
Submitted:
29 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
The present study examined metabolites in Northern wild rice (WLD), the impact of WLD supplemented high fat (HF) diet on gut microbiota, short chain fatty acids (SCFAs) and correlation with metabolism or inflammation versus HF+white rice (WHR) in mice. C57BL/6J mice received HF diet supplemented with 26g% WHR, 26g% WLD, or 13g% WHR+13g% WLD (WTWD) for 12 weeks. The levels of fasting plasma glucose, total cholesterol, triglycerides, insulin resistance, inflammatory cytokines and monocyte adhesion were lower, and the abundances of fecal Lactobacillus gasseri species bacteria and propionic acid were higher in HF+WLD diet-fed mice compared to HF+WHR diet-fed mice (p<0.05). The anti-inflammatory effects of HF+WTWD diet were weaker than that in HF+WLD diet, but were greater than that in HF+WHR diet-fed mice (p<0.05). Relative abundances of fecal Lactobacillus gasseri and propionic acid in HF+WLD fed-mice were higher than that in HF+WHR fed mice. The abundance of fecal L. gasseri and propionic acid negatively correlated with metabolic and inflammatory markers (p<0.05). The findings suggest that WLD dose-dependently attenuated metabolic and inflammatory disorders in HF diet-fed mice. Interactions between WLD components and gut microbiota may upregulate fecal propionic acid and contribute to metabolic and anti-inflammatory benefits of WLD diet-induced insulin resistant mice.
Keywords:
wild rice
; gut microbiota
; short chain fatty acids
; high fat diet
; mice
; metabolism
; chronic inflammation
; Lactobacillus gasseri
; propionic acid
1. Introduction
Northern wild rice (Zizania palustris, WLD) is an annual plant naturally growing in shallow water in Great Lake and Canadian Prairie regions. WLD was a traditional food of Indigenous people in North America and known as Indian rice or manoomin [1]. WLD is not a type of rice, but it has been considered as a kind of healthy whole grain [2]. WLD grain contains significantly more contents of proteins, fiber, vitamins and antioxidants compared to refined white rice (WHR) [3].
Previous studies demonstrated that WLD reduced hypercholesterolemia, hypertriglyceridemia and the extents of atherosclerotic lesions in low-density lipoprotein (LDL) receptor-knockout mice compared to WHR [4,5,6]. WHR is the most commonly consumed rice product in worldwide regions. The refining process removed most nutrients from the out layer of brown rice. Intake of WHR is associated with increased risks for type 2 diabetes (T2D) [7,8]. T2D is characterized by hyperglycemia, insulin resistance, obesity and chronic low-grade inflammation [9]. Our recent study demonstrated that the replacement of WHR with WLD reduced hyperglycemia, hyperlipidemia, insulin resistance and inflammatory markers in high fat (HF) diet-induced insulin resistant mice [10]. The underlying mechanism for the metabolic and anti-inflammatory effects of WLD remains to be determined.
Gastrointestinal tract or gut is the major organ in the body to absorb nutrients from foods and plays critical roles in metabolism and inflammation [11]. In addition, gut is the largest reservoir for microbes in the body. Most of gut bacteria are not harmful to the body, and many of them are even beneficial to health since they help to generate healthy nutrients from foods, modulate metabolism, and inhibit inflammation and the overgrowth of harmful microbes in the gut. The diversities and composition of bacteria in gut was affected by HF diet [12]. Gut bacteria are essential for HF diet-induced obesity [13]. Multiple groups of bacteria promote the digest of insoluble fiber from foods and generate short chain fatty acids (SCFAs) in gut. SCFAs are involved in the modulation of metabolism and inflammation in the body [14].
The present study examined the metabolites in WLD versus WHR, and the effects of the supplementation of two dosages of WLD to HF diet on metabolism, vascular inflammation, pro-inflammatory cytokines, gut microbiota and fecal SCFAs in mice compared to WHR-supplemented HF diet. Correlations between fecal microbiota, SCFAs and circulatory metabolic or inflammatory cytokines in the mice were further investigated.
2. Materials and Methods
2.1. Dietary Components
WLD was obtained from Floating Leaf Wild Rice Inc. (Springfield, MB). Asian white rice was obtained from local grocery store. Carbohydrate-free HF diet powder (cat#: D12492px11) was received from Research Diets (New Brunswick, NJ) which contains 47% of fat (91% from lard) and 35% of protein (weight/weight or w/w). WLD and WHR were grinded and passed through a 0.5 mm mesh. After the supplementation of 26 g% weight of rice powder, experimental HF diet was pelleted, which contained 35 g% fat accounting for 60% of the total calories, 26 g% protein for 20% of calories and 26 g% carbohydrate for the remaining 20% of calories in the diet.
2.2. Animals
C57 BL/6J mice (male, 6 weeks of age) were obtained from the Jackson Laboratory (Bar Harbor, ME). Mice were held in stainless steel cages in air-conditioned room and received regular rodent chow for one week for stabilization. The mice were randomly divided into 3 groups (n=5/group) after the stabilization. WHR diet group received HF diet supplemented with 26 g% of WHR (w/w). WTWD diet group was fed with HF diet supplemented with 13 g% of WHR and 13 g% of WLD. WLD diet group received HF diet supplemented with 26 g% of WLD. The dietary intervention lasted for 12 weeks.
2.3. Animal Monitoring and Sample Collection
Body weights and food intake were recorded at the onset and before the end of dietary intervention. Blood was collected from the saphenous vein to measure blood biochemical variables. At the end of the intervention, mice were euthanized via isoflurane (5%, v/v) inhalation. Abdominal aortae were surgically harvested and submerged in ice-cold Hank’s balance salt solution (HBSS) in culture dishes for ex vivo monocyte adhesion assay as previously described [15]. The protocol of the animal experiment was approved by the Animal Management and Protocol Committee at the University of Manitoba.
2.4. Measurements of Circulatory Glucose, Triglycerides and Cholesterol
The levels of plasma glucose, total cholesterol and triglycerides of mice after an overnight fasting were analysed using Sekisui Diagnostics SL reagents (Charlottetown, PE, Canada).
2.5. Measurements of Plasma Insulin and Pro-Inflammatory Cytokines
The levels of insulin, tumor necrosis factor-α (TNFα), plasminogen activator inhibitor-1 (PAI-1) and monocyte chemotactic protein-1 (MCP-1) antigens in plasma were measured using enzyme-linked immunosorbent assay (ELISA) kits from EMD Millipore (Billerica, MA for insulin), BD Bioscience (San Diego, CA for TNFα), Thermo Scientific (Ottawa, ON for MCP-1), and Oxford Biomedical Research (Oxford, MI for PAI-1). Homeostatic model assessment-insulin resistance (HOMA-IR) was calculated from plasma glucose and insulin in simultaneously collected blood samples using a mouse-specific formula [16].
2.6. Monocyte Adhesion Assay
Mouse abdominal aortae were isolated at the end of dietary regimen and submerged in ice-cold HBSS in 100 mm culture dishes. Fluorescently labeled WEHI-274.1 mouse monocytes (1x105) were added to each dish and incubated in Roswell Park Memorial Institute 1640 medium at room temperature with freshly isolated aorta for 30 min on a rotating mixer. Non-adhered monocytes were carefully removed via washes using ice-cold HBSS twice. Monocytes adhered on the surface of aorta were counted under microscopy (10X magnification). The averages of adhered monocytes on the intima of an aorta in five fields were applied for data analysis [15].
2.7. Fecal Bacteria DNA Extraction and 16S rRNA Gene Sequencing
Feces were collected from cages of mice individually hosted for overnight. DNA was extracted from feces using Power Fecal DNA Isolation Kit (QIAGEN, Germantown, MD) and quantified using a NanoDrop 2000 spectrophotometer (Thermo Scientific). Fecal DNA was amplified using primers targeting the V4-V5 region of bacterial DNA sequence [515F (5′-GTGYCAGCMGCCGCGGTAA) and 926R (5′-CCGYCAATTYMTTTRAGTTT)]. The DNA amplicons were normalized using Charm Biotech 96-well normalization kit for 16S rRNA gene sequencing on an Illumina MiSeq sequencer in Integrated Microbiome Resource at the Dalhousie University as previously described [17].
2.8. Bioinformatics Analyses of Gut Microbiota
The fastq file containing raw gut microbiome data was demultiplexed and then was trimmed to remove primers. Trimmed reads were imported into Quantitative Insights into Microbial Ecology 2 (QIIME2) platform. Diversity metrics (Core-metrics-phylogenetic) within QIIME2 were used to evaluate α- and β-diversity of gut microbiota [17].
2.9. Analysis of Fecal SCFAs
Fecal fatty acids were extracted using propyl chloroformate and derivatized with a reaction system containing propanol, pyridine and water. The analysis of SCFAs was conducted on an Agilent 7890A gas chromatography coupled with an Agilent 5975A inert XL EI/CI mass spectrometry in Microbiome Insights (Vancouver, BC, Canada) through customer service as previously described [18].
2.10. Extraction and Metabolomics Sample Analysis of WR
WLD and WHR were milled separately and passed through a 60 mesh filter. Rice powders (100 mg in triplicates) were extracted with 2 mL of methanol: water (3:2, v/v) and vortexed for 2 min, and then sonicated for 40 min at 4˚C. After a centrifugation (14,000 rpm) for 10 minutes at 4˚C using HERMLE 16/018 rotor (Hermle Labortechnik GmbH, Siemensstr, Wehingen, Germany), the supernatant was dried under nitrogen and stored at -80°C. Prior to injections, dried extract were reconstituted with 200 µL of water: acetonitrile (4:1, v/v) containing 300 ng norvaline. Analyses were conducted in a high performance liquid chromatography (HPLC) system (1290 Infinity Agilent Ltd., Santa Clara, CA) coupled with a 6538 UHD Accurate LC-Quadrupole Time-Of-Flight (QTOF)-mass spectrometry (MS) (Agilent Technologies, Santa Clara, CA) with dual electro-spray ionization (ESI) source. A 2.1 mm x 100 mm Agilent ZORBAX SB-Aq column (Agilent Technologies) was maintained at 60°C for chromatographic separation of samples using water (A) and acetonitrile (B) containing 0.1% formic acid. Run time for the reactions was set at 10 minutes with gradient of 0±6 min 2% B; 6±8.50 min 60% B; 8.50±8.60 min 2% B and 8.60±10 min 2% B were used to re-equilibrate the column. For each individual sample analysis, 2 μL of extract was injected with a flow rate maintained at 0.7 mL/min.
2.11. Metabolomics Data Acquisition
Metabolomic data acquisitions were completed in both positive (+) and negative (-) ESI modes. MS spectra were collected within the range of 50±1700 m/z and known references masses of 121.0508 and 922.0097 (ESI+) and 112.9860 and 1033.9880 (ESI-) utilized during all runs. The liquid chromatography-QTOF-MS metabolomics data workflow followed standard protocols and utilized multiple algorithms incorporated in Agilent Mass Hunter Qualitative (MHQ, 7.01 and Mass Profiler Professional software through custom service in the Canadian Centre for Agri-Food Research in Health and Medicine [10]. Kyoto Encyclopedia of Genes and Genomes (KEGG) database was used to assign potential physiological functions.
2.12. Statistics
Quantitative data was presented as means±standard deviation (SD). Probabilities from data >2 groups were analyzed using the one-way analysis of variance assay (ANOVA), followed by Kruskal-Wallis test paired with post-hoc Tukey test or Pairwise Wilcoxon test for probability between two groups. Liner discriminant analysis Effect Size (LEfSe) of gut microbiota was analyzed using Galaxy module. Correlations between two sets of variables were assessed using linear regression analysis. OriginPro 2021 software was used for plotting and statistical analysis. Probability <0.05 were considered as significant.
3. Results
3.1. Metabolites in WLD versus WHR
A total of 1,892 metabolites were identified in WLD and WHR via metabolomic analysis. A group of metabolites presented in10-20-fold more abundant in WLD than in WHR (p<0.001, Figure 1), which potentially regulate glucose metabolism, insulin, vascular tone, inflammation or with known therapeutic effects for diabetes or its complications. The metabolites were listed decently according to fold increase of their abundances in WLD versus WHR, which include amino acids or derivatives (homoarginine, L-glutamate), nucleotides (adenosine 5-monophosphate or AMP), glucose metabolism intermediate, glycan or sugar [α-D-glucose 1, 6-phophate or Glc-1-6-BP, N-Acetyl-D-mannosamine 6-phosphate or mannc 6-P, hyaluronic acid, 6-(β-D-glucosaminyl)-1D-myo-inositol or Glu-MI], or other natural products with capacities for regulating glucose metabolism or inflammation [pyridoxamine, α-L-rhamnopyranosyl-(1-2)-β-D-galactopyranosyl-(1)-β-D-glucopyranoside (RGG), aminomethylphosphonic acid (AMPA) and 8-amino-7-oxononanoate (KAPA)] (data available up on request).
3.2. Effects of WLD on Glucose, Lipids and Insulin Resistance in HF Diet-Fed Mice
The level of fasting plasma glucose (FPG) in mice fed with HF diet supplemented with 26% WLD for 12 weeks was significantly lower than that in mice fed with HF+26% WHR diet (p<0.01, Figure 2A). The levels of total cholesterol and triglycerides in mice treated with WLD or WTWD diet (HF+13% WHR+13% WLD) were significantly lower than that of WHR diet fed mice (p<0.05 or 0.01, Figure 2B). WLD diet and WTWD diet dose-dependently reduced fasting plasma insulin and HOMA-IR compared to WHR diet (Figure 2C and Figure 2D). No significant difference in body weights (Figure 2E) or food intake (data not shown) was detected among mice treated with the different diets.
3.3. Effects of WLD on Circulatory Inflammatory Cytokines and Monocyte Adhesion
WLD diet dose-dependently reduced the levels of pro-inflammatory cytokines, PAI-1, MCP-1 and TNFα in plasma, and ex vivo monocyte adhesion to the intima of mouse aortae compared to that in WHR diet-fed mice (p<0.01, Figure 3A-Figure 3C). WLD diet induced significantly greater inhibition on the pro-inflammatory mediators and monocyte adhesion compared to mice fed with WTWD diet (p<0.01, Figure 3D).
3.4. Influence of WLD on Diversities of Gut Microbiota
No significant difference in the levels of Shannon index, a common variable used for assessing richness and evenness of α-diversity in gut microbiota, was detected in feces from the three groups of mice receiving different dietary interventions (Figure 4A). The plot of principal component analysis demonstrated that the β-diversity in WLD diet-fed mice completely separated from WHR diet-fed mice, and that of WTWD diet-fed mice partially overlapped with that of WHR diet-fed mice as expected (Figure 4B).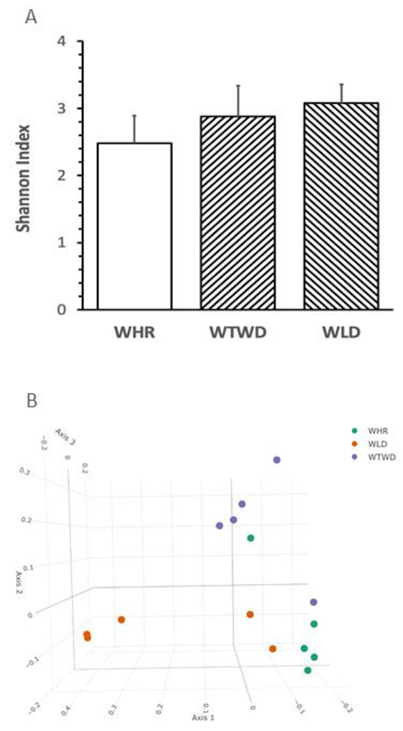
3.5. Influence of WLD on Taxonomic Composition of Gut Microbiota
The major phylum bacteria in feces of all three groups of mice were Firmicutes and Bacteroidetes, but no significance was detected in the relative abundances of the two predominant phylum bacteria in feces of the three groups of mice receiving HF diet supplemented with different types of rice. The abundances of Actinobacteria phylum bacteria in WTWD diet-fed mice were significantly lower than that in WLD diet-fed mice (p<0.01). No significant difference was detected between WHR and WLD or WTWD diet-fed mice. The abundances of fecal Vernrucomicrobia phylum bacteria in WLD diet-fed mice were significantly lower than that in WHR or WTWD diet-fed mice (p<0.05 or 0.001), and that in WTWD diet-fed mice were significantly higher than that in WHR diet-fed mice (p<0.05, Figure 4C). The abundances of fecal Akkermansiaceae family bacteria in WTWD diet-fed mice were significantly higher than WLD diet-fed mice (p<0.01). The abundances of Ruminococcaceae family bacteria in WLD diet-fed mice were significantly higher than that in WHR or WTWD diet-fed mice (p<0.05 or 0.001). The abundances of Ruminococcaceae in WTWD diet-fed mice were lower than WHR diet fed mice (p<0.05, Figure 4D). The abundances of Akkermansia genus bacteria in WLD diet fed mice were significantly lower that in WHR or WTWD diet fed mice (p<0.05 or 0.001). The abundances of fecal Bacteroides and Lactobacillus genus bacteria were significantly higher than that in WHR diet fed mice (p<0.05). The abundances of Lachnospiraceae NK4A136 genus bacteria in WTWD diet-fed mice were significantly lower than that in WHR or WLD diet-fed mice (p<0.05, Figure 4E). The abundances of fecal Lactobacillus gasseri (L. gasseri) species bacteria in WLD-diet fed mice were significantly higher than that in WHR or WTWD diet-fed mice (p<0.001, Figure 4F).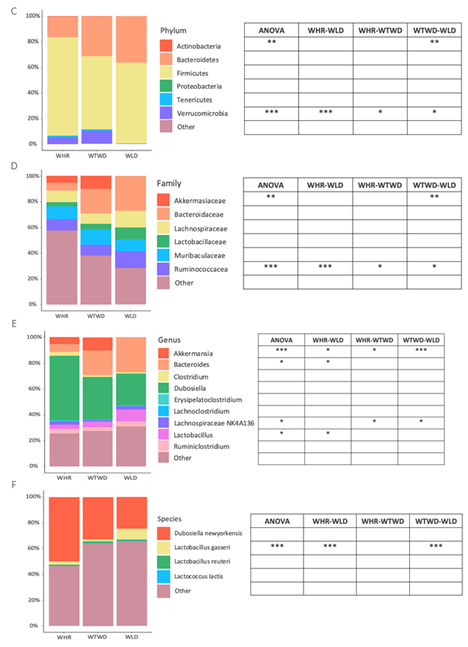
The results of LEfSe analysis demonstrated distinct patterns of gut microbiota in mice received HF+WHR, WLD or WTWD diet. The gut microbiota of HF+WHR diet-fed mice was characterized by the enrichment in Bifidobacterium phylum, Bifidobacteriaceae, Clostridiaceae and Peptococaceae family bacteria. The gut microbiota of HF+WLD diet-fed mice was highlighted by Bacteriodaceae, Lactobacillaceae family bacteria and Bacteroides and Lactobacillus genus bacteria. The gut microbiota of HF+WTWD diet-fed mice was featured by Verruconicrobia phylum, Akkermansiaceae family, Ruminococaceae family and Akkermansia genus bacteria (Figure 4G and Figure 4H).
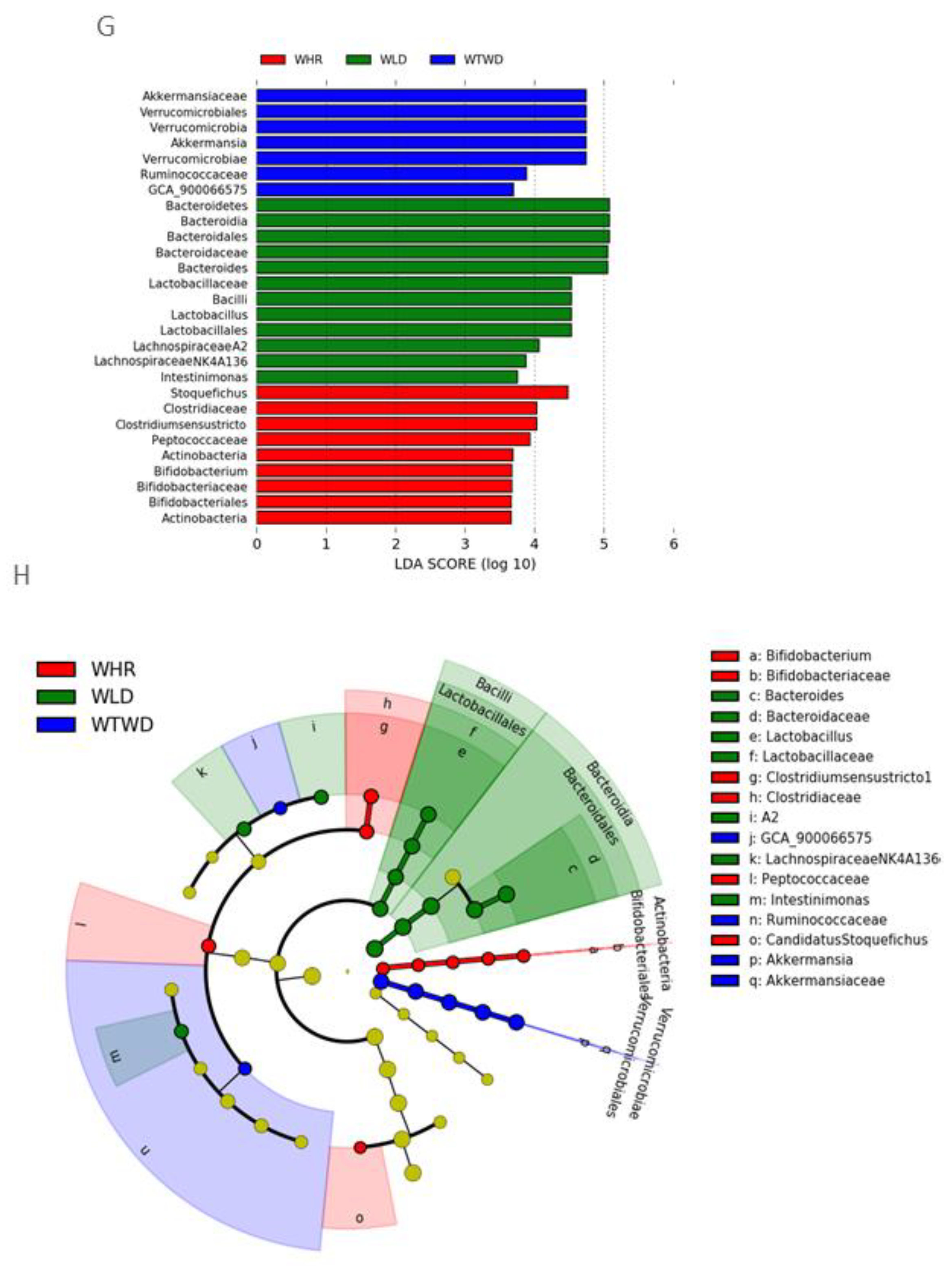
Figure 4.
Effects of WLD and WHR on gut microbiota in HF diet-fed mice. The regimen for the dietary intervention and animal groups was the same as described in the legend of Figure 2. Fecal samples were collected from individually housed mice at the end of dietary intervention. Gut microbiota was analysed using 16S rRNA gene sequencing. A: Shannon index for α-diversity of gut microbiota. Values were expressed in mean±SD (n=5/group); B: Principal component analysis for β-diversity was plotted based on Bray-Curtis dissimilarities between all sample sets (weighted by taxon abundance). C: Heat map for relative abundances of major types of phylum bacteria detected in feces of the mice (right) and results of statistical analysis between different dietary groups (left). D: Relative abundance of major family bacteria and statistical analysis results. E: Relative abundance of major genus bacteria and statistical analysis results. F: Relative abundance of major species bacteria and statistical analysis results. *, **, ***: p<0.05, 0.01 or 0.001 (ANOVA, Tukey test). G and H: liner discriminant analysis effect size of gut microbiota in HF-diet-fed mice supplemented with WLD, WTWD or WHR.
Figure 4.
Effects of WLD and WHR on gut microbiota in HF diet-fed mice. The regimen for the dietary intervention and animal groups was the same as described in the legend of Figure 2. Fecal samples were collected from individually housed mice at the end of dietary intervention. Gut microbiota was analysed using 16S rRNA gene sequencing. A: Shannon index for α-diversity of gut microbiota. Values were expressed in mean±SD (n=5/group); B: Principal component analysis for β-diversity was plotted based on Bray-Curtis dissimilarities between all sample sets (weighted by taxon abundance). C: Heat map for relative abundances of major types of phylum bacteria detected in feces of the mice (right) and results of statistical analysis between different dietary groups (left). D: Relative abundance of major family bacteria and statistical analysis results. E: Relative abundance of major genus bacteria and statistical analysis results. F: Relative abundance of major species bacteria and statistical analysis results. *, **, ***: p<0.05, 0.01 or 0.001 (ANOVA, Tukey test). G and H: liner discriminant analysis effect size of gut microbiota in HF-diet-fed mice supplemented with WLD, WTWD or WHR.
3.6. Impact of WLD on Fecal SCFAs and Correlation with Metabolic or Inflammatory Variables
The abundance of propionic acid in feces of mice receiving WLD diet was significantly higher than that in WHR diet-fed mice (p<0.05, Figure 5B). The abundances of other fecal SCFAs were not significantly different among three groups (Figure 5A, Figure 5C-Figure 5G). The relative abundance of propionic acid negatively correlated with the circulatory levels of FPG, total cholesterol, PAI-1 and MCP-1 in the mice (p<0.05, Figure 6).
3.7. Correlation between SCFAs and Gut Microbiota
The abundance of fecal isobutyric acid positively correlated with fecal Bacteroidetes phylum bacteria (p<0.05). The abundance of fecal Valeric acid positively correlated with fecal Actinobacteria phylum bacteria (p<0.05, Figure 7A). The abundances of fecal butyric acid positively correlated with fecal Muribaculaceae family bacteria (p<0.05). The abundance of fecal hexanoic acid positively correlated with fecal Clostridiaceae family bacteria (p<0.05, Figure 7B). The abundance of fecal hexanoic acid positively correlated with fecal Clostridum genus bacteria (p<0.05, Figure 7C). The abundances of SCFAs did not significantly correlate with detected species bacteria in the mice (Figure 7D).
3.8. Correlation between Fecal and Body Weights, Metabolic or Inflammatory Variables
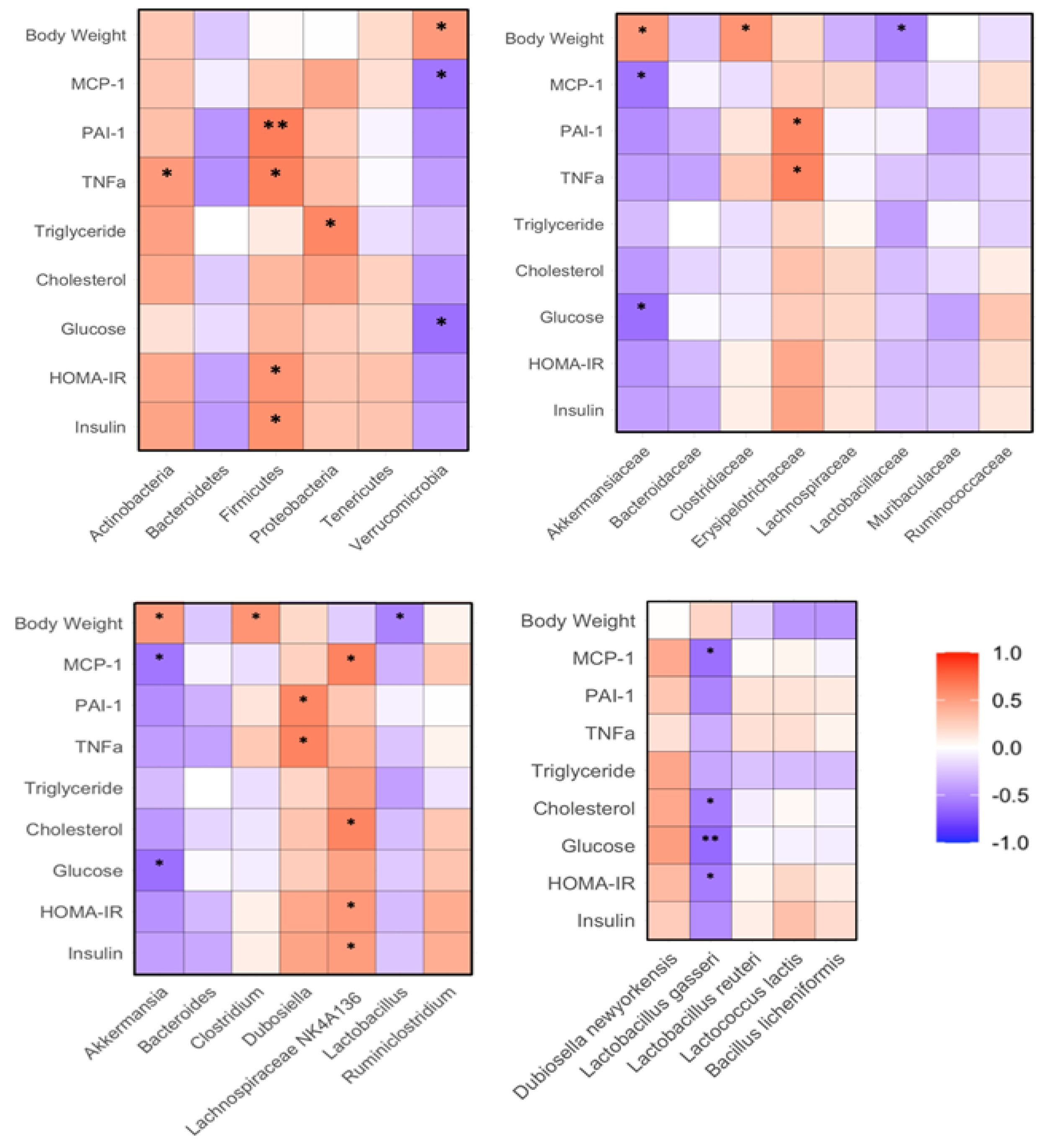
The relative abundances of fecal Firmicutes positively corrected with insulin, HOMA-IR, TNFα and PAI-1 in the mice (p<0.05 or 0.01). The abundances of fecal Verrucomicrobia negatively correlated with FPG and MCP-1, while as positively correlated with body weight of the mice (p<0.05, Figure 8A). The abundances of fecal Akkermansiaceae family bacteria negatively correlated with FPG and MCP-1, but positively correlated with body weights of the mice (p<0.05). The abundances of fecal Erysipelotrichaceae family bacteria positively correlated with PAI-1 and TNFα (p<0.05, Figure 8B). The abundance of fecal Akkermansia genus bacteria negatively correlated with FPG and MCP-1 and positively correlated with body weights of the mice (p<0.05). The abundances of Dubosiella genus bacteria positively correlated with PAI-1 and TNFα levels. The abundance of fecal Lachnosipiraceae NK4A136 genus bacteria positively correlated with insulin, HOMA-IR, cholesterol and MCP-1 (p<0.05, Figure 8C). The abundance of fecal L. gasseri negatively correlated with HOMA-IR, FPG, cholesterol and MCP-1 of the mice (p<0.05, Figure 8D).
4. Discussion
The results of the present study demonstrated that the supplementation of WLD to HF diet dose-dependently attenuated circulatory metabolic and pro-inflammatory markers in mice compared to those receiving HF+WHR diet. The abundances of probiotic L. gasseri species bacteria in the feces of mice fed with HF+WLD diet were significantly greater than that in mice fed with HF+WHR or HF+WTWD diet. The abundance of fecal propionic acid in HF+WLD diet-fed mice was significantly higher than HF+WHR diet-fed mice. The fecal abundances of both L. gasseri and propionic acid negatively correlated with cholesterol, pro-inflammatory cytokines, FPG or HOMA-IR in the mice. The results of metabolomic analysis demonstrated that WLD contained substantially higher abundances of metabolites with capacity to modulate glucose metabolism compared to WHR, including AMP, homoarginine, Glc-1, 6-BP, Glu-MI and L-glutamate.
Previous studies by Moghadasian et al. demonstrated that WLD treatment induced attenuation of atherosclerosis was associated with increased abundances of fecal bacteria, including Lactobacillus, and fatty acids, including butyric acid or longer chain fatty acids, in LDL receptor-deficient mice [19]. Hou et al reported that Asian wild rice (Zizania latifolia) intake reduced liver steatosis, insulin resistance and gut dybiosis in HF diet-fed mice [20]. The present study for the first time examined the effects of WLD on SCFAs including acetic acid and propionate acid and relationship between SCFA and gut microbiome in HF diet-fed mice. The findings of the present study demonstrated that WLD diet significantly increased the abundances of fecal propionic acid, and the relative abundance of fecal propionic acid negatively correlated with metabolic and inflammatory markers in HF-diet induced insulin resistant mice.
The results of LEfSe analysis indicated the feces of WLD diet-fed mice was enriched with Lactobacillaceae family and Lactobarcillus genus bacteria. The abundances of L. gasseri species bacteria in the feces of mice fed with WLD diet were significantly higher than that in mice receiving WHR or WTWD diet. Although the negative correlation between the abundances of fecal L. gasseri and propionate acid did not reach a significant level, but both L. gasseri and propionate acid negatively correlated to FPG, cholesterol and MCP-1 in the mice. Several species of Lactobacillus, including L. gasseri, have been recognized as potent probiotics for the management of diabetes and obesity in clinical trials [21,22]. The findings of the present study suggest that the increases of the abundance of L. gasseri in the gut may contribute to metabolic benefits and anti-inflammatory effects of WLD. Relationship between L. gasseri and the production of propionate acid is warrant to be investigated.
In addition, the abundances of fecal Bacteroides genus bacteria and Ruminococcaceae family bacteria in WLD diet-fed mice were significantly higher than that in WHR diet-fed mice. The abundances of fecal isobutyric acid positively correlated with Bacteroidetes phylum bacteria in the mice, but there was no detectable correlation between fecal isobutyric acid and metabolic or inflammatory variables in mice.
Previous studies demonstrated that fiber-rich diet reduced FPG and insulin resistance in T2D patients, which was associated with enhancement of a variety of gut bacteria capable to generate SCFAs and modulate glucose metabolism [23,24]. Our group previously reported that fiber-rich brown rice and germinated brown rice attenuated FPG, lipids and inflammatory markers, in HF diet-fed mice [13]. WLD contains 3-times higher level of fiber than that in WHR [25,26]. The precise role of fiber in the beneficial effect of WLD diet on metabolism, inflammation, gut microbiota and SCFA production are warrant to be further clarified in subsequent experiments.
WLD contains near 20-fold higher levels of AMP compared to WHR. AMP is a known agonist of AMP-activated protein kinase (AMPK). AMPK plays crucial modulatory roles in glucose, lipid metabolism, insulin resistance, and inflammation [27]. Previous study demonstrated that the colonization of Lactobacillus rhamnosus GG upregulated the phosphorylated AMPK in colon of mice [28]. Propionic acid reduced gluconeogenesis in HepG2 hepatocytes via the activation of AMPK pathway [29]. Propionic acid inhibited insulin induced de novo lipogenesis and increased glucose uptake in primary rat adipocytes [30]. Treatment of human adipose tissue explants with propionic acid resulted in downregulations of inflammatory cytokines and upregulations of lipoprotein lipase and glycose transporter [31]. Besides, WLD also contains abundant amounts of L-glutamate [32], AMPA [33], KAPA [34,35], homoarginine [36,37], and Glc-1, 6-BP [38,39] which are involved in the regulation of glucose metabolism or insulin resistance, pyridoxamine or vitamin 6 [40,41], RGG [42] and hyaluronic acid [43], which were known to benefit in the management of diabetes, inflammation or diabetic complications. Those metabolites enriched in WLD may partially contribute to the anti-diabetic and anti-inflammatory benefits of WLD in HF diet induced insulin resistant mice.
The anti-insulin resistance and anti-inflammatory effects of WLD in HF diet-fed mice were consistent with previous reports [10]. The present study added a half-dosage of WLD (WTWD) diet in the dietary regimen to determine the effectiveness of a lower and more feasible dosage of WLD on metabolism, inflammation, gut microbiota and SCFA production in mice. The results demonstrated that the WTWD diet, containing half-dosage of WLD compared to the WLD diet, significantly reduced triglycerides, total cholesterol, insulin resistance, pro-inflammatory cytokines and monocyte adhesion compared to WHR diet. The metabolic and anti-inflammatory effects of the WTWD diet were relatively weaker than the WLD diet as expected. The results suggest that weaker, but effective, metabolic and anti-inflammatory benefits may be achieved using a lower dosage of WLD (representing 10% of daily calorie intake) supplemented in HF diet in mice, which may be useful for future regimen design in human trials.
The limitation of the present study includes that the effects of WLD on the metabolism, inflammation and gut microbiota were only assessed in male, but not in female, mice. Future investigations may be required to compare the effects of WLD in female animals. The present study did not investigate the biological activities of compounds enriched in WLD. The effect and regimen for using WLD in diabetic patients need to be verified in randomized controlled clinical trials.
In conclusion, WLD supplementation in HF diet reduced FPG, lipids, insulin, HOMA-IR, monocyte adhesion and inflammatory cytokines related to monocyte adhesion compared to that in mice receiving WHR diet, which was associated with increased abundances of probiotic bacteria, L. gasseri, and propionic acid in feces of mice receiving WLD diet. The abundance of fecal L. gasseri and propionic acid negatively correlated with the metabolic and pro-inflammatory cytokines in the peripheral circulation of the mice. Multiple compounds enriched in WLD may independently modulate glucose metabolism and inflammation. The findings suggest that oral administration of WLD may improve metabolism and mitigate chronic inflammation in mice through multiple pathways, including, but not limited to, the modulation of gut microbiota, SCFA production and AMPK activation in high fat diet fed mice.
Conflicts of Interests
Authors have no conflict interest to the contents of the manuscript.
Acknowledgments
The authors appreciate the support from operating grant from Diabetes Canada (OG-3-15-4889-GS).
Abbreviations Used
AMP: adenosine monophosphate.
AMPA: aminomethylphosphonic acid.
AMPK: AMP-activated protein kinase.
ANOVA: analysis of variance assay.
ELISA: enzyme-linked immunosorbent assay.
FPG: fasting plasma glucose.
G1c-1: 6-BP: α-D-Glucose 1, 6-biphosphate.
GlcNAc-1P: N-Acetyl-alpha-D-glucosamine 1-phosphate.
Glu-MI: 6-(alpha-D-Glucosaminyl)-1D-myo-inositol.
HBSS: Hank’s balance salt solution.
HF: high fat (diet).
HOMA-IR: Homeostatic model assessment-insulin resistance.
KAPA: 8-amino-7-oxononanoate.
MCP-1: monocyte chemotactic protein-1.
PAI-1: plasminogen activator inhibitor-1.
RGG: α-L-rhamnopyranosyl-(1-2)-β-D-galactopyranosyl-(1)-β-D-glucopyranoside.
SCFA: short chain fatty acid.
TNFα: tumor necrosis factor-α.
WHR: white rice.
WLD: wild rice.
WTWD: 13g % of WHR and 13g % of WLD supplemented in HF diet.
References
- Stack Whitney, K. Manoomin: The Taming of Wild Rice in the Great Lakes Region. Environment & Society Portal, Arcadia, no.2. Rachel Carson Center for Environment and Society. [CrossRef]
- Kellogg, E.A. The Evolutionary History of Ehrhartoideae, Oryzeae, and Oryza. Rice 2009, 2, 1–14. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture. Wild rice, cooked. https://fdc.nal.usda.gov/fdc-app.html#/food-details/168897/nutrients. Accessed May 10, 2024.
- Surendiran, G. , Goh, C., Le, K., Zhao, Z., Askarian, F., Othman, R., Nicholson, T., Moghadasian, P., Wang, Y.J., Aliani. M, Shen, G.X., Beta, T., Moghadasian, M.H. Wild rice (Zizania palustris L.) prevents atherogenesis in LDL receptor knockout mice. Atherosclerosis 2013, 230, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Moghadasian, M.H. , Alsaif, M., Le, K., Gangadaran, S., Masisi, K., Beta, T., Shen, G.X. (2016). Combination effects of wild rice and phytosterols on prevention of atherosclerosis in LDL receptor knockout mice. J Nutr Bioch 2016, 33, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Moghadasian, M.H. , Zhao, R., Ghazawwi, N., Le, K., Apea-Bah, F.B., Beta, T., Shen, G.X. Inhibitory Effects of North American Wild Rice on Monocyte Adhesion and Inflammatory Modulators in Low-Density Lipoprotein Receptor-Knockout Mice. J Agric Food Chem 2017, 65, 9054–9060. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.A. , Pan, A., Malik, V., Sun, Q. White rice consumption and risk of type 2 diabetes: meta-analysis and systematic review. Br Med J 2012, 344, e1454. [Google Scholar] [CrossRef] [PubMed]
- Jannasch, F. , Kröger, J., Schulze, M.B. Dietary Patterns and Type 2 Diabetes: A Systematic Literature Review and Meta-Analysis of Prospective Studies. J Nutr 2017, 147, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E. , Herrero, L., Naaz, A. Obesity, inflammation, and insulin resistance. Gastroenterology 2007, 132, 2169–2180. [Google Scholar] [CrossRef]
- Zhao, R. , Wan, P., Shariati-Ievari, S., Michel, A., Shen, G.X. North American Wild Rice Attenuated Hyperglycemia in High Fat-Induced Obese Mice: Involvement of AMP-Activated Protein Kinase. J Agri Food Chem 2020, 68, 8855–8862. [Google Scholar] [CrossRef]
- Netto Candido, T.L. , Bressan, J., Alfenas, R.C.G. Dysbiosis and metabolic endotoxemia induced by high-fat diet. Nutr Hosp 2018, 35, 1432–1440. [Google Scholar]
- Turnbaugh, P.J. , Backhed, F., Fulton, L., Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, N. , Gasbarrini, A., Ponziani, F.R. Intestinal Barrier in Human Health and Disease. Int J Environ Res Public Health 2021, 18, 12836. [Google Scholar] [CrossRef] [PubMed]
- He, J. , Zhang, P., Shen, L., Niu, L., Tan, Y., Chen, L., Zhao, Y., Bai, L., Hao, X., Li, X., Zhang, S., Zhu, L. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int J Mol S 21, 6356. [CrossRef] [PubMed]
- Zhao, R. , Ren, S., Moghadasain, M.H., Rempel, J.D., Shen, G.X. Involvement of Fibrinolytic Regulators in Adhesion of Monocytes to Vascular Endothelial Cells Induced by Glycated LDL and to Aorta from Diabetic Mice. J Leukocy Biol 2014, 95, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R. , Le, K., Li, W., Ren, S., Moghadasian, M.H., Beta, T., Shen, G.X. Effects of Saskatoon Berry Powder on Monocyte Adhesion to Vascular Wall of Leptin Receptor-Deficient Diabetic Mice. J Nutr Biochem 2014, 25, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Huang, F. , Zhao, R., Xia, M., Shen, G.X. Impact of Cyanidin-3-Glucoside on Gut Microbiota and Relationship with Metabolism and Inflammation in High Fat-High Sucrose Diet-Induced Insulin Resistant Mice. Microorganisms 2020, 8, 1238. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X. , Qiu, Y., Zhong, W., Baxter, S., Su, M., Li, Q., Xie, G., Ore, B.M., Qiao, S., Spencer, M.D., Zeisel, S.H., Zhou, Z., Zhao, A., Jia, W.A. A targeted metabolomic protocol for short-chain fatty acids and branched-chain amino acids. Metabolomics 2013, 9, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Moghadasian, M.H. , Kaur, R., Kostal, K., Joshi, A.A., Molaei, M., Le, K., Fischer, G., Bonomini, F., Favero, G., Rezzani, R., Gregorchuk, B.S.J., Leung-Shing, V., Wuzinski, M., Seo, A.I., Bay, D.C. Anti-Atherosclerotic Properties of Wild Rice in Low-Density Lipoprotein Receptor Knockout Mice: The Gut Microbiome, Cytokines, and Metabolomics Study. Nutrients 2019, 11, 2894. [Google Scholar] [PubMed]
- Hou, X.D. , Yan, N., Du, Y.M., Liang, H., Zhang, Z.F., Yuan, X.L. Consumption of Wild Rice (Zizania latifolia) Prevents Metabolic Associated Fatty Liver Disease through the Modulation of the Gut Microbiota in Mice Model. Int J Mol Sci 2020, 21, 5375. [Google Scholar] [CrossRef]
- Łagowska, K. , Malinowska, A.M., Zawieja, B., Zawieja, E. Improvement of glucose metabolism in pregnant women through probiotic supplementation depends on gestational diabetes status: meta-analysis. Sci Rep 2020, 10, 17796. [Google Scholar] [CrossRef]
- Álvarez-Arraño, V. , Martín-Peláez, S. Effects of Probiotics and Synbiotics on Weight Loss in Subjects with Overweight or Obesity: A Systematic Review. Nutrients 2021, 13, 3627. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L. , Zhang, F., Ding, X., Wu, G., Lam, Y.Y., Wang, X., Fu, H., Xue, X., Lu, C., Ma, J., Yu, L., Xu, C., Ren, Z., Xu, Y., Xu, S., Shen, H., Zhu X, Shi, Y., Shen, Q., Dong, W., Liu, R., Ling, Y., Zeng, Y., Wang, X., Zhang, Q., Wang, J., Wang, L., Wu, Y., Zeng, B., Wei, H., Zhang, M., Peng, Y., Zhang, C. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [PubMed]
- Fu, J. , Xu, K., Ni, X., Li, X., Zhu, X., Xu, W. Habitual Dietary Fiber Intake, Fecal Microbiota, and Hemoglobin A1c Level in Chinese Patients with Type 2 Diabetes. Nutrients 2022, 14, 1003. [Google Scholar] [CrossRef]
- Lazyplant. How much fiber in brown, white & wild rice? https://lazyplant.com/fiber-rice/. Accessed May 3, 2024.
- Dr. Axe. Is wild rice is the best rice of all? Learn how to cook. https://draxe.com/nutrition/wild-rice/. Accessed May 3, 2024.
- Mohseni, R. , Teimouri, M., Safaei, M., Arab Sadeghabadi Z. AMP-activated protein kinase is a key regulator of obesity-associated factors. Cell Biochem Funct 2023, 41, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, T. , Song, X., An, Y., Wu, X., Zhang, W., Li, J., Sun, Y., Jin, G., Liu, X., Guo, Z., Wang, B., Lei, P., Cao, H. Lactobacillus rhamnosus GG Colonization in Early Life Ameliorates Inflammaging of Offspring by Activating SIRT1/AMPK/PGC-1α Pathway. Oxid Med Cell Longev 2021, 2021, 3328505. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H. , Ishii, M., Akagawa, M. Propionate suppresses hepatic gluconeogenesis via GPR43/AMPK signaling pathway. Arch Biochem Biophys 2019, 672, 108057. [Google Scholar] [CrossRef] [PubMed]
- Heimann, E. , Nyman, M., Degerman, E. Propionic acid and butyric acid inhibit lipolysis and de novo lipogenesis and increase insulin-stimulated glucose uptake in primary rat adipocytes. Adipocyte 2014, 4, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Al-Lahham, S. , Roelofsen, H., Rezaee, F., Weening, D., Hoek, A., Vonk, R., Venema, K. Propionic acid affects immune status and metabolism in adipose tissue from overweight subjects. Eur J Clin Invest 2012, 42, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Barnes, T. , Di, K. M., Filip Vlavcheski, Quadrilatero, J., Evangelia Litsa Tsiani, Mourtzakis, M. Glutamate increases glucose uptake in L6 myotubes in a concentration- and time-dependent manner that is mediated by AMPK. Applied Physiology, Nutrition and Metabolism/Applied Physiology, Nutrition, and Metabolism 2018, 43, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Sadanandan Jayanarayanan, Anju, T. R., Soman Smijin, Cheramadathikudiyil Skaria Paulose. Vitamin D3 supplementation increases insulin level by regulating altered IP3 and AMPA receptor expression in the pancreatic islets of streptozotocin-induced diabetic rat. ˜the œJournal of Nutritional Biochemistry 2015, 26, 1041–1049.
- Fan, S. , De Feng Li, Da Cheng Wang, Fleming, J., Zhang, H., Zhou, Y., Zhou, L., Zhou, J., Chen, T., Chen, G., Xian En Zhang, Bi, L. Structure and function of Mycobacterium smegmatis 7-keto-8-aminopelargonic acid (KAPA) synthase. Int J Biochem Cell Biol 2015, 58, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Romero-Navarro, G. , Cabrera-Valladares, G., German, M. S., Matschinsky, F. M., Velazquez, A., Wang, J., & Fernandez-Mejia, C. Biotin Regulation of Pancreatic Glucokinase and Insulin in Primary Cultured Rat Islets and in Biotin- Deficient Rats1. Endocrinology 2019, 140, 4595–4600. [Google Scholar]
- Henningsson, R. , Lundquist, I. Arginine-induced insulin release is decreased and glucagon increased in parallel with islet NO production. Am Physiol-Endocr aMetabolism 1998, 275, E500–E506. [Google Scholar]
- Pilz, S. , Meinitzer, A., Gaksch, M., Grübler, M., Verheyen, N., Drechsler, C., Hartaigh, B. ó, Lang, F., Alesutan, I., Voelkl, J., März, W., Tomaschitz, A. Homoarginine in the renal and cardiovascular systems. Amino Acids 2015, 47, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Katz, A. , Bogardus, C. Relationship between carbohydrate oxidation and G-1,6-P2 in human skeletal muscle during euglycemic hyperinsulinemia. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology/American Journal of Physiology. Regulatory, Integrative, and Comparative Physiology 1991, 260, R113–R119. [Google Scholar] [CrossRef] [PubMed]
- Katz, A. , Bogardus, C. Insulin-mediated increase in glucose 1,6-bisphosphate is attenuated in skeletal muscle of insulin-resistant man. Metabolism, 1990, 39, 1300–1304. [Google Scholar] [CrossRef] [PubMed]
- Maessen, D. E. , Brouwers, O., Gaens, K. H., Wouters, K., Cleutjens, J. P., Janssen, B. J., Miyata, T., Stehouwer, C. D., Schalkwijk, C. G. Delayed Intervention With Pyridoxamine Improves Metabolic Function and Prevents Adipose Tissue Inflammation and Insulin Resistance in High-Fat Diet–Induced Obese Mice. Diabetes 2015, 65, 956–966. [Google Scholar] [PubMed]
- Dakshinamurti, K. Vitamins and their derivatives in the prevention and treatment of metabolic syndrome diseases (diabetes). Can J Physiol and Pharmac. 2015, 93, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Salib, J. , Michael, H., Eskande, E. Anti-diabetic properties of flavonoid compounds isolated from Hyphaene thebaica epicarp on alloxan induced diabetic rats. Pharmacognosy Research 2013, 5, 22. [Google Scholar] [CrossRef]
- Litwiniuk, M. , Krejner, A., Speyrer, M.S., Gauto, A.R., Grzela, T. Hyaluronic Acid in Inflammation and Tissue Regeneration. Hyaluronic Acid in Inflammation and Tissue Regeneration. Wounds 2016, 28, 78–88. [Google Scholar] [PubMed]
Figure 1.
Major metabolites in Northern wild rice (WLD) compared to white rice (WHR). Fold increase of metabolites related to glucose metabolism and diabetes in WLD versus WHR detected by HPLC-MS. AMP: adenosine 5-monophosphate; Glc-1-6-BP :α-D-glucose 1; 6-phophate; mannc 6-P: N-Acetyl-D-mannosamine 6-phosphate; Glu-MI: 6-(β-D-glucosaminyl)-1D-myo-inositol; RGG: α-L-rhamnopyranosyl-(1-2)-β-D-galactopyranosyl-(1)-β-D-glucopyranoside ; AMPA: aminomethylphosphonic acid; KAPA: 8-amino-7-oxononanoate.
Figure 1.
Major metabolites in Northern wild rice (WLD) compared to white rice (WHR). Fold increase of metabolites related to glucose metabolism and diabetes in WLD versus WHR detected by HPLC-MS. AMP: adenosine 5-monophosphate; Glc-1-6-BP :α-D-glucose 1; 6-phophate; mannc 6-P: N-Acetyl-D-mannosamine 6-phosphate; Glu-MI: 6-(β-D-glucosaminyl)-1D-myo-inositol; RGG: α-L-rhamnopyranosyl-(1-2)-β-D-galactopyranosyl-(1)-β-D-glucopyranoside ; AMPA: aminomethylphosphonic acid; KAPA: 8-amino-7-oxononanoate.
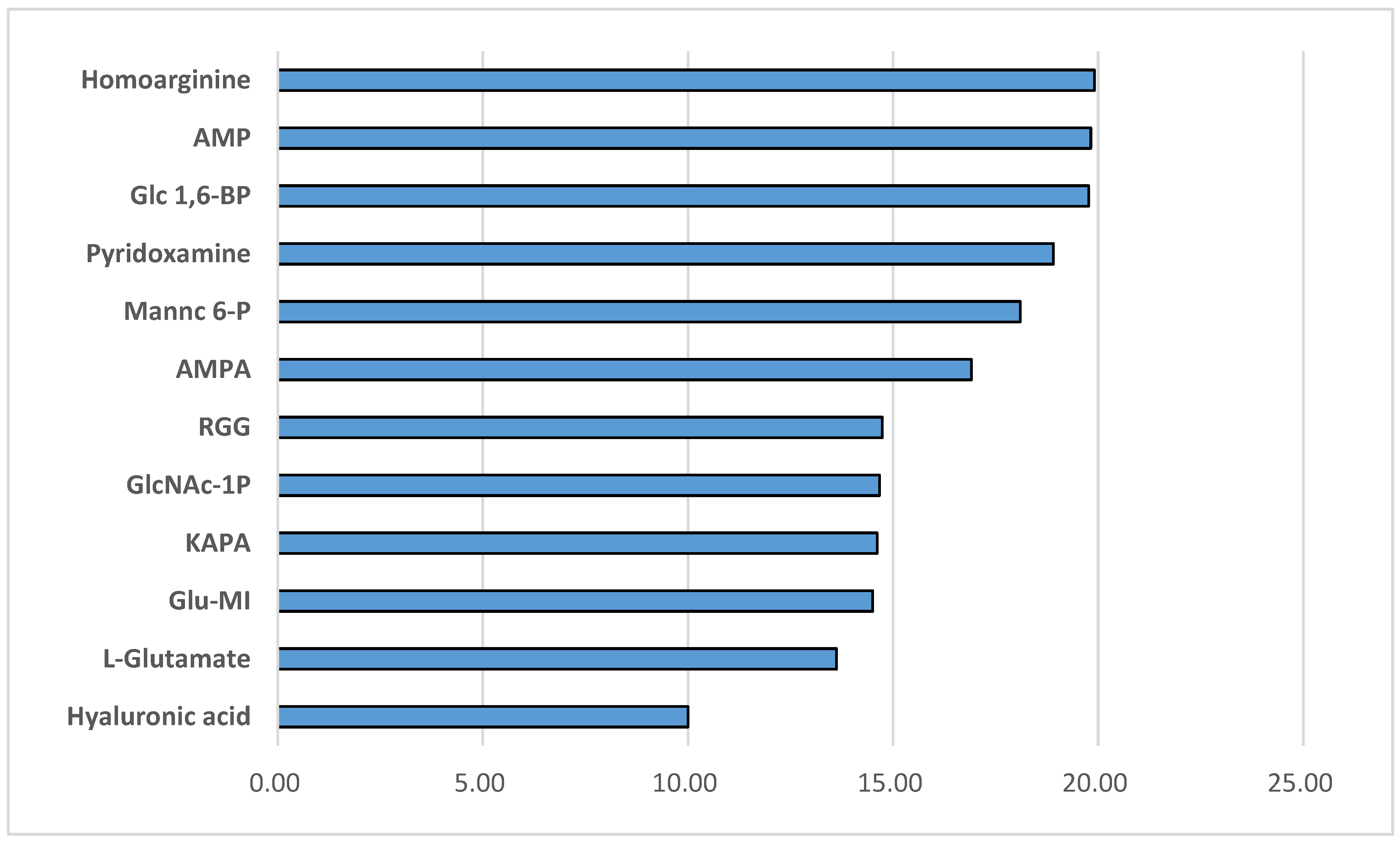
Figure 2.
Effects of WLD and WHR supplementation on metabolism in mice receiving high fat (HF) diet. C57 BL/6J mice (male, 6 weeks of age) were fed with HF diet supplemented with 26 g% (weight/weight) of WHR, 26g% of WLD or 13 g% of WHR+13 g% of WLD (WTWD) for 12 weeks. Blood samples were withdrawn via saphenous vein at onset after an overnight fasting for biochemical analysis. The levels of glucose, total cholesterol, triglycerides and insulin were measured in plasma of mice after 12 weeks of intervention following a fasting. Body weights were measured at one day before tissue harvesting. A: fasting plasma glucose. B: total cholesterol and triglycerides. C: insulin; D: homeostatic model assessment-insulin resistance (HOMA-IR); E: body weight. The values were expressed in mean±SD mg/dL (n=5/group). **: p<0.01 versus WHR group (ANAVA, Tukey test); +: p<0.05 versus WTWD group.
Figure 2.
Effects of WLD and WHR supplementation on metabolism in mice receiving high fat (HF) diet. C57 BL/6J mice (male, 6 weeks of age) were fed with HF diet supplemented with 26 g% (weight/weight) of WHR, 26g% of WLD or 13 g% of WHR+13 g% of WLD (WTWD) for 12 weeks. Blood samples were withdrawn via saphenous vein at onset after an overnight fasting for biochemical analysis. The levels of glucose, total cholesterol, triglycerides and insulin were measured in plasma of mice after 12 weeks of intervention following a fasting. Body weights were measured at one day before tissue harvesting. A: fasting plasma glucose. B: total cholesterol and triglycerides. C: insulin; D: homeostatic model assessment-insulin resistance (HOMA-IR); E: body weight. The values were expressed in mean±SD mg/dL (n=5/group). **: p<0.01 versus WHR group (ANAVA, Tukey test); +: p<0.05 versus WTWD group.
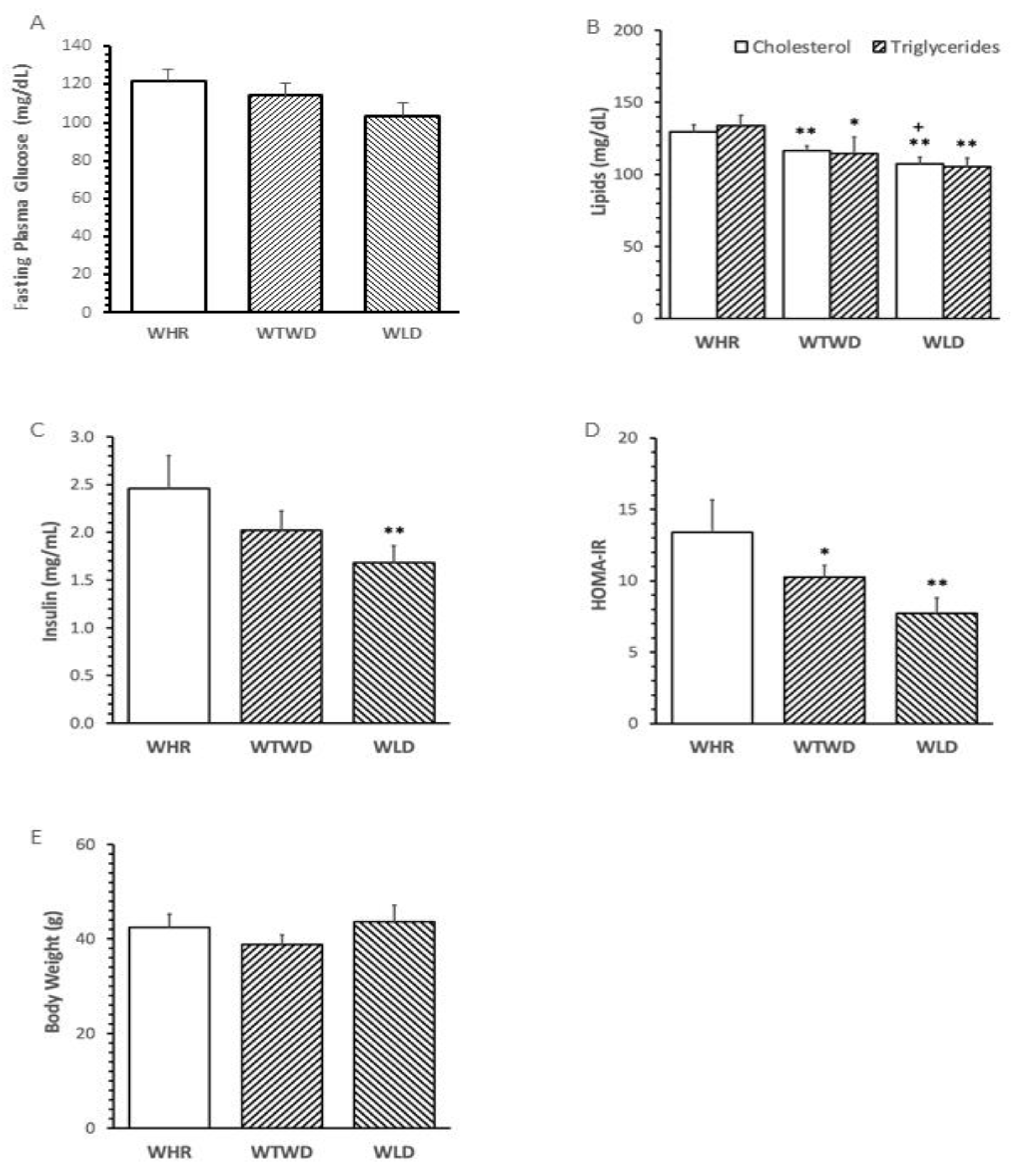
Figure 3.
Effects of WLD and WHR on inflammatory cytokines and monocyte adhesion in HF diet-fed mice. The regimen for the dietary intervention and animal groups was the same as described in the legend of Figure 2. Plasma was isolated from blood withdrawn from mice at the end of dietary intervention. A-C: The levels of tissue necrosis factor-α (TNFα), plasminogen activator inhibitor-1 (PAI-1), monocyte chemotactic protein-1 (MCP-1) and were measured using ELISA. Values were expressed in mean±SD pg/mL, ng/mL or cells/field (n=5/group). D: Monocyte adhesion to aorta was assessed as described in the Methods. **: p<0.01 versus WHR group; +, ++: p<0.05 or 0.01 versus WTWD group (ANOVA, Tukey test).
Figure 3.
Effects of WLD and WHR on inflammatory cytokines and monocyte adhesion in HF diet-fed mice. The regimen for the dietary intervention and animal groups was the same as described in the legend of Figure 2. Plasma was isolated from blood withdrawn from mice at the end of dietary intervention. A-C: The levels of tissue necrosis factor-α (TNFα), plasminogen activator inhibitor-1 (PAI-1), monocyte chemotactic protein-1 (MCP-1) and were measured using ELISA. Values were expressed in mean±SD pg/mL, ng/mL or cells/field (n=5/group). D: Monocyte adhesion to aorta was assessed as described in the Methods. **: p<0.01 versus WHR group; +, ++: p<0.05 or 0.01 versus WTWD group (ANOVA, Tukey test).
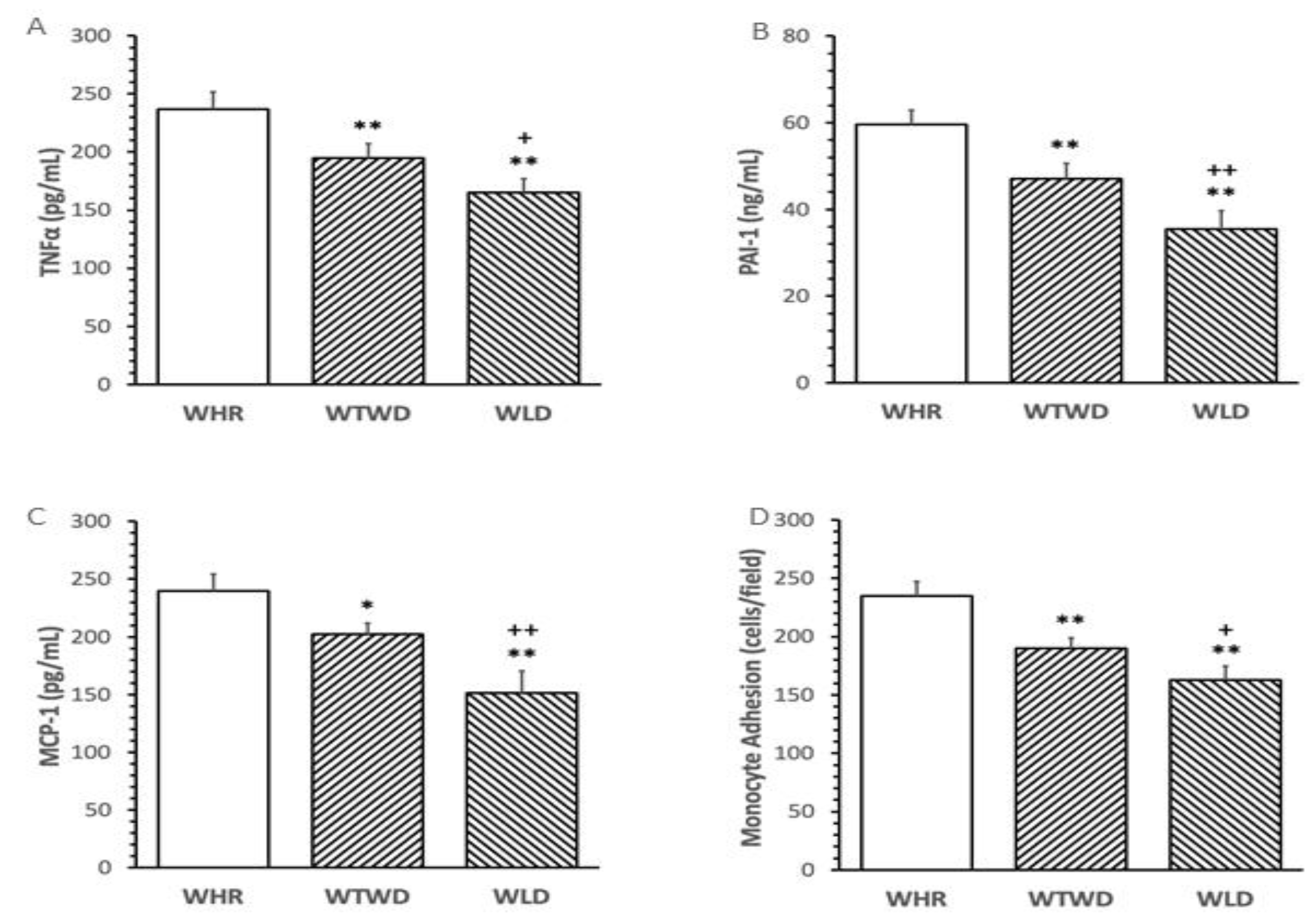
Figure 5.
Effects of WLD and WHR on relative abundances of fecal short chain fatty acids (SCFAs) in HF diet-fed mice. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. SCFAs were analysed using GC-MS as described in the Methods. A: acetic acid; B: propionic acid; C: isobutyric acid; D. butyric acid; E: isovaleric acid; F. valeric acid; G: hexanoic acid. Values were expressed in mean±SD (n=5/group). *: p<0.05 versus WHR group (ANOVA, Tukey test).
Figure 5.
Effects of WLD and WHR on relative abundances of fecal short chain fatty acids (SCFAs) in HF diet-fed mice. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. SCFAs were analysed using GC-MS as described in the Methods. A: acetic acid; B: propionic acid; C: isobutyric acid; D. butyric acid; E: isovaleric acid; F. valeric acid; G: hexanoic acid. Values were expressed in mean±SD (n=5/group). *: p<0.05 versus WHR group (ANOVA, Tukey test).
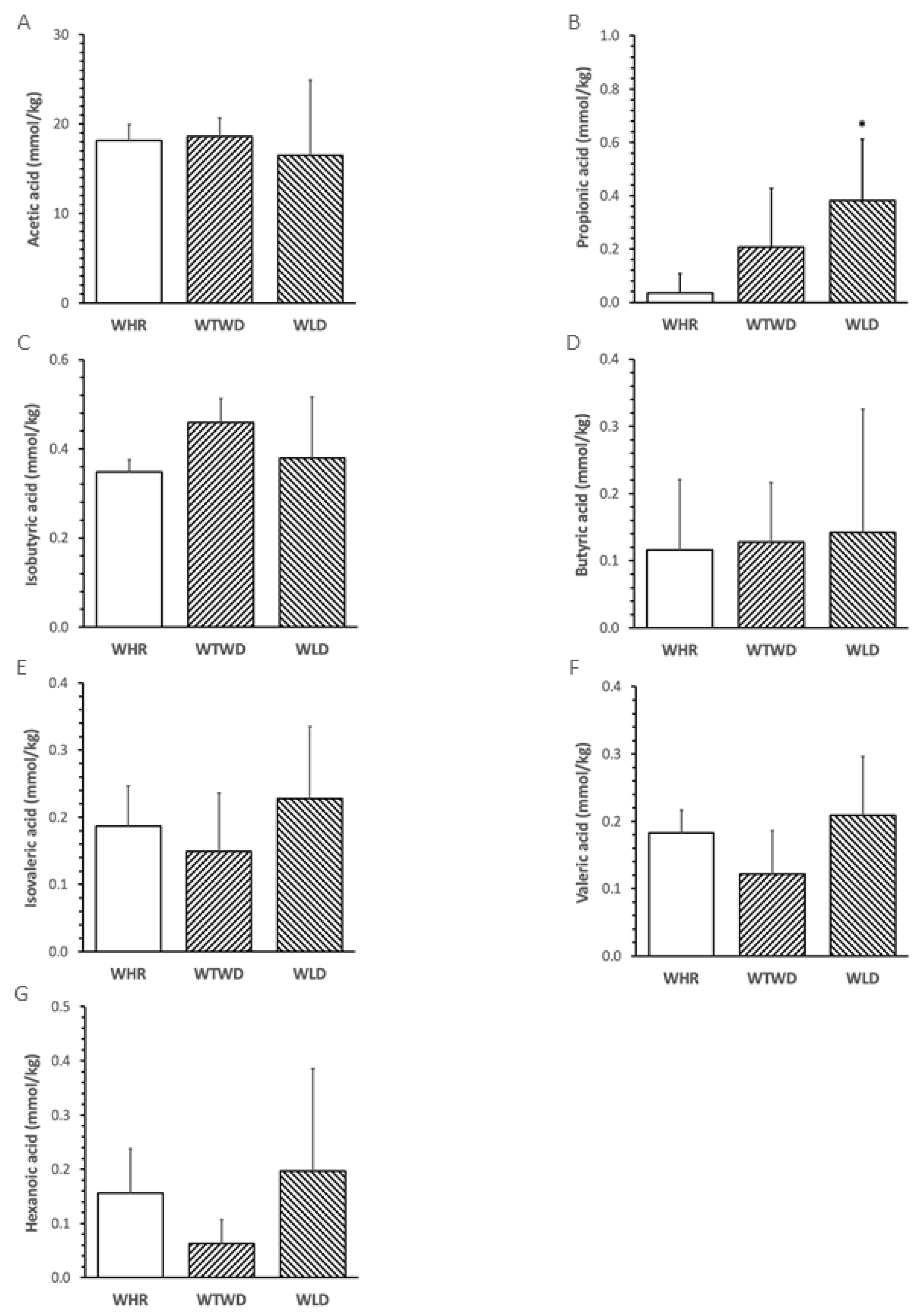
Figure 6.
Correlation between SCFAs and metabolic or inflammatory variables in mice received HF diet supplemented with WLD or WHR. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. SCFA measurement were conducted as that described in the Methods. *: p<0.05 between SCFA and biochemical variable using linear regression analysis (n=15).
Figure 6.
Correlation between SCFAs and metabolic or inflammatory variables in mice received HF diet supplemented with WLD or WHR. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. SCFA measurement were conducted as that described in the Methods. *: p<0.05 between SCFA and biochemical variable using linear regression analysis (n=15).
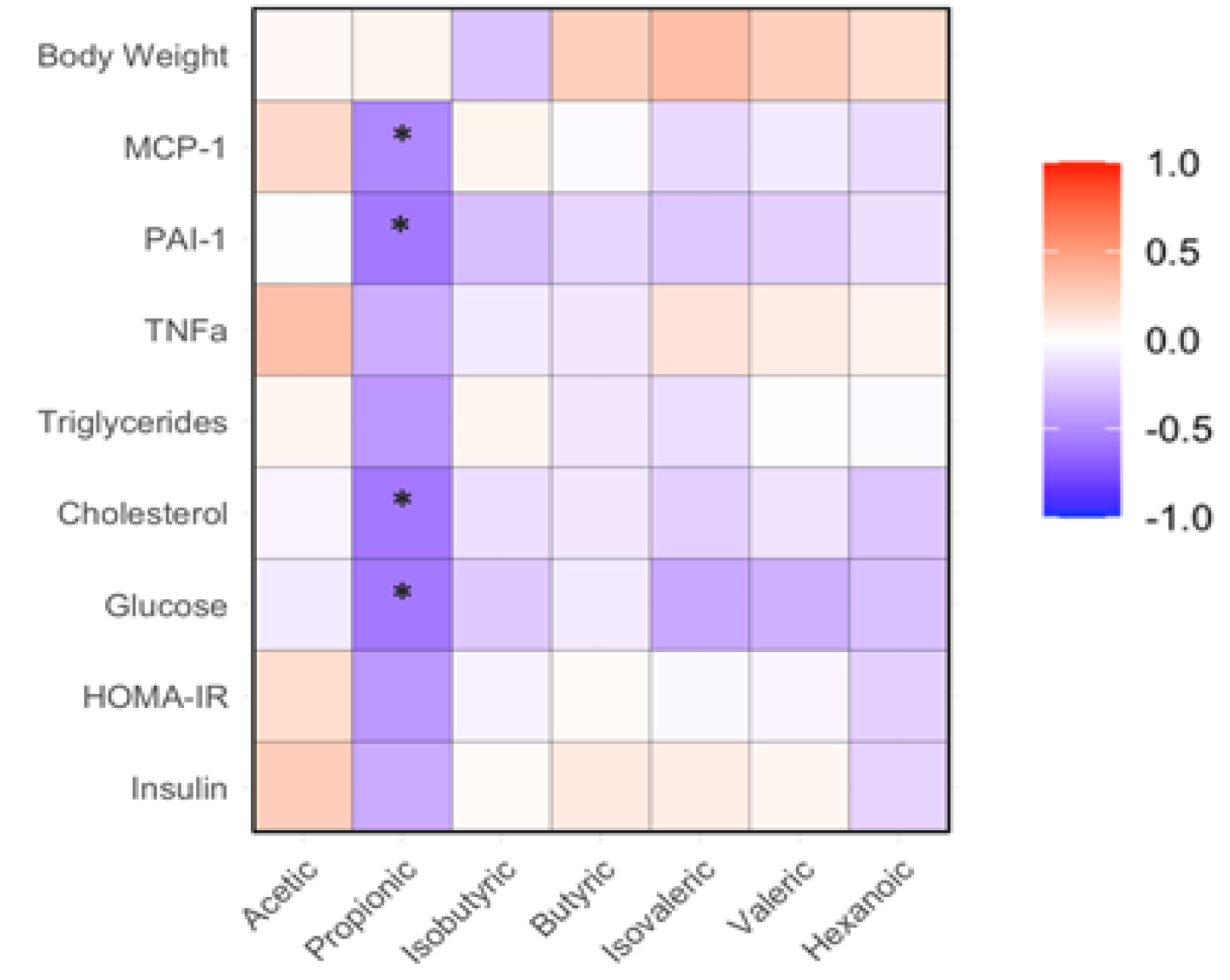
Figure 7.
Correlation between fecal SCFAs and gut microbiota in HF diet-fed mice. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. Bacterial gene sequencing and SCFAs analysis were conducted as described in the Methods. A: correlation between SCFAs and phylum bacteria; B: correlation between SCFAs and family bacteria; C: correlation between SCFAs and genus bacteria; D: correlation between SCFAs and species bacteria. *: p<0.05 between SCFA and biochemical variable using linear regression analysis (n=15).
Figure 7.
Correlation between fecal SCFAs and gut microbiota in HF diet-fed mice. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. Bacterial gene sequencing and SCFAs analysis were conducted as described in the Methods. A: correlation between SCFAs and phylum bacteria; B: correlation between SCFAs and family bacteria; C: correlation between SCFAs and genus bacteria; D: correlation between SCFAs and species bacteria. *: p<0.05 between SCFA and biochemical variable using linear regression analysis (n=15).
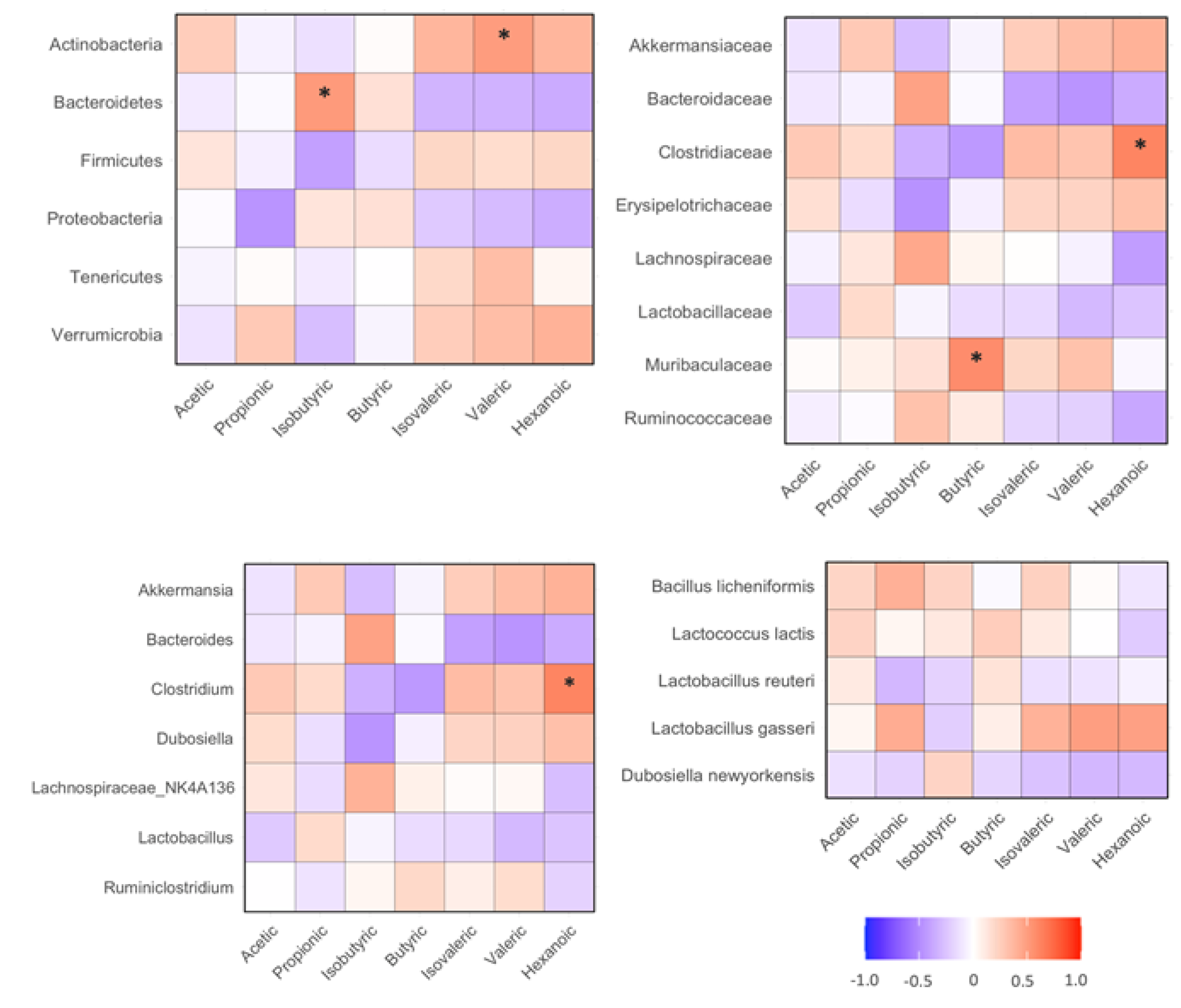
Figure 8.
Correlation between gut microbiota and biochemical variables in HF diet-fed mice. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. Bacterial gene sequencing, biochemical analysis and body weights were conducted as described in the Methods. A: Correlation between phylum bacteria, biochemical variables or body weights; B: correlation between family bacteria, biochemical variables or body weights; C: correlation between genus bacteria, biochemical variables or body weights; D: correlation between species bacteria, biochemical variables or body weights. *, **: p<0.05 or 0.01 between gut bacteria and biochemical variable or body weight using linear regression analysis (n=15).
Figure 8.
Correlation between gut microbiota and biochemical variables in HF diet-fed mice. The regimen of dietary intervention and animal groups was the same as described in the legend of Figure 2. Bacterial gene sequencing, biochemical analysis and body weights were conducted as described in the Methods. A: Correlation between phylum bacteria, biochemical variables or body weights; B: correlation between family bacteria, biochemical variables or body weights; C: correlation between genus bacteria, biochemical variables or body weights; D: correlation between species bacteria, biochemical variables or body weights. *, **: p<0.05 or 0.01 between gut bacteria and biochemical variable or body weight using linear regression analysis (n=15).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



