
Submitted:
31 July 2024
Posted:
01 August 2024
You are already at the latest version
Abstract
In July 2017, a family of three members; a 46-year-old male, a 45-year-old female and their 8-year-old daughter, returned to South Africa from Thailand. They presented symptoms consistent with mosquito-borne diseases, including fever, headaches, severe body aches and nausea. Mosquito bites in all family members suggested recent exposure to arthropod-borne viruses. Dengue virus 1 (Genus Orthoflavivirus) was isolated (isolate no. SA397) from the serum of the 45-year-old female via intracerebral injection in neonatal mice and subsequent passage in VeroE6 cells. Phylogenetic analysis of this strain indicated close genetic identity with cosmopolitan genotype 1 DENV1 strains from Southeast Asia, assigned to major lineage K, minor lineage 1 (DENV1I_K.1), such as GZ8H (99.92%) collected in November 2018 from China and DV1I-TM19-74 isolate (99.72%) identified in Bangkok, Thailand, in 2019. Serum samples from the 46-year-old male yielded a virus isolate that could not be confirmed as DENV1, prompting unbiased metagenomic sequencing for virus identification and characterization. Illumina sequencing identified multiple segments of a Mammalian orthoreovirus (MRV), designated Human/SA395/SA/2017. Genomic and phylogenetic analysis classified Human/SA395/SA/2017 as MRV-3 and assigned a tentative genotype MRV-3d based on the S1 segment. Genomic analyses suggested Human/SA395/SA/2017 may have originated from reassortments of segments among swine, bat, and human MRVs. The closest identity of the viral attachment protein σ1 (S1) was related to a human isolate identified from Tahiti, French Polynesia, in 1960. This indicates ongoing circulation and cocirculation of Southeast Asian and Polynesian strains, but detailed knowledge is hampered by limited availability of genomic surveillance. This case represents a rare concurrent importation of two distinct viruses with different transmission routes in the same family with similar clinical presentations. It highlights the complexities of diagnosing diseases with similar sequelae in travellers returning from tropical areas.
Keywords:
dengue virus
; mammalian orthoreovirus
; MRV
; imported infections
; clinical metagenomics
; metagenomics
1. Introduction
The global movement of people significantly contributes to the importation of exotic viral diseases into non-endemic regions. This phenomenon is particularly pronounced in the global spread of arboviruses, viruses transmitted by arthropods such as mosquitoes and ticks. Dengue virus (DENV), a leading member of this group, is a leading arthropod-borne virus affecting humans worldwide [1]. Dengue fever, caused by DENV, is a significant public health concern, affecting an estimated 390 million people each year and endemic to 120 countries, primarily in Southeast Asia, South Asia, and South America [2]. Dengue fever typically manifests with high fever, severe headache, myalgia, arthralgia, nausea, emesis, and cutaneous rashes. Severe cases escalate to dengue hemorrhagic fever with increased bleeding, thrombocytopenia, and plasma leakage [3,4].
Dengue virus is primarily transmitted to humans primarily by the bite of female mosquitoes of the genus Aedes [5]. From 2000 to 2016, 176 laboratory-confirmed dengue cases were identified in returned travelers, underscoring the risk of disease importation through international travel in South Africa [6]. Historically, South Africa has experienced at least three outbreaks of autochthonous dengue transmission, such as those in KwaZulu-Natal in 1897, 1901 and notably in 1926/1927, which resulted in 50,000 cases and 60 fatalities. Yet, DENV did not become endemic following these outbreaks [6].
Mammalian orthoreovirus (MRV) is a non-enveloped, segmented double-stranded RNA virus of the family Reoviridae, genus Orthoreovirus [7]. MRVs are generalist and ubiquitous pathogens with a broad mammalian host range and are divided into three major genotypes and serotypes (MRV-1, MRV-2, and MRV-3). A fourth serotype (MRV-4) exists with only one isolate, Ndelle reovirus [8]. MRV typing is primarily distinguished by the S1 segment, which encodes the viral attachment protein σ1 and non-structural protein σ1s, believed to govern tissue tropism and host range [9]. Three serotypes have been delineated based on the capacity of type-specific antisera to neutralize virus infectivity and inhibit hemagglutination. The prototype isolates are type 1 Lang (T1L), type 2 Jones (T2J), type 3 Dearing (T3D) and Abney (T3A).
The MRV genome comprises ten linear double-stranded RNA (dsRNA) segments in equimolar ratios within the virion. The segments have terminal non-translated regions that vary in length, and the major open reading frames (ORFs) range from 1059 to 3867 base pairs. The untranslated regions are compatible with other serotypes, allowing for extensive inter-serotype reassortment and generating mosaics of viruses with assorted segments from bat, porcine, bovine, and human hosts.
The zoonotic potential of MRVs is notable, as they are commonly spread via fecal-oral or respiratory routes and are frequently found in wastewater surveillance worldwide [10,11,12]. In South Africa, a divergent Orthoreovirus was identified from Eucampsipoda africana bat flies [13], but no MRVs have been reported, nor have serologic surveillance studies been undertaken. MRVs are usually associated with asymptomatic or mild respiratory [14,15] and enteric infections such as acute or persistent gastroenteritis [16,17,18,19,20]. Severe cases, however, have been documented in children associated with necrotizing encephalopathy [21] and meningitis [22]. MRVs have been identified extensively in pigs and bats in Europe [23,24,25], and generally, MRV causes limited pathology in infected bats [26]. Two independent experimental infection studies of MRV3 in pigs have demonstrated 100% mortality in neonatal pigs, with development of acute gastroenteritis and severe diarrhea within 72 hours of infection [27,28].
Here, we report the isolation and genetic characterization of DENV1 and MRV3 from a family returning to South Africa from Thailand in July 2017. This case represents a rare instance of concurrent importation of two different viruses with distinct transmission routes, highlighting the complexity associated with diagnosing tropical diseases in returning travelers, and the impact of global travel on pathogen spread
2. Materials and Methods
Sample Preparation, Virus Isolation and Sequencing
Clotted blood samples were submitted to the Arbovirus Reference Laboratory, Centre for Emerging, Zoonotic and Parasitic Diseases, National Institute for Communicable Diseases in Sandringham for arbovirus investigations. The serum was separated by centrifugation. Serology for arbovirus antibodies was performed using a hemagglutination inhibition screening assay [29] to detect antibodies to both endemic and exotic arboviruses, including dengue, chikungunya, West Nile and Sindbis. Virus isolation by intracerebral inoculation of suckling mice is routinely performed on suspected arbovirus cases at the Arbovirus Reference Laboratory as part of passive arbovirus surveillance for existing, new and emerging viruses.
Virus isolation was attempted by intracerebral inoculation of suckling mice (NIH strain) with the patient serum. After mice succumbed to virus infection, brain homogenates were clarified and passaged in VeroE6 cells. For the SA397 sample, intracerebral inoculation of suckling mice for two passages was performed and passaged once on VeroE6 (ATCC No. CRL-1586) green monkey kidney cells. For the SA397 sample, intracerebral inoculation of suckling mice was undertaken for three passages and two passages of VeroE6 cells, as previously described [30].
Viral RNA was extracted from the clarified supernatants and mouse homogenates using the Qiagen Viral RNA Mini Kit. Virus cDNA was prepared as described before [31]. Sequencing libraries were prepared using the Nextera DNA library preparation kit recommended by the manufacturer (Illumina, USA) and sequenced on the MiSeq Illumina platform. Random hexamer and adapter sequences were removed from the reads using fastp (v0.23.2) [32] under default settings. Clean reads were assembled using SPAdes (v3.15.5, --rnaviral flag), and viral contigs were identified using BLASTx as previously described [33]. SPAdes assembled DENV-1 and MRV-3 genome segments were remapped to the reference using BWA-MEM (v0.7.13-r1126) with default settings, and coverage was manually inspected using the Integrative Genomics Viewer (v2.3) [34]. Nucleotide sequences determined in this study were screened for vector contamination using NCBI VecScreen (https://www.ncbi.nlm.nih.gov/tools/vecscreen/) and have been deposited in GenBank under the accession numbers PP953510-PP953519.
Phylogenetic Analysis
For phylogenetic placement of the DENV1 strain, 43 DENV1 genotype I (major lineages A,B,D,H,J,K), 4 genotype II, 3 genotype III, 6 genotype IV, 3 genotype V, 4 genotype VI were downloaded from Genbank and aligned using MAFFT (v7.475, L-INS-i method). The DENV subtyping tool, as per the new Hill et al., 2024 nomenclature [35], was used to identify the genotypes and lineages with GenomeDetective (https://www.genomedetective.com/app/typingtool/dengue/) [36,37]. For phylogenetic placement of the S1 segment of the SA strain 41 strains of MRV3, MRV2 (10 isolates), and MRV1 (8 isolates) were aligned, and metadata such as collection date, host collection and country of origin were taken from Genbank or original publications. As MRV is a studied tool for oncolytic therapy [38] all isolates passed extensively in cells or adapted to cell lines were excluded from the phylogenetic inference. The resultant DENV1 and MRV S1 multiple sequence alignments were used to construct a consensus maximum-likelihood phylogenetic tree using IQ-TREE2 (v2.1.2) using the GTR+F+G4 nucleotide substitution model for (MORV S1) and TIM2+F+G4 (DENV1) as selected using the Bayesian Information Criterion in the IQ-TREE2 ModelFinder [39]. The resultant consensus tree from combined bootstrap trees was visualised using FigTree v1.4 (http://tree.bio.ed.ac.uk/software/figtree/).
3. Results
Case Presentation
In July 2017, a family of three from the Western Cape who had recently returned to South Africa from Thailand between 30/06/2017 and 16/07/2017 presented with symptoms consistent with mosquito-borne diseases (Table 1). Date of onset of symptoms was only recorded for one patient (SA397) as 16 July (date of arrival back in South Africa), and blood samples were collected from all three patients on 17 July (one day after arrival in South Africa). The family consisted of a 46-year-old male (SA395), an 8-year-old female (SA396), and a 45-year-old female (SA397). They all reported fever along with severe body aches. Patient SA396 also experienced nausea. Each family member confirmed mosquito bites during their travel. The absence of a rash, a common sign in many mosquito-borne illnesses, was noted in all three cases, and no one was hospitalized. Serology towards arboviruses using hemagglutination inhibition assays (HAI) revealed no HAI inhibition against DENV in any samples, which aligns with antibodies not being detectable yet during the acute phase of infection (patient SA397). The only positive in HAI was a low titer of antibodies in SA396 against the Chikungunya virus (CHIKV, Alphavirus), but this was not further investigated due to no virus being isolated from the patient sample
Assembly and Phylogenetic Position of Imported DENV-1 Genome
Sera samples from patient SA397 were propagated in suckling mice for two passages and once in VeroE6 cells and cross referenced with deep sequencing conducted on RNA isolated from the serum directly. Unbiased deep sequencing and de novo assembly revealed a single contig 10649 nt in length with a coding complete 3392 amino acid polyprotein sequence and recovery of 94 and 379nt of the 5′ and 3′ UTRs, respectively. BLASTn of the recovered DENV1 contig against deposited strains revealed the virus was most closely related in nucleotide identity (99.92%) to an unpublished DENV1 GZ8H/2019002/2018 human isolate (GenbankID: MN869904) collected on the 30th November 2018 China and 99.72% identity with the DV1I-TM19-74 isolate identified in Bangkok, Thailand in 2019. Phylogenetic inference of the imported case with other closely related DENV1 sequences revealed that the imported strain clustered with major lineage K, minor lineage 1 (DENV1I_K.1) isolates from Southeast Asia (Figure 1), with most closely related strains from Thailand, China and Cambodia [40]. It is noted that many of the closely related DENV1 sequences have metadata that indicates they are also imported cases into China from Thailand [41].
Assembly and Phylogenetic Position of Imported MRV Genome
Sera samples from patient SA395 were propagated in suckling mice for three passages and three times in VeroE6 cells and subjected to unbiased deep sequencing for the second and third VeroE6 passages. De novo assembly of the combined sequencing data revealed an almost complete MRV3 virus with coding complete segments of 9 segments recovered (Table 1) and a 3197/3929 (81%) of the L2 segment recovered (Figure 2A). We recovered complete sequences, including conserved terminal sequences 5′-GCUA and UCAUC-3′ for M2/3 and S3/4 segments. BLASTn sequence analysis of the assembled segments with their closest relative cross-referenced with the MRV serotype that the closely related segment has been identified from suggests that Human/SA395/SA/2017 MRV-3 may have arisen through multiple reassortment events of MRV-3s from varied origins but mostly closely related to bat, porcine and human MRV isolates.
MRVs are divided into three genotypes (MRV1-3), and genotyping is determined by segment S1. Our S1 tree (Figure 2) recapitulated the overall topology of other S1 strain phylogenies [18]. Based on S1 phylogeny, the Human/SA395/SA/2017 MRV-3 strain belongs to an unassigned lineage of the MRV3 bearing the highest nucleotide 93.64% identity with the human Tahiti MRV3 strain (GenbankID: L37679) isolated from a child in Tahiti, French Polynesia in 1960 [42] (Table 2). Given the close genetic relationship of SA397 and the Tahiti strain isolated in 1960, it’s also likely that a 4th genotype of MRV-3 (3d) has been circulating in Southeast Asia and Oceania with limited genomic detection.
4. Discussion
In this study, we characterized the importation of two exotic viruses, DENV1 and MRV, into South Africa from a family recently returned from Thailand. Through metagenomics and phylogenetics, we identified the DENV1 strain as originating from a common Southeast Asian lineage DENV1I_K.1. The MRV strain was identified as a novel reassorted MRV-3c, with segments closely related to porcine, human and bat segments suggesting this, and its S1 segment most closely related to a human isolate from Tahiti, French Polynesia from the 1960s. Investigating imported viruses, such as DENV and MRV, is crucial in understanding their origins, circulation, and potential for disease outbreaks. Such studies provide valuable insights into viral spread and evolution dynamics.
In 2019, Southeast Asia experienced a severe dengue outbreak, highlighting the persistent public health challenge posed by dengue fever in the region. Cambodia, for instance, reported over 10,000 cases annually, with more than 20,000 cases by July 2019 [48]. While DENV is not currently circulating in South Africa, several species of mosquitoes, including Ae. aegypti, are capable of vectoring DENV1 [49]. However, recent data on the vector competence of local mosquito populations for DENV1 are lacking. Only one study has indicated that South African colonies of Ae. aegypti could potentially support DENV1/2 and yellow fever transmission, with the Durban population having the greatest vectorial capability [50].
In contrast to DENV1, the origin and circulation of MRVs are less well understood due to limited genetic epidemiology. Our analysis revealed high divergence in the L1 segment of the South African MRV strain compared to other MRV isolates in GenBank (88.57% similarity). The L1 segment was most closely related to a porcine MRV while other segments were closely related to bat and human segments This suggests long-term circulation of MRV in Southeast Asia and Oceania, given the limited sampling and identity of MRV isolates from these regions.
Although the MRV was likely acquired in Thailand, the lack of comprehensive genetic surveillance data prevents us from confirming this with certainty. The possibility of local acquisition of MRV in South Africa cannot be ruled out. It is unclear what the potential route of virus transmission was to the patient, as the focus of the epi information collected was on potential mosquito transmission, and all patients reported mosquito bites. No further information was collected at the time that could be used to elucidate potential routes of transmission. This uncertainty underscores the need for further research into the virus’s ecology, epidemiology, and evolution, emphasizing the necessity of comprehensive genetic surveillance to understand MRV’s geographic distribution.
This study also highlights the capacity of MRVs to circulate silently in human and animal populations. It contributes to the expanding database on MRV’s geographic distribution and molecular diversity.
Continued surveillance of returned travelers using metagenomic approaches is critical for the early identification of exotic viruses. Metagenomics allows for the comprehensive detection of known and novel pathogens, providing a powerful tool for monitoring the importation of viruses into non-endemic regions. Integrating metagenomic surveillance with traditional serological methods can enhance our ability to respond promptly to emerging infectious diseases, thereby mitigating their potential impact on public health.
Author Contributions
Conceptualization, P.J.V.V, R.H.P. and J.T.P.; methodology, P.J.V.V and R.H.P.; formal analysis, P.J.V.V and R.H.P; investigation, P.J.V.V, R.H.P. and J.T.P.; resources, P.J.V.V, R.H.P. and J.T.P.; data curation, P.J.V.V and R.H.P; writing—original draft preparation, R.H.P.; writing—review and editing, P.J.V.V, R.H.P. and J.T.P.; visualization, R.H.P.; supervision, J.T.P.; funding acquisition, J.T.P All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Institute for Communicable Diseases, NICD-NHLS through its strategic funding to conduct disease surveillance.
Institutional Review Board Statement
Patient samples were tested as part of the Arbovirus Reference Laboratory’s routine diagnostic and surveillance efforts as part of its reference laboratory mandate. Communicable disease surveillance and outbreak investigation activities of the National Institute for Communicable Diseases (Johannesburg, South Africa) are approved by the Human Research Ethics Committee of the University of the Witwatersrand, Johannesburg, South Africa (M160667). Inoculation of suckling mice is covered under NICD Animal Ethics Committee Clearance number AEC 126-11.
Informed Consent Statement
Patient consent was inferred due to routine diagnostic and surveillance investigation.
Data Availability Statement
Nucleotide sequences determined in this study have been deposited in GenBank under the accession numbers MRV: PP953510-PP953519 and DENV1 PQ097697.
Acknowledgments
The Australian Galaxy service (https://usegalaxy.org.au/) provided compute resources. The National Institute for Communicable Diseases, NICD-NHLS supported this study through its strategic funding to conduct disease surveillance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; Myers, M.F.; George, D.B.; Jaenisch, T.; Wint, G.R.; Simmons, C.P.; Scott, T.W.; Farrar, J.J.; Hay, S.I. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhan, J.; Chen, L.; Chen, H.; Cheng, S. Global, regional, and national dengue burden from 1990 to 2017: A systematic analysis based on the global burden of disease study 2017. EClinicalMedicine 2021, 32, 100712. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Saito, M.; Mapua, C.A.; Natividad, F.F. Dengue illness: clinical features and pathogenesis. J Infect Chemother 2007, 13, 125–133. [Google Scholar] [CrossRef]
- Low, J.G.; Ong, A.; Tan, L.K.; Chaterji, S.; Chow, A.; Lim, W.Y.; Lee, K.W.; Chua, R.; Chua, C.R.; Tan, S.W.; Cheung, Y.B.; Hibberd, M.L.; Vasudevan, S.G.; Ng, L.C.; Leo, Y.S.; Ooi, E.E. The early clinical features of dengue in adults: challenges for early clinical diagnosis. PLoS Negl Trop Dis 2011, 5, e1191. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; Hendrickx, G.; Schaffner, F.; Elyazar, I.R.; Teng, H.J.; Brady, O.J.; Messina, J.P.; Pigott, D.M.; Scott, T.W.; Smith, D.L.; Wint, G.R.; Golding, N.; Hay, S.I. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Msimang, V.; Kemp, A.; Jansen van Vuren, P.; Weyer, J.; Paweska, J.T. Increased importation of dengue cases into South Africa: a risk for establishment of local endemicity? NICD The National Institute For Communicable Diseases: 01-04-2018, 2018. [Google Scholar]
- Matthijnssens, J.; Attoui, H.; Banyai, K.; Brussaard, C.P.D.; Danthi, P.; Del Vas, M.; Dermody, T.S.; Duncan, R.; Fang, Q.; Johne, R.; Mertens, P.P.C.; Mohd Jaafar, F.; Patton, J.T.; Sasaya, T.; Suzuki, N.; Wei, T. ICTV Virus Taxonomy Profile: Spinareoviridae 2022. J Gen Virol 2022, 103, 11. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Biagini, P.; Stirling, J.; Mertens, P.P.; Cantaloube, J.F.; Meyer, A.; de Micco, P.; de Lamballerie, X. Sequence characterization of Ndelle virus genome segments 1, 5, 7, 8, and 10: evidence for reassignment to the genus Orthoreovirus, family Reoviridae. Biochem Biophys Res Commun 2001, 287, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Ooms, L.S.; Kobayashi, T.; Dermody, T.S.; Chappell, J.D. A post-entry step in the mammalian orthoreovirus replication cycle is a determinant of cell tropism. J Biol Chem 2010, 285, 41604–41613. [Google Scholar] [CrossRef]
- Ichikawa, A.; Katayama, M.; Lai, H.; Wataru, S.; Takenaka-Uema, A.; Horimoto, T.; Murakami, S. Isolation and genetic characterization of a mammalian orthoreovirus from Vespertilio sinensis in Japan. Arch Virol 2023, 168, 165. [Google Scholar] [CrossRef]
- Lim, M.C.; Wang, Y.F.; Huang, S.W.; Yang, J.Y.; Wang, J.R. High Incidence of Mammalian Orthoreovirus Identified by Environmental Surveillance in Taiwan. PLoS One 2015, 10, e0142745. [Google Scholar] [CrossRef]
- Sedmak, G.; Bina, D.; Macdonald, J.; Couillard, L. Nine-year study of the occurrence of culturable viruses in source water for two drinking water treatment plants and the influent and effluent of a Wastewater Treatment Plant in Milwaukee, Wisconsin (August 1994 through July 2003). Appl Environ Microbiol 2005, 71, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Jansen van Vuren, P.; Wiley, M.; Palacios, G.; Storm, N.; McCulloch, S.; Markotter, W.; Birkhead, M.; Kemp, A.; Paweska, J.T. Isolation of a Novel Fusogenic Orthoreovirus from Eucampsipoda africana Bat Flies in South Africa. Viruses 2016, 8, 65. [Google Scholar] [CrossRef]
- Chua, K.B.; Voon, K.; Crameri, G.; Tan, H.S.; Rosli, J.; McEachern, J.A.; Suluraju, S.; Yu, M.; Wang, L.F. Identification and characterization of a new orthoreovirus from patients with acute respiratory infections. PLoS One 2008, 3, e3803. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Voon, K.; Yu, M.; Keniscope, C.; Abdul Rasid, K.; Wang, L.F. Investigation of a potential zoonotic transmission of orthoreovirus associated with acute influenza-like illness in an adult patient. PLoS One 2011, 6, e25434. [Google Scholar] [CrossRef]
- Giordano, M.O.; Martinez, L.C.; Isa, M.B.; Ferreyra, L.J.; Canna, F.; Pavan, J.V.; Paez, M.; Notario, R.; Nates, S.V. Twenty year study of the occurrence of reovirus infection in hospitalized children with acute gastroenteritis in Argentina. Pediatr Infect Dis J 2002, 21, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Mikuletic, T.; Steyer, A.; Kotar, T.; Zorec, T.M.; Poljak, M. A novel reassortant mammalian orthoreovirus with a divergent S1 genome segment identified in a traveler with diarrhea. Infect Genet Evol 2019, 73, 378–383. [Google Scholar] [CrossRef]
- Rosa, U.A.; Ribeiro, G.O.; Villanova, F.; Luchs, A.; Milagres, F.A.P.; Komninakis, S.V.; Tahmasebi, R.; Lobato, M.; Brustulin, R.; Chagas, R.T.D.; Abrao, M.; Soares, C.; Tinker, R.J.; Pandey, R.P.; Raj, V.S.; Sabino, E.C.; Deng, X.; Delwart, E.; Costa, A.C.D.; Leal, E. First identification of mammalian orthoreovirus type 3 by gut virome analysis in diarrheic child in Brazil. Sci Rep 2019, 9, 18599. [Google Scholar] [CrossRef] [PubMed]
- Steyer, A.; Gutierrez-Aguire, I.; Kolenc, M.; Koren, S.; Kutnjak, D.; Pokorn, M.; Poljsak-Prijatelj, M.; Racki, N.; Ravnikar, M.; Sagadin, M.; Fratnik Steyer, A.; Toplak, N. High similarity of novel orthoreovirus detected in a child hospitalized with acute gastroenteritis to mammalian orthoreoviruses found in bats in Europe. J Clin Microbiol 2013, 51, 3818–3825. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, D.W.; Capaul, R.; Prader, S.; Zagordi, O.; Geissberger, F.D.; Kugler, M.; Knorr, M.; Berger, C.; Gungor, T.; Reichenbach, J.; Shah, C.; Boni, J.; Zbinden, A.; Trkola, A.; Pachlopnik Schmid, J.; Huber, M. Persistent mammalian orthoreovirus, coxsackievirus and adenovirus co-infection in a child with a primary immunodeficiency detected by metagenomic sequencing: a case report. BMC Infect Dis 2018, 18, 33. [Google Scholar] [CrossRef] [PubMed]
- Ouattara, L.A.; Barin, F.; Barthez, M.A.; Bonnaud, B.; Roingeard, P.; Goudeau, A.; Castelnau, P.; Vernet, G.; Paranhos-Baccala, G.; Komurian-Pradel, F. Novel human reovirus isolated from children with acute necrotizing encephalopathy. Emerg Infect Dis 2011, 17, 1436–1444. [Google Scholar] [CrossRef]
- Tyler, K.L.; Barton, E.S.; Ibach, M.L.; Robinson, C.; Campbell, J.A.; O’Donnell, S.M.; Valyi-Nagy, T.; Clarke, P.; Wetzel, J.D.; Dermody, T.S. Isolation and molecular characterization of a novel type 3 reovirus from a child with meningitis. J Infect Dis 2004, 189, 1664–1675. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Beato, M.S.; Cavicchio, L.; Lavazza, A.; Chiapponi, C.; Leopardi, S.; Baioni, L.; De Benedictis, P.; Moreno, A. First identification of mammalian orthoreovirus type 3 in diarrheic pigs in Europe. Virol J 2016, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Naglic, T.; Rihtaric, D.; Hostnik, P.; Toplak, N.; Koren, S.; Kuhar, U.; Jamnikar-Ciglenecki, U.; Kutnjak, D.; Steyer, A. Identification of novel reassortant mammalian orthoreoviruses from bats in Slovenia. BMC Vet Res 2018, 14, 264. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Lesnik, R.; Brinkmann, A.; Ebinger, A.; Radonic, A.; Nitsche, A.; Muhldorfer, K.; Wibbelt, G.; Kurth, A. Isolation and characterization of three mammalian orthoreoviruses from European bats. PLoS One 2012, 7, e43106. [Google Scholar] [CrossRef] [PubMed]
- Colombino, E.; Lelli, D.; Canziani, S.; Quaranta, G.; Guidetti, C.; Leopardi, S.; Robetto, S.; De Benedictis, P.; Orusa, R.; Mauthe von Degerfeld, M.; Capucchio, M.T. Main causes of death of free-ranging bats in Turin province (North-Western Italy): gross and histological findings and emergent virus surveillance. BMC Vet Res 2023, 19, 200. [Google Scholar] [CrossRef]
- Qin, P.; Li, H.; Wang, J.W.; Wang, B.; Xie, R.H.; Xu, H.; Zhao, L.Y.; Li, L.; Pan, Y.; Song, Y.; Huang, Y.W. Genetic and pathogenic characterization of a novel reassortant mammalian orthoreovirus 3 (MRV3) from a diarrheic piglet and seroepidemiological survey of MRV3 in diarrheic pigs from east China. Vet Microbiol 2017, 208, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Thimmasandra Narayanappa, A.; Sooryanarain, H.; Deventhiran, J.; Cao, D.; Ammayappan Venkatachalam, B.; Kambiranda, D.; LeRoith, T.; Heffron, C.L.; Lindstrom, N.; Hall, K.; Jobst, P.; Sexton, C.; Meng, X.J.; Elankumaran, S. A novel pathogenic Mammalian orthoreovirus from diarrheic pigs and Swine blood meal in the United States. mBio 2015, 6, e00593–15. [Google Scholar] [CrossRef] [PubMed]
- Beaty, B.J.; Calisher, C.H.; Shope, R.E. Arboviruses. In: Diagnostic procedures for viral, rickettsial, and chlamydial infections, 7th ed. Washington (DC): American Public Health Association; 1995. p. 189–212.
- Jansen van Vuren, P.; Parry, R.; Khromykh, A.A.; Paweska, J.T. A 1958 Isolate of Kedougou Virus (KEDV) from Ndumu, South Africa, Expands the Geographic and Temporal Range of KEDV in Africa. Viruses 2021, 13, 7. [Google Scholar] [CrossRef]
- Djikeng, A.; Halpin, R.; Kuzmickas, R.; Depasse, J.; Feldblyum, J.; Sengamalay, N.; Afonso, C.; Zhang, X.; Anderson, N.G.; Ghedin, E.; Spiro, D.J. Viral genome sequencing by random priming methods. BMC Genomics 2008, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Parry, R.; James, M.E.; Asgari, S. Uncovering the Worldwide Diversity and Evolution of the Virome of the Mosquitoes Aedes aegypti and Aedes albopictus. Microorganisms 2021, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nature biotechnology 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Hill, V.; Cleemput, S.; Fonseca, V.; Tegally, H.; Brito, A.F.; Gifford, R.; Tran, V.T.; Kien, D.T.H.; Huynh, T.; Yacoub, S.; Dieng, I.; Ndiaye, M.; Balde, D.; Diagne, M.M.; Faye, O.; Salvato, R.; Wallau, G.L.; Gregianini, T.S.; Godinho, F.M.S.; Vogels, C.B.F.; Breban, M.I.; Leguia, M.; Jagtap, S.; Roy, R.; Hapuarachchi, C.; Mwanyika, G.; Giovanetti, M.; Alcantara, L.C.J.; Faria, N.R.; Carrington, C.V.F.; Hanley, K.A.; Holmes, E.C.; Dumon, W.; de Oliveira, T.; Grubaugh, N.D. A new lineage nomenclature to aid genomic surveillance of dengue virus. medRxiv 2024, 2024.05.16.24307504.
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.M.; Deforche, K.; de Oliveira, T. Genome Detective: an automated system for virus identification from high-throughput sequencing data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, V.; Libin, P.J.K.; Theys, K.; Faria, N.R.; Nunes, M.R.T.; Restovic, M.I.; Freire, M.; Giovanetti, M.; Cuypers, L.; Nowe, A.; Abecasis, A.; Deforche, K.; Santiago, G.A.; Siqueira, I.C.; San, E.J.; Machado, K.C.B.; Azevedo, V.; Filippis, A.M.B.; Cunha, R.V.D.; Pybus, O.G.; Vandamme, A.M.; Alcantara, L.C.J.; de Oliveira, T. A computational method for the identification of Dengue, Zika and Chikungunya virus species and genotypes. PLoS Negl Trop Dis 2019, 13, e0007231. [Google Scholar] [CrossRef] [PubMed]
- Siew, Z.Y.; Loh, A.; Segeran, S.; Leong, P.P.; Voon, K. Oncolytic Reoviruses: Can These Emerging Zoonotic Reoviruses Be Tamed and Utilized? DNA Cell Biol 2023, 42, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol Biol Evol 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Bohl, J.A.; Lay, S.; Chea, S.; Ahyong, V.; Parker, D.M.; Gallagher, S.; Fintzi, J.; Man, S.; Ponce, A.; Sreng, S.; Kong, D.; Oliveira, F.; Kalantar, K.; Tan, M.; Fahsbender, L.; Sheu, J.; Neff, N.; Detweiler, A.M.; Yek, C.; Ly, S.; Sath, R.; Huch, C.; Kry, H.; Leang, R.; Huy, R.; Lon, C.; Tato, C.M.; DeRisi, J.L.; Manning, J.E. Discovering disease-causing pathogens in resource-scarce Southeast Asia using a global metagenomic pathogen monitoring system. Proc Natl Acad Sci U S A 2022, 119, e2115285119. [Google Scholar] [CrossRef] [PubMed]
- Poltep, K.; Phadungsombat, J.; Nakayama, E.E.; Kosoltanapiwat, N.; Hanboonkunupakarn, B.; Wiriyarat, W.; Shioda, T.; Leaungwutiwong, P. Genetic Diversity of Dengue Virus in Clinical Specimens from Bangkok, Thailand, during 2018-2020: Co-Circulation of All Four Serotypes with Multiple Genotypes and/or Clades. Trop Med Infect Dis 2021, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Dermody, T.S.; Nibert, M.L.; Bassel-Duby, R.; Fields, B.N. Sequence diversity in S1 genes and S1 translation products of 11 serotype 3 reovirus strains. J Virol 1990, 64, 4842–4850. [Google Scholar] [CrossRef]
- He XiaoMing He, X.; Yao HuoChun Yao, H.; Zhang HongBiao Zhang, H.; Lin Tao Lin, T.; Yuan ShiShan Yuan, S.; Long JinXue Long, J.; Ding Chan Ding, C. Isolation and identification of a strain of porcine reovirus serotype 1 in China. 2013, 43, 22–29.
- Wang, L.; Fu, S.; Cao, L.; Lei, W.; Cao, Y.; Song, J.; Tang, Q.; Zhang, H.; Feng, Y.; Yang, W.; Liang, G. Isolation and identification of a natural reassortant mammalian orthoreovirus from least horseshoe bat in China. PLoS One 2015, 10, e0118598. [Google Scholar] [CrossRef] [PubMed]
- Lo, V.T.; Yoon, S.W.; Noh, J.Y.; Jang, S.S.; Na, W.; Song, D.; Jeong, D.G.; Kim, H.K. Characterization of replication and variations in genome segments of a bat reovirus, BatMRV/B19-02, by RNA-seq in infected Vero-E6 cells. Arch Virol 2022, 167, 2133–2142. [Google Scholar] [CrossRef]
- Rosen, L.; Hovis, J.F.; Mastrota, F.M.; Bell, J.A.; Huebner, R.J. Observations on a newly recognized virus (Abney) of the reovirus family. Am J Hyg 1960, 71, 258–265. [Google Scholar] [PubMed]
- Hrdy, D.B.; Rosen, L.; Fields, B.N. Polymorphism of the migration of double-stranded RNA genome segments of reovirus isolates from humans, cattle, and mice. J Virol 1979, 31, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.; Wang, J.; Wang, S.; Wu, J.; Zhou, Y.; Wang, X.; Luo, F.; Tu, X.; Chen, Q.; Huang, Y.; Ju, W.; Peng, X.; Rao, J.; Wang, L.; Jiang, N.; Ai, J.; Zhang, W. Emergence and Autochthonous Transmission of Dengue Virus Type I in a Low-Epidemic Region in Southeast China. Front Cell Infect Microbiol 2021, 11, 638785. [Google Scholar] [CrossRef] [PubMed]
- Kemp, A.; Jupp, P.G. Potential for dengue in South Africa: mosquito ecology with particular reference to Aedes aegypti. J Am Mosq Control Assoc 1991, 7, 574–583. [Google Scholar] [PubMed]
- Jupp, P.G.; Kemp, A. The potential for dengue in South Africa: vector competence tests with dengue 1 and 2 viruses and 6 mosquito species. Trans R Soc Trop Med Hyg 1993, 87, 639–643. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of imported DENV1 SA strain in 2017. Maximum-likelihood (ML) tree was constructed based on aligned DENV1 strains with nucleotide substitution model: TIM2 + F + I + G4 with 1000 ultrafast bootstrap replicates. DENV1 are indicated as the 6 major genotypes (I-VI) highlighted in different colours, with major and minor lineages if assigned on the label. Sequences accession number, country, and reported year are indicated, and the imported South African case is shown with arrowhead. Bootstrap support values are shown on nodes exceeding 75. Tree is midpoint rooted. Branch length corresponds to nucleotide substitutions per site.
Figure 1.
Phylogenetic analysis of imported DENV1 SA strain in 2017. Maximum-likelihood (ML) tree was constructed based on aligned DENV1 strains with nucleotide substitution model: TIM2 + F + I + G4 with 1000 ultrafast bootstrap replicates. DENV1 are indicated as the 6 major genotypes (I-VI) highlighted in different colours, with major and minor lineages if assigned on the label. Sequences accession number, country, and reported year are indicated, and the imported South African case is shown with arrowhead. Bootstrap support values are shown on nodes exceeding 75. Tree is midpoint rooted. Branch length corresponds to nucleotide substitutions per site.

Figure 2.
Genome organisation, coverage and phylogenetic inference of the S1 segment of the South African Human/SA395/SA/2017 MRV-3 strain. A) Coverage and genome organisation of the 10 MOV3 segments B) Consensus maximum likelihood phylogeny GTR+F+G4 model with a bootstrap of 1000 replicates. Accession number, host species, country and year are indicated for each strain. MRV-3 lineages are indicated in lowercase a,b and c, and MRV-2 and MRV-1 serotypes are indicated. South African Human/SA395/SA/2017 MRV-3 strain is indicated with an arrowhead. Bootstrap support values are shown on nodes exceeding 75. Tree is midpoint rooted. Branch length corresponds to nucleotide substitutions per site.
Figure 2.
Genome organisation, coverage and phylogenetic inference of the S1 segment of the South African Human/SA395/SA/2017 MRV-3 strain. A) Coverage and genome organisation of the 10 MOV3 segments B) Consensus maximum likelihood phylogeny GTR+F+G4 model with a bootstrap of 1000 replicates. Accession number, host species, country and year are indicated for each strain. MRV-3 lineages are indicated in lowercase a,b and c, and MRV-2 and MRV-1 serotypes are indicated. South African Human/SA395/SA/2017 MRV-3 strain is indicated with an arrowhead. Bootstrap support values are shown on nodes exceeding 75. Tree is midpoint rooted. Branch length corresponds to nucleotide substitutions per site.
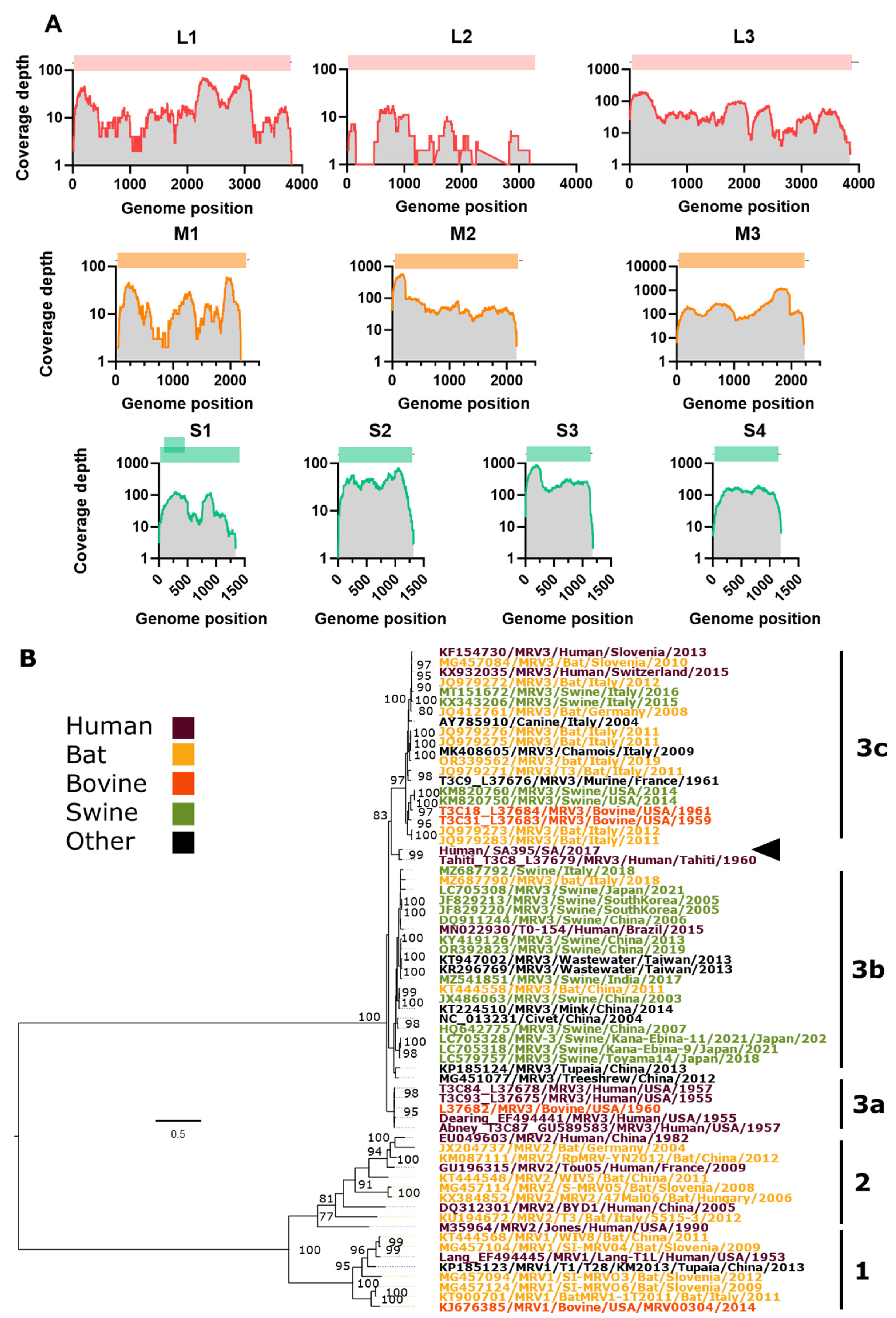

Table 1.
Clinical features of patients recently returned from Thailand.
| Date of blood collection | Case # | Age | Sex | Symptoms | Clinical features and complications |
|---|---|---|---|---|---|
| 17-July-2017 | SA395 | 46 | M | Onset (date not recorded): Fever (38.7°C) without rash, headache, severe body aches | No blood hematology test results were reported. The patients were not hospitalized. No previous history of dengue or yellow fever vaccination. All patients reported mosquito bites. |
| SA396 | 8 | F | Onset (date not recorded): Fever (38.5°C) without rash, severe body aches, nausea | ||
| SA397 | 45 | F | Onset (16/07/2017): Fever (39°C) without rash, severe body aches, nausea |
Table 2.
Highest nucleotide identities for each gene segment of the imported Human/SA395/SA/2017 MRV-3 strain.
Table 2.
Highest nucleotide identities for each gene segment of the imported Human/SA395/SA/2017 MRV-3 strain.
| Segment Protein |
Nucleotide (nt), Amino acid (aa) length |
Closest strain, GenbankID, (%) | MRV Serotype |
Location, Date | Host | Ref |
|---|---|---|---|---|---|---|
| L1 λ3 protein |
3822nt 1267aa |
SHR-A JX415466 88.57% |
MRV-1 | China, 2011 |
Pig | [43] |
| L2 λ2 protein |
2992/3918nt (76.3%) 993/1299aa (76.4%) |
AP-151 MN022938 93.99% |
MRV-3 | Brazil, 2015 |
Human | [18] |
| L3 λ1 protein |
3901nt 1275aa |
RpMRV-YN2012 KM087107 92.72% |
MRV-2 | Yunnan province, China, 2012 | Rhinolophus pusillus (bat) | [44] |
| M1 μ2 protein |
2295nt 736aa |
BatMRV/B19-02 MW582625 96.19% |
MRV-1 | Jeju Island, South Korea, 2019 | Miniopterus schreibersii (bat) | [45] |
| M2 μ1 protein |
2203nt 708aa |
Abney T3C87 GU589581 93.64% |
MRV-3 | Washington, D.C, USA, 1957 | Human | [46] |
| M3 μNS protein |
2241nt 721aa |
BatMRV/B19-02 MW582627 96.90% |
MRV-1 | Jeju Island, South Korea, 2019 | Miniopterus schreibersii (bat) | [45] |
| S1 σ1/σ1s protein |
1380nt σ1—455aa σ1s—120aa |
Tahiti L37679 93.57% |
MRV-3 | Tahiti, French Polynesia, 1960 | Human | [42] |
| S2 σ2 protein |
1323nt 427aa |
Lang L19774 98.11% |
MRV-1 | Ohio, USA, 1953 | Human | [47] |
| S3 σNS protein |
1198nt 366aa |
BatMRV/B19-02 MW582630 96.79% |
MRV-1 | Jeju Island, South Korea, 2019 | Miniopterus schreibersii (bat) | [45] |
| S4 σ3 protein |
1196nt 365aa |
SI-MRV07 MG999585 92.47% |
MRV-3 | Slovenia, 2017* | Human | [17] |
*Patient recently travelled from Thailand and Myanmar.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



