
Submitted:
31 July 2024
Posted:
01 August 2024
You are already at the latest version
Abstract
Apolipoprotein A (ApoA), an integral component of high-density lipoprotein (HDL) particles, plays a vital role in the transport and metabolism of lipids. It is implicated in both cardiovascular (CVD) and neurological diseases. In the context of CVD, ApoAs are pivotal in facilitating the reverse transport of cholesterol, mitigating inflammation, and preserving endothelial function. These char- acteristics highlight the crucial role of ApoA in averting the onset of CVDs. Moreover, ApoA has been implicated in the pathogenesis of several diseases, such as Alzheimer's disease (AD), Parkin- son's disease (PD), and multiple sclerosis (MS). Apart from its role in lipid metabolism, recent stud- ies have indicated the involvement of ApoA in neuroprotection, amyloid-beta clearance, and regu- lation of neuroinflammation. This paper addresses the research question: "How do different isoforms of Apolipoprotein A influence the development and progression of cardiovascular and neurological diseases, and what are the potential therapeutic implications of targeting ApoA in these conditions?" By providing a comprehensive review of current knowledge, this paper empha- sizes the importance of ApoA in understanding the intricate connections between cardiovascular and neurological health, suggesting that ApoA-targeted interventions may help treat diseases in both domains.
Keywords:
Apolipoproteins
; cardiovascular diseases
; neurological diseases
; atherosclerosis
; choles- terol
; Alzheimer's disease
1. Introduction
Apolipoprotein A (ApoA) serves as a versatile structural protein and an essential component of high-density lipoproteins (HDL) [1,2,3]. Its biosynthesis occurs predominantly in tissues such as the liver and the small intestines. ApoA plays a pivotal role in lipid metabolism by facilitating the transfer of excess cholesterol from peripheral tissues to the liver, thereby preventing cholesterol accumulation in arterial walls and neuronal tissues through reverse cholesterol transport [3,4,5]. Beyond lipid transport, ApoA has also been associated with several other biological processes, including inflammation, immune response, and antioxidant activity [6,7,8,9,10].
In humans, the isoforms of Apolipoprotein A (ApoA) —ApoA1, ApoA2, ApoA4, and ApoA5—are distinct protein products encoded by separate paralogous genes. These genes exhibit significant genetic diversity and variability. Notably, ApoA1, ApoA2, and ApoA5 share a common gene structure with three introns positioned at identical locations. However, ApoA4 differs in that it lacks its initial intron. This genetic arrangement suggests that the isoforms are not alternative splicing products but rather arise from distinct genes. These genetic differences have been linked to cardiovascular and neurological disease progression by altering ApoA expression and function [11,12,13,14,15,16].
Several studies suggest that specific ApoA isoforms may be associated with a heightened risk of cardiovascular disease (CVD) [11,12,13]. In particular, ApoA1M, a polymorphic variant of the APOA1 gene, is correlated with a higher risk of CVD, while ApoA1F, also a polymorphic variant of the APOA1 gene, is associated with a reduced risk of CVD. In addition to CVD, ApoA isoforms have also been implicated in the onset of neurodegenerative diseases like Alzheimer's disease (AD) [14,15,16,17,18]. ApoA1 has neuroprotective properties and may potentially inhibit the onset of AD [17].
Therefore, by mapping the multifaceted functions of ApoA, we may gain insights into the underlying mechanisms of these cardiovascular and neurological disorders, thereby identifying prospective therapeutic avenues. This review aims to discern the impact of different ApoA isoforms on cardiovascular and neurological diseases, guided by the research question: "How do different isoforms of Apolipoprotein A influence the development and progression of cardiovascular and neurological diseases, and what are the potential therapeutic implications of targeting ApoA in these conditions?"
2. Structure and Function of Apolipoprotein A (ApoA)
ApoA is one of the members of the apolipoprotein family, predominantly synthesized in the liver and small intestines. In addition to ApoA, other prominent members include ApoB, ApoC, and ApoE. These proteins collectively contribute to the regulation of lipid homeostasis and are crucial for normal cellular functions as well as implications in disease states. ApoA proteins are amphipathic, meaning they have both hydrophobic and hydrophilic regions. This structural feature is essential for their role in lipid transport. As shown in Table 1 ApoA has several isoforms that differ in their structure and function, contributing to a critical role in both normal and diseased cells. Therefore, the subsequent sections will discuss their roles in both contexts.
2.1. Structure and Function of Apolipoprotein A1 in Normal Cells
The most commonly identified isoform, ApoA1, is a protein predominantly associated with high-density lipoprotein (HDL), a major class of lipoproteins responsible for transporting lipids, including cholesterol and phospholipids, throughout the body. The ApoA1 gene, located on chromosome 11q23-q24, encodes the human ApoA1 protein with a mass of 28.1 kDa and precisely 243 amino acids [1,19]. ApoA1 has a helical structure with an N-terminal helix and a C-terminal helix, connected by a long, flexible loop region. The N-terminal helix is hydrophobic and interacts with lipids, whereas the C-terminal helix is hydrophilic and interacts with other proteins and lipids in the HDL particle [19]. ApoA1 plays a crucial role in lipid metabolism, particularly in promoting retrograde cholesterol transport.
In the process of retrograde cholesterol transport, as illustrated in Figure 1, ApoA1 plays a pivotal role. ApoA1 binds with the ATP-binding cassette transporter A1 (ABCA1) and acts as a cofactor for lecithin-cholesterol acyltransferase (LCAT). This journey begins with the synthesis of ApoA1 in the liver, where it rapidly gathers cholesterol and phospholipids from both the liver itself and peripheral cells, including macrophages, utilizing ABCA1. Moreover, macrophages efflux free cholesterol to HDL particles through the ATP-binding cassette (ABC) sub-family G member 1 (ABCG) transporter and the scavenger receptor class B type 1 (SR-BI). Once within discoidal HDL particles, LCAT activity transforms free cholesterol into cholesteryl ester, resulting in the maturation of spherical HDL particles. These mature HDL particles can efficiently transport cholesterol back to the liver, either directly through SR-BI or indirectly through the cholesteryl ester transfer protein (CETP). The liver can then uptake this cholesterol via the LDL receptor. Ultimately, hepatic cholesterol can take one of two routes: it can be excreted into the bile as either free cholesterol or bile acids, or it can be combined into lipoprotein particles and released back into the bloodstream [20]. This intricate process accentuates the critical role of ApoA1 in mediating plasma cholesterol transport and maintaining cellular cholesterol balance.
Within the structural framework of ApoA1, there is a distinct protein region known as the “Kringle”, consisting of 80 amino acids. This Kringle structure is not exclusive to ApoA1; it is also present in plasminogen, angiostatin, hepatocyte growth factor, prothrombin, urokinase, and tissue-type plasminogen activator, among others [21]. These proteins participate in a variety of vascular processes, including coagulation, blood clotting, fibrinolysis, and angiogenesis [21]. ApoA1 shares molecular similarities with the stabilizing factor for prostacyclin (PGI2), indicating that it may possess anti-clotting properties [1]. The alpha-helix structure of ApoA1 enables it to bind to unstable PGI2 molecules and stabilize them, resulting in an essential protective anti-aggregatory activity against platelet thrombus formation at sites of vascular injury [1]. This antiatherogenic function may prevent coronary artery disease. The structural flexibility and diverse functions of ApoA1 emphasize its critical role in preserving vascular health.
The therapeutic potential of apolipoprotein A-I (ApoA-I) in cardiovascular diseases has been explored through various approaches, including IV reconstituted ApoA1 infusion, mimetic peptides, and the stimulation of ApoA-I transcription. [22] First approach involves the use of reconstituted high-density lipoprotein (rHDL). Clinical trials have shown that intravenous administration of liposomic complexes containing human proapolipoprotein A-I in patients with familial hypercholesterolemia increased faecal excretion of cholesterol and biliary acids by over 30%. Subsequent trials with purified human apoA-I and synthetic phospholipids, indicated a 3.4% reduction in atherosclerotic plaque volume in acute coronary syndrome patients. However, development was halted due to liver toxicity concerns. Subsequent second-generation rHDL products emerged and they are currently under evaluation.
Another promising avenue involves apoA-I mimetic peptides, which are designed to replicate the antioxidant and cholesterol-binding functions of ApoA-I. Notably, such peptides have shown that they bind oxidized phospholipids more effectively than ApoA-I itself and reduce the HDL inflammatory index upon oral administration. However, the variant which requires intravenous or subcutaneous administration due to degradation in the intestine, did not yield significant improvements in HDL inflammatory index and even showed a trend towards elevated C-reactive protein levels in trials.
Additionally, stimulating endogenous ApoA-I synthesis has been investigated, as seen in the ASSERT study, where an oral inducer was tested over 12 weeks in 225 patients. While ApoA-I changes were not statistically significant, there was a trend toward increased HDL-C concentrations, suggesting that chronic treatment might be necessary for substantial effects. These strategies underscore the potential of ApoA-I in cardiovascular therapy, although challenges remain to be addressed.
2.2. Structure and Function of Apolipoprotein A2 (ApoA2) in Normal Cells
Apolipoprotein A2 (ApoA2) is another important protein associated with high-density lipoprotein (HDL) in the context of lipid metabolism and cardiovascular health. This protein is encoded by the ApoA2 gene located in chromosome 1q23.3. When combined with apolipoprotein D, ApoA2 can exist as a monomer, homodimer, or hybrid, and these forms can be detected in plasma. One of its key roles is the regulation of triglyceride degradation, at least in part, via HDL proteome regulation of lipoprotein lipase activity [23]. Genetic abnormalities within this gene can potentially lead to ApoA2 insufficiency or hypercholesterolemia [23]. The diverse structural configurations of ApoA2, and its pivotal role in regulating triglyceride degradation highlight its crucial contribution to lipid metabolism and the maintenance of optimal cholesterol levels, underscoring its significance in cardiovascular health.
2.3. Structure and Function of Apolipoprotein A4 (ApoA4) in Normal Cells
Human ApoA4, the largest apolipoprotein with a molecular weight of 46 kDa, is a significant component of postprandial lymph and plasma chylomicrons [23]. It is predominantly linked to triglyceride-rich lipoproteins, including chylomicrons and very low-density lipoproteins (VLDL) [24]. The majority of ApoA4 synthesis occurs in the jejunum, with additional production in the duodenum and ileum. It is additionally identified in the hypothalamus and the solitary tract of the brainstem, along with dendritic cells of the immune system [23]. Structurally, ApoA4 shares some similarities with the N-terminal structure of ApoA1, including a putative amphipathic α-helical repeat that is crucial for lipid binding and self-association. However, the helical bundles exhibit noticeable structural differences. ApoA4 is composed of 22 amino acid peptide repeats that can form amphipathic helices [25]. In the absence of lipids, ApoA4 self-assembles into homodimers. Each monomeric ApoA4 protein has a well-organized center core and less organized N- and C-termini. Helix-A (aa 74–94), helix-B (aa 96–204), helix-C (aa 206–255), and helix-D (aa 265–312) are the four 4-helix bundles forming the center core helices (amino acids 64–335) [25]. Many different cellular processes, such as fat absorption and chylomicron assembly, have been attributed to ApoA4. As a major HDL constituent, ApoA4 has been implicated in reverse cholesterol transfer, satiety, antioxidation, regulation of gut inflammation and most recently, the clearance of B-amyloid in the brain. ApoA4 promotes the transport and metabolism of dietary LYlipase (LPL), an enzyme responsible for the breakdown of triglycerides, resulting in the release of free fatty acids and their subsequent absorption by tissues [26]. This process contributes to the maintenance of lipid homeostasis and the regulation of plasma triglyceride levels. Therefore, any alteration in ApoA4 can have a major impact on antiatherogenic capabilities and trigger the development of cardiovascular disease or neurodegenerative diseases.
2.4. Structure and Function of Apolipoprotein A5 (ApoA5) in Normal Cells
Apolipoprotein A5 (ApoA5) is encoded by the ApoA5 gene, yielding a protein with 366 amino acid residues, with the mature ApoA5 comprising 343 amino acids. Structurally, ApoA5 is characterized as a hydrophobic protein predominantly composed of amphipathic α-helix secondary structures, rendering it insoluble in aqueous solutions at neutral pH. Notably, the N-terminal residues 1–146 form a helix bundle, displaying water solubility in the absence of lipids, whereas the full-length protein is water-insoluble. ApoA5 interacts with lipoproteins, particularly exhibiting a preference for high-density lipoprotein (HDL) over very low-density lipoprotein (VLDL). The protein achieves solubility in the presence of lipids, primarily due to the lipid-binding properties of the C-terminal end (residues 293–343). Functionally, ApoA5 plays a critical role in lipid metabolism, with specific residues, particularly within the region 192–238, identified as crucial for lipid binding and activation of lipoprotein lipase (LPL). Additionally, ApoA5 interacts with the LPL-anchoring protein GPIHBP1 (glycosylphosphatidylinositol-anchored high-density lipoprotein binding protein 1), involving positively charged residues (R210, K211, K215, K217) in the heparin-binding domain [27]. These structural and functional attributes highlight ApoA5 significance in lipid metabolism and its interactions with key components, contributing to our understanding of its role in maintaining cardiovascular health.
The Role of Apolipoprotein A in Diseases
As shown in, Table 2 in addition to regulating normal human health, each ApoA can also induce disease when altered. Numerous studies indicate that ApoA isoforms play a major role in neurodegenerative and cardiovascular diseases; moreover, some studies have linked them to diabetes, renal failure, hypercholesterolemia, and autoimmune disorders. The subsequent sections will explore the effects of these isoforms on a variety of disorders.
3. The Role of Apolipoprotein A in Cardiovascular Diseases
Cardiovascular diseases (CVDs) continue to be a major global health concern, accounting for a considerable proportion of morbidity and mortality [37]. Dyslipidemia is a significant risk factor for developing and progressing CVDs, marked by abnormal lipid levels, particularly low-density lipoprotein cholesterol (LDL-C) [38].
3.1. Apolipoprotein A1 in Coronary Artery Disease (CAD)
CAD, which is influenced by lipid metabolism disorders and genetic susceptibility, is mitigated by the protective effect of ApoA1. The alpha-helix structure of ApoA1 facilitates binding with prostaglandin I2 (PGI2), thereby extending the half-life of PGI2 through HDL and nascent HDL reconstituted from ApoA1 and phospholipids. This stabilization of PGI2 prevents platelet thrombi accumulation at vascular damage sites. The protective impact of HDL against coronary artery disease may be partially attributed to this mechanism [29].
Mutations or disruptions in the ApoA1 gene can lead to significant deficiencies in plasma HDL levels, similar to deficiencies seen in patients with genetic deficiencies in ABCA1 or LCAT. These deficiencies are associated with an increased risk of CAD [28]. Current research indicates that HDL and its key structural component, ApoA1, found in human atheroma, undergo dysfunctional and considerable oxidation due to myeloperoxidase (MPO), a pro-inflammatory enzyme. This oxidative modification impairs ABCA1 macrophage transport, activates inflammatory pathways, and increases CAD risk. Additionally, the cholesterol-accepting capability of ApoA1/HDL is compromised by MPO-induced oxidation [30,31].
Genetic studies have identified the ApoA1-75 A allele variant as linked to a reduced risk of CAD due to elevated serum ApoA1 and HDL-C levels [32]. Conversely, the ApoA-I E136X variant is associated with HDL-C deficiency and early onset of CAD [33]. The therapeutic implication lies in exploring strategies to enhance ApoA1's protective effects. Elevating serum ApoA1 and HDL-C levels through pharmacological approaches or lifestyle modifications could be a viable approach to reduce CAD risk. Additionally, understanding and addressing genetic variations in ApoA1 may provide targeted interventions to regulate lipid metabolism and decrease susceptibility to CAD.
3.2. Apolipoprotein A1 in Hypertension (HTN)
While research on the relationship between ApoA1 and HTN remains limited, a cross-sectional study conducted on the Chinese population in Wuhan aimed to explore this relationship. The study results indicated that higher ApoA1 levels in the blood might be linked to a lower chance of having hypertension among individuals with CAD, suggesting a potential protective role of ApoA1 against hypertension [34]. While more extensive investigations are required, the outcomes of this imply a potential protective association of higher ApoA1 levels with reduced hypertension prevalence in those with CAD, warranting further exploration.
3.3. Apolipoprotein A2
It is widely acknowledged that reduced HDL levels correlate with an increased risk of CVD. However, a comprehensive understanding of HDL metabolism is still lacking. Among the components of HDL, ApoA2 stands out as the second most prevalent apolipoprotein. A study designed to test the hypothesis that ApoA2 isoforms are inversely correlated with myocardial infarction concluded that none of the ApoA2 isoforms (ApoA2-ATQ/ATQ, ApoA2-ATQ/AT, and ApoA2-AT/AT) exhibited significant associations with myocardial infarction risk. Consequently, the precise role of ApoA2 in CVD remains ambiguous, particularly in terms of whether it functions as an antiatherogenic or proatherogenic protein [35].
Moreover, higher levels of apolipoprotein E (ApoE) are associated with the onset of cardiovascular disease in women with increased levels of C-reactive protein (CRP), regardless of whether they have low or high HDL levels. To determine if ApoA2 has an impact on this ApoE-related risk across these two groups, a study was undertaken. This research determined that ApoA2 significantly influences the relationship between ApoE and the likelihood of developing CVD in women with high levels of both HDL and CRP [36].
3.4. Apolipoprotein A4
ApoA4 plays a pivotal role in lipid metabolism and is associated with the onset of CVDs, though the precise mechanisms are still being investigated [39]. There is evidence suggesting that ApoA4 influences the development and progression of atherosclerosis. Research indicates that Antiatherogenic characteristics oversee various stages of atherosclerosis as illustrated in Figure 2. One significant function of ApoA4 is promoting reverse cholesterol transport, which enhances cholesterol efflux from peripheral tissues, including macrophage foam cells within plaques [40]. This process reduces plaque formation and helps regress existing plaques. Additionally, ApoA4 has anti-inflammatory properties, as it inhibits the expression of adhesion molecules and pro-inflammatory cytokines in endothelial cells, leading to decreased leukocyte recruitment and vascular inflammation [41]. Collectively, these attributes contribute to reducing atherosclerosis, ultimately reducing the likelihood of developing myocardial infarction (MI) and other CVDs. Researchers have observed that an excess of ApoA4 in the body hinders the progression of atherosclerosis, suggesting that elevated ApoA4 levels could serve as a protective barrier against the initiation and expansion of plaque in the blood vessels [42,43].
Moreover, Single Nucleotide Polymorphisms (SNPs) within the ApoA4 gene have been linked to altered lipid profiles and increased risk of CVD, including MI. Specific mutations or polymorphisms in the ApoA4 gene, such as the polymorphism (Gln360His), have been tied to elevated levels of low-density lipoprotein cholesterol (LDL-C) and triglycerides (TG), both recognized contributors to CVD risk. A meta-analysis specifically focused on the ApoA4 gene SNP (Gln360His) and its relation to CAD risk. The findings, drawn from an assessment of multiple studies, revealed that individuals carrying the HIS allele of the ApoA4 gene faced a higher risk of CAD compared to those with the Gln/Gln genotype [44].
Although ApoA4 has a protective effect against atherosclerosis and the development of MI and CVD, it has the opposite effect in diabetes. Glycated ApoA4 was found to induce atherogenesis in Type 2 diabetes patients with CAD [45]. Through the activation of the nuclear receptor NR4A3, gamma-ApoA4 (g-ApoA4) elicited proinflammatory responses in vitro and promoted the progression of atherosclerosis in ApoE-deficient (ApoE–/–) mice [45]. When compared to unaltered g-ApoA4, variants of g-ApoA4 with mutations (K-A) at sites frequently susceptible to glycation exhibited diminished proinflammatory and atherogenic effects in both in vivo and in vitro settings. As a result, the glycation of ApoA4 was correlated with the severity of CAD in individuals with Type 2 Diabetes Mellitus (T2DM).
Furthermore, increased levels of ApoA4 have been shown to contribute to an increased risk of developing cardiac amyloidosis [46]. Given its role in lipid metabolism and atherosclerosis, ApoA4 emerges as a promising therapeutic target for managing and preventing CVDs. Strategies aimed at boosting ApoA4 levels or improving its functional characteristics might contribute to a decreased risk of CVD. Mimetic peptides of ApoA4 that replicate its beneficial effects have been the subject of preclinical study. These peptides can increase reverse cholesterol transfer and modify lipid metabolism, consequently alleviating the impact of atherosclerosis [47]. In addition, pharmacological approaches focused on augmenting the endogenous expression of ApoA4 or enhancing its stability and efficacy might pave the way for novel CVD therapeutic avenues
3.5. Apolipoprotein A5
ApoA5 plays a key role in lipid metabolism, primarily by enhancing the activity of lipoprotein lipase (LPL) which in turn facilitates the rapid removal of TG-rich lipoproteins from circulation. Consequently, any dysregulations or genetic variations of ApoA5 have been associated with elevated plasma TG levels, recognized as a CVD risk factor [48].
The growth and progression of atherosclerosis are interlinked with ApoA5. Studies indicate that a deficiency or malfunction in ApoA5 deficiency or dysfunction correlates with a surge in atherosclerotic lesions in animal models [49]. Furthermore, VLDL, which are TG-rich lipoproteins, have been correlated with an increase in CVD risk. ApoA5, via its synergistic actions with LPL and hepatic lipase (HL), accelerates the breakdown of VLDL particles [50]. Owing to ApoA5's enhanced TG hydrolysis within VLDL, the result is the formation of smaller, less atherogenic lipoprotein particles.
Within the ApoA5, specific variants play a pivotal role in modulating plasma TG levels. Notably minor alleles of three single nucleotide polymorphisms (SNPs) -rs662799, rs3135506, and rs3135507 have been associated with impaired lipoprotein lipolysis, leading to raised VLDL levels, thus heightening an individual's susceptibility to CVDs [51]. Given these insights, interventions aimed at modifying ApoA5's expression or functionality present as potential therapeutic avenues to decrease TG-rich lipoprotein concentrations, subsequently mitigating CVD risk.
4. The Role of Apolipoprotein A1 in Neurological Diseases
ApoA1 has garnered increasing attention in the context of various neurological diseases including stroke, Alzheimer's disease (AD), Parkinson's disease (PD), cerebral amyloid angiopathy (CAA), general paresis, frontotemporal dementia (FTD), amyotrophic lateral sclerosis (ALS), multiple sclerosis (MS), and spinocerebellar degeneration (SCD).
4.1. Apolipoprotein A1 in Stroke
Stroke remains a leading global cause of mortality and disability. ApoA1's relevance to stroke lies in its profound impact on lipid homeostasis and its anti-inflammatory properties. ApoA1, through its ability to promote cholesterol removal from atherosclerotic plaques by interacting with ATP-binding cassette transporters like ABCA1, may play a crucial part in countering atherosclerosis, which in turn can diminish the risk of stroke. The neuroprotective fact of ApoA1 becomes particularly conspicuous when there is an alteration or reduction of its concentration. A lower ratio of apolipoprotein A1 to apolipoprotein B is associated with conditions like ischemic stroke and intracranial atherosclerotic disease. Studies suggest that diminished ApoA1 levels and elevated ApoB levels, coupled with a higher ApoB/A1 ratio, are risk determinants for first-time ischemic strokes. Intriguingly, while ApoA1 provides a shield against ischemic strokes, this protective scenario inverts hemorrhagic strokes. In this context, increased ApoA1 concentrations might indeed raise the probability of a first-time hemorrhagic stroke [52]. However, many studies rely on observational data, which can be influenced by confounding factors. For instance, genetic predispositions and lifestyle factors are not always adequately controlled, leading to potential biases. Additionally, there is variability in how ApoA1 levels are measured, which can affect the reliability of results. Future studies should adopt more rigorous methodologies to confirm these associations and elucidate underlying mechanisms.
4.2. Apolipoprotein A1 in Alzheimer's Disease (AD)
Alzheimer's disease (AD), characterized by the accumulation of cerebral neurd-beta (Aβ) and the formation of neurofibrillary tangles, is a crippling neurodegenerative condition. The potential involvement of ApoA1 in the clearance of Aβ from the brain, a pivotal process in the development of AD, has caught the attention of researchers. ApoA1 seems to facilitate Aβ transport across the blood-brain barrier, promoting its removal and possibly offering a protective mechanism against AD. A study led by Endres et al. observed a decline in ApoA1 levels in peripheral samples, such as serum or plasma, of AD patients. In contrast, an elevated concentration of ApoA1 was associated with a reduced risk of AD onset [53]. This study further highlighted a substantial positive link between heightened ApoA1 levels and the Mini-mental state examination (MMSE) score, an assessment tool gauging various cognitive functionalities including orientation, memory, attention, recall, and language skills [53]. Another investigation pinpointed a decrease in ApoA1 in the cerebrospinal fluid (CSF) of AD patients, which correlated with reduced cognitive performance and heightened AD disease severity [54].
While several studies report an inverse relationship between ApoA1 levels and AD risk, these findings are not universally consistent. The quality of evidence varies, with many studies lacking large sample sizes and long-term follow-up. Moreover, there is a need for standardization in the assessment of cognitive outcomes and ApoA1 measurements. Some studies use peripheral blood levels, while others focus on cerebrospinal fluid, leading to inconsistent findings. Future research should prioritize longitudinal studies with diverse populations to better understand the temporal relationship between ApoA1 and AD.
Given these insights, therapeutic avenues geared towards amplifying ApoA1-driven Aβ clearance could represent a promising direction for AD treatment. In a particular study, the intravenous administration of human recombinant ApoA1 Milano was shown to curtail beta-amyloid deposition in the brain, as demonstrated in the APP23-transgenic mouse model of AD [55].
4.3. Apolipoprotein A1 in Cerebral Amyloid Angiopathy (CAA)
CAA is defined by the accumulation of Aβ deposition within the walls of cerebral blood vessels, causing the vessel to become fragile and prone to hemorrhagic complications. The potential function of ApoA1 in aiding the removal of Aβ from the brain might also play a significant role in CAA. Studies have shown that a lack of ApoA1 exacerbates CAA and cognitive impairment. In contrast, overexpression of ApoA1, when driven by its native promoter in organs such as the liver and intestine, leads to a reduction in CAA, diminished neuroinflammation, and improved cognitive function [56]. Additionally, the absence of ApoA1 increases cortical amyloid deposition, cerebral amyloid angiopathy, as well as inflammation in areas like the cortical and hippocampal, especially in the APP/PS1 mouse model [56]. Consequently, therapeutic interventions aimed at enhancing APOA1-assisted Aβ clearance may offer a promising approach to mitigating the complications associated with CAA.
4.4. Apolipoprotein A1 in Parkinson's Disease (PD)
PD, primarily known as a movement disorder, also reveals interconnections with lipid metabolism in its development. ApoA1, with its intricate role in lipid transport, may exert an influence on alpha-synuclein aggregation, a hallmark of PD. It was observed that individuals with PD have lower plasma ApoA1 in plasma. Furthermore, in these patients, plasma ApoA1 concentrations show a correlation with their specific genotype at the ApoA1 promoter polymorphism, identified as rs670 [57]. Notably, a decrease in ApoA1 levels also indicated a more pronounced deficit in dopamine production in PD patients [58]. This information highlights the possible interplay between ApoA1 and the pathogenesis of PD, suggesting new avenues for research and potential therapeutic strategies.
4.5. Apolipoprotein A1 in General Paresis
General paresis, a neuropsychiatric manifestation of late-stage syphilis, results in cognitive impairment. The role of ApoA1 in this scenario is largely undetermined. However, considering ApoA1's known involvement in modulating immunity and decreasing inflammation, it is worthwhile to investigate any potential links. Notably, serum levels of ApoA1 were found to be lowered in patients suffering from general paresis. In contrast, individuals with syphilis but not showing signs of neurosyphilis had ApoA1 levels similar to those of healthy subjects [59]. A sustained decrease in ApoA1 concentration, coupled with a higher ApoB/ApoA-1 ratio, might be instrumental in the development of syphilitic dementia [59]. This suggests that monitoring and understanding lipid markers could offer insights into the progression and management of neurosyphilis and its complications.
4.6. Apolipoprotein A1 in Frontotemporal Dementia (FTD)
FTD, marked by degeneration of the frontotemporal lobes, results in changes in personality and language deficits. While ApoA1's main function is rooted in lipid metabolism, disruptions in lipid balance have been linked to neurodegenerative mechanisms, indirectly suggesting a potential role for ApoA1 in FTD. In particular, decreased plasma ApoA1 levels were identified in patients with a behavioral variant of FTD [60]. This underscores the possibility that lipid regulatory proteins, like ApoA1, may have broader implications in neurodegenerative conditions beyond their primary metabolic functions.
4.7. Apolipoprotein A1 in Amyotrophic Lateral Sclerosis (ALS)
ALS, a degenerative condition that targets motor neurons, might have ties to ApoA1 due to its involvement in lipid regulation and its anti-inflammatory attributes. Notably, elevated levels of HDL and ApoA1 in the bloodstream have been correlated with a decreased likelihood of ALS onset [61]. Delving deeper into how ApoA1 may either exacerbate or alleviate the mechanisms underlying ALS presents a crucial direction for future research. This highlights the potential of lipid modulators like ApoA1 in offering insights into the prevention and management of neurodegenerative diseases like ALS.
4.8. Apolipoprotein A1 in Multiple Sclerosis (MS)
MS, an autoimmune disorder, is marked by inflammation and demyelination within the central nervous system. The anti-inflammatory properties of ApoA1 could play a protective role by influencing the immune response and curbing neuroinflammation [62]. Research indicates that individuals with progressive MS have decreased plasma ApoA1 levels when compared to those with stable relapsing-remitting multiple sclerosis (RRMS) and to age-similar healthy controls [63]. This suggests that variations in ApoA1 levels might be significant in the disease progression and dynamics of MS, underlining the potential importance of this protein in understanding and managing the disease.
4.9. Apolipoprotein A1 in Spinocerebellar Degeneration (SCD)
SCD encompasses a group of hereditary disorders marked by progressive cerebellar dysfunction. While ApoA1 is predominantly associated with lipid metabolism, its auxiliary roles in neuronal well-being and inflammation might be pertinent to SCD. Research has indicated that diminished ApoA1 levels might adversely affect neuronal health, potentially playing a role in the progression of SCD [64]. This connection underscores the multifaceted influence of proteins like ApoA1 beyond their primary functions and emphasizes the importance of exploring their potential impacts on various neurological disorders.
5. Apolipoprotein A2 in Neurological Diseases
ApoA2 is primarily recognized for its role in lipid metabolism and cardiovascular health, but emerging research suggests its involvement in various neurological disorders including stroke, Alzheimer's disease (AD), frontotemporal dementia (FTD), and multiple sclerosis (MS).
5.1. Apolipoprotein A2 in Stroke
ApoA2, which is part of the complex tapestry of lipid metabolism, holds significance in the pathophysiology of stroke. ApoA2's regulatory influence on cholesterol transport and metabolism is central to vascular health. An imbalance, particularly elevated levels of ApoA2, is believed to intensify the risk for atherosclerosis, a condition where plaque builds up inside the arteries, which is a well-established precursor to stroke. Beyond its direct role in lipid metabolism, ApoA2's potential involvement in inflammatory pathways and its contributions to oxidative stress are of concern. These processes can escalate neuronal injuries, especially in the aftermath of ischemic strokes where blood flow to the brain is interrupted [65]. Recognizing the association between ApoA2 and stroke risk accentuates the multifaceted interplay between lipid regulation, inflammation, and cerebrovascular health.
5.2. Apolipoprotein A2 in Alzheimer's Disease (AD)
The presence of ApoA2 in Aβ plaques and its potential interaction with Aβ is intriguing. Aβ plaques are a hallmark pathological feature of Alzheimer's disease (AD), and molecules that interact with Aβ can influence its aggregation dynamics and thus the pathophysiology of AD. Several factors, such as antioxidant levels, inflammatory markers, and apolipoproteins, lead to considerable variability in the pace of cognitive decline among the elderly population. The study involving 440 elderly Japanese males provides valuable insights into the relationship between serum levels of apolipoproteins (ApoA1 and ApoA2) and cognitive function. Given that both ApoA1 and ApoA2 were found to be reduced in individuals with late onset nonfamilial AD, suggests that these apolipoproteins might play protective roles against cognitive decline [66].
While the exact mechanisms by which ApoA1 and ApoA2 influence cognitive function and AD progression remain to be fully elucidated, their association with cognitive impairment and AD underscores their potential importance in understanding the pathophysiology of neurodegenerative diseases. Furthermore, they might serve as potential therapeutic targets or biomarkers for AD and related disorders in the future.
5.3. Apolipoprotein A2 in Frontotemporal Dementia (FTD)
Behavioral variant frontotemporal dementia (bvFTD) stands as a prevalent contributor to early-onset dementia. The pathological hallmark of bvFTD involves disruptive lipid regulation that affects survival. A study revealed diminished levels of ApoA1 and ApoA2 within HDL among individuals with bvFTD when compared to control subjects. This study marks the first apolipoprotein assessment specific to bvFTD, indicating potential alterations in HDL function and an elevated risk of CVD in individuals affected by bvFTD [67].
5.4. Apolipoprotein A2 in Multiple Sclerosis (MS)
The link between cholesterol balance and nerve cell damage in multiple sclerosis (MS) has not been extensively studied. To understand this relationship better, a study was carried out comparing markers of nerve cell damage (like serum neurofilament light chain levels) in the cerebrospinal fluid (CSF) with cholesterol-related markers in MS. This study involved 133 early-stage MS patients. The results showed that lower serum levels of ApoA2 and ApoA1 were associated with increased nerve cell damage, as indicated by CSF neurofilament light chain levels. This finding hints at a possible protective function of ApoA2 in the progression of MS [68].
6. Apolipoprotein A4 in Neurological Diseases
ApoA4 may have a broader impact on the health and function of neurons in addition to its function in lipid metabolism and transport. Numerous studies have revealed that ApoA4 is present and regulated in several neurological diseases.
6.1. Apolipoprotein A4 in Stroke
The αIIbβ3 integrin found on platelets and its associated molecules are pivotal for thrombosis and maintaining homeostasis, and they play significant roles in cardiovascular and neurological events including myocardial infarction and stroke. A study revealed that the binding of ApoA4 to αIIbβ3 integrin relies on specific aspartic acid residues at the start of ApoA4 and is further influenced by interactions withinAApoA4's C-terminus. ApoA4 acts as a brake on platelet aggregation and excessive platelet activity that can occur postprandially. Remarkably, human ApoA4 levels in the bloodstream exhibit a circadian rhythm that inversely correlates with platelet aggregation and cardiovascular incidents. Consequently, ApoA4 has been identified as a newfound partner of αIIbβ3 integrin and serves as a natural inhibitor against blood clot formation [69].
6.2. Apolipoprotein A4 in Alzheimer's Disease:
Research is increasingly pointing towards a potential role of ApoA4 in the development and progression of Alzheimer's disease (AD). ApoA4 seems to influence the aggregation and clearance of amyloid-beta, a key player in AD pathology. Observations have shown decreased serum levels of ApoA4 in individuals diagnosed with AD, hinting at a potential relationship between ApoA4 concentration and the disease’s development [70]. While the exact mechanisms remain to be elucidated, ApoA4 may serve a neuroprotective function, safeguarding neurons from damage and staving off neurodegenerative changes.
6.3. Apolipoprotein A4 in Parkinson's Disease (PD)
PD is characterized by the loss of dopaminergic neurons in the substantia nigra and the accumulation of α -synuclein aggregates. While primarily considered a neurodegenerative disorder, recent studies propose that ApoA4 may participate in maintaining mitochondrial function and proteostasis [71]. Dysregulation in these processes is implicated in PD pathophysiology, hinting at a potential link between ApoA4 and PD. Interestingly, in one study ApoA4 was found to be predominantly detected in serum control groups but rare in PD patients, indicating dysregulation of ApoA4 in PD [71]. The link between ApoA4 and the potential modulation of Parkinson's disease (PD) progression is intriguing, especially given its suggested roles in mitochondrial functionality and proteostasis. If ApoA4 does indeed influence these cellular processes, it may have a bearing on PD onset and progression, since mitochondrial dysfunction and protein aggregation are hallmark features of the disease. Thus, understanding ApoA4's exact role in these processes could shed light on novel therapeutic avenues or preventative measures for PD.
6.4. Apolipoprotein A4 in Psychiatric Disorders:
Beyond neurodegenerative diseases, ApoA4 appears to have relevance in psychiatric disorders. The findings indicating diminished serum ApoA4 levels in patients with schizophrenia (SCZ) and depression offer intriguing insights into the potential involvement of ApoA4 in neuropsychiatric disorders. If ApoA4 indeed plays a role in the neurobiology and pathophysiology of these conditions, it highlights the broader spectrum of ApoA4's influence beyond its primary function in lipid metabolism [72,73,74].
The observed correlation between declining ApoA4 levels and the severity of depression underscores its potential utility as a biomarker. If consistent patterns emerge from subsequent studies, clinicians might leverage ApoA4 levels to diagnose, monitor, or even guide therapeutic strategies for depression. Furthermore, the association between ApoA4 individuals' emotional and behavioral responses to stress suggests a potentially deeper role for ApoA4 in mood regulation and stress resilience.
However, it is crucial to note that while these associations are compelling, they do not necessarily indicate causation. Further research is needed to delineate whether ApoA4 plays a direct role in these neuropsychiatric conditions or if its alterations are merely a consequence of other underlying processes.
6.5. Apolipoprotein A4 in Guillain-Barré Syndrome (GBS) and Chronic Inflammatory Demyelinating Polyneuropathy (CIDP)
The absence of changes in ApoA4 levels in the cerebrospinal fluid (CSF) of patients with Guillain-Barré syndrome (GBS) and chronic inflammatory demyelinating polyneuropathy (CIDP) provides important insights into the specificity ApoA4's involvement in various neurological conditions. While ApoA4 has been implicated in certain neurodegenerative and psychiatric disorders, its stability in the CSF of GBS and CIDP patients suggests that its potential influence is not universal across all neurological diseases [75].
This distinction emphasizes the importance of understanding the nuanced roles of biomolecules in different pathological settings. Just because a molecule is altered in one disease does not mean it will be affected in another, even if both diseases fall under the broader category of neurological disorder. It is essential to recognize that each neurological disease has its unique pathophysiology and molecular signature.
The findings further underscore the importance of targeted research in deciphering the specific roles and implications of molecules like ApoA4 in diverse medical conditions. It will be interesting to see how future research defines the limitApoA4's involvement in nervous system disorders.
7. The Role of Apolipoprotein A5 in Neurological Diseases
7.1. Apolipoprotein A5 in Stroke:
The ApoA1-C3-A5 gene cluster has been posited to be a pivotal player in the etiology of ischemic stroke, given its integral role in lipid metabolism and subsequent vascular implications. A study focusing on the northern Chinese Han population provided further insights into this hypothesis. This investigation specifically sought to elucidate any associations between the ApoA1-C3-A5 gene cluster and the incidence of ischemic stroke. Their findings revealed that three specific single nucleotide polymorphisms (SNPs) - namely rs670, rs2854116, and rs662799 - within this gene cluster indeed bear significant associations with the occurrence of ischemic stroke in the studied demographic. This observation underscores the genetic underpinnings of ischemic stroke and highlights the potential of genetic screening or interventions targeting these SNPs to mitigate stroke risk in specific populations [76].
7.2. Apolipoprotein A5 in Alzheimer's Disease (AD):
Elevated cholesterol levels in the blood have been implicated as a potential risk factor in AD. Some studies have pointed to specific genetic variations in proteins that regulate cholesterol metabolism as potential contributors to AD onset. However, the role of the ApoA5 gene, known to be involved in lipid metabolism, was brought into question. A study examining the -1131T>C polymorphism of the ApoA5 gene in 106 AD patients found no association between this genetic variation and the development of AD [77]. This suggests that while cholesterol regulation may play a role in AD, not all related genetic variants are necessarily risk factors for the disease.
8. Therapeutic Implications
8.1. Existing Therapies
Current therapies targeting ApoA1 include lifestyle modifications and pharmacological interventions aimed at increasing HDL-C levels. Statins, fibrates, and niacin are commonly used, but their efficacy in directly modulating ApoA1 levels and improving clinical outcomes in neurological diseases remains unclear. Emerging therapies, such as recombinant ApoA1 Milano, have shown promise in preclinical models of AD by reducing Aβ deposition. However, clinical trials are needed to validate these findings in humans.
8.2. Future Research Directions
As diagnostic biomarkers, apolipoproteins could facilitate the early detection of diseases and the development of personalized treatment plans, thereby introducing a new era of precision medicine that is based on genetic variations associated with apolipoproteins. Additionally, exploring the interactions between ApoA1 and other apolipoproteins, such as ApoA4 and ApoA5, could provide insights into comprehensive lipid management strategies. Investigating the role of ApoA1 in conjunction with anti-inflammatory and antioxidant therapies may also yield synergistic benefits for patients with neurological and cardiovascular diseases.
9. Conclusion
This review emphasizes the importance of ApoA in cardiovascular and neurological conditions. It highlights the significance of ApoA in the regulation of lipid metabolism and transport, which are essential for cardiovascular health, as well as its involvement in neurodegenerative processes. Furthermore, the impact of ApoA on cerebrovascular health and stroke risk accentuates its significance in neurological conditions and stroke risk. The goal of future research is to explore the potential of ApoA as a therapeutic strategy for the prevention and treatment of these conditions.
Author Contributions
RM and ZN contributed to manuscript writing and tabulations. All authors, contributing to various capacities, critically reviewed the manuscript and collectively approved it for publication.
Funding
open access publication of this article is covered by Qatar National Library.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mangaraj, M.; Nanda, R.; Panda, S. Apolipoprotein A-I: A Molecule of Diverse Function. Indian J. Clin. Biochem. 2015, 31, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Gauthamadasa, K.; Rosales, C.; Pownall, H.J.; Macha, S.; Jerome, W.G.; Huang, R.; Silva, R.A.G.D. Speciated Human High-Density Lipoprotein Protein Proximity Profiles. Biochemistry 2010, 49, 10656–10665. [Google Scholar] [CrossRef] [PubMed]
- Darabi, M.; Guillas-Baudouin, I.; Le Goff, W.; Chapman, M.J.; Kontush, A. Therapeutic applications of reconstituted HDL: When structure meets function. Pharmacol. Ther. 2015, 157, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Kohan, A.B.; Lo, C.-M.; Liu, M.; Howles, P.; Tso, P. Apolipoprotein A-IV: a protein intimately involved in metabolism. J. Lipid Res. 2015, 56, 1403–1418. [Google Scholar] [CrossRef] [PubMed]
- I Kamboh, M.; Friedlaender, J.S.; I Ahn, Y.; E Ferrell, R. A common deletion polymorphism in the apolipoprotein A4 gene and its significance in lipid metabolism. Arter. Thromb. A J. Vasc. Biol. 1994, 14, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.; Lee, D.; Hagaman, J.; Maeda, N. Marked reduction of high density lipoprotein cholesterol in mice genetically modified to lack apolipoprotein A-I. . Proc. Natl. Acad. Sci. 1992, 89, 7134–7138. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, J.M.; Cassidy, D.K.; Civeira, F.; Bisgaier, C.L.; Schaefer, E.J. Familial apolipoprotein A-I, C-III, and AIV deficiency and premature atherosclerosis due to deletion of a gene complex on chromosome 11. J Biol Chem. 1989, 264, 16339–16342. [Google Scholar] [CrossRef] [PubMed]
- Do, R.; Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Gao, C.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J.; et al. Common variants associated with plasma triglycerides and risk for coronary artery disease. Nat. Genet. 2013, 45, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Kato, H. [Apolipoprotein A I-C III-A IV deficiency]. Nihon Rinsho. 1994, 52, 3253–3256. [Google Scholar]
- Xiong, X.; Duan, Z.; Zhou, H.; Huang, G.; Niu, L.; Luo, Z.; et al. Correlation of apolipoprotein A-I with T cell subsets and interferon-γ in coronary artery disease. Immun Inflamm Dis. 2023, 11, e797. [Google Scholar] [CrossRef]
- Patsch, W.; Sharrett, A.R.; Chen, I.Y.; Lin-Lee, Y.C.; Brown, S.A.; Gotto, A.M., Jr.; et al. Associations of allelic differences at the A-I/C-III/A-IV gene cluster with carotid artery intima-media thickness and plasma lipid transport in hypercholesterolemic-hypertriglyceridemic humans. Arterioscler Thromb. 1994, 14, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, K.; Mashiba, S.; Hashimoto, H.; Kojima, S.; Matsuno, S.; Takeya, M.; et al. Low-density lipoprotein (LDL), which includes apolipoprotein A-I (apoAI-LDL) as a novel marker of coronary artery disease. Clin Chim Acta 2008, 397, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Wilson, C.J.; Mei, X.; Wales, T.E.; Engen, J.R.; Gursky, O. Structural Stability and Local Dynamics in Disease-Causing Mutants of Human Apolipoprotein A-I: What Makes the Protein Amyloidogenic? J Mol Biol. 2016, 428, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Obici, L.; Franceschini, G.; Calabresi, L.; Giorgetti, S.; Stoppini, M.; Merlini, G.; Bellotti, V. Structure, function and amyloidogenic propensity of apolipoprotein A-I. Amyloid 2006, 13, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Hamidi Asl, K.; Liepnieks, J.J.; Nakamura, M.; Parker, F.; Benson, M.D. A novel apolipoprotein A-1 variant, Arg173Pro, associated with cardiac and cutaneous amyloidosis. Biochem Biophys Res Commun. 1999, 257, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Moshe-Lilie, O.; Heitner, S.; Karam, C. Unique skin manifestations of hereditary apolipoprotein A-1 amyloidosis. Oxf. Med Case Rep. 2020, 2020, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.L.; Cao, D.; Lu, H.; Mans, R.A.; Su, Y.R.; Jungbauer, L.; Linton, M.F.; Fazio, S.; LaDu, M.J.; Li, L. Overexpression of Human Apolipoprotein A-I Preserves Cognitive Function and Attenuates Neuroinflammation and Cerebral Amyloid Angiopathy in a Mouse Model of Alzheimer Disease. J. Biol. Chem. 2010, 285, 36958–36968. [Google Scholar] [CrossRef] [PubMed]
- Lefterov, I.; Fitz, N.F.; Cronican, A.A.; Fogg, A.; Lefterov, P.; Kodali, R.; et al. Apolipoprotein A-I deficiency increases cerebral amyloid angiopathy and cognitive deficits in APP/PS1DeltaE9 mice. J Biol Chem. 2010, 285, 36945–36957. [Google Scholar] [CrossRef]
- Jacobs-Cachá, C.; Puig-Gay, N.; Helm, D.; Rettel, M.; Sellarès, J.; Meseguer, A.; Savitski, M.M.; Moreso, F.J.; Soler, M.J.; Seron, D.; et al. A misprocessed form of Apolipoprotein A-I is specifically associated with recurrent Focal Segmental Glomerulosclerosis. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Brewer, H.B.; Davidson, W.S.; Fayad, Z.A.; Fuster, V.; Goldstein, J.; Hellerstein, M.; Jiang, X.-C.; Phillips, M.C.; Rader, D.J.; et al. Cholesterol Efflux and Atheroprotection: Advancing the Concept of Reverse Cholesterol Transport. Circulation 2012, 125, 1905–1919. [Google Scholar] [CrossRef]
- Santonastaso, A.; Maggi, M.; De Jonge, H.; Scotti, C. High resolution structure of human apolipoprotein (a) kringle IV type 2: beyond the lysine binding site. J. Lipid Res. 2020, 61, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Verdier, C.; Martinez, L.O.; Ferrières, J.; Elbaz, M.; Genoux, A.; Perret, B. Targeting high-density lipoproteins: Update on a promising therapy. Arch. Cardiovasc. Dis. 2013, 106, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Kohan, A.B.; Lo, C.-M.; Liu, M.; Howles, P.; Tso, P. Apolipoprotein A-IV: a protein intimately involved in metabolism. J. Lipid Res. 2015, 56, 1403–1418. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Catapano, A.L.; Baigent, C.; Ray, K.K.; Cannon, C.P. Emerging therapies in dyslipidemia management. European Heart Journal 2020, 41, 997–1004. [Google Scholar]
- I Kamboh, M.; Friedlaender, J.S.; I Ahn, Y.; E Ferrell, R. A common deletion polymorphism in the apolipoprotein A4 gene and its significance in lipid metabolism. Arter. Thromb. A J. Vasc. Biol. 1994, 14, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Dallinga-Thie, G.M.; Bu, G.; de Vries, R. LDL receptor deficiency results in decreased cell proliferation and presynaptic bouton density in the murine hippocampus. Front Aging Neurosci. 2016, 8, 304. [Google Scholar]
- Nilsson, S.K.; Heeren, J.; Olivecrona, G.; Merkel, M. Apolipoprotein A-V; a potent triglyceride reducer. Atherosclerosis 2011, 219, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, A. (2"14) "Apolipoprotein A-I mutations and clinical evaluation," The HDL Handbook, pp. 9–. [CrossRef]
- Yui, Y.; Aoyama, T.; Morishita, H.; Takahashi, M.; Takatsu, Y.; Kawai, C. Serum prostacyclin stabilizing factor is identical to apolipoprotein A-I (Apo A-I). A novel function of Apo A-I. J. Clin. Investig. 1988, 82, 803–807. [Google Scholar] [CrossRef]
- Huang, Y.; DiDonato, J.A.; Levison, B.S.; Schmitt, D.; Li, L.; Wu, Y.; Buffa, J.; Kim, T.; Gerstenecker, G.S.; Gu, X.; et al. An abundant dysfunctional apolipoprotein A1 in human atheroma. Nat. Med. 2014, 20, 193–203. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Brewer, H.B., Jr.; Ansell, B.J.; Barter, P.; Chapman, M.J.; Heinecke, J.W.; Kontush, A.; Tall, A.R.; Webb, N.R. Dysfunctional HDL and atherosclerotic cardiovascular disease. Nat. Rev. Cardiol. 2016, 13, 48–60. [Google Scholar] [CrossRef]
- Liao, B.; Cheng, K.; Dong, S.; Liu, H.; Xu, Z. Effect of apolipoprotein A1 genetic polymorphisms on lipid profiles and the risk of coronary artery disease. Diagn. Pathol. 2015, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Dastani, Z.; Dangoisse, C.; Boucher, B.; Desbiens, K.; Krimbou, L.; Dufour, R.; Hegele, R.A.; Pajukanta, P.; Engert, J.C.; Genest, J.; et al. A novel nonsense apolipoprotein A-I mutation (apoA-IE136X) causes low HDL cholesterol in French Canadians. Atherosclerosis 2006, 185, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhang, Y.; Hu, P.; Wang, J.; Lu, N. Serum apolipoprotein A1 rather than apolipoprotein B is associated with hypertension prevalence in Chinese people with coronary artery disease. Blood Press. Monit. 2022, 27, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Kihara, T.; Yamagishi, K.; Honda, K.; Ikeda, A.; Yatsuya, H.; Saito, I.; Kokubo, Y.; Yamaji, T.; Shimazu, T.; Sawada, N.; et al. Apolipoprotein A2 Isoforms in Relation to the Risk of Myocardial Infarction: A Nested Case-Control Analysis in the JPHC Study. J. Atheroscler. Thromb. 2021, 28, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, J.P.; Bakker, S.J.L.; Sparks, C.E.; Dullaart, R.P.F. Apolipoprotein A-II Influences Apolipoprotein E-Linked Cardiovascular Disease Risk in Women with High Levels of HDL Cholesterol and C-Reactive Protein. PLOS ONE 2012, 7, e39110. [Google Scholar] [CrossRef] [PubMed]
- Mendis S, Davis S, Norrving B; World Health Organization. World Health Organization (WHO) global atlas on cardiovascular disease prevention and control. World Health Organization; 2019.
- Grundy, S.M.; Stone, N.J.; Bailey, A.L.; et al. 2018 AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA guideline on the management of blood cholesterol: A report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. J Am Coll Cardiol. 2019, . 73, e285–e350. [Google Scholar] [CrossRef]
- Vine, D.F.; Wang, Y.; Xia, S. Blood-brain barrier disruption: Adjuvant therapy for enhancement of CNS drug delivery. Transl Stroke Res. 2019, 10, 144–156. [Google Scholar]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Leducq Transatlantic Network on Atherothrombosis. Inflammation in atherosclerosis: From pathophysiology to practice. J Am Coll Cardiol. 2019, 54, 2129–2138. [Google Scholar] [CrossRef]
- Furtado, J.D.; Campos, H.; Appel, L.J. The need for cardiovascular outcome trials of low-density lipoprotein cholesterol-lowering therapies: The PCSK9 inhibitors and beyond. Circulation 2019, 139, 1948–1957. [Google Scholar]
- Lee, H.Y.; Birkenfeld, A.L.; Jornayvaz, F.R. Apolipoprotein CIII: A hepatokine and beyond. Diabetologia 2019, 61, 529–534. [Google Scholar]
- Qu, J.; Ko, C.-W.; Tso, P.; Bhargava, A. Apolipoprotein A-IV: A Multifunctional Protein Involved in Protection against Atherosclerosis and Diabetes. Cells 2019, 8, 319. [Google Scholar] [CrossRef]
- Xu, L.-B.; Zhou, Y.-F.; Yao, J.-L.; Sun, S.-J.; Rui, Q.; Yang, X.-J.; Li, X.-B. Apolipoprotein A1 polymorphisms and risk of coronary artery disease: a meta-analysis. Arch. Med Sci. 2017, 4, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Shen, Y.; Li, Q.R.; Ding, F.H.; Wang, X.Q.; Liu, H.J.; Yan, X.X.; Wang, L.J.; Yang, K.; Wang, H.B.; et al. Glycated Apolipoprotein A-IV Induces Atherogenesis in Patients With CAD in Type 2 Diabetes. Circ. 2017, 70, 2006–2019. [Google Scholar] [CrossRef] [PubMed]
- Bois, M.C.; Dasari, S.; Mills, J.R.; Theis, J.; Highsmith, W.E.; Vrana, J.A.; Grogan, M.; Dispenzieri, A.; Kurtin, P.J.; Maleszewski, J.J. Apolipoprotein A-IV–Associated Cardiac Amyloidosis. Circ. 2017, 69, 2248–2249. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Dong, R.; Yan, X.; Li, M.; Wang, S. Apolipoprotein A4 (APOA4): A novel therapeutic target for cardiovascular diseases. Eur J Med Chem. 2021, 211, 113116. [Google Scholar]
- Fruchart-Najib, J.; Baugé, E.; Niculescu, L.S.; Pham, T.; Thomas, B.; Rommens, C.; Majd, Z.; Brewer, B.; Pennacchio, L.A.; Fruchart, J.C. Mechanism of triglyceride lowering in mice expressing human apolipoprotein A5. Biochem Biophys Res Commun. 2004, 319, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Zhang, Z.; Yao, C.; Zhao, S. Emerging evidences for the opposite role of apolipoprotein C3 and apolipoprotein A5 in lipid metabolism and coronary artery disease. Lipids Heal. Dis. 2019, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-Q.; Demissie, S.; Cupples, L.A.; Zhu, Y.; Adiconis, X.; Parnell, L.D.; Corella, D.; Ordovas, J.M. Influence of the APOA5 locus on plasma triglyceride, lipoprotein subclasses, and CVD risk in the Framingham Heart Study. J. Lipid Res. 2004, 45, 2096–2105. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K.; Heeren, J.; Olivecrona, G.; Merkel, M. Apolipoprotein A-V; a potent triglyceride reducer. Atherosclerosis 2011, 219, 15–21. [Google Scholar] [CrossRef]
- Dong, H.; Chen, W.; Wang, X.; et al. Apolipoprotein A1, B levels, and their ratio and the risk of a first stroke: a meta-analysis and case-control study. Metab Brain Dis. 2015, 30, 1319–1330. [Google Scholar] [CrossRef]
- Endres, K. Apolipoprotein A1, the neglected relative of Apolipoprotein E and its potential role in Alzheimer’s disease. Neural Regen. Res. 2021, 16, 2141–2148. [Google Scholar] [CrossRef] [PubMed]
- Johansson, P.; Almqvist, E.G.; Bjerke, M.; Wallin, A.; Johansson, J.-O.; Andreasson, U.; Blennow, K.; Zetterberg, H.; Svensson, J. Reduced Cerebrospinal Fluid Concentration of Apolipoprotein A-I in Patients with Alzheimer’s Disease. J. Alzheimer's Dis. 2017, 59, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Retana, S.F.-D.; Montañola, A.; Marazuela, P.; De La Cuesta, M.; Batlle, A.; Fatar, M.; Grudzenski, S.; Montaner, J.; Hernández-Guillamon, M. Intravenous treatment with human recombinant ApoA-I Milano reduces beta amyloid cerebral deposition in the APP23-transgenic mouse model of Alzheimer's disease. Neurobiol. Aging 2017, 60, 116–128. [Google Scholar] [CrossRef]
- Button, E.B.; Boyce, G.K.; Wilkinson, A.; Stukas, S.; Hayat, A.; Fan, J.; Wadsworth, B.J.; Robert, J.; Martens, K.M.; Wellington, C.L. ApoA-I deficiency increases cortical amyloid deposition, cerebral amyloid angiopathy, cortical and hippocampal astrogliosis, and amyloid-associated astrocyte reactivity in APP/PS1 mice. Alzheimer's Res. Ther. 2019, 11, 44. [Google Scholar] [CrossRef]
- Swanson, C.R.; Li, K.; Unger, T.L.; Gallagher, M.D.; Van Deerlin, V.M.; Agarwal, P.; Leverenz, J.; Roberts, J.; Samii, A.; Gross, R.G.; et al. Lower plasma apolipoprotein A1 levels are found in Parkinson's disease and associate with apolipoprotein A1 genotype. Mov. Disord. 2014, 30, 805–812. [Google Scholar] [CrossRef]
- Qiang, J.K.; Wong, Y.C.; Siderowf, A.; Hurtig, H.I.; Xie, S.X.; Lee, V.M.; Trojanowski, J.Q.; Yearout, D.; Leverenz, J.B.; Montine, T.J.; et al. Plasma apolipoprotein A1 as a biomarker for Parkinson disease. Ann. Neurol. 2013, 74, 119–127. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, Y.F.; Liu, M.; Ma, L.L.; Peng, F.H.; Huang, Q.L.; Ma, X.M.; Chen, X.H. Syphilitic dementia and lipid metabolism. Eur. J. Neurol. 2016, 23, 1541–1547. [Google Scholar] [CrossRef]
- Kim, W.S.; He, Y.; Phan, K.; Ahmed, R.M.; Rye, K.-A.; Piguet, O.; Hodges, J.R.; Halliday, G.M. Altered High Density Lipoprotein Composition in Behavioral Variant Frontotemporal Dementia. Front. Neurosci. 2018, 12, 847. [Google Scholar] [CrossRef]
- Thompson, A.G.; Talbot, K.; Turner, M.R. Higher blood high density lipoprotein and apolipoprotein A1 levels are associated with reduced risk of developing amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 2021, 93, 75–81. [Google Scholar] [CrossRef]
- Meyers, L.; Groover, C.J.; Douglas, J.; Lee, S.; Brand, D.; Levin, M.C.; Gardner, L.A. A role for Apolipoprotein A-I in the pathogenesis of multiple sclerosis. J. Neuroimmunol. 2014, 277, 176–185. [Google Scholar] [CrossRef]
- Gardner, L.A.; Levin, M.C. Importance of Apolipoprotein A-I in Multiple Sclerosis. Front. Pharmacol. 2015, 6, 278. [Google Scholar] [CrossRef] [PubMed]
- Bodha, B. Cholesterol metabolism in spinocerebellar ataxias: Trying to unravel the underlying mechanism in the neurodegenerative disorders. Student Theses Faculty of Science and Engineering. https://fse.studenttheses.ub.rug.nl/21357/.
- Kihara, T.; Yamagishi, K.; Honda, K.; Ikeda, A.; Yatsuya, H.; Saito, I.; Kokubo, Y.; Yamaji, T.; Shimazu, T.; Sawada, N.; et al. Apolipoprotein A2 Isoforms in Relation to the Risk of Myocardial Infarction: A Nested Case-Control Analysis in the JPHC Study. J. Atheroscler. Thromb. 2021, 28, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Li, J.; Bao, Z.; Ruan, Q.; Yu, Z. Serum Levels of ApoA1 and ApoA2 Are Associated with Cognitive Status in Older Men. BioMed Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; He, Y.; Phan, K.; Ahmed, R.M.; Rye, K.-A.; Piguet, O.; Hodges, J.R.; Halliday, G.M. Altered High Density Lipoprotein Composition in Behavioral Variant Frontotemporal Dementia. Front. Neurosci. 2018, 12, 847. [Google Scholar] [CrossRef] [PubMed]
- McComb, M.; Krikheli, M.; Uher, T.; Browne, R.W.; Srpova, B.; Oechtering, J.; Maceski, A.M.; Tyblova, M.; Jakimovski, D.; Ramasamy, D.P.; et al. Neuroprotective associations of apolipoproteins A-I and A-II with neurofilament levels in early multiple sclerosis. J. Clin. Lipidol. 2020, 14, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.R.; Wang, Y.; Adili, R.; Ju, L.; Spring, C.M.; Jin, J.W.; Yang, H.; Neves, M.A.D.; Chen, P.; Yang, Y.; et al. Apolipoprotein A-IV binds αIIbβ3 integrin and inhibits thrombosis. Nat. Commun. 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, W.; Cao, X.; Wang, Y.; Zhu, C.; Guan, J. Identification of Serum Biomarkers in Patients with Alzheimer’s Disease by 2D-DIGE Proteomics. Gerontology 2022, 68, 686–698. [Google Scholar] [CrossRef]
- Lu, W.; Wan, X.; Liu, B.; Rong, X.; Zhu, L.; Li, P.; Li, J.; Wang, L.; Cui, L.; Wang, X. Specific Changes of Serum Proteins in Parkinson's Disease Patients. PLOS ONE 2014, 9, e95684. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, X.; Sun, L.; Qing, Y.; Hu, X.; Jiang, J.; Wang, D.; Cui, G.; Gao, Y.; Zhang, E.; et al. Decreased serum apolipoprotein A4 as a potential peripheral biomarker for patients with schizophrenia. J. Psychiatr. Res. 2021, 137, 14–21. [Google Scholar] [CrossRef]
- Sun, B.; Li, D.; Hou, X.; Li, W.; Gou, Y.; Hu, F.; Li, W.; Shi, X. A novel electrochemical immunosensor for the highly sensitive and selective detection of the depression marker human apolipoprotein A4. Bioelectrochemistry 2020, 135, 107542. [Google Scholar] [CrossRef]
- Sadeghi, M.; Roohafza, H.; Afshar, H.; Rajabi, F.; Ramzani, M.; Shemirani, H.; Sarafzadeghan, N. Relationship between depression and apolipoproteins A and B: a case–control study. Clinics 2011, 66, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-L.; Zhang, X.-M.; Mao, X.-J.; Deng, H.; Li, H.-F.; Press, R.; Fredrikson, S.; Zhu, J. Altered cerebrospinal fluid index of prealbumin, fibrinogen, and haptoglobin in patients with Guillain-Barré syndrome and chronic inflammatory demyelinating polyneuropathy. Acta Neurol. Scand. 2011, 125, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Li, L.; Deng, S.; He, Z. The association between apolipoprotein A1-C3-A5 gene cluster promoter polymorphisms and risk of ischemic stroke in the northern Chinese Han population. J. Int. Med Res. 2017, 45, 2042–2052. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, F.A.F.; de Labio, R.W.; Rigolin, V.d.O.S.; Minett, T.; Bertolucci, P.H.F.; Smith, M.d.A.C.; Payão, S.L.M. Apolipoprotein A-V gene polymorphism –1131T>C and Alzheimer's disease. J. Alzheimer's Dis. 2006, 10, 365–369. [Google Scholar] [CrossRef]
Figure 1.
The reverse cholesterol transport process.
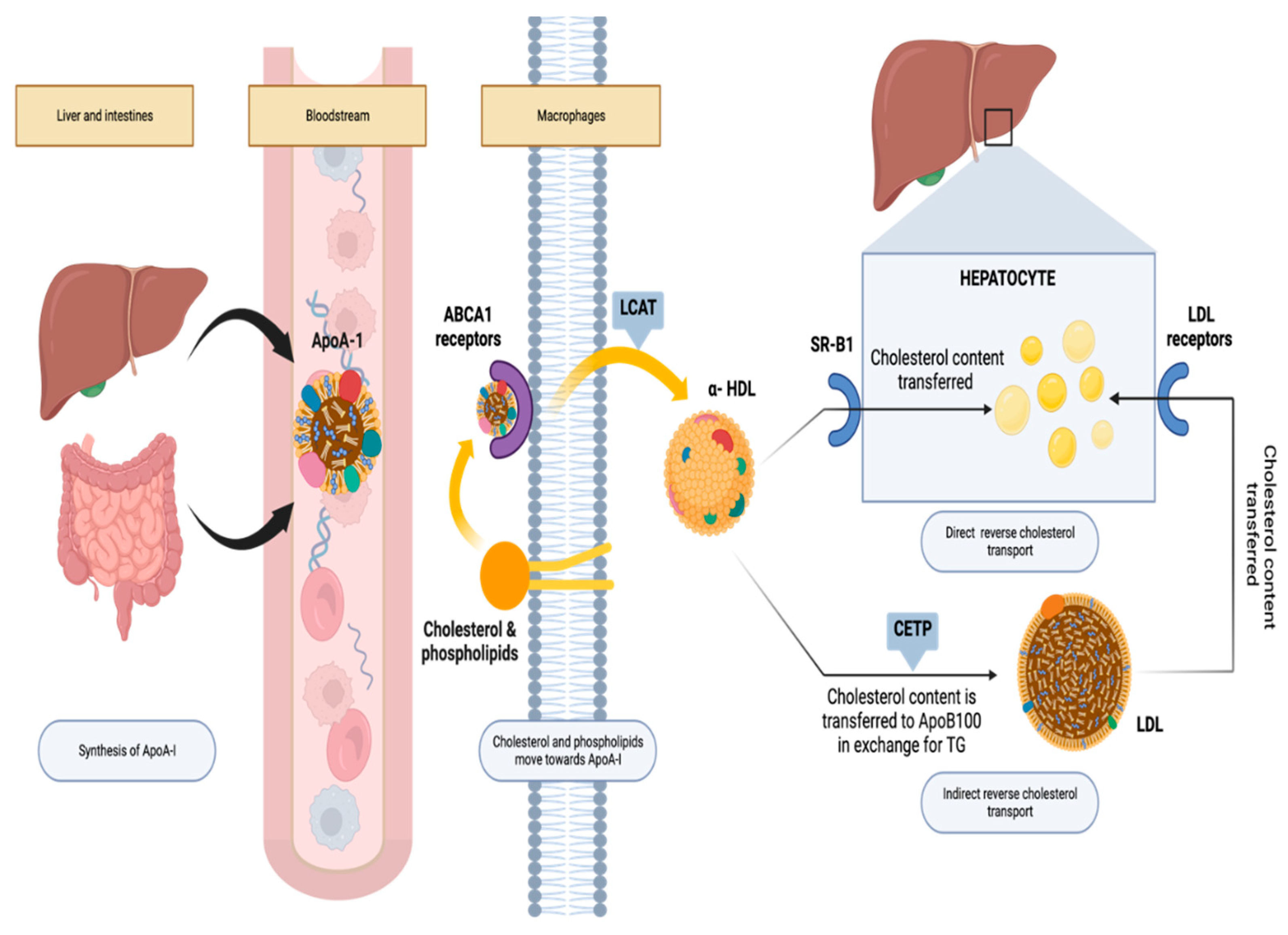
Figure 2.
The metabolic pathway of ApoA4 containing lipoproteins and potential anti-atherogenic mechanisms associated with ApoA4. After fat absorption, ApoA4 is produced and transported with chylomicrons through the lymphatic system into the bloodstream. In circulation, it enhances lipoprotein lipase (LPL) activity when combined with ApoC2. While most ApoA4 separates from chylomicron remnants (CR), around 25% transfers to high-density lipoproteins (HDL), with the remainder circulating in a lipid-free state. (1) ApoA4 is implicated in reverse cholesterol transport, potentially by stimulating LCAT activity, aiding in cholesterol transfer from foam cells or peripheral cells to HDL. (2) It may lower oxidized low-density lipoprotein (Ox-LDL) production, which is continually taken up by macrophages, forming foam cells. (3) ApoA4 exhibits anti-inflammatory properties, potentially suppressing P-selectin-mediated leukocyte and platelet adherence to endothelial cells and interacting with αIIbβ3 integrin to inhibit platelet aggregation.
Figure 2.
The metabolic pathway of ApoA4 containing lipoproteins and potential anti-atherogenic mechanisms associated with ApoA4. After fat absorption, ApoA4 is produced and transported with chylomicrons through the lymphatic system into the bloodstream. In circulation, it enhances lipoprotein lipase (LPL) activity when combined with ApoC2. While most ApoA4 separates from chylomicron remnants (CR), around 25% transfers to high-density lipoproteins (HDL), with the remainder circulating in a lipid-free state. (1) ApoA4 is implicated in reverse cholesterol transport, potentially by stimulating LCAT activity, aiding in cholesterol transfer from foam cells or peripheral cells to HDL. (2) It may lower oxidized low-density lipoprotein (Ox-LDL) production, which is continually taken up by macrophages, forming foam cells. (3) ApoA4 exhibits anti-inflammatory properties, potentially suppressing P-selectin-mediated leukocyte and platelet adherence to endothelial cells and interacting with αIIbβ3 integrin to inhibit platelet aggregation.
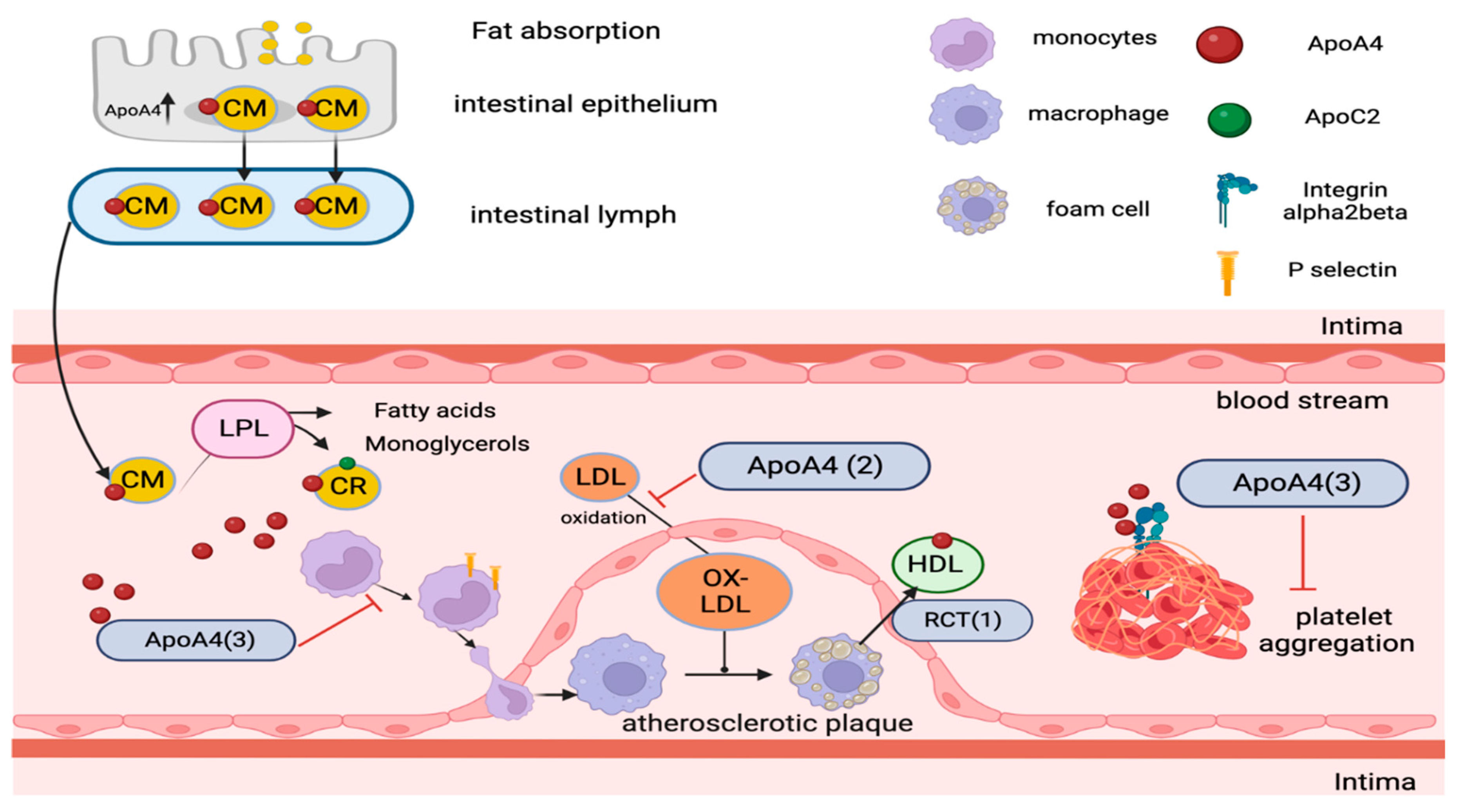

Table 1.
An overview of each isoform's gene, the number of introns typically present in their gene structure (with ApoA4 having one fewer), their primary function in lipid metabolism, and a notable feature that distinguishes each isoform.
Table 1.
An overview of each isoform's gene, the number of introns typically present in their gene structure (with ApoA4 having one fewer), their primary function in lipid metabolism, and a notable feature that distinguishes each isoform.
| Isoform | Gene | Number of Introns | Primary Function | Notable Feature |
|---|---|---|---|---|
| ApoA1 | APOA1 | 3 | Cholesterol efflux, major component of HDL | Most abundant protein in HDL particles |
| ApoA2 | APOA2 | 3 | HDL formation and function, modulates enzyme activity | Exists as a dimer in HDL particles |
| ApoA4 | APOA4 | 2 | Triglyceride metabolism, chylomicron formation | Lacks initial intron found in other isoforms |
| ApoA5 | APOA5 | 3 | Modulates lipoprotein lipase activity, triglyceride hydrolysis | Influences plasma triglyceride levels |
Table 2.
Role of ApoA isoforms in Cardiac and Neural Abnormalities.
| Isoform | disease | Increase (↑) or decrease (↓) | Reference Number |
|---|---|---|---|
| ApoA1 | Coronary artery disease (CAD) | ↓ | 28-32 |
| Hypertension (HTN) | ↓ | 33 | |
| Stroke | ↓ | 51 | |
| Parkinson’s disease (PD) | ↓ | 56-57 | |
| Alzheimer’s disease (AD) | ↓ | 52-54 | |
| Cerebral amyloid angiopathy (CAA) | ↓ | 55 | |
| General paresis | ↓ | 58 | |
| Frontotemporal dementia (FTD) | ↓ | 59 | |
| Amyotrophic later sclerosis (ALS) | ↓ | 60 | |
| Multiple sclerosis (MS) | ↓ | 61-62 | |
| Spinocerebellar degeneration (SCD) | ↓ | 63 | |
| ApoA2 | Myocardial infarction (MI) | ↓ | 34-35 |
| Stroke | ↓ | 64 | |
| Alzheimer’s disease (AD) | ↓ | 65 | |
| Frontotemporal dementia (FTD) | ↓ | 66 | |
| Multiple sclerosis (MS) | ↓ | 67 | |
| ApoA4 | Myocardial infarction (MI) | ↓ | 36-43 |
| Atherosclerosis | ↓ | 36-43 | |
| Alzheimer’s disease (AD) | ↓ | 68 | |
| Parkinson’s disease (PD) | ↓ | 69 | |
| Cardiac amyloidosis | ↑ | 45 | |
| Schizophrenia | ↓ | 70 | |
| Depression | ↓ | 71-72 | |
| Guillain-Barré syndrome (GBS) | N/A | 73 | |
| ApoA5 | Coronary artery disease (CAD) | ↓ | 47-50 |
| Hypertriglyceridemia | ↓ | 47-50 | |
| Multiple sclerosis (MS) | ↑ (gene variation) | 47-50 | |
| Stroke | ↑ (gene variation) | 74 | |
| Alzheimer’s disease | N/A | 75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



