
Submitted:
31 July 2024
Posted:
01 August 2024
You are already at the latest version
Abstract
Metals play vital roles in biological systems, with iron/heme being essential for cellular and metabolic functions necessary for survival and/or virulence in many bacterial pathogens. Given the rise of bacterial resistance to current antibiotics, there is an urgent need for the development of non-toxic and novel antibiotics that do not contribute to resistance to other antibiotics. Gallium, which mimics iron, has emerged as a promising antimicrobial agent, offering a novel approach to combat bacterial infections. Gallium does not have any known functions in biological systems. Gallium exerts its effects primarily by replacing iron in redox enzymes, effectively inhibiting bacterial growth by targeting multiple iron/heme-dependent biological processes and suppressing the development of drug resistance. The aim of this review is to highlight recent findings on the mechanisms of action of gallium and provide further insights into the development of gallium-based compounds. Understanding the mechanisms underlying gallium’s biological activities will enhance its therapeutic application across various medical fields, offering promising avenues for the treatment of infectious diseases, cancer, and inflammatory conditions.
Keywords:
Gallium
; Antimicrobial activity
; Reactive oxygen species
; Iron/heme uptake
Introduction
The discovery of antibiotics had a huge impact on humanity, saving countless lives from bacterial infections. However, overuse and misuse have led to the development of bacterial resistance to drugs, resulting in increased fatalities. The rise of drug-resistant bacteria has posed a significant challenge in treating bacterial infections, highlighting the need for the development of new antibiotics that do not promote bacterial resistance.
The use of metals in medicine dates back many centuries. Although essential metals can be toxic to cells in high concentrations, non-essential metals like silver (Ag), mercury (Hg), gold (Au), titanium (Ti), and tellurium (Te) are highly toxic to most bacteria and possess potent microbicidal properties even at very low concentrations.[1,2,3] Over the past two hundred years, medical practitioners have employed oxides of Te, Mg, and As, as well as salts of Cu and Hg, to treat various diseases such as leprosy, tuberculosis, gonorrhea, and syphilis.[2] The practice of using metals in medicine was common until Nobel laureate Sir Alexander Fleming discovered antibiotics in the 1920s, which led to a rapid decline in these applications. However, with the increasing prevalence of bacterial resistance to existing antibiotics, there is a critical need to develop novel, non-toxic antibiotics that do not contribute to resistance against other antibiotics. Consequently, metallodrugs have gained attention for combating drug-resistant bacteria. Among these, gallium [Ga(III)], a semi-metallic element, is utilized in various applications and devices, including electronics, semiconductors, and medical imaging. Ga(III) has shown therapeutic effects in the treatment of various disorders, including cancer, hemostasis, bone-related diseases, and autoimmune diseases.[4] Gallium nitrate (Ga(NO3)3) in citrate buffer was approved by the Food and Drug Administration (FDA) for the treatment of hypercalcemia secondary to malignancy in 2003.
Remarkably, Ga(III) also exhibits broad-spectrum antimicrobial activity against many pathogens. Gallium shares several chemical and physical properties similar to iron, such as ionization potential, atomic radius, and electron configuration. In octahedral coordination geometry, the ionic radius of Ga3+ is 0.62Å, while that of Fe3+ (high spin) is 0.645Å.[5] The tetrahedral ionic radius of Ga3+ and Fe3+ are 0.47 Å and 0.49 Å, respectively.[6] These properties enable Ga(III) to mimic Fe(III), allowing it to bind to the binding sites on proteins where Fe(III) binds. However, unlike iron, Ga(III) does not undergo redox recycling, whereas Fe(III) is readily reduced to Fe(II) under physiological conditions. Ga(III) has shown antimicrobial activity against various bacterial pathogens because microorganisms cannot distinguish between the two metals. Enzymes substituted with Ga(III) cannot perform essential functions necessary for bacterial survival, resulting in the disruption of iron-dependent metabolic processes, including iron acquisition/utilization, electron transport, oxidative stress defense, and DNA synthesis.[6]
Other antimicrobial metals, such as Ag and Cu, exhibit antimicrobial activity by reacting with reactive functional groups or by generating reactive oxygen species (ROS) from reactions with water and air. These mechanisms have led to the development of bacterial resistance through an efflux pump, conversion of the toxic metal to a less toxic form by modification, or sequestration by a metal-binding protein or ligands.[2] In contrast, Ga(III) exerts its antimicrobial activity by mimicking Fe(III), an essential element for bacterial survival. This mechanism likely makes it much more difficult for bacteria to develop resistance to gallium, as reducing Ga(III) uptake would lead to the reduction of iron uptake, which is vital for bacterial growth.[7,8]
Due to multi-targeting antimicrobial activity against bacteria, including drug-resistant strains, Ga(III) has garnered significant attention as a novel antibacterial strategy. Consequently, Ga(III)-based compounds, including combination therapy using different forms of Ga(III), have been developed in a variety of ways to increase therapeutic efficacy and improve their low bioavailability. Ga(III) is hydrolyzed into various hydroxide species, such as insoluble Ga(OH)3 and soluble Ga(OH)4, under physiological conditions.[9] The formation of Ga(III) complexes is one way to improve solubility and protect Ga(III) from hydrolysis, thereby increasing its bioavailability. For example, gallium nitrate (GaniteTM), which is no longer available for clinical use in the US for reasons other than safety or effectiveness, needs to be administrated intravenously due to hydrolysis.[10]
The usefulness of an antibiotic is increased if it has good solubility and bioavailability when administered orally. To increase gallium’s oral bioavailability, gallium-based compounds have been prepared by complexing Ga(III) ions with various ligands.[11] Gallium maltolate (GaM), composed of three maltolate ligands bidentately bound to a gallium ion, exhibits some activity as both an anticancer and antimicrobial agent following oral administration.[12] Researchers continue to explore its therapeutic applications, and GaM is still under investigation for the treatment of relapsed and refractory glioblastoma. Another oral gallium complex, tris(8-quinolinolato)gallium(III) (KP46), was synthesized for anticancer therapy and showed high hydrolytic stability.[12,13,14] Although KP46 exerted promising activity against renal cancer and melanoma cells,[15] it has poor bioavailability, possibly due to its low water solubility.[13] Duffin et al. also synthesized alkyl gallium quinolinolate complexes, which exhibited good activity against the motile promastigote form of Leishmania major with micromolar ranges of IC50.[16]
These gallium complexes demonstrate increased stability, solubility, and bioavailability, effectively delivering the antimicrobial agent Ga(III) to target sites and enhancing its therapeutic potential. These complexes also reduce potential side effects associated with free Ga(III) ions. To further improve the efficacy and safety of Ga(III)-based antimicrobial therapies, researchers are investigating various ligands for Ga(III) complexation. They are also developing innovative delivery systems, such as nanomaterials, gallium alloys, and liquid metals for controlled release and targeted delivery of Ga(III).[7,17,18]
This review summarizes the mechanisms of action of Ga(III) to offer insights into enhancing the antibacterial activity of Ga(III)-based compounds. Understanding the mechanisms underlying gallium’s biological activities will enhance its therapeutic therapies for the treatment of infectious diseases.
2. Mechanisms of Action of Ga(III)-Based Compounds
2.1. Iron Acquisition Pathways as a Target for Gallium Antimicrobials
Iron plays multiple roles in biological processes, including DNA synthesis, electron transport, catalysis, oxygen transport, and cellular respiration.[19] Bacteria have evolved numerous strategies to acquire iron from their environment, including animal hosts, to ensure their survival and proliferation.[20] Bacteria possess various iron uptake mechanisms to compete with host cells and systems for iron, addressing the challenges posed by the low bioavailability of iron and the host’s nutritional immunity, the latter referring to biological processes designed to sequester iron from invading microbes. These bacterial iron uptake mechanisms include siderophore/hemophore-mediated uptake, ferrous iron transporters (Feo system), ferric iron transporters, and heme receptors (Figure 1).
Pathogenic bacteria upregulate siderophore biosynthesis and iron-trafficking pathways in iron-deficient environments. Bacterial siderophores, ferric iron (Fe3+) chelators, sequester and deliver Fe3+ into bacteria from the environment.[21,22] These siderophores can compete with and, in some cases, remove Fe3+ from lactoferrin, transferrin, and other iron-binding proteins due to their higher affinity for Fe3+.[23,24,25] Researchers have exploited siderophores to deliver therapeutic agents into pathogens, including Ga(III) ions and antibiotics. A siderophore desferrioxamine (DFO), produced by Streptomyces pilosus, was used to deliver Ga(III) to bacteria. The Ga(III)-DFO complex demonstrated bactericidal activity against P. aeruginosa and inhibited biofilm formation.[26] Similarly, Ga(III) loaded pyochelin, a siderophore from P. aeruginosa, significantly enhanced the inhibitory effect on P. aeruginosa growth compared to gallium nitrate.[27] Pandy. et al. synthesized and demonstrated that a Ga(III)-ciprofloxacin-functionalized desferrichrome complex (gallium-sideromycin conjugate) exhibits broad-spectrum inhibitory activity against Gram-negative and positive bacteria.[28] This complex is internalized via FuhA-mediated transport. Thus, siderophore-mediated drug delivery is a promising strategy for enhancing antibacterial effects by increasing the uptake and activity of the drug.
Like Gram-positive and Gram-negative bacteria, iron acquisition by M. tuberculosis involves both siderophore-mediated iron acquisition and heme-iron acquisition. M. tuberculosis relies on siderophores (exochelins, carboxymycobactin, and mycobactin) to obtain iron from its environment for its growth and virulence.[29,30,31] Thus, targeting siderophore biosynthesis pathways is a rational approach to interrupting siderophore-mediated iron acquisition. Many studies provide detailed research on developing inhibitors for these pathways, particularly the mycobactin biosynthesis pathway.[32,33,34] As mentioned above, another approach for developing antitubercular drugs is to use siderophore as a vehicle to enhance the uptake of a compound with antimicrobial activity. A sideromycin, mycobactin-artemisinin conjugate, shows inhibitory activity against multi-drug resistant and extremely-drug resistant strains of M. tuberculosis and malaria.[35]
Currently, no studies on complexes of Ga(III) with siderophores or Ga(III)-sideromycins from M. tuberculosis as antitubercular agents have been reported. However, Ga-transferrin demonstrated a Fe-reversible, concentration-dependent growth inhibition of M. tuberculosis strains and M. avium complex (MAC) when the organisms grew extracellularly or within human macrophages.[36]
2.2. Heme Acquisition Pathways as a Target for Antimicrobials
In the human host, the most abundant source of iron is heme, a complex of iron with protoporphyrin. Heme is an important prosthetic group of heme proteins, such as hemoglobin, myoglobin, and other heme-containing enzymes responsible for oxygen transport, electron transfer, and redox reactions. Most heme is found in hemeproteins, including hemoglobin in erythrocytes and myoglobin in muscle tissue.[20,37] These hemeproteins and free heme are primary targets for bacterial pathogens to obtain iron.
While siderophores recognize and bind ferric ions in the environment, they cannot remove iron from the heme. Consequently, many bacteria secrete hemophores to capture heme and heme analogs (Figure 1). The heme can be either used directly to meet the metabolic needs of the organism for heme or broken down, thereby releasing iron, which is then available for use. The mechanisms for sequestering free heme or heme within hemeproteins and transporting them are diverse among bacterial species.[38] Several reviews have been reported describing heme-uptake mechanisms in Gram-positive and Gram-negative bacteria, as well as mycobacteria.[21,38,39,40,41] Here, we provide a brief overview of heme uptake by M. tuberculosis. In M. tuberculosis, several genes have been identified that are involved in heme uptake, allowing the mycobacterium to use heme as an alternative iron source. Proteins like Rv0203, mmpL11, and mhuD are implicated in heme utilization. Rv0203 is a secreted heme-binding protein, but it is not able to transfer heme from hemoglobin to inner membrane proteins such as MmpL3 and MmpL11 due to its low heme affinity compared to hemophore HasA from P. aeruginosa.[42,43]
Interestingly, it was reported that proline-proline-glutamate (PPE) proteins characterized in M. tuberculosis are involved in heme uptake. The PPE proteins are unique to mycobacteria. Tullius et al. found that PPE37 is an essential protein for efficient heme uptake as an iron source by M. tuberculosis,[44] while Mitra et al. identified novel heme-binding proteins, including PPE36, PPE62, and FecB2 (Rv0265c), with distinct roles in heme utilization.[45] PPE36 and PPE62 act as cell surface receptors for heme. The deletion of the ppe36 gene of M. tuberculosis significantly reduced heme utilization, suggesting that PPE36 is crucial for efficient heme utilization in vitro.[45] The periplasmic protein FecB2 was suggested to possibly play a role in both ferric siderophore and heme acquisition pathways. However, these proteins have low heme-binding affinity in bacterial heme uptake pathways compared to other proteins, such as IsdC and HasA.[46,47]
Due to their high structural homology to heme, these heme-uptake pathways have inspired the development of non-iron metalloporphyrins as antimicrobials.[11,48] These metalloporphyrins target either hemeproteins or pathways involved in heme uptake. Many non-iron metalloporphyrins, which mimic heme, were tested against the sexually transmitted pathogens (Neisseria gonorrhoeae and Haemophilus ducreyi), with gallium protoporphyrin IX (GaPP) showing the highest activity without causing the major side effect in vivo.[49] Likewise, GaPP inhibits the growth of P. aeruginosa by binding cytochrome in a place of heme, disrupting electron transfer during oxidative phosphorylation.[50] Recently, Zhang et al. demonstrated that water-soluble cationic gallium protoporphyrin (Ga-CHP) exerts broad-spectrum antibacterial activity against both Gram-positive and Gram-negative bacteria, including multidrug-resistant strains.[51] Additionally, Ga-CHP acts as a photosensitizer, enhancing its antibacterial efficacy through synergistic photodynamic therapy in vitro and in vivo.
Similar to siderophore-antibiotic conjugates, non-iron metalloporphyrins can be linked with antibiotics to enhance the delivery of drugs across the membrane, thereby increasing the permeability of the lipid membrane for a specific antibiotic. While no metalloporphyrin-antibiotic conjugates have been identified that exploit the bacterial heme-uptake pathways for significantly enhanced dual actions, porphyrin-antibiotic conjugates have attracted attention for their use in antibacterial photodynamic therapy (PDT). One research group prepared a 5,10,15,20-tetrakis (para-aminophenyl) porphyrin-vancomycin conjugate and demonstrated that the conjugate as a photosensitizer exerts effective and selective inhibitory activity against Staphylococcus aureus over Escherichia coli under white light.[52] A conjugate of 5(4′-carboxyphenyl)-10,15,20-triphenylporphyrin (cTPP) with the cationic antimicrobial peptide, apidaecin, exhibited broad-spectrum antibacterial activity when activated by light, effectively inhibiting bacterial growth, including P. aeruginosa, and S. aureus.[53]
2.3. Potential Targets for Ga(III)-Based Compounds
2.3.1. Ribonucleotide Reductase (RNR)
One strategy for developing new antibacterial drugs focuses on finding more selective compounds that target multiple pathways to minimize drug resistance. RNR catalyzes the transformation of ribonucleotides to deoxyribonucleotides, which are necessary for DNA synthesis and essential for cell proliferation, via a mechanism involving protein radicals.[54,55] RNR consists of two subunits, α (or R1) and β (or R2). The R2 subunit contains an indispensable binuclear iron center, where Ga(III) ion replaces iron, thereby disrupting the catalysis. There are several studies supporting the enzymatic inhibition by Ga(III). A molecular modeling study revealed that the redox-inactive Ga(III) replaces the redox-active Fe(III), leading to the inhibition of the catalysis.[56] In addition to the substitution of Fe(III) in the R2 subunit, it has been suggested that gallium nitrate can inhibit enzyme activity by forming Ga(III)-nucleotide diphosphates (NDP) complexes.[57,58,59] Density functional theory (DFT)/polarizable continuum model (PCM) study revealed that Ga(III) ion forms complexes with the free NDPs, which are not/poorly recognizable by the RNR, thus inhibiting the RNR activity in malignant cells.[58] Overall, it appears that Ga(III) inhibits RNR activity via several mechanisms, including iron deprivation and the formation of unfavorable Ga(III)-NDP complexes as substrates to RNR.
Gallium nitrate showed promising results when used in mice infected with M. tuberculosis.[60] This study demonstrated that the Ga(III) antimycobacterial activity paralleled its ability to inhibit several enzymes, including RNR and aconitase (Figure 2). In addition to bacteria, gallium nitrate inhibits eukaryotic cell proliferation by inhibiting RNR activity, accounting in part for its efficacy as an anticancer drug.[61] Consistent with the result observed in M. tuberculosis, gallium nitrate has been shown to inhibit P. aeruginosa RNR by up to ~40% at concentrations that result in growth inhibition.[8] The authors noted that increasing the gallium concentration did not lead to further inhibition, suggesting that gallium may specifically inhibit one of the two classes of P. aeruginosa RNR.[62]

Table 1.
Gallium(III) compounds and their pathogen-specific targets.
| Ga(III) | Target | Bacteria | Reference |
|---|---|---|---|
| Ga(NO3)3 | Aconitase | M. tuberculosis, M. abscessus | [60,63] |
| Ribonucleotide reductase | M. tuberculosis, P. aeruginosa PAO1 | [8,60] | |
| Catalase | M. abscessus, P. aeruginosa PAO1, K.pneumoniae, F. novicida | [8,64,65,66] | |
| Superoxide dismutases | P. aeruginosa 103 | [64] | |
| GaPP | Catalase | MRSA, K. pneumoniae, P. aeruginosa 103, M. abscessus. | [63,64,65,67] |
| Superoxide dismutases | P. aeruginosa 103 | [64] | |
| Aconitase | M. abscessus | [63] |
GaPP: Ga(III) protoporphyrin IX.
2.3.2. Oxidative Phosphorylation and Cytochrome Oxidases
The heme mimetic gallium protoporphyrin IX (GaPP) has been shown to possess broad antibacterial activity.[11,68] Stojiljkovic et al. were the first to demonstrate that non-iron metalloporphyrin compounds possess such activity, with GaPP being the most effective.[48] The authors proposed that bacteria internalize the metalloporphyrin via heme-uptake pathways. Once inside the cell, the metalloporphyrin binds to cytochromes, disrupting the electron transfer in oxidative phosphorylation (Figure 2). This disruption causes incomplete O2 reduction and generation of ROS.[48] Arrest of cellular respiration was also observed in P. aeruginosa, where GaPP inhibits the aerobic growth of P. aeruginosa by targeting cytochrome oxidases.[50] The authors investigated all five terminal oxidases of P. aeruginosa to determine which ones are responsible for GaPP sensitivity, as they require heme as a cofactor. Their findings indicated that the Cco-1, Cco-2, and Cio terminal oxidases are the primary targets for GaPP.
2.3.3. RNA Polymerase
Iron-targeting gallium compounds demonstrate antimicrobial activity through multiple mechanisms of action that remain unclear. The combined study of metalloproteomics with metabolomics and transcriptomics revealed that gallium nitrate binds RpoB and RpoC, two subunits of RNA polymerase in P. aeruginosa, leading to the reduction of metabolic rates and energy utilization by suppressing RNA synthesis.[69] Furthermore, the combination of gallium nitrate with acetate was found to be synergistic in inhibiting the growth of persister cells and was efficacious in murine P. aeruginosa infection models, attenuating bacterial virulence.
2.3.4. Aconitase
Several studies imply that Ga(III)-exposed bacteria exhibit reduced activity of multiple iron-utilizing enzymes, including aconitase, catalase, and iron superoxide dismutase. Aconitase is a Fe-sulfur enzyme that catalyzes the isomerization of citrate to isocitrate via cis-aconitate in the tricarboxylic acid cycle. The activity of aconitase from M. tuberculosis is reduced when exposed to gallium nitrate.[60] Likewise, an inhibitory effect of gallium on aconitase was observed in M. abscessus, with both gallium nitrate and GaPP inhibiting its aconitase activity.[63] The authors proposed a plausible explanation that Ga(III) ion, released from the breakdown of GaPP by bacterial heme oxygenase, may prevent the formation of the iron-sulfur cluster essential for aconitase catalysis. An alternative explanation for the reduced aconitase activity is that ROS generated by Ga(III) compounds, including GaPP, might disrupt the enzyme’s iron-sulfur cluster (see discussion below). Also, synergistic inhibition of aconitase was observed when M. abscessus was exposed to a combination of gallium nitrate and GaPP.[63]
However, the reduction of aconitase activity by Ga(III) seems to be pathogen-dependent. Gallium nitrate did not have a significant impact on P. aeruginosa aconitase activity.[8] The inhibition of aconitase activity by Ga(III) may vary due to the differences in gene regulation associated with the inhibitory activity of Ga(III) compounds between mycobacteria and Gram-negative bacteria. Thus, further study is needed to provide insight into inhibiting aconitase activity by Ga(III) in these bacterial types, including Gram-positive bacteria.
2.3.5. Oxidative Stress and Antioxidant Enzymes
Antioxidant enzymes play essential roles in protecting the cells from oxidative stress caused by ROS that damage cellular components such as DNA, proteins, and lipids. Bacteria have evolved various antioxidant enzymes, including catalases, superoxide dismutases, and peroxidases, to defend the cells from these ROS. Recent studies have shown that Ga(III) leads to ROS accumulation through two mechanisms: direct inhibition of antioxidant enzymes or indirect promotion of ROS production (Figure 3). Zemke et al. demonstrated that ROS might play a role in the antibacterial mechanisms of Ga(III) by showing that gallium nitrate exhibits a better antibacterial effect on aerobic bacteria compared to anaerobic bacteria.[70] They also demonstrated the synergistic effect of gallium nitrate and sodium nitrite as a NO donor in inhibiting P. aeruginosa growth and preventing biofilm growth under both aerobic and anaerobic conditions, indicating the involvement of ROS induced by gallium nitrate. Another study with P. fluorescens supported that Ga(III) disrupts Fe metabolism and induces oxidative stress that a reductive activity can neutralize via the overexpression of NADPH-producing enzymes.[71]
However, intracellular ROS levels measured by the ROS fluorescent probe H2DCFDA indicated that gallium nitrate did not significantly affect ROS levels in P. aeruginosa.[64,72] Despite this, gallium nitrate exhibits bacteriostatic growth inhibitory activity against P. aeruginosa, which was accompanied by decreased ATP production.[72] Thus, further studies are required to identify the cellular targets for Ga(III) ion and provide new insights into its mechanism of antibacterial action.
2.3.6. Catalase
The antibacterial effects of gallium compounds, such as Ga(III) ions or Ga(III) porphyrins, have been suggested to be partly due to the inhibition of bacterial antioxidant enzymes, resulting in an accumulation of ROS. Catalase is a key antioxidant found in many organisms, including bacteria. It converts hydrogen peroxide (H2O2) into water and oxygen.[73,74] Catalase is a tetrameric enzyme containing four Fe(II)-bound hemes, which is essential for its catalytic activity. Thus, catalase is a potential target for non-iron metalloporphyrins, including Ga(III) porphyrins, resulting in the disruption of antioxidant activity.[48,63]
Brugna et al. developed an excellent bacterial system to determine the mechanism of antibacterial non-iron metalloporphyrins.[75] The Gram-positive bacteria Enterococcus faecalis does not depend on heme for growth, leading to resistance to non-iron metalloporphyrins. The authors exploited this feature for the production of non-iron metalloporphyrin-substituted catalases. They found that catalases containing non-metalloporphyrins, especially GaPP, showed significantly reduced activity compared to heme-substituted catalase. In addition, an H2O2 consumption assay and/or native gel studies confirmed that GaPP inhibits catalase in S. aureus, P. aeruginosa, and M. abscessus.[63,64,67] These results suggest that GaPP binds to apo-catalase, rendering it inactive in the decomposition of H2O2.
A contradictory observation has been reported regarding the effect of gallium nitrate on catalase activities. Gallium nitrate exhibited pathogen/strain-dependent inhibitory activity on catalase. Gallium nitrate, which is not cofactor for catalase, reduced catalase activity in P. aeruginosa PAO1and Francisella novicida,[8,65,66] while no inhibition was observed in Staphylococcus aureus, P. aeruginosa PA103, and M. abscessus.[63,64,67] In case of P. aeruginosa strains, the authors suggested that this discrepancy might be due to varying iron concentration in the media used to grow the bacteria, indicating further study is needed to explore strain-specific or growth medium-related differences in antimicrobial activity of Ga(III) ion.[64] However, those catalase activity assays, including H2O2 consumption and native gel assays, were performed using lysates from Ga(III)-treated bacteria, whose measurements might have been affected by other chemical substances or enzymes in the lysates, resulting in inaccurate results. Guo et al. reported more convincing mechanistic studies by performing RNA sequencing (RNA-seq).[76] They demonstrated that the transcriptional levels of two superoxide dismutase-encoding genes, sodB and sodC, and two catalase-encoding genes, katE and katG, were downregulated by gallium nitrate, leading to intracellular accumulation of ROS in K. pneumoniae. Thus, it appears that gallium nitrate may alter transcriptional gene expressions rather than directly inhibiting catalase.
2.3.7. Superoxide Dismutase
The superoxide dismutases are additional key bacterial antioxidant enzymes.[77,78] Superoxide dismutases (SOD) are metalloenzymes that catalyze the dismutation of superoxide to H2O2 and O2 using a redox-active metal. To date, four isoforms of SODs have been identified: MnSOD, FeSOD, Cu/ZnSOD, and NiSOD.[79,80] MnSOD and FeSOD are typically located in the cytosol, while Cu/ZnSOD is found in periplasmic space. Since Ga(III) ion can substitute for Fe(III) in many redox enzymes, it is thought that Ga(III) may interfere with bacterial SODs by replacing a metal, most likely iron, in the SOD, leading to ROS accumulation within bacteria.
The SODs of P. aeruginosa and K. pneumoniae have been investigated as potential targets for GaPP and gallium nitrate.[8,64,65] Similar to catalase, these studies yielded inconsistent results: Gallium nitrate inhibited SOD activity from P. aeruginosa PA103 but did not from P. aeruginosa PAO1 and K. pneumoniae. In addition, a study with Ga(III)-chelated transferrin or lactoferrin reduced FeSOD activity in F. novicida.[66] The inconsistencies could be attributed to differences in iron concentrations in the growth media, pathogen-specific variations, or interference from the assays using bacterial lysates. Thus, SOD, as one of the potential targets for Ga(III), needs to be evaluated by performing alternative assays, such as RNA sequencing and proteomics.
2.3.8. Peroxidases
Bacterial peroxidases play a crucial role in protecting cells from oxidative damage. These enzymes convert H2O2 and organic peroxides into water and alcohols, respectively. Catalase-peroxidase (KatG) and alkyl hydroperoxide reductase (ahpC) limit oxidative stress in M. tuberculosis. KatG is a bifunctional heme-containing enzyme that protects M. tuberculosis from H2O2. Notably, unlike other mycobacteria, M. tuberculosis relies solely on KatG for H2O2 detoxification. In addition, KatG also possesses peroxynitritase activity,[81] indicating that KatG is an important virulence factor. Given this unique dependency on KatG for protection, Ga(III) porphyrins, acting as a heme mimic, could serve as potential antitubercular agents against M. tuberculosis by targeting this enzyme. However, the effectiveness and mechanisms of Ga(III) porphyrins as antitubercular agents, particularly in vivo, remain to be fully elucidated.
Interestingly, gallium nitrate enhanced the iron-containing peroxidase activity of Chrorella pyrenoidosa in both iron-deficient and normal iron-supplied cultures.[82] Similarly, the effects of gallium curcumin and gallium diacetylcurcumin on the structure, function, and oxidative stability of the horseradish peroxidase (HRP) enzyme were evaluated.[83] These gallium complexes increased the antioxidant activity of the peroxidase enzyme, suggesting that the complexes exhibit the potential for cancer treatment, but they have no significant antibacterial activity.
3. Conclusion and Prospective Views
Iron is an essential element for pathogenic bacterial survival in human hosts, playing a critical role in multiple biological processes. Due to their similar physicochemical properties to iron, gallium-based compounds have emerged as potential antimicrobial agents. Thus, gallium, acting as a multitarget antimicrobial agent, has been demonstrated to exert a broad antibacterial activity by interrupting iron/heme metabolism. Notably, it shows inhibitory activity against pathogenic bacteria that are resistant to current antibiotics. Numerous enzymes have been identified as targets for Ga(III) ions and Ga(III) porphyrins. One compelling mechanism of action for Ga(III)-based compounds is their ability to increase the production of ROS in bacteria by disrupting enzymatic antioxidants, such as SODs, catalases, peroxidases, or non-enzymatic antioxidant processes. Thus, Ga(III) enhances the host’s oxidative defenses during the immune response to pathogenic infection. However, it appears that Ga(III)’s inhibitory activity and mechanism(s) of action are pathogen-dependent (Table 1). For example, E. coli SOD and catalase mutants exert hypersensitivity to GaPP due to the overproduction of ROS.[48] In contrast, MRSA catalase and SOD transposants do not show increased susceptibility to GaPP and gallium nitrate.[67] Furthermore, MRSA transposon mutants with defects in antioxidant activity, such as thiol peroxidase, alkyl hydroperoxide reductase (AhpC), and staphyloxanthin, exhibit susceptibility to these gallium compounds comparable to that of MRSA USA300. These findings suggest that Ga(III)’s antimicrobial activity involves targeting multiple iron/heme-dependent biological processes and that ROS produced by Ga(III) is not the only factor contributing to bacterial killing. While gallium shows promise as an antimicrobial agent, further research is needed to elucidate the mechanism of the antibacterial activity of gallium-based compounds to improve their efficacy as antibiotics.
References
- Turner, RJ. Metal-based antimicrobial strategies. Microb Biotechnol. 2017, 10, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Turner, RJ. The good, the bad, and the ugly of metals as antimicrobials. Biometals. 2024, 37, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Frei, A; Verderosa, AD; Elliott, AG; Zuegg, J; Blaskovich, MAT. Metals to combat antimicrobial resistance. Nat Rev Chem. 2023, 7, 202–224. [Google Scholar] [CrossRef] [PubMed]
- Kurtuldu F, Mutlu N, Boccaccini AR, Galusek D. Gallium containing bioactive materials: A review of anticancer, antibacterial, and osteogenic properties. Bioact Mater. 2022, 17: 125-146.
- Bernstein, LR. Mechanisms of therapeutic activity for gallium. Pharmacol Rev. 1998, 50, 665–682. [Google Scholar] [PubMed]
- Chitambar, CR. Gallium and its competing roles with iron in biological systems. Biochim Biophys Acta. 2016, 1863, 2044–2053. [Google Scholar] [CrossRef] [PubMed]
- Li, F; Liu, F; Huang, K; Yang, S. Advancement of Gallium and Gallium-Based Compounds as Antimicrobial Agents. Front Bioeng Biotechnol. 2022, 10, 827960. [Google Scholar] [CrossRef] [PubMed]
- Goss CH, Kaneko Y, Khuu L, et al. Gallium disrupts bacterial iron metabolism and has therapeutic effects in mice and humans with lung infections. Sci Transl Med. 2018;10(460).
- Hacht, B. Gallium(III) Ion Hydrolysis under Physiological Conditions. Bulletin of The Korean Chemical Society. 2008, 29, 372–376. [Google Scholar]
- Chitambar, CR. The therapeutic potential of iron-targeting gallium compounds in human disease: From basic research to clinical application. Pharmacol Res. 2017, 115, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Kelson, AB; Carnevali, M; Truong-Le, V. Gallium-based anti-infectives: targeting microbial iron-uptake mechanisms. Curr Opin Pharmacol. 2013, 13, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, LR; Tanner, T; Godfrey, C; Noll, B. Chemistry and pharmacokinetics of gallium maltolate, a compound with high oral gallium bioavailability. Met Based Drugs. 2000, 7, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Collery, P; Domingo, JL; Keppler, BK. Preclinical toxicology and tissue gallium distribution of a novel antitumour gallium compound: tris (8-quinolinolato) gallium (III). Anticancer Res. 1996, 16, 687–691. [Google Scholar] [PubMed]
- Darwesh AMF, Imberti C, Bartnicka JJ, et al. In Vivo Trafficking of the Anticancer Drug Tris(8-Quinolinolato) Gallium (III) (KP46) by Gallium-68/67 PET/SPECT Imaging. Molecules. 2023, 28.
- Valiahdi, SM; Heffeter, P; Jakupec, MA; et al. The gallium complex KP46 exerts strong activity against primary explanted melanoma cells and induces apoptosis in melanoma cell lines. Melanoma Res. 2009, 19, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Duffin, RN; Blair, VL; Kedzierski, L; Andrews, PC. Alkyl gallium(III) quinolinolates: A new class of highly selective anti-leishmanial agents. Eur J Med Chem. 2020, 186, 111895. [Google Scholar] [CrossRef] [PubMed]
- Shi, F; Ma, S; Liu, S; et al. A novel antimicrobial strategy for bacterial infections: Gallium-based materials. Colloid and Interface Science Communications. 2023, 56, 100735. [Google Scholar] [CrossRef]
- Leong, M; Parker, CJ; Shaw, ZL; et al. Metallic Gallium Droplets Exhibit Poor Antibacterial Properties. ACS Appl Mater Interfaces. 2024, 16, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, N; Hurrell, R; Kelishadi, R. Review on iron and its importance for human health. J Res Med Sci. 2014, 19, 164–174. [Google Scholar] [PubMed]
- Cassat, JE; Skaar, EP. Iron in infection and immunity. Cell Host Microbe. 2013, 13, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Wandersman C, Delepelaire P. Bacterial iron sources: from siderophores to hemophores. Annu Rev Microbiol. 2004, 58: 611-647.
- Wilson, BR; Bogdan, AR; Miyazawa, M; Hashimoto, K; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Holden, VI; Bachman, MA. Diverging roles of bacterial siderophores during infection. Metallomics. 2015, 7, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Parrow, NL; Fleming, RE; Minnick, MF. Sequestration and scavenging of iron in infection. Infect Immun. 2013, 81, 3503–3514. [Google Scholar] [CrossRef] [PubMed]
- Sebbane, F; Jarrett, C; Gardner, D; Long, D; Hinnebusch, BJ. Role of the Yersinia pestis yersiniabactin iron acquisition system in the incidence of flea-borne plague. PLoS One. 2010, 5, e14379. [Google Scholar] [CrossRef] [PubMed]
- Banin, E; Lozinski, A; Brady, KM; et al. The potential of desferrioxamine-gallium as an anti-Pseudomonas therapeutic agent. Proc Natl Acad Sci U S A. 2008, 105, 16761–16766. [Google Scholar] [CrossRef] [PubMed]
- Frangipani, E; Bonchi, C; Minandri, F; Imperi, F; Visca, P. Pyochelin potentiates the inhibitory activity of gallium on Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2014, 58, 5572–5575. [Google Scholar] [CrossRef]
- Pandey, A; Savino, C; Ahn, SH; Yang, Z; Van Lanen, SG; Boros, E. Theranostic Gallium Siderophore Ciprofloxacin Conjugate with Broad Spectrum Antibiotic Potency. J Med Chem. 2019, 62, 9947–9960. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. Iron, mycobacteria and tuberculosis. Tuberculosis (Edinb). 2004, 84, 110–130. [Google Scholar] [CrossRef] [PubMed]
- Gobin, J; Horwitz, MA. Exochelins of Mycobacterium tuberculosis remove iron from human iron-binding proteins and donate iron to mycobactins in the M. tuberculosis cell wall. J Exp Med. 1996, 183, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Gobin, J; Moore, CH; Reeve JR, Jr; Wong, DK; Gibson, BW; Horwitz, MA. Iron acquisition by Mycobacterium tuberculosis: isolation and characterization of a family of iron-binding exochelins. Proc Natl Acad Sci U S A. 1995, 92, 5189–5193. [Google Scholar] [CrossRef] [PubMed]
- Meneghetti, F; Villa, S; Gelain, A; et al. Iron Acquisition Pathways as Targets for Antitubercular Drugs. Curr Med Chem. 2016, 23, 4009–4026. [Google Scholar] [CrossRef] [PubMed]
- Shyam, M; Shilkar, D; Verma, H; et al. The Mycobactin Biosynthesis Pathway: A Prospective Therapeutic Target in the Battle against Tuberculosis. J Med Chem. 2021, 64, 71–100. [Google Scholar] [CrossRef]
- Shyam, M; Shilkar, D; Rakshit, G; Jayaprakash, V. Approaches for targeting the mycobactin biosynthesis pathway for novel anti-tubercular drug discovery: where we stand. Expert Opin Drug Discov. 2022, 17, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Miller, MJ; Walz, AJ; Zhu, H; et al. Design, synthesis, and study of a mycobactin-artemisinin conjugate that has selective and potent activity against tuberculosis and malaria. J Am Chem Soc. 2011, 133, 2076–2079. [Google Scholar] [CrossRef] [PubMed]
- Olakanmi, O; Britigan, BE; Schlesinger, LS. Gallium disrupts iron metabolism of mycobacteria residing within human macrophages. Infect Immun. 2000, 68, 5619–5627. [Google Scholar] [CrossRef] [PubMed]
- Tandara, L; Salamunic, I. Iron metabolism: current facts and future directions. Biochem Med (Zagreb). 2012, 22, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Contreras, H; Chim, N; Credali, A; Goulding, CW. Heme uptake in bacterial pathogens. Current Opinion in Chemical Biology. 2014, 19, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y; Guo, M. Bacterial heme-transport proteins and their heme-coordination modes. Arch Biochem Biophys. 2009, 481, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Richard KL, Kelley BR, Johnson JG. Heme Uptake and Utilization by Gram-Negative Bacterial Pathogens. Front Cell Infect Microbiol. 2019, 9: 81.
- Chao, A; Sieminski, PJ; Owens, CP; Goulding, CW. Iron Acquisition in Mycobacterium tuberculosis. Chem Rev. 2019, 119, 1193–1220. [Google Scholar] [CrossRef] [PubMed]
- Owens, CP; Du, J; Dawson, JH; Goulding, CW. Characterization of heme ligation properties of Rv0203, a secreted heme binding protein involved in Mycobacterium tuberculosis heme uptake. Biochemistry. 2012, 51, 1518–1531. [Google Scholar] [CrossRef] [PubMed]
- Owens, CP; Chim, N; Graves, AB; et al. The Mycobacterium tuberculosis secreted protein Rv0203 transfers heme to membrane proteins MmpL3 and MmpL11. J Biol Chem. 2013, 288, 21714–21728. [Google Scholar] [CrossRef] [PubMed]
- Tullius MV, Nava S, Horwitz MA. PPE37 Is Essential for Mycobacterium tuberculosis Heme-Iron Acquisition (HIA), and a Defective PPE37 in Mycobacterium bovis BCG Prevents HIA. Infect Immun. 2019, 87.
- Mitra, A; Speer, A; Lin, K; Ehrt, S; Niederweis, M. PPE Surface Proteins Are Required for Heme Utilization by Mycobacterium tuberculosis. PPE Surface Proteins Are Required for Heme Utilization by Mycobacterium tuberculosis. MBio. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Liu, M; Tanaka, WN; Zhu, H; Xie, G; Dooley, DM; Lei, B. Direct hemin transfer from IsdA to IsdC in the iron-regulated surface determinant (Isd) heme acquisition system of Staphylococcus aureus. J Biol Chem. 2008, 283, 6668–6676. [Google Scholar] [CrossRef] [PubMed]
- Deniau, C; Gilli, R; Izadi-Pruneyre, N; et al. Thermodynamics of heme binding to the HasA(SM) hemophore: effect of mutations at three key residues for heme uptake. Biochemistry. 2003, 42, 10627–10633. [Google Scholar] [CrossRef] [PubMed]
- Stojiljkovic, I; Kumar, V; Srinivasan, N. Non-iron metalloporphyrins: potent antibacterial compounds that exploit haem/Hb uptake systems of pathogenic bacteria. Mol Microbiol. 1999, 31, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Bozja, J; Yi, K; Shafer, WM; Stojiljkovic, I. Porphyrin-based compounds exert antibacterial action against the sexually transmitted pathogens Neisseria gonorrhoeae and Haemophilus ducreyi. Int J Antimicrob Agents. 2004, 24, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Hijazi S, Visca P, Frangipani E. Gallium-Protoporphyrin IX Inhibits Pseudomonas aeruginosa Growth by Targeting Cytochromes. Front Cell Infect Microbiol. 2017, 7.
- Zhang, H; Li, Q; Qi, X; et al. Iron-blocking antibacterial therapy with cationic heme-mimetic gallium porphyrin photosensitizer for combating antibiotic resistance and enhancing photodynamic antibacterial activity. Chemical Engineering Journal. 2023, 451, 138261. [Google Scholar] [CrossRef]
- Zhai L, Yang K-W. Porphyrin-vancomycin: A highly promising conjugate for the identification and photodynamic inactivation of antibiotic resistant Gram-positive pathogens. Dyes and Pigments. 2015, 120: 228-238.
- Dosselli, R; Gobbo, M; Bolognini, E; Campestrini, S; Reddi, E. Porphyrin−Apidaecin Conjugate as a New Broad Spectrum Antibacterial Agent. ACS Medicinal Chemistry Letters. 2010, 1, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, P; Reichard, P. Ribonucleotide reductases. Annu Rev Biochem. 2006, 75, 681–706. [Google Scholar] [CrossRef] [PubMed]
- Sintchak, MD; Arjara, G; Kellogg, BA; Stubbe, J; Drennan, CL. The crystal structure of class II ribonucleotide reductase reveals how an allosterically regulated monomer mimics a dimer. Nat Struct Biol. 2002, 9, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, V; Angelova, S; Markova, N; Dudev, T. Gallium as a Therapeutic Agent: A Thermodynamic Evaluation of the Competition between Ga(3+) and Fe(3+) Ions in Metalloproteins. J Phys Chem B. 2016, 120, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Chitambar, CR; Antholine, WE. Iron-targeting antitumor activity of gallium compounds and novel insights into triapine(®)-metal complexes. Antioxid Redox Signal. 2013, 18, 956–972. [Google Scholar] [CrossRef] [PubMed]
- Kircheva, N; Dudev, T. Novel Insights into Gallium’s Mechanism of Therapeutic Action: A DFT/PCM Study of the Interaction between Ga(3+) and Ribonucleotide Reductase Substrates. J Phys Chem B. 2019, 123, 5444–5451. [Google Scholar] [CrossRef] [PubMed]
- Marzilli, LG; De Castro, B; Caradonna, JP; Stewart, RC; Van Vuuren, CP. Nucleoside complexing. A Raman and carbon-13 NMR spectroscopic study of the binding of hard and soft metal species. Journal of the American Chemical Society. 1980, 102, 916–924. [Google Scholar] [CrossRef]
- Olakanmi, O; Kesavalu, B; Pasula, R; Abdalla, MY; Schlesinger, LS; Britigan, BE. Gallium nitrate is efficacious in murine models of tuberculosis and inhibits key bacterial Fe-dependent enzymes. Antimicrob Agents Chemother. 2013, 57, 6074–6080. [Google Scholar] [CrossRef] [PubMed]
- Smith BD, Karp JE. Ribonucleotide reductase: an old target with new potential. Leuk Res. Vol 27. England; 2003:1075-1076.
- Jordan, A; Torrents, E; Sala, I; Hellman, U; Gibert, I; Reichard, P. Ribonucleotide reduction in Pseudomonas species: simultaneous presence of active enzymes from different classes. J Bacteriol. 1999, 181, 3974–3980. [Google Scholar] [CrossRef] [PubMed]
- Choi S-r, Switzer B, Britigan BE, Narayanasamy P. Gallium Porphyrin and Gallium Nitrate Synergistically Inhibit Mycobacterial Species by Targeting Different Aspects of Iron/Heme Metabolism. ACS Infectious Diseases 2020.
- Scott, ZW; Choi, SR; Talmon, GA; Britigan, BE; Narayanasamy, P. Combining Gallium Protoporphyrin and Gallium Nitrate Enhances In Vitro and In Vivo Efficacy against Pseudomonas aeruginosa: Role of Inhibition of Bacterial Antioxidant Enzymes and Resultant Increase in Cytotoxic Reactive Oxygen Species. ACS Infect Dis. 2022, 8, 2096–2105. [Google Scholar] [CrossRef] [PubMed]
- Scott Z, Choi S-r, Britigan BE, Narayanasamy P. Dual Gallium Drug Treatment Against Carbapenem-Resistant Klebsiella Pneumoniae: Efficacy and Potential Mechanism(s) of Action and Resistance. Advanced Therapeutics. 2024, n/a(n/a), 2400147.
- Olakanmi, O; Gunn, JS; Su, S; Soni, S; Hassett, DJ; Britigan, BE. Gallium disrupts iron uptake by intracellular and extracellular Francisella strains and exhibits therapeutic efficacy in a murine pulmonary infection model. Antimicrob Agents Chemother. 2010, 54, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Choi, SR; Talmon, GA; Hearne, K; et al. Combination Therapy with Gallium Protoporphyrin and Gallium Nitrate Exhibits Enhanced Antimicrobial Activity In Vitro and In Vivo against Methicillin-Resistant Staphylococcus aureus. Mol Pharm. 2023, 20, 4058–4070. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, S; Visaggio, D; Pirolo, M; Frangipani, E; Bernstein, L; Visca, P. Antimicrobial Activity of Gallium Compounds on ESKAPE Pathogens. Antimicrobial Activity of Gallium Compounds on ESKAPE Pathogens. Front Cell Infect Microbiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y; Han, B; Xie, Y; et al. Combination of gallium(iii) with acetate for combating antibiotic resistant Pseudomonas aeruginosa. Chem Sci. 2019, 10, 6099–6106. [Google Scholar] [CrossRef] [PubMed]
- Zemke AC, Madison CJ, Kasturiarachi N, Pearce LL, Peterson J. Antimicrobial Synergism Toward Pseudomonas aeruginosa by Gallium(III) and Inorganic Nitrite. Front Microbiol. 2020, 11: 2113.
- Bériault, R; Hamel, R; Chenier, D; Mailloux, RJ; Joly, H; Appanna, VD. The overexpression of NADPH-producing enzymes counters the oxidative stress evoked by gallium, an iron mimetic. Biometals. 2007, 20, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Wang Y, Yang X, Zhang S, et al. Comparative proteomics unveils the bacteriostatic mechanisms of Ga(III) on the regulation of metabolic pathways in Pseudomonas aeruginosa. J Proteomics. 2023, 289: 105011.
- Deisseroth, A; Dounce, AL. Catalase: Physical and chemical properties, mechanism of catalysis, and physiological role. Physiol Rev. 1970, 50, 319–375. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A; Yan, LJ; Jana, CK; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid Med Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef] [PubMed]
- Brugna, M; Tasse, L; Hederstedt, L. In vivo production of catalase containing haem analogues. Febs j. 2010, 277, 2663–2672. [Google Scholar] [CrossRef] [PubMed]
- Guo, M; Tian, P; Li, Q; et al. Gallium Nitrate Enhances Antimicrobial Activity of Colistin against Klebsiella pneumoniae by Inducing Reactive Oxygen Species Accumulation. Microbiol Spectr. 2023, 11, e0033423. [Google Scholar] [CrossRef] [PubMed]
- Fasnacht, M; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front Mol Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef] [PubMed]
- Broxton, CN; Culotta, VC. SOD Enzymes and Microbial Pathogens: Surviving the Oxidative Storm of Infection. PLoS Pathog. 2016, 12, e1005295. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y; Abreu, IA; Cabelli, DE; et al. Superoxide dismutases and superoxide reductases. Chem Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef] [PubMed]
- Najmuldeen, H; Alghamdi, R; Alghofaili, F; Yesilkaya, H. Functional assessment of microbial superoxide dismutase isozymes suggests a differential role for each isozyme. Free Radic Biol Med. 2019, 134, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Wengenack, NL; Jensen, MP; Rusnak, F; Stern, MK. Mycobacterium tuberculosis KatG is a peroxynitritase. Biochem Biophys Res Commun. 1999, 256, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Simon, L; Kiss, F; Bakó, A; et al. Effect of gallium on photosynthetic pigments and peroxidase activity of Chlorella pyrenoidosa. Journal of Plant Nutrition. 1989, 12, 1123–1140. [Google Scholar] [CrossRef]
- Jahangoshaei, P; Hassani, L; Mohammadi, F; Hamidi, A; Mohammadi, K. Investigating the effect of gallium curcumin and gallium diacetylcurcumin complexes on the structure, function and oxidative stability of the peroxidase enzyme and their anticancer and antibacterial activities. J Biol Inorg Chem. 2015, 20, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Bacterial iron/heme uptake pathways.
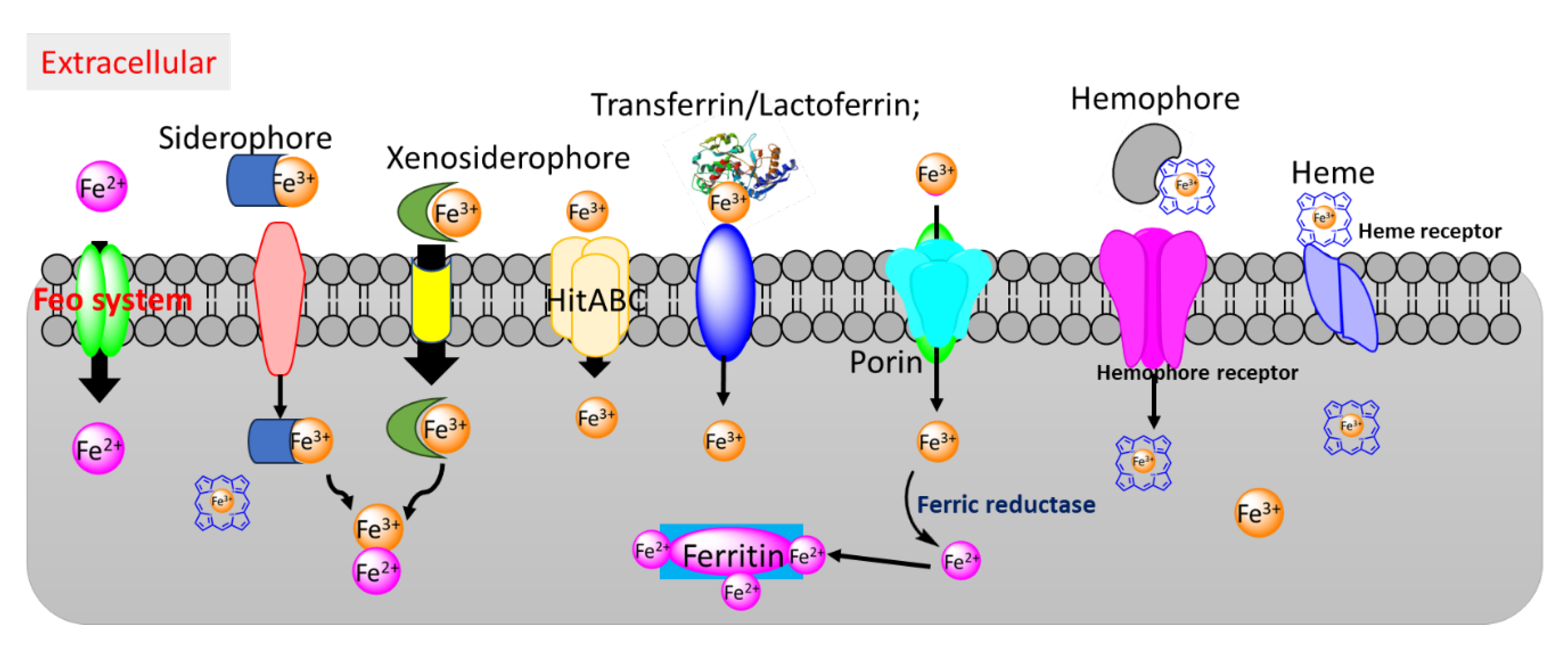
Figure 2.
Schematic diagram of the uptake of gallium-based compounds through iron/heme acquisition pathways and their proposed intracellular targets.
Figure 2.
Schematic diagram of the uptake of gallium-based compounds through iron/heme acquisition pathways and their proposed intracellular targets.
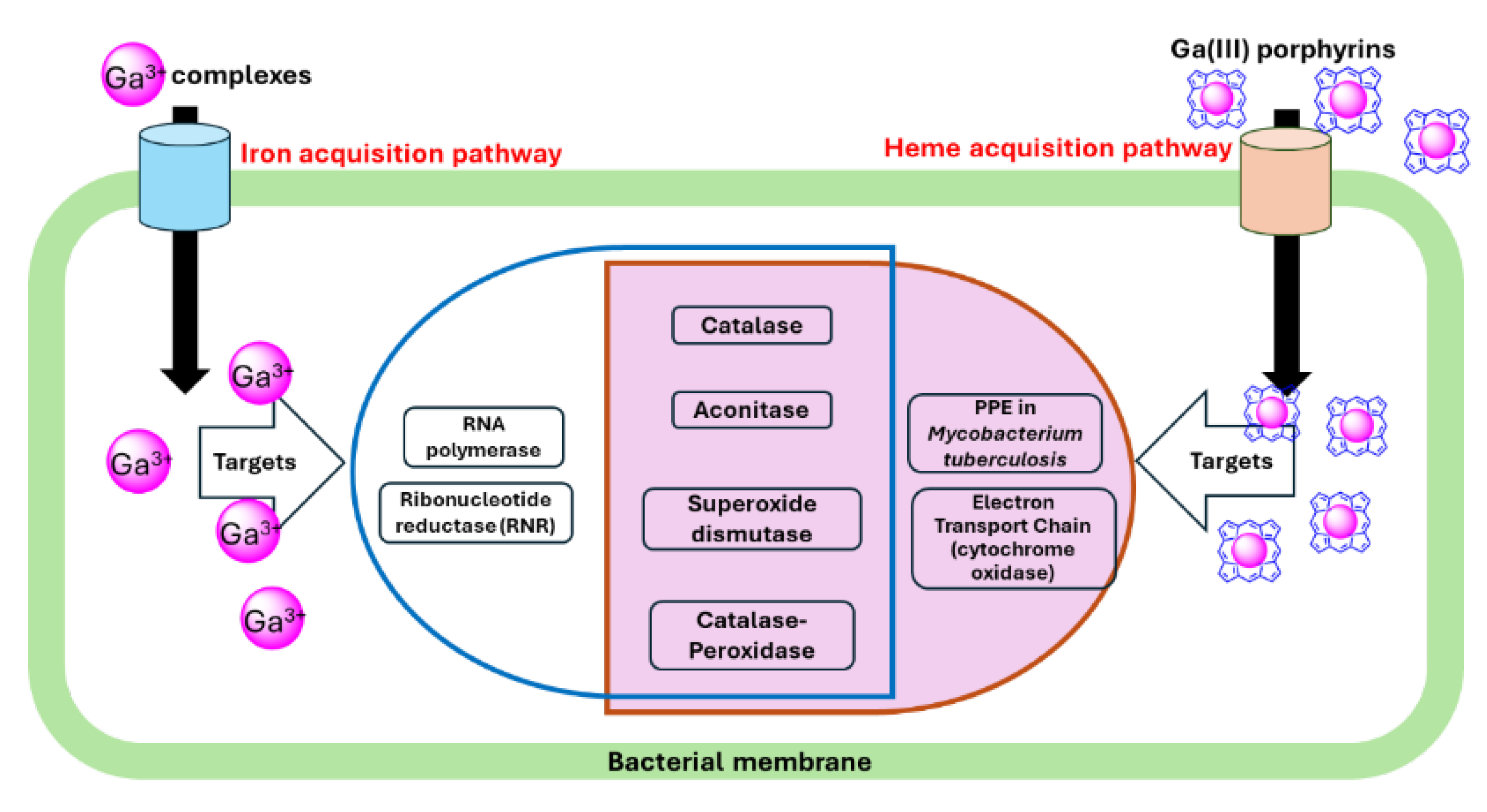
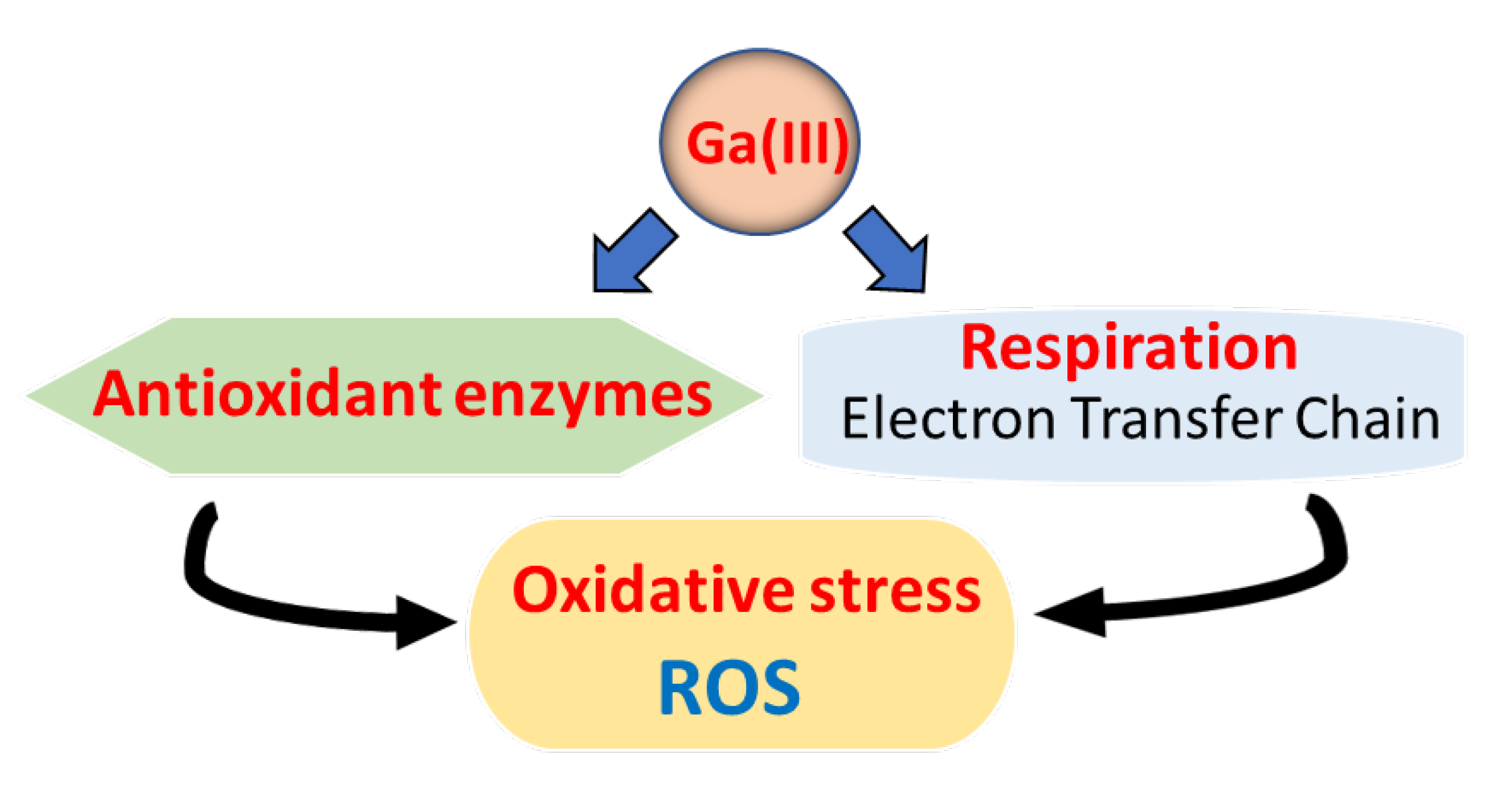
Figure 3.
Reactive oxygen species (ROS) induced by Ga(III) in bacteria. Ga(III) compounds can inhibit antioxidant enzymes, accumulating ROS within bacteria. In addition, Ga(III) porphyrin, such as GaPP, disrupts the electron transfer chain (ETS) by replacing heme in respiratory enzymes, such as cytochrome c. This leads to incomplete oxygen reduction, generating superoxide and other ROS.
Figure 3.
Reactive oxygen species (ROS) induced by Ga(III) in bacteria. Ga(III) compounds can inhibit antioxidant enzymes, accumulating ROS within bacteria. In addition, Ga(III) porphyrin, such as GaPP, disrupts the electron transfer chain (ETS) by replacing heme in respiratory enzymes, such as cytochrome c. This leads to incomplete oxygen reduction, generating superoxide and other ROS.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



