Submitted:
01 August 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
Microsporidia are intracellular parasites infecting a wide range of vertebrates and invertebrates. In bees, they cause a diarrheal disease called nosematosis, which represents a growing problem contributing to the decline of bee colonies. This parasitic disease infects the gastrointestinal tract of bees, disrupts their immune system, weakens them, and shortens their lifespan. There are two known species of this microsporidial parasite with a global distribution: Vairimorpha apis and Vairimorpha ceranae, with current research indicating that the species V. ceranae is gradually replacing V. apis. In this study, we focused on the detection and occurrence of Vairimorpha spp. in bee colonies kept in different environments, as well as on comparing the impact of hive location on its occurrence. The methods used included sampling from winter bee carcasses, microscopic examination of bee gut contents, and subsequent molecular duplex PCR analysis. Altogether, we examined 82 hives located at 4 different sites and focused on the impact of transportation as an abiotic factor disturbing bees leading to the development of Vairimorpha spp. disease. The results indicate an increased occurrence of this parasite in hives located in urban sites and hives situated in intravillage locations, which may be attributed to transportation and associated vibrations as an abiotic stress factor. Detection revealed that all samples were positive for Vairimorpha ceranae.
Keywords:
Vairimorpha spp.
; diagnostic
; duplex PCR
; Slovakia
1. Introduction
Annual losses in bee colonies over the past few decades have been a major problem for all beekeepers. The syndrome called CCD - Colony Collapse Disorder appears to be the result of the accumulation of stressors that chronically weaken bee colonies. The immune system, metabolic pathways, and cognitive processes gradually weaken in the body under chronic stress until exhaustion and eventual failure (Chrousos 2009; Even, Devaud, Barron 2012).
Current epigenetic and nutrigenomic studies highlight the importance of interactions between microflora, climate, pathogens, toxins, nutrition (diversity and quality), and colony resilience to environmental changes (Dequenne, Philippart de Foy, Cani 2022).
Microorganisms responsible for the mortality of bee colonies include a wide range of bacteria, viruses, fungi, and protists, many of which are serious bee pathogens (Engel et al., 2016). Paenibacillus larvae and Melissococcus plutonius are highly prevalent bacterial pathogens causing American and European foulbrood (Genersch 2010; Forsgren 2010). If left untreated, they typically have a lethal outcome for the colony. In addition to bacterial pathogens, more than twenty different bee viruses have been described so far (McMenamin and Genersch, 2015), with most of them capable of causing various physiological changes (including severe physical deformities), behavioral changes, and reduced lifespan (Ryabov et al., 2023; Nguyen et al., 2024). However, individual hosts vary in susceptibility to specific infections, with many viruses persisting chronically and asymptomatically in bee colonies (De Miranda et al, 2013). The synergistic negative effects on bee health can result from combined exposure to pathogens and pesticides or concurrent infection by multiple pathogens. Many viral pathogens of bees also infect wasps or bumblebees, indicating a broader range of host species with overlapping geographic ranges (Streicher et al., 2024; Klocek et al., 2023).
Studies show that infection with microsporidia Vairimorpha spp., especially Vairimorpha apis and Vairimorpha ceranae is another important factor that contributes to the decline of the honey bee population. V. ceranae was originally discovered in Asian honey bees (Apis ceranae) and over the past few decades has spread to Western honey bee (A. mellifera) colonies worldwide (Parrella et al., 2024; Blot et al., 2023; Higes et al., 2006). V. ceranae has been found in hemolymph and various tissues, but it primarily infects the midgut of adult honey bees. By infecting individual bees in a colony, their lifespan is shortened, which results in a decrease in the colony's ability to provide and store food and thus maintain a healthy population (Alberoni et al., 2023). V. ceranae infection can not only disrupt the integrity of the midgut, but also affect energy metabolism and thus suppress the immune response of bees (Alberoni et al., 2023). Some researchers disagree that V. ceranae is more virulent than V. apis and may thus be responsible for bee colony losses (Liu et al., 2024; Li et al., 2024). They believe that the virulence of Vairimorpha spp. other factors also contribute (Zbrozek et al., 2023; Zhang et al, 2021). Thus, V. ceranae is generally considered to be the main biotic factor seriously threatening honey bees together with the ectoparasitic mite Varroa destructor. The latter parasitizes the fat body of honeybees and thus causes a shortening of the life span, a decrease in body weight and a reduced width of the acini of the hypopharyngeal glands (SD Ramsey et al., 2019; Bruckner et al., 2023). In addition, V. destructor can also transmit viruses such as deformed wing virus (DWV) (Oddie et al., 2023).
A growing body of evidence suggests that pesticides, specifically neonicotinoid insecticides, are among the main triggers of honey bee colony declines. After reviewing the risk of this group of insecticides to bee health, they were banned in Europe by the European Food Safety Authority (EFSA) (Bass and Field, 2018). However, several researchers have confirmed that bees show large differences in their sensitivity to different insecticides – including compounds belonging to the same class (Beadle et al., 2019; Iwasa et al., 2004; Manjon et al., 2018; Reid et al., 2020). For example, according to the official categories of the US Environmental Protection Agency (USEPA et al., 2014), in acute contact bioassays, honey bees (Apis mellifera) are >1000 times less sensitive to the neonicotinoid thiacloprid, while this group is classified as "virtually non-toxic". " and to the neonicotinoid imidacloprid are extremely sensitive, therefore this group is classified as "highly toxic". Also, while many pyrethroid insecticides are highly toxic to insect pollinators, tau-fluvalinate exhibits such low acute toxicity to bees that beekeepers use it as an in-hive treatment against parasitic Varroa mites (Bass et al., 2024). However, even lower doses of pesticides can have a significant impact on bee colonies. They can cause sublethal effects (e.g. reduced mobility, learning, memory, orientation, thermoregulation, foraging), which results in a decrease in the function of the entire colony and indirectly in their decline (Hester et al., 2023; Raine and Rundlöf 2024).
Currently, climate change is not only a threat to us humans, but also worries bees all over the world (Van Espen et al., 2023). Increased temperature brings many negative effects on food reserves (including honey production, increase in reserves for colonies and the composition of sugars in honey, it also affects interactions between plants and pollinators by reducing their visitation and the strength of interactions, it even causes colony mortality (bees die earlier, they are less resistant against winter, as their thermal tolerance decreases, they suffer from heat fluctuations) (Zapata-Hernández et al., 2024; Palmer-Young et al., 2023; Glass and Harrison, 2024; Manlik et al., 2023). A decrease in temperatures can cause increased mortality of colonies. Longer exposure to reduced temperatures affects their immunity and microbiome (Butolo et al., 2022). An increase in precipitation, which had a negative impact mainly on the hive's homeostasis (its internal temperature and relative humidity of the breeding ground and the forage area), food supply (the richness and diversity of pollen decreases, there is an increase in the weight of the bee colony and the average annual yield, there are also changes in the microbiome and a decrease in the interaction between plants and pollinators (Switanek et al., 2017; Abou-Shaara et al., 2017). A decrease in precipitation may result in reduced honey production and also reduce interactions between plants and pollinators (Jaworski et al., 2022). Changes in precipitation are also a starter for the reproduction and distribution of various pests, the emergence of diseases and changes in the geographical distribution of different lines of A. mellifera (Abou-Shaara and Darwish, 2021). However, the opposite view is held by Langowska et al., 2017, who argue that rising temperatures could be beneficial for bees, as they would extend spring foraging periods and thus be better prepared to take advantage of summer nectar and pollen flows.
Climate change and habitat fragmentation result in the inability of bees to gather enough food for their needs. Colonies need pollen to grow and reproduce, so a severe lack of food can cause a colony to fail drastically. This failure includes traits suggestive of social breakdown, including cannibalism and looting of resources from weaker colonies, as well as reduced brood care (Corby-Harris et al., 2022; Tong et al., 2019; Corby-Harris et al., 2022). The nutritional health of bees depends on the availability and collection of nectar, pollen, as well as their quantity and quality to meet the main nutritional requirements of carbohydrates, proteins, lipids, vitamins and minerals (Castaños et al., 2023). Mayack and Naug (2009) demonstrated a correlation between bees infected with the protozoan Vairimorpha ceranae and a higher level of hunger. Thus, these infected bees experience nutritional stress, which is a consequence of many pathogenic infections. Based on the feeling of great hunger, the bees are more inclined to go out of the hive for food. However, if bees infected in this way go in search of food in low-energy states, it is more likely that they will die during this act and will not return to the hive. The foraging behavior of honey bees is largely regulated by the nutritional status of individuals, independent of the amount of stored food in the colony. The nutritional and energetic stress of individuals due to infection thus provides a possible mechanism for the disappearance of bees from hives that still have intact honey reserves. If nutritional stress on individuals is a major contributor to colony loss, its effects are likely to be felt most severely at times and places where it is difficult for bees to find suitable food (Naug 2009).
On a global level, the extensive use of plastics has contributed to the production of a huge amount of hazardous waste and thus represents one of the most significant problems of today. Ubiquitous microplastics represent a potential threat to various species of plants and animals, therefore it also has an impact on bee colonies (Lin et al., 2024), specifically in the form of a decrease in the diversity of the intestinal microbiome, changes in the structure of the microbiome and changes in the expression of genes related to detoxification and immunity (Wang et al. al., 2021b; Balzani et al., 2022; Buteler et al., 2022). Also, atmospheric PM, primarily emitted by automobile traffic, combustion engines, but also coal mining and, last but not least, agricultural residues, affect worker bee navigation, prolong the duration of foraging and affect bee survival (Thimmegowda et al. 2020; Cho et al. 2021). Heavy metals such as copper, lead, or manganese also have a lethal or sublethal effect on bees (Al Naggar et al. 2020). The activity of pollinators was also largely influenced by noise, turbulence and dust on roadsides (Dargas et al. 2016; Phillips et al. 2021).
Many microorganisms are contained in various parts of larvae and adult bees, their food and plastic, and they play an important role in food digestion, pollination and antagonistic effect against various pathogens. In this context, bees are mainly associated with fungi, which can provide material for pollen degradation or help the maturation of royal jelly, while they can also be a source of food (Cui et al., 2022; Khan et al., 2020). Honey bees are therefore a potential model of fungus-host-symbiont interactions. Fungi are numerous in bee bread and nectar, but also from healthy honey bees Gilliam et al. (1979) isolated four fungal species, with Candida species detected predominantly from bees fed herbicide and antibiotics. Research in nosemosis field tests has revealed that the degree of Vairimorpha ceranae infection is related to yeast growth in the honey bee gut, suggesting that exposure to stresses such as infection can lead to structural changes in the gut microbial community (Yun et al., 2018). In this study, we aimed to compare the survivability of honey bees in different environments after infection with Vairimorpha species, taking into account different stressful factors.
A rodent in the hive can also be such a stressful factor. In picture no. 1 we see a shrew, which moved into the hive during the winter. During the spring inspection of bee colonies, we found a non-terrestrial infection in most of the bees, until finally this bee colony fell completely. So we think that this rodent caused stress in the bees, which culminated to a lethal end.

Image no. 1.
The shrew at the bottom of the hive.
2. Material and Methods
2.1. Collection of Samples
In the month of March 2024, we took samples of winter carcasses of bees (Fig. 2) in the number of 30 pcs per bee colony from several locations where bee colonies were located. According to the type of biotope, we divided them into the following categories: A) urban biotope, B) rural biotope in the intravillage, C) rural biotope in the extravillage and D) forest biotope (Fig. 3 and Tab. 1):
2.1.1. Location no. 1: Municipal Bees (Košice - altitude 206 masl, Košice- Barca - altitude 210 masl, Michalovce - Altitude 113 masl):
All urban bee habitats are located between houses or blocks, within 5 meters of a busy road. Within 3 kilometers from Košice and 2 kilometers from Košice-Barca, as the crow flies, the largest eastern Slovakian plant U.S. is located. Steel Košice, producing mainly steel and sheets; at a distance of max. 1 km from both locations is the thermal power plant TEKO Košice. The company KOSIT, dedicated to the collection and processing of municipal waste, has its headquarters in the incinerator directly in the Košice-Barca area, as well as the industrial park Immopark, which produces electronic and mechanical components for the automotive industry. The company CASSPOS a.s. in Michalovce is focused on production various steel products and generally focuses on aluminum processing. Within this site there are still several smaller industrial plants, we have listed only the largest and closest ones.
Food for bees is provided in the form of various types of flowers found in city parks. Also, people living in houses or apartments grow garden crops, fruit trees, balcony flowers (but most of them are sprayed and fertilized).
2.1.2. Location no. 2: Inner Village (Lipníky - Altitude 280 masl)
Site located in the immediate vicinity of road 1: class, where there is heavy traffic. The nearest industrial areas are 14 km away (ZEOCEM Bystré) - processing of clinoptilolite and 15 km to Prešov, where there are several plants, the largest of which is a clothing plant and an automobile plant. This area is characterized by agricultural activity, where rapeseed (which is sprayed) is grown for 3 km as the crow flies. Another option for bee food is gardens where people grow fruits, vegetables and ornamental plants. This is a locality with seasonal use of the laying conditions of the site, mainly in the first half of the beekeeping season (March - June of the calendar year) for obtaining mainly light (spring) types of honey, honey from fruit trees such as willow and also goldenrod honey.
2.1.3. Location no. 3: Extravillain of the Village (Okrúhle- Altitude 280 masl)
The beehives are located in the garden behind the house, 500 m away from the 1st class road (with heavy traffic). The nearest plant is approx. 30 km from ZEOCEM Bystré and 50 km from Bukóza Vranov nad Topľou, approx. 30 km from Bardejov - textile industry. Agricultural crops are grown in the surroundings: organic peas, wheat (but bees do not pollinate them); in addition, only fruit trees and horticultural crops. It is a permanent habitat of bee colonies in the cadastre of Okrúhle village, district Svidník, in the area of the Ondavská vrchovina, in close proximity to the Forest Park called "Kerta v Okrúhlom". This location is mainly suitable for obtaining forest and mixed honeys. Picturesque forests alternate with mountain meadows and pastures. The principles of ecological agriculture are applied in this area, which strictly exclude the use of any artificial fertilizers or other agrochemicals. The phytodiversity of this park is very rich (40-50 species of trees): for example, Acer platanoides, Fraxinus excelsior, Tilia cordata, Tilia plathyphyllos, Quercus petraea, Quercus robur, Picea abies, Pinus silvestris and Larix decidua.
2.1.4. Location no. 4: Forest Bees (Komárnik- Altitude 420 masl)
It is a site located in the immediate vicinity of the forest, 2 km as the crow flies from a 1st class road (with heavy traffic). It is a locality of the Eastern Carpathians Protected Landscape Area, Dukla Forest Community of European Importance. 80% of the area consists of forest communities of the Eastern Carpathians, represented mainly by fir, beech and valuable deciduous trees. The nearest plant is over 100 km away. Agriculture in the area is not widespread, only hay is cut. Bees only use the surrounding leafy conifers and meadow flowers for laying eggs. In connection with the climate, the temperature is lower than in cities and more constant without higher fluctuations. The health status of bee colonies is checked annually by the relevant Regional Veterinary and Food Administration of Vranov and Prešov by taking samples of honeydew, which are evaluated by the State Veterinary and Food Institute in Dolní Kubín. It is a temporary habitat for bee colonies located in the region under Duklau, in the cadastre of the municipality of Nižný Komárnik, district Svidnik. This location is specific for its microclimatic conditions. It is part of the Eastern Carpathian Protected Landscape Area and at the same time the Dukla Area of European Importance. The dominant plant communities in the area are beech-fir flowering forests with a high proportion of white fir (Abies alba). The National Nature Reserve (hereinafter referred to as "NPR") Komárnická jedlina is located in close proximity to the site. These are intact fir-beech forests with admixtures of rare trees such as maples, elms, ash trees and linden trees. This NPR is classified as a European speciality. The territory was declared for the protection of biotopes of European importance, rare preserved relics of the natural fir-beech forests of the Eastern Carpathians with a very significant thickness and age differentiation. The 5th degree of nature protection with a continuous no-intervention regime applies here. This rare reserve of national importance is important for ensuring species diversity, ecological stability of the region and, last but not least, for the possibility of observing natural evolutionary processes in nature. This site is located in the outskirts of the village of Nižný Komárnik, 2 km from the international road. The radius of action of the bee colonies at this site is limited to 1.5 km due to the ruggedness of the relief. 80% of the area in the range consists of forest communities and the remaining 20% are meadow biotopes and riparian vegetation of the local stream "Šivárna." The territory is also sporadically visited by the brown bear (Ursus arctos) and therefore the bees must be protected by electric protection and fencing in accordance with § 45 par. 1 letter b) c) d) Decree of the Ministry of the Environment of the Slovak Republic No. 170/2021 of April 19, 2021, implementing Act No. 543/2002 Coll. on nature and landscape protection, as amended. No. 170/2021 Coll.
In all beehives, we collected bee carcasses from the bottom of the hive, at the same time we observed and evaluated the strength of infection of the beehives, according to the number of occupied frames, by checking the strips of grind on the underlays.


Image no. 2.
Dead bees at the bottom of the hive.
Image no. 3.
Observed habitats: A) Urban habitat; B) Rural site; intravilan; C) Rural location of extravillans; D) Forest habitat.
Image no. 3.
Observed habitats: A) Urban habitat; B) Rural site; intravilan; C) Rural location of extravillans; D) Forest habitat.

Table 1.
Number of bee colonies per individual site.
| Habitat according to the location of bee colonies | Numbers of bee colonies |
|---|---|
| A) City bees | 22 |
| B) Rural bees in the inner city | 27 |
| C) Rural bees in extravilla | 22 |
| D) Forest habitat | 11 |
2.2. Sample Processing and Detection of Vairimorpha spp.
2.2.1. Microscopic Diagnostics
We cut the abdomens from the bee carcass samples and mixed them in a mortar with distilled water, then crushed the abdomens. We followed the Pohl 2005 methodology.
We observed the samples under a microscope at 40x magnification and counted the number of spores in the specimen. We counted spores in 5 fields of view and after averaging, we evaluated the strength of the infection according to Table no. 2
Table 2.
The strength of the Nozemosis infection in relation to the number of spores.
| Strength of infection | Number of spores |
|---|---|
| Negative sample | 0 in the field of view |
| Weak nozem infection + | 1-19 spores per field of view |
| Moderately strong nozem infection ++ | 20-100 spores per field of view |
| Strong nozem infection +++ | Over 100 spores per field of view |
2.2.2. Molecular Diagnostics
We isolated DNA from crushed bee abdomen samples using the commercially available DNA -sorb- AM isolation kit from AmpliSense and following the manufacturer's instructions. We used a PRECELLYS tissue homogenizer with a program of 6500 rpm for 2x 45 seconds to break the hard shell of the spores.
In the next step, we created a PCR mix for each sample, which consisted of PCR water, master mix and 10 µM mediums for both species of Vairimorpha spp. (table no. 3). We added the template to the mix and vortexed thoroughly.
Table 3.
Components of the PCR mix and their amounts.
| Components of the PCR mix | Quantity |
|---|---|
| PCR water | 11,5 µl |
| Firepol Master Mix (Solis Biodine) | 4 µl |
| APIS FOR(5′-GGGGCCATGTGTTTGACGTACTATGTA-3′) | 0,5 µl |
| APIS REV (5′-GGGGGGCGTTTAAAAATGTGAAACAACTATG-3′) | 0,5 µl |
| MITOC FOR (5′CGGCGACGATGATGATGATGAAAATATTAA-3′) | 0,5 µl |
| MITOC REV (5′-CCCGGTCATTCTCAAAAAAAACCG-3´) | 0,5 µl |
| Template | 2,5 µl |
Duplex PCR was performed using a VWR RISTRETTO thermal cycler. The initial denaturation took place for 4 min. at 95 °C, followed by 28 cycles of denaturation at 95 °C (25 s), annealing at 58 °C (45 s) and polymerization at 72 °C (2 min). The final extension step lasted 7 min. at 72 ° C. To prove the presence of DNA in the investigated samples, we used gel electrophoresis, using a 1.5% agarose gel. After evaluating the results with a UV transilluminator, the DNA concentration was measured with a NanoDrop and the PCR products were sent for sequencing. The obtained sequences were compared with the sequences stored in the gene bank using the BLAST program.
3. Results
3.1. Microscopic Diagnostics
Using a light microscope with 400x magnification, we detected the presence or absence of spores of Vairimorpha spp. in bee colonies. In 41 (50%) samples we detected spores of Vairimorpha spp. and in 41 (50%) samples there were no spores found (Fig. no. 5 and Tab. no. 4)
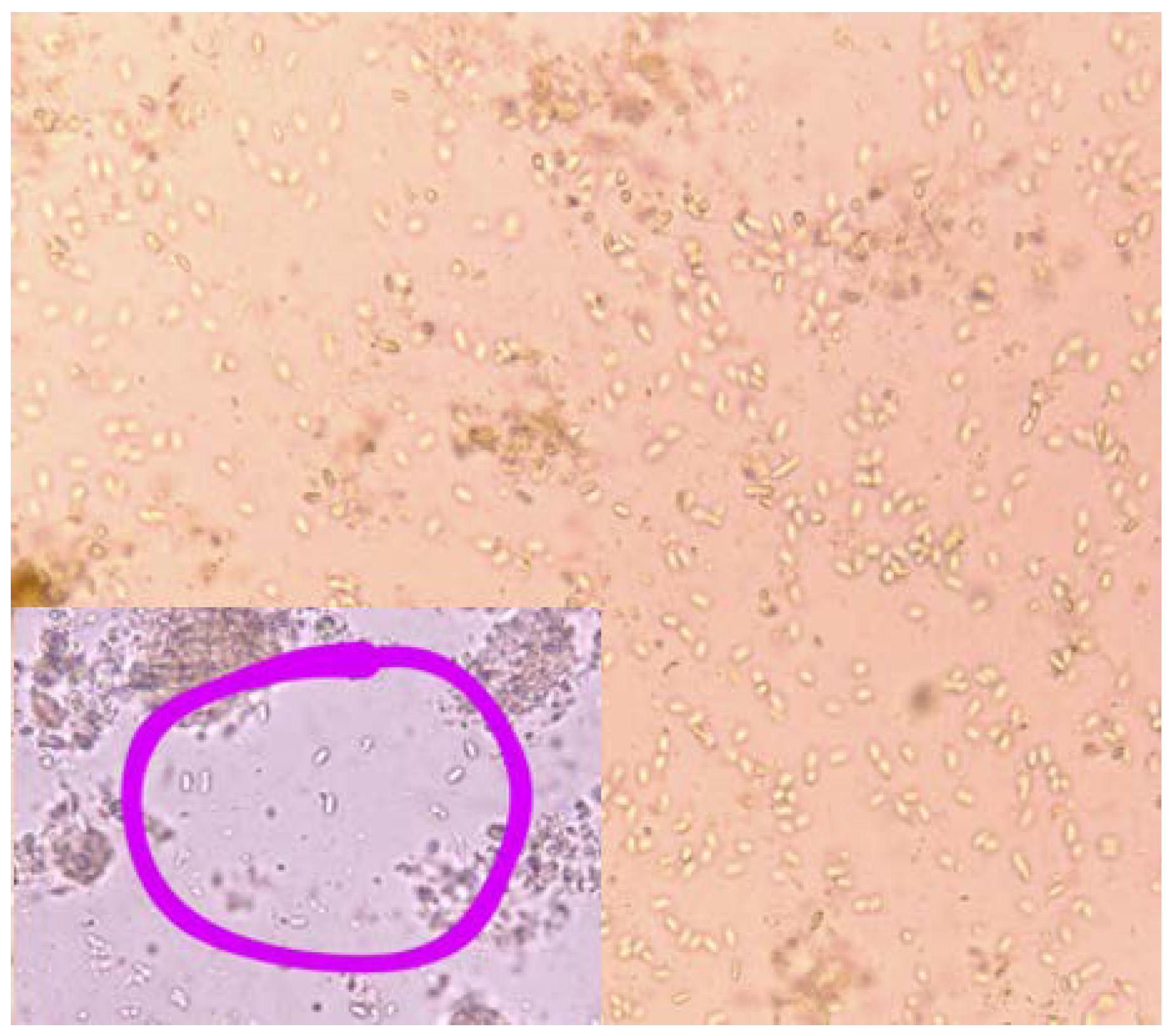
Image no. 5.
Spores under the microscope at 400x magnification.
Table 4.
Evaluation of negative and positive samples with differentiation of infection strength.
| negative | + | ++ | +++ | Positive | |
|---|---|---|---|---|---|
| City | 5 | 5 | 3 | 9 | 17/22 |
| Inner city | 7 | 6 | 6 | 8 | 20/27 |
| Extravilla | 18 | 0 | 0 | 4 | 4/22 |
| Forest | 11 | 0 | 0 | 0 | 0/11 |
| Total count | 41 | 11 | 9 | 21 | 41/82 |
3.2. Molecular Diagnostics
The results of the microscopic method correlated with the results of molecular diagnostics, except for one case of a sample taken from a city district, where we evaluated one sample microscopically as negative, and after examination with molecular methods, we found it positive. In all positive samples for the detection of Vairimorpha spp., which underwent DNA isolation and subsequent amplification with the help of primers for both species Vairimorpha apis (amplicon 321 bp) and Vairimorpha ceranae (amplicon 218 bp) after loading on a 1.5% agarose gel and subsequent visualization the species Vairimorpha ceranae was detected using a UV transilluminator.
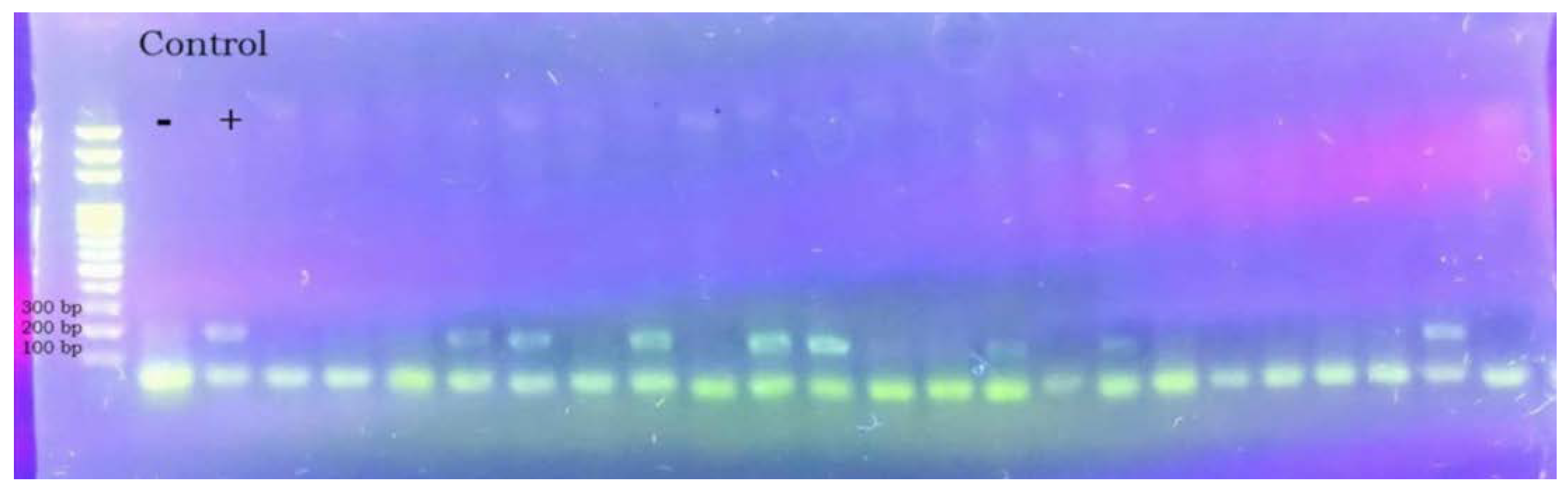
Image no. 6.
Samples positive for the disease Vairimorpha ceranae (amplicon 218 bp).
3.3. Proportion of Positive Cases with Respect to Habitat
After examination of urban bees, 18 (81.81%) samples were positive and 4 (18.19%) were negative. Of the rural bees located at the site in the inner city, 20 (74%) samples were positive and 7 (16%) were negative. Of the samples of rural bees located at the site in the extravillan, 4 (18.18%) were positive and 18 (81.82%) were negative. Of the 11 samples of bees located in the forest habitat, all bee colonies (100%) were free of Vairimorpha spp. (see table no. 5 and picture no. 7)
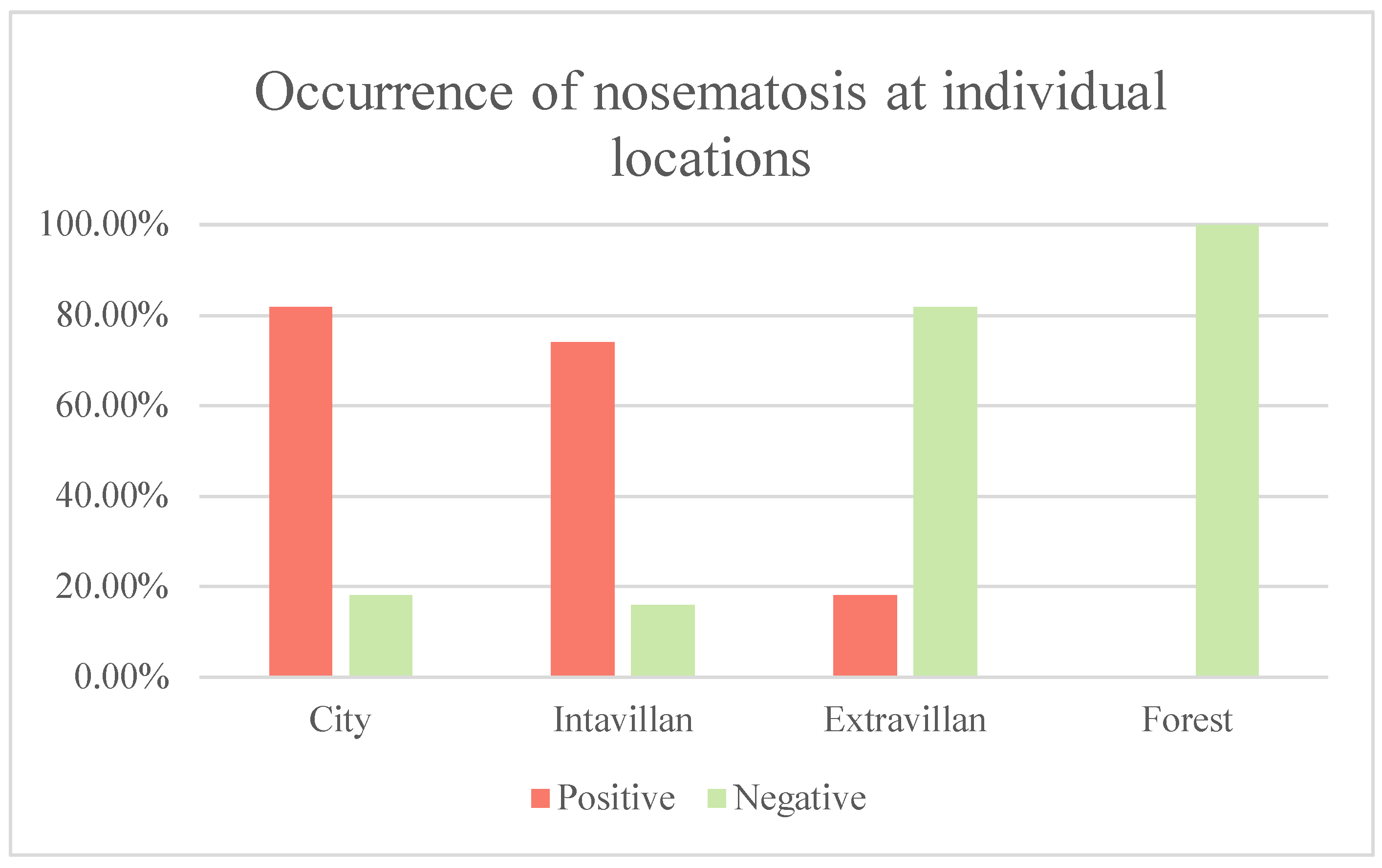
Table 5.
Proportion of positivity and negativity of bee colonies on Vairimorpha spp. to a specific location.
Table 5.
Proportion of positivity and negativity of bee colonies on Vairimorpha spp. to a specific location.
| Location | Number of positive colonies | Number of negative colonies |
|---|---|---|
| City bees | 18 | 4 |
| Rural bees - intravillan | 20 | 7 |
| Rural bees - extravillan | 4 | 18 |
| Forest bees | 0 | 11 |
Image no. 7.
Graph of the incidence of nosematosis at individual sites.
4. Discussion
On the infection of bees with Vairimorpha spp. several factors are involved, which must be followed in order to preserve a large bee colony. The most important prevention against this bee disease includes suitable environmental temperature, adequate number of bees in the hives, selection of habitat for wintering honey bees enabling earlier and more frequent flying, which would prevent overcrowding of the hive environment and the spread of infection at the same time. Replenishment of winter supplies, hygienic feeding and watering, regular disinfection of bee colonies, filtration, burning of dead individuals and sanitation are also important (Galajda et al., 2021). Failure to comply with the conditions for proper beekeeping can lead to bacterial or viral infection, which are other important factors in the reduction of the bee population (Genersch 2010; Forsgren 2010; Ryabov et al., 2023; Nguyen et al., 2024). Even today, very popular pesticides are an important factor affecting the health of bees (Hester et al., 2023; Raine and Rundlöf 2024). In our study, we therefore focused on comparing the occurrence of nosematosis in bees occurring in different habitats. As the results show, the infection most often occurred in bees kept in cities (81.81%), followed by the intravillage area of the village (74%), extravillage area (18.18%), and we detected no bees positive for the Vairimorpha species in the forest habitat. It can therefore be concluded that the urban environment was the least suitable for bee colonies due to the highest level of stress factors influencing the occurrence of nosematosis. Several researchers also dealt with the issue of comparing urban and rural environments. It has been concluded that urban environments are suitable in terms of flower diversity and higher temperature (Prendergast et al., 2022), while the situation of bees in the countryside is correlated with the city (Amado De Santis and Chacoff, 2020). That urban areas are probably more suitable for bees than agricultural areas was proven in their work by Geslin et al. (2013), who noted a large difference between food supply diversity and bee richness. Mahé et al. (2021) in their study found a higher concentration of insecticides in urban foragers and a higher content of metals in their larvae compared to rural bees. Also, bees living near the main Italian highway Autostrada A1 showed contamination with nanoscale Fe oxides/hydroxides and barite, as well as collected pollen and honey produced by the bee colony (Papa et al., 2021). Another study (Taylor et al., 2023) investigating trace element contamination in honey bees inhabiting urban areas around the largest and longest operating nickel smelter in the South Pacific at Nouméa, New Caledonia reported elevated concentrations of potentially toxic trace elements including cobalt, chromium and nickel in bees. Concentrations of these metals decreased with distance from smelting operations, suggesting a relationship between environmental contamination and uptake of trace elements by bees. The bee colonies studied by us were influenced not only by industrial centers, but also by climate, pesticides, food options, and last but not least, vibrations and shocks caused by road traffic. With this, we would like to appeal to beekeepers and the scientific community to pay more attention to this issue and to investigate road traffic as a stress factor affecting the health of bee colonies.
5. Conclusions
Based on the global decline of pollinators and insects in general (Potts et al. 2016; McDermott 2021), the health of bees has long been the subject of debate in both the beekeeping and scientific communities (Hassler et al. 2021). Productive beekeeping requires that bee colonies have enough pasture from spring to fall. Regular migration significantly supports the development of the bee colony, improves its health and the supply of bee colonies. Many studies deal with various biotic and abiotic stressors affecting bee colonies. However, based on our results, we believe that further research would be needed to investigate other potential factors that may plausibly influence bee physiology and thus lead to the spread of Vairimorpha spp. We think that the impact of road traffic, especially in terms of vibrations and shocks, is generally under-researched so far. However, in general, the combined effect of multiple stressors is certainly more harmful than a single stressor (Goulson et al. 2015; Goulson and Nicholls 2022). It can be argued that the interaction between multiple stressors that vary spatially and temporally is a key factor underlying the issue of global honey bee health.
Funding
This paper was created with the support of the grant projects GP MŠVVaŠ SR VEGA no. 1/0161/23, and APVV-21-0185. .
References
- Abou-Shaara, H.F., Darwish, A.A.E. (2021). Expected prevalence of the facultative parasitoid Megaselia scalaris of honey bees in Africa and the Mediterranean region under climate change conditions. Int J Trop Insect Sci, 41, 3137–3145. [CrossRef]
- Abou-Shaara, H.F., Owayss, A.A., Ibrahim, Y.Y., et al. (2017). A review of impacts of temperature and relative humidity on various activities of honey bees. Insect. Soc., 64, 455–463. [CrossRef]
- Alberoni, D., Di Gioia, D., Baffoni, L. (2023). Alterations in the Microbiota of Caged Honeybees in the Presence of Nosema ceranae Infection and Related Changes in Functionality. Microb Ecol, 86(1), 601-616. [CrossRef]
- Al Naggar Y, Dabour K, Masry S, Sadek A, Naiem E, Giesy JP. Sublethal effects of chronic exposure to CdO or PbO nanoparticles or their binary mixture on the honey bee (Apis millefera L.). Environ Sci Pollut Res Int. 2020 Jun;27(16):19004-19015. Epub 2018 Oct 3. [CrossRef] [PubMed]
- Amado De Santis, A.A., Chacoff, N.P. (2020). Urbanization Affects Composition but Not Richness of Flower Visitors in the Yungas of Argentina. Neotrop Entomol, 49, 568–577. [CrossRef]
- Balzani P, Galeotti G, Scheggi S, Masoni A, Santini G, Baracchi D. Acute and chronic ingestion of polyethylene (PE) microplastics has mild effects on honey bee health and cognition. Environ Pollut. 2022 Jul 15;305:119318. Epub 2022 Apr 18. [CrossRef] [PubMed]
- Bass, C., & Field, L. M. (2018). Neonicotinoids. Current biology, 28(14), R772-R773. [CrossRef]
- Bass, Chris, Hayward, Angela, Troczka, Bartlomiej J., Haas, Julian, Nauen, Ralf. (2024). The molecular determinants of pesticide sensitivity in bee pollinators, Science of The Total Environment, Volume 915, 170174, ISSN 0048-9697. [CrossRef]
- Beadle, K., Singh, K. S., Troczka, B. J., Randall, E., Zaworra, M., Zimmer, C. T.,... & Bass, Blot, N., Clémencet, J., Jourda, C., Lefeuvre, P., Warrit, N., Esnault, O., Delatte, H. (2023). Geographic population structure of the honeybee microsporidian parasite Vairimorpha (Nosema) ceranae in the South West Indian Ocean. Sci Rep, 13(1), 12122. [CrossRef]
- Bruckner, S., Straub, L., Neumann, P., Williams, G.R. (2023). Negative but antagonistic effects of neonicotinoid insecticides and ectoparasitic mites Varroa destructor on Apis mellifera honey bee food glands. Chemosphere, 313, 137535. [CrossRef]
- Buteler M, Alma AM, Stadler T, Gingold AC, Manattini MC, Lozada M. Acute toxicity of microplastic fibers to honeybees and effects on foraging behavior. Sci Total Environ. 2022 May 20;822:153320. Epub 2022 Jan 22. [CrossRef] [PubMed]
- Butolo, N.P., Azevedo, P., Alencar, L.D., Malaspina, O., Nocelli, R.C.F. (2021). Impact of low temperatures on the immune system of honeybees. Journal of Thermal Biology, 101, 103082. [CrossRef]
- Castelli, L., Branchiccela, B., Romero, H., et al. (2022). Seasonal Dynamics of the Honey Bee Gut Microbiota in Colonies Under Subtropical Climate. Microb Ecol, 83, 492–500. [CrossRef]
- Castaños, C.E., Boyce, M.C., Bates, T., Millar, A.H., Flematti, G., Lawler, N.G., et al. (2023). Lipidomic features of honey bee and colony health during limited supplementary feeding. Insect Molecular Biology, 32(6), 658–675. [CrossRef]
- Cho Y, Jeong S, Lee D, Kim SW, Park RJ, Gibson L, Zheng C, Park CR. Foraging trip duration of honeybee increases during a poor air quality episode and the increase persists thereafter. Ecol Evol. 2021 Jan 23;11(4):1492-1500. [CrossRef] [PubMed] [PubMed Central]
- Chrousos, G.P. (2009). Stress and disorders of the stress system. Nat Rev Endocrinol, 5, 374–381. [CrossRef]
- Corby-Harris, Vanessa, Snyder, Lucy, Meador, Charlotte, Watkins-DeJong, Emily, Obernesser, Bethany T., Brown, Nicholas, Carroll, Mark J. (2022). Diet and pheromones interact to shape honey bee (Apis mellifera) worker physiology, Journal of Insect Physiology, Volume 143, 104442, ISSN 0022-1910. [CrossRef]
- Cui, P., Kong, K., Yao, Y., Huang, Z., Shi, S., Liu, P., Huang, Y., Abbas, N., Yu, L., Zhang, Y. (2022). Community composition, bacterial symbionts, antibacterial and antioxidant activities of honeybee-associated fungi. BMC Microbiol, 22(1), 168. [CrossRef]
- Dargas JH, Chaves SR, Fischer E. Pollination of lark daisy on roadsides declines as traffic speed increases along an Amazonian highway. Plant Biol (Stuttg). 2016 May;18(3):542-4. Epub 2016 Feb 21. [CrossRef] [PubMed]
- De Miranda, J.R., Bailey, L., Ball, B.V., Blanchard, P., Budge, G.E., Chejanovsky, N., Chen, Y., Gauthier, L., Genersch, E., de Graaf, D.C., Ribière, M., Ryabov, E., De Smet, L., van der Steen, J.J.M. (2013). Standard methods for virus research in Apis mellifera. J Apic Res, 52, 1–56. [CrossRef]
- Dequenne, I., Philippart de Foy, J. M., & Cani, P. D. (2022). Developing Strategies to Help Bee Colony Resilience in Changing Environments. Animals (Basel), 12(23), 3396. [CrossRef]
- Dhruba Naug, Nutritional stress due to habitat loss may explain recent honeybee colony collapses, Biological Conservation, Volume 142, Issue 10, 2009, Pages 2369-2372, ISSN 0006-3207. [CrossRef]
- Engel, P., Kwong, W.K., McFrederick, Q., Anderson, K.E., Barribeau, S.M., Chandler, J.A., Cornman, R.S., Dainat, J., de Miranda, J.R., Doublet, V., Emery, O., Evans, J.D., Farinelli, L., Flenniken, M.L., Granberg, F., Grasis, J.A., Gauthier, L., Hayer, J., Koch, H., Kocher, S., Martinson, V.G., Moran, N., Munoz-Torres, M., Newton, I., Paxton, R.J., Powell, E., Sadd, B.M., Schmid-Hempel, P., Schmid-Hempel, R., Song, S.J., Schwarz, R.S., vanEngelsdorp, D., Dainat, B. (2016). The Bee Microbiome: Impact on Bee Health and Model for Evolution and Ecology of Host-Microbe Interactions. mBio, 7(2), e02164-15. [CrossRef]
- Engel P, Kwong WK, McFrederick Q, Anderson KE, Barribeau SM, Chandler JA, Cornman RS, Dainat J, de Miranda JR, Doublet V, Emery O, Evans JD, Farinelli L, Flenniken ML, Granberg F, Grasis JA, Gauthier L, Hayer J, Koch H, Kocher S, Martinson VG, Moran N, Munoz-Torres M, Newton I, Paxton RJ, Powell E, Sadd BM, Schmid-Hempel P, Schmid-Hempel R, Song SJ, Schwarz RS, vanEngelsdorp D, Dainat B. The Bee Microbiome: Impact on Bee Health and Model for Evolution and Ecology of Host-Microbe Interactions. mBio. 2016 Apr 26;7(2):e02164-15. [CrossRef] [PubMed] [PubMed Central]
- Even, N., Devaud, J.M., Barron, A.B. (2012). General Stress Responses in the Honey Bee. Insects, 3(4), 1271-98. [CrossRef]
- Forsgren, E. (2010). European foulbrood in honey bees. J Invertebr Pathol, 103(Suppl 1), S5–S9. [CrossRef]
- Galajda, R., Valenčáková, A., Sučik, M., Kandráčová, P. (2021). Nosema Disease of European Honey Bees. J Fungi (Basel), 7(9), 714. [CrossRef]
- Genersch, E. (2010). American foulbrood in honeybees and its causative agent, Paenibacillus larvae. J Invertebr Pathol, 103(Suppl 1), S10–S19. [CrossRef]
- Geslin B, Gauzens B, Thébault E, Dajoz I (2013) Plant Pollinator Networks along a Gradient of Urbanisation. PLOS ONE 8(5): e63421. [CrossRef]
- Glass, J.R., Harrison, J.F. (2024). A thermal performance curve perspective explains decades of disagreements over how air temperature affects the flight metabolism of honey bees. J Exp Biol, 227(7), jeb246926. [CrossRef]
- Goulson D, Nicholls E, Botías C, Rotheray EL. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. 2015 Mar 27;347(6229):1255957. [CrossRef] [PubMed]
- Goulson D, Nicholls E. Anthropogenic influences on bee foraging. Science. 2022 Mar 4;375(6584):970-972. Epub 2022 Mar 3. [CrossRef] [PubMed]
- Hassler, Edgar E., Joseph A. Cazier, Brandon Hopkins, James T. Wilkes, Kiefer Smith, and Max Rünzel. (2021). "A century of discovery: Mining 100 years of honey bee research." Journal of Apicultural Research 60, no. 1: 3-12. [CrossRef]
- Hester, K.P., Stoner, K.A., Eitzer, B.D., Koethe, R.W., Lehmann, D.M. (2023). Pesticide residues in honey bee (Apis mellifera) pollen collected in two ornamental plant nurseries in Connecticut: Implications for bee health and risk assessment, Environmental Pollution, Volume 333, 2023, 122037, ISSN 0269-7491. [CrossRef]
- Higes, M., Martín, R., Meana, A. (2006). Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J Invertebr Pathol, 92(2), 93–95. [CrossRef]
- Iwasa, T., Motoyama, N., Ambrose, J. T., & Roe, R. M. (2004). Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop protection, 23(5), 371-378. [CrossRef]
- Jaworski, C. C., Geslin, B., Zakardjian, M., Lecareux, C., Boulogne, I., Raynaud, X.,... & Tison, L. (2023). In-hive disturbances linked to neonicotinoids exposure in honey bees. Scientific reports, 13(1), 3747.
- Khan KA, Al-Ghamdi AA, Ghramh HA, Ansari MJ, Ali H, Alamri SA, Al-Kahtani SN, Adgaba N, Qasim M, Hafeez M. (2020). Structural diversity and functional variability of gut microbial communities associated with honey bees. Microb Pathog. Jan;138:103793. Epub 2019 Oct 15. [CrossRef] [PubMed]
- Kit S. Prendergast, Sean Tomlinson, Kingsley W. Dixon, Philip W. Bateman, Myles H.M. (2022). Menz, Urban native vegetation remnants support more diverse native bee communities than residential gardens in Australia's southwest biodiversity hotspot, Biological Conservation, Volume 265, 109408, ISSN 0006-3207. [CrossRef]
- Klocek D, Grybchuk D, Macedo DH, Galan A, Votýpka J, Schmid-Hempel R, Schmid-Hempel P, Yurchenko V, Kostygov AY. (2023). RNA viruses of Crithidia bombi, a parasite of bumblebees. J Invertebr Pathol. Nov;201:107991. Epub 2023 Sep 13. [CrossRef] [PubMed]
- Langowska, A., Zawilak, M., Sparks, T.H. et al. (2017). Long-term effect of temperature on honey yield and honeybee phenology. Int J Biometeorol 61, 1125–1132. [CrossRef]
- Laurino, D., Manino, A., Patetta, A., Porporato, M. (2011). Toxicity of neonicotinoid insecticides to honey bees: laboratory tests. Bulletin of insectology, 64(1), 107-113.
- Levin, M.D., Bohart, G.E. (1955). Effect of Cold Storage on the Activity and Life Span of Two Osmia Lignaria Subspecies (Hymenoptera: Megachilidae). Journal of Economic Entomology, 48(3), 326–329. [CrossRef]
- Li YH, Chen YH, Chang FM, Wu MC, Nai YS. (2024). Monitoring the Season-Prevalence Relationship of Vairimorpha ceranae in Honey Bees (Apis mellifera) over One Year and the Primary Assessment of Probiotic Treatment in Taichung, Taiwan. Insects. Mar 19;15(3):204. [CrossRef] [PubMed] [PubMed Central]
- Lin Z, Shen S, Wang K, Ji T (2024). Biotic and abiotic stresses on honeybee health. Integrative Zoology 19, 442–457. [CrossRef]
- Mahé, C., Jumarie, C., Boily, M. (2021). The countryside or the city: Which environment is better for the honeybee?, Environmental Research, Volume 195, 110784, ISSN 0013-9351. [CrossRef]
- Manjon, C., Troczka, B.J., Zaworra, M., Beadle, K., Randall, E., Hertlein, G.,... & Bass, C. (2018). Unravelling the molecular determinants of bee sensitivity to neonicotinoid insecticides. Current Biology, 28(7), 1137-1143.
- Manlik, O., Mundra, S., Schmid-Hempel, R., & Schmid-Hempel, P. (2023). Impact of climate change on parasite infection of an important pollinator depends on host genotypes. Global Change Biology, 29, 69–80. [CrossRef]
- Mayack, Christopher, Naug, Dhruba (2009). Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection, Journal of Invertebrate Pathology, Volume 100, Issue 3, Pages 185-188, ISSN 0022-2011. [CrossRef]
- McDermott A. (2021). News Feature: To understand the plight of insects, entomologists look to the past. Proc Natl Acad Sci U S A. Jan 12;118(2):e2018499117. Epub 2020 Dec 16. [CrossRef] [PubMed] [PubMed Central]
- McMenamin AJ, Genersch E. (2015). Honey bee colony losses and associated viruses. Curr Opin Insect Sci 8:121–129. [CrossRef]
- Mutinelli, F. (2011). The spread of pathogens through trade in honey bees and their products (including queen bees and semen): overview and recent developments. Rev Sci Tech, 30(1), 257-271. [CrossRef]
- Murilhas, A. (2005). Aethina tumida arrives in Portugal. Will it be eradicated? Eur Bee Newsl, 7, 7–9.
- Nagaraja, N., Raja, D.S. (2021). Honey bee diseases and pests management strategies in the Indian subcontinent: An update. J Invertebr Pathol, 184, 107648. [CrossRef]
- Nazzi, F., Pennacchio, F. (2014). Disentangling multiple interactions in the hive ecosystem. Trends Parasitol, 30(12), 556–561. [CrossRef]
- Nguyen TT, Yoo MS, Lee HS, Truong AT, Youn SY, Lee SJ, Kim J, Cho YS. First detection and prevalence of Apis mellifera filamentous virus in Apis mellifera and Varroa destructor in the Republic of Korea. Sci Rep. 2024 Jun 19;14(1):14105. [CrossRef] [PubMed] [PubMed Central]
- Oddie MAY, Lanz S, Dahle B, Yañez O, Neumann P. (2023). Virus infections in honeybee colonies naturally surviving ectoparasitic mite vectors. PLoS One. 2023 Dec 15;18(12):e0289883. [CrossRef] [PubMed] [PubMed Central]
- Palmer-Young EC, Markowitz LM, Huang W-F, Evans JD. High temperatures augment inhibition of parasites by a honey bee gut symbiont. Appl Environ Microbiol. 2023 Oct 31;89(10):e0102323. Epub 2023 Oct 4. [CrossRef] [PubMed] [PubMed Central]
- Papa G, Capitani G, Capri E, Pellecchia M, Negri I. Vehicle-derived ultrafine particulate contaminating bees and bee products. Sci Total Environ. 2021 Jan 1;750:141700. Epub 2020 Aug 14. [CrossRef] [PubMed]
- Parrella P, Elikan AB, Kogan HV, Wague F, Marshalleck CA, Snow JW. (2024). Bleomycin reduces Vairimorpha (Nosema) ceranae infection in honey bees with some evident host toxicity. Microbiol Spectr. Feb 6;12(2):e0334923. Epub 2024 Jan 5. [CrossRef] [PubMed] [PubMed Central]
- Phillips, Benjamin B., James M. Bullock, Kevin J. Gaston, Karen A. Hudson-Edwards, Meg Bamford, Dave Cruse, Lynn V. Dicks, Carmen Falagan, Claire Wallace, and Juliet L. Osborne. "Impacts of multiple pollutants on pollinator activity in road verges." Journal of Applied Ecology 58, no. 5 (2021): 1017-1029. [CrossRef]
- Pohl, F., 2005. Bienen- krankheiten. Franckh- Kosmos Verlags- GmbH & Co KG, Stuttgart. P115-116, ISBN 978-3-440-10407-1.
- Potts, S. G., Roberts, S. P. M., Dean, R., Marris, G., Brown, M. A., Jones, R., Neumann, P., & Settele, J. (2010). Declines of managed honey bees and beekeepers in Europe. Journal of Apicultural Research, 49(1), 15-22. [CrossRef]
- Potts, S.G., Biesmeijer, J.C., Kremen, C., et al. (2010). Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol, 25, 345–353. [CrossRef]
- Prendergast, K.S., Dixon, K.W. & Bateman, P.W. (2022) A global review of determinants of native bee assemblages in urbanised landscapes. Insect Conservation and Diversity, 15(4), 385–405. [CrossRef]
- Raine NE, Rundlöf M. (2023). Pesticide Exposure and Effects on Non-Apis Bees. Annu Rev Entomol. 2024 Jan 25;69:551-576. Epub 2023 Oct 12. [CrossRef] [PubMed]
- Ramsey, SD, Ochoa, R, Bauchan, G, Gulbronson, C, Mowery, JD, Cohen A. et al. (2019).
- Varroa destructor feeds primarily on the fat tissue of the bee body and not on the hemolymph Proc Natl Acad Sci USA, 116 ( 2019 ), s. 1792 - 1801. [CrossRef]
- Reid, R. J., Troczka, B. J., Kor, L., Randall, E., Williamson, M. S., Field, L. M.,... & Davies, T. E. (2020). Assessing the acute toxicity of insecticides to the buff-tailed bumblebee (Bombus terrestris audax). Pesticide biochemistry and physiology, 166, 104562. [CrossRef]
- Ryabov EV, Nearman AJ, Nessa A, Grubbs K, Sallmann B, Fahey R, Wilson ME, Rennich KD, Steinhauer N, Fauvel AM, Chen Y, Evans JD, vanEngelsdorp D. (2023). Apis mellifera Solinvivirus-1, a Novel Honey Bee Virus That Remained Undetected for over a Decade, Is Widespread in the USA. Viruses. 2023 Jul 21;15(7):1597. [CrossRef] [PubMed] [PubMed Central]
- Streicher T, Brinker P, Tragust S, Paxton RJ. Host Barriers Limit Viral Spread in a Spillover Host: A Study of Deformed Wing Virus in the Bumblebee Bombus terrestris. Viruses. 2024 Apr 15;16(4):607. [CrossRef] [PubMed] [PubMed Central]
- Switanek, Matthew, Crailsheim, Karl, Truhetz, Heimo, Brodschneider, Robert. (2017). Modelling seasonal effects of temperature and precipitation on honey bee winter mortality in a temperate climate, Science of The Total Environment, Volume 579, Pages 1581-1587, ISSN 0048-9697. [CrossRef]
- Thimmegowda GG, Mullen S, Sottilare K, Sharma A, Mohanta R, Brockmann A, Dhandapany PS, Olsson SB. (2020). A field-based quantitative analysis of sublethal effects of air pollution on pollinators. Proc Natl Acad Sci U S A. 2020 Aug 25;117(34):20653-20661. Epub 2020 Aug 10. [CrossRef] [PubMed] [PubMed Central]
- Tong L, Nieh JC, Tosi S. Combined nutritional stress and a new systemic pesticide (flupyradifurone, Sivanto®) reduce bee survival, food consumption, flight success, and thermoregulation. Chemosphere. 2019 Dec;237:124408. Epub 2019 Jul 20. [CrossRef] [PubMed]
- USEPA, Health Canada Pest Management Regulatory Agency, and the California Department of Pesticide Regulation, 2014. Guidance for Assessing Pesticide Risks to Bees. https://www.epa.gov/sites/production/files/2014-06/documents/pollinator_risk_assessment_guidance_06_19_14.pdf 59.
- Vanderplanck, M., Gerbaux, P., Mougel, F., Logan, M., Moerman, R., Garofalo, F., Rasmont, P., De Pauw, E., Michez, D. (2019). Influence of pollen quality on male and female reproductive performance in a solitary bee. Sci Rep, 9, 11688. [CrossRef]
- Van Espen, Marie, Williams, James H., Alves, Fátima, Hung, Yung, de Graaf, Dirk C., Verbeke, Wim (2023). Beekeeping in Europe facing climate change: A mixed methods study on perceived impacts and the need to adapt according to stakeholders and beekeepers, Science of The Total Environment, Volume 888, 164255, ISSN 0048-9697.
- Wang K, Li J, Zhao L, Mu X, Wang C, Wang M, Xue X, Qi S, Wu L. (2021). Gut microbiota protects honey bees (Apis mellifera L.) against polystyrene microplastics exposure risks. J Hazard Mater. 2021 Jan 15;402:123828. Epub 2020 Sep 5. [CrossRef] [PubMed]
- Yun JH, Jung MJ, Kim PS, Bae JW. (2018). Social status shapes the bacterial and fungal gut communities of the honey bee. Sci Rep. 2018 Jan 31;8(1):2019. [CrossRef] [PubMed] [PubMed Central]
- Zapata-Hernández, G., Gajardo-Rojas, M., Calderón-Seguel, M., Muñoz, A. A., Yáñez, K. P., Requier, F., Fontúrbel, F. E., Ormeño-Arriagada, P. I., & Arrieta, H. (2024). Advances and knowledge gaps on climate change impacts on honey bees and beekeeping: A systematic review. Global Change Biology, 30, e17219. [CrossRef]
- Zbrozek M, Fearon ML, Weise C, Tibbetts EA. (2023). Honeybee visitation to shared flowers increases Vairimorpha ceranae prevalence in bumblebees. Ecol Evol. Sep 20;13(9):e10528. [CrossRef] [PubMed] [PubMed Central]
- Zhang Y, Su M, Wang L, Huang S, Su S, Huang WF. (2021). Vairimorpha (Nosema) ceranae Infection Alters Honey Bee Microbiota Composition and Sustains the Survival of Adult Honey Bees. Biology (Basel). Sep 13;10(9):905. [CrossRef] [PubMed] [PubMed Central]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



