage related genes) | 12.5 | nd | |
| B7H15_11045 | PVL, phi PVL orf 51-like protein | 16.5 | 13 | |
| Cross bridge synthesis | ||||
| B7H15_13360 | FmhA, fem-like factor | 1.02 | 1.7 | |
| B7H15_06500 | FmhC, fem-like factor | 0.98 | - 2.3 | |
| Fatty acid biosynthesis | ||||
| B7H15_06405 | FabD, ACP S-malonyltransferase | 1.2 | 1.5 | |
| B7H15_05075 | FabF, ß-ketoacyl-ACP synthase II | - 1.5 | - 1.2 | |
| B7H15_06410 | FabG, 3-oxoacyl-acyl-carrier-protein reductase | - 1.2 | - 1.1 | |
| B7H15_05210 | FabI, enoyl-ACP reducatse | 1.5 | 1.7 | |
| B7H15_11660 | FabZ, 3-hydroxyacyl-ACP dehydratase | 2.2 | 1.6 | |
| B7H15_07130 | PlsY, glycerol-3-phosphate acyltransferase | - 1.2 | - 1.5 | |
| B7H15_06400 | PlsX, phosphate acyltransferase | - 1.5 | 1 | |
| B7H15_06385 | FakA, fatty acid kinase | 1.1 | 1 | |
| B7H15_04195 | FakB1, fatty acid binding protein 1 | - 1.2 | 1 | |
| B7H15_07520 | FakB2, fatty acid binding protein 2 | 1 | 1 | |
| Carboxylating Enzymes | ||||
| B7H15_09750 | PckA, PEP carboxylase | - 5 | - 100 | |
| B7H15_08395 | AccC, acetyl-coA carboxylase | 1.2 | 2 | |
| Cell wall thickness in nm (mean ± SD) | ||
| Strain | Ambient air | 5% CO2 |
| JE2 parent | 15.37 ± 2.64 | 12.05 ± 2.44 |
| JE2ΔmpsABC | 33.43 ± 7.71 | 15.00 ± 2.78 |
(-) indicates a downregulation of gene expression in the mutant compared to the parent. nd, not determine.

Table 1.
CW thickness measurements of S. aureus JE2 parent and JE2ΔmpsABC exposed to ambient air and 5% CO2.
Table 1.
CW thickness measurements of S. aureus JE2 parent and JE2ΔmpsABC exposed to ambient air and 5% CO2.
| Cell wall thickness in nm (mean ± SD) | ||
| Strain | Ambient air | 5% CO2 |
| JE2 parent | 15.37 ± 2.64 | 12.05 ± 2.44 |
| JE2ΔmpsABC | 33.43 ± 7.71 | 15.00 ± 2.78 |
Quantification of the CW thickness (nm) was determined using the program ImageJ (Fiji software). Each value represents the mean ± standard deviations (SD) from a total of 20 cells for each strain (JE2 parent and ΔmpsABC) in each condition (ambient air and 5% CO2).
Table 2.
Selected up-/downregulated genes in ΔmpsABC vs. JE2 parent after 4 and 8 h.
| Protein Number | Function | Fold change Mutant vs. Parent |
||
|---|---|---|---|---|
| 4h | 8h | |||
| Cell wall (CW) lytic enzymes | ||||
| B7H15_11645 | SceD, lytic transglycosylase | 592.3 | 517.1 | |
| B7H15_14290 | IsaA, lytic transglycosylase | 4.3 | 2.1 | |
| B7H15_12775 | SsaA, secretory antigen | 20 | 5.2 | |
| B7H15_10875 | Amidase, 251 aa, prophage-encoded | 16.8 | 9.9 | |
| B7H15_01500 | LytM, glycine-glycine endopeptidase | 14.8 | 8.4 | |
| B7H15_03720 | LysM, peptidoglycan-binding domain-containing protein, probably autolysin | 8 | 2 | |
| Wall teichoic acid (WTA) biosynthesis | ||||
| B7H15_05355 | TarM, poly(ribitol-phosphate) α-N-acetylglucosaminyltransferase | - 43.3 | - 61.5 | |
| CW anchored proteins | ||||
| B7H15_00750 | SasD, cell-wall-anchored protein | - 27.3 | - 3.2 | |
| B7H15_14775 | SasA, serine-rich repeat glycoprotein adhesin | - 23.4 | - 15 | |
| B7H15_13880 | SIRK signal domain/LPXTG anchor domain surface protein | - 22.2 | - 3.3 | |
| B7H15_00135 | AdsA, LPXTG-anchored adenosine synthase | - 11.5 | - 4.1 | |
| B7H15_00620 | Spa, staphylococcal protein A | - 6.4 | - 1.6 | |
| Secreted enzymes | ||||
| B7H15_01535 | Peptidase C51 domain-containing protein | - 64.6 | - 31 | |
| B7H15_14690 | Aureolysin, zinc metalloproteinase | - 44.6 | - 1.2 | |
| B7H15_00545 | Phospholipase C, phosphatidylinositol | - 23.1 | - 1.7 | |
| B7H15_01765 | Lip2(geh), triacylglycerol lipase | - 20.3 | - 200 | |
| Transporters | ||||
| B7H15_01540 | Type VII secretion effector EsxA (and all other type VII secretion protein genes) | - 56.5 | - 102 | |
| B7H15_01675 | Formate/nitrite transporter family protein | - 33.7 | - 1.7 | |
| B7H15_01120 | Peptide ABC transporter substrate-binding protein | - 15.3 | - 4.7 | |
| B7H15_01110 | ABC transporter permease | - 14.9 | - 6 | |
| B7H15_00420 | YdhK family protein, lipoprotein, putative | - 13.5 | - 29 | |
| B7H15_00415 | Copper-translocating P-type ATPase | - 11.2 | - 21.6 | |
| B7H15_03550 | Metal ABC transporter permease | - 11.2 | - 176 | |
| Toxins | ||||
| B7H15_02365 | PSM-α3, phenol-soluble modulin | - 479 | - 6360 | |
| B7H15_11280 | δ-lysin, phenol-soluble modulin | - 336 | - 11215 | |
| B7H15_06120 | PSM-ß, phenol-soluble modulin | - 235 | - 5200 | |
| B7H15_06050 | Hla, α-hemolysin | - 61 | - 2408 | |
| Regulators | ||||
| B7H15_14830 | IcaR, ica operon transcriptional repressor | - 20.7 | - 15.6 | |
| B7H15_11300 | AgrA, accessory gene regulator protein A | - 25 | - 91 | |
| B7H15_11285 | AgrB, accessory gene regulator protein B | - 24 | - 89 | |
| B7H15_11290 | AgrD, accessory gene regulator protein D | - 24 | - 69 | |
| B7H15_11295 | AgrC, accessory gene regulator protein C | - 28 | - 90 | |
| B7H15_03430 | SarA, staphylococcal accessory regulator | - 1.7 | - 3 | |
| B7H15_06535 | CodY, transcriptional repressor | - 1.2 | - 1.6 | |
| Prophage genes | ||||
| B7H15_10915 | Phage tail family protein (and most phage related genes) | 12.5 | nd | |
| B7H15_11045 | PVL, phi PVL orf 51-like protein | 16.5 | 13 | |
| Cross bridge synthesis | ||||
| B7H15_13360 | FmhA, fem-like factor | 1.02 | 1.7 | |
| B7H15_06500 | FmhC, fem-like factor | 0.98 | - 2.3 | |
| Fatty acid biosynthesis | ||||
| B7H15_06405 | FabD, ACP S-malonyltransferase | 1.2 | 1.5 | |
| B7H15_05075 | FabF, ß-ketoacyl-ACP synthase II | - 1.5 | - 1.2 | |
| B7H15_06410 | FabG, 3-oxoacyl-acyl-carrier-protein reductase | - 1.2 | - 1.1 | |
| B7H15_05210 | FabI, enoyl-ACP reducatse | 1.5 | 1.7 | |
| B7H15_11660 | FabZ, 3-hydroxyacyl-ACP dehydratase | 2.2 | 1.6 | |
| B7H15_07130 | PlsY, glycerol-3-phosphate acyltransferase | - 1.2 | - 1.5 | |
| B7H15_06400 | PlsX, phosphate acyltransferase | - 1.5 | 1 | |
| B7H15_06385 | FakA, fatty acid kinase | 1.1 | 1 | |
| B7H15_04195 | FakB1, fatty acid binding protein 1 | - 1.2 | 1 | |
| B7H15_07520 | FakB2, fatty acid binding protein 2 | 1 | 1 | |
| Carboxylating Enzymes | ||||
| B7H15_09750 | PckA, PEP carboxylase | - 5 | - 100 | |
| B7H15_08395 | AccC, acetyl-coA carboxylase | 1.2 | 2 | |
(-) indicates a downregulation of gene expression in the mutant compared to the parent. nd, not determine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



