
Submitted:
01 August 2024
Posted:
02 August 2024
You are already at the latest version
Abstract
Probiotics are living microbes that provide benefits to the host. The increasing data on health promotion following probiotics administration triggered growing interest among researchers and pharmaceutical companies looking for novel therapeutic or preventive tools. Infections of the lower genital tract in females are caused by a wide range of pathogens, they are widespread, and represent one of the main areas for the use of probiotics and postbiotics. Vulvovaginal candidiasis (VVC) affects 75% of women in their reproductive age at least once during their lifetime. Of these women 5-8% develop the recurrent form (RVVC). The disease onset is triggered by the overgrowth of Candida species on the vaginal mucosa. Here, we evaluated the beneficial properties of the Bacillus coagulans spore-producing LMG S-24828 against C. albicans and C. parapsilosis. Our results show that both alive B. coagulans and its Cell-Free-Supernatant (CFS) exerted antifungal activity against both fungi. Moreover, alive B. coagulans reduced hyphal formation, inhibited C. albicans adhesion to vaginal epithelial cells and showed co-aggregation capacity. Overall, B. coagulans LMG S-24828 exerted a protective effect on vaginal epithelial cells infected with C. albicans. These data suggest that B. coagulans LMG S-24828 may provide benefits in the context of Candida vaginal infections.
Keywords:
Bacillus coagulans
; Probiotics
; Candida
; Vaginal epithelial cells
1. Introduction
Vulvovaginal candidiasis (VVC) is a disease widely spread among the female population. Three-fourths of women worldwide are estimated to be affected by VVC at least once during their reproductive age. Moreover, 5-8% of them develop the recurrent form (RVVC), consisting of more than 4 episodes of VVC per year. Both VVC and RVVC have a significant impact on the life quality and mental wellbeing of the approximately 138 million affected women [1,2]. VVC is a symptomatic inflammation of the vagina, and it is mainly caused by Candida albicans (C. albicans). Infections due to other species of the genus Candida, such as C. parapsilosis, C. glabrata, C. tropicalis, and C. krusei, are less common, but they are more frequently associated with the recurrent form. VVC symptoms, that include itching, burning, discharge, and dyspareunia, are caused by a hyperinflammatory response to Candida rather than by the presence of the fungus, which normally dwells in the vagina as a commensal [3]. As a colonizer of the healthy vaginal niches, Candida dwells on the vaginal mucosa in form of yeast and its proliferation and pathogenicity are constantly inhibited by the immune system and by resident bacteria, such as Lactobacillus spp. The pH of a normal vaginal environment typically ranges between 3.5 and 4.5, because of the lactic acid produced by lactobacilli [4,5]. Alterations in microbial composition and immune system dysfunctions may increase the risk of Candida infection. Pregnancy, diabetes mellitus, hormones administered as oral contraceptives, wide-spectrum antibiotic therapy and genetic predisposition are among the main VVC predisposing factors [6]. The VVC onset is linked to C. albicans morphological transition from yeast to hyphal form. The subsequent epithelial invasion and cell damage trigger an intense immune and inflammatory response; the latter ultimately causes the symptoms of the disease.
VVC episodes are typically treated with oral or topical azole antifungals (mainly fluconazole). However, despite the therapy effectiveness grants relief from symptoms, in about 50% of cases reinfection occurs after 6 months. For RVVC, the elective therapy requires continuous treatment with fluconazole for 6 months. Nevertheless, more than half of the women show symptoms relapse few months after the end of fluconazole treatment. Overall, azole-based treatment has a limited long-term efficacy and consequently high costs [7,8]. Moreover, the occurrence of clinical isolates resistant to azoles further indicates the need to define alternative therapeutic approaches [9].
In this regard, interest in probiotics and on their role in human health is increasing in recent years, because of their excellent performance in preventing and treating several diseases and of the increasing demand for natural medicines by consumers [10]. Probiotics are living microorganisms that, when consumed in appropriate quantities, provide health benefits to the host, as defined by the World Health Organization Antimicrobial activity against pathogens, modulation of the immune system, and enhancement of essential nutrients bioavailability are among the major benefits granted by probiotics consumption. In addition, engineered microbes may be used as therapeutic vectors, to deliver vaccines, antibiotics, enzymes, or other drugs, representing thus a promising research field for the treatment of several diseases. Probiotics have been demonstrated to interact also with host microbiota. The latter may influence several organs, typically by immunomodulatory signals. Indeed, connections between the gut microbiota and the brain, liver, lung, skin and vagina have been already demonstrated [11,12]. Probiotics are being studied also in psychiatry as a novel therapeutic approach aimed to treat conditions such as mood disorders, anxiety, and depression [13,14].
Probiotics are typically Gram-positive bacteria, mainly belonging to the genera Lactobacillus and Bifidobacterium. Recently, other genera have been demonstrated to show probiotic activity [15,16]. Spore-producing bacteria, such as several species belonging to the genus Bacillus, have been frequently employed in commercial formulations due to their properties. Indeed, they can survive various industrial processes, and they remain stable at room temperature over non-sporulating probiotic species [17]. Most probiotic formulations are taken orally. In spore-based formulation probiotics, spore germination and bacterial proliferation occur on the gut mucosa surface.
Evidence on the benefits of probiotics in VVC treatment or prevention is yet to be fully demonstrated, but preclinical studies and clinical trials suggest a beneficial role of several probiotics against Candida vaginal infections [18,19]. Although there is no sufficient evidence to support the replacement of antifungals with probiotics to date, the latter represent an interesting approach at least to prevent the recurrent forms and an aid to restore the vaginal eubiosis [20,21].
Here, we showed in vitro that Bacillus coagulans LMG S-24828 (recently reclassified as Heyndrickxia coagulans and marketed as Weizy® by Giellepi S.p.A.), counteracts Candida virulence and exerts a beneficial effect against vaginal epithelial cells infection by C. albicans. These promising preliminary data suggest that this bacterial strain has all the potentialities to be employed as a spore-forming probiotic for counteracting Candida spp. vaginal infections.
2. Materials and Methods
2.1. Microbial Strains and Growth Conditions
The strain of Bacillus coagulans LMG S-24828 (Weizy®) was provided by Giellepi SpA (Milan, Italy) in the form of lyophilized spores. Spores were stored at 4 °C and germination was obtained by incubation in Tryptic Soy Broth (TSB) (Condalab, Spain) for 24 hours at 37 °C, under agitation. Bacteria, obtained from spores’ germination, were used in their exponential growth phase for each experiment. The reference strains Candida albicans SC5314 (ATCC MYA-2876) and Candida parapsilosis CLIB214 (ATCC 22019) were employed. Both strains had been stored in frozen stocks at -80 °C in Sabouraud Dextrose Broth (SDB) (Condalab, Spain) supplemented with 15% glycerol. After thawing, the fungi were grown in Yeast Extract-Peptone-Dextrose broth (YPD) (Scharlab S.L., Spain) and incubated at 37 °C under aerobic conditions for 24 hours. Fungi in the exponential growth phase were used in the experiments.
2.2. pH Measurement of Microbial Culture
Bacillus coagulans (5x106 CFU/mL) and C. albicans or C. parapsilosis (both at 5x103 CFU/mL) were added to a medium consisting of 5 ml of YPD broth and 5 ml of TSB. The co-culture was incubated at 37 °C for 24 hours, under agitation. Samples containing only the bacterium, only the fungus, or sterile media were also included as controls. After incubation, samples were centrifuged at 4,000 rpm for 15 minutes and sterilized by filtration; then, pH was measured using a pHmeter (Hanna Instruments, Italy).
2.3. Analysis of Bacillus Coagulans Effect on Candida Growth
The impact of B. coagulans on C. albicans and C. parapsilosis growth was evaluated by incubating the fungi with or without the bacterium. Five hundred microliters of B. coagulans (5x106 CFU/mL) in TSB were seeded in a 24-well plate with 500 µL of C. albicans or C. parapsilosis (both at 5x103 CFU/mL) in YPD broth. As a control sample, both Candida species were incubated with sterile TSB without B. coagulans. To assess acidification contribution, 500 µL of both fungi were seeded with 500 µL of TSB that had been acidified with chloridric acid (Mallinckrodt Baker, USA) to pH 4.4. The plate was incubated at 37 °C for 24 hours. Then, culture supernatants were collected together with adherent fungi, that were detached by washing the wells with Soybean-Casein-Digest-Lecithin-Polysorbate-80 broth (SCDLP80, Biotec, Italy). The Candida burden was quantified by serial diluting the samples and seeding them on Sabouraud Dextrose Agar (SDA) (Condalab, Spain) supplemented with chloramphenicol (50 mg/l) (Biolife Italiana Srl, Italy). Colony Forming Units (CFU) were counted after 24-48 hours of incubation at 37 °C.
2.4. Preparation of Cell-Free-Supernatant (CFS) from Bacillus Coagulans and Its Effect on Candida Growth
The B. coagulans broth culture obtained from spore germination, as detailed above, was centrifuged at 4,000 rpm at 4 °C for 15 minutes. Then, the supernatant was discarded, and the pellet was resuspended in 1 ml of TSB. Optical density (OD) at the wavelength of 595 nm was measured using a spectrophotometer (Sunrise, Tecan, Switzerland), and bacterial concentration was calculated from the OD values by means of a stored standard curve. Then, 5 ml of TSB containing B. coagulans (1x108 CFU/mL) were incubated at 37 °C for 24 hours, under agitation. At the end of the incubation, CFS was obtained by centrifugation of the bacterial culture at 4,000 rpm at 4 °C for 15 minutes, as previously described [22]. Briefly, the supernatants were collected, filter-sterilized with 0.22 µm syringe filters (Corning Incorporated, Germany) and stored at -80 °C until their use. The sterility of CFS was confirmed by incubating an aliquot of each CFS at 37 °C for 72 hours and then seeding it on Tryptic Soy Agar (TSA). The effect of CFS on C. albicans and C. parapsilosis growth was evaluated by seeding 100 µl of YPD containing C. albicans or C. parapsilosis (1x104 CFU/mL) in a 96-well plate, together with 100 µL of CFS (or the same volume of TSB in the control samples). The plate was incubated at 37°C and the cultures OD were kinetically measured (reading cycle: 2 hours) for 18 hours at the wavelength of 570 nm by a spectrophotometer (Sunrise, Tecan, Switzerland).
2.5. Bacillus Coagulans Impact on C. albicans Hyphae Formation
Filamentation assay was performed on chamber slides (Nunc Lab-Tek II, Thermo Fisher Scientific, USA). One hundred µL of C. albicans in YPD broth supplemented with 10% of fetal bovine serum (FBS) (SIAL Srl, Italy) were seeded in the well of the chamber slide along with 100 µl of B. coagulans in TSB supplemented with 10% FBS. As a control sample, the fungus was incubated with sterile TSB supplemented with 10% FBS. The chamber slide was placed at 37 °C with 5% CO2 for 4 hours. Fifteen minutes before the end of the incubation, 40 µL of 1% Uvitex 2B fluorescent dye (Polysciences, Inc, PA, USA) were added to each well. After the end of incubation, wells were gently washed twice with 200 µL of phosphate buffered saline (PBS) that had been warmed at 37 °C. Then, samples were fixed with 1% paraformaldehyde (200 µl/well, 30 minutes of incubation at 4 °C). Finally, wells were washed twice with 200 µL of cold PBS (4 °C) and they were allowed to dry. The chamber slide was then disassembled by removing the plastic frame that kept the wells isolated, and the slide surface was treated with ProLong Gold Antifade Reagent (Molecular Probes, Invitrogen, St. Louis, Mo, USA). The fluorescence of fungi was visualized by epifluorescence microscope Nikon Eclypse 90i (Nikon Instruments, Tokyo, Japan). Yeast cells and hyphal fragments were counted (at least three fields for each experimental condition) and the percentage of hyphal fragments in each sample was calculated.
2.6. Evaluation of Bacillus Coagulans Ability to Co-Aggregate with C. albicans
To establish if B. coagulans was able to co-aggregate with C. albicans, 50 µL of yeasts resuspended in PBS (2x107 CFU/mL) and 50 µL of B. coagulans suspension in PBS (2x109 CFU/mL) were seeded in a U-bottom 96-well plate (Corning Incorporated, USA) and incubated in gentle agitation at 37 °C for 1 hour. Samples containing the bacteria, or the fungus alone were also included as controls. Co-aggregation rate was visually evaluated and scores from 0 to 4 were assigned, according to previous report [19]. Briefly, score 0 means no aggregation; score 1 is characterized by aggregates with small clusters; score 2 is characterized by aggregates with larger numbers of yeasts; score 3 includes clumps visible with the naked eye, containing large numbers of yeast cells; score 4, the maximum, is assigned when large clumps become visible with the naked eye at the center of the well.
2.7. Long-Term Effect of Bacillus Coagulans on Candida Metabolism
To evaluate whether the B. coagulans - Candida co-incubation could determine a long-term effect on fungal metabolic activity, 500 µL of the C. albicans or C. parapsilosis (both 1x105 CFU/mL) in YPD broth were seeded in a 24-well plate (Sarstedt, Germany), 500 µL of B. coagulans (1x108 CFU/mL) in TSB were added to each well and incubation was carried out at 37 °C for 24 hours. In control samples, sterile TSB was added. At the end of incubation time, an aliquot of each sample was collected and seeded on SDA supplemented with chloramphenicol to exclude B. coagulans growth. Plates were incubated at 37°C for 24 hours. Plates were then washed to collect the grown fungi and 100 μl of both C. albicans and C. parapsilosis suspensions (1x106 CFU/mL) were seeded in DMEM supplemented with 5% FBS in a 96-well plate and incubated at 37 °C for 24 hours. Then, the medium was removed from each well, and 100 µL of 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT, Sigma Aldrich, USA) solution, supplemented with menadione (1 μM, Sigma-Aldrich, USA), were added. The plate was covered with tinfoil and incubated at 37 °C for 3 hours. Finally, 80 µL were collected from each well, transferred to another 96-well plate and the color intensity was quantified by measuring the OD at 492 nm wavelength.
2.8. Establishment of the Monolayer of Vaginal and Intestinal Epithelial Cells
Two different human epithelial cell lines were employed: A-431 cell line from vaginal epithelial squamous cell carcinoma (ATCC CLR-1555) and the CaCo-2 cell line from colon-rectal adenocarcinoma (ATCC HTB-37). Both cell lines were cultured in DMEM (SIAL Srl, Italy) supplemented with L-glutamine (2 nM) (Euroclone SpA, Italy), penicillin (100 U/ml) (Euroclone SpA, Italy), streptomycin (100 μL/ml) (Euroclone SpA, Italy), ciprofloxacin (20 mg/ml) (Euroclone SpA, Italy) and FBS (Fetal Bovine Serum, SIAL Srl, Italy). The cell lines were kept viable by sub-culturing twice a week and keeping them at 37 °C with 5% CO2. Epithelial cell monolayers were obtained by seeding the cells in a 24-well plate (Sarstedt, Germany) (5x105/mL, 1 mL/well) and by incubating the plate for 1 day (A-431) and 2 days (CaCo-2) at 37 °C with 5% CO2. Before being infected, the epithelial cell monolayers were washed with warmed PBS. DMEM supplemented with 10% FBS was employed for monolayer establishment, whereas DMEM supplemented with 5% FBS was used for the experiments. In the experiments including B. coagulans, antibiotics were not added to the medium.
2.9. Effect of Bacillus Coagulans on C. albicans-Induced Vaginal Epithelial Cell Damage
Five hundred microliters of B. coagulans were seeded on a vaginal epithelial cell monolayer (bacteria/vaginal cells, Multiplicity-Of-Infection (MOI 100/1) and incubated at 37 °C with 5% CO2 for 6 hours. Five hundred microliters of C. albicans were then added to each well (C. albicans/vaginal cells, MOI 1/1), then the plate was incubated at 37 °C with 5% CO2 for further 18 hours. After the incubation, lactate dehydrogenase (LDH) release in the surrounding medium was measured by a commercial kit (Hoffmann-La Roche, Switzerland). The percentage of damage was calculated following the manufacturer's instructions. Due to LDH activity impairment caused by acidification, a bacteria-acid-producer-specific protocol was employed for samples containing B. coagulans, as previously described [23]. Briefly, the culture medium of each well was removed, and wells were washed with warmed HBSS (Gibco, USA). Vaginal epithelial cells were then lysed with 0.1% Triton X-100 and the LDH assay was performed. Cell damage was calculated as the complementary percentage of the cell viability obtained by considering OD values from uninfected cells as 100%.
2.10. Impact of Bacillus Coagulans on C. albicans Adhesion to Vaginal Epithelial Cells
Five hundred microliters of B. coagulans (bacteria/vaginal cells, MOI 100/1) and 500 µL of C. albicans (C. albicans/vaginal cells, MOI 1/1) were simultaneously added to a vaginal epithelial cells monolayer and incubated at 37 °C with 5% CO2 for 2 hours. Cells devoid of bacteria and infected with the fungus were used as control. After the incubation, the culture medium was removed, and the wells were gently washed with warm PBS to remove non-adherent fungi. Vaginal epithelial cells were then lysed by adding 1 mL of 0.2% Triton X-100. Then, serial dilutions were performed and plated on SDA supplemented with chloramphenicol. CFU were counted after 24-48 hours of incubation at 37 °C. The percentage of fungal adhesion in samples containing C. albicans and B. coagulans was calculated by considering the mean CFU values of C. albicans control samples as 100%. Adhesion inhibition value was calculated by subtracting the percentage of adhesion from 100.
2.11. β-Defensin-2 Production after C. albicans Infection in the Presence of Bacillus Coagulans
The release of β-defensin-2 by vaginal epithelial cells, that had been pre-colonized with B. coagulans for 6 hours (bacteria/vaginal cells, MOI 100/1) and then infected with C. albicans (C. albicans/vaginal cells, MOI 1/1) for further 18 hours, were quantified by using a commercial ELISA kit (MyBioSource, USA) according to Manufacturers’ instructions.
2.12. Evaluation of Bacillus Coagulans Spores’ Capacity to Germinate on the Intestinal Epithelial Cells
The CaCo-2 intestinal epithelial cell monolayer was challenged with 1 mL of DMEM-TSB (1:1) containing B. coagulans spores and incubated at 37 °C plus 5% CO2 for 24 hours. Cells incubated with sterile medium were included as control. After incubation, wells were visualized by an inverted-optical microscope (Nikon Eclipse TS100, Japan). Then, monolayers were lysed by adding 100 μl of 0.2% Triton X-100 to each well. After pipetting, 20 μl were deposited on a microscope glass slide and a Gram staining (Liofilchem, Italy) was performed. Slides were visualized using the Nikon Eclypse 90i (Nikon Instruments, Tokyo, Japan) microscope.
2.13. Statistical Analysis
All statistical analyses were carried out using GraphPad Prism 10 software. Gaussian distribution of data was confirmed by the Shapiro-Wilk test. Statistical analysis of normally distributed data was performed by using the unpaired, two-tailed, Student’s t-test or the one-way ANOVA test followed by the uncorrected Fisher’s LSD multiple comparisons tests. Data with no Gaussian distribution was analyzed by the Kruskal-Wallis test followed by the uncorrected Dunn’s multiple comparisons test. The specific statistical analysis performed in each experiment has been detailed in the figure legends. Values of *p < 0.05, ***p < 0.001, and ****p < 0.0001 were considered statistically significant.
3. Results
3.1. Bacillus Coagulans Exerts Anti-Fungal Activity against C. albicans and C. parapsilosis
To investigate a direct antifungal effect of Bacillus coagulans LMG S-24828 (B. coagulans), the latter was co-incubated with C. albicans and C. parapsilosis and pH values and fungal load were evaluated. The assessment of the pH values revealed that B. coagulans induced acidification of the medium to a final pH of 4.4. The co-incubation of B. coagulans with C. albicans or C. parapsilosis did not prevent such acidification. Differently, when C. albicans and C. parapsilosis were cultured alone, the pH of the medium returned values between 6.0 and 6.5, similar to the pH of the sterile medium (Figure 1A). Next, the antiproliferative activity of B. coagulans against fungal cells was evaluated by determining the fungal load in samples where C. albicans or C. parapsilosis had been incubated in the presence or absence of B. coagulans. In addition, to establish if the acidification per se could affect fungal growth, C. albicans and C. parapsilosis were also incubated in a sterile acidified medium (pH 4.4). The results show that B. coagulans significantly reduced fungal proliferation: the growth of both Candida species was massively affected, with a mean inhibition rate of 93.6% for C. albicans (Figure 1B) and 98.9% for C. parapsilosis (Figure 1C). Differently, the bare acidification of the medium did not affect the fungal growth (Figure 1B-C).
A Cell-Free Supernatant (CFS), obtained from a 24-hour B. coagulans culture, was employed to test if B. coagulans metabolites, including organic acids, could impair C. albicans and C. parapsilosis growth. Fungal cultures were grown in presence or absence of B. coagulans CFS and their growth was kinetically monitored and assessed by optical density. Finally, growth curves were drawn. For both Candida species, the curves show that fungal growth was reduced when the fungi were incubated with B. coagulans CFS (Figure 2A-B). The analysis of the area under the curves returned significantly lower OD values when both Candida species were grown with B. coagulans CFS, thus supporting the B. coagulans CFS inhibitory effect (Figure 2C-D).
3.2. Bacillus Coagulans impairs C. albicans Adhesion to Vaginal Epithelial Cells, Reduces Hyphae Formation and Promotes C. albicans Co-Aggregation
To investigate if B. coagulans could impair fungal adhesion, a vaginal epithelial cell monolayer was challenged for 2 hours with C. albicans or with C. albicans plus B. coagulans. After removing the non-adherent fungi, epithelial cells were lysed, and the remaining suspensions containing only alive fungal cells were serially diluted and plated on SDA supplemented with chloramphenicol, to inhibit bacterial growth. As shown in Figure 3A, we obtained an average 17.5 % of inhibition in the capacity of C. albicans to adhere to vaginal epithelial cells in the presence of B. coagulans, as compared to the C. albicans capacity to adhere to vaginal epithelial cells alone. The capacity of B. coagulans to aggregate fungal cells, is one of the hypothesized mechanisms used by the bacterium to hinder C. albicans adhesion to the epithelium. By its aggregation capacity, B. coagulans forces C. albicans to a closer contact with potential antifungal metabolites, therefore preventing fungal adhesion to target host sites. Here, we evaluated the aggregation between B. coagulans and C. albicans by visual assessment: after 1 hour of coincubation a large clump, indicating coaggregation between bacteria and fungi, was visible at the center of the well and, according to the scoring system described by Pericolini E. and coworkers [19], it was assigned a score 4. Interestingly, B. coagulans was also capable to self-aggregate even though less efficiently (score 2) (Figure 3B). Moreover, a hyphal formation assay was performed to investigate if B. coagulans could affect C. albicans dimorphic transition. After 4 hours co-incubation of B. coagulans and C. albicans in a medium supplemented with 10% Fetal Bovine Serum (FBS), a significant decrease in the percentage of hyphal fragments could be observed in samples where C. albicans was grown with B. coagulans, as compared to samples where C. albicans was grown alone (Figure 3C). Figure 3D shows a heatmap representing the percentage of hyphal fragments in each different experiment performed. Figure 3E shows representative images of the fluorescence microscopy analysis, revealing fewer and shorter hyphal fragments when the fungus was co-incubated with B. coagulans.
3.3. Beneficial Effects of Bacillus Coagulans against C. albicans Vaginal Epithelial Cells Infection
Next, we tested a possible protective role of B. coagulans against the damage induced by C. albicans on vaginal epithelial cells. After 6 hours of colonization with B. coagulans, the vaginal epithelial cells were infected with C. albicans, and the incubation lasted for further 18 hours (for a total of 24 hours). After incubation, the cell damage was then quantified by measurement of lactate dehydrogenase (LDH). Our results show a significant reduction of C. albicans-induced cell damage in Candida-infected vaginal epithelial cells pre-colonized with B. coagulans, whereas in vaginal epithelial cells that had not undergone the pre-incubation with B. coagulans, a significantly higher level of cell damage could be observed (Figure 4A).
In the same experimental setting, we also tested the capacity of B. coagulans to improve vaginal epithelial cell response to C. albicans by analyzing the release of the antimicrobial peptide (AMP) β-defensin-2 by vaginal epithelial cells pre-colonized with B. coagulans. The results show that B. coagulans induced per se a significant production of β-defensin-2 by vaginal epithelial cells. β-defensin-2 production was also increased in vaginal epithelial cells pre-colonized with B. coagulans and then challenged with C. albicans, thus demonstrating that B. coagulans is able to potentiate the capacity of the infected epithelium to produce AMPs (Figure 4B).
3.4. Additional Bacillus Coagulans Features for Its Potential Use in a Probiotic Formulation to Treat Candida Vaginal Infections
The persistence of the anti-Candida effects after removal of B. coagulans was investigated. To this end, B. coagulans was co-incubated with C. albicans or C. parapsilosis for 24 hours. Then, B. coagulans was removed by seeding the samples on chloramphenicol - supplemented agar medium for further 24 hours to allow the growth of Candida colonies only. Finally, Candida was picked up and grown in fresh medium overnight and subsequently tested by XTT assay. Fungi non-precultured with B. coagulans and grown in sterile medium were employed as control. The results show the lack of a long-term effect on fungal metabolic activity in both C. albicans (Figure 5A) and C. parapsilosis (Figure 5B) after B. coagulans removal, indeed no differences in metabolic activity has been observed by comparing the two different Candida cultures. Last, we evaluated the capacity of B. coagulans spores to germinate on human intestinal epithelial cells (i.e., CaCo-2 cell line). The results show that a productive germination occurred after 24 hours from spore’s inoculum. The Gram staining confirmed the presence of Gram-positive bacilli, which were absent in the untreated CaCo-2, used as negative control (Figure 5C).
4. Discussion
Probiotics are living microbes that confer a health benefit to the human host. The anti-pathogen activity, the immunomodulatory effect, and the tolerated proliferation within the human organism are some of the acknowledged key features of probiotics. Research on probiotics has rapidly increased over the last few decades, and probiotic administration is considered as a promising therapeutic approach to treat various diseases. Yet very little information is available to date on probiotics efficacy in treatment and prevention of VVC. In the present study, we have investigated the probiotic potentiality of Bacillus coagulans (B. coagulans) LMG S-24828 (Weizy®) in the contest of Candida-associated vaginal infections. We started by evaluating the capacity of B. coagulans to affect Candida virulence traits related to invasiveness and pathogenicity, such as fungal growth, adhesion to epithelial cells and dimorphic transition. By employing a vaginal epithelial cell infection model, we assessed the potential beneficial and immunomodulatory properties of B. coagulans in the context of vaginal infection by C. albicans. A healthy vaginal environment is characterized by an acidic pH with values around 4. The acidification is granted by resident bacteria, such as lactobacilli, that produce lactic acid as the end-product of their fermentative metabolism. In addition, several other species, included in commercial probiotic formulation contribute to the acidification of the vaginal environment, thus maintaining or restoring eubiotic conditions. Recently, it has been demonstrated that B. coagulans too is able to produce lactic acid [24,25]. In line with such evidence, here we show that B. coagulans is capable to induce acidification to pH values similar to those occurring in an eubiotic vaginal milieu; in addition, such acidification capacity is not hindered by co-culturing B. coagulans with C. albicans or C. parapsilosis. Such capacity to provide an acidic environment is a necessary feature (albeit not sufficient per se) of every probiotic formulation intended for the recovery of vaginal eubiosis, which explains the importance of this result.
Candida can physiologically colonize a healthy vaginal mucosa without inducing symptoms, thanks to a constant inhibition of its overgrowth granted by the immune system and by the resident microbiota [26]. Therefore, antiproliferative activity against Candida is another pivotal attribute of probiotic formulations intended to treat or prevent vaginal infections. Our results show that B. coagulans massively affects Candida growth capacity, and this effect is not due only to the mere acidification of the medium, since both C. albicans and C. parapsilosis have been able to grow on B. coagulans-free medium, acidified to pH of 4.4. Interestingly, the evaluation of anti-proliferative activity mediated by Cell-Free Supernatant (CFS) obtained from B. coagulans culture, showed that even metabolites produced by this bacterial strain can reduce fungal growth. Therefore, it is possible that the acidification induced by B. coagulans per se may affect Candida, but that other metabolites must be involved to obtain an effective impairment of fungal growth. This would explain why Candida growth is not inhibited in a B. coagulans-free acidified medium. A similar effect has been described also for C. parapsilosis [22]. Besides supporting the probiotic features of B. coagulans LMG S-24828, these findings also suggest possible postbiotic characteristics.
The capacity to adhere is the first step for Candida to begin the infection process. Here, we show that B. coagulans impairs C. albicans adhesion and this seems to be due mostly to its capacity to co-aggregate Candida. Indeed, co-aggregation is an important mechanism used by probiotics to clear microbial pathogens: through co-aggregation, the probiotics can create a competitive microenvironment that compromises the capacity of pathogens to adhere to epithelial cells. This is, ultimately, an effective means for preventing Candida adhesion and infection [19].
As a dimorphic fungus, C. albicans can exist in two different morphologies: yeast cells or hyphal forms. When C. albicans dwells in the vaginal niche of healthy women as a harmless colonizer, the yeast form is prevalent and, in such form, Candida is well tolerated by the genital mucosa. Differently, the hyphal form becomes prevalent during symptomatic VVC episodes. The elongation of hyphal structures causes an invasion of epithelial cells, induces neutrophil recruitment and triggers a wide inflammatory response due to the high levels of cellular damage [27]. Moreover, the presence of hyphae is linked to the production of candidalysin, the only toxin demonstrated to be produced by C. albicans hyphae, that exerts cytolytic activity and induces cell damage through the induction of mtROS by vaginal epithelial cells [28,29]. Here, we demonstrate that the co-culture of C. albicans with B. coagulans dampens the hyphal elongation capacity, as shown by the observation of shorter and fewer hyphal fragments by fluorescence microscopy. In such impairment effect on C. albicans dimorphic transition, an important role may be played by the acidification induced by B. coagulans. Indeed, several studies report that acidic pH values inhibit C. albicans to switch from yeast to hyphal form [30,31]
To contextualize the observed anti-Candida effects of B. coagulans in the VVC scenario, we have employed an in vitro infection model consisting of a vaginal epithelial cell monolayer infected with C. albicans in the presence or absence of B. coagulans. By this model, we have demonstrated that upon fungal challenge the presence of the Bacillus correlates with a reduction of epithelial cell damage induced by the fungus. In addition, we have assessed the production of AMPs by vaginal epithelial cells, in order to establish if B. coagulans is capable to alert them of an impending danger and to stimulate a response. Indeed, our data show that following pre-colonization with B. coagulans, the epithelial cells get activated and increase the release of the alarmin β-defensin-2, a molecule with a potent antimicrobial activity against Gram negative bacteria and Candida [32,33].
VVC cases due to non-albicans Candida species have always been uncommon, yet they are progressively increasing. Non-albicans Candida species have been demonstrated to become resistant to antifungals at a higher rate than C. albicans [34]; hence, to prevent a Candida infection by the use of probiotics may provide beneficial effects especially against drug-resistant species. For this reason, notwithstanding the main goal of this project is the demonstration of the beneficial effects of B. coagulans LMG S-24828 against C. albicans, a C. parapsilosis reference strain was also included in some of the experiments described above. Interestingly, the effects of B. coagulans on C. parapsilosis are superimposable to the effects on C. albicans. Indeed, the hindering of both C. albicans and C. parapsilosis growth to a similar extent, suggests that the effects of B. coagulans LMG S-24828 are not species-specific, and they might affect a broad spectrum of Candida species.
A complex crosstalk links the gut to the vaginal microbiota, involving a continuous bacterial species translocation through these body sites. Indeed, the origin of vagina colonizer bacteria has been traced to the rectum, which also acts as a reservoir. In addition to the migration made possible by the anatomical proximity, also bacterial hematogenous transfer from the gut to the uterus has been described [12]. Such crosstalk is an essential feature, because most probiotic formulations are typically assumed by ingestion, and the proliferation of beneficial bacteria, as well as the germination of the spores, occur on the surface of intestinal mucosa. For spore-based probiotic formulations in particular, an efficient germination on intestinal epithelial cells is essential in targeting beneficial bacteria to the gut. Our results show that the spores of B. coagulans LMG S-24828 can germinate on the intestinal epithelial cell line CaCo-2, which makes this strain suitable to be employed as an effective probiotic to prevent and treat vaginal epithelial infections.
5. Conclusions
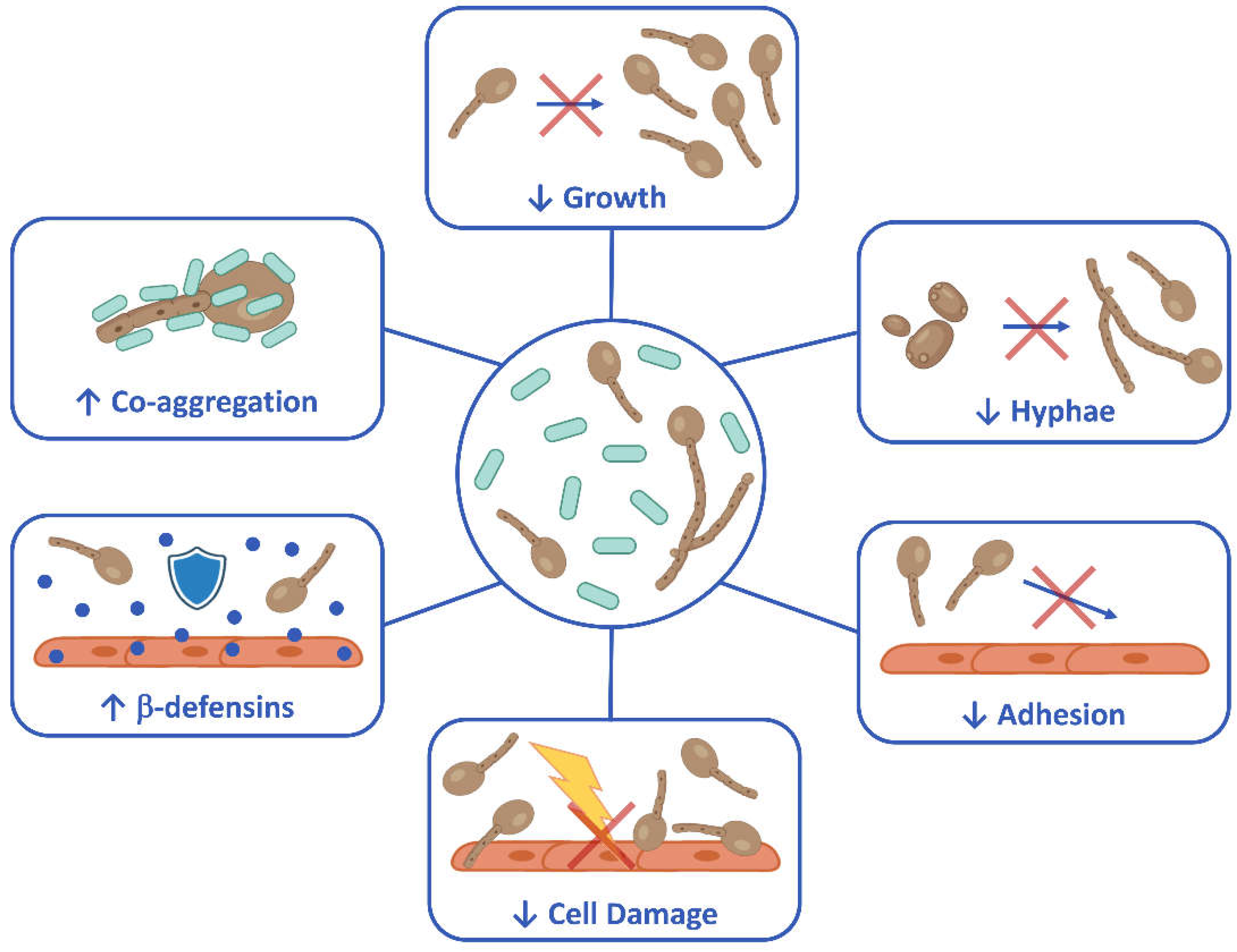
The spore-forming B. coagulans LMG S-24828 has shown a marked antifungal activity against C. albicans and C. parapsilosis, resulting in a protective effect toward vaginal epithelial cells infected by fungi. Specifically, as summarized in Figure 6, the inhibition of fungal growth both by the bacteria and by their secreted molecules, the inhibition of hyphae formation, the reduction of fungal adhesion, the capacity to co-aggregate with C. albicans, as well as the effects on the epithelial cells (reduction of Candida-induced cell damage and the increase of β-defensin-2 secretion), are all features exhibited by B. coagulans in this in vitro study. Taken together, the data presented here suggest that B. coagulans LMG S-24828 has beneficial effects that makes this bacterium a potential therapeutic tool for the prevention and treatment of Candida vaginal infection. Further studies are warranted to better characterize its probiotic and postbiotic activities.
Author Contributions
L.S., A.A., N.P., F.D.S. and E.P. have contributed significantly to this work, including conceptualization, original draft, review and editing. R.I., C.S., R.R. and S.K. contributed to the final review and editing. L.S. and N.P. were responsible for methodology, laboratory investigation and research. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding; however, Giellepi S.p.A. (Milan, Italy) provided the B. coagulans LMG S-24828 spores, as described in the Methods section. E.P. was supported by FAR instrumentation 2022.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
Financial support was not received to conduct this research; apart from Giellepi S.p.A. (Milan, Italy) providing the B. coagulans LMG S-24828 spores, all materials and equipment used in this research came from supplies purchased by E.P. Additionally, F.D.S. provided medical advice to Giellepi S.p.A. (Milan, Italy) limited to clinical research issues. R.R. is employed at Giellepi S.p.A.
References
- Zhu, Y.-X.; Li, T.; Fan, S.-R.; Liu, X.-P.; Liang, Y.-H.; Liu, P. Health-Related Quality of Life as Measured with the Short-Form 36 (SF-36) Questionnaire in Patients with Recurrent Vulvovaginal Candidiasis. Health Qual Life Outcomes. 2016, 14, 65. [Google Scholar] [CrossRef]
- Sobel, J.D. Recurrent Vulvovaginal Candidiasis. Am. J. Obstet. Gynecol. 2016, 214, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ardizzoni, A.; Wheeler, R.T.; Pericolini, E. It Takes Two to Tango: How a Dysregulation of the Innate Immunity, Coupled With Candida Virulence, Triggers VVC Onset. Front. Microbiol. 2021, 12, 692491. [Google Scholar] [CrossRef]
- Ceccarani, C.; Foschi, C.; Parolin, C.; D’Antuono, A.; Gaspari, V.; Consolandi, C.; Laghi, L.; Camboni, T.; Vitali, B.; Severgnini, M.; et al. Diversity of Vaginal Microbiome and Metabolome during Genital Infections. Sci. Rep. 2019, 9, 14095. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PLoS ON. 2013, 8, e80074. [Google Scholar] [CrossRef]
- Gaziano, R.; Sabbatini, S.; Monari, C. The Interplay between Candida Albicans, Vaginal Mucosa, Host Immunity and Resident Microbiota in Health and Disease: An Overview and Future Perspectives. Microorganisms. 2023, 11, 1211. [Google Scholar] [CrossRef]
- Satora, M.; Grunwald, A.; Zaremba, B.; Frankowska, K.; Żak, K.; Tarkowski, R.; Kułak, K. Treatment of Vulvovaginal Candidiasis—An Overview of Guidelines and the Latest Treatment Methods. JCM. 2023, 12, 5376. [Google Scholar] [CrossRef]
- Sobel, J.D.; Wiesenfeld, H.C.; Martens, M.; Danna, P.; Hooton, T.M.; Rompalo, A.; Sperling, M.; Livengood, C.; Horowitz, B.; Von Thron, J.; et al. Maintenance Fluconazole Therapy for Recurrent Vulvovaginal Candidiasis. N. Engl. J. Med. 2004, 351, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D. Resistance to Fluconazole of Candida Albicans in Vaginal Isolates: A 10-Year Study in a Clinical Referral Center. Antimicrob. Agents Chemother. 2023, 67, e00181-23. [Google Scholar] [CrossRef]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic Characteristics of Bacillus Coagulans and Associated Implications for Human Health and Diseases. J. Funct. Foods. 2020, 64, 103643. [Google Scholar] [CrossRef]
- Sánchez-Pellicer, P.; Navarro-Moratalla, L.; Núñez-Delegido, E.; Ruzafa-Costas, B.; Agüera-Santos, J.; Navarro-López, V. Acne, Microbiome, and Probiotics: The Gut–Skin Axis. Microorganisms. 2022, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Amabebe, E.; Anumba, D.O.C. Female Gut and Genital Tract Microbiota-Induced Crosstalk and Differential Effects of Short-Chain Fatty Acids on Immune Sequelae. Front. Immunol. 2020, 11, 2184. [Google Scholar] [CrossRef] [PubMed]
- Parvin, T.; Sadras, S.R. Advanced Probiotics: Bioengineering and Their Therapeutic Application. Mol. Biol. Rep. 2024, 51, 361. [Google Scholar] [CrossRef] [PubMed]
- Merkouris, E.; Mavroudi, T.; Miliotas, D.; Tsiptsios, D.; Serdari, A.; Christidi, F.; Doskas, T.K.; Mueller, C.; Tsamakis, K. Probiotics’ Effects in the Treatment of Anxiety and Depression: A Comprehensive Review of 2014–2023 Clinical Trials. Microorganisms. 2024, 12, 411. [Google Scholar] [CrossRef] [PubMed]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells. 2023, 12, 184. [Google Scholar] [CrossRef] [PubMed]
- Fijan, S.; Frauwallner, A.; Langerholc, T.; Krebs, B.; Ter Haar (Née Younes), J.A.; Heschl, A.; Mičetić Turk, D.; Rogelj, I. Efficacy of Using Probiotics with Antagonistic Activity against Pathogens of Wound Infections: An Integrative Review of Literature. Biomed Res. Int. 2019, 2019, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus As Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, E.; Pericolini, E.; Ballet, N.; Roselletti, E.; Sabbatini, S.; Mosci, P.; Decherf, A.C.; Pélerin, F.; Perito, S.; Jüsten, P.; et al. Saccharomyces Cerevisiae-Based Probiotic as Novel Anti-Fungal and Anti-Inflammatory Agent for Therapy of Vaginal Candidiasis. Benef. Microbes. 2018, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Pericolini, E.; Gabrielli, E.; Ballet, N.; Sabbatini, S.; Roselletti, E.; Cayzeele Decherf, A.; Pélerin, F.; Luciano, E.; Perito, S.; Jüsten, P.; et al. Therapeutic Activity of a Saccharomyces Cerevisiae-Based Probiotic and Inactivated Whole Yeast on Vaginal Candidiasis. Virulence. 2017, 8, 74–90. [Google Scholar] [CrossRef]
- Pedro, N.A.; Mira, N.P. A Molecular View on the Interference Established between Vaginal Lactobacilli and Pathogenic Candida Species: Challenges and Opportunities for the Development of New Therapies. Microbiol. Res. 2024, 281, 127628. [Google Scholar] [CrossRef]
- Palmeira-de-Oliveira, R.; Palmeira-de-Oliveira, A.; Martinez-de-Oliveira, J. New Strategies for Local Treatment of Vaginal Infections. Adv. Drug Deliv. Rev. 2015, 92, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, L.; Sala, A.; Ardizzoni, A.; De Seta, F.; Singh, D.K.; Gacser, A.; Blasi, E.; Pericolini, E. Lactobacillus Acidophilus, L. Plantarum, L. Rhamnosus, and L. Reuteri Cell-Free Supernatants Inhibit Candida Parapsilosis Pathogenic Potential upon Infection of Vaginal Epithelial Cells Monolayer and in a Transwell Coculture System In Vitro. Microbiol. Spectr. 2022, 10, e02696-21. [Google Scholar] [CrossRef] [PubMed]
- Van Den Bossche, S.; Vandeplassche, E.; Ostyn, L.; Coenye, T.; Crabbé, A. Bacterial Interference With Lactate Dehydrogenase Assay Leads to an Underestimation of Cytotoxicity. Front. Cell. Infect. Microbiol. 2020, 10, 494. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Fujiwara, Y.; Thongbhubate, K.; Maeda, M.; Kanaori, K. Spore-Forming Lactic Acid-Producing Bacterium Bacillus Coagulans Synthesizes and Excretes Spermidine into the Extracellular Space. J. Agric. Food Chem. 2023, 71, 9868–9876. [Google Scholar] [CrossRef] [PubMed]
- Konuray, G.; Erginkaya, Z. Potential Use of Bacillus Coagulans in the Food Industry. Foods. 2018, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, S.N.; Yamang, H.; Lorenz, M.C.; Chew, S.Y.; Than, L.T.L. Role of Vaginal Mucosa, Host Immunity and Microbiota in Vulvovaginal Candidiasis. Pathogens. 2022, 11, 618. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhou, X.; Ren, B.; Cheng, L. The Regulation of Hyphae Growth in Candida Albicans. Virulence. 2020, 11, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Naglik, J.R.; Gaffen, S.L.; Hube, B. Candidalysin: Discovery and Function in Candida Albicans Infections. Curr. Opin. Microbiol. 2019, 52, 100–109. [Google Scholar] [CrossRef]
- Spaggiari, L.; Ardizzoni, A.; Ricchi, F.; Pedretti, N.; Squartini Ramos, C.A.; Squartini Ramos, G.B.; Kenno, S.; De Seta, F.; Pericolini, E. Fungal Burden, Dimorphic Transition and Candidalysin: Role in Candida Albicans-Induced Vaginal Cell Damage and Mitochondrial Activation in Vitro. PLoS One. 2024, 19, e0303449. [Google Scholar] [CrossRef]
- Jang, S.J.; Lee, K.; Kwon, B.; You, H.J.; Ko, G. Vaginal Lactobacilli Inhibit Growth and Hyphae Formation of Candida Albicans. Sci. Rep. 2019, 9, 8121. [Google Scholar] [CrossRef]
- Saporito-Irwin, S.M.; Birse, C.E.; Sypherd, P.S.; Fonzi, W.A. PHR1, a pH-Regulated Gene of Candida Albicans, Is Required for Morphogenesis. Mol. Cell Biol. 1995, 15, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.-M.; Harder, J. Human Beta-Defensin-2. Int. J. Biochem. Cell Biol. 1999, 31, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Järvå, M.; Phan, T.K.; Lay, F.T.; Caria, S.; Kvansakul, M.; Hulett, M.D. Human β-Defensin 2 Kills Candida Albicans through Phosphatidylinositol 4,5-Bisphosphate–Mediated Membrane Permeabilization. Sci. Adv. 2018, 4, eaat0979. [Google Scholar] [CrossRef] [PubMed]
- Makanjuola, O.; Bongomin, F.; Fayemiwo, S. An Update on the Roles of Non-Albicans Candida Species in Vulvovaginitis. J. Fungi. 2018, 4, 121. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Anti-Candida effect exerted by B. coagulans. (A) Mono-cultures and co-cultures of C. albicans (Ca), C. parapsilosis (Cp) and B. coagulans (Bc) pH values after 24 hours of incubation at 37 °C. The chart reports the mean values pH ± SEM from 3 different experiments. The range highlighted in light green represents the mean pH levels of the healthy vaginal environment. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. Ca vs Bc + Ca **** p < 0.0001. Cp vs Bc + Cp **** p < 0.0001. Effect of B. coagulans (Bc) on C. albicans (Ca) (B) and C. parapsilosis (Cp) (C) growth capacity and acidification contribution upon 24 hours of incubation at 37 °C. The graph shows the mean C. albicans (CFUx105/mL) ± SEM from three different experiments. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. Ca vs Bc+ Ca **** p < 0.0001. Cp vs Bc + Cp *** p < 0.001.
Figure 1.
Anti-Candida effect exerted by B. coagulans. (A) Mono-cultures and co-cultures of C. albicans (Ca), C. parapsilosis (Cp) and B. coagulans (Bc) pH values after 24 hours of incubation at 37 °C. The chart reports the mean values pH ± SEM from 3 different experiments. The range highlighted in light green represents the mean pH levels of the healthy vaginal environment. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. Ca vs Bc + Ca **** p < 0.0001. Cp vs Bc + Cp **** p < 0.0001. Effect of B. coagulans (Bc) on C. albicans (Ca) (B) and C. parapsilosis (Cp) (C) growth capacity and acidification contribution upon 24 hours of incubation at 37 °C. The graph shows the mean C. albicans (CFUx105/mL) ± SEM from three different experiments. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. Ca vs Bc+ Ca **** p < 0.0001. Cp vs Bc + Cp *** p < 0.001.
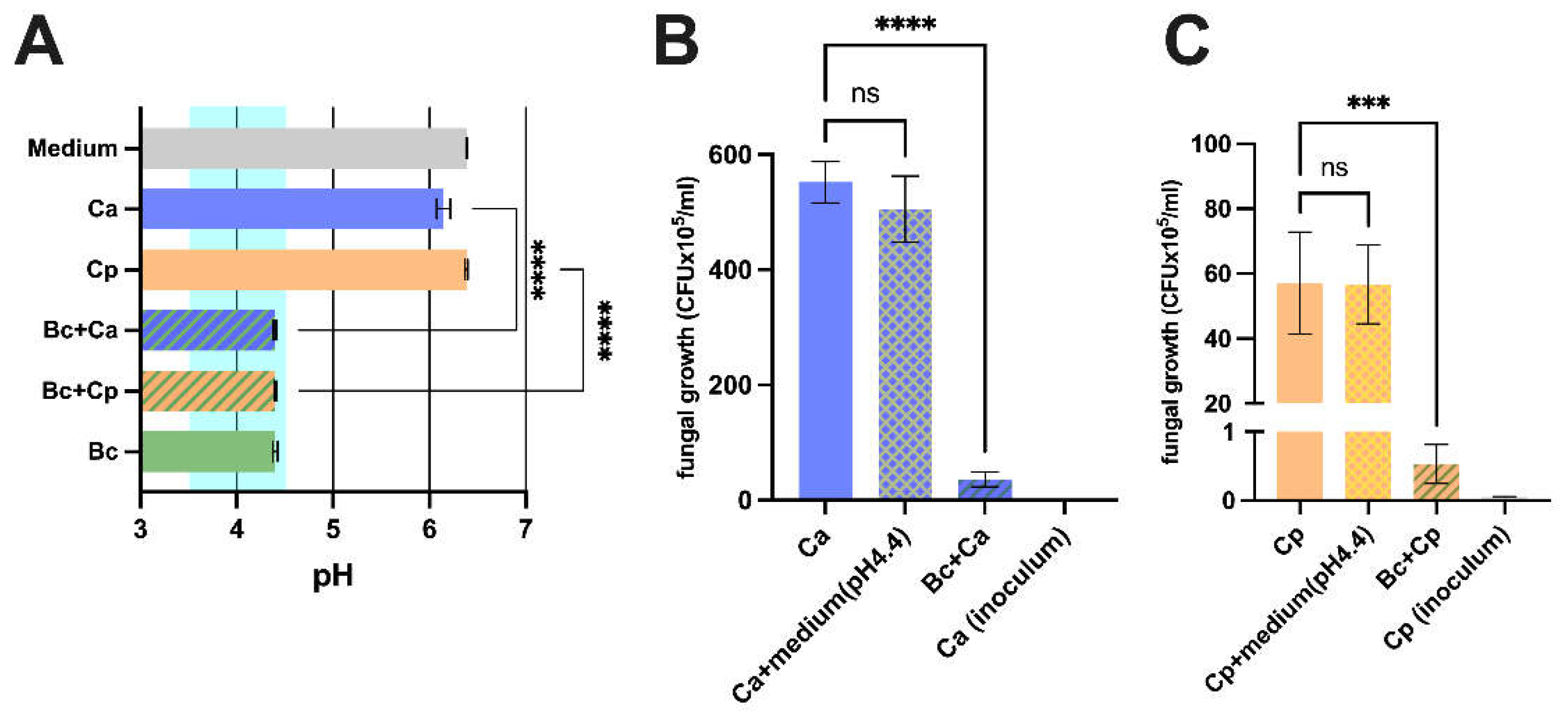
Figure 2.
Kinetic measurement of C. albicans (Ca) (A) or C. parapsilosis (B) growth when cultivated with B. coagulans CFS (Bc CFS) or a sterile medium at 37 °C. Culture OD at 570 nm wavelength was automatically detected every 120 minutes for a total 18 hours. The graphs report the mean OD values ± SEM from triplicate samples of three different experiments. The Area Under the Curve (AUC) analysis was performed on kinetic data from C. albicans (C) and C. parapsilosis (D) samples. Statistical analysis was performed on AUC values through the unpaired two-tailed Student’s t-test. Ca vs Bc CFS + Ca and Cp vs Bc CFS + Cp * p < 0.05.
Figure 2.
Kinetic measurement of C. albicans (Ca) (A) or C. parapsilosis (B) growth when cultivated with B. coagulans CFS (Bc CFS) or a sterile medium at 37 °C. Culture OD at 570 nm wavelength was automatically detected every 120 minutes for a total 18 hours. The graphs report the mean OD values ± SEM from triplicate samples of three different experiments. The Area Under the Curve (AUC) analysis was performed on kinetic data from C. albicans (C) and C. parapsilosis (D) samples. Statistical analysis was performed on AUC values through the unpaired two-tailed Student’s t-test. Ca vs Bc CFS + Ca and Cp vs Bc CFS + Cp * p < 0.05.
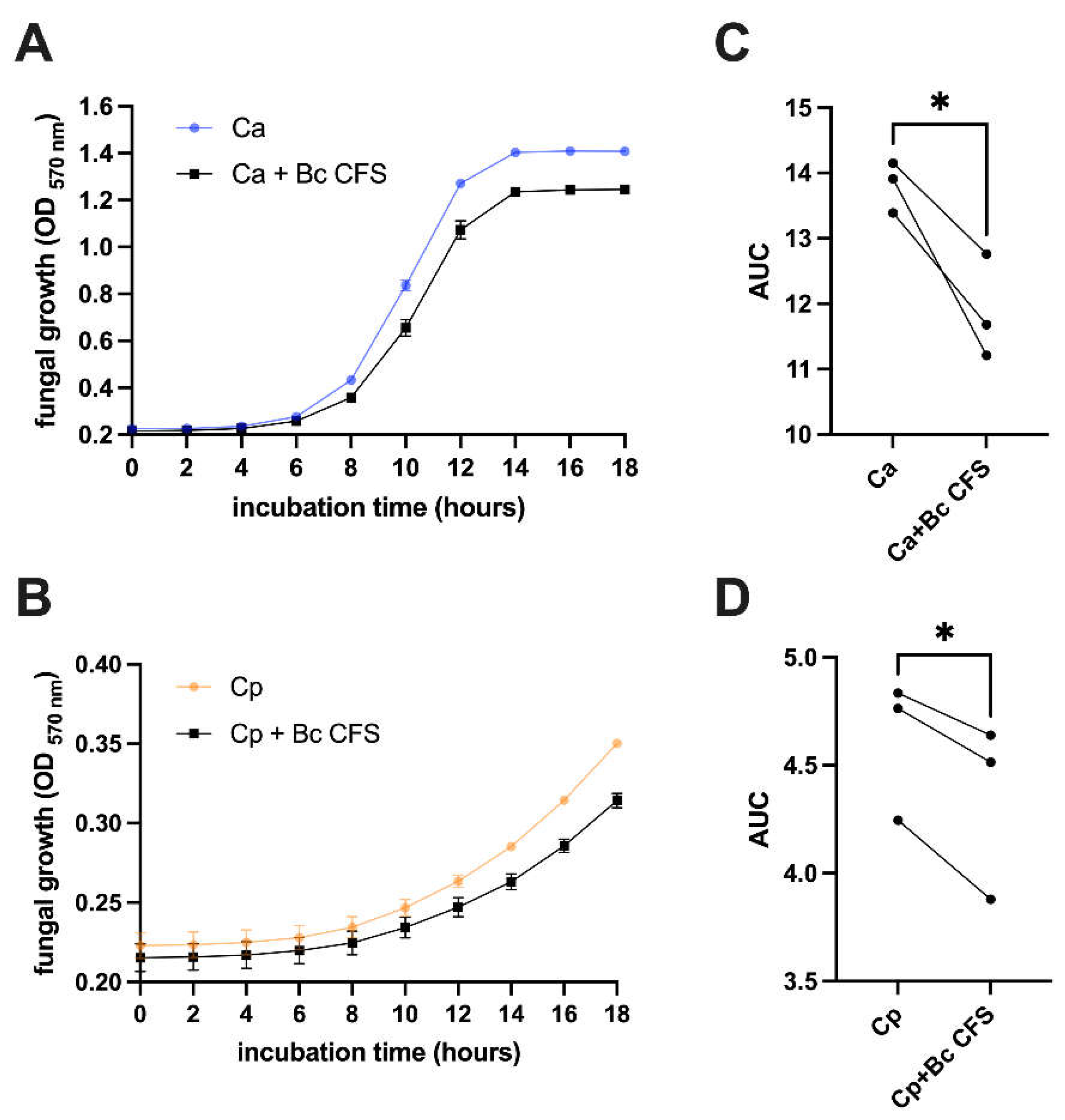
Figure 3.
Evaluation of C. albicans (Ca) adhesion capacity to a vaginal epithelial cell monolayer in the presence of B. coagulans (Bc) (A). The histogram graph shows the average % ± SEM of fungal adhesion inhibition exerted by B. coagulans (Bc). Data are from 3 independent experiments. (B) Assessment of B. coagulans (Bc) capacity to co-aggregates with C. albicans (Ca) after 1 hour of co-incubation. Boxes in the heatmap represent the score assigned to each sample in three independents experiments. 0: no aggregation; 1: aggregates with small clusters; 2: aggregates with larger numbers of yeasts; 3: clumps visible with the naked eye containing large numbers of yeast cells; 4: maximum score for large clumps visible with the naked eye in the well center. (C-E) Effect of B. coagulans (Bc) on C. albicans (Ca) hyphal formation upon 4 hours of co-incubation. Hyphal fragments were optically counted by fluorescent microscopy imaging. The fungal cell wall was stained with Uvitex 2B fluorescent dye. (C) The bars-chart reports the mean percentage ± SEM of hyphal fragments counted in three different fields from 3 independent experiments. Statistical analysis was performed by the unpaired, two-tailed Student t-test. Ca vs Bc + Ca * p < 0.05. (D) The heatmap shows the % of hyphal fragments counted in each field; the squares' color indicates the abundance of hyphae in the field (red: high % hyphal fragments; green: low % hyphal fragments). (E) Representative images from Fluorescence microscopy analysis are shown from C. albicans (Ca) or C. albicans plus B. coagulans (Bc + Ca) taken at 40X magnification.
Figure 3.
Evaluation of C. albicans (Ca) adhesion capacity to a vaginal epithelial cell monolayer in the presence of B. coagulans (Bc) (A). The histogram graph shows the average % ± SEM of fungal adhesion inhibition exerted by B. coagulans (Bc). Data are from 3 independent experiments. (B) Assessment of B. coagulans (Bc) capacity to co-aggregates with C. albicans (Ca) after 1 hour of co-incubation. Boxes in the heatmap represent the score assigned to each sample in three independents experiments. 0: no aggregation; 1: aggregates with small clusters; 2: aggregates with larger numbers of yeasts; 3: clumps visible with the naked eye containing large numbers of yeast cells; 4: maximum score for large clumps visible with the naked eye in the well center. (C-E) Effect of B. coagulans (Bc) on C. albicans (Ca) hyphal formation upon 4 hours of co-incubation. Hyphal fragments were optically counted by fluorescent microscopy imaging. The fungal cell wall was stained with Uvitex 2B fluorescent dye. (C) The bars-chart reports the mean percentage ± SEM of hyphal fragments counted in three different fields from 3 independent experiments. Statistical analysis was performed by the unpaired, two-tailed Student t-test. Ca vs Bc + Ca * p < 0.05. (D) The heatmap shows the % of hyphal fragments counted in each field; the squares' color indicates the abundance of hyphae in the field (red: high % hyphal fragments; green: low % hyphal fragments). (E) Representative images from Fluorescence microscopy analysis are shown from C. albicans (Ca) or C. albicans plus B. coagulans (Bc + Ca) taken at 40X magnification.
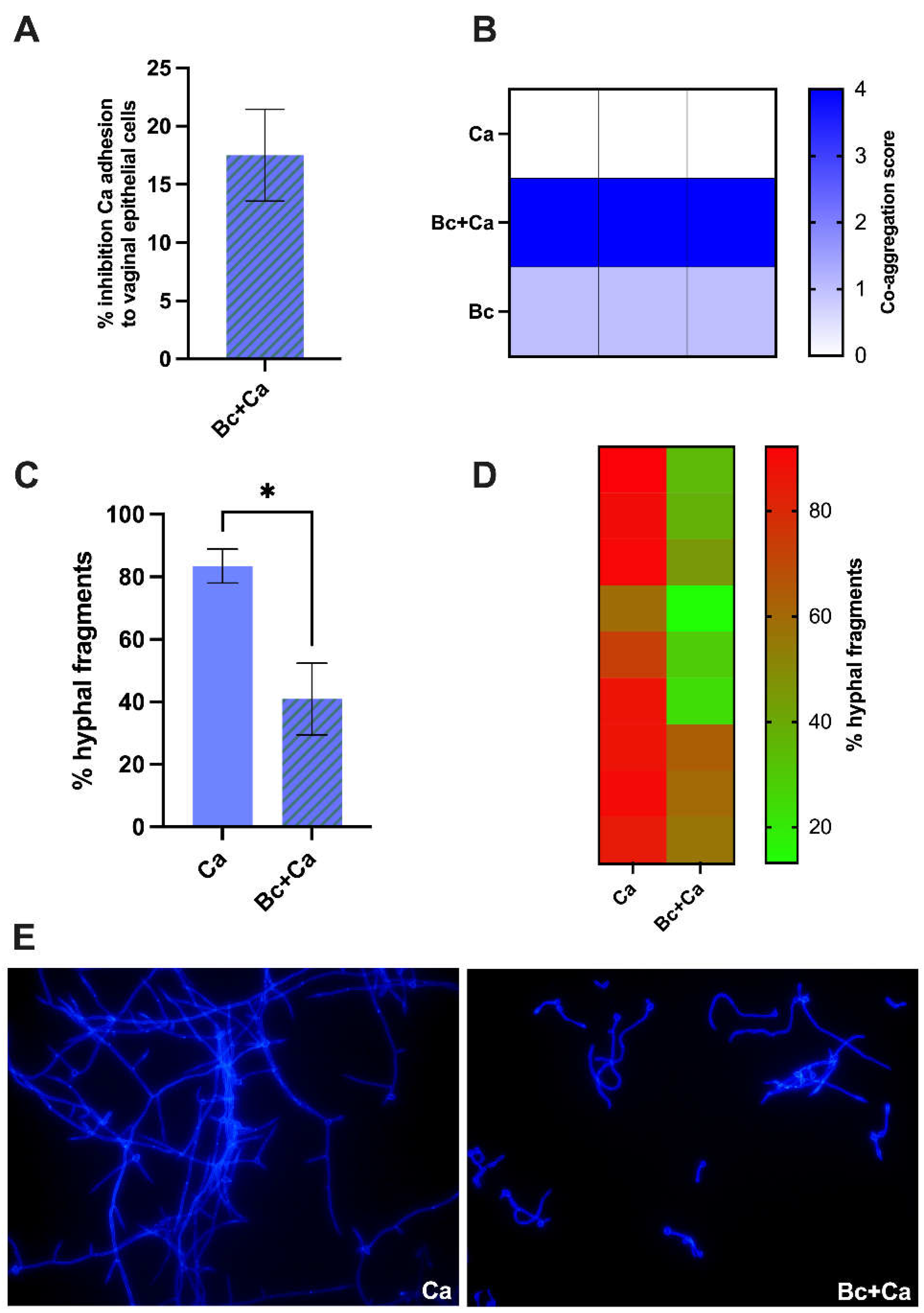
Figure 4.
(A) Percentage of vaginal cell damage pre-colonized or not by B. coagulans (Bc) for 6 hours and infected for further 18 hours with C. albicans (Ca). The chart reports the average percentage of cell damage ± SEM of triplicate samples from three different experiments. Statistical analysis was performed by the unpaired, two-tailed Student t-test. Ca vs Bc + Ca * p < 0.05. (B) Production of β-defensin-2 by vaginal epithelial cells pre-colonized or not by B. coagulans (Bc) for 6 hours and infected for further 18 hours with C. albicans (Ca). Uninfected cells (Ctrl), and cells colonized by the bacterium without C. albicans were also included in the experiments. The graph reports the mean ± SEM from three independent experiments. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. Untreated cells vs Bc pre-colonized cells * p < 0.05.
Figure 4.
(A) Percentage of vaginal cell damage pre-colonized or not by B. coagulans (Bc) for 6 hours and infected for further 18 hours with C. albicans (Ca). The chart reports the average percentage of cell damage ± SEM of triplicate samples from three different experiments. Statistical analysis was performed by the unpaired, two-tailed Student t-test. Ca vs Bc + Ca * p < 0.05. (B) Production of β-defensin-2 by vaginal epithelial cells pre-colonized or not by B. coagulans (Bc) for 6 hours and infected for further 18 hours with C. albicans (Ca). Uninfected cells (Ctrl), and cells colonized by the bacterium without C. albicans were also included in the experiments. The graph reports the mean ± SEM from three independent experiments. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. Untreated cells vs Bc pre-colonized cells * p < 0.05.
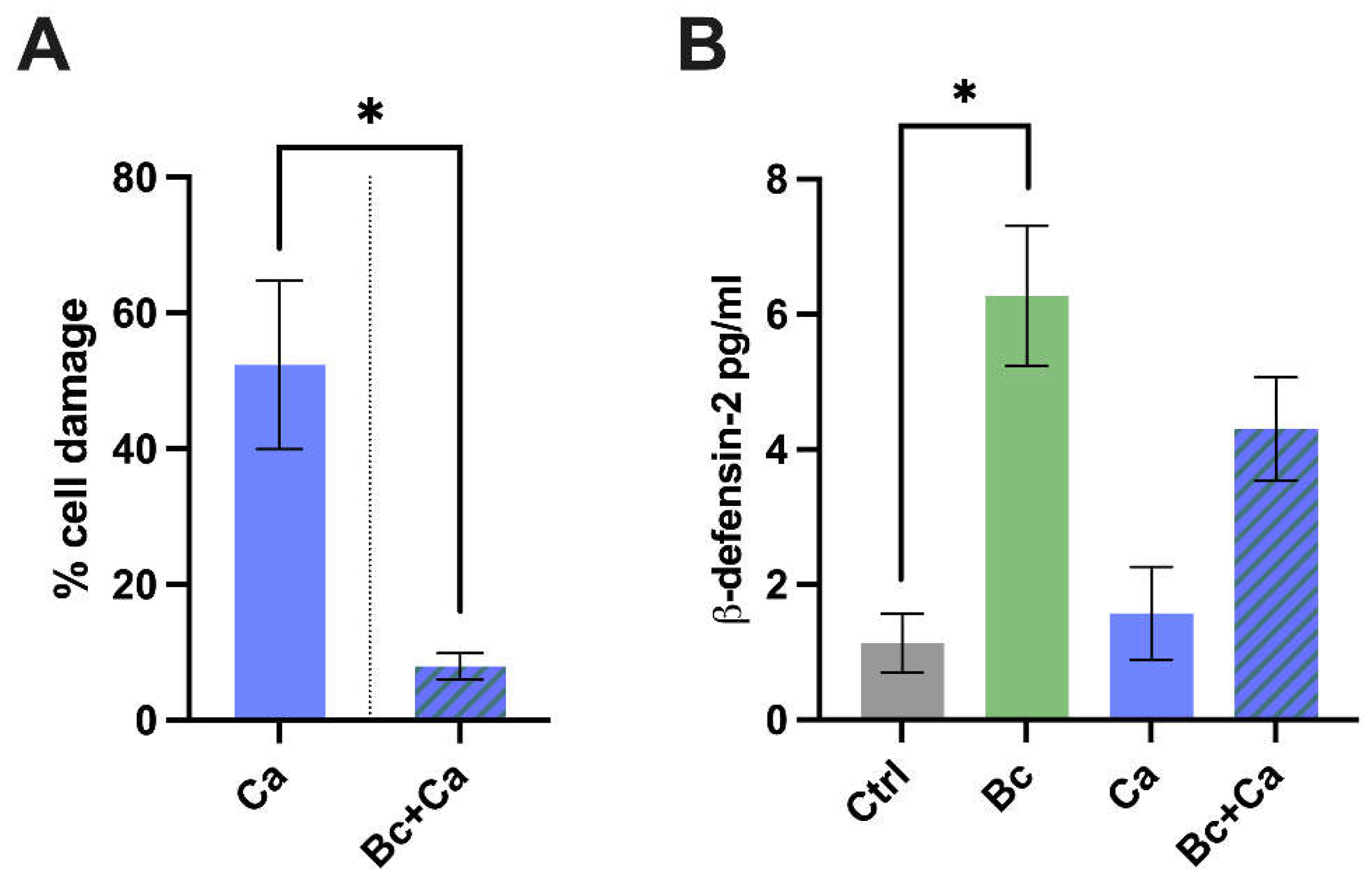
Figure 5.
Evaluation of antifungal effect permanency upon B. coagulans removal. C. albicans (Ca) (A) and C. parapsilosis (Cp) (B) metabolic activity quantification after being incubated with B. coagulans or sterile medium for 24 hours and subsequent fungal isolation and cultivation for 24 hours in the lack of bacteria. The graphs show the mean OD at 492 nm wavelength ± SEM from triplicate sample of 3 different experiments. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. (C) Capacity of B. coagulans spores to germinate on intestinal epithelial cells CaCo-2. Bacterial spores were seeded on an intestinal epithelial cell monolayer of CaCo-2 and incubated at 37 °C + 5% CO2 for 24 hours. After incubation, monolayers were photographed (upper images) and subsequently lysed. A Gram staining was then performed to visualize the presence of germinated B. coagulans (Bc) (lower images).
Figure 5.
Evaluation of antifungal effect permanency upon B. coagulans removal. C. albicans (Ca) (A) and C. parapsilosis (Cp) (B) metabolic activity quantification after being incubated with B. coagulans or sterile medium for 24 hours and subsequent fungal isolation and cultivation for 24 hours in the lack of bacteria. The graphs show the mean OD at 492 nm wavelength ± SEM from triplicate sample of 3 different experiments. Statistical analysis was performed by the one-way ANOVA test followed by the uncorrected Fisher's LSD test. (C) Capacity of B. coagulans spores to germinate on intestinal epithelial cells CaCo-2. Bacterial spores were seeded on an intestinal epithelial cell monolayer of CaCo-2 and incubated at 37 °C + 5% CO2 for 24 hours. After incubation, monolayers were photographed (upper images) and subsequently lysed. A Gram staining was then performed to visualize the presence of germinated B. coagulans (Bc) (lower images).
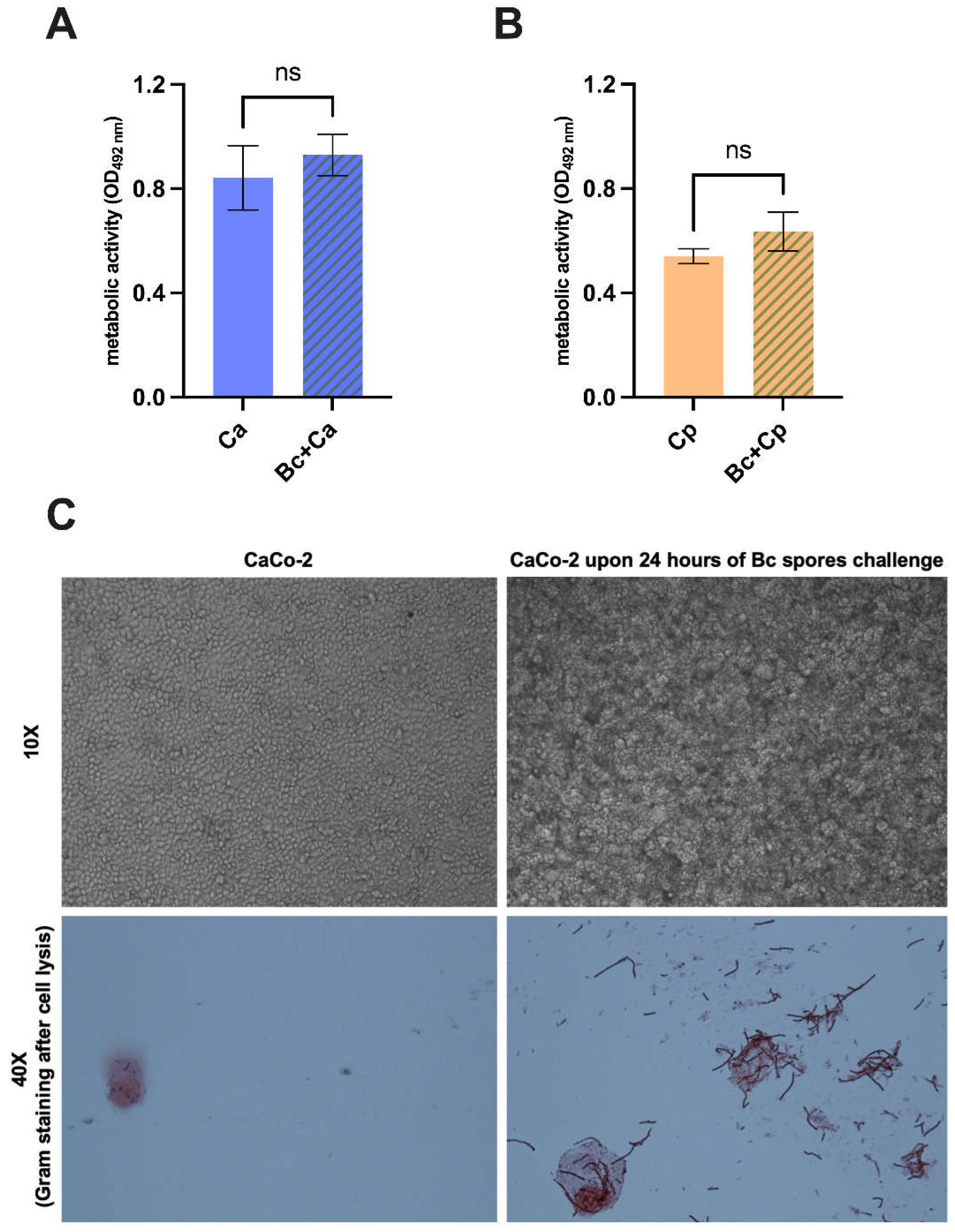
Figure 6.
Schematic representation of biological activities of B. coagulans LMG S-24828 against Candida. Created with BioRender.com.
Figure 6.
Schematic representation of biological activities of B. coagulans LMG S-24828 against Candida. Created with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



