
Submitted:
30 July 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
This study evaluated the effect of pig diet supplementation with an Olea europaea L. extract on the muscle fatty acid composition. Ten commercial hybrid pigs (Large White x (Landrace x Duroc)) were randomly assigned to two experimental treatments and fed either a standard diet (C) or diet with olive leaves extract (OL). For 90 days, the OL group received the olive leaf extract (300 mg/head/day), which mainly comprises oleuropein diglucoside, luteolin-7-glucoside, and verbascoside. The phenolic content, antioxidant activity, bioactive molecules’ characterization, and the fatty acids profile of meat and diet were performed. The results indicated that while the OL supplementation did not significantly alter the overall chemical composition of the meat or the pigs' performance, it did modify the lipid profile of the meat. The OL diet results in a significant reduction in SFA and an increase in oleic acid in meat, which correspondingly elevates the total MUFA content. Dietary treatment reduced the sum of n-6 PUFA in the meat of the OL group, and resulted in a significantly lower n-6/n-3 PUFA ratio. Therefore, these findings suggest that incorporating olive leaf extract into the pig diet enhances the nutritional and functional quality of the meat by improving its fatty acid profile.
Keywords:
Olea europaea L. polyphenols
; fatty acids
; olive leaves
; functional feed
; pigs
; antioxidants
; bioactive molecules
; meat
; pig performances
; Hedysarum coronarium.
1. Introduction
In recent years, the principles shown in the European Commission's action plan (2006) on reducing the use of antibiotics in animal breeding and considering ecological transition approaches, to transform 'no-food' matrices into important resources and not waste, stimulate scientific research into innovative solutions for a better balance between the environment, food systems, biodiversity, and circularity of resources [1]. Specifically, the use of agro-industrial coproducts in animal feed, from the olive oil sector, which holds significant economic and social importance in the Mediterranean area [2], offers a promising natural alternative. Due to their high polyphenol content, their co-products can enhance performance parameters, improve animal welfare, and preserve food quality, especially meat and meat products [3,4,5,6].
Olive oil co-products, such as olive leaves, are rich in polyphenols, natural compounds with high biological value. When included in the diets of monogastric animals (pigs), these compounds exhibit various beneficial effects, including anti-inflammatory, antioxidant, antibacterial, antiproliferative, and antifungal activities [7,8,9]. Among the bioactive molecules found in olive leaves, oleuropein, and hydroxytyrosol are the most abundant [10] and notable for their beneficial properties. Oleuropein is known to localize within biomembranes and alongside hydroxytyrosol, acts as a scavenger of peroxyl radicals near the membrane surface, while also interfering with their chain propagation [11]. Paiva-Martins et al. [12] highlight that nutritional interventions involving the use of bioactive molecules extracted from plant matrices, olive supply chain co-products, are crucial for enhancing the fatty acid profile, antioxidant content, and lipid stability of pig meat. In this context, feeding animals with natural antioxidants could be used as a vehicle for these compounds in the circulatory system, facilitating their distribution and retention in tissues, thereby positively influencing meat quality characteristics [13].
Incorporating a phenolic extract with high antioxidant content into the diet, in association with bioactive molecules from pasture grasses such as Sulla, represents a novel strategy in farm animals, to improve their growth performance, animal welfare, and production quality. This approach aligns with the goals of sustainable agriculture and animal breeding. According to the literature, the consumption of pasture grasses rich in antioxidants, especially vitamins, carotenoids, and flavonoids can protect meat against oxidation, thereby improving meat quality and animal welfare [14]. Specifically, Sulla (H. coronarium L.), a short-lived perennial legume native to the Mediterranean area and cultivated as a biennial forage for grazing and/or hay or silage production, has demonstrated positive effects on the productivity of several animal species. Its high fodder quality, palatability and beneficial impacts on livestock make it a valuable component in animal nutrition [15,16,17].
The positive effects of this plant species can be attributed to its high protein content, substantial levels of secondary metabolites (phenolic compounds), favorable ratio of degradable to structural carbohydrates, presence of proanthocyanidins, and moderate content of condensed tannins (CT). These attributes vary significantly according to species and growth stages [15,17,18]. The intake of CT is associated with the transfer of phenolic compounds to various tissues thus contributing to an increase in the antioxidant capacity and oxidative stability of animal products [19,20]. Pasture grasses like Sulla, offer high functional value in the diet, boosting the overall beneficial activity [21]. Rodríguez-Hernández et al. [18] highlight the antiparasitic effect of this species due to its condensed tannins content (20-110 g/kg dry matter) and phenolic compounds belonging to the flavonoid group type and contained in various parts of the plant. The beneficial effects of these molecules depend on their physical and chemical structure and their dietary content. Notably, high levels of condensed tannins (60-120 g/kg dry matter) can have adverse effects on non-ruminant livestock [14]
Therefore, the presence of secondary metabolites in this plant species could offer valuable support to limit drug use and improve livestock health [14]. Given the lack of studies in pigs on the combined effects of Sulla intake and antioxidant compounds extracted from olive oil supply chain coproducts, this study aims to address this gap. The objective is to formulate a feed functionalized with molecules from O. europaea L. for pigs, combined with a diet incorporating pasture grasses, to enhance precision feeding. This perspective aligns with the European Union plans and programs, including the One Health holistic approach, the European Green Deal, and Farm To Fork strategies which emphasize sustainable and integrated food systems.
In recent years, there has been a growing interest among consumers not only for the nutritional but also the functional properties of animal-derived foods destined for human consumption, particularly meat. Specifically, the fatty acid composition of meat influences several quality parameters, such as its nutritional and functional quality, shelf-life, and rheological characteristics [12,17]. The composition and content of the fatty acids in pig meat have garnered attention due to their impact on human health [22]. Recently, the presence of a high saturated fatty acid (SFA) content in meat has driven scientific research toward identifying and supplementing livestock diets with natural bioactive molecules, rich in antioxidants, and possessing a well-balanced lipid profile [23]. In this regard, incorporating olive oil coproducts into farm animal diets has demonstrated improvements in animal welfare, shelf life, and quality of animal products attributed to their polyunsaturated fatty acid (PUFA) and polyphenol content [24,25,26,27].
Several studies highlighted the benefits of incorporating long-chain n-3 PUFAs into the diet, establishing an optimal n-6/n-3 PUFA ratio of 3:1/4:1 to improve the health status not only of animals but also of humans as end consumers [12]. Indeed, it has been shown that fatty acid content and composition are directly influenced by dietary intake [28,29]. Considering this, Ponnampalam et al. [28] emphasize that diet plays a pivotal role in lipid metabolism and fatty acid (FA) synthesis in both animals and humans, and the effects of fatty acids are determined by their energy concentration and type. In addition, pasture- and forage-based diets are rich in omega-3 (n-3) short-chain PUFAs, while concentrated feed diets are common sources of omega-6 (n-6) short-chain PUFAs. The presence of n-3 and n-6 PUFAs in animals results from their direct consumption (feed) or the synthesis of longer-chain PUFAs from short-chain precursors in the body through desaturation and elongation processes. The consumption of n-3 PUFAs is known to improve health and welfare status of both animals and humans due to their various biological, biochemical, pathological, and pharmacological effects. In contrast, high levels of n-6 PUFA consumption can be potentially harmful [28].
Based on these considerations, we hypothesize that supplementing pig diets with molecules derived from the olive oil supply chain can improve the fatty acid profile of pigs without altering their growth performance.
2. Materials and Methods
Animal procedures were reviewed and approved by the Ethical Animal Care and Use Committee of the University of Naples “Federico II” (Protocol No. 99607-2017).
2.1. Plant Samples
Olive leaves of the Ortice cultivar were collected in March 2024 in an olive grove located in the Valley of the Middle Volturno in the Campania Region (Italy) (41°12′46″ N 14°24′07″E; 110 m above sea level in Ruviano (CE)). The choice of this cultivar is due to the high number of bioactive molecules with significant functional value [10]. Leaves were collected and were air-dried according to the procedure reported in Di Meo et al. [10]. Plants of Sulla (S, Hedysarium coronarium) in the vegetative state were collected from the “Carmine Campone” farm located in Castelpoto (Benevento, Italy). The plant samples were identified by Prof. Antonino Pollio of the University of Naples (Table 1).
2.2. Animals and Experimental Design
The study was conducted at the “Carmine Campone” farm located in Castelpoto (Benevento, Italy) (41°06′28′′N 14°40′18′′ E; 285 m above sea level) on ten commercial hybrid pigs (Large White x (Landrace x Duroc)) in the finishing period which was randomly assigned to two experimental diets. Each dietary group consisted of three replicates. The experimental treatments were: (1) a standard control diet (C); (2) a C diet supplemented with O. europaea L. leaves extract (300 mg/head/day) (OL). The study lasted 90 days. In the farm, 10 pigs were selected from the same group of animals, all fed equal amounts of the same standard diet. Animals were weighed (167.6 ± 8.38 kg initial body weight and 10 ± 1 months of age), individually identified, and allocated in individual pens. Specifically, all the pigs were fed twice daily (7.30 a.m. and 4.30 p.m.) during the study period with two dietary treatments formulated to contain: standard diet (control group (C); 5 pigs), standard diet, and olive leaves extract on the diet fed (experimental group (OL), 5 pigs). Feed and water were offered ad libitum during the study, with daily records and continuous access to fresh water. Pigs were weighed every 15 days from the beginning to the end of the experimental trial to determine the average daily gain (ADG). All the animals were slaughtered on the same day in a commercial abattoir, according to the European Union welfare guidelines (Council Regulation (EC) No. 1099/2009). Animals were electrically stunned and exsanguinated. The carcass weight was recorded, and 24 h after slaughter, Longissimus dorsi, diaphragm, Semimembranosus, and Psoas major muscles were removed from each carcass, then immediately transported to the laboratory and refrigerated for further analysis.
2.3. Pig Performance Assays
Pigs' weight and total dry matter intake were recorded for 90 days. Data on daily weight gain and feed conversion ratio were collected for the whole experimental period.
2.4. Standard Diet and Muscle Chemical Composition Analysis
Pigs were fed a standard diet containing maize, soya protein, wheat, barley, bran, and pasture grasses Sulla (H. coronarium L.), available for pasture-fed pigs. The diets were formulated to be isonitrogenous and isoenergetic. Individual feedstuff from each animal category was sampled weekly and analyzed according to the AOAC method [30]. The composition and amounts of the standard diet are shown in Table 2 and Table 3. The standard diet offered to the pigs of the OL group had the same ingredients as the diet supplied to the control group pigs (Table 2).
In addition, in the muscle samples, moisture, ash, crude protein, and crude fat were analyzed according to the AOAC methods [30].
2.5. Functional Diet: Preparation and Analysis
Dietary supplementation was carried out by adding dried olive leaves extract of the Ortice cultivar, in powder form, into the standard feed ration. The functional feed was administered to the pigs in the OL group at a concentration of 300 mg/head/day. Diet and dried olive leaf extracts were obtained by microwave-assisted extraction (MAE), following the method reported by Di Meo et al. [10,24,25]. In dried leaves, standard diet, enriched diet, and pasture grasses Sulla, total polyphenols (TPC) and flavonoids (TFC) content and the antioxidant activity were determined as reported in Di Meo et al. [10]; additionally, phenolic compounds in the olive leaves and Sulla extracts, were characterized by using HPLC-UV.
2.6. HPLC-UV Analysis
2.6.1. Chemicals
Authentic phenolic compounds: vanillic acid, gallic acid, tyrosol, 3-hydroxytyrosol, p-coumaric acid, caffeic acid, ferulic acid, ellagic acid, oleuropein, chlorogenic acid, kaempferol, quercitin, rutin, verbascoside, luteolin-7-glucoside, and oleuropein diglucoside (Figure 2) were purchased from Merck KGaA (Darmstadt, Germania). HPLC was used to check the purity of these compounds and one peak was given by each compound. All solvents used were of HPLC grade.
2.6.2. Preparation of Extracts with Different Solvents
Leaves of Ortice olive tree (25.0 g) and H. coronarium plants (25.0 g) were frozen at -80 °C, powdered in a mortar, and left to infuse for three days in different solvents to obtain the extract with hexane (H), methylene chloride (MC), acetone (A), methanol (M), and water (W) [31]. The organic extracts were filtered on Whatman paper n° 1, dried with Na2SO4, and concentrated under vacuum, yielding 0.4, 1.3, 1.0, and 2.2 g of residual material, respectively; instead, an aliquot (25 mL) of the aqueous fraction was lyophilized to obtain 0.3 g (2.4 g in total) of crude extract (Figure 1).
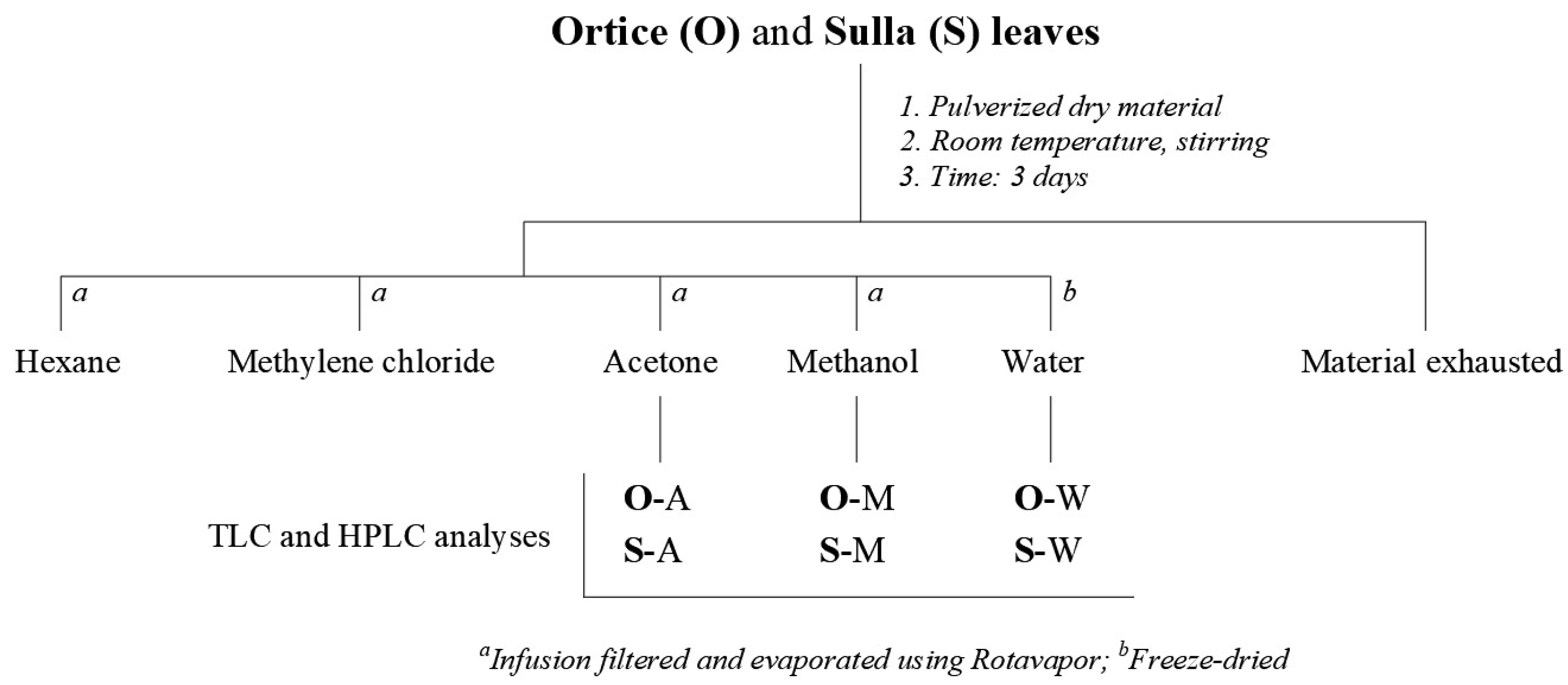
Figure 1.
Extraction of plant material.
The organic fractions A and M and the aqueous residue W obtained both from leaves of the olive tree Ortice and form plants H. coronarium were injected into HPLC obtaining the chromatographic profiles to be compared with the standards considered. The methanolic extracts (O-M and S-M) were the most interesting in terms of abundance and number of compounds identified by comparison with the reference standards and are illustrated in Figure 3.
2.6.3. General Experimental Procedures
The high-performance liquid chromatography (HPLC) apparatus consisted of a System Gold 127 Beckman pump, a System Gold 166 UV detector Beckman, and a Shimadzu Chromatopac C-R6A recorder. Analytical HPLC was performed using a Phenomenex column InertClone 5 um ODS(2) 150 A (150 x 4.60 mm), and eluted with HCOOH (A, 0.1 % in H2O) and acetonitrile (B), starting with 5 % B for 5 min and followed by the installation of a gradient to obtain 100 % B over 30 min, at a solvent flow rate of 1 mL/min. Analytical thin layer chromatography (TLC) was performed on Kieselgel 60 F254 or RP-18 F254 plates with 0.2 mm layer thickness (Merck). Spots were visualized by UV light or by spraying with H2SO4/AcOH/H2O (5:10:4). The plates were then heated for 3 min at 120 °C. Preparative TLC was performed on Kieselgel 60 F254 plates, with 0.5- or 1-mm film thickness (Merck). Flash column chromatography (FCC) was performed on Kieselgel 60, 230-400 mesh (Merck), at medium pressure. Column chromatography (CC) was performed on Kieselgel 60, 70-240 mesh (Merck).
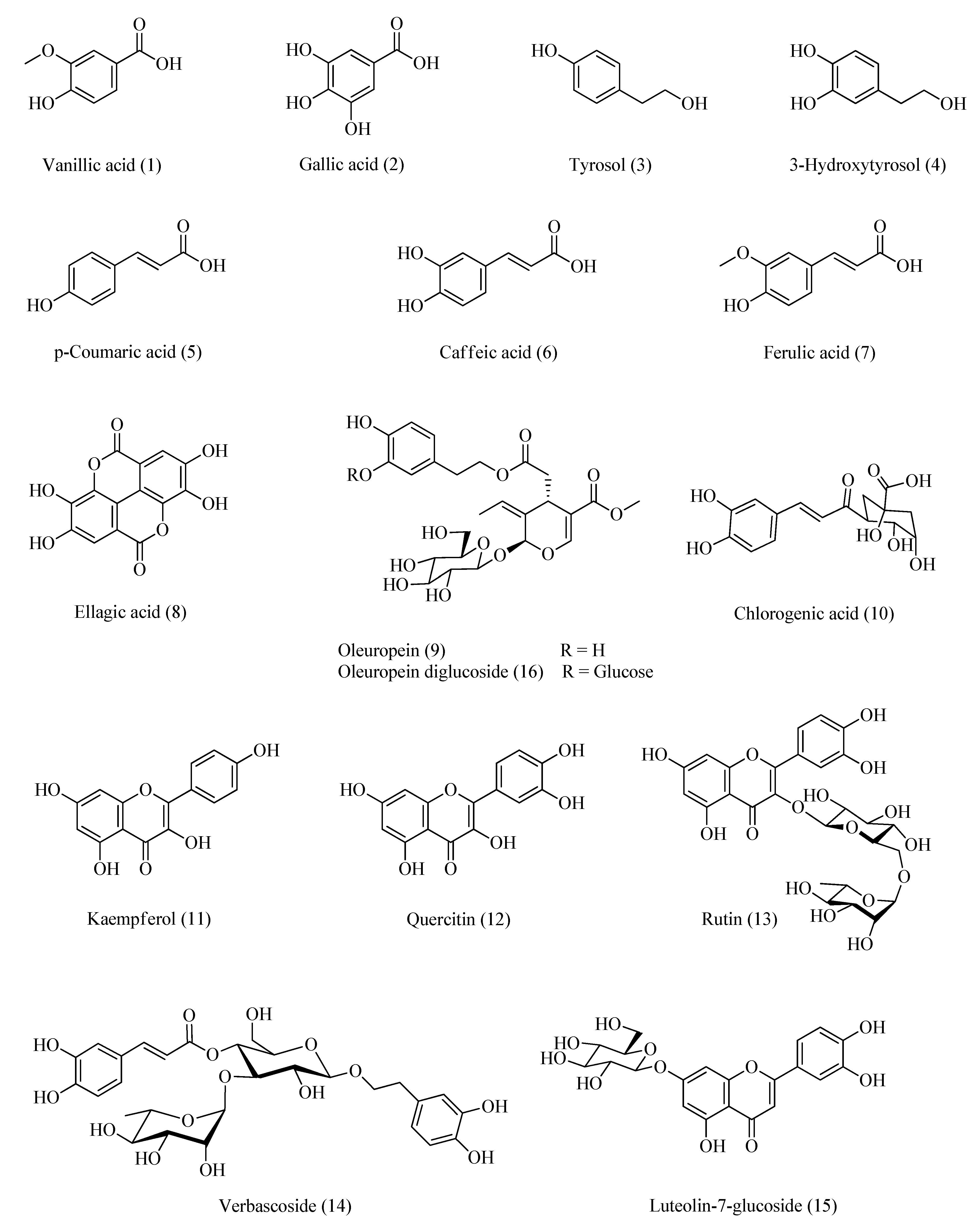
Figure 2.
Standards injected to HPLC.
2.7. Fatty Acids Analysis of Meat and Diet
Different types of pig muscles (Longissimus dorsi, Diaphragm, Semimembranosus, and Psoas major) were selected. A cold extraction protocol (an in-house method) was used for lipid extraction from food matrices. The homogenized and finely chopped sample is weighed, added to CH2Cl2, sonicated for 10 min, and placed under agitation for 2 days. So, the sample was filtered, dried over sodium sulfate, and subsequently evaporated to dryness under a nitrogen flow. Feed was extracted with methylene chloride and the organic phase was dried under a slight flow of nitrogen. The sample was dried for 1 h over P2O5 and then treated for 20 min at 60 °C with a solution of boron trifluoride/methanol 10% (1.3 M, 0.5 mL) and 100 µL of dimethoxypropane [32]. Finally, each solution was extracted twice with hexane, and the organic phase was dried. The fatty acid methyl esters were redissolved in hexane, filtered on a millet, and injected into a gas chromatograph [24,25].
The gas chromatographic analysis of the fatty acid profile involves the conversion of fatty acids into methyl esters (FAMEs). The transmethylation reaction in the presence of anhydrous methanol can occur with either basic or acidic catalysis. For basic transmethylation, an aliquot of the sample accurately weighed, is transesterified using the following protocol. The extracted fat is dissolved in n-hexane (containing internal standards), and then treated firstly with a 2N solution of KOH in methanol and then vortexed for 1 min at room temperature. The reaction is stopped after 5 min by adding NaHSO4·H2O. After phase separation, the supernatant (hydrocarbon phase), containing the methyl esters of fatty acids, is transferred into a vial and then injected into the gas chromatograph. The analysis is performed in triplicate. For acid transmethylation, an aliquot of methylating reagent (acetyl chloride slowly added to anhydrous methanol) containing internal standards is added to the previously separated lipid fraction. The transmethylation reaction takes place at 60 °C for 1 h with the reaction vials being agitated at 5-min intervals. After cooling, 2 mL of saturated aqueous solution in hexane and 5 mL of hexane is added, agitated for 1 min, and the hexane phase is recovered. The hexane phase is washed twice with 5 mL of saturated aqueous solution in hexane. The recovered organic phases are dried with 0.5 g of anhydrous sodium sulfate for 1 h. The hexane phase (supernatant) is transferred into a vial and injected into the gas chromatograph. The gas chromatography (GC) apparatus consisted of an Agilent GC 8890A instrument equipped with a split/splitless inlet and FID detector and an Agilent 7693A automatic liquid sampler. Analysis was performed using a capillary column Agilent HP-88, (100 m × 0.25 mm, 0.20 µm film thickness). The following parameters were set during the experiments: injected sample volume: 1 mL, introduced into injector using autosampler heated to 250 °C, with a split ratio of 100:1. Nitrogenous as the carrier gas, 40 psi, constant pressure mode. Oven ramp program: 100 °C (13 min), 10 °C/min to 180 °C (6 min), 1 °C/min to 200 °C (20 min), 4 °C/min to 230 °C (7 min) detector temperature 280 °C, with H2: 40 mL/min, air: 400 mL/min, make up gas: 25 mL/min. A mixture standard of 37 FAMEs was diluted to 50–100 ng/µL for each component and used to test system repeatability. All components were well resolved. The overlaid chromatograms from six injections showed excellent area and RT repeatability. The fatty acids were identified by comparing the retention times with those of reference standards and quantified relative to the internal standards.
2.8. Statistical Analysis
Statistical analysis was carried out using GraphPad Prism (GraphPad, San Diego, CA, USA) version 8.0 for Windows. Prior to statistical analysis, the normality of variable distributions was assessed and Levene’s test was applied to evaluate the homogeneity of variances. Data on pig meat chemical composition and animal performance were analyzed using a one-way ANOVA (Newman–Keuls post-test) to evaluate the effect of the dietary treatment. Data on function analysis of diet and to compare fatty acids of diet and meat with each other and the total content of saturated, monounsaturated, and polyunsaturated fatty acids were analyzed using a Two-way ANOVA analysis (Tukey test). The data are expressed as mean ± SEM and a p-value <0.05 was considered the minimum statistical significance.
3. Results
3.1. Pig Performances and Chemical Composition of Muscle
Table 4 shows the results obtained from the in vivo performance for both control and treated pigs. Statistical analysis reveals no significant differences between dietary treatments for the analyzed parameters, including final body weight (p = 0.261), carcass weight (p = 0.563), dry matter intake (DMI; p = 0.674), average daily gain (ADG; p = 0.633) and feed-conversion ratio (FCR; p = 0.105). Furthermore, no significant differences were observed in the chemical composition of the different muscles between the two groups (C and OL). The average values for moisture, raw protein, ether extract, and ash content are reported in Table 4 with no significant differences found (p > 0.05) across these parameters.
3.2. Functional Diet Analysis
3.2.1. Total Polyphenols Content, Total Flavonoid Content, and Antioxidant Activity
The results of total polyphenols and flavonoid content were quantified as mg gallic acid equivalent/g dry weight (mg GAE/g DW) and mg quercetin equivalent/g dry weight (mg QE/g DW), respectively. Antioxidant activity was assessed based on the percentage inhibition of free radicals. Table 5 details the total phenolic content and antioxidant activity of diet, Sulla, and olive leaf extract. The polyphenols and flavonoids content of the enriched diet (33.59 ± 0.88 mg GAE/g; 2.98 ± 0.32 mg QE/g) increased statistically significantly compared to the standard diet (8.14 ± 0.30 mg GAE/g; 1.71 ± 0.20 mg QE/g) (p < 0.05) and Sulla extract (17.47 ± 0.52 mg GAE/g; 3.02 ± 0.26 mg QE/g) (p < 0.001). Additionally, the antioxidant capacity of olive leaf extract (51.10 ± 2.98%), enriched diet (standard diet + Ortice olive leaf extract; 70.20 ± 2.36%), and Sulla (63.20 ± 2.00%) shows a statistically significant increase compared to the standard diet (28.21 ± 1.20%). These findings highlight the substantial functional benefits of the functional diet with O. europaea L. polyphenols due to its high polyphenols content and enhanced radical scavenging activity. Additionally, the data suggest that pasture grasses, such as Sulla, available to the pigs, contribute significantly to the diet’s phenolic compound content, underscoring their value as a natural resource.
3.2.2. HPLC-UV Analysis
The main compounds identified in the phenolic extracts (Ortice olive leaves and Sulla plant) are shown in Figure 3. The chromatographic profiles of the methanolic extracts of Ortice leaves (Figure 3A) highlighted the presence of several key phenolic compounds: tyrosol (3) and its 3-hydroxy derivative (4), acid p-coumaric (5), oleuropein (9) and its diglucoside (16), chlorogenic acid (10), two flavonoids quercetin (12) and luteolin-7-glucoside (15). Additionally, phenylpropanoid verbascoside (14) was also identifies [33]. The methanolic extract of the Sulla plant predominantly contains tyrosol (3) and its 3-hydroxy derivative (4), caffeic acid (6) and ellagic acid (8), and, finally, oleuropein (9) (Figure 3B).
Notably, Ortice olive leaf extract exhibited the highest relative concentration of verbascoside (14), luteolin-7-glucoside (15), and oleuropein diglucoside (16), while Sulla was particularly rich in caffeic (6) and ellagic acids (8). The relative concentrations of these phenolic compounds were determined by comparing retention times and absorption spectra with those of pure standards as shown in Table 6.
The chromatographic analysis highlights the qualitative molecular difference between Ortice olive leaf extract and Sulla; differences between highlights the Sulla and olive leaf extract, emphasizing the distinctive phenolic profiles of each. These findings underscore the potential benefits of supplementing pig’s diet with O. europaea extracts, particularly when combined with forage sources like Sulla. The data suggest that the bioactive molecules of Sulla associated with the phenolic compounds from the olive leaf extract significantly contribute to the enhancement of the diet’s antioxidant properties.
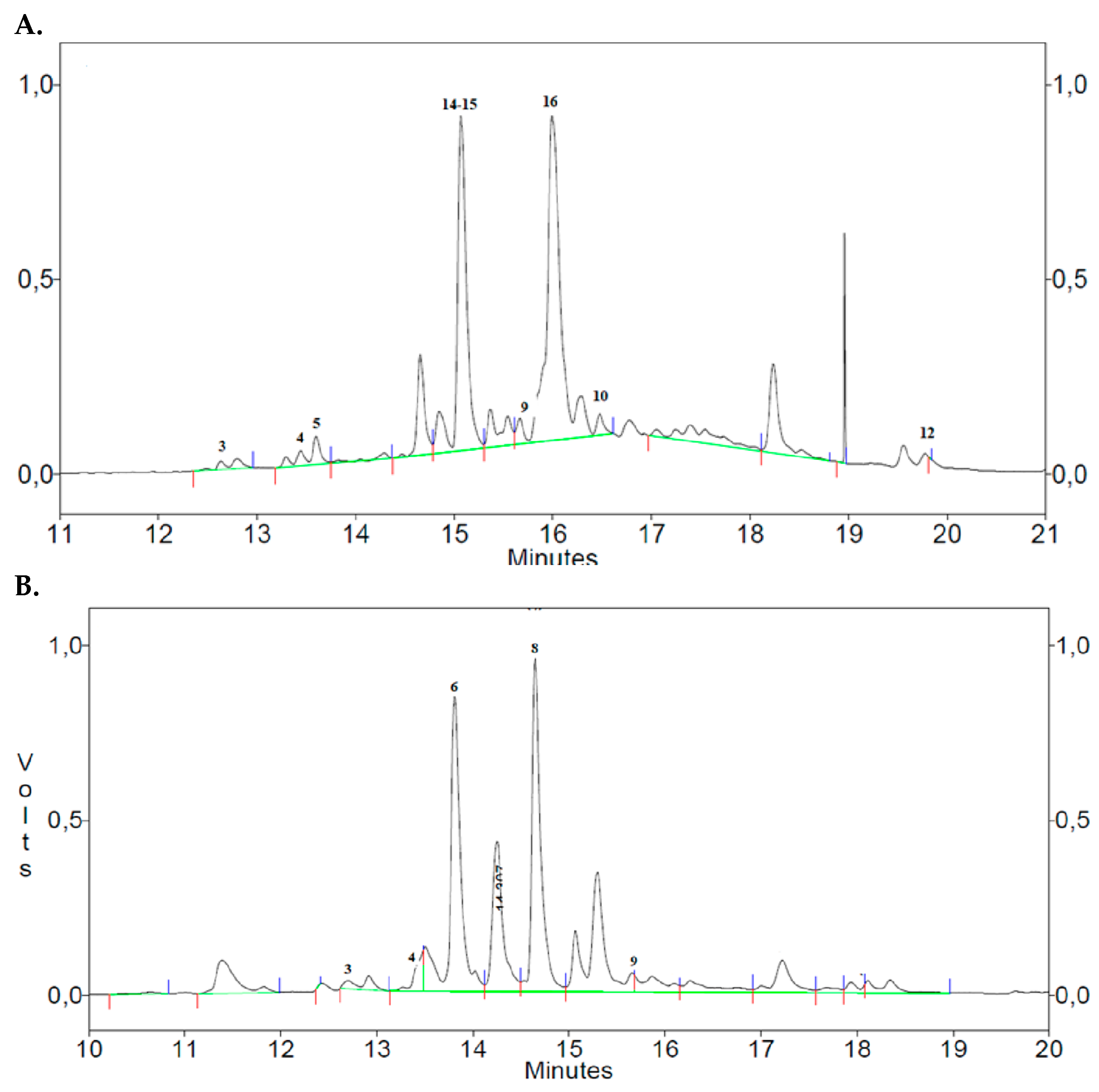
Figure 3.
Chromatographic profiles of methanol extracts of Ortice olive leaves (A) and plant Sulla (B).
Figure 3.
Chromatographic profiles of methanol extracts of Ortice olive leaves (A) and plant Sulla (B).
3.3. Fatty Acid Profile
3.3.1. Dietary Fatty Acid Profile
Table 7 shows the content of individual fatty acids in the standard diet, Ortice olive leaf extract, enriched diet, and Sulla plant extract. The data indicated that dietary supplementation with olive leaf extract and Sulla intake significantly reduced saturated fatty acid (SFA) content (p < 0.001). In addition, Ortice olive leaf extract was associated with a significant increase in monounsaturated fatty acids (MUFA) due to its high oleic acid content (C18:1 n9; p < 0.001)), while Sulla determined an increase in PUFA (p < 0.001) by balancing and significantly reducing the n-6/n-3 ratio (p < 0.001).
3.3.2. Muscle Fatty Acid Profile
The effects of the dietary treatment on individual and total fatty acids in pig muscles are shown in Table 8. In the present study, the supplementation of olive leaf extract resulted in a reduction of SFA and a statistically significant increase of MUFA (p < 0.001) across all muscle types. Furthermore, no significant differences were found in the fatty acid composition between the different muscles. Within the SFA category, the OL diet statistically significantly reduced the concentration of C18:0 (p < 0.001). In the MUFA class, it resulted in a significant increase in the concentration of C18:1n9 (p < 0. 001). Conversely, in the PUFA class, the OL diet led to significant reductions only for certain fatty acids, such as linoleic (C18:2n6; p < 0.05) and α-linolenic acid (C18:3n3; p < 0.05), while other fatty acids do not show significant changes. As a result, the MUFA/SFA ratio increased significantly (p < 0.001) with OL treatment compared to the control. The OL diet resulted in a statistically significant reduction of the total n-6 PUFA content in the meat (p < 0.001) and improved the n-6/n-3 ratio (p < 0.001) compared to the control diet.
4. Discussion
In this study, the use of dried olive leaves from the Ortice cultivar—native to the study region—was selected due to their high concentration of bioactive compounds and potent antioxidant activity [10]. Leaf drying decreases water activity, reduces the enzymatic destruction of certain polyphenols, and increases oleuropein synthesis [12,34]. Considering the high chemical stability of dry leaf powder when mixed with feed, the extract was provided to pigs throughout the experimental period as a dry powder mixed with the diet; it was mixed as a powder to prevent the animals from rejecting olive leaves in the diet [12]. The decision to supplement the pigs` diet with O. europaea molecules, in addition to molecules present in the pasture grasses (Sulla), resulted from a functional screening of the compounds and the analysis of the lipid profile of the extracts. The extract of O. europaea L. shows a high content of oleuropein, the most abundant compound in olive leaves, followed by 3-hydroxytyrosol, luteolin 7-O-glucosides, and verbascoside [35,36]. Pereira et al. [37] also identified oleuropein and lueolin-7-O-glucoside as the most abundant phenolic compounds in olive leaf extracts. In particular, these phenolic compounds are known to impart several beneficial effects in animals of zootechnical interest, including antioxidant, anti-inflammatory, antiproliferative, antibacterial, and antifungal properties [3,25,38].
Furthermore, Sulla, a fodder usually fed to ruminants [17,19], was found to be particularly appealing to the pigs in our study; it has a high content of condensed tannins with an anti-parasitic effect [18], and it also contains phenolic compounds with significant antioxidant capacity, including quercetin, kaempferol, myricetin and the isoflavone formononetin [15]. In addition, molecular characterization of the Sulla extract shows, consistent with findings by Molinu et al. [39], revealed the presence of phenolic acids such as caffeic and ellagic acid, known for their beneficial effects in zootechnical animals [40,41,42,43]. Wang et al. [41] point out that a diet rich in ellagic acid enhances antioxidant capacity, digestive enzyme activity, immune function, intestinal function, and growth performance in broilers. Similarly, caffeic acid has been shown to improve the intestinal barrier function, modulate gut microbiota and its metabolites, reduce inflammatory responses, and decrease oxidative stress, thereby enhancing growth performance in piglets [42]. Despite these benefits, there is a notable gap in the scientific literature regarding the antioxidant capacity and individual phenolic compounds quantification of the Sulla extract. Based on these, we aim to better understand the molecular profile of this short-lived perennial legume, native to southern Italy, and assess its potential benefits for pigs. Considering the several beneficial effects of the molecules of O. europaea extract and due to the diversity in the bioactive molecules composition of Sulla compared to these extracts, we decided to supplement the diet and formulate a functional feed to maximize the range of potential effects.
In this study, the supplementary level of 300 mg/head/day of O. europaea extract was selected based on prior research emphasizing its antioxidant properties [10,22]. Several studies report that the antioxidant effects are known to be influenced by the concentration and type of polyphenols integrated into the diet [5,23,44,45]. Our study confirmed that the polyphenol content in O. europaea L. extracts is consistent with previous reports [10,46]. Variations in phenolic composition among olive leaf extracts could be attributed to differences in the analytical extraction procedures and/or sample origin or harvesting period [10,36,47,48]. Specifically, the polyphenol content of the Ortice cultivar in our study (19.13 mg GAE/g) aligns with earlier findings indicating that the content for this cultivar averages around 16 mg GAE/g [10].
Furthermore, our findings show that supplementation with olive leaf extract provided no change in the growth performance of the pigs, as reported even by Leskovec et al. [49]. Similarly, Botsoglou et al. [47] reported that dietary supplementation with fish oil and olive leaves did not adversely affect pig growth rates; contrary to the results of Paiva-Martins et al. [27] who observed different outcomes with olive leaf supplementation. One of the most expected results of incorporating olive leaf into pig diets was its effect on the fatty acid profile of meat. In our study, positive effects were observed in the fatty acid composition showing a reduction in SFA, an increase in MUFA, and a positive balance of PUFA (Omega6 and Omega3). Specifically in our study, the enriched diet led to a reduction in n-6 PUFAs and an increase in n-3, mainly attributed to C18:2 n6 and C18:3 n3 PUFAs, respectively.
Wood and Enser [50] report that PUFAs, which cannot be synthesized, exhibit tissue concentrations that respond rapidly to dietary changes: in contrast, major SFAs and MUFAs, being synthesized endogenously, show less sensitivity to dietary variations. However, our study observed that increased dietary inclusion of olive leaf extract promoted a significant reduction in SFA content and a linear increase in the MUFA levels, in agreement with the findings of Joven et al. [51]. Joven et al. [51], also reported that diets enriched with olive cakes results in higher MUFA and lower PUFA concentrations in pork, consistent with our results. The increase in MUFA in pigs’ meat, mainly due to the high content of C18:1n9 in olive leaf extract (36.18 g/100 g), supports previous research indicating that dietary oleic acid can influence the quality of pork by improving its sensory characteristics and oxidative stability [12,47]. Numerous studies have reported effects in pigs [52], in broilers [53], and in rabbits [54]. Losacco et al. [54] evaluated the effects of the use of olive by-products as a dietary supplement for rabbit production and health highlighting an increase of MUFA in meat, especially oleic acid, showing that the content of intramuscular oleic acid and MUFA of rabbits fed different olive pomaces were proportional to the oleic acid content of the by-product. These effects are caused by the presence of oleic acid in the diet, as it is known that fatty acid deposition in pigs is mainly influenced by the lipid composition of the diet [55].
In the present study, the overall PUFA content in meat was largely unaffected by dietary treatment, except for the levels of C18:2n6 and C18:3n3, where significant variations were shown. Literature indicates that diets high in PUFAs can enhance meat`s susceptibility to oxidation [50]. This observation underscored the advantage of using O. europaea molecules, which are low in PUFAs, as they positively influence the lipid profile of the meat. However, Sarmiento-García et al. [56] emphasize that fatty acid content and composition could vary depending on multiple factors, such as the type of muscle analyzed, genetic variation (with important differences between commercial and indigenous pigs), age, or slaughter weight. Pugliese et al. [57] found that the age of pigs can affect the deposition of MUFA, with indigenous breeds showing an increased capacity for MUFA accumulation.
Supplementing pig diets with moderate levels of olive leaves, even at low dosages is a beneficial source of biologically active compounds and could increase phenolic content in meat [3]. In addition, it may also positively affect pig performance and fatty acid composition by decreasing the SFA content and increasing the MUFA, especially the proportion of oleic acid, which is also essential for human health [51].
This pilot study provides promising new perspectives on the use of functional feed on the fatty acid profile of pigs’ meat. Nevertheless, the limited sample size due to the low availability of animals on the farm constrains the generalizability of these findings. Consequently, further research with a larger number of pigs and varying concentrations of olive leaf extract is underway to provide a more comprehensive assessment of its beneficial effects.
5. Conclusions and Future Implications
This paper is the first study on the inclusion of a combined functional diet in the diet of finishing pigs. Our research highlights that: i) Sulla extract and olive leaf extract have a high antioxidant capacity and high polyphenol content; ii) the functional diet had no significant effects on the growth performance of pigs and the chemical composition of meat; iii) the functional diet improve the fatty acid profile of pigs’ meat.
Our results suggest that the combined use of fodder plants such as Sulla and bioactive molecules coming from olive leaf extracts in the pig feed ration contributes to guaranteeing suitable production performance and improving the meat fatty acid profile with the correct balancing of SFA, MUFA, and PUFA. In particular, this highlights that the use of O. europaea extracts in pig feeding can be practiced without negative effects on the animals' performance.
In conclusion, the introduction of sustainable pig feeding models could represent a valid strategy for the ecological transition of territories with large olive and cereal livestock production in inland Mediterranean areas.
A diet with Sulla fodder, cereals, and specific amounts of bioactive molecules from O. europaea, in addition to improving the lipid profile of pigs’ meat, offers further insights into the application of strategies for the recovery and use of beneficial molecules for the formulation of functional feeds in terms of resource circularity in olive-growing areas.
Finally, olive tree leaf extracts, due to their verified antioxidant, antimicrobial, and antiviral activity and as stimulators of the immune response, represent a great potential for industrial applications in various sectors, including the feed, pharmaceutical, and cosmetic industries, and are a valid support in reducing antibiotic resistance in animals of veterinary interest. This represents a real strategic approach to the recovery of an agro-industrial residue in line with the circular economy concept to stimulate the feed industry to produce functional feeds for large-scale distribution.
Based on these premises, the scientific community has started a large amount of research aimed at investigating the effects of these molecules on the functional quality, rheological characteristics, and oxidative stability of meat as well as the morpho-functional aspects of various organs and tissues involved in animal welfare. Further research is needed to confirm the results obtained in this study and to extend knowledge about the effects of feed functionalized with natural bioactive compounds.
Author Contributions
Conceptualization, M.C.D.M, M.R. an L.D.A.; methodology, M.C.D.M, I.L. and A.Z.; software, M.C.D.M.; validation, R.S., L.D.A., M.R. and E.V.; formal analysis, M.C.D.M. and I.L.; investigation, M.C.D.M. and E.V.; resources, E.V.; data curation, M.C.D.M., M.D.N. and A.R.B.; writing—original draft preparation, M.C.D.M.; writing—review and editing, P.V., A.Z. and E.V.; visualization, R.S., M.R. and L.D.A.; supervision, E.V. and P.D.G.; project administration, E.V.; funding acquisition, E.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project “RossoPiccante: Modelli innovativi di suinicoltura sostenibile per la produzione della salsiccia Rosso Piccante di Castelpoto”—PSR Campania 2014/2020 Misura 16.1–Azione 2–CUP B82C21000760002, and by the project “S.OIL: Strategie innovative di gestione sostenibile dell’olivicoltura sannita e valorizzazione delle molecole bioattive dei sottoprodotti e co-prodotti” - PSR Campania 2014/2020 Misura 16.1 -Azione 2–CUP B59H23000020006.
Institutional Review Board Statement
The animal study protocol was approved by the Ethical Animal Care and Use Committee of the University of Naples Federico II in Italy (Protocol No. 99607-2017). The authors confirm that the ethical policies of the journal, as noted on the journal’s author guidelines page, have been adhered to and the appropriate ethical review committee approval has been received, and confirm that they have followed EU standards for the protection of animals used for scientific purposes and feed legislation.
Informed Consent Statement
Animal procedures were reviewed and approved by the Ethical Animal Care and Use Committee of the University of Naples “Federico II” (Protocol No. 99607-2017).
Data Availability Statement
All the data are available in the manuscript.
Acknowledgments
We acknowledge the “Carmine Campone” farm (Castelpoto, Italy) for providing the pigs for the trial and AIPRAS Onlus (Associazione Italiana per la Promozione delle Ricerche sull’Ambiente e la Salute umana) for support of this investigation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Malenica, D.; Kass, M.; Bhat, R. Sustainable Management and Valorization of Agri-Food Industrial Wastes and By-Products as Animal Feed: For Ruminants, Non-Ruminants and as Poultry Feed. Sustainability 2023, 15, 117. [Google Scholar] [CrossRef]
- Hukerdi, Y. J.; Nasri, M. F.; Rashidi, L.; Ganjkhanlou, M.; Emami, A. Effects of dietary olive leaves on performance, carcass traits, meat stability and antioxidant status of fattening Mahabadi male kids. Meat Sci. 2019, 153, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Ferlisi, F.; Tang, J.; Cappelli, K.; Trabalza-Marinucci, M. Dietary supplementation with olive oil co-products rich in polyphenols: a novel nutraceutical approach in monogastric animal nutrition. Front. Vet. Sci. 2023, 10, 1272274. [Google Scholar] [CrossRef] [PubMed]
- Mallamaci, R.; Budriesi, R.; Clodoveo, M.L.; Biotti, G.; Micucci, M.; Ragusa, A.; Curci, F.; Muraglia, M.; Corbo, F.; Franchini, C. Olive Tree in Circular Economy as a Source of Secondary Metabolites Active for Human and Animal Health Beyond Oxidative Stress and Inflammation. Molecules 2021, 26, 1072. [Google Scholar] [CrossRef] [PubMed]
- Munekata, P. E. S.; Rocchetti, G.; Pateiro, M.; Lucini, L.; Domínguez, R.; Lorenzo, J. M. Addition of plant extracts to meat and meat products to extend shelf-life and health-promoting attributes: An overview. Curr. Opin. Food Sci. 2020, 31, 81–87. [Google Scholar] [CrossRef]
- Berbel, J.; Posadillo, A. Review and Analysis of Alternatives for the Valorisation of Agro-Industrial Olive Oil By-Products. Sustainability 2018, 10, 237. [Google Scholar] [CrossRef]
- Mahfuz S, Shang Q, Piao X. Phenolic compounds as natural feed additives in poultry and swine diets: a review. J Anim Sci Biotechnol. 2021, 12, 48. [Google Scholar] [CrossRef]
- Di Domenico, M.; Feola, A.; Ambrosio, P.; Pinto, F.; Galasso, G.; Zarrelli, A.; Di Fabio, G.; Porcelli, M.; Scacco, S.; Inghingolo, F.; Quagliuolo, L.; Ballini, A.; Boccellino, M. Antioxidant Effect of Beer Polyphenols and Their Bioavailability in Dental-Derived Stem Cells (D-dSCs) and Human Intestinal Epithelial Lines (Caco-2) Cells. Stem Cells Int. 2020, 1, 8835813. [Google Scholar] [CrossRef] [PubMed]
- Vastolo, A.; Calabró, S.; Liotta, L.; Musco, N.; Di Rosa, A.R.; Cutrignelli, M.I.; Chiofalo, B. In Vitro Fermentation and Chemical Characteristics of Mediterranean By-Products for SwineNutrition. Animals 2019, 9, 556. [Google Scholar] [CrossRef]
- Di Meo, M.C; De Cristofaro, G.A.; Imperatore, R.; Rocco, M.; Giaquinto, D.; Palladino, A.; Zotti, T.; Vito, P.; Paolucci, M.; Varricchio, E. Microwave-Assisted Extraction of Olive Leaf from Five Italian Cultivars: Effects of Harvest-Time and Extraction Conditions on Phenolic Compounds and In Vitro Antioxidant Properties. ACS Food Sci. Technol. 2021, 2, 31–40. [Google Scholar] [CrossRef]
- Saija, A.; Trombetta, D.; Tomaino, A.; Lo Cascio, R.; Princi, P.; Uccella, N; Bonina, F.; Castelli, F. `In vitro’ evaluation of the antioxidant activity and biomembrane interaction of the plant phenols oleuropein and hydroxytyrosol. Int J Pharm. [CrossRef]
- Paiva-Martins, F.; Barbosa, S.; Pinheiro, V.; Mourão, J. L.; Outor-Monteiro, D. The effect of olive leaves supplementation on the feed digestibility, growth performances of pigs and quality of pork meat. Meat Sci. 2009, 82(4), 438–443. [Google Scholar] [CrossRef] [PubMed]
- Qwele, K.; Hugo, A.; Oyedemi, S. O.; Moyo, B.; Masika, P. J.; Muchenje, V. Chemical composition, fatty acid content and antioxidant potential of meat from goats supplemented with Moringa (Moringa oleifera) leaves, sunflower cake and grass hay. Meat Sci. 2013, 93(3), 455–4. [Google Scholar] [CrossRef] [PubMed]
- Poutaraud, A.; Michelot-Antalik, A.; Plantureux, S. Grasslands: a source of secondary metabolites for livestock health. J. Agric. Food Chem 2017, 65(31), 6535–6553. [Google Scholar] [CrossRef] [PubMed]
- Tava, A.; Biazzi, E.; Ronga, D.; Mella, M.; Doria, F.; D’Addabbo, T.; Candido, V.; Avato, P. Chemical identification of specialized metabolites from sulla (Hedysarumcoronarium L.) collected in southern Italy. Molecules 2021, 26, 4606. [Google Scholar] [CrossRef] [PubMed]
- Rivero, M. J.; Rodríguez-Estévez, V.; Pietrosemoli, S.; Carballo, C.; Cooke, A. S.; Kongsted, A. G. Forage consumption and its effects on the performance of growing swine—Discussed in relation to European wild boar (Sus Scrofa L.) in semi-extensive systems: A review. Animals 2019, 9, 457. [Google Scholar] [CrossRef]
- Priolo, A.; Bella, M.; Lanza, M.; Galofaro, V.; Biondi, L.; Barbagallo, D.; Ben Salem, H.; Pennisi, P. Carcass and meat quality of lambs fed fresh sulla (Hedysarum coronarium L.) with or without polyethylene glycol or concentrate. Small Rumin. Res. 2005, 59, 281–288. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, P.; Reyes-Palomo, C.; Sanz-Fernández, S.; Rufino-Moya, P.J.; Zafra, R.; Martínez-Moreno, F.J.; Rodríguez-Estévez, V.; Díaz-Gaona, C. Antiparasitic Tannin-Rich Plants from the South of Europe for Grazing Livestock: A Review. Animals 2023, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Ponte, M.; Maniaci, G.; Di Grigoli, A.; Gannuscio, R.; Ashkezary, M.R.; Addis, M.; Pipi, M.; Alabiso, M.; Todaro, M.; Bonanno, A. Feeding Dairy Ewes with Fresh or Dehydrated Sulla (Sulla coronarium L.) Forage. 2. Effects on Cheese Enrichment in Bioactive Molecules. Animals 2022, 12, 2462. [Google Scholar] [CrossRef]
- Gladine, C.; Rock, E.; Morand, C.; Cauchart, D.; Durand, D. Bioavailability and antioxidant capacity of plant extracts rich in polyphenols, given as a single acute dose, in sheep made highly susceptible to lipoperoxidation. Br. J. Nutr. 2007, 98, 691–701. [Google Scholar] [CrossRef]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Botsoglou, E.; Govaris, A.; Ambrosiadis, I.; Fletouris, D. Lipid and protein oxidation of α-linolenic acid-enriched pork during refrigerated storage as influenced by diet supplementation with olive leaves (Olea europea L.) or α-tocopheryl acetate. Meat Sci. 2012, 92, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Difonzo, G.; Troilo, M.; Squeo, G.; Pasqualone, A.; Caponio, F. Functional compounds from olive pomace to obtain high-added value foods–a review. J. Sci. Food Agr. 2021, 101(1), 15–26. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, M.C.; Salzano, A.; Zotti, T.; Palladino, A.; Giaquinto, D.; Maruccio, L.; Romanucci, R.; Rocco, M.; Zarrelli, A.; D’Occhio, M.J. , Campanile, G.; Varricchio, E. Plasma fatty acid profile in Italian Holstein-Friesian dairy cows supplemented with natural polyphenols from the olive plant Olea Europaea L. Vet. Anim. Sci. 2023, 21, 100298. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, M. C.; Giacco, A.; Zarrelli, A.; Mandrone, V. M.; D’Angelo, L.; Silvestri, E.; De Girolamo, P.; Varricchio, E. Effects of Olea europaea L. Polyphenols on the Animal Welfare and Milk Quality in Dairy Cows. Animals 2023, 13(20), 3225. [Google Scholar] [CrossRef] [PubMed]
- Paiva-Martins, F.; Barbosa, S.; Silva, M.; Monteiro, D.; Pinheiro, V.; Mourão, J.L.; Fernandes, J.; Rocha, S.; Belo, L.; Santos-Silva, A. The effect of olive leaf supplementation on the constituents of blood and oxidative stability of red blood cells. J. Funct. Foods 2014, 9, 271–279. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Ribeirinha, T.; Silva, A.; Gonçalves, R.; Pinheiro, V.; Mourão, J. L.; Outor-Monteiro, D. Effects of the dietary incorporation of olive leaves on growth performance, digestibility, blood parameters and meat quality of growing pigs. J. Sci. Food Agric. 2014, 94(14), 3023–3029. [Google Scholar] [CrossRef]
- Ponnampalam, E. N.; Sinclair, A. J.; Holman, B. W. The sources, synthesis and biological actions of omega-3 and omega-6 fatty acids in red meat: An overview. Foods 2021, 10(6), 1358. [Google Scholar] [CrossRef] [PubMed]
- Dugan, M. E.; Vahmani, P.; Turner, T. D.; Mapiye, C.; Juárez, M.; Prieto, N.; Beaulieu, A. D.; Zijlstra, R. T.; Patience, J. F.; Aalhus, J. L. Pork as a source of omega-3 (n-3) fatty acids. J. Clin. Med. 2015, 4(12), 1999–2011. [Google Scholar] [CrossRef]
- AOAC—Association of Official Analytical Chemists. Official Methods of Analysis, 16th ed.; AOAC: Washington, DC, USA, 1995. [Google Scholar]
- DellaGreca, M.; Previtera, L.; Purcaro, R.; Zarrelli, A. Cinnamic ester derivatives from Oxalis pes-caprae (Bermuda buttercup). J. Nat. Prod. 2007, 70, 1664–1667. [Google Scholar] [CrossRef]
- Crescenzo, R.; Bianco, F.; Mazzoli, A.; Giacco, A.; Cancelliere, R.; Di Fabio, G.; Zarrelli, A.; Liverini, G.; Iossa, S. Fat quality influences the obesogenic effect of high-fat diets. Nutrients 2015, 7, 9475–9491. [Google Scholar] [CrossRef] [PubMed]
- Pollio, A.; Zarrelli, A.; Romanucci, V.; Di Mauro, A.; Barra, F.; Pinto, G.; Crescenzi, E.; Roscetto, E.; Palumbo, G. Polyphenolic profile and targeted bioactivity of methanolic extracts from Mediterranean ethnomedicinal plants on human cancer cell lines. Molecules 2016, 21, 395. [Google Scholar] [CrossRef] [PubMed]
- Paiva-Martins, F.; Gordon, M. H. Isolation and characterization of the antioxidant component 3,4-dihydroxyphenylethyl 4-formyl-3-formylmethyl-4-hexanoate from olive (Olea europaea). J. Agric. Food Chem. 2001, 49, 4214–4219. [Google Scholar] [CrossRef] [PubMed]
- Savournin, C.; Baghdikian, B.; Elias, R.; Dargouth-Kesraoui, F.; Boukef, K.; Balansard, G. Rapid high-performance liquid chromatography analysis for the quantitative determination of oleuropein in Olea europaea leaves. J Agric Food Chem. 2001, 49, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Benavente-Garcıa, O.; Castillo, J.; Lorente, J.; Ortuño, A. D. R. J.; Del Rio, J. A. Antioxidant activity of phenolics extracted from Olea europaea L. leaves. Food Chem. 2000, 68(4), 457–462. [Google Scholar] [CrossRef]
- Pereira, A.P.; Ferreira, I.C.; Marcelino, F.; Valentão, P.; Andrade, P.B.; Seabra, R.; Estevinho, L.; Bento, A.; Pereira, J.A. Phenolic Compounds and Antimicrobial Activity of Olive (Olea europaea L. Cv. Cobrançosa) Leaves. Molecules 2007, 12, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Pino, F.; Millan-Linares, MC.; Villanueva-Lazo, A.; Fernandez-Prior, Á.; Montserrat-de-la-Paz, S. In vivo evidences of the health-promoting properties of bioactive compounds obtained from olive by-products and their use as food ingredient. Crit Rev Food Sci Nutr. 2023, 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Molinu, M.G.; Sulas, L.; Campesi, G.; Re, G.A.; Sanna, F.; Piluzza, G. Subterranean Clover and Sulla as Valuable and Complementary Sources of Bioactive Compounds for Rainfed Mediterranean Farming Systems. Plants 2023, 12, 417. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Deng, C.; Chen, Q.; Zhao, S.; Li, P.; Wu, T.; Hou, Y.; Yi, D. Protective effects and mechanisms of ellagic acid on intestinal injury in piglets infected with porcine epidemic diarrhea virus. Front. Immunol. 2024, 15, 1323866. [Google Scholar] [CrossRef]
- Wang, F.; Chen, J.; Yin, Y.; Yang, M.; Xiao, Y.; Cheng, Y.; Yin, L.; Fu, C. The effects of dietary ellagic acid supplementation on growth performance, immune response, antioxidant activity, digestive enzyme activities, and intestinal functions in yellow-feathered broilers. J. Anim. Sci. 2022, 100(12), skac301. [Google Scholar] [CrossRef]
- Wen, X.; Zhou, M.; Lu, Q.; Liu, B.; Shi, X.; Zhao, J. Addition of ellagic acid improved the immune ability and delayed the apoptosis of ovarian granulosa cells of Guizhou black goat. Anim. Prod. Sci. 2023, 64(1), AN23310. [Google Scholar] [CrossRef]
- Echegaray, N.; Munekata, P.E.S.; Centeno, J.A.; Domínguez, R.; Pateiro, M.; Carballo, J.; Lorenzo, J.M. Total Phenol Content and Antioxidant Activity of Different Celta Pig Carcass Locations as Affected by the Finishing Diet (Chestnuts or Commercial Feed). Antioxidants 2021, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Serra, V.; Salvatori, G.; Pastorelli, G. Dietary Polyphenol Supplementation in Food Producing Animals: Effects on the Quality of Derived Products. Animals 2021, 11, 401. [Google Scholar] [CrossRef] [PubMed]
- DeJong, S. & Lanari, M. C. Extracts of olive polyphenols improve lipid stability in cooked beef and pork: Contribution of individual phenolics to the antioxidant activity of the extract. Food Chem. 2009, 116, 892–897. [Google Scholar] [CrossRef]
- Botsoglou, E.; Govaris, A.; Christaki, E.; Botsoglou, N. Effect of dietary olive leaves and/or α-tocopheryl acetate supplementation on microbial growth and lipid oxidation of turkey breast fillets during refrigerated storage. Food Chem. 2010, 121(1), 17–22. [Google Scholar] [CrossRef]
- Botsoglou, E.; Govaris, A.; Ambrosiadis, I.; Fletouris, D.; Papageorgiou, G. Effect of olive leaf (Olea europea L.) extracts on protein and lipid oxidation in cooked pork meat patties enriched with n-3 fatty acids. J Sci Food Agric. 2014, 94, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Mylonaki, S.; Kiassos, E.; Makris, D. P.; Kefalas, P. Optimisation of the extraction of olive (Olea europaea) leaf phenolics using water/ethanol-based solvent systems and response surface methodology. Anal Bioanal Chem. 2008, 392, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Leskovec, J.; Rezar, V.; Nemec Svete, A.; Salobir, J.; Levart, A. Antioxidative Effects of Olive Polyphenols Compared to Vitamin E in Piglets Fed a Diet Rich in N-3 PUFA. Animals 2019, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Wood, J. D.; Enser, M.; Fisher, A. V.; Nute, G. R.; Sheard, P. R.; Richardson, R. I. ; Hughes,S.I.; Whittington, F. M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Joven, M.; Pintos, E.; Latorre, M. A.; Suárez-Belloch, J.; Guada, J. A.; Fondevila, M. Effect of replacing barley by increasing levels of olive cake in the diet of finishing pigs: Growth performances, digestibility, carcass, meat and fat quality. Anim Feed Sci Technol. 2014, 197, 185–193. [Google Scholar] [CrossRef]
- Tsala, A.; Mpekelis, V.; Karvelis, G.; Tsikakis, P.; Goliomytis, M.; Simitzis, P. Effects of Dried Olive Pulp Dietary Supplementation on Quality Characteristics and Antioxidant Capacity of Pig Meat. Foods 2020, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- Dal Bosco, A.; Mugnai, C.; Ruggeri, S.; Mattioli, S.; Castellini, C. Fatty acid composition of meat and estimated indices of lipid metabolism in different poultry genotypes reared under organic system. Poultry Sci. 2012, 91(8), 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Losacco, C.; Laudadio, V.; Schiavitto, M.; Tufarelli, V. Perspectives and advantages of using olive (Olea europaea) by-products as a dietary supplement for rabbit production and health. S. Afr. J. Anim. Sci. 2023, 53(5), 737–754. [Google Scholar] [CrossRef]
- Ferrer, P.; Calvet, S.; García-Rebollar, P.; de Blas, C.; Jiménez-Belenguer, A. I.; Hernández, P.; Piquer, O.; Cerisuelo, A. Partially defatted olive cake in finishing pig diets: Implications on performance, faecal microbiota, carcass quality, slurry composition and gas emission. Animal 2020, 14(2), 426–434. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-García, A.; Vieira-Aller, C. Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review. Animals 2023, 13, 1696. [Google Scholar] [CrossRef]
- Pugliese, C.; Sirtori, F. Quality of Meat and Meat Products Produced from Southern European Pig Breeds. Meat Sci. 2012, 90, 511–518. [Google Scholar] [CrossRef]

Table 1.
Plants under study.
| Family name | Cultivar name | Scientific name | English name |
|---|---|---|---|
| Oleaceae | Ortice | Olea europaea | Olive tree |
| Brassicaceae | - | Hedysarium coronarium | Wild Radish |
Table 2.
Ingredients of standard diet and pasture grasses (% dry matter (DM)).
| Components | % DM |
|---|---|
| Standard Diet | |
| Maize | 45.00 |
| Barley | 27.00 |
| Wheat | 15.00 |
| Soy Protein 46% | 8.00 |
| Bran | 5.00 |
| Pasture grasses | |
| Sulla a | 18.24 |
a 500 g/head/day of pasture grasses Sulla (H. coronarium L.).
Table 3.
Chemical composition of standard diet and pasture grasses on fresh matter (F.M.) and dry matter (D.M.).
Table 3.
Chemical composition of standard diet and pasture grasses on fresh matter (F.M.) and dry matter (D.M.).
| a. Standard Diet | ||
| Parameters | DM [%] | |
| Moisture | 11.15 | |
| Dry matter | 88.85 | |
| Crude protein | 13.00 | |
| Ash | 1.94 | |
| Crude Fats | 2.78 | |
| Raw fibre | 4.05 | |
| Starch | 52.17 | |
| b. Sulla (H. coronarium L.) | ||
| Parameters | F.M. [%] | D.M. [%] |
| Moisture | 81.76 | |
| Dry matter | 18.24 | |
| Crude protein | 4.56 | 25.00 |
| Ash | 2.07 | 11.36 |
| NDF 1 | 6.57 | 36.00 |
| ADF 2 | 3.79 | 20.80 |
1 NDF: neutral detergent fibre; 2 ADF: acid detergent fibre.
Table 4.
Pig performances and chemical composition of muscle (g/100 g wet weight).
| Dietary treatment1 | SEM6 | p-value | ||
|---|---|---|---|---|
| Control | OL | |||
| Final BW2, kg | 204 | 200 | 2.016 | 0.261 |
| Carcass weight, kg | 200 | 185 | 5.560 | 0.563 |
| Total DMI3, g/d | 3.54 | 3.26 | 0.156 | 0.674 |
| ADG4, g/d | 750 | 650 | 9.261 | 0.633 |
| FCR 5, g DMI/g ADG | 4.72 | 5.02 | 0.192 | 0.105 |
| Chemical composition | ||||
| Moisture | 71.65 | 72.20 | 0.245 | 0.321 |
| Crude protein | 23.78 | 23.54 | 0.112 | 0.055 |
| Ether extract | 3.06 | 3.01 | 0.091 | 0.186 |
| Ash | 1.51 | 1.50 | 0.067 | 0.156 |
1 Treatments were standard diet (control) and standard diet + O. europaea L. leaves extract (OL); 2 BW = body weight; 3 DMI = dry matter intake; 4 ADG = average daily gain; 5 FCR = feed conversion ratio; 6 SEM = standard error of means.
Table 5.
Total phenolic content and antioxidant activity (% DPPH inhibition) in standard and enriched diet, Ortice olive leaf, and Sulla extracts. 1.
Table 5.
Total phenolic content and antioxidant activity (% DPPH inhibition) in standard and enriched diet, Ortice olive leaf, and Sulla extracts. 1.
| Phenolic Extracts | TPC (mg GAE/g) | TFC (mg QE/g) | Inhibition % |
|---|---|---|---|
| Standard Diet | 8.14 ± 0.30 | 1.71 ± 0.20 | 28.21 ± 1.20 |
| Olive leaves extract | 19.13 ± 0.75 | 3.50 ± 0.43 | 51.10 ± 2.98 |
| Enriched Diet | 33.59 ± 0.88 | 2.98 ± 0.32 | 70.20 ± 2.36 |
| Sulla (H. coronarium) | 17.47 ± 0.52 | 3.02 ± 0.26 | 63.20 ± 2.00 |
1 Data are expressed as mean ± SD (n=3). Two-way ANOVA analysis between TPC, TFC, and inhibition % in phenolic extracts. TPC and TFC vs Inhibition% in the standard diet, olive leaves extract, enriched diet, and Sulla ** p < 0.01.
Table 6.
Phenolic acids contents (mg/g of dry extract) of Ortice olive leaves (A) and Sulla plant (B) with retention time and r2. Values are mean ± SD (n = 3).
Table 6.
Phenolic acids contents (mg/g of dry extract) of Ortice olive leaves (A) and Sulla plant (B) with retention time and r2. Values are mean ± SD (n = 3).
| A.Ortice olive leaves | ||||||
| No. Peak | Compound | λ (nm) | RT (min) | r2 | Phenolic compounds concentration (mg/g) | |
| 14 | Verbascoside | 280 | 14 | 0.98 | 0.098 ± 0.05 | |
| 15 | Luteolin-7-glucoside | 280 | 15 | 0.99 | 0.098 ± 0.04 | |
| 16 | Oleuropein diglucoside | 280 | 16 | 0.99 | 0.100 ± 0.09 | |
| B. Sulla plant | ||||||
| No. Peak | Compound | λ (nm) | RT (min) | r2 | Phenolic compounds concentration (mg/g) | |
| 6 | Caffeic acid | 280 | 14 | 0.99 | 0.068 ± 0.04 | |
| 8 | Ellagic acid | 280 | 14.7 | 0.99 | 0.096 ± 0.01 | |
Table 7.
Fatty acids profile of the standard diet, Ortice olive leaf extract, enriched diet and Sulla extract.1.
Table 7.
Fatty acids profile of the standard diet, Ortice olive leaf extract, enriched diet and Sulla extract.1.
| Fatty acid | Items | Standard diet (g/100 g) |
Ortice olive leaf extract (g/100 g) |
Enriched diet (g/100g) |
Sulla extract |
SEM | p-value |
|---|---|---|---|---|---|---|---|
| Myristic | C14:0 | 0.88 | 1.14 | 0.70 | 0.51 | 0.13 | ns |
| Palmitic | C16:0 | 39.44 | 38.00 | 37.01 | 26.11 | 2.64 | <0.001 |
| Trans-Palmitoleic | C16:1n7t | 0.09 | 0.06 | 0.05 | 0.00 | 0.02 | ns |
| Palmitoleic | C16:1n7 | 0.15 | 0.25 | 0.18 | 0.70 | 0.13 | ns |
| Stearic | C18:0 | 9.73 | 10.55 | 11.01 | 13.37 | 0.78 | <0.001 |
| Oleic | C18:1n9 | 16.35 | 36.18 | 19.30 | 2.56 | 6.90 | <0.001 |
| Trans-Linoleic | C18:2n6t | 0.30 | 0.13 | 0.18 | 0.00 | 0.06 | ns |
| Linoleic | C18:2n6 | 30.96 | 11.28 | 29.11 | 9.27 | 5.73 | <0.001 |
| Gamma-Linolenic | C18:3n6 | 0.16 | 0.02 | 0.12 | 0.00 | 0.04 | ns |
| Eicosenoic | C20:1n9 | 0.10 | 0.18 | 0.08 | 0.23 | 0.03 | ns |
| Alpha-Linolenic | C18:3n3 | 0.08 | 0.06 | 1.01 | 46.59 | 11.55 | <0.001 |
| Eicosadienoic | C20:2n6 | 0.19 | 0.24 | 0.08 | 0.00 | 0.05 | ns |
| Dihomo-γ-linolenic | C20:3n6 | 0.12 | 0.05 | 0.10 | 0.00 | 0.03 | ns |
| Arachidonic | C20:4n6 | 0.77 | 1.01 | 0.68 | 0.38 | 0.13 | ns |
| Lignoceric | C24:0 | 0.15 | 0.36 | 0.14 | 0.05 | 0.07 | ns |
| Eicosapentaenoic (EPA) | C20:5n3 | 0.23 | 0.09 | 0.05 | 0.11 | 0.04 | ns |
| Nervonic | C24:1n9 | 0.06 | 0.04 | 0.06 | 0.00 | 0.01 | ns |
| Docosahexaenoic (DHA) | C22:6n3 | 0.01 | 0.04 | 0.01 | 0.12 | 0.03 | ns |
| Total | |||||||
| ∑SFA | 50.20 | 50.05 | 48.86 | 40.04 | 2.11 | <0.001 | |
| ∑MUFA | 16.84 | 36.80 | 19.67 | 3.49 | 5.93 | <0.001 | |
| ∑PUFA | 32.96 | 13.15 | 31.47 | 56.47 | 7.69 | <0.001 | |
| ∑n-6 PUFA | 32.50 | 12.73 | 30.27 | 9.65 | 5.09 | <0.001 | |
| ∑n-3 PUFA | 0.32 | 0.19 | 1.07 | 46.82 | 10.89 | <0.001 | |
| n-6/n-3 PUFA |
101.56 | 67.00 | 28.29 | 0.21 | 19.20 | <0.001 | |
| ∑SFA/ ∑MUFA |
2.98 | 1.36 | 2.48 | 11.47 | 2.01 | <0.001 | |
| ∑SFA/ ∑PUFA |
1.52 | 3.81 | 1.55 | 0.71 | 0.58 | <0.001 |
1 Abbreviation: MUFA, monounsaturated fatty acid; SFA, saturated fatty acid; PUFA, polyunsaturated fatty acid; n-6, omega6; n-3, omega3; SEM, standard error of the mean; ns, not significant. Note: Two-way ANOVA in the standard diet, Ortice olive leaf extract, enriched diet, and Sulla.
Table 8.
Fatty acids profile of Longissimus dorsi, Diaphragm, Semimembranosus, and Psoas major muscles.1
Table 8.
Fatty acids profile of Longissimus dorsi, Diaphragm, Semimembranosus, and Psoas major muscles.1
| Musculus Longissimus dorsi | |||||
|---|---|---|---|---|---|
| Fatty acid | Items | Control diet | OL diet | SEM | p-value |
| Myristic | C14:0 | 2.17 | 1.67 | 0.18 | ns |
| Palmitic | C16:0 | 29.16 | 28.98 | 0.06 | ns |
| Trans-Palmitoleic | C16:1n7t | 0.02 | 0.02 | 0.00 | ns |
| Palmitoleic | C16:1n7 | 0.15 | 0.12 | 0.01 | ns |
| Stearic | C18:0 | 7.82 | 5.35 | 0.87 | <0.001 |
| Trans-Oleic | C18:1n9t | 0.01 | 0.05 | 0.01 | ns |
| Oleic | C18:1n9 | 51.16 | 55.63 | 1.59 | <0.001 |
| Trans-Linoleic | C18:2n6t | 0.03 | 0.03 | 0.00 | ns |
| Linoleic | C18:2n6 | 8.12 | 6.26 | 0.66 | <0.05 |
| Gamma-Linolenic | C18:3n6 | 0.27 | 0.30 | 0.01 | ns |
| Eicosenoic | C20:1n9 | 0.27 | 0.21 | 0.02 | ns |
| Alpha-Linolenic | C18:3n3 | 0.03 | 0.04 | 0.00 | ns |
| Eicosadienoic | C20:2n6 | 0.18 | 0.21 | 0.01 | ns |
| Dihomo-γ-linolenic | C20:3n6 | 0.06 | 0.05 | 0.00 | ns |
| Arachidonic | C20:4n6 | 0.46 | 0.40 | 0.02 | ns |
| Lignoceric | C24:0 | 0.00 | 0.49 | 0.17 | ns |
| Eicosapentaenoic (EPA) | C20:5n3 | 0.01 | 0.01 | 0.00 | ns |
| Nervonic | C24:1n9 | 0.01 | 0.01 | 0.00 | ns |
| Docosatetraenoic | C22:4n6 | 0.07 | 0.06 | 0.00 | ns |
| Docosatetraenoic-N6 | C22:5n6 | 0.01 | 0.01 | 0.00 | ns |
| Docosatetraenoic-N3 | C22:5n3 | 0.04 | 0.04 | 0.00 | ns |
| Docosahexaenoic (DHA) | C22:6n3 | 0.01 | 0.11 | 0.04 | ns |
| Total | |||||
| SFA | 39.14 | 33.97 | 1.84 | <0.001 | |
| MUFA | 51.61 | 60.29 | 3.08 | <0.001 | |
| PUFA | 9.25 | 5.75 | 1.24 | <0.05 | |
| Omega6 | 8.99 | 5.19 | 1.35 | <0.01 | |
| Omega3 | 0.08 | 0.33 | 0.09 | ns | |
| n6/n3 | 119.80 | 15.95 | 36.82 | <0.001 | |
| Musculus Diaphragm | |||||
| Fatty acid | Items | Control diet | OL diet | SEM | p-value |
| Myristic | C14:0 | 1.14 | 1.33 | 0.07 | ns |
| Palmitic | C16:0 | 25.35 | 24.61 | 0.26 | ns |
| Trans-Palmitoleic | C16:1n7t | 0.01 | 0.02 | 0.00 | ns |
| Palmitoleic | C16:1n7 | 1.83 | 1.99 | 0.06 | ns |
| Stearic | C18:0 | 14.47 | 13.30 | 0.41 | <0.05 |
| Trans-Oleic | C18:1n9t | 0.90 | 1.06 | 0.06 | ns |
| Oleic | C18:1n9 | 35.02 | 37.07 | 0.73 | <0.001 |
| Trans-Linoleic | C18:2n6t | 0.02 | 0.04 | 0.01 | ns |
| Linoleic | C18:2n6 | 14.30 | 13.33 | 0.34 | ns |
| Gamma-Linolenic | C18:3n6 | 0.01 | 0.01 | 0.00 | ns |
| Eicosenoic | C20:1n9 | 0.68 | 0.98 | 0.11 | ns |
| Alpha-Linolenic | C18:3n3 | 5.31 | 5.32 | 0.00 | ns |
| Eicosadienoic | C20:2n6 | 0.09 | 0.06 | 0.01 | ns |
| Dihomo-γ-linolenic | C20:3n6 | 0.08 | 0.09 | 0.00 | ns |
| Arachidonic | C20:4n6 | 0.32 | 0.27 | 0.02 | ns |
| Lignoceric | C24:0 | 0.32 | 0.38 | 0.02 | ns |
| Eicosapentaenoic (EPA) | C20:5n3 | 0.03 | 0.04 | 0.01 | ns |
| Nervonic | C24:1n9 | 0.03 | 0.02 | 0.00 | ns |
| Docosatetraenoic | C22:4n6 | 0.01 | 0.01 | 0.00 | ns |
| Docosatetraenoic-N6 | C22:5n6 | 0.02 | 0.02 | 0.00 | ns |
| Docosatetraenoic-N3 | C22:5n3 | 0.01 | 0.01 | 0.00 | ns |
| Docosahexaenoic (DHA) | C22:6n3 | 0.10 | 0.10 | 0.00 | ns |
| Total | |||||
| SFA | 38.20 | 39.71 | 0.54 | ns | |
| MUFA | 44.17 | 41.16 | 1.07 | <0.05 | |
| PUFA | 20.28 | 17.98 | 0.81 | <0.05 | |
| Omega6 | 14.73 | 12.95 | 0.63 | ns | |
| Omega3 | 5.45 | 5.00 | 0.16 | ns | |
| n6/n3 | 2.70 | 2.59 | 0.04 | ns | |
| Musculus Semimembranosus | |||||
| Fatty acid | Items | Control diet | OL diet | SEM | p-value |
| Myristic | C14:0 | 2.10 | 1.67 | 0.15 | ns |
| Palmitic | C16:0 | 23.41 | 22.23 | 0.42 | <0.001 |
| Trans-palmitoleic | C16:1n7t | 0.22 | 0.20 | 0.01 | ns |
| Palmitoleic | C16:1n7 | 2.37 | 2.88 | 0.18 | ns |
| Stearic | C18:0 | 12.14 | 11.18 | 0.34 | <0.05 |
| Trans-Oleic | C18:1n9t | 0.02 | 0.32 | 0.11 | ns |
| Oleic | C18:1n9 | 37.05 | 39.19 | 0.76 | <0.001 |
| Trans-Linoleic | C18:2n6t | 0.22 | 0.28 | 0.02 | ns |
| Linoleic | C18:2n6 | 13.77 | 11.00 | 0.98 | <0.05 |
| Gamma-Linolenic | C18:3n6 | 0.00 | 0.06 | 0.02 | ns |
| Eicosenoic | C20:1n9 | 0.00 | 0.00 | 0.00 | ns |
| Alpha-Linolenic | C18:n3 | 3.04 | 4.24 | 0.42 | <0.05 |
| Eicosadienoic | C20:2n6 | 0.65 | 1.05 | 0.14 | ns |
| Dihomo-γ-linolenic | C20:3n6 | 0.01 | 0.01 | 0.00 | ns |
| Arachidonic | C20:4n6 | 0.51 | 1.00 | 0.17 | ns |
| Lignoceric | C24:0 | 3.46 | 3.57 | 0.04 | ns |
| Eicosapentaenoic (EPA) | C20:5n3 | 0.12 | 0.12 | 0.00 | ns |
| Nervonic | C24:1n9 | 0.10 | 0.14 | 0.02 | ns |
| Docosatetraenoic | C22:4n6 | 0.01 | 0.01 | 0.00 | ns |
| Docosatetraenoic-N6 | C22:5n6 | 0.45 | 0.52 | 0.02 | ns |
| Docosatetraenoic-N3 | C22:5n3 | 0.26 | 0.27 | 0.01 | ns |
| Docosahexaenoic (DHA) | C22:6n3 | 0.13 | 0.10 | 0.01 | ns |
| Total | |||||
| SFA | 41.11 | 36.39 | 1.67 | <0.001 | |
| MUFA | 39.75 | 45.34 | 1.98 | <0.001 | |
| PUFA | 19.15 | 18.28 | 0.31 | ns | |
| Omega6 | 14.52 | 11.80 | 0.96 | <0.01 | |
| Omega3 | 3.54 | 4.92 | 0.49 | ns | |
| n6/n3 | 4.11 | 2.40 | 0.61 | ns | |
| Musculus Psoas Major | |||||
| Fatty acid | Items | Control diet | OL diet | SEM | p-value |
| Myristic | C14:0 | 1.70 | 2.00 | 0.11 | ns |
| Palmitic | C16:0 | 24.57 | 24.54 | 0.01 | ns |
| Trans-Palmitoleic | C16:1n 7t | 0.02 | 0.05 | 0.01 | ns |
| Palmitoleic | C16:1 7 | 3.42 | 2.63 | 0.28 | ns |
| Stearic | C18:0 | 9.94 | 7.97 | 0.70 | <0.001 |
| Trans-Oleic | C18:1n9t | 0.01 | 0.02 | 0.00 | ns |
| Oleic | C18:1n9 | 50.47 | 55.37 | 1.74 | <0.001 |
| Trans-Linoleic | C18:2n6t | 0.01 | 0.05 | 0.01 | ns |
| Linoleic | C18:2n6 | 7.80 | 5.77 | 0.72 | <0.05 |
| Gamma-Linolenic | C18:3n6 | 0.08 | 0.04 | 0.01 | ns |
| Eicosenoic | C20:1n9 | 0.01 | 0.05 | 0.01 | ns |
| Alpha-Linolenic | C18:3n3 | 0.09 | 0.07 | 0.01 | ns |
| Eicosadienoic | C20:2n6 | 0.82 | 0.26 | 0.20 | ns |
| Dihomo-γ-linolenic | C20:3n6 | 0.13 | 0.10 | 0.01 | ns |
| Arachidonic | C20:4n6 | 0.52 | 0.42 | 0.03 | ns |
| Lignoceric | C24:0 | 0.02 | 0.42 | 0.14 | ns |
| Eicosapentaenoic (EPA) | C20:5n3 | 0.08 | 0.05 | 0.01 | ns |
| Nervonic | C24:1n9 | 0.11 | 0.06 | 0.02 | ns |
| Docosatetraenoic | C22:4n6 | 0.12 | 0.05 | 0.02 | ns |
| Docosatetraenoic-N6 | C22:5n6 | 0.01 | 0.00 | 0.00 | ns |
| Docosatetraenoic-N3 | C22:5n3 | 0.00 | 0.01 | 0.00 | ns |
| Docosahexaenoic (DHA) | C22:6n3 | 0.01 | 0.01 | 0.00 | ns |
| Total | |||||
| SFA | 36.24 | 34.15 | 0.74 | ns | |
| MUFA | 54.06 | 61.25 | 2.55 | <0.001 | |
| PUFA | 9.70 | 4.61 | 2.25 | <0.001 | |
| Omega 6 | 8.67 | 3.81 | 1.73 | <0.001 | |
| Omega 3 | 0.19 | 0.60 | 0.15 | ns | |
| n6/n3 | 45.66 | 6.34 | 11.88 | <0.001 | |
1 Abbreviation: MUFA, monounsaturated fatty acid; SFA, saturated fatty acid; PUFA, polyunsaturated fatty acid; n-6, omega6; n-3, omega3; SEM, standard error of the mean; ns, not significant. Note: Two-way ANOVA in the Longissimus dorsi, Diaphragm, Semimembranosus, and Psoas major muscles in control and OL diet.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



