
Submitted:
29 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
The USH1G protein SANS is a small multifunctional scaffold protein. It is involved in several different cellular processes, such as intracellular transport, in the cytoplasm, or splicing of pre-mRNA, in the cell nucleus. Here, we aimed to gain insight into the regulation of the subcellular localization and the nuclear-cytoplasmic shuttling of SANS and its paralogue ANKS4B, not yet reported in the nucleus. We identified karyopherins mediating the nuclear import and export by screening the nuclear interactome of SANS. Sequence analyses predicted in silico evolutionarily conserved nuclear localization sequences (NLSs) and nuclear export sequences (NESs) in SANS, but only NESs in ANKS4B, which are suitable for karyopherin binding.Quantifying the nuclear-cytoplasmic localization of wild-type SANS and NLS/NES mutants, we experimentally confirmed in silico predicted NLS and NES functioning in the nuclear-cytoplasmic shuttling in situ in cells. The comparison of SANS and its paralogue ANKS4B revealed substantial differences in the interaction with the nuclear splicing protein PRPF31 and in their nuclear localization. Finally, our results on pathogenic USH1G/SANS mutants suggest that the loss of NLSs and NESs and thereby the ability to control nuclear-cytoplasmic shuttling is disease-relevant.
Keywords:
usher syndrome
; USH1G
; SANS
; ANKS4B
; karyopherin
; nuclear-cytoplasmic shuttling
1. Introduction
SANS (scaffold protein containing ankyrin repeats and SAM domain) is encoded by the USH1G gene [1]. Pathogenic variants of USH1G lead to the human Usher syndrome (USH), the most common form of combined hereditary deaf-blindness in humans [2,3]. SANS is a small 52 kDa protein, which consists of 461 amino acids. It is composed of three N-terminal ankyrin repeats (ANK1-3), a central domain (CENT), a sterile alpha motif (SAM), and a PDZ binding motif at the C-terminal end (Figure 1A) [1]. Based on its different functional properties the CENT can be further divided into three subdomains, CENTn1, CENTn2, and CENTc [4]. Context-dependent binding of numerous proteins to its different domains characterizes SANS as a potent scaffold protein [5,6,7,8,9,10,11,12]. In auditory hair cells, SANS plays a crucial role in arranging the four other USH type-1 proteins in the mechanosensitive tip-link complex of the stereocilia [13].
In photoreceptor cells of the retina, SANS functions have been associated with ciliogenesis and the intracellular transport of cargoes to their photosensitive outer segment [6,7,8,9,10,11]. In addition to these cytoplasm-associated processes, SANS has also been found in the nucleus [7,12,14]. In the nucleus, SANS has been recently linked to the control of pre-mRNA splicing and the activation of the spliceosome by interacting with several spliceosomal components [4,12]. However, the question of how the distribution of SANS between the two cellular compartments the cytoplasm and the nucleus is regulated in the cell remained open.
Interestingly, SANS has a paralogue protein in ANKS4B (ankyrin repeat and SAM domain-containing protein 4B) [14]. Both scaffold proteins are composed of the same principal domain structure (Figure 1A); possess high, up to 80% amino acid sequence homology, and share common binding partners, such as the USH1C protein harmonin [5,14,15,16]. ANKS4B and SANS are both located at the tips of microvilli, the brush border microvilli of intestinal enterocytes and the stereocilia, highly modified microvilli of inner ear hair cells, respectively [13,17]. However, while both are found in the cytoplasm, ANKS4B has not yet been reported in the nucleus [14,18]. Moreover, the mechanisms underlying this difference in nuclear-cytoplasmic partitioning have yet to be determined and is one of the questions we address here.
Nuclear transport is a fundamental cellular process that regulates the localization of macromolecules within the nuclear or cytoplasmic compartments. In humans, approximately 60 proteins participate in nuclear transport, including Ran system proteins that ensure directed and rapid transport, components that form nuclear pore complexes (NPCs) in the nuclear membrane, and karyopherins that transport cargoes through the NPCs [19]. For bidirectional shuttling of nuclear cargo proteins, karyopherins, namely importins and exportins are needed [20]. These bind specifically to linear elements in the nuclear cargos, which are called nuclear localization sequences (NLSs) and nuclear export sequences (NESs), respectively.
In the present study, we aimed to shed light on the nuclear-cytoplasmic shuttling of SANS and its paralogue ANKS4B. We screened the SANS interactome to identify importins and exportins and used in silico prediction tools to find conserved NLSs and NESs in SANS protein sequences of several vertebrates. We validated the nuclear-cytoplasmic shuttling potential of wild-type and mutated NLS/NES versions of SANS in HeLa and HEK293T cells by quantifying the nuclear-cytoplasmic localization, which confirmed predicted NLSs and NESs to be important for correct nuclear-cytoplasmic shuttling of SANS. In contrast, strict localization of the SANS paralogue ANKS4B in the cytoplasm is ensured in the absence of any NLS by several NESs. Finally, we provide evidence that the dysregulation and disruption of SANS´ nuclear-cytoplasmic shuttling is relevant for the development of the USH disease in USH1G patients.
2. Materials and Methods
2.1. DNA Constructs

2.2. PCR Primers
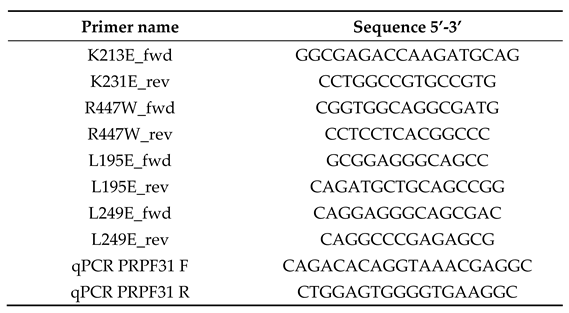
2.3. Antibodies
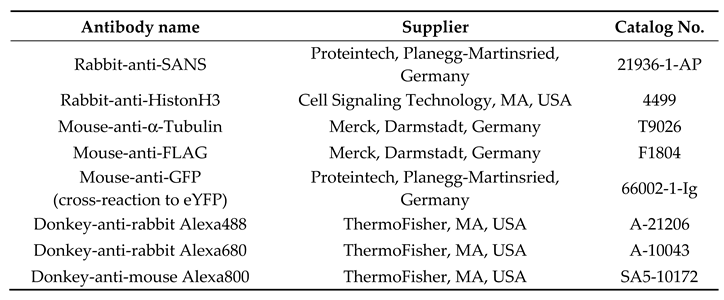
2.4. Cloning
NLS/NES mutants were created by Q5® Site-Directed Mutagenesis Kit according to the manufacturers protocol (NEB, Frankfurt am Main, #E0554S) with pDONR-SANS as a template. pDONR-ANKS4B was kindly provided by Katja Luck [21]. GatewayTM LR reaction into the appropriate destination vector was performed according to the manufacturers protocol (ThermoFisher, Massachusetts, #11791020).
2.5. Cell Lines and Culture
Dulbecco’s modified Eagle’s medium (DMEM) (ThermoFisher, Massachusetts, #31966021) containing 10% heat-inactivated fetal calf serum (FCS) (Cytiva, 489 #SV30160.03, Freiburg) was used to culture HeLa (kindly provided by AG Doorman, IMP) and HEK293T cells (ATCC: CRL-3216).
2.5. Immunocytochemistry
Cells were fixed with 4% paraformaldehyde in PBS for 10 min at RT, washed with PBS, permeabilized with PBST (0.1% Triton-X100 (Roth, Karlsruhe, # 3051.1)) 5 min at RT and blocked with 0.1% ovalbumin, 0.5% fish gelatine in PBS for 1 h at RT. Primary antibodies were incubated overnight at 4 °C, followed by washing with PBS and secondary antibody in a blocking solution containing DAPI (1 mg/mL) (Roth, Karlsruhe, #6335.1) incubations 1 h at RT. After washing, cells were mounted in Mowiol (Roth, Karlsruhe, #0713.2).
2.6. Fluorescence (co-)Localization Observation
Cells were seeded in 6 well plates with a density of 250.000 cells/well. Cells were transfected with LipofectamineTM LTX as described by the manufacturer (ThermoFisher, Massachusetts, #15338100) and incubated for 24 h. If indicated cells were treated with 5 nM Leptomycin-B (Selleck Chemicals GmbH, Munich, #S7580) or 40 µM Importazole (Merck, Darmstadt, #401105) for 4h before fixation. If indicated cells were treated with LipofectamineTM RNAiMAX for 24 h as described by the manufacturer (ThermoFisher, Massachusetts, #13778075). siRNA for PRPF31 (IDT, Leuven, #TriFECTa®Kit DsiRNA Duplex, 5’-AGCUAUGGGAUAGUAAGAUGUUUGC-3’) was used. Knockdown efficiency was validated by qPCR as described previously [12]. Cells were fixed with 4% paraformaldehyde in PBS for 10 min at RT and counterstained for 10 min in DAPI (1 mg/mL) (Roth, Karlsruhe, #6335.1). After washing, cells were mounted in Mowiol (Roth, Karlsruhe, #0713.2).
2.7. Fluorescence Microscopy
Fixed cells were observed with a Leica TCS SP5 or Zeiss LSM900. Images were analyzed with Fiji [22] (https://imagej.net/software/fiji/downloads). Intensity plot was performed over the indicated region of interest with the Fiji Plugin Plot Profile. Z-projections were performed over the indicated region of interest with the Fiji Plugin Orthogonal Views. Pearson coefficient was performed with the Fiji Plugin Coloc 2.
2.8. In Silico Prediction
The amino acid sequence of the protein of interest was used from uniprot. The following sequences were used: Homo sapiens SANS (Q495M9); Macaca nemestrina SANS (A0A2K6BVI9); Mus musculus SANS (Q80T11); Danio rerio SANS (A0A8M1RQW2); Xenopus laevis SANS (A0A8J0TLE4); H. sapiens ANKS4B (Q8N8V4). NLSs were predicted with a given amino acid sequence by NLStradamus [23]. NESs were predicted with a given amino acid sequence by LocNES [24]. Homology of SANS NLS/NES was aligned with Blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE =Proteins, accessed on 02nd April 2024). Homology of SANS and ANKS4B was calculated from the alignment of Blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE =Proteins, accessed on 01st March 2024).
2.9. CellProfiler Based Nuclear/Cytoplasmic Ratio Quantification
Images were preanalyzed with Fiji and then analyzed automatically with CellProfiler [25]. For analysis, we used the pipeline “Human cells” (https://cellprofiler-examples.s3.amazonaws.com/ExampleHuman.zip) with adjustments available on Github: https://github.com/LabWolfrum/Fritze_et_al_2024_SANS_Nuclear_localization
In short, three tif files (protein of interest, DAPI, Brightfield) were used as input. Primary and secondary structures were identified with CellProfiler and the mean intensity of the eYFP channel of the nucleus and cytoplasm was used for further statistical analysis.
2.10. CellFractionation Assay
Cells were seeded in 6 well plates with a density of 250.000 cells/well. Cells were transfected with LipofectamineTM LTX as described by the manufacturer (ThermoFisher, Massachusetts, #15338100) and incubated for 24h. Cells were harvested and lysed into nuclear and cytoplasmic fractions with the commercial kit NE-PERTM (ThermoFisher, Massachusetts, #78833). Protein amount was measured with a BCA assay, and proteins were eluted with 5x Laemmli buffer, separated by SDS-PAGE followed by total protein measurement (RevertTM 700 Total Protein Staining) (LICOR, Nebraska, 926-11010) and Western blotting. Relative intensity was calculated with Fiji for the GFP (cross-reaction for eYFP) band and normalized to total protein and HistoneH3 band.
2.11. Fluorescence Resonance Energy Transfer (FRET) Acceptor Photobleaching Assay
HeLa cells were seeded in 6 well plates with a density of 250.000 cells/well. Cells were transfected with LipofectamineTM LTX as described by the manufacturer (ThermoFisher, Massachusetts, #15338100). Fixed HeLa cells were analyzed with a Leica TCS SP8 and FRET acceptor photobleaching was performed following the Leica protocol (https://downloads.leica-microsystems.com/TCS%20SP8/Application%20Note/FRET_AB_with%20SP8-AppLetter_EN.pdf). eCFP was used as donor (D) and eYFP as acceptor (A). The acceptor was bleached at 100% laser intensity for 10 repeats. FRET efficiency was calculated via: . To exclude cellular structure the mean of six regions of interest (ROI) in the bleached area was calculated and the FRET efficiency of an unbleached region (≥ 3 µm away from bleached ROI) was subtracted from the mean. Bleach efficiency was calculated via the formula . Only ROIs with > 60% bleach efficiency were used for analysis. Data was normalized to 1 by the positive control eCFP-c-eYFP.
2.12. SANS Nuclear Interactome Analysis
SANS nuclear interactome analysis in HEK293T cells [12] were used as input for the Cytoscape plugin ClueGO according to their gene names based on HGNC. Gene Ontology (GO) term enrichment analysis was performed by ClueGO v2.3.3.
2.13. Statistics
Statistical analysis was performed with R-Studio [26]. The statistical methods are listed in corresponding individual legends. Results are shown from at least 3 separate experiments. Significance was determined as: *p≤0.05, **p≤0.009, ***p. ≤0.0009.
3. Results
3.1. Subcellular Localization of SANS
Recent work indicated that SANS shuttles from the cytoplasm into the nucleus [7,12,14]. We confirmed cytoplasmic and nuclear localization of endogenous SANS in HeLa cells by immunocytochemistry using anti-SANS antibodies (Figure 1B) and in HeLa cells transfected with eYFP-SANS and 3xFLAG-SANS monitoring eYFP fluorescence or indirect anti-FLAG immunofluorescence, respectively (Figure 1C). Quantification of eYFP-SANS fluorescence in cytoplasmic and nuclear compartments by CellProfiler revealed that approximately 30% of SANS was present in the cytoplasm and 70% in the nucleus in HeLa cells (Figure 1D). Quantitative analysis of the compartmental distribution in eYFP-SANS expressing HEK293T cells confirmed that most SANS (77%) was localized in the nucleus (Figure S1A). Next, we biochemically validated the compartmental distribution of SANS by cytoplasmic-nuclear cell fractionations in eYFP-SANS transfected HeLa cells (Figure 1E). Western blot analysis of the nuclear and cytoplasmic fractions showed that approximately 70% of SANS was present in the nucleus confirming the obtained in situ data.
Taken together our experimental data consistently demonstrated that SANS is predominantly localized in the nucleus.
3.2. Identification of Karyopherins in the Nuclear Interactome of SANS
To identify potential karyopherins that bind to SANS, we screened the previously published nuclear interactome of SANS in HEK293T cells [12] for importins and exportins. We identified in the GO-term gene clusters “nuclear import” (GO:0051170) and “nuclear export” (GO:0051168) (Cytoscape plugin ClueGO, https://apps.cytoscape.org/apps/cluego, accessed on 6th of June 2024; Table S1) eight nuclear importins and three exportins including exportin-1/CRM1, the main nuclear protein exporter (Table 1).
Subsequently, we investigated their role by applying commercially available importin and exportin inhibitors to eYFP-SANS transfected HeLa cells followed by confocal microscopy analysis and quantification with CellProfiler [25]. Applying the importin-β inhibitor Importazole (IPZ) to transfected HeLa had no effect on eYFP-SANS subcellular localization (Figure S2A-B). Further, we investigated the role of the exportin CRM1 in SANS nuclear export by treating eYFP-SANS HeLa cells with the CRM1 inhibitor Leptomycin-B (LMB) [31]. Confocal microscopy and CellProfiler-based quantification of the subcellular localization of SANS revealed that applications of LMB to the cells resulted in a significant increase in the nuclear localization of SANS when compared to the untreated control (Figure 2A,B). In contrast, treatments with the LMB solvent DMSO had no effect on the nuclear localization. Combined, the treatment revealed that the core export of SANS is mediated by CRM1.
3.3. In Silico Prediction and Evolutionary Conservation of Nuclear Localization Sequences (NLSs) and Nuclear Export Sequences (NESs) in SANS
For nuclear transport karyopherins bind to nuclear localization sequences (NLSs) and nuclear export sequences (NESs) present in the cargo molecules [20]. To identify NLSs and NESs in the human SANS sequence we used the prediction tools NLStradamus [23] and LocNES [24], respectively (Table 2). In addition, the predicted consensus sequences were scored from 0 to 1.
NLStradamus predicted two NLSs within SANS, located in the CENTn2 (NLS_1213-224) and SAM domains (NLS_2436-447), with scores of 0.733 and 0.679, respectively, both above the NLS threshold of 0.6 [23] (Figure 3A). LocNES identified three NESs within SANS, two in the CENTn2 domain (NES_1181-195 and NES_2235-249) and one in the SAM domain (NES_3406-420), with scores ranging from 0.250 to 0.462, all above the NES threshold of 0.1 [24] (Figure 3A). In addition, we classified the predicted NESs into distinct classes of NES motifs binding to the exportin CRM1 (Table 2).
Sequence alignments of the identified NLSs and NESs motifs of SANS across five diverse vertebrates from human to frog, namely H. sapiens, M. nemestrina, M. musculus, D. rerio, and X. laevis revealed high to medium conservation of the motifs (Figure 3B-F). Notably, D. rerio and X. laevis have two and three Ush1g genes encoding SANS, respectively. However, all sequences of the predicted NLSs and NESs in the multiple genes of the respective species are identical.
Both NLSs are 100% conserved throughout all the three mammals. In NLS_1213-224 of D. rerio and X. laevis we observed exchanges of two positive charged amino acids, namely arginine to lysins. However, in NLS_2436-447 of both species, exchanges between charged and uncharged amino acids were found.
The CRM1-consensus motif of NES_3406-420 is 100% conserved throughout all vertebrate species tested. In contrast, NES_1181-195 was conserved in all vertebrates except for X. laevis. NES_2235-249 was also conserved in all tested species but differed in its starting residue in M. musculus, D. rerio and X. laevis. Taken together, in silico analyses predicted two NLSs and three NES/CRM1 motifs in SANS that differ in their probabilities and evolutionary conservation in vertebrates.
3.4. Analysis of the Nuclear Localization Efficiency of Predicted NLS in SANS in the Cell
Next, we examined the efficiency of the two predicted NLSs of SANS in cells. Using site-specific mutagenesis, we mutated positively charged residues to uncharged or negatively charged residues in the NLSs of human SANS that should prevent karyopherin binding to the mutated NLS (Table 2, Figure 4A). eYFP-tagged wild-type SANS and SANS NLS mutants (SANSK213E/ΔNLS_1213-224, SANSR447W/ΔNLS_2436-447) were transfected into HeLa cells (Figure 4B) and HEK293T cells (Figure S1A) and the subcellular distribution of the SANS variants was then assessed by confocal microscopy. Confocal images, their z-projections, and fluorescence intensity plots of eYFP-tagged SANSK213E and SANSR447W indicated an increase of SANS in the cytoplasm of HeLa and HEK293T cells. CellProfiler-based quantification of the SANS localization showed a significant decrease in nuclear localization of SANSK213E (ΔNLS_1213-224) compared to wild-type SANS in both HeLa and HEK293T cells (Figure 4C, Figure S1B). Analysis of SANSR447W (ΔNLS_2436-447) showed a decrease in its nuclear localization that was not significant in HeLa cells but was slightly significant in HEK293T cells (Figure 4C; Figure S1B). In summary, our findings indicate that NLS_1213-224 has a stronger role in the nuclear localization of SANS than NLS_2436-447.
3.5. Analysis of the Nuclear Export Efficiency of Predicted NES in SANS in the Cell
We examined the nuclear export efficiency of the predicted NESs of SANS in the cell. We designed and generated SANS NES mutants by exchanging hydrophobic residue leucine (L) to glutamic acid (E) by site-specific mutagenesis for SANS NES_1181-195 and SANS NES_2235-249 (Table 2, Figure 5A). This resulted in SANSL195E (ΔNES_1181-195) and SANSL249E (ΔNES_2235-249), which were no longer predicted as a NES by LocNES. The two NES mutants were tagged with eYFP, transfected into HeLa and HEK293T cells, and examined by confocal microscopy (Figure 5B, Figure S3A). Unexpectedly, eYFP-SANSL249E formed prominent cytoplasmic aggregates, while eYFP-SANSL195E predominantly localized in small nuclear droplets, as shown by fluorescence intensity plotting and z-projection. CellProfiler-based quantifications showed no significant difference in the nuclear-cytoplasmic ratio of eYFP-SANSL249E (ΔNES_2235-249) compared with eYFP-SANS (Figure 5C, Figure S3B). In contrast, the quantification for eYFP-SANSL195E (ΔNES_1181-195) exhibited a significant increase in the nuclear localization compared to eYFP-SANS. These findings suggest that NES_1181-195 but not NES_2235-249 plays a role in SANS nuclear export.
Unfortunately, we failed to design any synthetic mutant for SANS NES_3406-420 since all genetic modifications introduced by site-specific mutagenesis in this region led to the emergence of a new NES in near proximity. Therefore, we decided to evaluate the role of SANS NES_3406-420 in deletion constructs lacking the domains containing NES_3406-420. The confirmed pathogenic frameshift variants of the human USH1G gene, SANSS278Pfs*71 and SANSS243* (www.LOVD.nl/USH1G, accessed on 04th January 2023), which result in a premature stop at the end of CENTn2 or CENTc domain of SANS, respectively, fulfill this criterium (Figure 6A). Confocal microscopy of eYFP-tagged SANSS278Pfs*71 and SANSS243* expressed in HeLa and HEK293T cells revealed enrichments of the pathogenic variants in the nucleus when compared to wild type SANS, as indicated by fluorescence intensity plotting and z-projection (Figure 6B, Figure S4A). CellProfiler-based quantifications confirmed the significant enrichment of both pathogenic variants in the nucleus compared to eYFP-SANS (Figure 6C, Figure S4B). The retention of SANSS278Pfs*71 and SANSS243* in the nucleus indicates that NES_3406-420 plays an additional pivotal role in SANS nuclear export.
In addition, we investigated the cellular localization of the frameshift mutation SANSV132Gfs*3, which lacks almost the entire CENTn1 and all following C-terminal domains including all NLS and NES (Figure 6A). eYFP-tagged SANSV132Gfs*3 localized predominantly in the nucleus of HeLa and HEK293T cells, as indicated by fluorescence intensity plotting and z-projection (Figure 6B, Figure S4A). CellProfiler-based quantifications confirmed the significant enrichment of SANSV132Gfs*3 in the nucleus compared to eYFP-SANS (Figure 6C, Figure S4B).
Taken together our findings suggest that NES_1181-195 and NES_3406-420 but not NES_2235-249 play a role in SANS nuclear export. Furthermore, present results on SANS pathogenic variants also suggest that alterations in nuclear shuttling may be part of the pathomechanisms leading to USH1G.
3.6. SANS and Its Paralogue ANKS4B Are Localized to Different Subcellular Sites
SANS and its paralogue ANKS4B have nearly the same principal domain structure (Figure 1A), possess a homology of up to 80% in their amino acid sequences, and share common binding partners, such as USH1C/harmonin [5,14,15,16]. Since the differences in cellular function of closely related proteins often depend on the different spatial arrangements in the cell, we tested whether SANS and ANKS4B differ in their cytoplasmic/nuclear localization.
To determine the subcellular localization of ANKS4B we analyzed eYFP-tagged ANKS4B expressing HeLa cells (Figure 7A, B). Confocal microscopy showed that eYFP-ANKS4B was localized prominently in the cytoplasm, also indicated by fluorescence intensity plotting and z-projections. CellProfiler-based quantification demonstrated significantly lower nuclear localization of eYFP-ANKS4B compared to eYFP-SANS (Figure 7B).
NLStradamus predicted no NLS for ANKS4B. However, LocNES predicted six distinct NESs of various CRM1 binding motif classes for ANKS4B spanning its SAM-domain (Table 3) and predicted scores higher than 0.1, the threshold of LocNES [23].
To test whether these motifs are relevant for the subcellular localization we treated eYFP-ANKS4B transfected HeLa cells with the CRM1 inhibitor LMB (Figure 7C). CellProfiler-based quantification demonstrated that the LMB treatment resulted in a significant increase of eYFP-ANKS4B in the nucleus, which was not observed in DMSO treated control cells (Figure 7D).
Taken together, in contrast to SANS, the SANS paralogue ANKS4B is predominantly localized in the cytoplasm guaranteed by nuclear export based on CRM1 exportins.
3.7. SANS and Its Paralog ANKS4B Differentially Interact with PRPF31
Next, we tested whether SANS and ANKS4B also differ in their binary interaction with the splicing molecule PRPF31 by fluorescence resonance energy transfer (FRET) acceptor photobleaching assays (Figure 8A, B) [4,32]. For this we co-transfected HeLa cells with eYFP-SANS or eYFP-ANKS4B, respectively, and eCFP-PRPF31. As controls, we used eYFP alone and harmonin-eCFP, which was previously shown to bind to ANKS4B and SANS [5,17]. FRET signals were normalized to 1 by the positive control eCFP-c-eYFP. The FRET pair eYFP-SANS-eCFP-PRPF31 generated significantly higher normalized FRET signals than the control pair eYFP-eCFP-PRPF31, confirming the binary binding of SANS and PRPF31 (Figure 8C) [4,12]. In contrast, the eYFP-ANKS4B-eCFP-PRPF31 FRET pair showed no increase in normalized FRET efficiencies when compared to the control eYFP-eCFP-PRPF31 pair. The height of the normalized FRET signals of eYFP-ANKS4B-harmonin-eCFP was in the range of the FRET signals of the FRET pair eYFP-SANS-eCFP-PRPF31 and significantly higher than the signals of the eYFP-eCFP-PRPF31 control pair, confirming the interaction of ANKS4B with harmonin as previously reported [17].
Next, we co-expressed eYFP-SANS or eYFP-ANKS4B with mRFP-PRPF31 or harmonin-mCherry, respectively in HeLa cells for subsequent confocal microscopy analyses (Figure 8C). Our analyses revealed that ANKS4B co-localized with harmonin in small droplets in the cytoplasm as observed before [14] but does not colocalize with PRPF31 in the nucleus which was strengthened by the negative Pearson coefficient (R = -0.14). In contrast, SANS co-localized with both harmonin and PRPF31, but in different compartments, namely in the cytoplasm and in the nucleus, respectively. Both co-localizations were supported by high positive Pearson coefficients (R = 0.96 and 0.92, respectively). The representative fluorescence intensity plots of tagged SANS and its interacting partners further demonstrated that SANS was almost completely found in the different compartments of its interacting partner. This finding indicated that SANS nuclear-cytoplasmic shuttling is additionally controlled by its interacting partners. However, further analyses with siRNA-based knockdown of PRPF31 showed no alteration of eYFP-SANS in HeLa cells (Figure S5).
In summary, our findings underscore the divergent nuclear localization/export sequences between SANS and ANKS4B paralog, resulting in differential subcellular localization patterns and cellular functions.
4. Discussion
The protein traffic across the nuclear pores is mostly mediated by members of karyopherin-β (or Kap) family commonly known as importins and exportins which specifically recognize NLSs and NESs of the cargo molecules [20]. In the present study, we provide several lines of evidence that the nuclear localization of SANS is regulated by an interplay of importins and exportins. We identified several importins and exportins in the previously described nuclear interactome of SANS [12]. In silico predictions revealed evolutionary conserved NLSs and NESs in the SANS sequences allowing the binding of importins and exportins. We experimentally confirmed the function of in silico predicted NLSs and NESs by quantifying the subcellular localization of SANS mutants in situ.
Multiple NLS and NES have been previously described for nuclear proteins, such as Fanconi anemia group A protein (FANCA) or enzyme 5-lipoxygenase (5-LO), respectively [33,34]. Our data on the two NLS suggests that NLS_1213-224 localized in the central domain of SANS is the major NLS for the nuclear localization of SANS. In comparison to the N-terminal NLS_2436-447, NLS_1213-224 was scored higher in the predictions [23]. In addition, sequence alignments across vertebrates suggests that NLS_2436-447 is less conserved. In NLS_2436-447 charged amino acids are exchanged to uncharged in lower vertebrates probably affecting the interaction with importins. Importin binding is based on interactions with positively charged amino acids in NLSs of nuclear proteins [20] and exchanges to uncharged residues have been shown to alter NLS binding to importins [35]. In our study, we also experimentally induced such modifications by site-specific mutations of the NLSs in SANS. The quantitative data obtained showed that the decrease in nuclear localization of SANS with a mutated NLS_1213-224 was highly significant in both HeLa and HEK293T cells. In contrast, mutated NLS_2436-447 did not lead to a significant decrease in the nuclear localization of SANS in HeLa cells and only to a decrease of lower significance in HEK293T cells. Differences between cell lines in the nuclear-cytoplasmic partitioning of proteins were previously found to be related to differences in the metabolism between the cell lines [36,37,38].
Based on these data, we hypothesize that importins preferentially bind to NLS_1213-224 of SANS for the import into the nucleus, e.g. to accomplish SANS’ nuclear role in pre-mRNA splicing. Interestingly, NLS_1213-224 is present in the central domain of SANS which represents the major binding site for numerous SANS interacting proteins [7,10,12]. Thus, the binding of cytoplasmic proteins such as myomegalin and whirlin to the central domain [7,10] is likely to compete with the binding of importins to the NLS_1213-224. The competition in binding between importins and other cytoplasmic binding partners as a mechanism controlling the nuclear-cytoplasmic shuttling of nuclear proteins has been described previously [39,40]. Accordingly, the binding competition of cytoplasmic interaction partners of SANS and importins to the NLS sites could guarantee its cytoplasmic localization and thus ensure its cytoplasmic functions, e.g. in intracellular transport or ciliogenesis [7,9,11].
In addition, the binding of nuclear proteins may also facilitate nuclear import in a process known as “piggybacking” [41,42]. Since SANS was not completely removed from the nucleus when NLS_1213-224 was mutated, and the overexpression of PRPF31 resulted in almost complete nuclear localization of co-expressed eYFP-SANS, we speculated that PRPF31 may play a piggyback role for SANS during nuclear import. However, since siRNA-mediated silencing of endogenous PRPF31 did not alter the nuclear-cytoplasmic shuttling of SANS we rejected this hypothesis. Alternatively, binding of importins to NLS_2436-447 or binding of the identified importins, found in the SANS interactome, to non-classical NLS, which are not predictable by prediction tools may compensate for an absence of NLS_1213-224.
Inhibition of the nuclear export by LMB suggests CRM1-dependent nuclear export of SANS. The analysis of the three predicted NES of SANS suggests NES_1181-195 of the CENTn2 domain and NES_3406-420 of the SAM domain as the major NES for the nuclear-cytoplasmic shuttling of SANS. All three predicted NESs are above the threshold for qualifying NESs and are highly conserved, except for NES_2235-249, which differs in 3 of 5 vertebrates in the first residue. As expected for potent NESs, the site-specific mutations resulting in a non-functional NES_1181-195 and the deletion variants of SANS lacking the NES_3406-420 led to a highly significant increase of their nuclear localization in HeLa and HEK293T cells. In contrast, mutated NES_2235-249 did not lead to a significant increase in its nuclear localization. Interestingly, NES_1181-195 and NES_3406-420 of SANS belong to different classes of CRM1 motifs that can differ in the binding affinity of CRM1 [43] and thereby, probably also in the nuclear export efficiency.
In comparison to SANS, its paralogue ANSK4B was far less abundant localized in the nucleus, which correlates with the lack of any known nuclear function of ANKS4B [14,17]. At first glance, the absence of ANKS4B might be caused because its sequence does not contain an NLS since none of the predicted tools applied, predicted a NLS for importin binding in ANKS4B, but several NESs suitable for CRM1 exportin binding. Interestingly, after inhibition of CRM1, more than 80% of the ANKS4B was located in the nucleus. This raises the question of how ANKS4B shuttles into the nucleus. In comparison to SANS (~51.5 kDa https://www.uniprot.org/uniprotkb/Q495M9/entry) ANKS4B (~46 kDa https://www.uniprot.org/uniprotkb/Q8N8V4/entry) is smaller, in the range of the size to diffuse passively through the nuclear pore complex commonly reported between 40 to 50 kDa [19]. However, we also found that the much larger eYFP-tagged ANKS4B ~75 kDa, which we monitored in our experiments, localized in the nucleus. The reason for this is possible that this size exclusion for free diffusion through the NPC is not as effective as assumed. Indeed, more recent findings indicate that larger macromolecules of > 100 kDa can also diffuse through the NPC [44]. Alternatively, as known for other proteins and already discussed above for SANS the nuclear import of ANKS4B may also be mediated by non-canonical NLS, which cannot be predicted by prediction tools [20,45]. Although we could not fully elucidate the regulation of ANKS4B import into the nucleus, our results confirm the previously shown predominantly cytoplasmic localization of ANKS4B. However, once ANKS4B enters the nucleus, it is immediately exported back into the cytoplasm mediated by the CRM1 exportin. This mechanism ensures that ANKS4B is available for its primary functions in the cytoplasm.
In our study, we provide evidence that nuclear-cytoplasmic shuttling of SANS and ANSK4B is coordinated by the selective binding of karyopherins to NLSs and NESs and determines their compartment-specific function. In the cell, protein functions and their regulations are commonly fine-tuned by post-translational modifications such as site-specific reversible phosphorylation [38,46,47]. Therefore, it is not surprising that reversible phosphorylation also emerges as an important process for regulating the nuclear availability of proteins [48]. Interestingly, we previously demonstrated that the kinase inhibitor D-ribofuranosylbenzimidazole (DRB) increases the abundance of SANS in the nucleus due to the inhibition of the phosphorylation at S422 (see Fig. S5 in [9]). S422 is a CK2 (casein kinase 2) phosphosite near NES_3406-420. Therefore, it is tempting to speculate that CK2-mediated phosphorylation not only inhibits MAGI2 from binding to the internal PDZ-binding motif (PBM) in SANS-SAM [9] but may also regulate the binding of CRM1 to the NES.
While for the SANS encoding USH1G gene, more than 70 pathogenic variants have been identified thus far leading to deaf-blindness in patients [49] (www.LOVD.nl/USH1G) to our knowledge defects in ANKS4B have not been associated with a disease [18]. We have previously demonstrated that pathogenic variants of USH1G/SANS variants can lead to the disruption of fundamental cellular processes in both the cytoplasm and the nucleus: While USH1G/SANS variants lead to altered ciliogenesis and intracellular transport in the cytoplasm [10,11], they cause defects in pre-mRNA splicing in the nucleus [12]. Here we show that these SANS variants are highly enriched in the nucleus due to the defect in nuclear-cytoplasmic shuttling. Probably the lack of NESs leads to defects in nuclear export.
Although the SANS variants are targeted into the nucleus, they cannot fulfill their functions in the splicing process there, as we have previously demonstrated [12]. Possibly, not all interaction partners that interact with SANS during proper activation of the spliceosome can interact with the mutated, truncated versions of the scaffold protein. Probably more importantly, due to its pathogenic mutations, SANS is no longer available for its functions in the cytoplasm. A combination of defects in cytoplasmic trafficking and ciliogenesis as well as in pre-RNA splicing in the nucleus may underlie the pathology leading to USH1G. This complex scenario may be present in the retinal cells in the eye. In the inner ear, USH1G-associated deafness is more likely caused by defects in the assembly of mechanosensitive tip-link complex in the stereocilia of auditory hair cells [13].
5. Conclusions
USH1G/SANS is a multifunctional scaffold protein that participates in various cellular processes executed in different compartments of the cell, namely in the cytoplasm or nucleus. To fulfill its specific tasks, efficient and controlled nuclear-cytoplasmic shuttling of the SANS protein is mediated by karyopherins binding to NLSs and NESs in the SANS. This is very different from its strict cytoplasmic paralogue ANKS4B, in which several NESs guarantee its localization and function in the cytoplasm. Finally, we provide evidence that the dysregulation and disruption of SANS´ nuclear-cytoplasmic shuttling can be relevant for the development of the USH disease in USH1G patients.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org. Supplementary_Figure_File.docx: all Figures S1-S5 referred to in this work; Table_S1.xlsx: GO-term analysis of SANS nuclear interactome from [12]; CellProfiler workflow available on GitHub: https://github.com/LabWolfrum/Fritze_et_al_2024_SANS_Nuclear_localization
Author Contributions
J.S.F. set up and carried out most of the experiments. F.F.S. carried out some cloning and experiments. J.S.F. and U.W. designed the studies and wrote the manuscript. All authors proofread the manuscript.
Funding
This study was supported by funds from the FAUN-Stiftung, Nuremberg, The Foundation Fighting Blindness (FFB) [PPA-0717-0719-RAD], and DFG (Project-No.: 521147274) to U.W..
Data Availability Statement
All raw data are available through contacting the corresponding author.
Acknowledgments
We thank Ronald Roepman (Human Genetics, Nijmegen) and Katja Luck (IMB, Mainz) for the contribution of several plasmids and constructs used in this study. We thank Christof Rickert (LMCF, JGU), the Microscopy Core Facility IMB, Mark Zorin (IMP, Mainz), Ulrike Maas (IMP, Mainz) and Joshua Linnert (IMP, Mainz) for their technical support. J.S.F. was enrolled in the International PhD Programme (IPP) Mainz.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Weil, D.; El-Amraoui, A.; Masmoudi, S.; Mustapha, M.; Kikkawa, Y.; Laine, S.; Delmaghani, S.; Adato, A.; Nadifi, S.; Zina, Z. Ben; et al. Usher Syndrome Type I G (USH1G) Is Caused by Mutations in the Gene Encoding SANS, a Protein That Associates with the USH1C Protein, Harmonin. Hum Mol Genet 2003, 12, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Fuster-Garcia, C.; Garcia-Bohorquez, B.; Rodriguez-Munoz, A.; Aller, E.; Jaijo, T.; Millan, J.M.; Garcia-Garcia, G. Usher Syndrome: Genetics of a Human Ciliopathy. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Delmaghani, S.; El-Amraoui, A. The Genetic and Phenotypic Landscapes of Usher Syndrome: From Disease Mechanisms to a New Classification. Hum Genet 2022, 141, 709–735. [Google Scholar] [CrossRef] [PubMed]
- Fritze, J.S.; Stiehler, F.F.; Wolfrum, U. Pathogenic Variants in USH1G/SANS Alter Protein Interaction with Pre-RNA Processing Factors PRPF6 and PRPF31 of the Spliceosome. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Adato, A.; Lefevre, G.; Delprat, B.; Michel, V.; Michalski, N.; Chardenoux, S.; Weil, D.; El-Amraoui, A.; Petit, C. Usherin, the Defective Protein in Usher Syndrome Type IIA, Is Likely to Be a Component of Interstereocilia Ankle Links in the Inner Ear Sensory Cells. Hum Mol Genet 2005, 14, 3921–3932. [Google Scholar] [CrossRef] [PubMed]
- Maerker, T.; van Wijk, E.; Overlack, N.; Kersten, F.F.J.; Mcgee, J.; Goldmann, T.; Sehn, E.; Roepman, R.; Walsh, E.J.; Kremer, H.; et al. A Novel Usher Protein Network at the Periciliary Reloading Point between Molecular Transport Machineries in Vertebrate Photoreceptor Cells. Hum Mol Genet 2008, 17, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Overlack, N.; Kilic, D.; Bauss, K.; Marker, T.; Kremer, H.; van Wijk, E.; Wolfrum, U.; Bauß, K.; Märker, T.; Kremer, H.; et al. Direct Interaction of the Usher Syndrome 1G Protein SANS and Myomegalin in the Retina. Biochim Biophys Acta 2011, 1813, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Papal, S.; Cortese, M.; Legendre, K.; Sorusch, N.; Dragavon, J.; Sahly, I.; Shorte, S.; Wolfrum, U.; Petit, C.; El-Amraoui, A. The Giant Spectrin BetaV Couples the Molecular Motors to Phototransduction and Usher Syndrome Type I Proteins along Their Trafficking Route. Hum Mol Genet 2013, 22, 3773–3788. [Google Scholar] [CrossRef] [PubMed]
- Bauss, K.; Knapp, B.; Jores, P.; Roepman, R.; Kremer, H.; Wijk, E. V; Marker, T.; Wolfrum, U. Phosphorylation of the Usher Syndrome 1G Protein SANS Controls Magi2-Mediated Endocytosis. Hum Mol Genet 2014, 23, 3923–3942. [Google Scholar] [CrossRef] [PubMed]
- Sorusch, N.; Baub, K.; Plutniok, J.; Samanta, A.; Knapp, B.; Nagel-Wolfrum, K.; Wolfrum, U. Characterization of the Ternary Usher Syndrome SANS/Ush2a/Whirlin Protein Complex. Hum Mol Genet 2017, 26, 1157–1172. [Google Scholar] [CrossRef] [PubMed]
- Sorusch, N.; Yildirim, A.; Knapp, B.; Janson, J.; Fleck, W.; Scharf, C.; Wolfrum, U. SANS (USH1G) Molecularly Links the Human Usher Syndrome Protein Network to the Intraflagellar Transport Module by Direct Binding to IFT-B Proteins. Front Cell Dev Biol 2019, 7, 216. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, A.; Mozaffari-Jovin, S.; Wallisch, A.K.; Schäfer, J.; Ludwig, S.E.J.; Urlaub, H.; Lührmann, R.; Wolfrum, U. SANS (USH1G) Regulates Pre-MRNA Splicing by Mediating the Intra-Nuclear Transfer of Tri-SnRNP Complexes. Nucleic Acids Res 2021, 49, 5845–5866. [Google Scholar] [CrossRef] [PubMed]
- Caberlotto, E.; Michel, V.; Foucher, I.; Bahloul, A.; Goodyear, R.J.; Pepermans, E.; Michalski, N.; Perfettini, I.; Alegria-Prévot, O.; Chardenoux, S.; et al. Usher Type 1G Protein sans Is a Critical Component of the Tip-Link Complex, a Structure Controlling Actin Polymerization in Stereocilia. Proc Natl Acad Sci U S A 2011, 108, 5825–5830. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, J.; Zhang, M. Myosin VII, USH1C, and ANKS4B or USH1G Together Form Condensed Molecular Assembly via Liquid-Liquid Phase Separation. Cell Rep 2019, 29, 974–986.e4. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Pan, L.; Chen, X.; Wu, L.; Zhang, M. The Structure of the Harmonin/sans Complex Reveals an Unexpected Interaction Mode of the Two Usher Syndrome Proteins. Proc Natl Acad Sci U S A 2010, 107, 4040–4045. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; He, Y.; Lu, Q.; Zhang, M. Mechanistic Basis of Organization of the Harmonin/USH1C-Mediated Brush Border Microvilli Tip-Link Complex. Dev Cell 2016, 36, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Crawley, S.W.; Weck, M.L.; Grega-Larson, N.E.; Shifrin, D.A.; Tyska, M.J. ANKS4B Is Essential for Intermicrovillar Adhesion Complex Formation. Dev Cell 2016, 36, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Graves, M.J.; Matoo, S.; Choi, M.S.; Storad, Z.A.; El Sheikh Idris, R.A.; Pickles, B.K.; Acharya, P.; Shinder, P.E.; Arvay, T.O.; Crawley, S.W. A Cryptic Sequence Targets the Adhesion Complex Scaffold ANKS4B to Apical Microvilli to Promote Enterocyte Brush Border Assembly. J Biol Chem 2020, 295, 12588–12604. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, L.; Chen, L.; Gong, B.; Jia, D.; Sun, Q. Nuclear Transport Proteins: Structure, Function, and Disease Relevance. Signal Transduct Target Ther 2023, 8. [Google Scholar] [CrossRef]
- Wing, C.E.; Fung, H.Y.J.; Chook, Y.M. Karyopherin-Mediated Nucleocytoplasmic Transport. Nat Rev Mol Cell Biol 2022, 23, 307–328. [Google Scholar] [CrossRef]
- Wiemann, S.; Pennacchio, C.; Hu, Y.; Hunter, P.; Harbers, M.; Amiet, A.; Bethel, G.; Busse, M.; Carninci, P.; Diekhans, M.; et al. The ORFeome Collaboration: A Genome-Scale Human ORF-Clone Resource. Nat Methods 2016, 13, 191–192. [Google Scholar] [CrossRef]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ Ecosystem: An Open Platform for Biomedical Image Analysis. Mol Reprod Dev 2015, 82, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Ba, A.N.N.; Pogoutse, A.; Provart, N.; Moses, A.M. NLStradamus: A Simple Hidden Markov Model for Nuclear Localization Signal Prediction. BMC Bioinformatics 2009, 10. [Google Scholar] [CrossRef]
- Xu, D.; Marquis, K.; Pei, J.; Fu, S.C.; Caʇatay, T.; Grishin, N. V.; Chook, Y.M. LocNES: A Computational Tool for Locating Classical NESs in CRM1 Cargo Proteins. Bioinformatics 2015, 31, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Stirling, D.R.; Swain-Bowden, M.J.; Lucas, A.M.; Carpenter, A.E.; Cimini, B.A.; Goodman, A. CellProfiler 4: Improvements in Speed, Utility and Usability. BMC Bioinformatics 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Positteam Download RStudio - Posit Available online:. Available online: https://posit.co/downloads/ (accessed on 24 July 2023).
- Zhou, Y.; Liu, F.; Xu, Q.; Yang, B.; Li, X.; Jiang, S.; Hu, L.; Zhang, X.; Zhu, L.; Li, Q.; et al. Inhibiting Importin 4-Mediated Nuclear Import of CEBPD Enhances Chemosensitivity by Repression of PRKDC-Driven DNA Damage Repair in Cervical Cancer. Oncogene 2020, 39, 5633–5648. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.; Neufeldt, C.J.; Pang, D.; Wilson, K.; Loewen-Dobler, D.; Joyce, M.A.; Wozniak, R.W.; Tyrrell, D.L.J. Functional Characterization of Nuclear Localization and Export Signals in Hepatitis C Virus Proteins and Their Role in the Membranous Web. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Pemberton, L.F.; Paschal, B.M. Mechanisms of Receptor-Mediated Nuclear Import and Nuclear Export. Traffic 2005, 6, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Shoubridge, C.; Tan, M.; Fullston, T.; Cloosterman, D.; Coman, D.; McGillivray, G.; Mancini, G.; Kleefstra, T.; Gécz, J. Mutations in the Nuclear Localization Sequence of the Aristaless Related Homeobox; Sequestration of Mutant ARX with IPO13 Disrupts Normal Subcellular Distribution of the Transcription Factor and Retards Cell Division. Pathogenetics 2010, 3. [Google Scholar] [CrossRef]
- Jans, D.A.; Martin, A.J.; Wagstaff, K.M. Inhibitors of Nuclear Transport. Curr Opin Cell Biol 2019, 58, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Karpova, T.S.; Baumann, C.T.; He, L.; Wu, X.; Grammer, A.; Lipsky, P.; Hager, G.L.; McNally, J.G. Fluorescence Resonance Energy Transfer from Cyan to Yellow Fluorescent Protein Detected by Acceptor Photobleaching Using Confocal Microscopy and a Single Laser. J Microsc 2003, 209, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Lou, M.; Pang, C.W.M.; Gerken, A.E.; Brock, T.G. Multiple Nuclear Localization Sequences Allow Modulation of 5-Lipoxygenase Nuclear Import. Traffic 2004, 5, 847–854. [Google Scholar] [CrossRef]
- Ferrer, M.; Rodríguez, J.A.; Spierings, E.A.; de Winter, J.P.; Giaccone, G.; Kruyt, F.A.E. Identification of Multiple Nuclear Export Sequences in Fanconi Anemia Group A Protein That Contribute to CRM1-Dependent Nuclear Export. Hum Mol Genet 2005, 14, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Di Antonio, V.; Bellucci, L.; Thomas, D.R.; Caporuscio, F.; Ciccarese, F.; Ghassabian, H.; Wagstaff, K.M.; Forwood, J.K.; Jans, D.A.; et al. Contribution of the Residue at Position 4 within Classical Nuclear Localization Signals to Modulating Interaction with Importins and Nuclear Targeting. Biochim Biophys Acta Mol Cell Res 2018, 1865, 1114–1129. [Google Scholar] [CrossRef] [PubMed]
- De Ganck, A.; Hubert, T.; Van Impe, K.; Geelen, D.; Vandekerckhove, J.; De Corte, V.; Gettemans, J. A Monopartite Nuclear Localization Sequence Regulates Nuclear Targeting of the Actin Binding Protein Myopodin. FEBS Lett 2005, 579, 6673–6680. [Google Scholar] [CrossRef] [PubMed]
- Kopp, M.; Rotan, O.; Papadopoulos, C.; Schulze, N.; Meyer, H.; Epple, M. Delivery of the Autofluorescent Protein R-Phycoerythrin by Calcium Phosphate Nanoparticles into Four Different Eukaryotic Cell Lines (HeLa, HEK293T, MG-63, MC3T3): Highly Efficient, but Leading to Endolysosomal Proteolysis in HeLa and MC3T3 Cells. PLoS One 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Esposito, A.; Choi, H.; Matarese, M.; Benedetti, V.; Di Malta, C.; Monfregola, J.; Medina, D.L.; Lippincott-Schwartz, J.; Ballabio, A. MTOR-Dependent Phosphorylation Controls TFEB Nuclear Export. Nat Commun 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, A.; Dunphy, W.G. Binding of 14-3-3 Proteins and Nuclear Export Control the Intracellular Localization of the Mitotic Inducer Cdc25. Genes Dev 1999, 13, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, D.A.; Karns, L.R.; VandenBerg, S.R.; Creutz, C.E. Control of the Nuclear-Cytoplasmic Partitioning of Annexin II by a Nuclear Export Signal and by P11 Binding. J Cell Sci 2001, 114, 3155–3166. [Google Scholar] [CrossRef] [PubMed]
- Macara, I.G. Transport into and out of the Nucleus. Microbiol Mol Biol Rev 2001, 65, 570–594. [Google Scholar] [CrossRef]
- Tessier, T.M.; Macneil, K.M.; Mymryk, J.S. Piggybacking on Classical Import and Other Non-Classical Mechanisms of Nuclear Import Appear Highly Prevalent within the Human Proteome. Biology (Basel) 2020, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.Y.J.; Fu, S.C.; Chook, Y.M. Nuclear Export Receptor CRM1 Recognizes Diverse Conformations in Nuclear Export Signals. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Timney, B.L.; Raveh, B.; Mironska, R.; Trivedi, J.M.; Kim, S.J.; Russel, D.; Wente, S.R.; Sali, A.; Rout, M.P. Simple Rules for Passive Diffusion through the Nuclear Pore Complex. J Cell Biol 2016, 215. [Google Scholar] [CrossRef]
- Bourgeois, B.; Hutten, S.; Gottschalk, B.; Hofweber, M.; Richter, G.; Sternat, J.; Abou-Ajram, C.; Göbl, C.; Leitinger, G.; Graier, W.F.; et al. Nonclassical Nuclear Localization Signals Mediate Nuclear Import of CIRBP. Proc Natl Acad Sci U S A 2020, 117, 8503–8514. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama, H.; Wynn, R.M.; Lee, W.R.; Fukasawa, M.; Mizuguchi, H.; Gardner, K.H.; Repa, J.J.; Uyeda, K. Regulation of Nuclear Import/Export of Carbohydrate Response Element-Binding Protein (ChREBP): Interaction of Anα-Helix of ChREBP with the 14-3-3 Proteins and Regulation by Phosphorylation. Journal of Biological Chemistry 2008, 283, 24899–24908. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, X.; Gusev, E.; Wang, C.; Fagotto, F. Regulation of the Phosphorylation and Nuclear Import and Export of β-Catenin by APC and Its Cancer-Related Truncated Form. J Cell Sci 2014, 127, 1647–1659. [Google Scholar] [CrossRef] [PubMed]
- Nardozzi, J.D.; Lott, K.; Cingolani, G. Phosphorylation Meets Nuclear Import: A Review. Cell Commun Signal 2010, 8. [Google Scholar] [CrossRef] [PubMed]
- Fokkema, I.F.A.C.; Taschner, P.E.M.; Schaafsma, G.C.P.; Celli, J.; Laros, J.F.J.; den Dunnen, J.T. LOVD v.2.0: The next Generation in Gene Variant Databases. Hum Mutat 2011, 32, 557–563. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Nuclear localization of SANS. (A) Domain structure of SANS and its paralogue ANKS4B: Both consist of three ankyrin repeats (ANK1-3), a central domain (CENT), divided in SANS into three parts (CENTn1, CENTn2, and CENTc), a sterile alpha motif (SAM), and a C-terminal type-I PDZ binding motif (PBM, asterisk). ANKS4B and SANS amino acid sequences are highly similar in their N- and C-terminal region. (B, C) Confocal microscopy of HeLa cells either stained for endogenous SANS (B) or transfected with eYFP-SANS, 3xFLAG-SANS (C), counterstained with DAPI. Endogenous SANS and 3xFLAG-SANS was visualized by indirect immunofluorescence of anti-SANS and anti-FLAG. (B, C) Endogenous SANS, eYFP-SANS, and 3xFLAG-SANS were localized in the cytoplasm and the nucleus. (D) Quantification of eYFP-SANS localization by CellProfiler in HeLa cells. eYFP-SANS is localized in both nucleus and cytoplasm. (E) Quantification of eYFP-SANS localization by Cell fractionation assay in HeLa cells. eYFP-SANS is localized similar to (D) in nucleus and cytoplasm. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
Figure 1.
Nuclear localization of SANS. (A) Domain structure of SANS and its paralogue ANKS4B: Both consist of three ankyrin repeats (ANK1-3), a central domain (CENT), divided in SANS into three parts (CENTn1, CENTn2, and CENTc), a sterile alpha motif (SAM), and a C-terminal type-I PDZ binding motif (PBM, asterisk). ANKS4B and SANS amino acid sequences are highly similar in their N- and C-terminal region. (B, C) Confocal microscopy of HeLa cells either stained for endogenous SANS (B) or transfected with eYFP-SANS, 3xFLAG-SANS (C), counterstained with DAPI. Endogenous SANS and 3xFLAG-SANS was visualized by indirect immunofluorescence of anti-SANS and anti-FLAG. (B, C) Endogenous SANS, eYFP-SANS, and 3xFLAG-SANS were localized in the cytoplasm and the nucleus. (D) Quantification of eYFP-SANS localization by CellProfiler in HeLa cells. eYFP-SANS is localized in both nucleus and cytoplasm. (E) Quantification of eYFP-SANS localization by Cell fractionation assay in HeLa cells. eYFP-SANS is localized similar to (D) in nucleus and cytoplasm. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
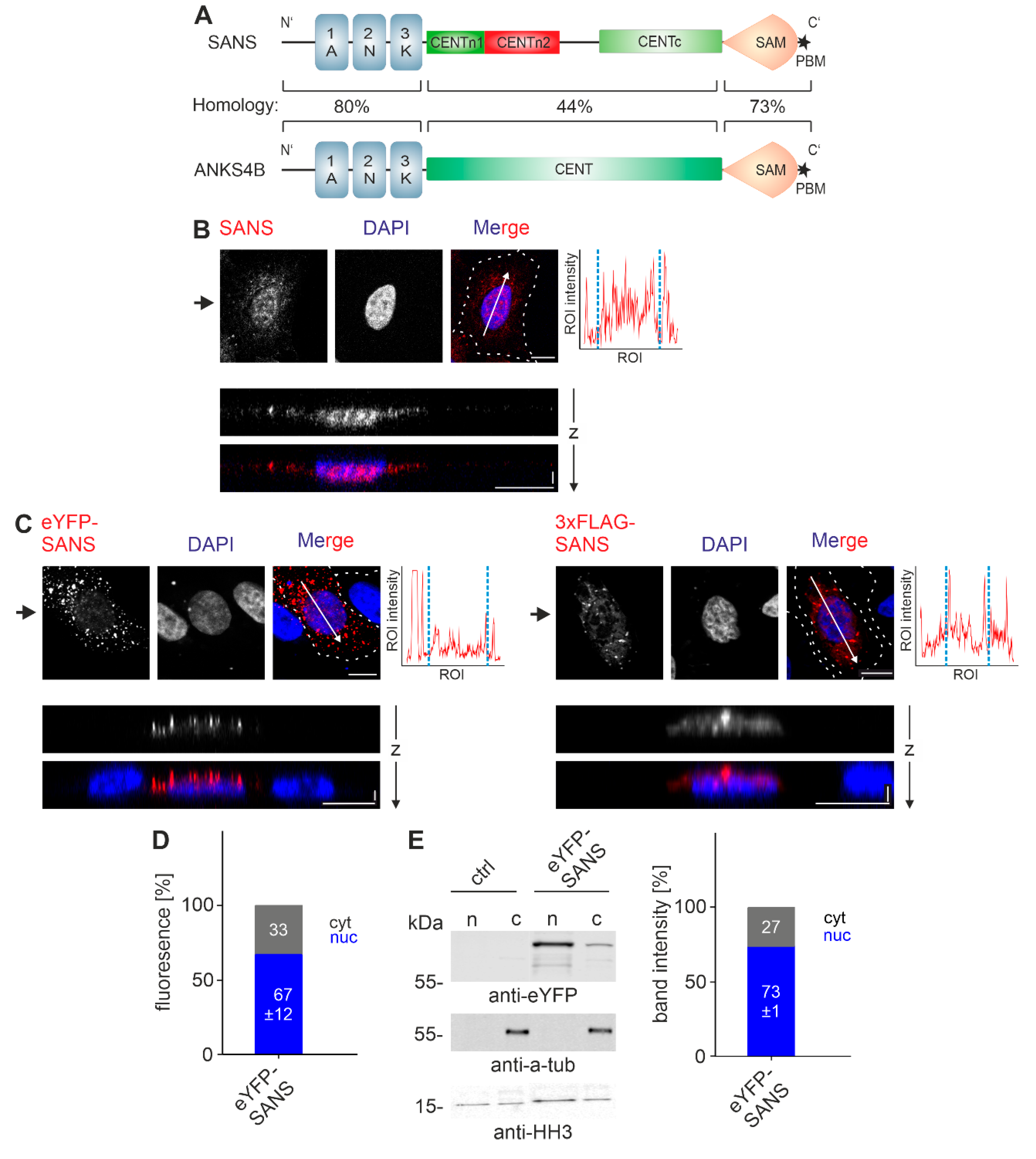
Figure 2.
Nuclear localization of SANS in HeLa after treatment with Leptomycin-B. (A) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) and treated with 5 nM CRM1-inhibitor LeptomycinB (LMB) or its solvent DMSO. (B) Quantification of (A) by CellProfiler. eYFP-SANS was significantly enriched in the nucleus after LMB treatment. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
Figure 2.
Nuclear localization of SANS in HeLa after treatment with Leptomycin-B. (A) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) and treated with 5 nM CRM1-inhibitor LeptomycinB (LMB) or its solvent DMSO. (B) Quantification of (A) by CellProfiler. eYFP-SANS was significantly enriched in the nucleus after LMB treatment. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
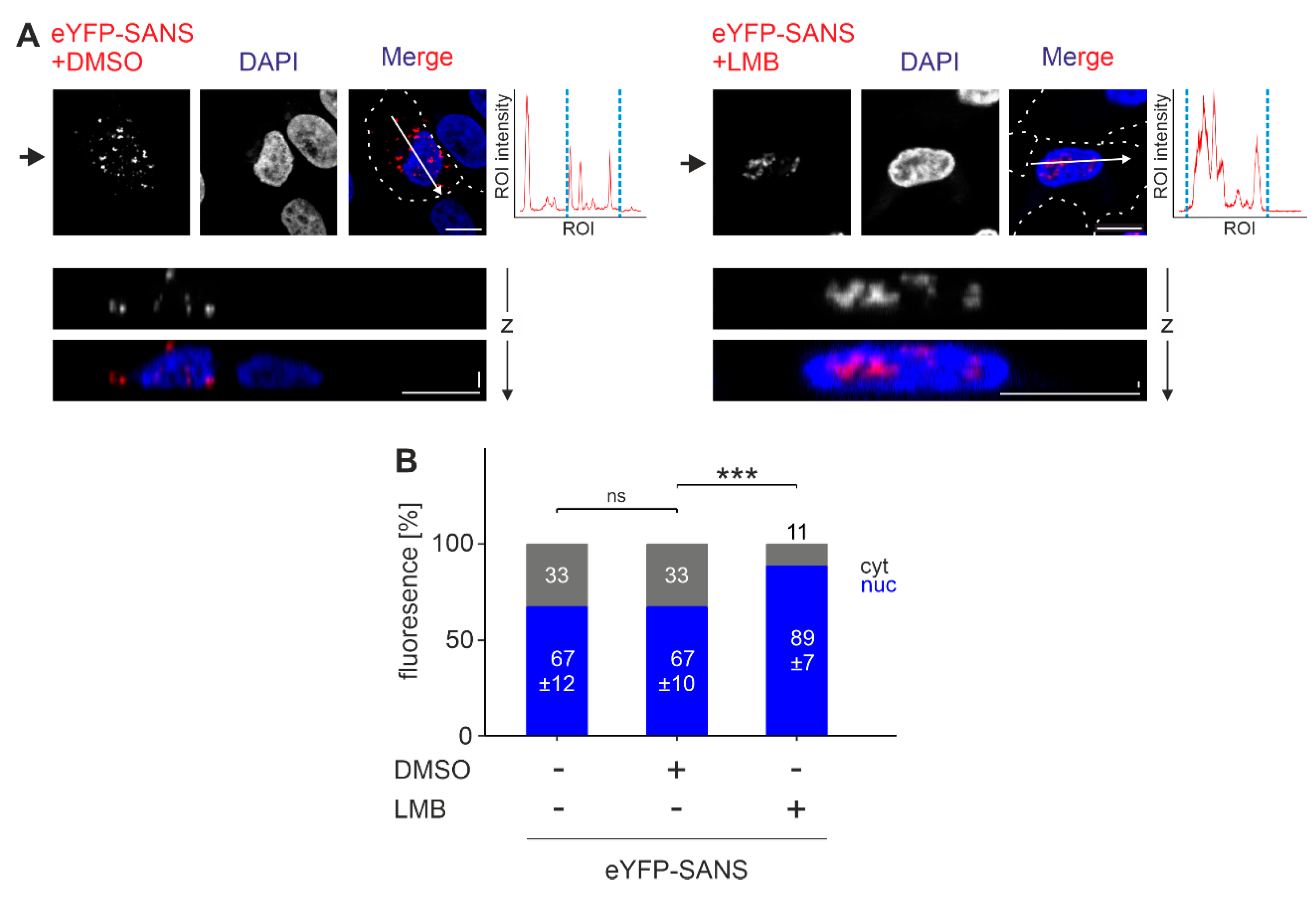
Figure 3.
Prediction of nuclear localization sequences (NLSs) and nuclear export sequences (NESs) of SANS. (A) Three NES ($; NES_1181-195, NES_2235-249, NES_3406-420) and two NLS (#; NLS_1213-224, NLS_2436-447) were predicted for SANS by NLStradamus and LocNES. (B-F) Conservation of SANS NLS_1213-224 (B), NLS_2436-447 (C), NES_1181-195 (D), NES_2235-249 (E) and NES_3406-420 (F) with blastp to Human SANS for five model organisms. In B-C, positive charged residues (red), arginine-to-lysin exchanges (purple), charge changes (green) are indicated in NLSs. In D-F, first line: position of consensus amino acids residues in CRM1-motif are indicated in red. Valid (purple) and invalid (green) residue changes in the consensus sequence of the CRM1 motif are indicated. ** and *** indicate that the genomes of D. rerio and X. laevis possess two and three Ush1g genes encoding for SANS orthologues, respectively, which do not differ at the NLS and NES sequences.
Figure 3.
Prediction of nuclear localization sequences (NLSs) and nuclear export sequences (NESs) of SANS. (A) Three NES ($; NES_1181-195, NES_2235-249, NES_3406-420) and two NLS (#; NLS_1213-224, NLS_2436-447) were predicted for SANS by NLStradamus and LocNES. (B-F) Conservation of SANS NLS_1213-224 (B), NLS_2436-447 (C), NES_1181-195 (D), NES_2235-249 (E) and NES_3406-420 (F) with blastp to Human SANS for five model organisms. In B-C, positive charged residues (red), arginine-to-lysin exchanges (purple), charge changes (green) are indicated in NLSs. In D-F, first line: position of consensus amino acids residues in CRM1-motif are indicated in red. Valid (purple) and invalid (green) residue changes in the consensus sequence of the CRM1 motif are indicated. ** and *** indicate that the genomes of D. rerio and X. laevis possess two and three Ush1g genes encoding for SANS orthologues, respectively, which do not differ at the NLS and NES sequences.
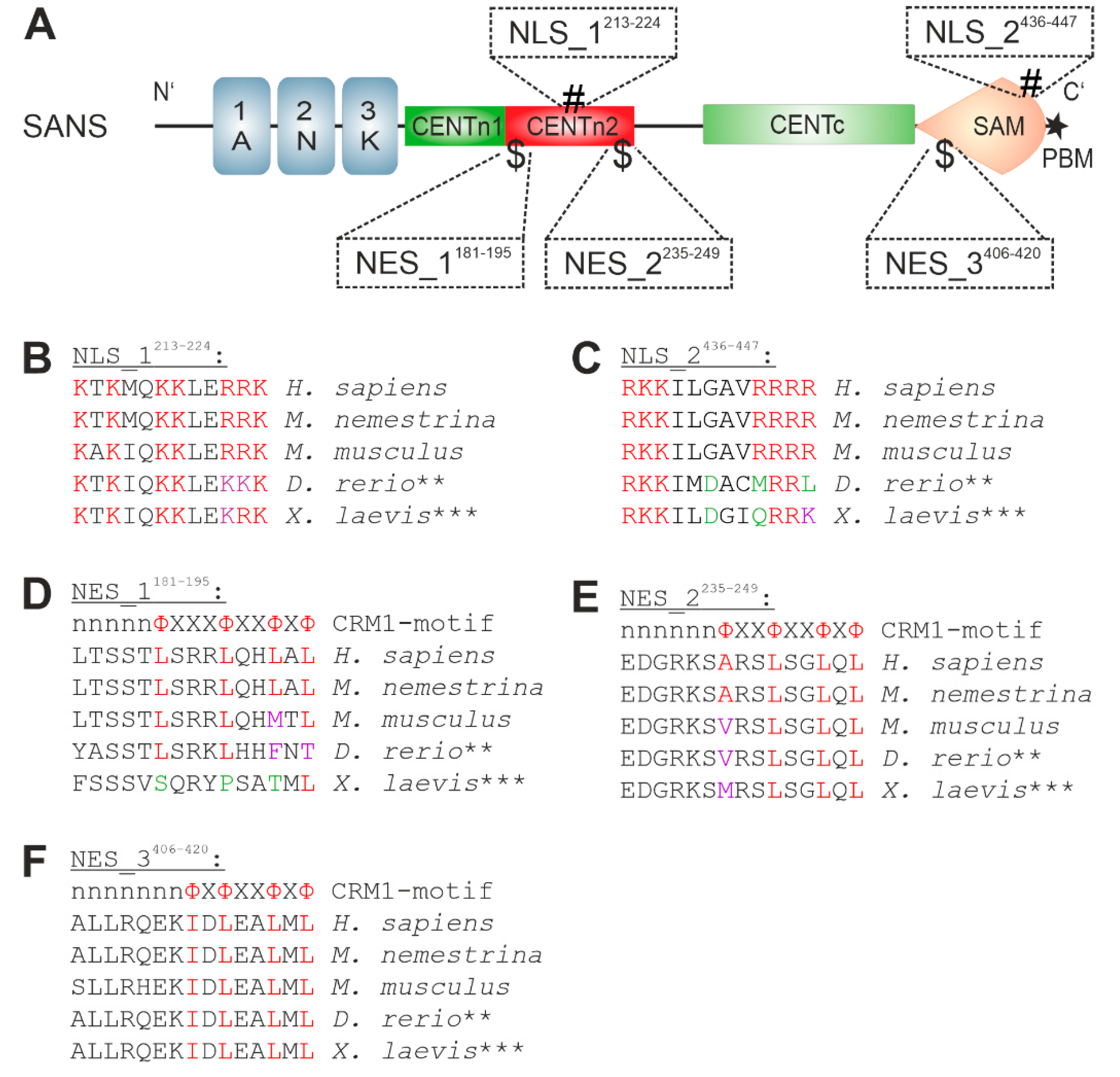
Figure 4.
Subcellular localization of SANS NLS mutants. (A) Domain structure of SANS, the two NLS (#) mutants K213E (ΔNLS_1213-224) and R447W (ΔNLS_2436-447) are indicated above the cartoon. (B) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) or SANS NLS mutants, counterstained with DAPI. (C) Quantification of (B) by CellProfiler. eYFP-SANSK213E differed significantly from eYFP-SANS. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
Figure 4.
Subcellular localization of SANS NLS mutants. (A) Domain structure of SANS, the two NLS (#) mutants K213E (ΔNLS_1213-224) and R447W (ΔNLS_2436-447) are indicated above the cartoon. (B) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) or SANS NLS mutants, counterstained with DAPI. (C) Quantification of (B) by CellProfiler. eYFP-SANSK213E differed significantly from eYFP-SANS. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.

Figure 5.
Localization of SANS NES mutants. (A) Domain structure of SANS, the two NES ($) mutants L195E (ΔNES_1181-195) and L249E (ΔNES_2235-249) are indicated below the cartoon. (B) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) or NES mutants, counterstained with DAPI. (C) Quantification of (B) by CellProfiler. eYFP-SANSL195E was significantly enriched in the nucleus compared to eYFP-SANS. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
Figure 5.
Localization of SANS NES mutants. (A) Domain structure of SANS, the two NES ($) mutants L195E (ΔNES_1181-195) and L249E (ΔNES_2235-249) are indicated below the cartoon. (B) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) or NES mutants, counterstained with DAPI. (C) Quantification of (B) by CellProfiler. eYFP-SANSL195E was significantly enriched in the nucleus compared to eYFP-SANS. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
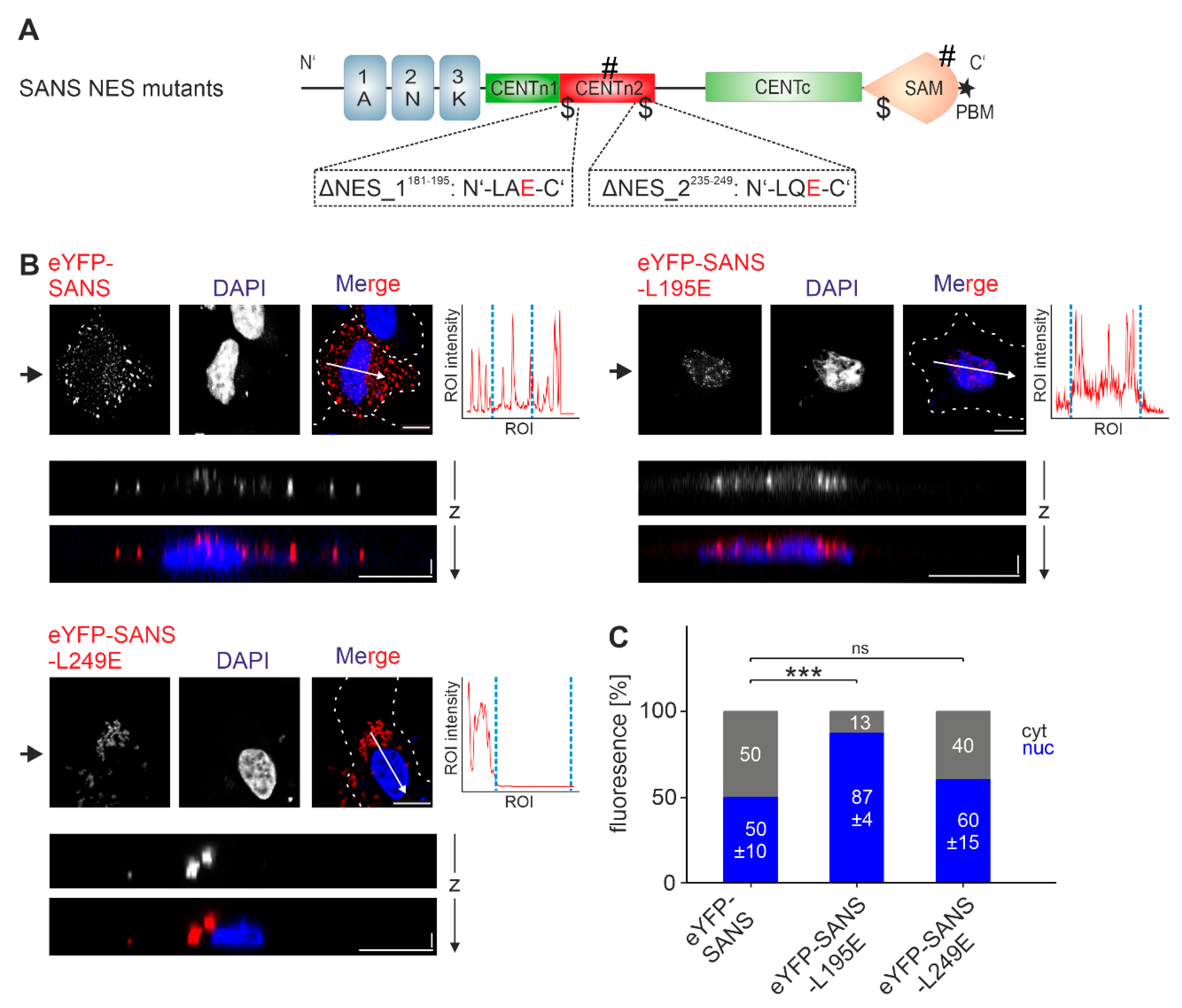
Figure 6.
Localization of pathogenic variant of SANS in HeLa cells. (A) The pathogenic variant SANSS278Pfs*71 and SANSV132Gfs*3 are frameshift mutations which lead to missense extension (red boxes and amino acid sequences below) and a premature stop in CENTc or CENTn1, respectively. The pathogenic variant SANSS243* has a premature stop codon after the CENTn2 domain. (B) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red), eYFP-SANSS278Pfs*71, eYFP-SANSS243* and eYFP-SANSV132Gfs*3, counterstained with DAPI. (C) Quantification of (B) with CellProfiler. All pathogenic variants were significantly enriched in the nucleus. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
Figure 6.
Localization of pathogenic variant of SANS in HeLa cells. (A) The pathogenic variant SANSS278Pfs*71 and SANSV132Gfs*3 are frameshift mutations which lead to missense extension (red boxes and amino acid sequences below) and a premature stop in CENTc or CENTn1, respectively. The pathogenic variant SANSS243* has a premature stop codon after the CENTn2 domain. (B) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red), eYFP-SANSS278Pfs*71, eYFP-SANSS243* and eYFP-SANSV132Gfs*3, counterstained with DAPI. (C) Quantification of (B) with CellProfiler. All pathogenic variants were significantly enriched in the nucleus. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for three independent experiments with a minimum of 75 cells.
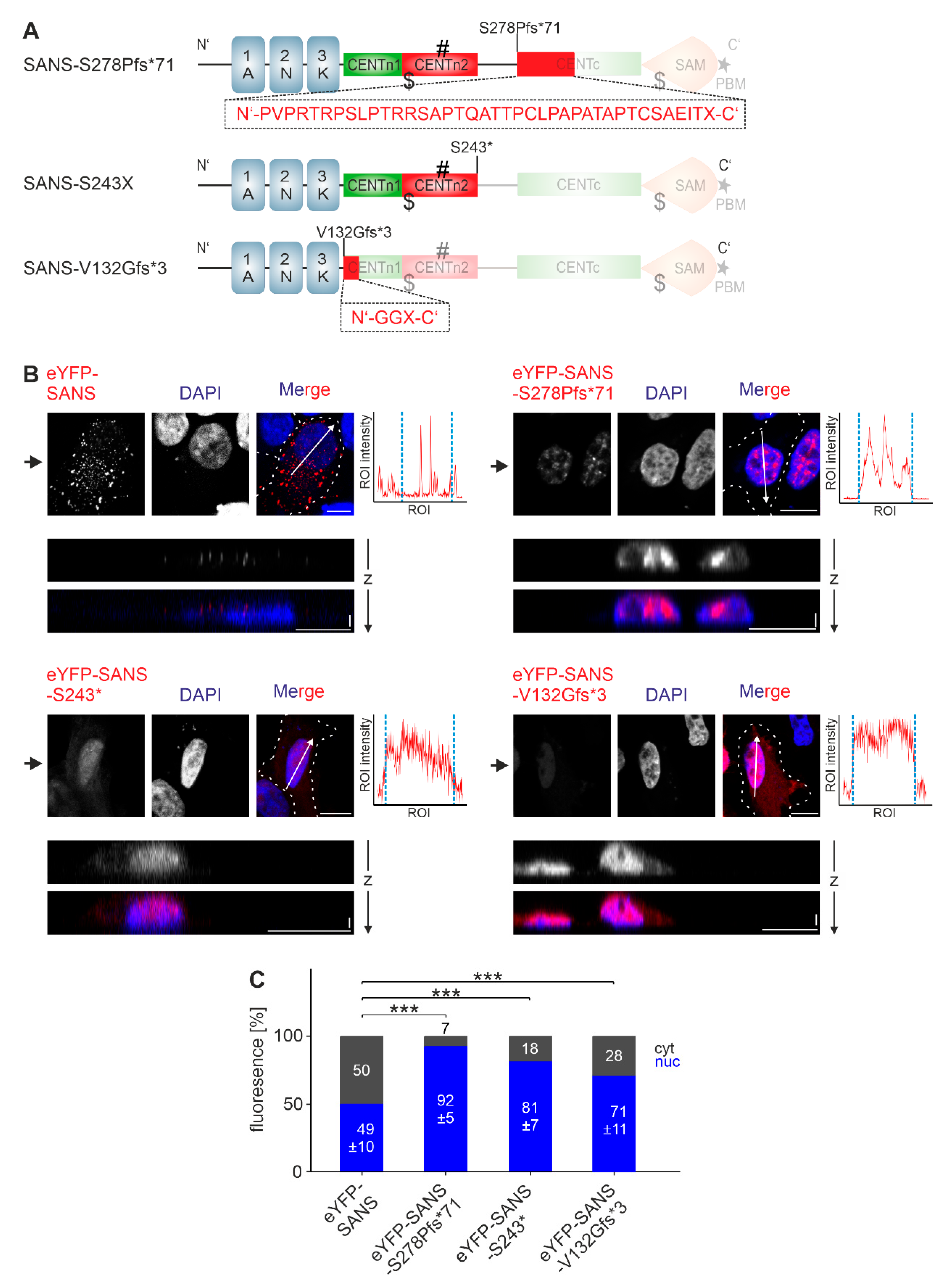
Figure 7.
Comparison of the nuclear-cytoplasmic localization of SANS and its paralogue ANKS4B in HeLa cells. (A) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) or eYFP-ANKS4B (red), counterstained with DAPI. (B) Quantification of (A) with CellProfiler. eYFP-ANKS4B was highly enriched in the cytoplasm compared to eYFP-SANS. (C) Confocal microscopy of HeLa cells transfected with eYFP-ANKS4B (red), counterstained with DAPI. Cells were treated with DMSO or 5 nM CRM1 inhibitor Leptomycin-B (LMB). (D) Quantification of (C) with CellProfiler. eYFP-ANKS4B was highly enriched in the nucleus after treatment with LMB. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for 3 independent experiments with a minimum of 75 cells.
Figure 7.
Comparison of the nuclear-cytoplasmic localization of SANS and its paralogue ANKS4B in HeLa cells. (A) Confocal microscopy of HeLa cells transfected with eYFP-SANS (red) or eYFP-ANKS4B (red), counterstained with DAPI. (B) Quantification of (A) with CellProfiler. eYFP-ANKS4B was highly enriched in the cytoplasm compared to eYFP-SANS. (C) Confocal microscopy of HeLa cells transfected with eYFP-ANKS4B (red), counterstained with DAPI. Cells were treated with DMSO or 5 nM CRM1 inhibitor Leptomycin-B (LMB). (D) Quantification of (C) with CellProfiler. eYFP-ANKS4B was highly enriched in the nucleus after treatment with LMB. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Students t-test was performed for 3 independent experiments with a minimum of 75 cells.
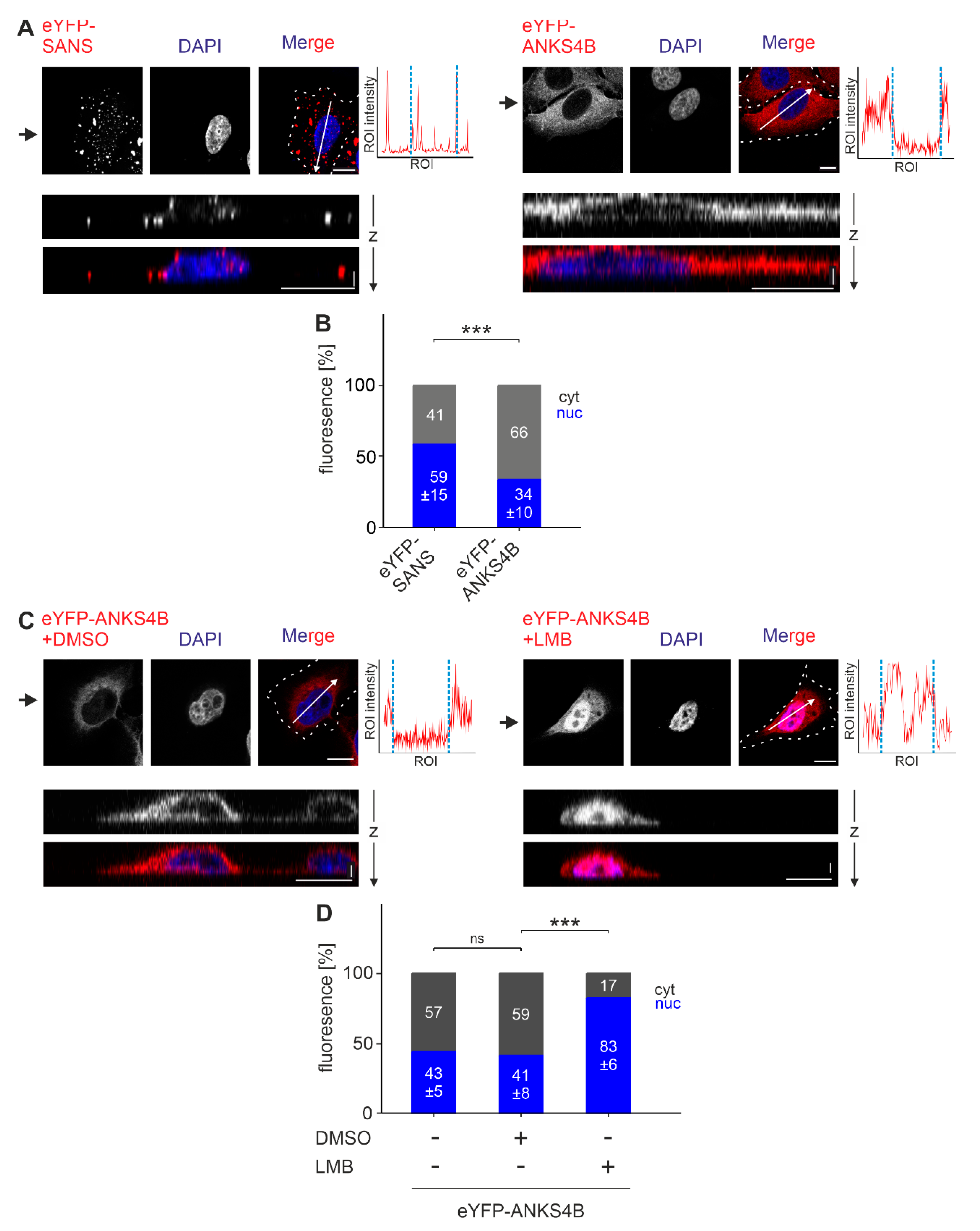
Figure 8.
ANKS4B binary interaction in the nucleus. (A) Illustration of the FRET acceptor bleach assay. Interaction of two proteins tagged with either eCFP (blue) or eYFP (yellow) leads to FRET (upper). The acceptor (eYFP) is bleached (flash symbol) (middle), which leads to increased emission of the donor (eCFP) (lower). (B) FRET assay in co-transfected HeLa cells. FRET efficiencies were normalized to 1 by the fused eCFP-c-eYFP FRET pair (positive control). FRET pair eYFP-ANKS4B-eCFP-PRPF31 show not a significant increase in the normalized FRET efficiencies when compared to eYFP FRET pair negative controls. Outliers are shown as dots above/below the boxplots. Dunn’s test after a Kruskal-Wallis test was performed for three independent experiments. (C) Confocal microscopy of HeLa cells co-transfected with eYFP-SANS (red) or eYFP-ANKS4B (red) and harmonin-mCherry (green) or mRFP-PRPF31 (green), counterstained with DAPI. eYFP-ANKS4B only co-localized with harmonin-mCherry but not with mRFP-PRPF31. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Pearson coefficient R values indicate co-localization.
Figure 8.
ANKS4B binary interaction in the nucleus. (A) Illustration of the FRET acceptor bleach assay. Interaction of two proteins tagged with either eCFP (blue) or eYFP (yellow) leads to FRET (upper). The acceptor (eYFP) is bleached (flash symbol) (middle), which leads to increased emission of the donor (eCFP) (lower). (B) FRET assay in co-transfected HeLa cells. FRET efficiencies were normalized to 1 by the fused eCFP-c-eYFP FRET pair (positive control). FRET pair eYFP-ANKS4B-eCFP-PRPF31 show not a significant increase in the normalized FRET efficiencies when compared to eYFP FRET pair negative controls. Outliers are shown as dots above/below the boxplots. Dunn’s test after a Kruskal-Wallis test was performed for three independent experiments. (C) Confocal microscopy of HeLa cells co-transfected with eYFP-SANS (red) or eYFP-ANKS4B (red) and harmonin-mCherry (green) or mRFP-PRPF31 (green), counterstained with DAPI. eYFP-ANKS4B only co-localized with harmonin-mCherry but not with mRFP-PRPF31. White arrows in merge images: regions of interests (ROI) of fluorescence intensity plots; blue dashed lines: DAPI positive nuclear extension; black arrows: position of Z-projections. Scale bars: horizontal = 10 µm; vertical = 2 µm. Pearson coefficient R values indicate co-localization.
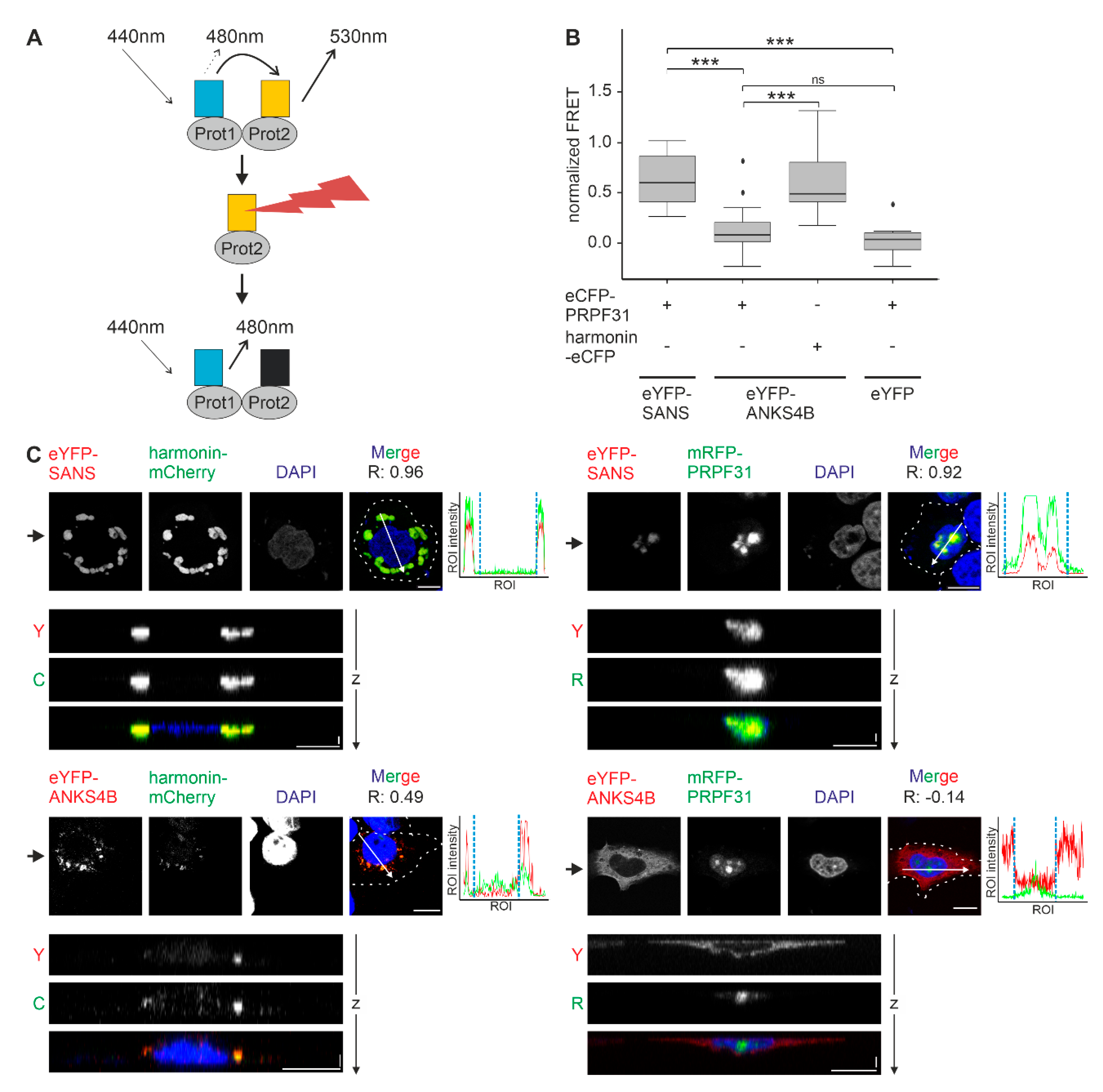

Table 1.
Nuclear exportins and importins identified as potential SANS interacting proteins in the nucleus [12].
Table 1.
Nuclear exportins and importins identified as potential SANS interacting proteins in the nucleus [12].
| Gene | Protein | Export/Import | NLS/NES Recognition |
|---|---|---|---|
| KPNB1 | Importin-β | Protein importer | classic NLS [20] non classic NLS [20] |
| IPO4 | Importin-4 | Protein importer | classic NLS [27] non classic NLS [20] |
| IPO5 | Importin-5 | Protein importer | classic NLS [28] non classic NLS [20] |
| IPO7 | Importin-7 | Protein importer | classic NLS [29] non classic NLS [20] |
| IPO8 | Importin-8 | Protein importer | classic NLS [29] non classic NLS [20] |
| IPO9 | Importin-9 | Protein importer | non classic NLS [20] |
| IPO11 | Importin-11 | Protein importer | non classic NLS [20] |
| IPO13 | Importin-13 | Protein importer | classic NLS [30] non classic NLS [30] |
| XPO1 | CRM1/Exportin-1 | Protein exporter | NES |
| XPO5 | Exportin-5 | dsRNA exporter | - |
| XPOT | Exportin-T | amino-acylated tRNAs export | - |
Table 2.
Nuclear localization (NLSs) and export sequences (NESs) of SANS predicted by LocNES and NLStradamus.
Table 2.
Nuclear localization (NLSs) and export sequences (NESs) of SANS predicted by LocNES and NLStradamus.
| NLS/NES | Sequence | Score | CRM1-class | NLS/NES Mutations |
|---|---|---|---|---|
| NLS_1213-224 | 213-KTKMQKKLERRK-224 | 0.733 | - | K213E: ETKMQKKLERRK |
| NLS_2436-447 | 436-RKKILGAVRRRR-447 | 0.679 | - | R447W: RKKILGAVRRRW |
| NES_1181-195 | 181-LTSSTLSRRLQHLAL-195 | 0.262 | 1a | L195E: LTSSTLSRRLQHLAE |
| NES_2235-249 | 235-EDGRKSARSLSGLQL-249 | 0.250 | 1b | L249E: EDGRKSARSLSGLQE |
| NES_3406-420 | 406-ALLRQEKIDLEALML-420 | 0.462 | 2 | n.a. |
Table 3.
Predicted nuclear export sequences (NES) of ANKS4B.
| NLS/NES | Sequence | Score | CRM1-Class |
|---|---|---|---|
| NES_1336-350 | 336-VEWEEDVVDATPLEV-350 | 0.191 | 1c |
| NES_2338-352 | 338-WEEDVVDATPLEVFL-352 | 0.199 | 1c |
| NES_3339-353 | 339-EEDVVDATPLEVFLL-353 | 0.205 | 1b |
| NES_4346-360 | 346-TPLEVFLLSQHLEEF-360 | 0.181 | 2 |
| NES_5349-363 | 349-EVFLLSQHLEEFLPI-363 | 0.104 | 3 |
| NES_6362-376 | 362-PIFKREQIDLEALLL-376 | 0.251 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



