
Submitted:
30 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
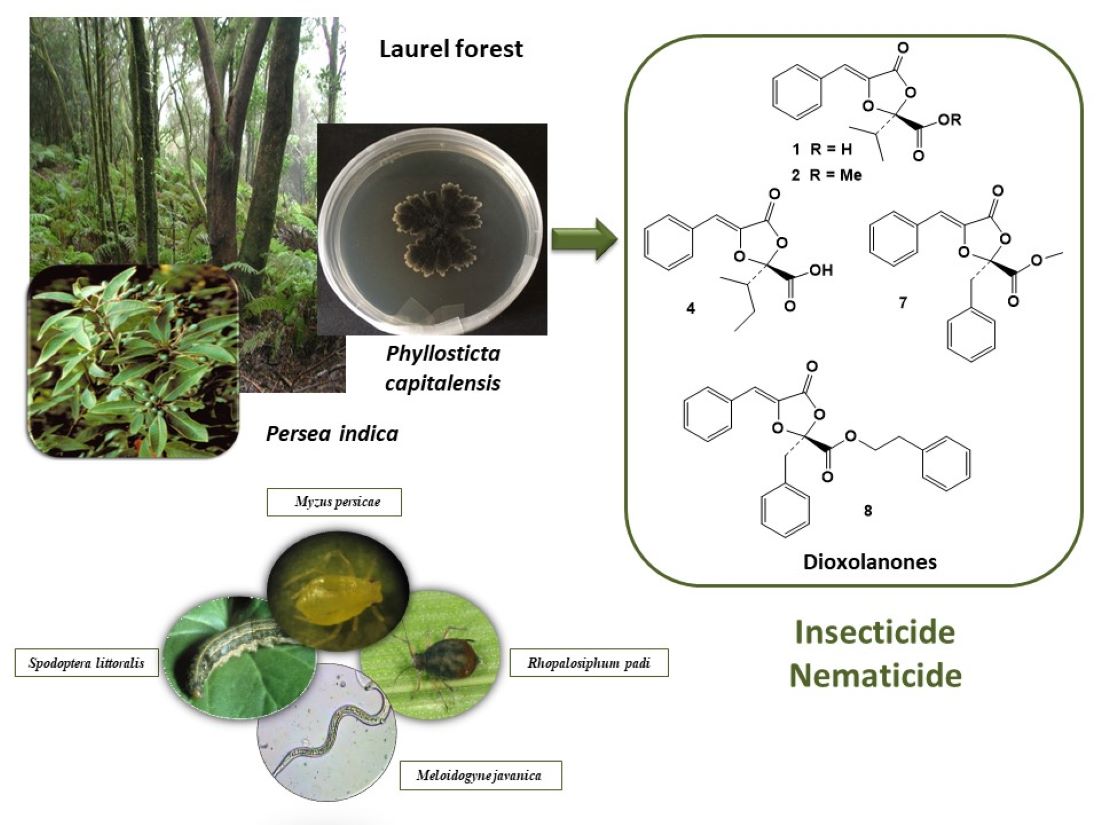
The search for natural product-based biopesticides from endophytic fungi is an effective tool to find new solutions. In this study, we studied a pre-selected fungal endophyte, isolate YCC4, from the paleoendemism Persea indica, along with compounds present in the extract and the identification of the insect antifeedant and nematicidal ones. The endophyte YCC4 was identified as Phyllosticta capitalensis by molecular analysis. The insect antifeedant activity was tested by choice bioassays against Spodoptera littoralis, Myzus persicae and Rhopalosiphum padi and the in vitro and in vivo mortality was tested against the root-knot nematode Meloiydogyne javanica. Since the extract resulted an effective insect antifeedant, a strong nematicidal and lacked phytotoxicity on tomato plants, a comprehensive phytochemical study was carried out. The chemical study resulted in the identification of two new metabolites, metguignardic acid (4) and (-)-epi-guignardone I (14), along the known dioxolanones guignardic acid (1), ethyl guignardate (3), guignardianones A (5), C (2), D (7) and E (6), phenguignardic acid methyl ester (8), and the meroterpenes guignardone A (9) and B (10) and guignarenone C (12), (-)-guignardone I (13) and the phyllomeroterpenoid B (15). Among these compounds, 1 and 4 were effective antifeedants against both insect species, while 2 was only active on the aphid M. persicae. The nematicidal compounds were 4, 7 and 8. This is the first report on the insect antifeedant or nematicidal effects of these dioxolanone-type compounds. Since the insect antifeedant and nematicidal activity of the P. capitalensis extract depends on the presence of dioxolanone components, future fermentation optimizations are needed to promote the biosynthesis of these compounds instead of the meroterpenes.
Keywords:
Phyllosticta capitalensis
; dioxolanones
; metguignardic acid
; meroterpenes
; 14‐epi‐Guignardone I
; nematicidal activity
; insecticidal activity
1. Introduction
Endophytic fungi are capable of producing a diverse range of bioactive compounds that exhibit a wide array of biological activities including, insecticidal, antioxidant, antifungal, antiviral, antibacterial, and cytotoxic properties. The secretion of these secondary metabolites by endophytic fungi is known to host plant defence response, enabling it to better cope with both biotic and abiotic stressors [1]. Thus, endophytic fungi represent a promising novel source for the biotecnological bioproduction of valuable active compounds [2].
From a biotechnological point of view, it is important to highlight that some endophytic fungi produce secondary metabolites previously found in their host plant. A gene transfer between endophyte and host plant during the establishment of the endophytic association could be the explanation [3]. There are several examples of pharmacologically important plant products obtained from fungal endophytes including paclitaxel (Taxol®) produced by Taxomyces andreanae from Taxus brevifolia, vincristine produced by Fusarium oxysporum from Catharanthus roseus, podophyllotoxin produced by Trametes hirsuta, Phialocephala fortinii and F. oxysporum isolates from Phialocephala hexandrum, P. peltatum and Juniperus recurve, and camptothecin produced by Fusarium solani from Camptotheca acuminata [3]. Therefore, the chemistry of the host plant is an important selection criteria for the isolation of fungal endophytes.
The Macaronesian laurel forest is an evergreen humid plant community composed of plants recruited from European/Mediterranean and tropical regions and modified due to temperature changes during the Pleistocene [4]. Laurel forest species host endophyte communities common to the tropical forests (Colletotrichum, Pestalotiopsis and Guignardia), being G. mangiferae, Glomerella acutata, Neofusicoccum parvum and Phomopsis sp. the dominant endophytes [5].
Persea indica, a perennial tree belonging to the Lauraceae family, is one of the dominant species of the Macaronesian laurel forest. P. indica contains ryanodane and isoryanodane diterpenes, alkene-γ-lactones and avocadofuranes in the aerial parts, avocadofuranes in the fruits [6], alkane-γ-lactones, alkyne-γ-lactones, avocadofurane precursors, cis- and trans-p-coumarate esters of (−)-borneol and small amounts of ryanoid diterpenes in the roots [7,8]. The ryanodane diterpenes are strong insect antifeedants against Spodoptera litura and S. littoralis [9,10], and among the compounds found in the roots, (−)-borneol cis-p-coumarate and (+)-majorenolide were moderately antifeedant and (+)-majorynolide was moderately nematicidal [8]. Given that the ryanoids are the most relevant compounds, the study of their accumulation in different plant parts (leaves, stems, roots) of a mature wild tree showed that the ryanodane epiryanodol was present in all the plant parts, with the stems and leaves having the highest amount [11]. In a previous study, an extract from the endophyte G. mangiferae A. J. Roy (anamorph Phyllosticta capitalensis P. Hennings), isolated from P. indica, showed strong insect antifeedant effects against S. littoralis and nematicidal activity against root-knot nematode Meloidogyne. javanica [5].
Therefore, in this work, the plant P. indica was chosen for the isolation of fungal endophytes with the potential ability to produce secondary metabolites with biopesticidal properties. The endophytic fungal strain YCC4 was isolated from P. indica leaves, identified and fermented on Czapek-Dox-Yeast liquid medium to give an ethyl acetate (EtOAc) extract with plant protection properties. The bioassay-guided fractionation of this extract gave five active compounds (1, 2, 4, 7, 8) that have been identified based on their spectroscopic data. The extract and pure compounds were tested against insect pests (Spodoptera littoralis, Myzus persicae and Rhopalosiphum padi) and the plant parasitic nematode M. javanica. Detailed findings are presented in this paper.
2. Results
The endophytic fungus YCC4 was isolated from leaves of the endemic Macaronesian species Persea indica and identified as Phyllosticta capitalensis Henn (Synonym: Guignardia manguiferae A.J. Roy). An EtOAc extract obtained from the fermentation of P. capitalensis was tested against insect pests (S. littoralis, M. persicae) and the root-knot nematode M. javanica.
The extract showed strong antifeedant effect against S. littoralis (EC50 = 9.77 μg/cm2), followed by M. persicae (EC50 = 17.2 μg/cm2), and it was not active on R. padi. The nematicidal effects of the extract against M. javanica J2 were very strong (24-72h mortality range of 96-100%), with effective LC50 concentration of 0.44 mg/mL at 72h. Furthermore, the extract also had effective egg hatching inhibition effects (64% inhibition respect to the control after 28 days) (Table 1). The extract was also tested for phytotoxic effects on seeds of mono- and dicotyledoneous plant species (Lolium perenne, Lactuca sativa and Solanum lycopersicum), at a dose of 0.4mg/mL. There were no significant effects on L. perenne germination, leaf and root growth (14, 34, 26% inhibition respectively after 7 days), a moderate promotion of L. sativa root growth (55%), and no effect on S. lycopersicum germination (14% inhibition after 7 days) or root growth (Table 1).
Given the strong in vitro nematicidal effects (on J2 mortality and egg hatching) of the extract and its low phytotoxicity on S. lycopersicum, an in vivo experiment in pots was conducted. YCC4 extract-treated soil at the LC50 concentration (0.44 mg/mL) significantly reduced the reproductive traits of the M. javanica population causing a strong reductions (>60%) of nematode egg masses, egg production, and IF and MR index, with respect to the untreated control after 2 months (Table 2).
The bioactive extract from endophyte P. capitalensis was submitted to a comprehensive phytochemical study. The chromatographic study of the extract led to the isolation of two new metabolites and thirteen known compounds (Figure 1 and Figure 2). By comparison of their 1H and 13C data with the literature, these known compounds were identified as the dioxolanones guignardic acid (1) [15,16], ethyl guignardate (3) [17], guignardianones A (5), C (2), D (7) and E (6) [18,19], phenguignardic acid methyl ester (8) [20,21], and the meroterpenes guignardone A (9) [22,23], guignardone B (10) [22], guignarenone B (11) and guignarenone C (= guignardone H) (12) [24,25], (-)-guignardone I (13) [25,26], phyllomeroterpenoid B (15) [21], and 3,4-dihydroxybenzoic acid.
Among the new compounds, the first one was named metguignardic acid and assigned structure 4. Its high resolution mass spectrum showed the molecular ion [M]+ at m/z 276.1001 in accordance with the formula C15H16O5. The 1H NMR spectrum of 4 was very similar to that of guignardic acid (1), also obtained from this culture. Analysis of the 1H NMR spectrum revealed the presence of two methyl groups at δ 0.99 (t, J = 7.4 Hz,H-17) and 1.07 (d, J = 6.8 Hz, H-15), one methine signal at δ 2.43 (1H, ddd, J = 10.1, 6.8, 3.2 Hz, H-14) and an olefinic proton at δ 6.52 (1H, s). Additionally, the protons of one methylene group were observed at δ 1.29 (1H, ddd, J = 13.5, 10.1, 7.3 Hz, H-16) and 1.66 (1H, ddd, J = 13.5, 7.4, 3.2 Hz-H-16), as well as resonances of five aromatic protons [7.34 (1H, m), 7.40 (2H, m), 7.67 (2H, dd, J = 7.1, 1.5 Hz)] of a mono-substituted benzene ring.
The 13C NMR spectrum (Table 3) and HSQC spectra confirmed these assignments by showing signals from two methyl groups, a methylene, a methine, a trisubstituted double bond at δ 110.1 (C-6) and 135.3 (C-4), which is conjugated with a lactone at δ 162.7 (C-5). An angular carbon bearing two oxygenated functions at δ 108.3 (C-2) and a carboxyl group at δ 169.5 (C-13) were also observed. The only difference with guignardic acid (1) was therefore, the appearance of a methylene group at δ 22.4 (C-16) indicating the presence on C-2 of an isobutyl group instead of an isopropyl one.
The HMBC spectrum was also in accordance with structure 4. Thus, correlations of H-15 and H-17 methyls with C-14 (δ 39.3) and C-16 (δ 22.4) were observed. Furthermore, H-15 methyl and H-16 methylene showed connectivities with C-2 (δ 108.3), which allowed the isobutyl group to be located in the molecule. The singlet at δ 6.52 (H-6) showed correlations with the aromatic carbons at δ 129.9 (C-8 and C-12), 135.3 (C-4) and 162.7 (C-5). Crosspeaks observed between two aromatic protons at δ 7.40 (H-9 and H-11) and a carbon at δ 132.1 permitted this last signal to be assigned to C-7.
(-)-14-epi-Guignardone I is another new natural product to which we have assigned structure 14 on the basis of the following considerations: Its HRMS showed the molecular ion [M]+ at m/z 310.1630 corresponding to C17H26O5, which indicated that is an isomeric compound of (-)-guignardone I (13). In its 1H NMR spectrum were observed signals of an angular methyl at δ 1.48 (H-11), two methyls at δ 1.21 and 1.37 (H-16 and H-17) on a same carbon bearing oxygen, and a methoxy group at δ 3.50 (H-7). Other observed resonances were the signals of coupled protons H-8 at δ 2.04 (1H, ddd, J =17.0, 10.9, 1.9 Hz, H-8β) and 2.65 (1H, dd, J = 17.0, 6.3 Hz, H-8α) and, two doublet of doublets at δ 2.18 (J = 13.4, 7.5, 6.0 Hz, H-5β) and 2.40 (J = 13.3, 4.8, 3.8 Hz, H-5α) of methylene group at C-5. Two methine signals at δ 4.29 (dd, J = 13.2, 5.4 Hz, H-4) and 3.73 (dd, J = 7.9, 3.8 Hz, H-6) were assigned to two geminal protons of hydroxyl and methoxy groups attached at C-4 and C-6, along the signal of proton H-9 at δ 1.88 (dd, J = 10.8, 5.9 Hz, H-9). Significant was the resonance at δ 2.13, at a lower field than in guignardone I (13) (δ 1.59), which was attributed to H-14 based on its HMBC correlations with C-9 (δ 40.8), C-13 (δ 18.9) and C-15 (δ 71.8). This fact led us to think that 14 was the C-14 epimer of (-)-guignardone I, which is rare in this type of meroterpenoids. This configuration was finally established considering the NOESY spectrum of 14, which showed a strong correlation between H-11 methyl with H-9 and H-14, indicating that they have an axial configuration in the α-face of the molecule. Correlations of H-5β with H-4 and H-6 were also observed. These data, along with the coupling constants of the signals corresponding to protons H-4 and H-6, showed an α disposition for the hydroxyl and methoxy group at C-4 and C-6 in the cyclohexenone ring [27]. The13C NMR spectrum (Table 3) also confirmed the structure 14 for (-)-14-epi-guignardone I, showing signals of three methyls, one methoxy group, four methylenes, two tertiary carbons at δ 66.0 (C-4) and 79.0 (C-6), two quaternaries carbons bearing oxygen at δ 71.8 (C-15) and 86.3 (C-10), and a carbonyl group at δ 194.6 (C-1). In addition, two olefinic carbons were observed at δ 107.8 (C-2) and 167.5 (C-3), the latter also bound to an oxygen atom.
Compounds 1-15 were tested for their biocidal effects against the insect and nematode targets (S. littoralis, M. persicae, M. javanica). Table 4 shows the insect antifeedant effects of these compounds, with 4 and 1 being effective against S. littoralis (EC50 values of 7.53 and 9.65 μg/cm2 respectively), while 1, 2 and 4 acted on M. persicae (EC50 values of 2.87, 1.74 and 11.45 μg/cm2, respectively).
When tested against M. javanica J2 in vitro, compounds 4, 7 and 8 were very active (LC50 values of 0.24, 0.11 and 0.23 mg/mL respectively), while 1 showed moderate effects (60% mortality at 1 mg/ml) (Table 5). The time-course experiment of the nematicide compounds (24, 48 and 72h) showed that 8 was the most active one, starting at 24h, followed by 4 (Table 5). Further in vitro tests on egg hatching inhibition showed that for the total inhibition rate values, compound 7 was the most active one (71.5% inhibition) followed by 8 (58.2%) and 4 (54.4%) (Figure 3). However, the time course observation showed that 7 induced an early inhibition at 0 d (61.2%) with a similar peak at 28 d (65%); compound 4 peaked at 28 d (83.8%) and compound 8 peaked at 14 d (51%) (Figure 3). Therefore, the time-course egg hatching inhibition caused by the extract correlated with the combined action of these compounds.
3. Discussion
The genus Phyllosticta (Ascomycetes) are a group of microscopic fungi that can colonize a variety of plant hosts, including several Citrus species. Some Phyllosticta species have the capacity to cause disease, including leaf spots and black spots on fruits, while others have only been observed as endophytes [25]. Phyllosticta capitalensis is an endophyte and weak plant pathogen with a worldwide distribution presently known from 70 plant families [26]. Phyllosticta species are potential biocontrol agents. For example, P. capitalensis isolated from Camellia sinensis showed antifungal effects in dual growth tests against Fusarium sp., Pestalotiosis sp. and Sclerotinia sclerotiorum [27]. Furthermore, endophytitc Phyllosticta sp. isolated from cucumber showed potential as a seed treatment agent for Meloidogyne incognita biocontrol [28]. P. capitalensis was previosusly described in P. indica (as Guignardia manguiferae) as the producer of a bioactive extract against insects and nematodes [5] as reported here, but the secondary metabolites responsible for such activities were unknown.
In this work, the secondary metabolites produced by the endophytic fungus P. capitalensis have been identified. The phytochemical study of this extract gave the known compounds guignardic acid (1), ethyl guignardate (3), guignardianones A (5), C (2), D (7) and E (6), phenguignardic acid methyl ester (8), and the meroterpenes guignardone A (9) and B (10) and guignarenone C (= guignardone H) (12), (-)-guignardone I (13) and the phyllomeroterpenoid B (15), along with the new C-14 epimer of (-)-guignardone I (14) and metguignardic acid (4).
The biogenetic origin of compound 4 could be explained as the fusion of the products from the oxidative deamination of the amino acids phenylalanine and isoleucine, similarly to that described for guignardic acid (1) [12], but replacing the amino acid valine for isoleucine, which would result in the formation of metguignardic acid (4) instead of guignardic acid (1). There are other dimers of dioxolanone type compounds in which the aminoacids valine or phenylalanine have been proposed as precursors in their formation instead of valine [13,16].
Phyllostica species and P. capitalensis are an important source of meroterpenes compounds. The meroterpenes guignardones A-C and guignarerones A-D [19], guignardones D-E and tricycloalternarene F [20], guignarenone C and guignardone J-K [29] have been isolated from P. capitalensis (G. mangiferae), isolated from Ilex cornuta, and guignardones J-O from Guignardia sp. [30]. Other compounds from P. capitalensis isolated from Smilax glabra, were 15-hydroxy-tricycloalternarene, guignardones A-B and guignardones P-S [31]. Guignardones F-I were obtained in cultures of an endophytic fungus A1, isolated from a mangrove plant [22,23,24,25,26]. P. capitalensis isolated from Cephalotaxus fortunei also yielded a large number of meroterpenes (guignardone B, F, G, C, M, N, O, H, I, J, K; 12-hydroxylated guignardone A, 13-hydroxylated guignardone A, 17-hydroxylated guignardone A, phyllostictone A-E [14]. A mangrove P. capitalensis endophyte gave guignardones A, J, M and 12-hydroxylated guignardone A [32] and guignardones U-X [33]. Additionally, another isolate of this fungal species produced the sesquiterpene-shikimate-conjugated spirocyclic meroterpenoids A and B [34]. Meroterpenes have been described as antibacterial and antinflammatory compounds [34]. However, in this work, we have not found any antifeedant or nematicidal effects for the meroterpenes produced by P. capitalensis isolated from P. indica.
In contrast to meroterpenes, a lower number of dioxolanone derivatives have been found in P. capitalensis. An isolate from from Smilax glabra gave guignardiones C-D [35], and an isolate from Cephalotaxus fortunei yielded the dioxolanones guignardic acid, 2-hydroxyethyl guignardate, ethyl guignardate and guignardianone C [14]. Guignardianone G was reported from a mangrove isolate of P. capitalensis [36]. Plant protection effects for dioxolanone-type compounds have been described. Guignardianone C showed broad-spectrum antifungal activities against the plant pathogens Rhizoctonia solani, Fusarium graminearum and Botrytis cinerea, but not guignardic or penguignardic acids [33]. Guignardic and alaguignardic acids were phytotoxic in a Vitis vinifera leaf-disk assay and also against Oryza sativa and Triticum aestivum, whereas guignardianones A-F were no active, indicating that the free carboxyl group or at least a polar moiety is important for the phytotoxic activity of the analyzed dioxolanone-type secondary metabolites [16]. Guignardianone C moderately inhibited the growth of L. sativa and L. perenne [14].
In this work, only dioxolanone derivatives were active. Guignardic (1) and metguignardic acid (4) were effective antifeedants against both insect species, while guignardione C (2) was only active on the aphid M. persicae. In this case, and similarly to the phytotoxic activity described before, the active compounds presented a free carboxyl group (1, 4) or a metyhylated group (2), in contrast with the inactive less polar ethyl derivative one (3). The nematicidal effects showed a different structure-activity relationship pattern. The presence of an aromatic phenylethyl substituent in C-2 determined a strong effect (7, 8), followed by an an isobutyl group (4). This is the first report on the insect antifeedant or nematicidal effects of these compounds.
4. Materials and Methods
4.1. General Experimental Procedures
Optical rotations were determined at room temperature on a Perkin Elmer 343 polarimeter (Perkin Elmer, Waltham, MA, USA). IR spectra were taken in Bruker IFS 66/S and Perkin-Elmer 1600 spectrometers. NMR spectra were measured on a Bruker AMX-500 spectrometer (1H 500 MHz/13C 125 MHz) with pulsed field gradient using the CDCl3 solvent (δH 7.26 and δC 77.0) as internal standard. EIMS and exact mass measurements were recorded on a Micromass Autospec instrument at 70 eV. The HRESIMS were acquired using a Micromass LCT Premier spectrometer. Preparative and semipreparative HPLC was carried out with a Beckman Coulter 125P equipped with a diode-array detector 168 (Beckman Coulter Life Sciences, Brea, CA, USA), preparative Interstil Prep-sil 20 mm x 250 mm, 10 µm particle size (Gasukuro Kogio) and semipreparative Beckman Ultrasphere silica 10 mm x 250 mm, 5 µm particle size columns. Silica gel 60 F254 (Merck, Darmstadt, Germany) and Sephadex LH-20 (Sigma-Aldrich, St. Lo Sigma-Aldrich, St. Louis, MO, USA) were used for column chromatography.
4.2. Isolation and Identification of Endophytic Fungus YCC4
The endophytic fungus Phyllosticta capitalensis Henn. was isolated from leaves of the endemic Macaronesian species Persea indica (L.) Spreng., collected at Las Mercedes in Parque Rural de Anaga (Tenerife, Canary Islands). The leaves were washed with tap water for 10 min to remove impurities. Surface disinfection was performed in a laminar flow hood by soaking in 70% ethanol for 1 minute, followed by 1% sodium hypochlorite for 10 minutes, and then washed again with 70% ethanol for 1 minute. The plant samples were finally washed with sterile distilled water for 1 min. Surface-sterilized samples were then cut into small segments (0.5 cm) and tissue segments were placed in Petri dishes containing sterile potato dextrose agar medium (PDA), supplemented with chloramphenicol (50 mg/ L) to inhibit bacterial growth. The plates were incubated at 27 ºC in a growth chamber for 3–15 days in darkness. The emerging fungal colonies were transferred to fresh PDA plates to obtain pure strain and further identification.
The isolated fungus was identified at the molecular level based on the amplification (PCR) and sequencing of the ribosomal ITS region of the rDNA, according to a molecular biological protocol described previously [37]. Briefly, genomic DNA (100–200 ng) was amplified (PTC-200 Thermal Cycler, MJ Research, San Diego, CA, USA), 25 µL final volume with AmpONE Taq DNA polymerase PCR kit (GeneAll, Seoul, Korea) with 35 cycles (95 °C, 1 min; 50 °C, 20 s; 72 °C, 1.5 min) after an initial denaturation (95 °C, 2 min) followed by a final extension (72 °C, 7 min). The PCR-amplified products were checked by agarose gel (1%) electrophoresis, purified using the EXO-SAP-IT kit (Affimetrix-USB; Thermo Fisher Scientific, Waltham, MS, USA), and sequenced on an AB 3500 Genetic Analyzer (Thermo Fisher Scientific, Waltham, MS, USA) at the University of La Laguna (La Laguna, Spain) genomic service. The fungal endophyte was identified as P. capitalensis Henn.by comparison of the ITS1-5.8S-ITS2 sequence of the rDNA with those deposited in the NCBI database (National Center for Biotechnology Information) using Basic Local Alignment Search Tool (nBLAST) (GenBank accession MT649668). A sample of this fungus was deposited in. CECT (Valencia, Spain) with number 20914 in accordance with the Budapest treaty.
4.3. Cultivation of YCC4 for Extract Preparation
P. capitalensis was cultivated in PDA medium in Petri dishes for 15 days at 26 oC. After this time, sterile distilled water (10 ml) was added, and the surface of the mycelium was gently scraped with a spatula. This mycelial suspension was poured into an Erlenmeyer flask (250 ml) with 100 ml of Czapek-Dox-Yeast liquid medium [Cz-L: NaNO3 (2 g/l), KH2PO4 (5 g/l), MgSO4 (0.5 g/l), FeSO4 (0.01 g/l), ZnSO4 (0.003 g/l), yeast extract (1 g/l) and glucose (60 g/l)], and cultivated at 26 oC continuously stirring (120 rpm) for 4 days to obtain 1 liter of pre-inoculum. Erlenmeyer flasks (100, 250 ml) with fresh medium (100 ml) were inoculated with 5 ml of the pre-inoculum. Then, after 20 days of fermentation under the same conditions, the culture medium was separated from the mycelium by filtration in a Büchner and extracted with EtOAc. Subsequently, the solvent was removed under reduced pressure to afford a dry crude extract (10.5 g).
4.4. Isolation and Compound Identification
The dry crude extract (10.5 g) was fractionated by vacuum liquid chromatography (VLC) over silica gel eluted with an increasingly polar gradient of n-Hexane/EtOAc and EtOAc/MeOH to obtain six fractions. Fraction 1 (n-Hexane/EtOAc 75:25) was separated into four subfractions (F1A-F1D) by silica gel column chromatography using n-hexane /EtOAc mixtures of increasing polarity (90:10–50:50). Guignardianone A (5) (23.0 mg), guignardianone C (2) (96.1 mg) and ethyl guignardate (3) (28 mg) were isolated from the less polar subfraction 1A (n-hexane/EtOAc, 90:10), using preparative high pressure liquid chromatography (HPLC) (Hex/EtOAc 97:3). Subfraccion 1B, obtained with n-hexane/EtOAc 15%, was subjected to (CC) on silica gel medium pressure column with n-hexane/EtOAc gradients (100:0-90:10) and, subsequently, by preparative HPLC using n-hexane/EtOAc (97:3) as mobile phase afforded penguignardic acid methyl ester (7) (36.5 mg) and guignardianone D (8) (89 mg). Similarly, preparative HPLC chromatography of subfraction 1C (n-hexane/EtOAc 85:15) eluted with n-hexane/EtOAc 20% gave guignardone A (9) (78.3 mg). In the most polar subfraction of the chromatography of fraction 1 (n-hexane/EtOAc 50:50), metguignardic acid (4) (204 mg) was isolated.
Guignardianone A (5) (430 mg) and guignardone A (9) were obtained again (80 mg) from fraction 2 by silica gel column chromatography with mixtures of increasing polarity of n-hexane/EtOAc and EtOAc/MeOH. In addition, guignardone B (10) (13.6 mg) and phyllomeroterpenoid B (15) (5.0 mg) were isolated in the fractions eluted with EtOAc. Further purification the most polar fractions (MeOH) by HPLC chromatography with n-hexane/EtOAc/MeOH (30:64:6) as mobile phase gave metguignardic acid (4) (30 mg) and guignardic acid (1) (118 mg).
Fraction 3, obtained eluting with n-hexane/EtOAc (50:50), was chromatographed on silica gel column with mixtures of increasing polarity of n-hexane/EtOAc and EtOAc/MeOH. After successive medium pressure column chromatographies and/or HPLC, guignardianone C (2) (49 mg), phenguignardic acid methyl ester (8) (2.4 mg), guignardianone E (6) (1.5 mg), guignarenone C [= guignardone H] (12) (14.6 mg), guignardone B (10) (19.2 mg) and 3,4-dihydroxybenzoic acid (34.3 mg) were isolated.
The most polar fractions 4-6 (EtOAc/MeOH 100:0-50:50) of the extract were subjected to silica gel column chromatography eluted with an increasingly polar gradient of CH2Cl2/MeOH (99:2-0:100). Less polar fractions (CH2Cl2/MeOH, 99:1-99:2) were purified by semipreparative HPLC using a mixture of CH2Cl2/EtOAc 50% to give guignarenone B (11) (2.2 mg). From fractions eluted with CH2Cl2/MeOH (99:5-90:20) were isolated (-)-14-epi-guignardone I (14) (6.2 mg) and (-)-guignardone I (13) (1.1 mg) by HPLC chromatography with n-hexane/EtOAc/MeOH (40:55:5) as eluent.
4.4.1. Guignardic Acid (1)
[α]D: -26.7 (c 0.25, CHCl3); 1H NMR (500 MHz, CDCl3): δ 1.10 and 1.09 (each 3H, d, J = 6.9 Hz, H-15 and H-16), 2.68 (1H, sept, J = 6.9 Hz, H-14), 6.53 (1H, s, H-6), 7.35 (1H, m, H-10), 7.39 (2H, m, H-9 and H-11), 7.68 (2H, br d, J = 7.1 Hz, H-8 and H-12); EIMS m/z: 262 [M]+ (5), 118 (100), 90 (38); 13C NMR data, see Table 3; HRMS [M]+ at m/z 262.0853, calculated for C14H14O5 262.0841.
4.4.2. Guignardianone C (2)
[α]D: -10.2 (c 0.25, CHCl3); 1H-RMN (500 MHz, CDCl3): δ 1.07 and 1.08 (6H, d, J = 6.9 Hz, H-15 and H-16), 2.69 (1H, sept, J = 6.9 Hz, H-14), 3.85 (3H, s, -OCH3), 6.50 (1H, s, H-6), 7.35 (1H, m, H-10), 7.41 (2H, m, H-9 and H-11), 7.68 (2H, d, J = 7.1 Hz, H-8 and H-12); EIMS m/z: 276 [M]+ (5), 118 (100), 90 (39), 71 (13); 13C NMR data, see Table 3; HRMS [M]+ at m/z 276.0999, calculated for C15H16O5 276.0998.
4.4.3. Metguignardic Acid (4)
[α]D -54.7 (c 0.3, CHCl3); IR (CHCl3) νmax: 2919, 1796, 1739, 1254, 1195, 1183 cm-1; 1H NMR (500 MHz, CDCl3): δ 0.99 (3H, t, J = 7.4 Hz, H-17), 1.07 (3H, d, J = 6.8 Hz, H-15), 1.29 (1H, ddd, J = 13.5, 10.1, 7.3 Hz, H-16), 1.66 (1H, ddd, J = 13.5, 7.4, 3.2 Hz, H-16), 2.43 (1H, ddd, J = 10.1, 6.8, 3.2 Hz, H-14), 6.52 (1H, s, H-6), 7.34 (1H, m, H-10), 7.40 (2H, m, H-9 and H-11), 7.67 (2H, dd, J = 7.1, 1.5 Hz, H-8 and H-12); EIMS: m/z 276 [M]+ (6), 118 (100), 90 (50); 13C NMR data, see Table 3; HRMS [M]+ at m/z 276.1001, calculated for C15H16O5 276.0998.
4.4.4. Phenguignardic Acid Methyl Ester (7)
[α]D +47.6 (c 0.14, CHCl3); IR (CHCl3) νmax: 2919, 2369, 2345, 1734, 1256, 1178 cm-1; 1H NMR (500 MHz, CDCl3): δ 3.52 and 3.57 (each 1H, d, J = 14.7 Hz, H-14), 3.85 (3H, s, -OCH3), 6.30 (1H, s, H-6), 7.24 (3H, m, H-17, H-18 and H-19), 7.28 (2H, m, H-16 and H-20), 7.36 (1H, m, H-10), 7.40 (2H, m, H-9 and H-11), 7.63 (2H, br d, J = 7.2 Hz, H-8 and H-12); 13C NMR data, see Table 3; EIMS: m/z 324 [M]+ (2), 264 (2), 118 (100), 91 (29); HRMS [M]+ at m/z 324.0987, calculated for C19H16O5 324.0998.
4.4.5. Guignardianone D (8)
[α]D: +40.3 (c 0.25, CHCl3); 1H-RMN (500 MHz, CDCl3): δ 2.96 (2H, t, J = 6.6 Hz, H-2’), 3.52 and 3.46 (each 1H, d, J = 14.7 Hz, H-14), 4.45 (2H, m, H-1’), 6.30 (1H, s, H-6), 7.15 (2H, m, H-4’ and H-8’), 7.19 (3H, m, H-5’, H-6’and H-7’), 7.24 (5H, br s, H-16, H-17, H-18, H-19 and H-20), 7.36 (1H, m, H-10), 7.41 (2H, m, H-9 and H-11), 7.62 (2H, d, J = 7.2 Hz, H-8 and H-12); EIMS: m/z 414 [M]+ (4), 265 (11), 118 (100), 105 (27), 91 (84); 13C NMR data, see Table 3; HRMS [M]+ at m/z 414.1465, calculated for C26H22O5 414.1467.
4.4.6. Guignarenone C (= Guignardone H) (11)
[α]D +82.8 (c 0.18, CHCl3); 1H NMR (500 MHz, CDCl3): δ 1.33 (3H, s, H-11), 1.54 (1H, dddd, J = 13.2, 11.1, 8.4, 3.9 Hz, H-13α), 1.66 (3H, br s, H-17), 1.79 (1H, ddd, J = 14.1, 11.2, 7.1 Hz, H-12α), 1.92 (1H, m, H-13β), 1.95 (1H, m, H-9), 2.13 (1H, m, H-12β), 2.17 (1H, m, H-8α), 2.20 (1H, m, H-14), 2.23 (1H, ddd, 13.8, 6.7, 5.3 Hz, H-5α), 2.33 (1H, dt, J = 16.2, 1.5 Hz, H-8β), 2.37 (1H, ddd, J = 13.8, 4.8, 3.7 Hz, H-5β), 3.47 (3H, s, -OMe), 3.70 (1H, dd, J = 6.7, 3.6 Hz, H-6), 4.25 (1H, t, J = 5.1 Hz, H-4), 4.63 (1H, dt, J = 1.8, 0.9 Hz, H-16), 4.73 (1H, t, J = 1.5 Hz, H-16); 13C NMR data, see Table 1; EIMS m/z: 292 [M]+ (1), 262 (100), 234 (18), 219 (10), 122 (32), 84 (42); 13C NMR data, see Table 3; HRMS [M]+ at m/z 292.1676, calculated for C17H24O4 292.1675.
4.4.7. (-)-Guignardone I (13)
[α]D -40.0 (c 0.3, MeOH); 1H NMR (500 MHz, CDCl3): δ 1.19 and 1.21 (each 3H, s, H-16 and 17), 1.34 (3H, s, H-11), 1.57 and 1.80 (each 1H, m, H-13β and H-13α), 1.59 (1H, m, H-14), 1.63 and 2.02 (each 1H, m, H-12β and H-12α), 2.06 (1H, td, J = 9.0, 1.5 Hz, H-9), 2.23 (1H, ddd, J = 13.7, 7.0, 5.5, H-5β), 2.26 (1H, ddd, J = 17.7, 7.5, 1.5 Hz, H-8β), 2.39 (1H, ddd, J = 13.7, 4.8, 3.7 Hz, H-5α), 2.64 (1H, d, J = 17.5 Hz, H-8α), 3.48 (3H, s, -OCH3), 3.72 (1H, dd, J = 6.8, 3.7 Hz, H-6), 4.27 (1H, br t, J = 5.1 Hz, H-4); EIMS: m/z 310 [M]+ (4), 280 (45), 262 (22), 209 (21), 194 (100), 179 (25), 166 (37), 122 (90), 81 (49), 59 (58); 13C NMR data, see Table 3; HRMS [M]+ at m/z 310.1769, calculated for C17H26O5 310.1780.
4.4.8. (-)-14-epi-Guignardone I (14)
[α]D + 6.25 (c 0.32, MeOH); νmax: 3448, 2969, 2929, 1654,1617, 1457, 1362, 1283, 1215, 1163, 1119, 1071 cm-1; 1H NMR (500 MHz, CDCl3): δ 1.21 and 1.37 (each 3H, s, H-16 and 17), 1.48 (3H, s, H-11), 1.69 and 1.94 (each 1H, m, H-13), 1.82 (2H, br t, J = 8 Hz, H-12), 1.88 (1H, dd, J = 10.8, 5.9 Hz, H-9), 2.04 (1H, ddd, J =17.0, 10.9, 1.9 Hz, H-8β), 2.13 (1H, m, H-14), 2.18 (1H, m, H-5β), 2.40 (1H, ddd, J = 13.3, 4.8, 3.8 Hz, H-5α), 2.65 (1H, dd, J = 17.0, 6.3 Hz, H-8α), 3,18 (d, J= 6.3, -OH), 3.50 (3H, s, - OCH3), 3.73 (1H, dd, J = 7.9, 3.8 Hz, H-6), 4.29 (1H, dt, J = 6.4, 4.6, H-4); 13C NMR data, see Table 3; HRESIMS [M+Na]+ at m/z 333.1669, calculated for C17H26O5Na 333.1678.
4.5. Nematicidal Activity
Meloidogyne javanica population was maintained on Solanum lycopersicum plants (var. Marmande) cultivated in pot cultures in environmentally controlled growth chambers (at 25 ±1 °C, >70% relative humidity). Egg masses of M. javanica were handpicked from infected tomato roots. Second-stage juveniles (J2) were obtained from hatched eggs by incubating egg masses in a water suspension at 25 ºC for 24 h.
4.5.1. In Vitro Effect on Juveniles
The extract, fractions and compounds were dissolved in distilled water containing 5% of a DMSO-Tween solution (0.5% Tween 20 in DMSO) and evaluated as described by Andres et al. (2017) [38]. The initial concentrations tested were of 1 and 0.5 mg mL for extract or pure compound respectively and four replicates were used for each test. Tests with mortality rates >90% at 72h, were further tested to assess J2 mortality after 24 and 48 h. The nematicidal activity data are presented as percent dead J2 corrected according to Scheider-Orelli’s formula. Five serial dilutions were used to calculate the effective lethal doses (LC50 and LC90) by Probit Analysis (STATGRAPHICS Centurion XVI, version 16.1.02).
4.5.2. In Vitro Effect on Egg Hatching
Egg masses (three) were placed in each well of a 96-well plate containing test solutions at LC90 concentrations. The control wells contained water/DMSO/Tween 20. Each experiment was replicated four times. The plates were covered and maintained in the dark at 25C for 5 days after which the hatched J2s were counted, and the treatments were replaced with sterilized distilled water. The hatched J2s from egg masses were monitored weekly for 1 month until egg hatching was finished in the control [38]. Relative hatch inhibition rates (compared with the controls) were calculated for each immersion time as follows:
Relative suppression rate (%) = (number of J2 in control — number of J2 in test solutions) / number of J2 in control × 100.
4.5.3. Effect on Infection and Reproduction of M. javanica Population in Tomato Plants Hatching
The extract was evaluated at 0.45 mg/mL (LC50) in 1% ethanol. The treatment (100 mL) was applied to a pot containing 1000 g of the moistened substrate (sterile sandy/loam soil mixture) at the time of nematode inoculation (2000 M. javanica eggs) and incubated for 5 days in a growth chamber (25 ºC, 60% relative humidity). After this period, 4-week-old tomato seedlings were transplanted, maintained for 60 days in a growth chamber (25 ºC, 60% relative humidity, 16 h photoperiod) and fertilized with 50 mL of a 0.3% solution of 20-20-20 (N-P-K) every 10 days. Six pots for each treatment were used, and the experiment was carried out in duplicate. At harvest, the whole root system from each pot was collected. Roots were washed free of soil, examined for determining the number of egg masses. Eggs from root egg masses were extracted by maceration in a 10% commercial bleach solution (40 g/L NaOCl) for 10 min, passed through a 70 μm aperture screen and collected in a 25 μm sieve for final counting. Relative suppression rate of the extract on eggs masses and number of eggs per plant was calculated. The infection frequency (IF: number of egg masses per plant divided by the number of eggs inoculated per pot) and the multiplication rate (MR: number of eggs per plant divided by the egg inoculum) were determined. Data from each treatment were transformed by Log10 (x), mean values were compared by Student t-test at p < 0.05 to determine significant differences in nematode population reproductive traits associated with treatment effects.
4.6. Antifeedant Activity
The insect colonies (Spodoptera littoralis, Myzus persicae and Rhopalosiphum padi) come from laboratory colonies reared on artificial diet and host plants (Capsicum annuum, Hordeum vulgare), respectively, at 22 ± 1 °C, >70% relative humidity and 16:8 h (L:D) photoperiod at ICA-CSIC.
The tests have been described before [11]. Briefly, the upper surface of leaf disks or fragments (1.0 cm2) of C. annuum and H. vulgare were treated with 10 µL of extract or compound at an initial dose of 10 or 5 µg/µL (100 or 50 µg/cm2) respectively. Two sixth-instar S. littoralis larvae (>24 h after molting) per Petri dish or 10 apterous aphid adults (24–48 h old) placed in a 2×2 cm ventilated plastic box (20) were allowed to feed at room temperature or in the growth chamber respectively. The experiments ended at 75% larval consumption of paired control or treatment disks for S. littoralis or after 24 h for aphids. Each experiment was repeated 2 times. Feeding inhibition (%FI), based on the disk surface consumption (digitalized with https://imagej.nih.gov/ij/ [39], and aphid settling inhibition (%SI), based on the number of aphids on each leaf fragment, were calculated as % FI/SI = [1 − (T/C) × 100], where T and C represent feeding/settling on treated and control leaf disks, respectively. The significance of these effects was analyzed by the nonparametric Wilcoxon paired signed-rank test. Tests with an FI/SI > 70% were further tested in dose-response experiments (range of activities between 100 and <50%, minimum of 3 doses) to calculate their effective dose EC50 from linear regression analysis (% FI/SI on Log-dose, STATGRAPHICS Centurion XVI, version 16.1.02).
5. Conclusions
The endophyte Phyllosticta capitalensis has been isolated from Persea indica. An EtOAc extract from this fungus showed strong insect antifeedant (against Spodoptera littoralis and Myzus persicae) and nematicidal (against Meloydogine javanica) effects in vitro and in vivo.
A comprehensive phytochemical study of this extract gave the known compounds guignardic acid (1), ethyl guignardate (3), guignardianones A (5), C (2), D (7) and E (6), phenguignardic acid methyl ester (8), and the meroterpenes guignardone A (9) and B (10) and guignarenone C (= guignardone H) (12), (-)-guignardone I (13) and the phyllomeroterpenoid B (15) along with the new C-14 epimer of (-)-guignardone I (14) and metguignardic acid (4). Among these compounds, only dioxolanone derivatives were active against the insect and nematode targets. Compounds 1 and 4 were effective antifeedants against both insect species, while 2 was only active on the aphid M. persicae. The nematicidal compounds were 4, 7, and 8. This is the first report on the insect antifeedant or nematicidal effects of these compounds. Since the insect antifeedant and nematicidal activity of the P. capitalensis extract depends on the presence of dioxolanone components, future fermentation optimizations are needed to promote the biosynthesis of these compounds instead of the meroterpenes.
6. Patents
Gonzalez-Coloma, A.; Díaz, C.E.; Andres, M.F.; Fraga, B.M.; Bolaños, P. et al. Biocidal products and use thereof for controlling phytopathogens and pest organism that harm plants. 2016, PCT Patent WO 2016/034751 A1
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Data sequence of Phyllosticta capitalensis; Figures S1–S4: 1H-NMR and 13C-NMR of compounds 1-2; Figures S5–S10: 1H-NMR,13C-NMR, DEPT, HSQC, COSY, HMBC spectra for compound 4; Figures S11–S14: 1H-NMR and 13C-NMR of compounds 7-8; Figures S15–S20: 1H-NMR,13C-NMR, HSQC, COSY, HMBC, NOESY spectra for compound 14.
Author Contributions
Conceptualization, C.E.D. and A.G-C.; Methodology, M.F.A., P.B., C.E.D., A.G-C.; Validation, M.F.A., C.E.D. and A.G.-C; Formal analysis, M.F.A., P.B., C.E.D., A.G-C.; Investigation, P.B., M.F.A, A.G.-C. and C.E.D.; Supervision, C.E.D. and A.G.-C.; Writing—Original draft, A.G-C. and C.E.D.; Writing—Review & editing, C.E.D. and A.G.-C. Funding acquisition, A.G.-C. and C.E.D.; All authors have read and agreed to the published version of the manuscript.
Funding
This work has been supported by grant CTQ2015-64049-C3-1R, MINECO/FEDER, Spain.
Acknowledgments
We gratefully acknowledge the deceased Prof. B.M. Fraga for his scientific contribution.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ayushi, S.; Kaushik, N.; Sharma, A.; Marzouk, T.; Djébali, N. Exploring the potential of endophytes and their metabolites for biocontrol activity. Biotech. 2022, 12, 277. [Google Scholar] [CrossRef]
- Morales-Sánchez, V.; Andres, M.F.; Díaz, C.E.; González-Coloma, A. Factors affecting the metabolite productions in endophytes: biotechnological approaches for production of metabolites. Curr Med Chem. 2020, 27, 1855–1873. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: unique plant inhabitants with great promises. Appl Microbiol Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef] [PubMed]
- Kondraskov, P.; Schütz, N.; Schüßler, C.; De Sequeira M., M.; Guerra A., S.; Caujapé-Castells, J.; Jaén-Molina, R.; Marrero-Rodríguez, A.; Koch, M.A.; Linder, P.; Kovar-Eder, J.; Thivet, M. Biogeography of mediterranean hotspot biodiversity: Reevaluating the tertiary relict’ hypothesis of Macaronesian laurel forests. PLoS ONE 2015, 10, 0132091. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.F.; Díaz, C.E.; Giménez, C.; Cabrera, R.; González-Coloma, A. Endophytic fungi as novel sources of biopesticides: the Macaronesian laurel forest, a case study. Phytochem Rev. 2017, 16, 1009–1022. [Google Scholar] [CrossRef]
- Fraga, B.M.; Terrero, D.; Gutiérrez, C.; González-Coloma, A. Minor diterpenes from Persea indica: Their antifeedant activity. Phytochemistry 2001, 56, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Fraga, B.M.; Terrero, D.; Bolaños, P.; Díaz, C.E. Diterpenes with new isoryanodane derived skeletons from Persea indica. Tetrahedron Lett. 2017, 58, 2261–2263. [Google Scholar] [CrossRef]
- Fraga, B.M.; Díaz, C.E.; Bolaños, P.; Bailén, M.; Andres, M.F.; González-Coloma, A. Alkane, alkene, alkyne-γ-lactones and ryanodane diterpenes from aeroponically grown Persea indica roots. Phytochemistry. [CrossRef]
- Fraga, B.M.; González-Coloma, A.; Gutiérrez, C.; Terrero, D. Insect antifeedant isoryanodane diterpenes from Persea indica. J. Nat. Prod. 1997, 60, 880–883. [Google Scholar] [CrossRef]
- González-Coloma, A.; Terrero, D.; Perales, A.; Escoubas, P.; Fraga, B.M. Insect antifeedant ryanodane diterpenes from Persea indica. J. Agric. Food Chem. 1996, 44, 296–300. [Google Scholar] [CrossRef]
- González-Coloma, A.; Andres, M.F.; Contreras, R.; Zúñiga, G.E.; Díaz, C.E. Sustainable production of insecticidal compounds from Persea indica. Plants 2022, 11(3), 418. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Heerklotz, K.; Drandarov, K.; Heerklotz, J.; Hesse, M.; Werner, C. Guignardic acid, a novel type of secondary metabolite produced by the endophytic fungus Guignardia sp., isolation, structure elucidation and asymmetric synthesis. Helv. Chim. Acta 2001, 84, 3766–3772. [Google Scholar] [CrossRef]
- Molitor, D.; Liermann, J.C.; Berkelmann-Löhnertz, B.; Buckel, I.; Opatz, T.; Thines, E. Phenguignardic acid and guignardic acid, phytotoxic secondary metabolites from Guignardia bidwellii. J. Nat. Prod. 2012, 75, 1265–1269. [Google Scholar] [CrossRef]
- Ma, K.L.; Wei, W.J.; Li, H.Y.; Son, Q.Y.; Dong, S.H. Meroterpenoids with diverse ring systems and dioxolanone-type secondary metabolites from Phyllosticta capitalensis and their phytotoxic activities. Tetrahedron 2019, 75, 4611–4619. [Google Scholar] [CrossRef]
- Andernach, L.; Sandjo, L.P.; Buckel, I.; Thines, E.; Opatz, T. Assignment of configuration in a series of dioxolanone-type secondary metabolites from Guignardia bidwellii. Eur. J. Org. Chem. 2013, 5946–5951. [Google Scholar] [CrossRef]
- Buckel, I.; Molitor, D.; C. Liermann, J.; Sandjo, L.; Berkelmann-Löhnertz, B.; Opatz, T.; Thines, E. Phytotoxic dioxolanone-type secondary metabolites from Guignardia bidwellii. Phytochemistry 2013, 89, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.Q.; Lin, Z.; Wang, Y.; Wang, J.; Zhou, X.; Yang, Bi; Liu, J.; Yang, X.; Wang, Y.; Liu, Y. New phenyl derivatives from endophytic fungus Aspergillus flavipes AIL8 derived from mangrove plant Acanthus ilicifolius. Fitoterapia 2014, 95, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.G.; Zhao, H.; Li, J.J.; Chen, S.M.; Mou, L.M.; Zou, J.; Chen, G.D.; Qin, S.Y.; Wang, C.X.; Hu, D.; Dao, X.S.; Gao, H. Phyllomeroterpenoids A-C, multi-biosynthetic pathway derived meroterpenoids from the TCM endophytic fungus Phyllosticta sp. and their antimicrobial activities. Sci. Rep. 2017, 7, 12925. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.H.; Liu, M.; Jiang, N.; Guo, Z.K.; Ma, J.; Zhang, J.; Song, Y.C; Tan, R.X. Guignardones A-C: three meroterpenes from Guignardia mangiferae. Eur. J. Org. Chem. 2010, 6348–6353. [Google Scholar] [CrossRef]
- Guimaräes, D.; Lopes, N.; Pupo, M. Meroterpenes isolated from the endophytic fungus Guignardia mangiferae. Phytochemistry Lett. 2012, 5, 519–523. [Google Scholar] [CrossRef]
- Sommart, U.; Rukachaisirikul, V.; Trisuwan, K.; Tadpetch, K.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Tricycloalternarene derivatives from the endophytic fungus Guignardia bidwellii PSU-G11. Phytochemistry Lett. 2012, 5, 139–143. [Google Scholar] [CrossRef]
- Kobayashi, T.; Takizawa, I.; Shinobe, A.; Kawamoto, Y.; Abe, H.; Ito, H. Asymmetric synthesis and structure revision of guignardone H and I: development of a chiral 1,3-diketone possessing C2 symmetry. Org. Lett. 2019, 21, 3008–3012. [Google Scholar] [CrossRef]
- Mei, W.L.; Zheng, B.; Zhao, Y.X.; Zhong, H.M.; Chen, X.L.W.; Zeng, Y.B.; Dong, W.H; Huang, J.L.; Proksch, P.; Dai, H.F. Meroterpenes from endophytic fungus A1 of mangrove plant Scyphiphora hydrophyllacea. Mar. Drugs 2012, 10, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Fraga, B.M.; Díaz, C.E. Proposal for structural revision of several disubstituted tricycloalternarenes. Phytochemistry, 1132. [Google Scholar] [CrossRef]
- Buijs, V.A.; Groenewald, J.Z.; Haridas, S.; LaButti, K.M.; Lipzen, A.; Martin, F.M.; Barry, K.; Grigoriev, I.V.; Crous, P.W.; Seidl, M.F. Enemy or ally: a genomic approach to elucidate the lifestyle of Phyllosticta citrichinaensis. G3 2022, 12, jkac061. [Google Scholar] [CrossRef] [PubMed]
- Wikee, S.; Udayanga, D.; Crous, P.W.; Chukeatirote, E.; McKenzie, E.H.C.; Bahkali, A.H.; Dai, D.; Hyde, K.D. Phyllosticta, an overview of current status of species recognition. Fungal Diversity 2011, 51, 43–61. [Google Scholar] [CrossRef]
- Thambugala, K.; Daranagama, D.; Kannangara, S. Biocontrol potential of endophytic fungi in tea (Camellia sinensis (L.) Kuntze) leaves against selected fungal phytopathogens. Malaysia. J. Microbiol. 2022, 18, 665–669. [Google Scholar] [CrossRef]
- Yan, X.-N.; Sikora, R.A.; Zheng, J.-W. Potential use of cucumber (Cucumis sativus L.) endophytic fungi as seed treatment agents against root-knot nematode Meloidogyne incognita. J. Zhejiang Uni. Sci. B 2011, 12, 219–225. [Google Scholar] [CrossRef]
- Han, W.B.; Dou1, H.; Yuan, W.H.; Gong, W.; Hou, Y.Y.; Ng, S.W.; Tan, R-X. Meroterpenes with toll-like receptor 3 regulating activity from endophytic fungus Guignardia mangiferae. Planta Med. 2015, 81, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Li, T.X.; Yang, M.H.; Wang, X.B.; Wang, Y.; Kong, L.Y. Synergistic antifungal meroterpenes and dioxolanone derivatives from the endophytic fungus Guignardia sp. J. Nat. Prod. 2015, 78, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.H.; Liang, F.L.; Wu, W.; Chen, Y.C.; Pan, Q.L.; Li, H.H.; Ye, W.; Liu, H.X.; Li, S.N.; Tan, G.H.; Zhang, W.M. Guignardones P–S, new meroterpenoids from the endophytic fungus Guignardia mangiferae A348 derived from the medicinal plant Smilax glabra. Molecules 2015, 20, 22900–22907. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xiong, B.; Xu, J. Chemical investigation of secondary metabolites produced by mangrove endophytic fungus Phyllosticta capitalensis. Nat. Prod. Res. 2021, 35, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhao, S.; Gu, C.; Tian, K.; Wang, Z.; Liu, F.; Ye, Y. Antifungal meroterpenes and dioxolanone derivatives from plant-associated endophytic fungus Phyllosticta sp. WGHL2. Fitoterapia 2021, 148, 104778. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.-X.; Liu, W.-Q.; Shi, Z.-X.; Zhu, H.-Y.; Fan, S.-Q.; Zhang, J.; Liu, W.-Y.; Xu, L.-J.; Ren, Q.-J.; Feng, F.; Xu, J. Meroterpenoids with divers’ rings systems from Phyllosticta capitalensis and their anti-inflammatory activity. Phytochemistry 2024, 217. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.L.; Li, D.L.; Chen, Y.C.; Tao, M.H.; Zhang, W.M.; Zhang, D.Z. Secondary metabolites of endophytic Guignardia mangiferae from Smilax glabra and their antitumor activities. Chin. Tradit. Herb. Drugs 2012, 43, 856–860. [Google Scholar]
- Zhu, X.; Liu, Y.; Hu, Y.; Lv, X.; Shi, Z.; Yu, Y.; Jiang, X.; Feng, F.; Xu, J. Neuroprotective activities of constituents from Phyllosticta capitalensis, an endophyte fungus of Loropetalum chinense var. rubrum. Chem. Biodiversity 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Morales-Sánchez, V.; Díaz, C.E.; Trujillo, E.; Olmeda, S.A.; Valcarcel, F.; Muñoz, R.; Andrés, M.F.; González-Coloma, A. Bioactive metabolites from the endophytic fungus Aspergillus sp. SPH2. J. Fungi 2021, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Andrés, M.F.; Rossa, G.E., Cassel; Santana, O.; Díaz, C.E.; González-Coloma, A. Biocidal effects of Piper hispidinervum (Piperaceae) essential oil and synergism among its main components. Food Chem Toxicol. 2017, 109, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Dioxolanone compounds of Phyllosticta capitalensis.
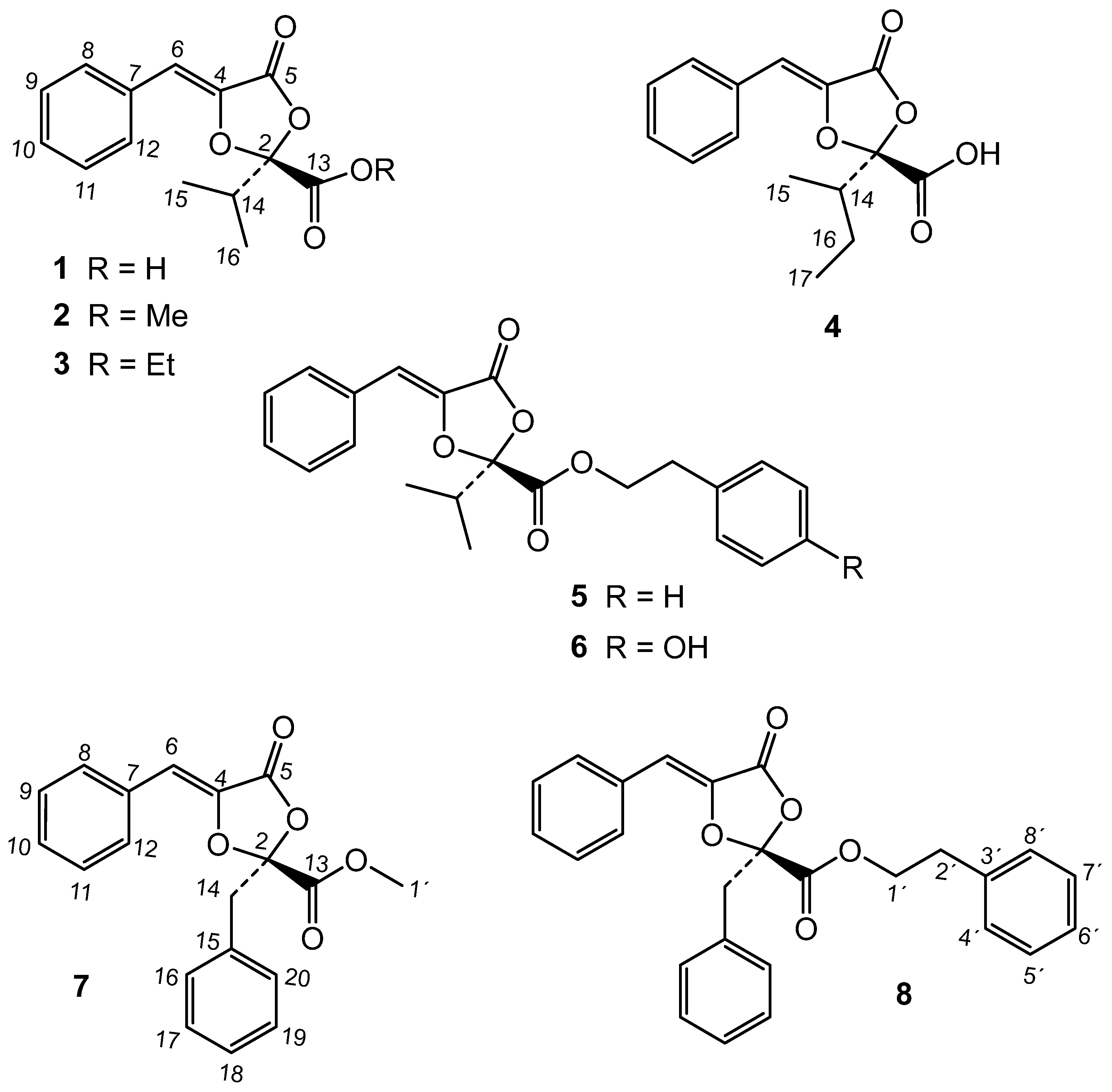
Figure 2.
Meroterpene compounds of Phyllosticta capitalensis.
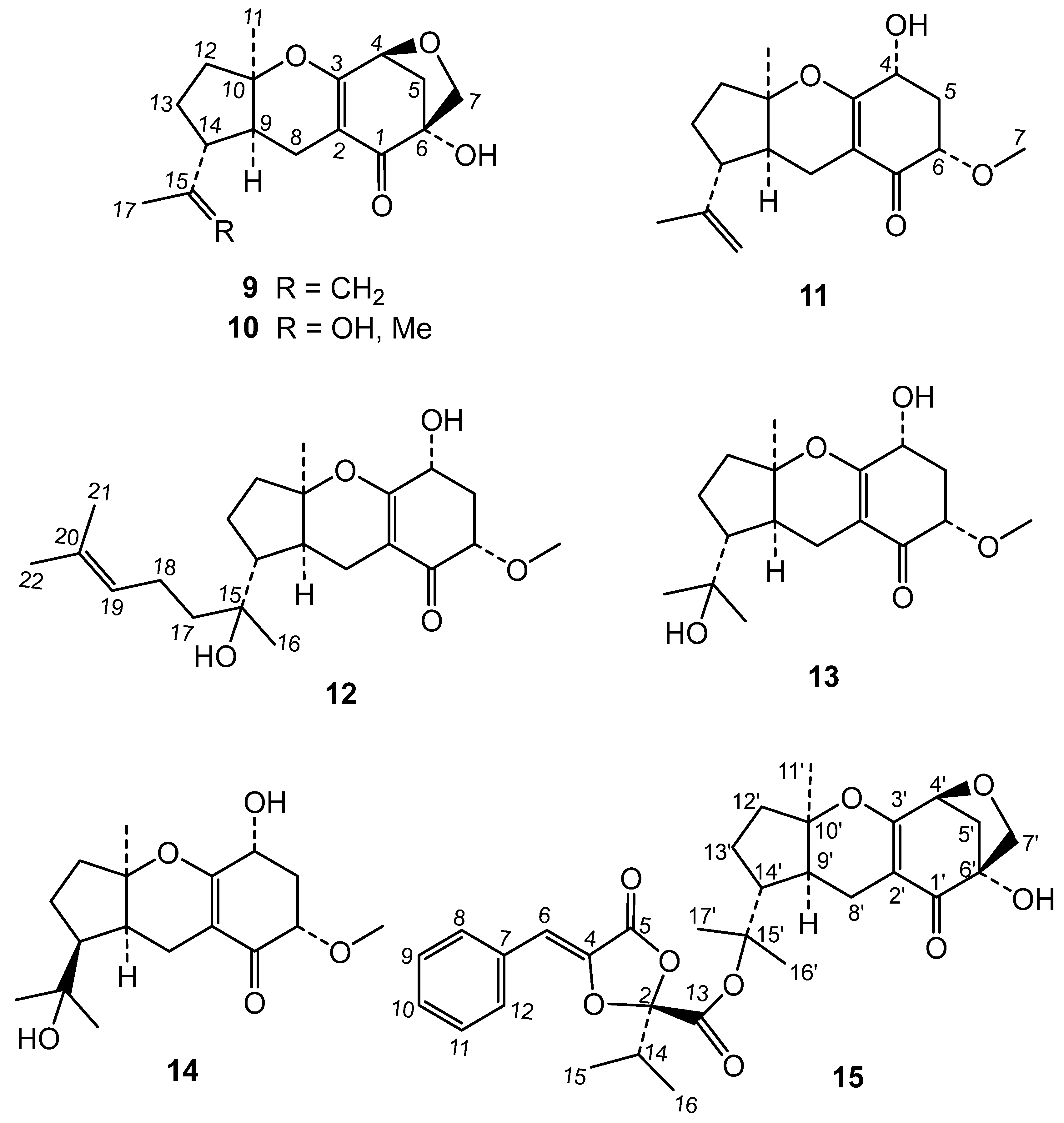
Figure 3.
Effects of compounds 4, 7 and 8 (extract in the background) on Meloidogyne javanica egg hatching. Time 0 d: 5 days of immersion in test solutions; time 7 d and subsequent: number of days of immersion in water after time 0 d. Each column represents the hatch inhibition rate for the respective treatment relative to the control.
Figure 3.
Effects of compounds 4, 7 and 8 (extract in the background) on Meloidogyne javanica egg hatching. Time 0 d: 5 days of immersion in test solutions; time 7 d and subsequent: number of days of immersion in water after time 0 d. Each column represents the hatch inhibition rate for the respective treatment relative to the control.
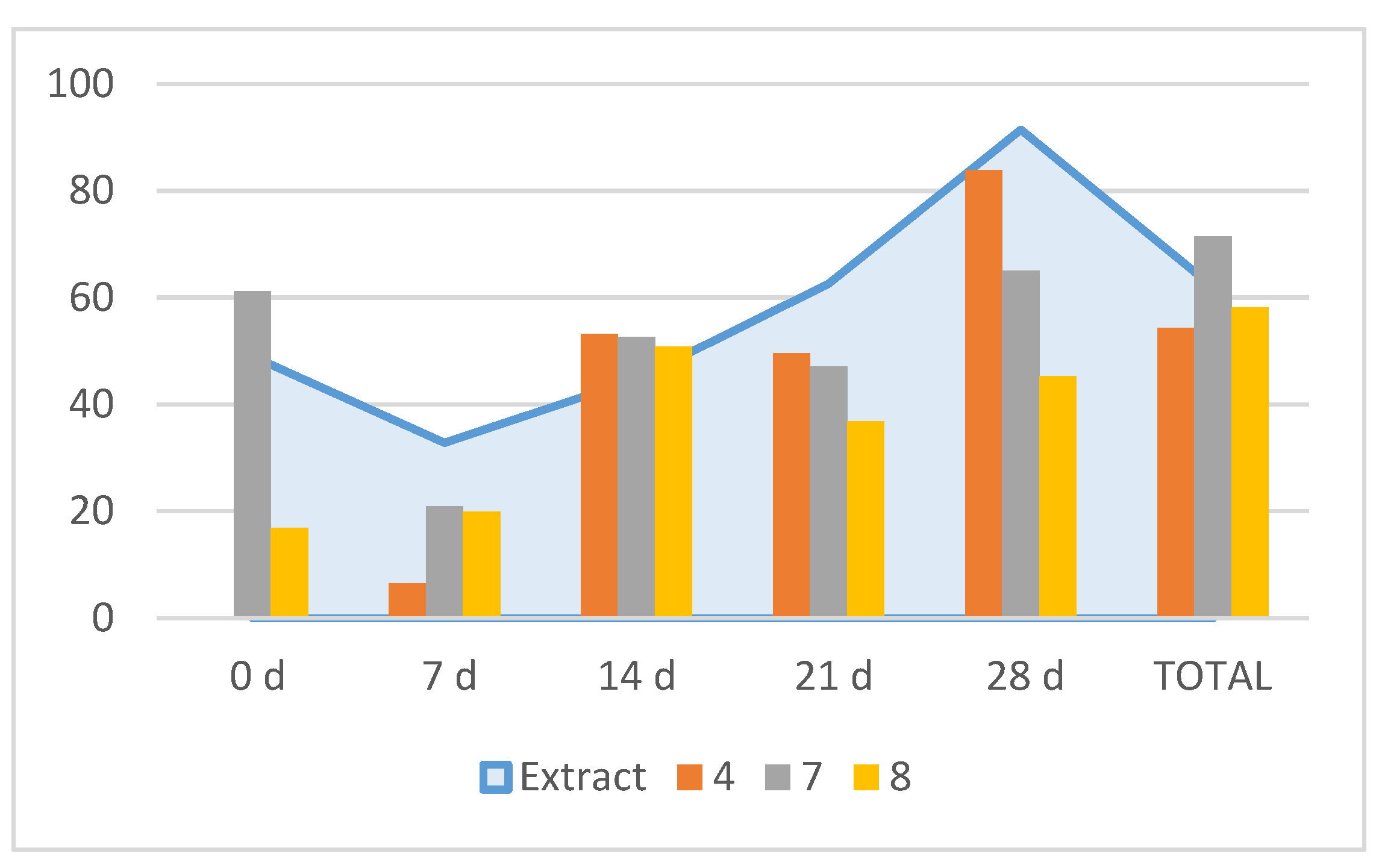

Table 1.
Summary of the bioactivities of the YCC4 extract on the insects (Spodoptera littoralis, Myzus persicae, Rhopalosiphum padi), the nematode Meloydogine javanica and the plant models Lolium perenne, Lactuca sativa and Solanum lycopersicum.
Table 1.
Summary of the bioactivities of the YCC4 extract on the insects (Spodoptera littoralis, Myzus persicae, Rhopalosiphum padi), the nematode Meloydogine javanica and the plant models Lolium perenne, Lactuca sativa and Solanum lycopersicum.
| Target | Action | Value |
|---|---|---|
| S. littoralis | Antifeedant | 9.77 (4.13-16.95)a |
| M. persicae | Antifeedant | 17.2 (4.13-23.16)a |
| R. padi | Antifeedant | >100 |
| M. javanica | J2 mortality | 0.44 (0.41-0.47)b |
| Egg hatching (28 days) | 64c | |
| L. perenne | Germination | 85.7±21.4d |
| Root growth | 74.4±7.1d | |
| Leaf growth | 65.8±7.8d | |
| L. sativa | Germination | 100d |
| Root growth | 155.3±22.9d | |
| S. lycopersicum | Germination | 85.7±21.4d |
| Root growth | 107.7± 13.1d |
aEffective dose EC50 (µg/cm2). bLethal concentration LC50 (mg/mL). cPercent inhibition. dPercent relative to the control.
Table 2.
In vivo effects of the Phyllosticta capitalensis extract on reproductive traits of Meloidogyne javanica in tomato plants, 60 days post inoculation, with 2000 eggs per plant, maintained in a growth chamber.
Table 2.
In vivo effects of the Phyllosticta capitalensis extract on reproductive traits of Meloidogyne javanica in tomato plants, 60 days post inoculation, with 2000 eggs per plant, maintained in a growth chamber.
| Treatment | egg masses/planta | RSb % |
eggs /plant x100 |
RSc % |
IFd | MRe |
|---|---|---|---|---|---|---|
| Extract | 47.4±6.2a | 66.6 | 335 ± 5.7a | 60 | 0.0237a | 16.6a |
| Control | 142 ± 22.5b | 832 ± 5.5b | 0.71b | 41.2b |
1 a Values are mean ±standard error of ten replicated plants. Values within the same column followed by different lower-case letter are significantly different according to Least Significant. b Relative suppression on eggs masses Difference (LSD) test (P <0.05). c Relative suppression on number of eggs per plant. d Infection Frecuency: egg masses per plant/egg inoculum. e Multiplication Rate: eggs per plant/egg inoculum.
Table 3.
13C NMR data of compounds 1-4, 7-8 and 11-14.
| Carbon | 1 | 2 | 4 | 7 | 8 | 11 | 13 | 14 |
| 1 | 195.0 | 194.9 | 194.6 | |||||
| 2 | 109.6 | 108.5 | 108.3 | 105.2 | 105.2 | 105.9 | 105.5 | 107.8 |
| 3 | 167.7 | 168.4 | 167.5 | |||||
| 4 | 136.0 | 135.7 | 135.3 | 135.3 | 135.2 | 65.7 | 65.8 | 66.0 |
| 5 | 163.5 | 162.8 | 162.7 | 162.3 | 162.3 | 34.5 | 34.6 | 34.8 |
| 6 | 109.4 | 109.6 | 110.1 | 109.5 | 109.6 | 79.1 | 79.1 | 79.0 |
| 7 | 132.2 | 132.3 | 132.1 | 132.2 | 132.2 | 58.4 | 58.4 | 58.5 |
| 8 | 129.9 | 129.9 | 129.9 | 129.8 | 129.8 | 16.1 | 18.7 | 16.1 |
| 9 | 128.7 | 128.8 | 128.8 | 128.8 | 128.7 | 43.2 | 41.3 | 40.8 |
| 10 | 129.0 | 129.1 | 129.2 | 129.8 | 129.0 | 87.7 | 88.9 | 86.3 |
| 11 | 128.7 | 128.8 | 128.8 | 128.8 | 128.7 | 22.3 | 22.2 | 30.5 |
| 12 | 129.9 | 129.9 | 129.9 | 129.0 | 129.8 | 37.4 | 38.4 | 34.7 |
| 13 | 169.5 | 166.0 | 169.5 | 165.2 | 165.8 | 26.9 | 24.6 | 18.9 |
| 14 | 33.1 | 33.0 | 39.3 | 40.7 | 40.8 | 48.9 | 51.1 | 49.6 |
| 15 | 14.4 | 14.5 | 11.1 | 130.6 | 130.6 | 145.5 | 73.0 | 71.8 |
| 16 | 15.2 | 15.2 | 22.4 | 131.0 | 131.0 | 111.2 | 27.7a | 27.4b |
| 17 | 11.5 | 128.6 | 128.5 | 19.2 | 28.7a | 29.5b | ||
| 18 | 127.8 | 127.9 | ||||||
| 19 | 128.6 | 128.5 | ||||||
| 20 | 131.0 | 131.0 | ||||||
| 1′ | 53.4 | 67.4 | 53.6 | |||||
| 2′ | 34.8 | |||||||
| 3′ | 136.8 | |||||||
| 4′/8′ | 128.7 | |||||||
| 5′/7′ | 128.4 | |||||||
| 6′ | 126.8 |
a,b These carbons can be interchanged.
Table 4.
Insect antifeedant effects of compounds 1-5, 7-12 and 14-15 tested at 50 µg/cm2 (data is expressed as average ± standard error) and effective doses EC50.
Table 4.
Insect antifeedant effects of compounds 1-5, 7-12 and 14-15 tested at 50 µg/cm2 (data is expressed as average ± standard error) and effective doses EC50.
| Chemical class | Compound |
S.littoralis %FIa± SE |
EC50c (µg/cm2) |
M. persicae %SIb ± SE |
EC50c (µg/cm2) |
|---|---|---|---|---|---|
| Dioxolanone | 1 | 84.4 ± 4.9 | 9.6 (5.3-17.5) | 82.0 ± 5.0 | 2.9 (1.0-7.9) |
| 2 | 50.3 ± 17.9 | 82.9 ± 3.7 | 1.7 (0.9-3.4) | ||
| 3 | 40.8 ± 15.3 | 29.1 ± 8.0 | |||
| 4 | 94.5 ± 1.94 | 7.5 (5.4-10.4) | 86.6 ± 3.5 | 11.4 (8.3-15.8) | |
| 5 | 52.0 ± 13.6 | 31.5 ± 8.3 | |||
| 7 | 20.0 ± 8.8 | 64.5 ± 7.9 | |||
| 8 | 28.3 ± 14.1 | 50.7 ± 7.7 | |||
| Meroterpene | 9 | 41.3 ± 16.1 | 37.3 ± 7.9 | ||
| 10 | 7.3 ± 7.3 | 47.0 ± 7.3 | |||
| 11 | 60.5 ± 16.9 | 44.1 ± 7.3 | |||
| 12 | 56.7 ± 14.0 | 35.2 ± 8.3 | |||
| 14 | 53.7 ± 12.5 | 39.5 ± 8.0 | |||
| Phyllomeroterpenoid | 15 | 55.2 ± 16.3 | 52.2 ± 8.5 |
Table 5.
Nematicidal activity against Meloidogyne javanica J2 of compounds 1-5, 7-12 and 14-15 (data is expressed as average ± standard error) and lethal doses (LC50 and LC90).
Table 5.
Nematicidal activity against Meloidogyne javanica J2 of compounds 1-5, 7-12 and 14-15 (data is expressed as average ± standard error) and lethal doses (LC50 and LC90).
| Compounda | Meloidogyne javanica (%)b | ||||
|---|---|---|---|---|---|
| 24 hc | 48 hc | 72 hc | LC50 mg/mLd (95% CL) |
LC90 mg/mLd (95% CL) |
|
| 1 | 60.98±3.42 | ||||
| 2 | 12.11 ±0.32 | ||||
| 3 | 8.60 ±2.81 | ||||
| 4 | 86.68 ± 0.95 | 88.30 ± 2.51 | 100.0 ± 0.0 | 0.24 (0.23-0.25) | 0.40 (0.39-0.43) |
| 5 | 5.78 ± 1.37 | ||||
| 7 | 40.98 ± 2.54 | 97.13 ± 0.51 | 100.0 ± 0.0 | 0.23 (0.22-0.24) | 0.43 (0.41-0.45) |
| 8 | 94.23 ± 3.59 | 100 ± 0.00 | 100.0 ± 0.0 | 0.11 (0.10-0.11) | 0.19 (0.18-0.20) |
| 9 | 49.09±0.78 | ||||
| 10 | 0.00 ±0.18 | ||||
| 11 | 0.00±0.63 | ||||
| 12 | 0.05±0.37 | ||||
| 14 | 0.00± 0.29 | ||||
| 15 | 7.45±1.79 | ||||
aValues (%) are means of four replicates. bTested at 0.5 mg/mL. cIncubation time. d Lethal doses (LC50 and LC90) and 95% confidence limits (CL).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



