
Submitted:
01 August 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
Background: Cancer stem cells (CSC) are small subpopulation of cancer cells that has the potential to self-renew and has the strong proliferative capacity that sustain tumorigenesis capability. This ability of the CSCs to escape immune responses make the CSCs a primary source of functionally altered, immune-resistant, chemo-resistant aggressive tumor cells. Mentioned characteristic determines the potential advantage of targeting CSCs for the treatment of solid tumor. Method: We downloaded different gene expression datasets of CSCs from the NCBI-GEO database and identified common gens by using suitable Venn tool. Then explored the prognostic significance of the particular genes in particular cancer along with analysis of expression of these genes at protein level in human carcinomas by using different webserver. Finally, using toxicogenomic database we selected several important drugs or chemicals. Result: In this study, we identified common targets between three types of solid tumor and the prognostic significance of the identified genes. The identified drugs capable of targeting the expression or signaling network of designated genes of CSC that may contribute in CSC targeted cancer therapy. Conclusion: We identified several CSC associated genes in human breast, lung and ovarian carcinomas and several drugs that can target the function of these genes. Our study also suggests that there is need for more experimental investigations to find out the actual functional activity and the mechanism of action of the genes of CSC.
Keywords:
Cancer Stem Cell
; Bioinformatics
; Breast cancer
; Lung adenocarcinoma
; Ovarian cancer
; Cancer biomarker
1. Introduction
Abnormal cell growth and series of enabling mutations creates clump of cells that are plastic, genetically malleable and could potentially breach the tissue barrier and spread to secondary sites within the body, a condition known as cancer. Cells multiplies via cell division and cells die due to senescence or when damaged or harbors irreparable DNA damage. The process that intrinsically induces cascade of events of cell death is known as programmed cell death or apoptosis. When cell cycle gets disrupted or the mechanism of programmed cell death fail to initiate death of cells with critical mutations or unrepaired DNA, unregulated, abnormal cell growth can result in formation of lump of tissues in the body known as tumors. Tumors can occur as benign (not malignant) or cancerous. The cancerous or malignant tumors can invade the neighboring tissues [1]. Cancer is the second most common cause of death worldwide [2].
Cancer Stem Cells (CSCs) are a group of self-renewing cells, which has the strong tumorigenic potential and unlimited proliferation capacity. CSCs originate from either differentiated cells or adult tissue of resident stem cell at the stage of tumor onset [3]. Tumor is a heterogeneous mixture of cells with unsynchronized cycling cells that are in different phases of cell cycle. Cells within the tumor mass also differ in terms of mutation, plasticity, level of dedifferentiation and response to stress. Compared to other cells inside a tumor, CSCs are more resistant to therapies and has great adaptability based on their surrounding environment [4] and these cells are much more dedifferentiated. Because of the significant role in resistance to tumor chemotherapy and radiation therapy, as well as tumor recurrence, CSCs has gained much attention [5]. These cancer stem cells maintain their self-renewal properties through different signaling pathways like TGFB signaling, Wnt/B catenin signaling, Notch signaling, Hedgehog signaling etc [6]. Chemotherapy or radiotherapy may not always completely destroy the CSCs, after treatment a small subset of residual CSC can survive, which can cause cancer relapse and ultimately lead to invasiveness and therapy resistance [7]. CSCs induce relapse, metastasis, radiation resistance and multidrug resistance, due to their capacity to remain arrested in G0 phase of cell cycle, and enter cell cycle at opportune moment. Ever since their discovery in leukemia in 1994, CSCs have been viewed as potential target for cancer treatment. There are many intracellular and extracellular factors that control the activity of CSCs, so these factors can serve as drug target [8]. Thus, in the past several years CSC targeted therapy gained increasing interest from research community, especially as potential target in aggressive metastatic cancer.
The most frequent type of cancer in women is breast cancer, following skin cancer. In 2020, more than 2 million new cases are registered globally. Based on GLOBOCAN 2020 data, breast cancer ranks fifth globally in terms of cancer-related fatalities [9]. Majority of the patients get to know about their disease in routine screening, other than that rest of the cases are discovered from breast lump, change in the sizes or discharging of nipple. Genetic mutations and DNA damage, which further activates the secretion of estrogen, are the cause of breast cancer [10].
Based on GLOBOCON 2018 data, in the category of lung cancer, 2.09 million new cases and 1.76 million deaths are registered globally. In both the genders, lung cancer is most common cancer and also the leading cause of cancer-related deaths. Specially in women, this is the third most frequent cause of cancer and second most lethal cancer [11]. 90% of the lung cancer cases are caused due to smoking. Men are affected more compared to women [12]. Ovarian cancer, another frequent type of cancer in women ranks eighth in terms of cancer related deaths, having less than 45% five-year survival rate [13]. Ovarian cancer can strike anyone at any age; persons over 50 are more likely to have it [14]. Right now, majority of the clinical studies concentrated on targeted methods, such as latest attempts at immune therapies.
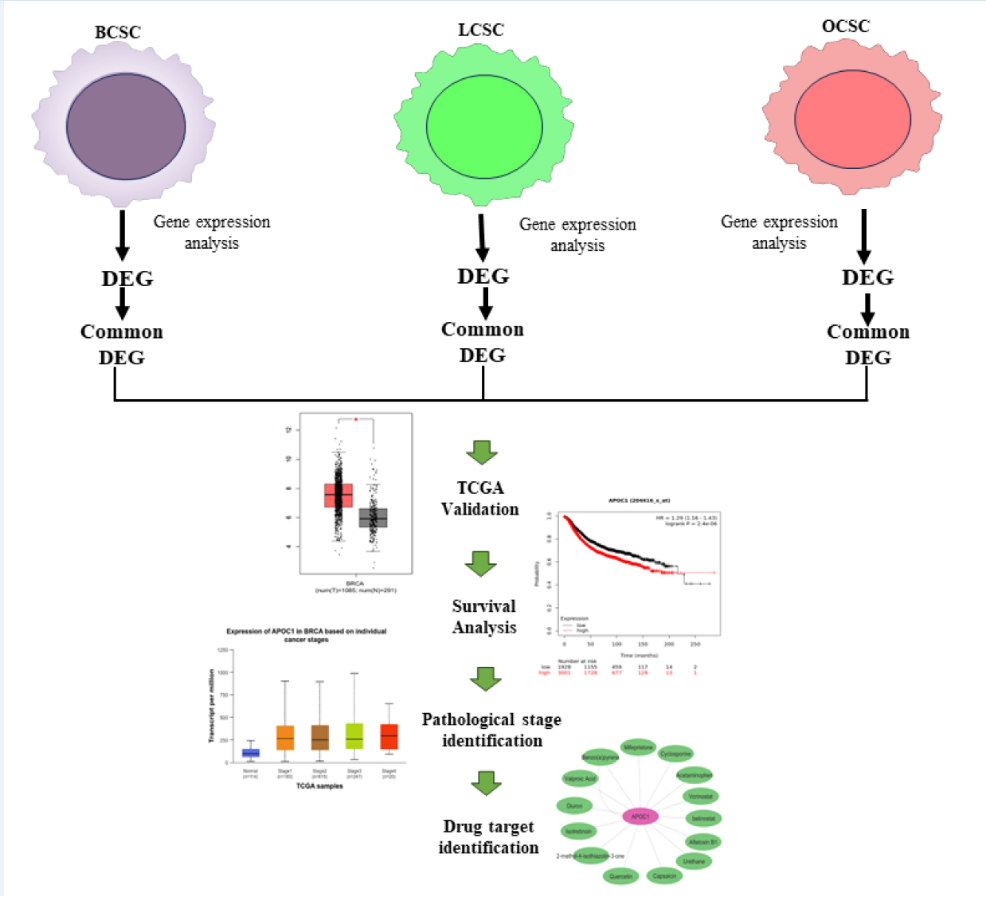
Use of bioinformatics for in silico studies in translational drug delivery is becoming more and more important in both pharmaceutical industries and academics [15]. The roots of bioinformatics can be found more than a half a century ago, although DNA sequencing wasn’t started at that time [16]. The rising amount of data generated through the drug discovery process can now be computationally utilized to address major difficulties. In this present study we focused on identifying CSC linked genes, expression of these genes, role as prognostic biomarker and potential target for drug intervention in breast, lung and ovarian cancer through in depth in silico analysis.
2. Materials and Methods
2.1. Collection of Datasets
Three datasets for breast, lung and ovarian cancer gene expression profiles were downloaded from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/ ). GEO is a database which is freely available for high throughput gene expression and other computational data sets about genomes. GEO is a joint collaboration of National Centre for Biotechnology Information (NCBI) and National Library of Medicine (NLM). It mainly collects the raw and processed data of experimental design for the purpose of studies in high-throughput gene expression and genomics [17]. Datasets utilized are described in Table 1.
2.2. Identification of DEGs
Identification of DEGs was done by using the LIMMA [18] and DESeq2 [19] packages of R Programming. Here we use the log2fold change with the cutoff of 0.5 and P-value kept under 0.05. Common genes were identified by using Bioinformatics and Evolutionary Genomics to generate the Venn diagram (https://bioinformatics.psb.ugent.be/webtools/Venn/).
2.3. Validation of the Genes from TCGA Database
The Cancer Genome Atlas (TCGA) database was utilized to achieve the validation of the genes. The Cancer Genome Atlas or TCGA is launched by The National Institute of Health (NIH) in 2005, for advancement of comprehensive understanding for genetics of cancer to discover innovative cancer therapies, methods and preventions using genome analysis technologies [20]. We used Gene Expression Profiling Interactive Analysis (GEPIA), which is a freely available web-based tool for fast and customizable outcomes based on TCGA. Features including differential expression analysis, profile plotting, correlation analysis, survival analysis and gene detection are offered by GEPIA [21].
2.4. Survival Analysis
We used Kaplan-Meier Plotter (KM Plotter) (https://kmplot.com/analysis/) to perform survival analysis based on gene expression levels of 3 type of cancers. It is an online global database, which has both survival data and gene expression data of ovarian, breast, lung and gastric cancers. Those data were used to evaluate the inter-relation between the prognostic significance and DEGs of cancers [22].
2.5. Identification of Pathological Stages
We used ULCAN (https://ualcan.path.uab.edu/) web server for the detection of the correlation of the genes with different stages of cancer [23]. ULCAN is an important web portal that analyze the TCGA gene expression data from different cancer type. This web portal performs different functions like analysis of relative expression of particular genes across normal and tumor samples and also analyze their expression in different tumors based on tumor grade, race, patients age, nodal metastatic status, body weight, individual cancer stages etc. From the ULCAN web tool we also analyze the relation of the genes with nodal metastatic status in particular cancer.
2.6. Identification of Protein Expression Level of the Genes by Using Human Protein Atlas (HPA)
The HPA database is used to identify the protein expression of the particular genes in particular cancer. The immunohistochemical images downloaded from HPA. HPA mainly contains various immunohistochemical images of various cancers. Based on the staining intensity the results were divided into low, moderate and high. In our study we use HPA database for the identification of protein expression of prognostic genes in particular cancer [24].
2.7. Immune Infiltration Analysis
TIMER2.0 is used to analyze the different immune cell infiltration level with the expression of the particular genes in particular cancer. TIMER 2.0 mainly uses the TCGA data of different cancer and estimates the level of different immune cell infiltration. Here we use CIBERSORT-ABS & TIMER algorithm to identify the M2 macrophage, Dendritic cell, Neutrophil infiltration [25].
2.8. Drug Target Identification
To identify disease targets for each type of cancers we searched in Comparative Toxicogenomics Database (CTD) [26]. CTD (https://ctdbase.org/) is free resource for literature-based database, which show correlation between chemicals, genes/proteins and diseases. It is a combination of cross-species reductionist data from the literature with high-throughput studies to improve knowledge about molecular actions of chemicals. After the identification of targeting drugs or chemicals, the drugs visualized by Cytoscape [27].
3. Results
3.1. Identification of DEGs
The used datasets of each cancer stem cells were taken and examined individually to find out the differentially expressed genes. Three types of cancer datasets are from breast cancer, lung cancer and ovarian cancer. The datasets of the breast cancers are GSE7513, GSE15192 and GSE136287. There were 7578 DEGs (2555 up-regulated and 3935 down-regulated) in GSE7513, 11509 DEGs (3950 up-regulated and 4207 down-regulated) in GSE15192 and 1312 DEGs (266 up-regulated and 73 down-regulated) in GSE136287. The datasets of lung cancers are GSE35603, GSE50627 and GSE160320. There were 10419 DEGs (5321 up-regulated and 5098 down-regulated) in GSE35603, 8100 DEGs (1809 up-regulated and 1832 down-regulated) in GSE50627 and 12482 DEGs (1528 up-regulated and 674 down-regulated) in GSE160320. Similarly, the datasets of ovarian cancers are GSE28799, GSE53759, GSE80373. There were 9881 DEGs (3410 up-regulated and 3422 down-regulated) in GSE28799, 5884 DEGs (1123 up-regulated and 813 down-regulated) in GSE53759 and 13308 DEGs (2310 up-regulated and 2054 down-regulated) in GSE80373.
Using Bioinformatics and Evolutionary Genomics, we achieved the Venn diagrams of common genes from 3 datasets of each type of cancers, where we got 5 up-regulated common genes from the breast cancer datasets, 12 up-regulated and 7 down-regulated common genes from the lung cancer datasets and 29 up-regulated and 20 down-regulated common genes from the ovarian datasets depicted in Figure 1. The common up and downregulated genes from each cancer are given in Supplementary Data S1.
3.2. Validation of the Genes from TCGA Database
We excluded those genes that did not have sufficient clinical information for our analysis. Our final study included total 12 genes (up and down-regulated) from 9 datasets. We have 1 up regulated gene from breast cancer, 1 upregulated and 1 downregulated gene from lung cancer and 4 upregulated and 5 downregulated genes from ovarian cancer. The summery of up and down regulated genes are depicted in Table 2. Validation of the genes from TCGA database are generated in box plots which depicted in Figure 2.
3.3. Survival Analysis
From our survival analysis, we found that, out of 12 genes of different types of cancer, only 8 genes (APOC1, SLC44A5, CAV2, PRRG4, ADCY7, AKAP12, TPM2, FLNC) have significant influence (p value<0.01) on the overall survival of patients of respective cancer and rest of the 4 genes (MECOM, SLC6A12, ABHD11, MKP-1) are not that significant. So, the eight prognostically significant genes used for the further analysis. As shown in Figure 3 higher expression of APOC1 is related to poor overall survival of breast cancer patients. In lung adenocarcinoma patients, higher expression of SLC44A5 is related to decreased overall survival and higher expression of Cav2 is associated with increased overalls survival. However, in case of ovarian cancer we have found opposite results. Like PRRG4 expression is upregulated in ovarian cancer as well as ovarian CSCs but higher expression of PRRG4 is associated with increased overall survival. On the other hand, ADCY7, AKAP12, TPM2 & FLNC are downregulated in ovarian cancer and also in ovarian CSCs but their higher expression is significantly associated with decreased overall survival.
3.4. Correlation between the Expression of CSC Associated Genes and Pathological Stages of Carcinomas
From the ULCAN web server we investigate the correlation between the expression of these eight genes with different pathological stages of particular cancer. Like expression of APOC1 is significantly increased (p value <0.01) in different stages of breast cancer from the normal persons but there were no significant changes between the stages (Figure 4A). There is total four stages are mentioned in this plot which are stage 1, stage 2, stage 3 and stage 4. The SLC44A5 expression is significantly upregulated and CAV2 expression is significantly downregulated (p value <0.01) in different stages of lung adenocarcinoma patients compared to normal individuals (Figure 4B & 4C). However, there was no significant variation between the stages of lung adenocarcinoma in case of the expression of SLC44A5 and CAV2. The other genes (PRRG4, ADCY7, AKAP12, TPM2 & FLNC) does not show any significant changes between the stages of ovarian serous cystadenocarcinoma. In case of ovarian serous cystadenocarcinoma, sufficient data is not available form the normal individuals, indicating need for more investigation to detail role of identified genes in ovarian cancer (Figure 4D, E, F, G, H).
From this web server we also investigate the expression of these genes between tumor and normal samples based on the nodal metastatic status. Nodal metastatic status indicated by N0-N3 label, N0 represents no lymph node metastasis, N1 represents metastasis to 1-3 lymph nodes, N2 represents metastasis in 4-9 lymph nodes and lastly N3 represents metastasis in 10 or more axillary lymph nodes. We found that in breast cancer, APOC1 is significantly increases (p value <0.01) in N0, N1, N2 and N3 stages from the normal healthy individuals (Figure 5A). In case of lung adenocarcinoma patients’ expression of SLC44A5 and CAV2 is significantly increased and decreased (p value <0.01) respectively, in all the nodal metastatic stages compared to normal persons (Figure 5B & 5C). In both breast cancer and lung adenocarcinoma, these three genes do not show any significant changes within the N0, N1, N2 and N3 stages. There was no data available for the genes of ovarian cancer.
3.5. Immunohistochemical Analysis
From the HPA database we explored the protein expression of the eight genes in respective cancers depicted in Figure 6 and found that APOC1 is not detected in normal breast but shows medium expression in breast cancer sample. The expression of SLC44A5 shows lower expression in normal lung tissue but it shows medium expression in lung adenocarcinoma patients. However, CAV2 expression is highly stained in normal lung and shows medium expression in case of patients of lung adenocarcinoma. From the TCGA data we have found that expression of PRRG4 is upregulated in ovarian cancer, from the HPA data supports the TCGA data that in normal ovary the expression of PRRG4 is not detected but ovarian cancer patients show high intensity of PRRG4. But the other genes of ovarian cancer like ADCY7, AKAP12, TPM2 & FLNC shows contrasting data. From the TCGA these four genes are found to be downregulated in ovarian cancer. From the HPA it has been found that expression of ADCY7 and FLNC is not detected in normal ovary but in case of ovarian cancer patients their expression is medium and low respectively. AKAP12 and TPM2 expression is high in both the normal as well as ovarian cancer samples.
3.6. Immune Infiltration Analysis
From the TIMER analysis we have found that expression of the identified genes either positively or negatively corelated with the immune cell infiltration in particular cancer which is represented in Figure 7. There is important significance of the infiltration of M2 macrophage, neutrophil and dendritic cells in the TME. Like Infiltration of M2 macrophages in the TME helps in the promotion of tumor growth and metastasis by releasing the various proangiogenic factors and also, they suppress the proliferation of T cell [28]. The neutrophils in the TME stimulates tumor angiogenesis by releasing MMP9. They induce the recruitment of M2 macrophages and Treg cells and also can inhibit the function of NK cells [29]. Tumor infiltrating dendritic cells have the capacity of immune suppression in the TME rather than stimulating the immune cells [30]. Here from our study, we have found that APOC1 in breast cancer (n = 1100) is positively corelated with the infiltration of M2 macrophage, Neutrophil and Dendritic cells. In Lung adenocarcinoma (n = 515), expression of CAV2 is positively corelated with the infiltration of M2 macrophage, Neutrophil and Dendritic cell. But expression of SLC44A5 is negatively corelated with the infiltration of these immune cells. On the other hand, M2 macrophage, Neutrophil and Dendritic cell infiltration is positively related with the expression of PRRG4 and ADCY7 in ovarian cancer (n = 303). Expression of AKAP12 is positively related with the infiltration of Neutrophil but negatively related with infiltration of Dendritic cell. TPM2 expression is positively related with the infiltration of M2 macrophage and Neutrophil, but their infiltration is not significantly related with the expression of FLNC in case of ovarian cancer. However, expression of AKAP12 and FLNC is negatively corelated with the infiltration of dendritic cell in patients of ovarian cancer.
3.7. Drug Target Identification
From the CTD we have identified several drugs that can influence the expression of particular genes. From the analysis we identified that APOC1, SLC44A5 and PRRG4 are upregulated so we searched for drugs that could decrease their expression (Figure 8A, B & D). Simultaneously, CAV2, ADCY7, AKAP12, TPM2 and FLNC genes are downregulated so here we searched for the drugs that could increase their expression (Figure 8C, E, F, G, H). We found Cyclosporine, Valproic Acid, Ivermectin and Acetaminophen increase the expression of ADCY7. Again, for increasing the expression of CAV2, we found Doxorubicin, Tretinoin, Valproic Acid and Cisplatin. And for decreasing the APOC1 expression, we found drugs such as Valproic Acid, Quercetin and Cyclosporin. We further found, Cyclosporin, Bisphenol S, Sunitinib and Quercetin, decrease the expression of SLC44A5. There also many other drugs that can target the expression of remaining genes. The datasheets derived from the CTD are given in Supplementary Data S2.
4. Discussion
CSC are small subpopulation of cancer cells, which have the capability of self-renewal and leads to the resistance to chemotherapy. Different signaling pathways like JAK-STAT, Hedgehog, TGFB, Notch, Wnt-B catenin, VEGF signaling pathways regulate the development of CSC. Different inhibitors, siRNA, monoclonal antibodies, enzyme inhibitors that intervenes with those signaling pathways can target the cancer stem cells. There are clinical trials ongoing targeting CSCs [31]. Breast, lung and ovarian cancer, originate from the epithelial cells and turn into most aggressive types of cancer by inducing the EMT [32,33,34]. There are different strategies available for targeting the stem cell of these cancers, although, most of these strategies are yet to be clinically approved and plasticity and biology of CSCs need to be explored in more detail for identification of novel targets. In this present study, we first downloaded the CSC gene expression datasets of these three different types of cancer and analyzed genes that are associated with CSCs from different data sets of respective cancer types in order to identify genes that can be important intervention target in these cancers. We tried to identify important genes that are expressed in cancer stem cells of these three types of cancer. Our analysis revealed that there are no common genes between these three types of cancer stem cells, implying that the genes of cancer stem cells are specific for cancer tissue type. Next, we identified common important up and down regulated genes in cancer stem cells of particular type of cancer i.e. breast, lung and ovarian cancer and investigated their relation with different pathological stages of respective type of cancer and also their prognostic value. Finally, we identified several potential candidate drugs that can target those particular genes of cancer stem cells. We validate the identified genes in TCGA data set of respective cancers and also carried out the survival analysis to identify the prognostic significance of these genes. Lastly, we analyzed these selected genes for drug targets, focusing on candidate drugs that can modulate the function of these genes in particular cancers and reduce the CSC mediated cancer proliferation and metastasis.
From our analysis, we identified APOC1(Apolipoprotein C1) gene in breast CSCs, SLC44A5 and CAV2 genes in lung CSCs and MECOM, SLC6A12, PRRG4, ABHD11, DUSP1(MKP-1), ADCY7, AKAP12, TPM2, FLNC in case of ovarian CSCs as some of the most important genes. Out of these twelve genes, eight genes (APOC1, SLC44A5, CAV2, PRRG4, ADCY7, AKAP12, TPM2, FLNC) show significant overall prognostic value in respective cancers.
APOC1 is recently been identified in various types of cancers, such as, colorectal cancer [35], cervical cancer [36] and also in breast cancer [37]. In breast cancer APOC1 induce cell proliferation and metastasis by inducing MAP kinase pathway. In our analysis we have identified APOC1 is upregulated in human breast CSC and also in tumor samples in TCGA database (Figure 2A). When we analyzed the effect of expression status of this gene on overall survival, we found that higher expression of this gene is linked to worse overall survival (Figure 3A). We further analyze the relation with pathological stages of breast cancer with expression of this gene, we found that its expression is significantly increased in different stages from the normal condition and also nodal metastatic status increased in N0, N1, N2, N3 stages of breast cancer. Protein expression data from HPA shows that in normal breast tissue it is not detected but breast cancer tissue shows moderate expression (Figure 6A). TIMER analysis reveals that in breast cancer patients, expression of APOC1 is positively corelated with the infiltration of M2 macrophages, Neutrophils as well as Dendritic cells which contributes in tumor promoting environment in breast cancer (Figure 7A). We identified drugs that can decrease the expression of APOC1 are Valproic acid, Cyclosporin, Quercetin, Isotretinoin amongst several others.
Differential expression of SLC44A5 (Solute Carrier Family 44 Member 5) and CAV2 (Caveolin 2) are found in lung cancer stem cells, where SLC44A5 shows upregulated expression and CAV2 shows downregulated expression. SLC44A5 is one type of choline transporter which plays important role in inducing cell viability and invasion of hepatocellular cancer cells and also inhibits apoptosis [38]. In lung cancer, we could not find any study regarding the role of this gene. From the TCGA data we found that its expression is significantly upregulated in lung adenocarcinoma sample compared to normal samples and its higher expression significantly associated with decreased overall survival of lung cancer patients. The expression of SLC44A5 is significantly increased in the various pathological stages of lung adenocarcinoma patients but there are no significant changes within the stages of lung adenocarcinoma (Figure 4B). Its expression also positively corelated with the nodal metastatic status of patients of lung adenocarcinoma. Several drugs like Cyclosporine, Sunitinib, Bisphenol A and Quercetin can decrease the activity or expression of this gene. Caveolae are small infoldings of the cell membrane and involved in endocytosis, cellular trafficking and also signal transduction. CAV 2 is involved in the inhibition of TGFB signaling pathway and reduce the proliferation of mouse lung endothelial cells [39]. From the TCGA data, we found that its expression is downregulated in lung adenocarcinoma patients and its higher expression is significantly related to increased overall survival, indicating that it could be an important prognostic marker for the patients of lung adenocarcinoma. Also, the expression of CAV2 is significantly decreased in the pathological stages and various nodal metastatic condition from the normal healthy individuals. Expression of CAV2 protein is found to be highly and moderately expressed in Normal lung tissue and lung adenocarcinoma patients respectively (Figure 6C). Immune infiltration analysis shows that infiltration of M2 macrophage, neutrophil and dendritic cells positively corelated with the expression of CAV2 but negatively corelated with the expression of SLC44A5 in case of lung adenocarcinoma patients. As this gene is downregulated in lung cancer, therefore, we tried to identify drugs that could increase its expression leading to perturbation in cancer proliferation. Drugs that can increase the expression of CAV2 includes Doxorubicin. Cisplatin, Tretinoin and Valproic acid.
Next, PRRG4 (Proline Rich and Gla Domain 4), ADCY7 (Adenylate cyclase 7), AKAP12 (A Kinase Anchor Protein 12), TPM2 (Tropomyosin 2), FLNC (Filamin C) genes are identified in ovarian cancer stem cells. PRRG4 is upregulated and ADCY7, AKAP12, TPM2 & FLNC are downregulated in ovarian cancer stem cells when compared with the non- stem cells. PRRG family proteins are involved in cell signalling and regulation, although the specific function of PRRG4 is not fully elucidated. Zhang et al., found that upregulated expression of PRRG4 induces breast cancer metastasis through downregulation of Robo1 via NEDD4 [40]. In another study it has been found that PRRG4 regulates the mitochondrial function and migratory behaviour of breast cancer cells by Src-STAT3-POLG axis [41]. In our study this gene is found to be upregulated in ovarian cancer stem cell but higher expression of this gene is significantly linked with increased overall survival which shows a paradoxical significance of this gene in ovarian cancer patients. HPA data shows PRRG4 is highly expressed in ovarian cancer whereas not detected in case of normal ovary tissue (Figure 6D). PRRG4 induces the M2 macrophage, neutrophil and dendritic cell infiltration in the ovarian cancer microenvironment. Valproic acid, Acetaminophen, Paclitaxel can reduce the activity of the gene in cancer.
ADCY7, AKAP12, TPM2 and FLNC genes show reduced expression in ovarian cancer stem cells and from the TCGA data. ADCY7 is the membrane protein of adenylate cyclase family and induce the signal transduction. There are reports that indicate ADCY7 is associated with poor prognosis of myeloid leukaemia patients [42]. AKAP12 act as a tumor suppressor gene in different types of cancer and can inhibit the proliferation, migration, invasion and angiogenesis of cancer cells as well as block cell cycle through the activation of protein kinase C [43]. Tropomyosins are the actin binding proteins that regulates the force generation and sensing stiffness. Downregulated expression of TPM2 has been found to enhance the proliferation and migration of colorectal cancer cell [44] and also promotes the breast cancer metastasis and chemoresistance [45]. FLNC is an actin crosslinking cytoskeletal protein that maintains the cellular morphology. In a study of gastric cancer, Filamin C supress the metastasis and angiogenesis by reducing the expression of MMP2 [46]. Paradoxically, from the Kaplan-Meier plot, we found that high expression of ADCY7, AKAP12, TPM2 & FLNC genes are significantly associated with decrease overall survival, indicating the need for detail study of the function of these genes in CSCs, tumor microenvironment and survival of cancer cells. There is lack of sufficient data for these genes regarding relation with the different pathological stages and nodal metastatic status of ovarian cancer patients. However, when we investigate the immune infiltration status of these genes in ovarian cancer, we found that ADCY 7 expression positively related the infiltration of M2 macrophage, neutrophil and dendritic cells but AKAP12 expression is positively corelated with the infiltration of only neutrophils. TPM2 expression also positively related with the M2 macrophage and neutrophil infiltration but not significantly related with dendritic cell infiltration whereas expression of FLNC is not significantly related with infiltration of M2 macrophage and neutrophil and negatively related with dendritic cell infiltration in ovarian cancer. From immunohistochemical images we have found that ADCY7 expression is not detected in normal ovary and moderately expressed in cancerous tissue. AKAP12 and TPM2 are highly expressed in both normal ovary and ovarian cancer. However, expression of FLNC is not detected in normal condition and in ovarian cancer it shows lower intensity. Drugs like Doxorubicin, Quercetin, Sunitinib, Cyclosporine, Fulvestrant can target the AKAP12 and cyclosporine, valproic acid, Acetaminophen can increase the effect of ADCY7. Also, Doxorubicin, Valproic acid, Bisphenol A can effectively target the TPM 2 and FLNC gene of ovarian cancer stem cells. From our study, Valproic acid, Sunitinib and Quercetin emerged as drug candidates that can target most of the CSC associated genes or signalling pathways in breast, ovarian and lung cancer. It is important to note that Quercetin has been actively explored as potential candidate drug in cancer but so far clinical studies have failed to replicate same level of impact shown in preclinical and in vitro studies [47] implying that literature and high throughput study based drug data bank may identify many well researched candidate drugs that failed to replicate promising results revealed in vitro/pre-clinical studies in clinical set up and careful screening must be conducted to identify novel promising candidate drugs or small molecules.
5. Conclusions
Overall, our analysis suggests that there are no significant CSC associated genes that has common expression pattern in ovarian, lung and breast cancer data set but we identified common genes amongst different data sets of ovarian, lung and breast cancer respectively. We identified several CSC associated genes that are promising prognostic biomarker for breast, lung and ovarian cancers and also identified several candidate drugs that can modulate the function and expression of these genes or their protein products. Our study also indicates need for thorough experimental analysis of role of these genes in different cancer stem cells, molecular mechanism of their action and relationship with metastasis, survival, chemoresistance and relapse of cancer.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary Data S1: Spreadsheet of common genes from DEG, Supplementary Data S2: Spreadsheet CTD results.
Author Contributions
Pallabi Mondal: Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing- Original draft. Poulami Singh: Software, Formal analysis, Methodology. Krishna Mahanti: Data curation, Software, Writing- review and editing. Sankar Bhattacharyya: Conceptualization, Data curation, Supervision, Writing – Review and editing. Authors also declare that no generative AI tool has been used to write or compose any part of the manuscript, no papermill service is involved and all author agreed to the communicated version of the manuscript.
Funding
This research received no external fundings.
Data Availability Statement
All dataset, software, plugins, versions and methodology described in detail in manuscript, additional detail analysis spreadsheets and information included in supplementary material. Any specific clarification or analysis file will be made available upon request.
Acknowledgments
S.B. thanks Department of Higher education and Department of Science and Technology (DST), Government of West Bengal for infrastructure Grants. P.M. thanks the Counsil of Scientific and Industrial Research & University Grants Commission (CSIR & UGC), India, for fellowship.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cooper GM. The Cell: A Molecular Approach. 2nd edition. Sunderland (MA): Sinauer Associates; 2000. The Development and Causes of Cancer. Available from: https://www.ncbi.nlm.nih.gov/books/NBK9963/.
- Nagai, H., & Kim, Y. H. (2017). Cancer prevention from the perspective of global cancer burden patterns. Journal of thoracic disease, 9(3), 448–451. [CrossRef]
- Walcher, L., Kistenmacher, A. K., Suo, H., Kitte, R., Dluczek, S., Strauß, A., Blaudszun, A. R., Yevsa, T., Fricke, S., & Kossatz-Boehlert, U. (2020). Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Frontiers in immunology, 11, 1280. [CrossRef]
- Najafi, M., Mortezaee, K., & Majidpoor, J. (2019). Cancer stem cell (CSC) resistance drivers. Life sciences, 234, 116781. [CrossRef]
- Biserova, K., Jakovlevs, A., Uljanovs, R., & Strumfa, I. (2021). Cancer Stem Cells: Significance in Origin, Pathogenesis and Treatment of Glioblastoma. Cells, 10(3), 621. [CrossRef]
- Ordaz-Ramos, A., Tellez-Jimenez, O., & Vazquez-Santillan, K. (2023). Signaling pathways governing the maintenance of breast cancer stem cells and their therapeutic implications. Frontiers in cell and developmental biology, 11, 1221175. [CrossRef]
- Bisht, S., Nigam, M., Kunjwal, S. S., Sergey, P., Mishra, A. P., & Sharifi-Rad, J. (2022). Cancer Stem Cells: From an Insight into the Basics to Recent Advances and Therapeutic Targeting. Stem cells international, 2022, 9653244. [CrossRef]
- Yang, L., Shi, P., Zhao, G., Xu, J., Peng, W., Zhang, J., Zhang, G., Wang, X., Dong, Z., Chen, F., & Cui, H. (2020). Targeting cancer stem cell pathways for cancer therapy. Signal transduction and targeted therapy, 5(1), 8. [CrossRef]
- Łukasiewicz, S., Czeczelewski, M., Forma, A., Baj, J., Sitarz, R., & Stanisławek, A. (2021). Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers, 13(17), 4287. [CrossRef]
- Menon G, Alkabban FM, Ferguson T. Breast Cancer. [Updated 2024 Feb 25]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK482286/.
- Bade, B. C., & Dela Cruz, C. S. (2020). Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clinics in chest medicine, 41(1), 1–24. [CrossRef]
- Siddiqui F, Vaqar S, Siddiqui AH. Lung Cancer. [Updated 2023 May 8]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK482357/.
- Webb, P. M., & Jordan, S. J. (2017). Epidemiology of epithelial ovarian cancer. Best practice & research. Clinical obstetrics & gynaecology, 41, 3–14. [CrossRef]
- Roett, M. A., & Evans, P. (2009). Ovarian cancer: an overview. American family physician, 80(6), 609–616.
- Wooller, S. K., Benstead-Hume, G., Chen, X., Ali, Y., & Pearl, F. M. G. (2017). Bioinformatics in translational drug discovery. Bioscience reports, 37(4), BSR20160180. [CrossRef]
- Gauthier, J., Vincent, A. T., Charette, S. J., & Derome, N. (2019). A brief history of bioinformatics. Briefings in bioinformatics, 20(6), 1981–1996. [CrossRef]
- Clough, E., & Barrett, T. (2016). The Gene Expression Omnibus Database. Methods in molecular biology (Clifton, N.J.), 1418, 93–110. [CrossRef]
- Diboun, I., Wernisch, L., Orengo, C. A., & Koltzenburg, M. (2006). Microarray analysis after RNA amplification can detect pronounced differences in gene expression using limma. BMC genomics, 7, 252. [CrossRef]
- Love, M. I., Huber, W., & Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome biology, 15(12), 550. [CrossRef]
- Tomczak, K., Czerwińska, P., & Wiznerowicz, M. (2015). The Cancer Genome Atlas (TCGA): an immeasurable source of knowledge. Contemporary oncology (Poznan, Poland), 19(1A), A68–A77. [CrossRef]
- Tang, Z., Li, C., Kang, B., Gao, G., Li, C., & Zhang, Z. (2017). GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic acids research, 45(W1), W98–W102. [CrossRef]
- Győrffy B. (2024). Integrated analysis of public datasets for the discovery and validation of survival-associated genes in solid tumors. Innovation (Cambridge (Mass.)), 5(3), 100625. [CrossRef]
- Chandrashekar, D. S., Bashel, B., Balasubramanya, S. A. H., Creighton, C. J., Ponce-Rodriguez, I., Chakravarthi, B. V. S. K., & Varambally, S. (2017). UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia (New York, N.Y.), 19(8), 649–658. [CrossRef]
- Asplund, A., Edqvist, P. H., Schwenk, J. M., & Pontén, F. (2012). Antibodies for profiling the human proteome-The Human Protein Atlas as a resource for cancer research. Proteomics, 12(13), 2067–2077. [CrossRef]
- Li, T., Fu, J., Zeng, Z., Cohen, D., Li, J., Chen, Q., Li, B., & Liu, X. S. (2020). TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic acids research, 48(W1), W509–W514. [CrossRef]
- Davis, A. P., Wiegers, T. C., Johnson, R. J., Sciaky, D., Wiegers, J., & Mattingly, C. J. (2023). Comparative Toxicogenomics Database (CTD): update 2023. Nucleic acids research, 51(D1), D1257–D1262. [CrossRef]
- Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., Amin, N., Schwikowski, B., & Ideker, T. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome research, 13(11), 2498–2504. [CrossRef]
- Lin, Y., Xu, J., & Lan, H. (2019). Tumor-associated macrophages in tumor metastasis: biological roles and clinical therapeutic applications. Journal of hematology & oncology, 12(1), 76. [CrossRef]
- Giese, M. A., Hind, L. E., & Huttenlocher, A. (2019). Neutrophil plasticity in the tumor microenvironment. Blood, 133(20), 2159–2167. [CrossRef]
- Tran Janco, J. M., Lamichhane, P., Karyampudi, L., & Knutson, K. L. (2015). Tumor-infiltrating dendritic cells in cancer pathogenesis. Journal of immunology (Baltimore, Md. : 1950), 194(7), 2985–2991. [CrossRef]
- Borlongan, M. C., & Wang, H. (2023). Profiling and targeting cancer stem cell signaling pathways for cancer therapeutics. Frontiers in cell and developmental biology, 11, 1125174. [CrossRef]
- Pal, A. K., Sharma, P., Zia, A., Siwan, D., Nandave, D., Nandave, M., & Gautam, R. K. (2022). Metabolomics and EMT Markers of Breast Cancer: A Crosstalk and Future Perspective. Pathophysiology : the official journal of the International Society for Pathophysiology, 29(2), 200–222. [CrossRef]
- Xiao, D., & He, J. (2010). Epithelial mesenchymal transition and lung cancer. Journal of thoracic disease, 2(3), 154–159. [CrossRef]
- Loret, N., Denys, H., Tummers, P., & Berx, G. (2019). The Role of Epithelial-to-Mesenchymal Plasticity in Ovarian Cancer Progression and Therapy Resistance. Cancers, 11(6), 838. [CrossRef]
- Tang, W., Liu, H., Li, X., Ooi, T. C., Rajab, N. F., Cao, H., & Sharif, R. (2023). Upregulation of APOC1 Promotes Colorectal Cancer Progression and Serves as a Potential Therapeutic Target Based on Bioinformatics Analysis. Journal of oncology, 2023, 2611105. [CrossRef]
- Shi, X., Wang, J., Dai, S., Qin, L., Zhou, J., & Chen, Y. (2020). Apolipoprotein C1 (APOC1): A Novel Diagnostic and Prognostic Biomarker for Cervical Cancer. OncoTargets and therapy, 13, 12881–12891. [CrossRef]
- Zhang, H., Wang, Y., Liu, C., Li, W., Zhou, F., Wang, X., & Zheng, J. (2022). The Apolipoprotein C1 is involved in breast cancer progression via EMT and MAPK/JNK pathway. Pathology, research and practice, 229, 153746. [CrossRef]
- Peng, G. Z., Ye, Q. F., Wang, R., Li, M. X., & Yang, Z. X. (2016). Knockdown by shRNA identifies SLC44A5 as a potential therapeutic target in hepatocellular carcinoma. Molecular medicine reports, 13(6), 4845–4852. [CrossRef]
- Xie, L., Vo-Ransdell, C., Abel, B., Willoughby, C., Jang, S., & Sowa, G. (2011). Caveolin-2 is a negative regulator of anti-proliferative function and signaling of transforming growth factor-β in endothelial cells. American journal of physiology. Cell physiology, 301(5), C1161–C1174. [CrossRef]
- Zhang, L., Qin, Y., Wu, G., Wang, J., Cao, J., Wang, Y., Wu, D., Yang, K., Zhao, Z., He, L., Lyu, J., Li, H., & Gu, H. (2020). PRRG4 promotes breast cancer metastasis through the recruitment of NEDD4 and downregulation of Robo1. Oncogene, 39(49), 7196–7208. [CrossRef]
- Wang, Y., Wang, J., Chen, L., Chen, Z., Wang, T., Xiong, S., Zhou, T., Wu, G., He, L., Cao, J., Liu, M., Li, H., & Gu, H. (2023). PRRG4 regulates mitochondrial function and promotes migratory behaviors of breast cancer cells through the Src-STAT3-POLG axis. Cancer cell international, 23(1), 323. [CrossRef]
- Chen, S. L., Hu, F., Wang, D. W., Qin, Z. Y., Liang, Y., & Dai, Y. J. (2020). Prognosis and regulation of an adenylyl cyclase network in acute myeloid leukemia. Aging, 12(12), 11864–11877. [CrossRef]
- Li H. (2022). Physiologic and pathophysiologic roles of AKAP12. Science progress, 105(3), 368504221109212. [CrossRef]
- Cui, J., Cai, Y., Hu, Y., Huang, Z., Luo, Y., Kaz, A. M., Yang, Z., Chen, D., Fan, X., Grady, W. M., & Wang, J. (2016). Epigenetic silencing of TPM2 contributes to colorectal cancer progression upon RhoA activation. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine, 37(9), 12477–12483. [CrossRef]
- Zhang, J., Zhang, J., Xu, S., Zhang, X., Wang, P., Wu, H., Xia, B., Zhang, G., Lei, B., Wan, L., Zhang, D., & Pang, D. (2018). Hypoxia-Induced TPM2 Methylation is Associated with Chemoresistance and Poor Prognosis in Breast Cancer. Cellular physiology and biochemistry : international journal of experimental cellular physiology, biochemistry, and pharmacology, 45(2), 692–705. [CrossRef]
- Qiao, J., Cui, S. J., Xu, L. L., Chen, S. J., Yao, J., Jiang, Y. H., Peng, G., Fang, C. Y., Yang, P. Y., & Liu, F. (2015). Filamin C, a dysregulated protein in cancer revealed by label-free quantitative proteomic analyses of human gastric cancer cells. Oncotarget, 6(2), 1171–1189. [CrossRef]
- Yang, D., Wang, T., Long, M., & Li, P. (2020). Quercetin: Its Main Pharmacological Activity and Potential Application in Clinical Medicine. Oxidative medicine and cellular longevity, 2020, 8825387. [CrossRef]
Figure 1.
Common Differentially Expressed Gene of different cancer stem cell gene expression datasets. A. Common up-regulated genes between three breast cancer stem cell gene expression datasets. (There are no common downregulated genes found in between three BCSC) B. Common upregulated C. Common down-regulated genes between three lung cancer stem cell gene expression datasets. D. Common up-regulated & E. Common down-regulated genes between three ovarian cancer stem cell gene expression datasets.
Figure 1.
Common Differentially Expressed Gene of different cancer stem cell gene expression datasets. A. Common up-regulated genes between three breast cancer stem cell gene expression datasets. (There are no common downregulated genes found in between three BCSC) B. Common upregulated C. Common down-regulated genes between three lung cancer stem cell gene expression datasets. D. Common up-regulated & E. Common down-regulated genes between three ovarian cancer stem cell gene expression datasets.
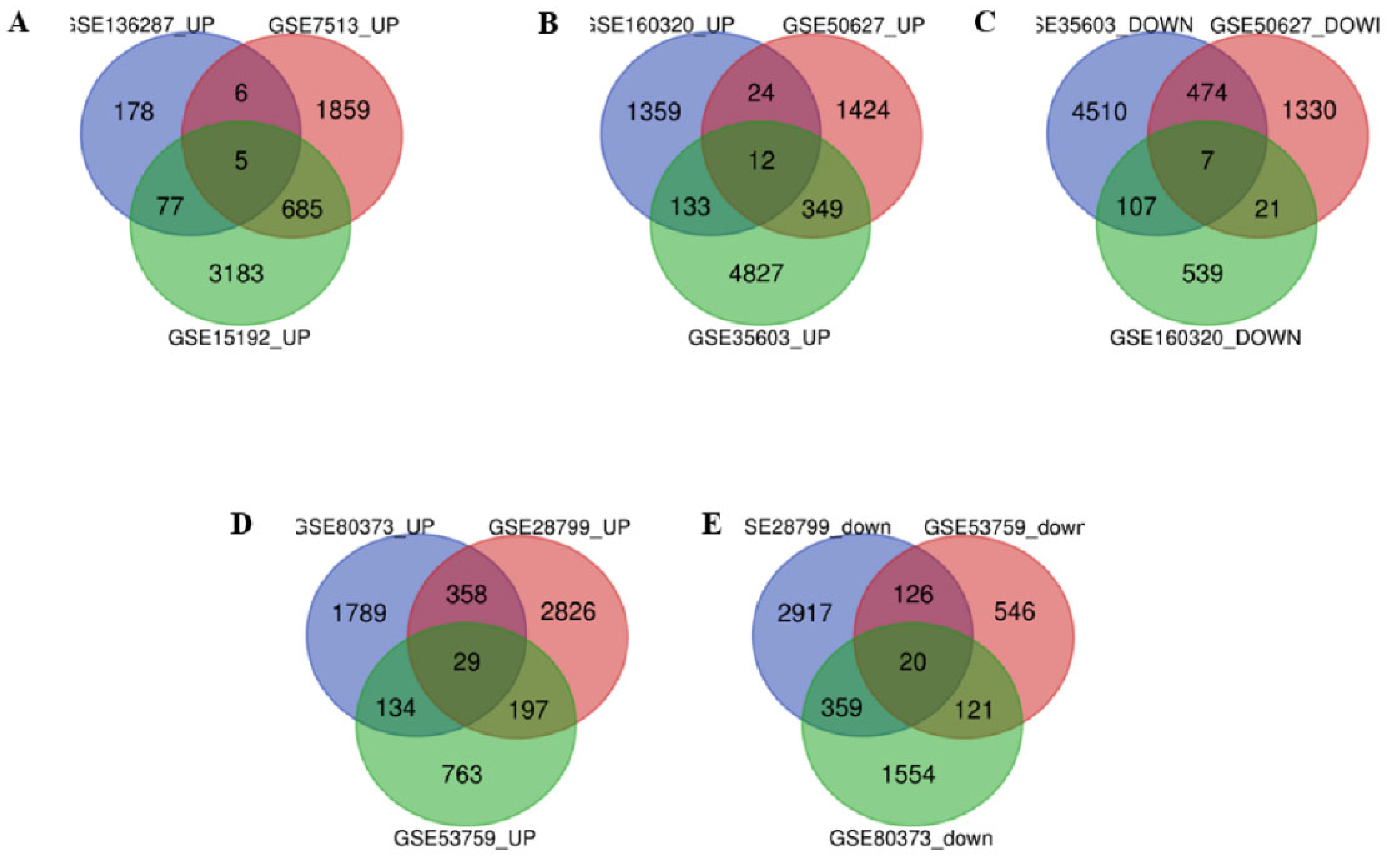
Figure 2.
Expression of the selected genes validated from TCGA by using GEPIA database in respective cancer. A. APOC1 in Breast cancer B. SLC44A5 and C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer. Here red colour represents tumor sample and black colour represents normal samples.
Figure 2.
Expression of the selected genes validated from TCGA by using GEPIA database in respective cancer. A. APOC1 in Breast cancer B. SLC44A5 and C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer. Here red colour represents tumor sample and black colour represents normal samples.
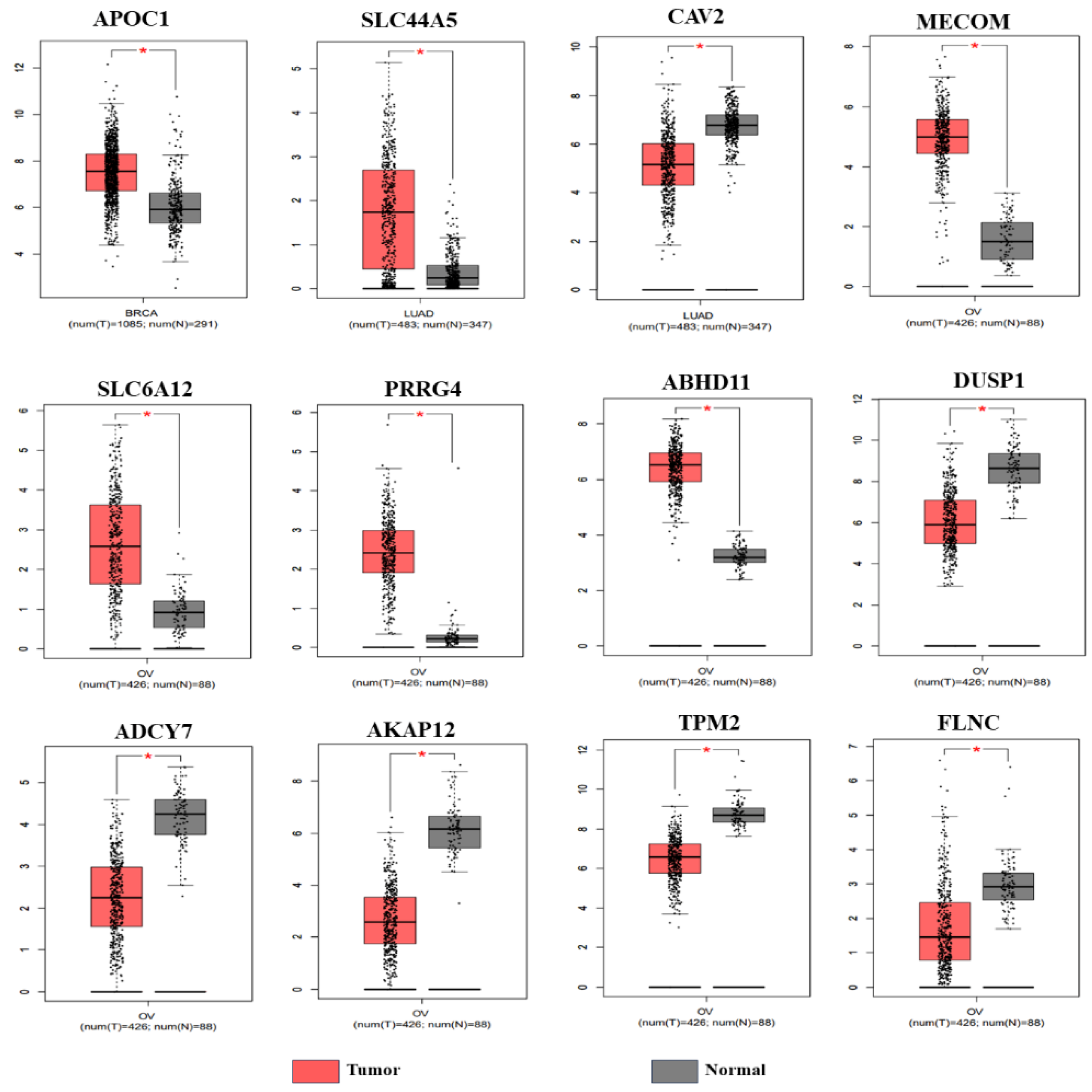
Figure 3.
Prognostic value of mRNA expression of the genes in breast, lung adenocarcinoma and ovarian cancer patients analyzed by KM plotter. A. APOC1 in Breast cancer B. SLC44A5 and C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer.
Figure 3.
Prognostic value of mRNA expression of the genes in breast, lung adenocarcinoma and ovarian cancer patients analyzed by KM plotter. A. APOC1 in Breast cancer B. SLC44A5 and C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer.
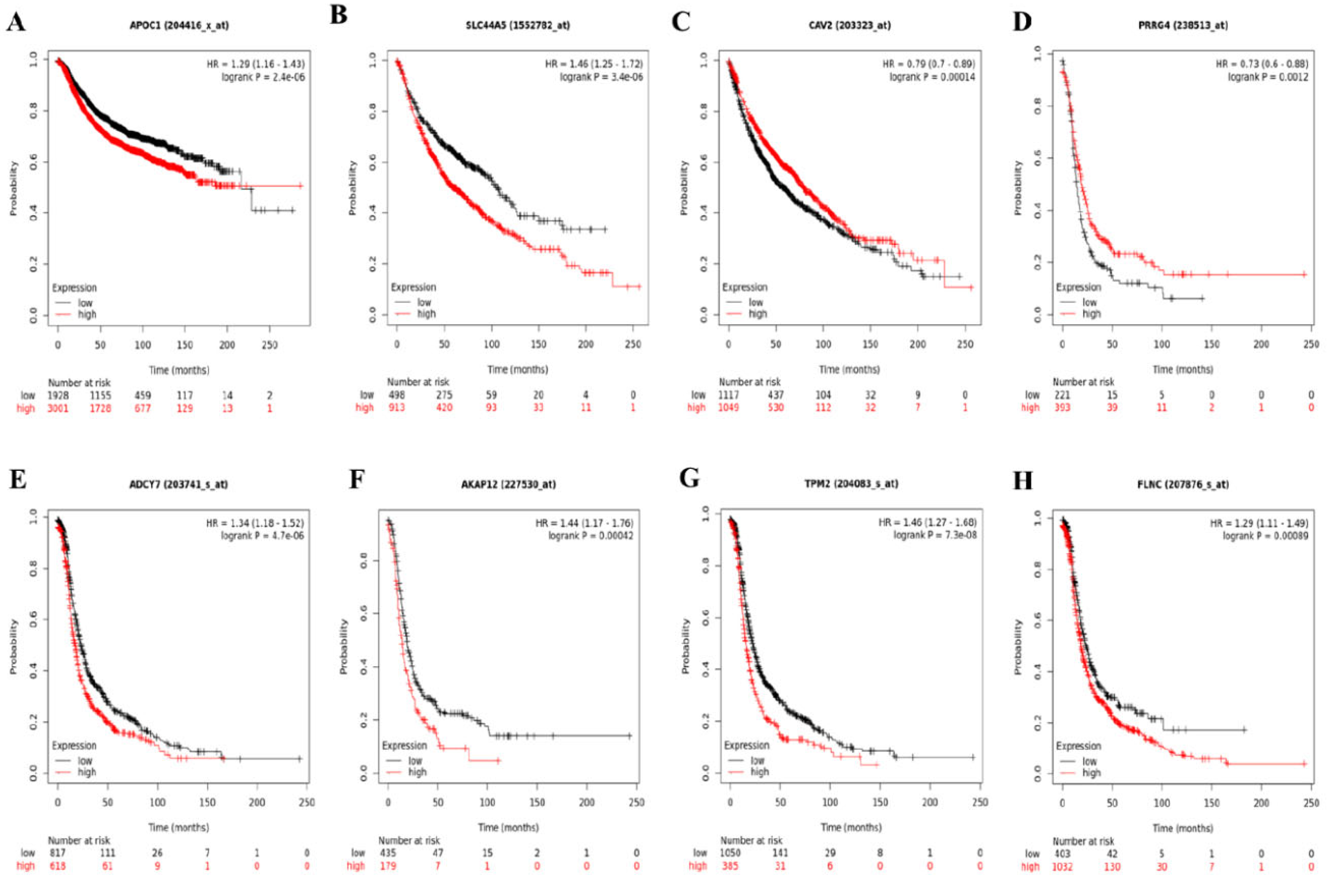
Figure 4.
Expression of the genes with the different stages of particular cancer. A. APOC1 in breast cancer B. SLC44A5 & C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer.
Figure 4.
Expression of the genes with the different stages of particular cancer. A. APOC1 in breast cancer B. SLC44A5 & C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer.
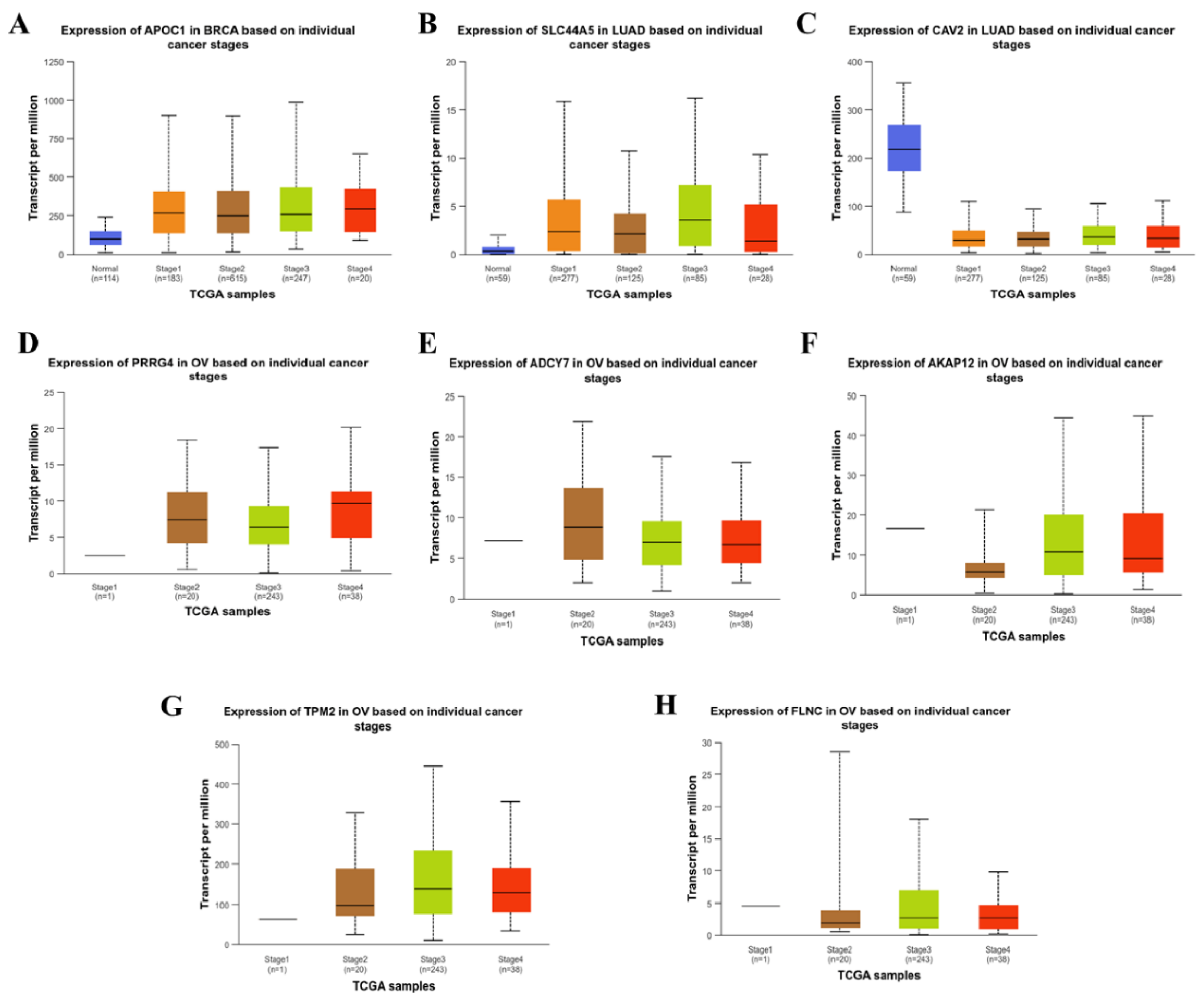
Figure 5.
Expression of the genes based on the nodal metastatic status of particular cancer patients. A. APOC1 in case of breast cancer patients B. SLC44A5 & C. CAV2 in case of Lung adenocarcinoma patients. There is no data regarding the nodal metastatic status of ovarian cancer patients.
Figure 5.
Expression of the genes based on the nodal metastatic status of particular cancer patients. A. APOC1 in case of breast cancer patients B. SLC44A5 & C. CAV2 in case of Lung adenocarcinoma patients. There is no data regarding the nodal metastatic status of ovarian cancer patients.
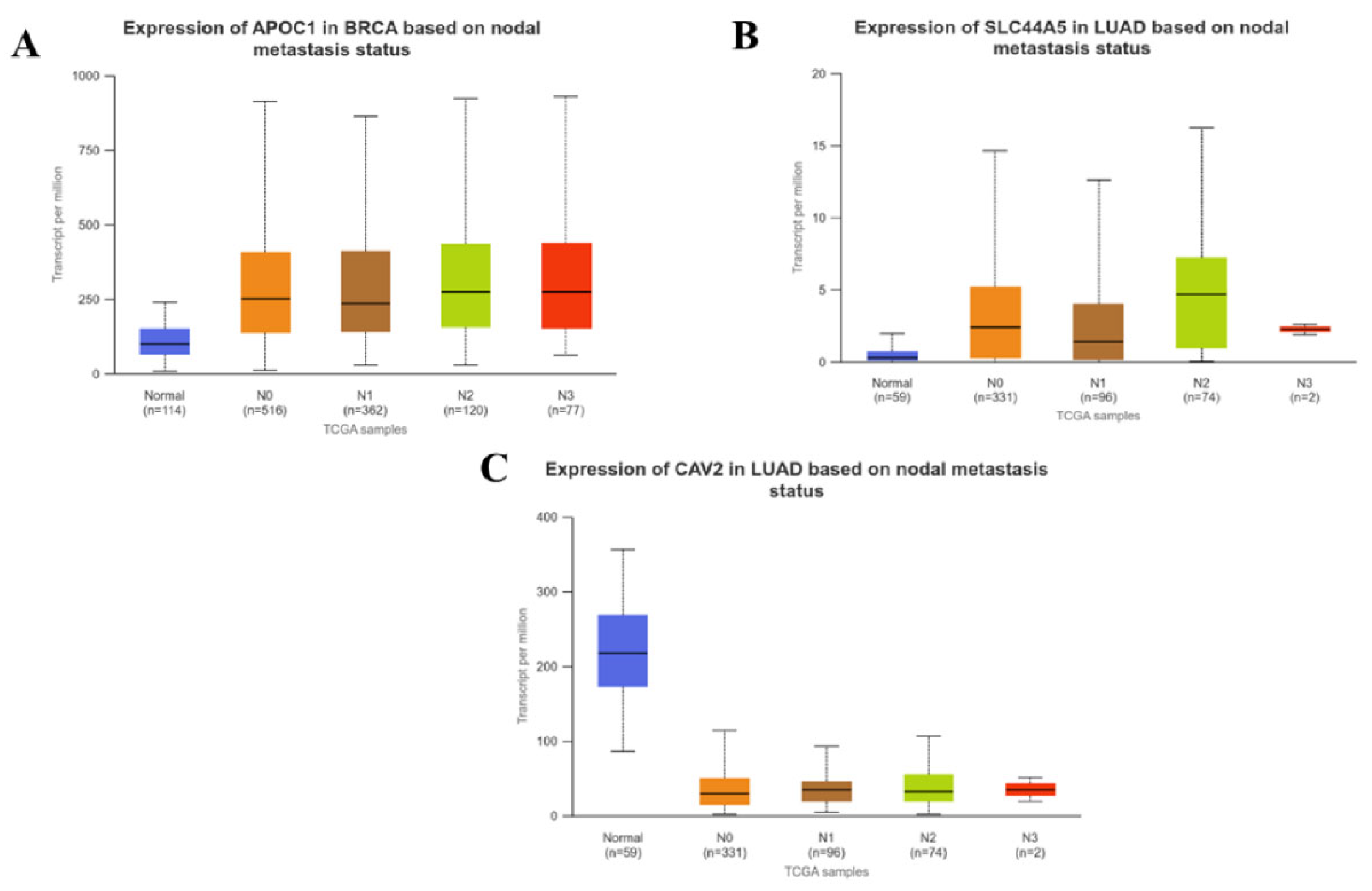
Figure 6.
Protein expression of the genes acquired from the Human Protein Atlas database. A. APOC1 in breast cancer B. SLC44A5 & C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer. Here staining is in four parts- Not detected, Low intensity, Moderate intensity and High intensity.
Figure 6.
Protein expression of the genes acquired from the Human Protein Atlas database. A. APOC1 in breast cancer B. SLC44A5 & C. CAV2 in lung adenocarcinoma D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer. Here staining is in four parts- Not detected, Low intensity, Moderate intensity and High intensity.
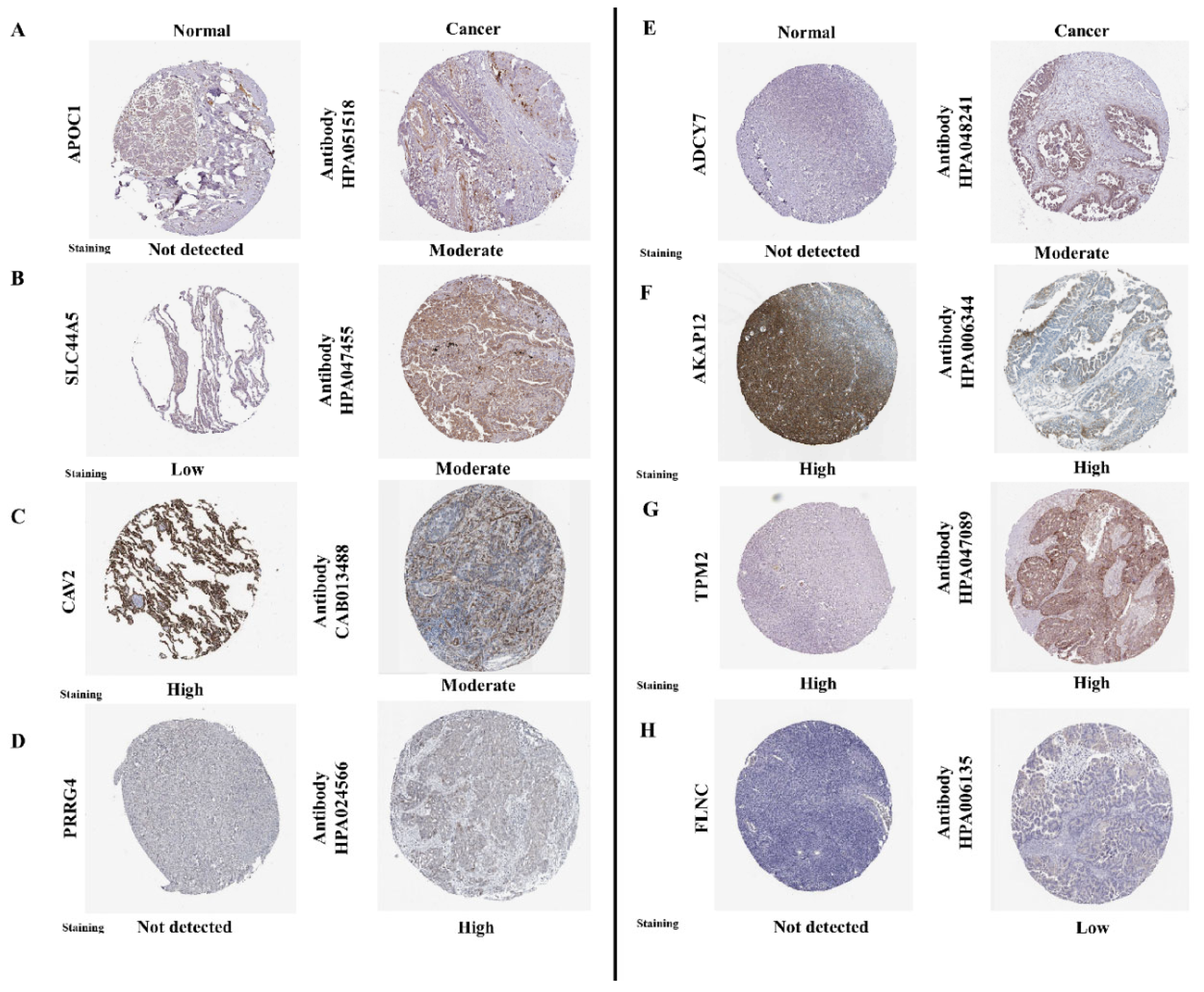
Figure 7.
Prediction of Immune cell infiltration with the expression of the genes in particular cancer. Correlation of immune cell infiltration with the expression of A. APOC1 in breast cancer B. SLC44A5 & C. CAV2 in lung adenocarcinoma and D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer.
Figure 7.
Prediction of Immune cell infiltration with the expression of the genes in particular cancer. Correlation of immune cell infiltration with the expression of A. APOC1 in breast cancer B. SLC44A5 & C. CAV2 in lung adenocarcinoma and D. PRRG4 E. ADCY7 F. AKAP12 G. TPM2 H. FLNC in ovarian cancer.
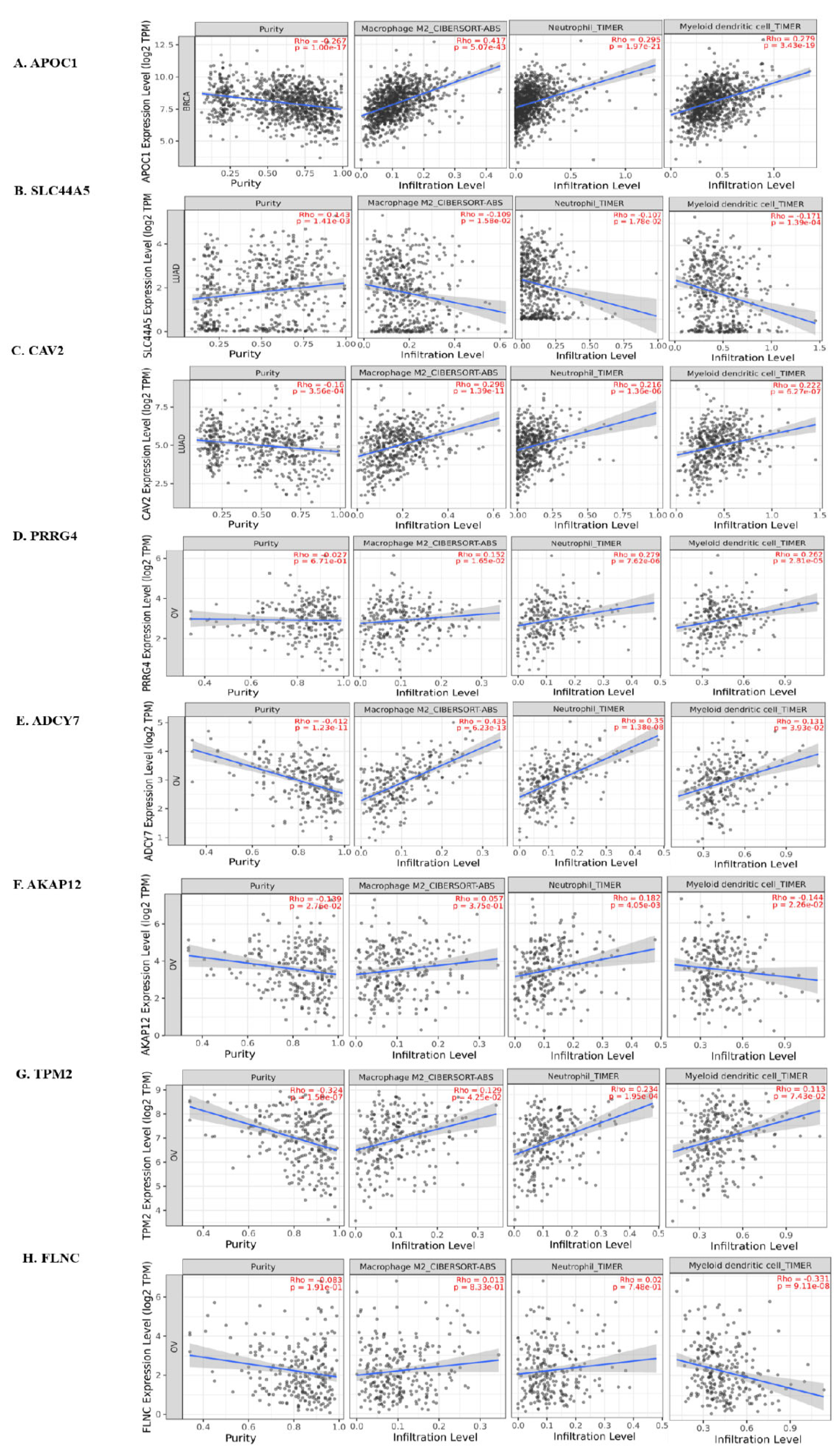
Figure 8.
Figure 8. Chemical-Gene interaction Network established with the prognostically targeting genes. A. APOC1 B. SLC44A5 C. CAV2 D. PRRG4 E. ADCY7 F. TPM2 G. AKAP12 H. FLNC.
Figure 8.
Figure 8. Chemical-Gene interaction Network established with the prognostically targeting genes. A. APOC1 B. SLC44A5 C. CAV2 D. PRRG4 E. ADCY7 F. TPM2 G. AKAP12 H. FLNC.
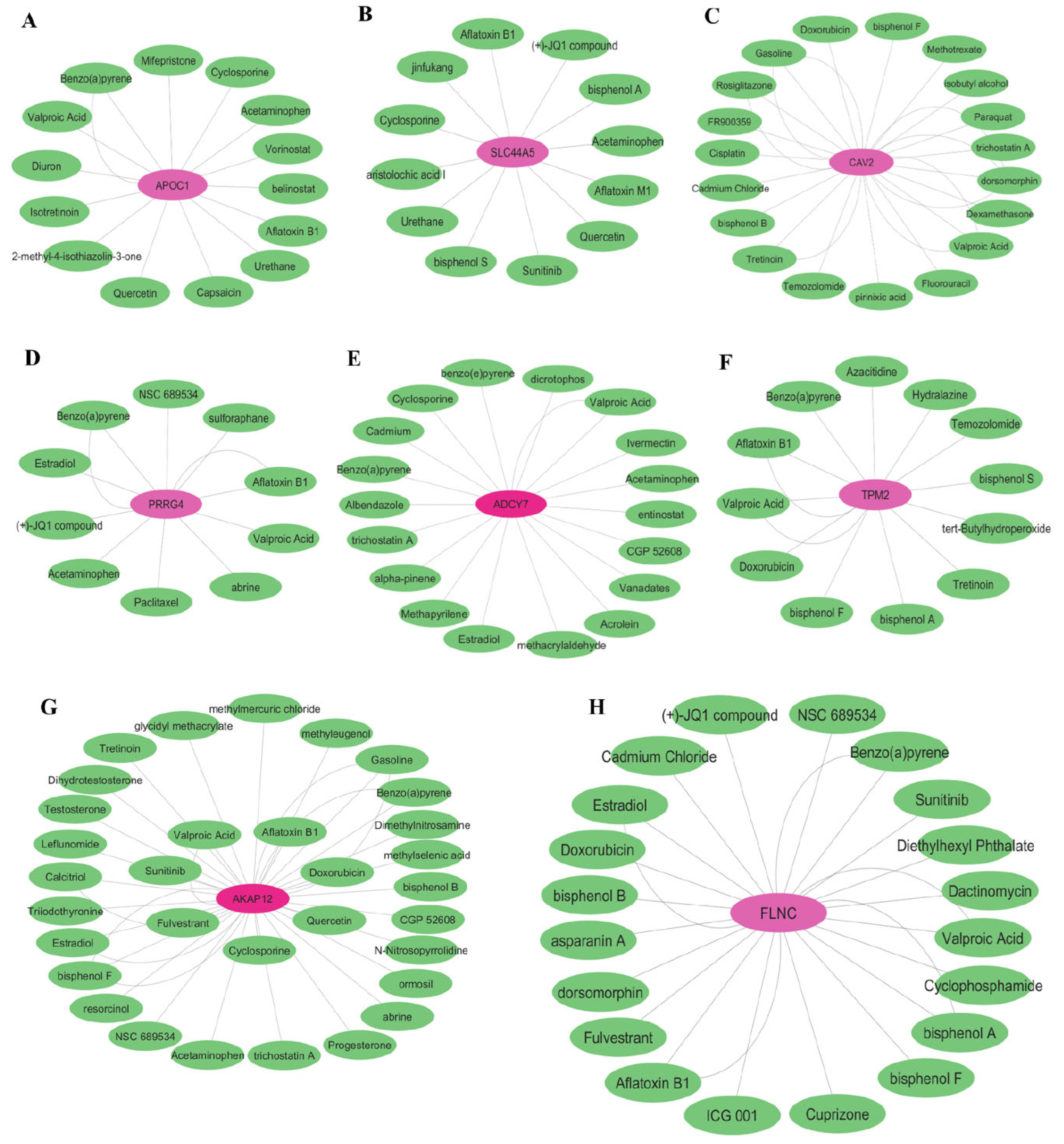

Table 1.
Datasets used in this study.
| Serial No. | Accession no. | Cancer Type | Sample groups used in this study | Platform |
|---|---|---|---|---|
| 1. | GSE7513 | Breast Cancer | Cancer Stem Cell (n=14) vs. Non-Cancer Stem Cell (n=15) | GPL570 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array |
| 2. | GSE15192 | Breast Cancer | Cancer Stem Cell (n=4) vs. Non-Cancer Stem Cell (n=4) | GPL570 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array |
| 3. | GSE136287 | Breast Cancer | Cancer Stem Cell (n=9) vs. Non-Cancer Stem Cell (n=9) | GPL6244 [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| 4. | GSE35603 | Lung Cancer | Cancer Stem Cell (n=3) vs. Parental Tumor cell (n=3) | GPL570 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array |
| 5. | GSE50627 | Lung Cancer | Normal Stem Cell (n=6) vs. Cancer Stem Cell (n=9) | GPL6244 [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| 6. | GSE160320 | Lung Cancer | Cancer Stem Cell (n=3) vs. Non-Cancer Stem Cell (n=3) | GPL26963 Agilent-085982 Arraystar human IncRNA V5 microarray |
| 7. | GSE28799 | Ovarian Cancer | Cancer Stem Cell (n=3) vs. Non-Cancer Stem Cell (n=3) | GPL570 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array |
| 8. | GSE53759 | Ovarian Cancer | Cancer Stem Cell (n=3) vs. Non-Cancer Stem Cell (n=3) | GPL6244 [HuGene-1_0-st] Affymetrix Human Gene 1.0 ST Array [transcript (gene) version] |
| 9. | GSE80373 | Ovarian Cancer | Cancer Stem Cell (n=4) vs. Non-Cancer Stem Cell (n=4) | GPL13667 [HG-U219] Affymetrix Human Genome U219 Array |
Table 2.
List of common up and down-regulated genes of three type of cancer.
| Cancer Type | Up-regulated common genes | Down-regulated common genes |
|---|---|---|
| Breast Cancer | VWA5A, LXN, CLIC4, APOC1, MCFD2. | ------- |
| Lung Cancer | ABCB7, SLC44A5, AIF1L, SYNE1, ID2, ID4, RPS6KA, PPM1D, TP53BP2, ANGPT1, RHOBT, SLCO4C1. | TRAM2, CAV2, CAP1, GLIPR1, TFPI2, PLAUR, MAN2A1 |
| Ovarian Cancer | ABCA3, DUSP4, EPHA4, ASAH1, FOXO1, MECOM, SLC6A12, PRRG4, ZDHHC14, AKR1C3, FRAS1, TSC22D1, MBD5, SLC5A3, PRDM1, ZFX, CTNS, AKR1B1, TGFBR3, AKR1C1, CAT, PKD2, C7ORF26, TMEM222, ARID5B, CCNG2, ABHD11, CDK19, PXK. | ITGA3, ANXA3, NFE2L3, GJC1, CYR61, DARS2, UGCG, DEPDC1B, DUSP1(MKP-1), BUB1, TBC1D1, ADCY7, TPX2, AURKA, AKAP12, TPM2, FLNC, PGM2, COTL1, HJURP. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



