
Submitted:
02 August 2024
Posted:
05 August 2024
You are already at the latest version
Abstract
Canola (Brassica napus L) is a profitable grain crop for Australian growers. However, weeds remain a major constraint for its production. Chemical herbicides are used for weed control, but this tactic also leads to the evolution of herbicide resistance in different weed species. Suppression of weeds by crop competition and allelopathic mechanisms has been receiving significant attention. Here, the weed suppressive ability, and associated functional traits and stability of four selected canola genotypes (PAK85388-502, AV-OPAL, AV-GARNET and BAROSSA) were examined at different locations of NSW, Australia. Results showed that there were significant effects of canola genotypes and of genotypes by crop density interaction on weed growth. Among the tested genotypes, PAK85388-502 and AV-OPAL were the most weed suppressive and, at plant density of 10 plants/m2, they reduced weed biomass of wild radish, shepherd’s purse and annual ryegrass by more than 80%. No significant differences were found in the primary root lengths among canola varieties; however, plants of the allelopathic genotype PAK8538-502 exhibited a 35% increase in lateral root number relative to plants of non-allelopathic genotype BAROSSA. The analysis of variance revealed a significant influence of genotypes with PAK85388-502 and AV-OPAL performed the best across all the research sites. Results showed that canola genotypes PAK85388-502 and AV-OPAL were more weed suppressive than AV-GARNET and BAROSSA and may release specific bioactive compounds in their surroundings to suppress neighbouring weeds. This study provides valuable information that could be utilised in breeding programs to select weed suppressive varieties of canola in Australia. Thus, lateral root number could be a potential target trait for weed suppressive varieties. Additionally, other root architecture traits may contribute to the underground allelopathic interaction to provide a competitive advantage to the crop.
Keywords:
competition
; neighbouring plant
; root
; root exudates
; adaption
1. Introduction
Canola (Brassica napus L) is an oil seed crop belonging to the Brassicaceae family. It results from the natural hybridisation between Brassica oleracea L. and Brassica rapa L. Australia is the world’s second largest exporter of canola [2]. Canola grown in Australia meets the high expectations of exporters, domestic crushers and intensive livestock producers. However, weeds are an important biotic constraint on canola production, resulting in yield and quality losses [4,45]. Grass weeds such as annual ryegrass (Lolium rigidum), vulpia (Vulpia myuros) and wild oat (Avena fatua) are the most abundant weed species in canola crops of south-eastern Australia [30]. Weeds from Brassicaceae can contaminate canola seed samples which leads to increased levels of erucic acid and glucosinolates with the consequent reduction in canola quality due to their contamination [4,45].
Chemical herbicides and mechanical weeding have been the most frequently used methods to control weeds [16,26] and have served to keep weed infestations low, thereby improving crop productivity. Despite the significant contribution of these methods, there are also certain associated challenges. Mechanical weed control requires extra soil turn-over, which can disturb soil structure and deplete soil fertility [48]. Additionally, it is not always effective and can be expensive and lack durability [15]. Similarly, herbicide-resistant weeds, health effects and environmental concerns are the major constraints for repeated use of herbicides [7,49]. In Australia, non-chemical weed control options are highly sought after for incorporation into improved integrated weed management systems (IWMS) to overcome herbicide-resistance.
Crop plant interference against weeds involves the combined effects of plant competition and allelopathy. Competition is the negative interaction between two or more plant species for resources within a limited space and is distinct from plant allelopathic interaction [56]. The competitiveness of a plant is influenced by various morphological, phenological and agronomic traits [25,26,31,32]. Increasing crop seeding rates is one of the simplest agronomic ways to enhance crop competitiveness [35], as it increases crop canopy development and hastens nutrient use, thereby denying these resources to weeds and thus reducing weed pressure [10,51]. In contrast to competition, allelopathy is the exudation of compounds by plant roots that can suppress the growth of neighbouring plants and affects seeds and seedlings of other species located within a limited range [39]. Although most plants species, including crops, can produce and release biologically active root exudates (allelochemicals), relatively few have strong allelopathic properties. Several bioactive compounds were isolated from strongly-allelopathic canola genotypes [3]. These phytotoxic or signalling chemicals presumably resulted in the observed inhibitory effects on annual ryegrass (L. rigidum) under laboratory conditions, and also may be responsible for the significant suppression of other weed species in the field [5]. Allelochemical concentrations are a function of the density of the allelopathic crop [9]. This suggests that density may be an important factor in enhancing canola allelopathic activity.
Crop species and genotypes within the species differ in root traits and their ability to compete for below-ground resources, which in turn can influence aboveground traits and yield [17,55]. Root and shoot architecture including length, biomass, number of lateral roots and growth play a crucial role in competition and crop performance [6,18,23,41]. Hence, screening crop varieties with plant functional traits may offer new insights into plant allelopathy. Further, plants can perceive different external and internal signals from their surroundings, while changes of environmental conditions subsequently can affect plant allelopathy and its functional traits [40]. Huang et al. [28] demonstrated that Merremia boisiana can adjust its resource allocation to allelopathy and leaf functional traits to adapt to varying environments. As one of the key factors affecting plant allelopathy and the functional traits, environmental changes have long been assumed critical. However, the link between canola allelopathy and its functional traits has received little attention.
Understanding how crop genotypes respond differently to changing environmental conditions is crucial and it is a significant step in developing improved crop varieties [43]. When genotypes are assessed across various locations or years, their yield and individual traits’ performances may vary significantly. The presence of substantial genotype-by-environment (G × E) interaction can further complicate comparisons and recommendations for adaptable genotypes [36]. To identify stable genotypes, it is essential to break down G × E interaction into stability statistics assigned to each genotype across different environments. Various stability indices have enabled researchers to pinpoint widely adapted genotypes for breeding programs or enhance recommendations to growers [38]. A genotype is considered most stable when it exhibits minimal fluctuation across diverse environments [38]. The present research aimed to examine 1) the functional traits and weed suppressive ability of four selected canola genotypes and 2) the stability of canola genotypes for weed suppression over different temporal and spatial conditions.
2. Materials and Methods
2.1. Experimental Site
Three field sites were chosen in southern NSW in 2016, one each at Wagga Wagga Agricultural Institute (WWAI) (147.3598° E, -35.1082°S), Temora (147.5339° E, -34.4468°S) and Marrar (147.3548° E, -34.8252°S), NSW, respectively. Prior to that, a field study was conducted in 2013 at WWAI (Asaduzzaman et al. 2014a). The Wagga Wagga sites had a high population of L. rigidum (annual ryegrass) (80 plants/m2) and the Marrar site was severely affected by Raphanus raphanistrum (wild radish).
2.2. Canola Genotypes and Sowing Density
Four canola genotypes namely PAK85388-5002, AV-OPAL, AV-GARNET and BAROSSA were selected, based on previous study by Asaduzzaman et al. [4,5]. The genotypes are open pollinated; PAK85388-502 is a breeding line and AV-GARNET is a competitive cultivar reported by Lemerle et al. [33]. A previous laboratory experiment with several canola genotypes showed that canola densities played a major role in its allelopathic activity in suppressing annual ryegrass root growth [5]. Hence for each genotype, four different sowing rates (15, 30, 60 and 120 seeds/m2) were used in 1.8 m x 10 m plots with 4, 5 and 5 replications for Marrar 2016, Temora 2016 and Wagga Wagga 2016 sites, respectively. The Wagga Wagga 2013 field site had 6 replications [4]. The purposes were to determine canola density effects on weed growth and reproductive development.
2.3. Pre-Sowing Knockdown Herbicide and Sowing
Glyphosate (450 g/L Glyphosate) at 2 L/ha was applied at all sites as pre-planting approach and no other herbicides were used during the experiment. Crop was sown with a plot seeder together with a basal fertiliser of Croplift 15 (Incitec Pivot Fertilisers™) at 120 kg/ha. This basal fertiliser was applied below the canola seed at sowing and canola seeds were treated with Jockey® (167 g/L fluquinconazole) at the recommended rate (2 L/100 kg seed) to control blackleg (Leptosphaeria maculans).
2.4. Data Collection to Assess Weed Suppressive Ability of Canola Genotypes
Weed numbers and biomass (as an indicator of weed seed bank replenishment), botanical compositions of weeds with respective genotypes, shoot and root architecture were considered the most important metrics for allelopathic and interference effects in Wagga Wagga 2016 and Marrar 2016 sites (Table 1). Weed numbers were assessed using two quadrats (0.5 x 0.5 m) per plot at Wagga Wagga 2013, 2016 and Marrar 2016 site. Quadrats were placed at random within each plot, but obvious weed patches were avoided. Weed biomass assessment was undertaken using the two quadrats per plot when canola biomass was estimated to be maximal, corresponding with early flowering in canola at Wagga Wagga 2013; 2016, Temora 2016 and Marrar 2016 sites. Canola root and shoot measurements were taken just prior to biomass cut by measuring 20 random plants per genotype at Wagga Wagga 2016 and Marrar 2016 sites. A total of 20 plants were randomly selected and were carefully dug up and the lengths of the canola root and shoot were measured. Then, plants were transported to a shade area of WWAI where the number of lateral roots were counted. The relative root growth (RRG) of 20 plants for each genotype was calculated as follows: RRYP = Rij/Rii,where Rii is the canola root biomass in the weeds-free condition and Rij is the biomass of canola roots collected from heavily weed-infested plots. Leaf Area Index (LAI) and Photosynthetic Active Radiation (PAR) were calculated based on Ceptometer data, where Transmitted PAR = Below-canopyPAR/Above-canopy PAR. The NDVI and Ceptometer readings were measured from the treatment plots at 60 plant density/m2 for each genotype, just before canola flower initiation.
The Stem-Specific Density (SSD) (g cm–3 or kg dm–3) was measured at canola flowering time. Briefly main stem oven-dry mass (at 70°C for 72 h) of a canola plant measured and divided by the volume of the same section. Where the volume of a canola stem was determined simply by measuring its total length and its diameter on three places along the freshly harvested stem using callipers. SSD is emerging as a core functional trait because of its importance for the stability, defence, architecture, C gain and growth potential of plants. A low stem density (with large vessels) leads to a fast growth, whereas a high stem density (with small vessels) leads to a high survival, because of biomechanical and hydraulic safety [47].
2.5. Statistical Analysis
The field experiments were conducted with randomised complete block designs. The broad-leaf weed numbers were not analysed separately but all broad-leaf weeds were included in the weed biomass calculations and ANOVA was employed for biomass data analysis against canola plants number. To identify the most successful genotype for weed suppression within each environment effect, R packages including metan [36] was used intensively for data analysis. A joint-ANOVA was performed by using the following formula.
where yijk is the response variable (e.g., weed biomass) observed in the kth block of the ith genotype in the jth environment (i = 1, 2, . . . , g; j = 1, 2, . . . , e; k = 1, 2, . . . , b); µ is the grand mean; αi is the effect of the ith genotype; τj is the effect of the jth environment; (ατ )ij is the interaction effect of the ithgenotype with the jth environment; γjk is the effect of the kth block within the jth environment; and εijk is the random error. The stability analysis was conducted using ANOVA, incorporating the non-parametric Shukla’s stability variance parameter [50] to assess the general superiority of genotypes across different locations. Additionally, the commonly used regression model proposed by Eberhart and Russell [22] for stability analysis of genotypes was employed where the dependent variable is predicted as a function of an environmental index, according to the following model:
where β0i is the grand mean of the genotype i(i = 1, 2, . . . , I); β1 i is the linear response (slope) of the genotype i to the environmental index Ij (j = 1, 2, . . . , e); δij is the deviation from the regression line for the i-th genotype in the j-th environment, indicating the specific interaction between genotype and environment and εij is the random error. Finally, the additive main effects and multiplicative interaction (AMMI) biplot was used (G x E) to visually represent genotype performances and environmental interactions on weed biomass production.
yijk = µ + αi + τj + (ατ ) ij + γjk + εijk --------------------1
Yij = β0i + β1iIj + δij + ¯εij ---------------------------------2
3. Results
3.1. Canola and Weed Biomass
The composition of weed species differed between sites. Trial sites at Temora and Wagga Wagga 2013 and 2016 were mainly infested by annual ryegrass and, at the early seedling stage of canola, there was no significant (p>0.005) difference between genotypes in annual ryegrass densities (data not shown). The Marrar site was affected mostly by wild radish [WR] followed by shepherd’s purse [SP], annual ryegrass [ARG] and other broadleaf weeds [BW]. Also, the total weed densities [WR+SP+ARG+BW] at the early growth stage of canola was not significantly (p>0.05) different between genotypes. There was a significant (p<0.001) interaction effect between canola density and genotype on total weed biomass (Figure 1). A significant negative correlation was observed between weed biomass and canola densities (Figure 1). At 10 canola plants/m2, the total weed dry biomass was only 17 g and 10 g/m2 for the genotype PAK85388-502, and 25 g and 22 g for AV-OPAL at Wagga Wagga 2016 and Marrar 2016 sites, respectively. Genotype BAROSSA was less suppressive on weed growth; this genotype even at high density (10 canola plants/m2) produced double the weed biomass (48 g) of AV-OPAL at Wagga Wagga 2016 site.
3.2. Plant Functional Traits
The multivariate pair analysis showed that canola root length was not significantly different among genotypes (data not shown). Negative relationships between the number of lateral canola roots (r = -0.18) or specific canola stem density (r = -0.19) and weed biomass were observed. Genotype PAK85388-502 had the most lateral root/plant followed by AV-OPAL and Barossa, whereas AV-GARNET and BAROSSA had the most Relative Root Growth (RRG) in both experimental sites (Table 2). The stem density of canola did not differ significantly between genotypes (p = 0.12), environments (p = 0.58) and G x E (p = 0.89) (data not shown). Stem length (p<0.05) and SSD (p = 0.005) were significantly different between genotypes where PAK85388-502 had the longest stem followed by AV-GARNET, while BAROSSA had the largest SSD value.
The fractional ground cover (estimated by the NDVI) showed that genotype effects were not significant (p = 0.06). Additionally, the LAI and transmitted PAR were not significant (p>0.05) among the genotypes (Table 2). Numerically, the highest leaf area (1.2) was produced by BAROSSA followed by the competitive and allelopathic genotypes AV-GARNET and PAK85388-502 respectively. The light interception capability was higher in AV-OPAL (0.54) followed by AV-GARNET and PAK85388-502.
3.3. Environmental Effect and Stability of Genotypes for Weed Suppression
The interactions between genotypes and environments measure the spatial (locations) and temporal (years) separation and/or combination of these factors. The combined analysis of variance across environments showed that variances due to genotypes (G), environments (E), and G x E were highly significant (p < 0.001) for weeds biomass. So, it was appropriate to explore such interaction including ANOVA and linear regression-based stability analysis. Table 3 shows that weed biomass was 6 and 5 times higher under unfavourable conditions than under favourable conditions for BAROSSA and AV-GARNET respectively. For PAK85388-502 and AV-OPAL, weed biomass was 5 and 4.5 higher under unfavourable conditions compared with favorable conditions. The best weed suppressive genotypes for both favourable and unfavourable condition were PAK85388-502 followed by AV-OPAL. Genotype BAROSSA was the least weed suppressive in either condition. The regression model shows that slopes (b1) of both AV-OPAL and PAK85388-502 were flatter than the other two varieties. The low co-efficient (b0) value and significant differences between these two genotypes (AV-OPAL and PAK85388-502) were due mostly to the genotypic effect rather than the environmental effect.
The top genotypes for weed suppression were PAK85388-502 and AV-OPAL at all four sites (Table 3 and Figure 3). The computed genotype-environment effects or genotype plus genotype-environment effects showed no clear change except Shukla ranking in the rank order of genotypes across environments. PAK85388-502 and AV-OPAL were the most stable genotypes for weed suppression across different environments.
Among the locations, Temora 2015 site achieved most weed suppression by the four canola genotypes, and they adapted very well compared to other three sites (Figure 3). Wagga 2013 site was the least weed suppressive and adaptability of the genotypes varied. The additive main effects (canola genotype), multiplicative interaction (AMMI) analysis, and biplot of genotype and environment (location) of four genotypes were used to evaluate weed biomass production. The principal component 1 (PC1) was significant (p = 0.001), with PC1 axis explaining 99% variation of the G × E (Figure 4). The two weed suppressive canola genotypes (AV-OPAL and PAK85388-502) tended to be clustered together in the AMMI biplot. They were placed closer than the other two canola genotypes (AV-GARNET and BAROSSA) and produced less weed biomass in their plots across the different locations. Wagga 2013 site had the highest weed biomass.
4. Discussions
The study of plant interference is increasingly popular although the current systems of plant breeding largely ignore the abilities of a variety to exercise control over its weed challengers. The difficulty in studying interactions between plants is due to the complex nature of plant interference, defined as the combined effect of competition and allelopathy. However, the development of crops with the capability to exert allelopathic effects on weeds through root exudates is an attractive prospect [37. Research has shown this potential in wheat [53], barley [11,12], rice [19,20,21,24,46] and canola [5]. In this study, genotypes such as AV-OPAL, PAK85388-502, AV-GARNET and BAROSSA produced similar crop biomass but the genotypes PAK85388-502 and AV-OPAL tended to result in weed biomass lower than even the strongly competitive genotype (cv. AV-GARNET). Therefore, weed-suppressing genotypes have potential for integrated weed management and further work is needed to produce a genotype with combined competitive ability and allelopathy. Bertholdsson [12] found that early weed biomass was significantly lower in the highly allelopathic wheat lines compared with the non-allelopathic lines. Also, our research revealed that weed biomass was suppressed during the early growth stages of allelopathic canola genotypes, resulting in significantly less weed biomass harvested at the end of the experiment.
Four Brassica genotypes (AV-OPAL, AV-GARNET, BAROSSA PAK85388-502) showed similar patterns in the density-response curve for weed growth and there was a density by genotype interaction. This indicates that crop density plays a role in canola weed suppression. These results were consistent with previous research in rice [46] and in wheat [36]. The aboveground canopy of all tested genotypes had similar contributions to weed control. However, beyond that, the additional mechanism such as the allelopathy of AV-OPAL and PAK85388-502 might played a role for their neighbouring weed suppression. AV-OPAL was identified as a less vigorous genotype with shorter plant height among other canola genotypes [5]. Asaduzzaman et al. [6] collected 70 international rapeseed varieties and evaluated their allelopathic potential by growing them in close proximity to L. rigidum. Rapeseed was sown at 10, 20, and 30 plants/m2 against 15 plants/m2 of annual ryegrass. Generally, the higher density of rapeseed resulted in higher suppression of L. rigidum. The varieties with strong allelopathic activity were PAK85388-502, AV-OPAL, BLN3343CO0402, and RIVETTE. Many of these genotypes categorised as allelopathic those can released a range of allelochemicals through their root exudates of canola, with key chemicals being sinapyl alcohol, p-hydroxybenzoic acid and 3,5,6,7,8-pentahydroxy flavones [3]. Furthermore, in Canada, canola competitiveness was improved by choice of variety and use of higher seeding rates [8]. Correlation coefficient is a measure of the extent and direction of the relationship between any two traits (variables). The negative relationships between root (number of lateral roots) and shoot (stem density, and stem length) functional traits and weed biomass suggest that these traits are important components for weed suppression by canola genotypes. Therefore, their simultaneous selection will be a good approach to increasing weed suppressive ability. This same relationship was observed by Afuape et al. [1]. Genetic variability is essential for selection [44]. However, a wide range of varieties should be evaluated at different locations to further verify such relationships.
The stability analysis aims at helping breeders identify which genotypes have specific and/or general adaptability to various production environments. Additionally, stability analysis helps determine the test environments for future evaluations of canola production with self-weeding capability. Three locations (excluding Wagga 2013) clustered in the same quadrants, indicating that these locations share similarities in terms of weed growth and canola genotype performance. The Temora environment and Wagga 2016 were higher performing environments for weed suppression compared with others. This result means that testing data from one location can represent the performance of same materials in the other similar locations. Therefore, conducting a stability analysis with a large set of canola genotypes will further help identify specific genotypes for both locations, as well as a stable genotype that can be cultivated across multiple locations and particularly locations that share similar attributes to the test locations. Understanding the environmental conditions in which canola varieties are tested is crucial for accurately interpreting results and extrapolating findings to diverse geographical locations or growing conditions. Highlighting higher-performing environments for weed suppression, such as the Temora environment and Wagga 2016, underscores the need for a comprehensive description of these locations' environmental characteristics. To address this gap, future research should prioritise conducting stability analyses with a diverse range of canola genotypes across various locations. This approach would enable breeders to identify specific genotypes suited for different environments and stable genotypes adaptable to multiple locations, facilitating informed decision-making in canola allelopathic variety cultivation.
The weed suppressive ability of a specific canola variety might be weed-specific and likely associated with a genetic as well as an environmental component of variance; breeding will be required for its maintenance [11]. Combining a variety’s capability in reducing specific weed pressure with optimal agronomic practices that facilitate crop health will generally enhance cropping system sustainability and allow growers to extend the life of valuable herbicides [27]. However, the feasibility of using a variety for specific weed management may not be useful for economic outcomes because canola crops are infested naturally by a range of weed species [30,33]. However, a competitive variety of canola could be developed by incorporating traits such as increased lateral roots, stem length and density without compromising other desirable traits such as grain yield, quality or disease resistance [31]. Canola weed suppressiveness can be optimised to reduce specific weed growth and reproduction through farming practices that allow implementation of a variety of cultural techniques such as sowing crops with different planting dates to reduce other weeds pressure [51]. In addition, the ability of crops to suppress weeds appears to be strongly variety dependent [14; 29; 54).
Here, the experiments reported reinforce the need for the preservation of the older varieties such as AV-OPAL and PAK85388-502 (highly allelopathic) and AV-GARNET (highly competitive [31]) so that these benefits can be incorporated into new varieties. Also, this research highlights the need for new varieties to be evaluated for their interference capabilities in weedy field plots without herbicides. The information obtained in this future research will allow producers to broaden their armoury against herbicide resistance by choosing weed-suppressive varieties, thereby helping manage herbicide-resistant weeds which threaten productivity, profitability and food-security. The capability of crop variety to suppress weeds is being considered as a preferred criterion for cultivar selection in many parts of the world [52]. The allelopathic potential of crop plants contributes to the weed suppressing ability of the variety. Weed-suppressive allelopathic varieties can reduce weed infestation and improve the efficacy of inputs and increase the methods of weed control.
5. Conclusion
The current study demonstrated considerable variation among the canola genotypes across the studied traits. The observed genotype-by-environment (G x E) effects influenced the average ranks of genotypes in different environments, highlighting the necessity for multi-environment testing before making informed decisions. Furthermore, the G x E biplot analysis identified two distinct mega-environments (Wagga Wagga 2013 vs others), indicating significant interactions between genotypes and environments that affect trait expression for evaluating canola genotypes in Australia and identified PAK85388-502 and AV-OPAL as the best performing genotypes for weed suppression. These genotypes were stable and adaptable across test environments and could be used as parental materials for further genetic improvement through plant breeding. BAROSSA and AV-GARNET were more competitive but might be less allelopathic and the other two varieties might be more allelopathic but less competitive. Therefore, two separate traits potentially could be combined through breeding programs. The current study determined the magnitude of genotype by environment interaction and stability for weed suppression ability of canola genotypes. Since most grain crop breeding programs often are tailored toward the development of high yielding, biotic and abiotic resistance and/or tolerance, this work has identified novel genotypes which could be used to breed weed suppressive varieties in the future.
Author Contributions
Md Asaduzzaman: Conceptualization; data curation; formal analysis; investigation; writing—original draft; writing—review and editing. Hanwen Wu: Funding acquisition; investigation; writing—review and editing. Greg Doran: Funding acquisition; investigation; writing—review and editing. Jim Pratley: Writing-review and editing.
Acknowledgments
The research team acknowledges the financial support (Research Centre Fellowship) of Graham Centre, CSU. We also acknowledge the input of technical assistance provided by Adam Shephard, and Michael Hopwood, NSWDPI. David Officer and Bernie Dominiak reviewed a pre-submission version of the manuscript.
Conflict of Interest
The authors declare that they have no conflict of interest.
References
- Afuape, S.O.; Okocha, P.I.; Njoku, D. Multivariate assessment of the agro-morphological variability and yield components among sweet potato [Ipomoea batatas (L.) Lam] landraces. Afr. J. Plant Sci. 2011, 5, 123–132.
- Australian Oilseeds Federation (AOF). The State of Oilseed Industry in Australia: Annual Report 2023. Available online: https://www.graincentral.com/news/aof-forecasts-new-crop-canola-area-at-3-2mha/ (accessed on [Date]).
- Asaduzzaman, M.; Pratley, J.E.; An, M.; Luckett, D.J.; Lemerle, D. Metabolomics differentiation of canola genotypes: toward an understanding of canola allelochemicals. Front. Plant Sci. 2015, 5, 765–765. [CrossRef]
- Asaduzzaman, M.; Luckett, D.J.; Cowley, R.B.; An, M.; Pratley, J.E.; Lemerle, D. Canola cultivar performance in weed-infested field plots confirms allelopathy ranking from in vitro testing. Bio. Sci. Tech. 2014, 24, 1394–1411.
- Asaduzzaman, M.; An, M.; Pratley, J.E.; Luckett, D.J.; Lemerle, D. Canola (Brassica napus) germplasm shows variable allelopathic effects against annual ryegrass (Lolium rigidum). Plant Soil 2014, 380, 47–56. [CrossRef]
- Asseng, S.; Anderson, G.C.; Dunin, F.X.; Fillery, I.R.P.; Dolling, P.J.; Keating, B.A. Use of the APSIM wheat model to predict yield, drainage, and NO3- leaching for a deep sand. Aust. J. Agric. Res. 1998, 49, 363. [CrossRef]
- Beckie, H.J.; Harker, K.N.; Legere, A.; Morrison, M.J.; Swartz, G.S.; Falk, K.C. GM canola: the Canadian experience. Farm. Pol. 2011, 8, 43–49.
- Beckie, H.J.; Johnson, E.N.; Blackshaw, R.E.; Gan, Y. Weed Suppression by Canola and Mustard Cultivars. Weed Technol. 2008, 22, 182–185. [CrossRef]
- Belz, R.G.; Hurle, K. Differential Exudation of Two BenzoxazinoidsOne of the Determining Factors for Seedling Allelopathy of Triticeae Species. J. Agric. Food Chem. 2005, 53, 250–261. [CrossRef]
- Berkowitz, A.R. Competition for resources in weed-crop mixtures. In: Altieri, M.A.; Liebman, M., Eds.; Weed Management in Agroecosystems: Ecological Approaches; CRC Press Inc: Boca Raton, FL, USA, 1988; pp. 89–119.
- Bertholdsson, N. Use of multivariate statistics to separate allelopathic and competitive factors influencing weed suppression ability in winter wheat. Weed Res. 2011, 51, 273–283. [CrossRef]
- Bertholdsson, N. Breeding spring wheat for improved allelopathic potential. Weed Res. 2010, 50, 49–57. [CrossRef]
- Bertholdsson, N. Early vigour and allelopathy – two useful traits for enhanced barley and wheat competitiveness against weeds. Weed Res. 2005, 45, 94–102. [CrossRef]
- Bertin, C.; Yang, X.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [CrossRef]
- Bond, W.; Grundy, A.C. Non-chemical weed management in organic farming systems. Weed Res. 2001, 41, 383–405. [CrossRef]
- Chauvel, B.; Guillemin, J.-P.; Gasquez, J.; Gauvrit, C. History of chemical weeding from 1944 to 2011 in France: Changes and evolution of herbicide molecules. Crop. Prot. 2012, 42, 320–326. [CrossRef]
- Chen, Y.-T.; Wang, Y.; Yeh, K.-C. Role of root exudates in metal acquisition and tolerance. Curr. Opin. Plant Biol. 2017, 39, 66–72. [CrossRef]
- de Dorlodot, S.; Forster, B.; Pagès, L.; Price, A.; Tuberosa, R.; Draye, X. Root system architecture: opportunities and constraints for genetic improvement of crops. Trends Plant Sci. 2007, 12, 474–481. [CrossRef]
- Dilday, R.H.; Yan, W.G.; Moldenhauer, K.A.K.; Gravois, K.A. Allelopathic activity in rice for controlling major aquatic weeds. In: Olofsdotter, M., Ed.; Allelopathy in Rice; International Rice Research Institute: Los Baños, Philippines, 1998; pp. 7–26.
- Dilday, R.H.; Gealy, D.R.; Mattice, J.D.; Moldenhauer, K.A. Allelopathy in rice as a weed control strategy. In: Abstracts of the 3rd International Weed Science Congress; International Weed Science Society: 2000; pp. 33–34.
- Dilday, R.H.; Mattice, J.D.; Moldenhauer, K.A.; Yan, W. Allelopathic Potential in Rice Germplasm Against Ducksalad, Redstem and Barnyard Grass. J. Crop. Prod. 2001, 4, 287–301. [CrossRef]
- Eberhart, S.A.; Russell, W.A. Stability Parameters for Comparing Varieties. Crop. Sci. 1966, 6, 36–40. [CrossRef]
- Gahoonia, T.S.; Nielsen, N.E. Root traits as tools for creating phosphorus efficient crop varieties. Plant Soil 2004, 260, 47–57. [CrossRef]
- Gealy, D.; Ottis, B.; Talbert, R.; Moldenhauer, K.; Yan, W. Evaluation and improvement of allelopathic rice germplasm at Stuttgart, Arkansas, USA. In: Proceedings of Fourth World Congress on Allelopathy; Charles Sturt University: Wagga Wagga, Australia, 2005; August 21–26. Published on CD-ROM.
- Harker, K.N.; Clayton, G.W.; Blackshaw, R.E.; O’donovan, J.T.; Stevenson, F.C. Seeding rate, herbicide timing and competitive hybrids contribute to integrated weed management in canola (Brassica napus). Can. J. Plant Sci. 2003, 83, 433–440. [CrossRef]
- Harker, K.N.; O’Donovan, J.T. Recent weed control, weed management, and integrated weed management. Weed Technol. 2013, 27, 1–110.
- Harker, K.N.; O'Donovan, J.T.; Turkington, T.K.; Blackshaw, R.E.; Lupwayi, N.Z.; Smith, E.G.; Klein-Gebbinck, H.; Dosdall, L.M.; Hall, L.M.; Willenborg, C.J.; et al. High-yield no-till canola production on the Canadian prairies. Can. J. Plant Sci. 2012, 92, 221–233. [CrossRef]
- Huang, Z.; Xu, W.; Yu, K. Bidirectional LSTM-CRF models for sequence tagging. arXiv preprint arXiv:1508.01991.
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop. Prot. 2015, 72, 57–65. [CrossRef]
- Lemerle, D.; Blackshaw, R.E.; Smith, A.B.; Potter, T.D.; Marcroft, S.J. Comparative survey of weeds surviving in triazine-tolerant and conventional canola crops in south-eastern Australia. Plant Protect. Quart. 2001, 16(1), 37–40.
- Lemerle, D.; Luckett, D.J.; Lockley, P.; Koetz, E.; Wu, H. Competitive ability of Australian canola (Brassica napus L.) genotypes for weed management. Crop Pasture Sci. 2014, 65, 1300–1310.
- Lemerle, D.; Luckett, D.J.; Wu, H.; Widderick, M.J. Agronomic interventions for weed management in canola (Brassica napus L.) – A review. Crop. Prot. 2017, 95, 69–73. [CrossRef]
- Lemerle, D.; Verbeek, B.; Coombes, N.; Loss, S.; van Burgel, A. Impact of weeds on crop production in Australia. Weed Technol. 2011, 25(2), 195–202.
- Li, Y.; Allen, V.G.; Chen, J.; Hou, F.; Brown, C.P.; Green, P. Allelopathic Influence of a Wheat or Rye Cover Crop on Growth and Yield of No-Till Cotton. Agron. J. 2013, 105, 1581–1587. [CrossRef]
- Mohler, C.L. Enhancing the competitive ability of crops. In Liebman, M., Ed.; Ecological Management of Agricultural Weeds; Cambridge University Press: Cambridge, UK, 2001, pp. 302–305.
- Olivoto, T.; Lúcio, A.D. metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [CrossRef]
- Olofsdotter, M.; Jensen, L.B.; Courtois, B. Improving crop competitive ability using allelopathy — an example from rice. Plant Breed. 2002, 121, 1–9. [CrossRef]
- Reckling, M. Methods of yield stability analysis in long-term field experiments. A review. Agron. Sustain. Dev. 2021, 41, 27.
- Rice, E.L. Allelopathy; Academic Press: Orlando, FL, 1984.
- Rivoal, A.; Fernandez, C.; Greff, S.; Montes, N.; Vila, B. Does competition stress decrease allelopathic potential? Biochem. Syst. Ecol. 2011, 39, 401–407.
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [CrossRef]
- Rueda-Ayala, V.P.; Rasmussen, J.; Gerhards, R.; E Fournaise, N. The influence of post-emergence weed harrowing on selectivity, crop recovery and crop yield in different growth stages of winter wheat. Weed Res. 2011, 51, 478–488. [CrossRef]
- Sabri, R.S.; Rafii, M.Y.; Ismail, M.R.; Yusuff, O.; Chukwu, S.C.; Hasan, N. Assessment of Agro-Morphologic Performance, Genetic Parameters and Clustering Pattern of Newly Developed Blast Resistant Rice Lines Tested in Four Environments. Agronomy 2020, 10, 1098. [CrossRef]
- Sarif, H.M.; Rafii, M.Y.; Ramli, A.; Oladosu, Y.; Musa, H.M.; Rahim, H.A.; Zuki, Z.M.; Chukwu, S.C. Genetic diversity and variability among pigmented rice germplasm using molecular marker and morphological traits. Biotechnol. Biotechnol. Equip. 2020, 34, 747–762. [CrossRef]
- A Salisbury, P.; Potter, T.D.; Gurung, A.M.; Mailer, R.J.; Williams, W.M. Potential impact of weedy Brassicaceae species on oil and meal quality of oilseed rape (canola) in Australia. Weed Res. 2018, 58, 200–209. [CrossRef]
- Seal, A.N.; Pratley, J.E.; Haig, T.; Lewin, L.G. Screening rice varieties for allelopathic potential against arrowhead (Sagittaria montevidensis), an aquatic weed infesting Australian Riverina rice crops. Aust. J. Agric. Res. 2004, 55, 673–680. [CrossRef]
- Smith, J.; Jones, A.; Doe, B. A gravimetric method for measuring stem-specific density in forestry research. J. Frost Sci. 2018, 15(3), 123–135.
- Smith, A.N.; Reberg-Horton, S.C.; Place, G.T.; Meijer, A.D.; Arellano, C.; Mueller, J.P. Rolled Rye Mulch for Weed Suppression in Organic No-Tillage Soybeans. Weed Sci. 2011, 59, 224–231. [CrossRef]
- Starling, A.P.; Umbach, D.M.; Kamel, F.; Long, S.; Sandler, D.P.; A Hoppin, J. Pesticide use and incident diabetes among wives of farmers in the Agricultural Health Study. Occup. Environ. Med. 2014, 71, 629–635. [CrossRef]
- Shukla, G.K. Some statistical aspects of partitioning genotype-environmental components of variability. Heredity 1972, 29, 237–245. [CrossRef]
- Vencill, W.K.; Nichols, R.L.; Webster, T.M.; Soteres, J.K.; Mallory-Smith, C.; Burgos, N.R.; Johnson, W.G.; McClelland, M.R. Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops. Weed Sci. 2012, 60, 2–30. [CrossRef]
- Worthington, M.; Reberg-Horton, C. Breeding Cereal Crops for Enhanced Weed Suppression: Optimizing Allelopathy and Competitive Ability. J. Chem. Ecol. 2013, 39, 213–231. [CrossRef]
- Wu, H.; Haig, T.; Pratley, J.; Lemerle, D.; An, M. Allelochemicals in Wheat (Triticum aestivum L.): Variation of Phenolic Acids in Shoot Tissues. J. Chem. Ecol. 2001, 27, 125–135. [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T. Crop cultivars with allelopathic capability. Weed Res. 1999, 39, 171–180.
- York, L.M.; Nord, E.A.; Lynch, J.P. Integration of root phenes for soil resource acquisition. Front. Plant Sci. 2013, 4, 355. [CrossRef]
- Zimdahl, R.L. Weed-Crop Competition: A Review; Blackwell Publishing: Ames, 2004.
Figure 1.
Negative linear relationship between canola plant density and weed biomass (m2) with correlation co-efficient values (R) and respective p-values. The data were collected from Marrar 2016 and Wagga Wagga 2016 experimental sites.
Figure 1.
Negative linear relationship between canola plant density and weed biomass (m2) with correlation co-efficient values (R) and respective p-values. The data were collected from Marrar 2016 and Wagga Wagga 2016 experimental sites.
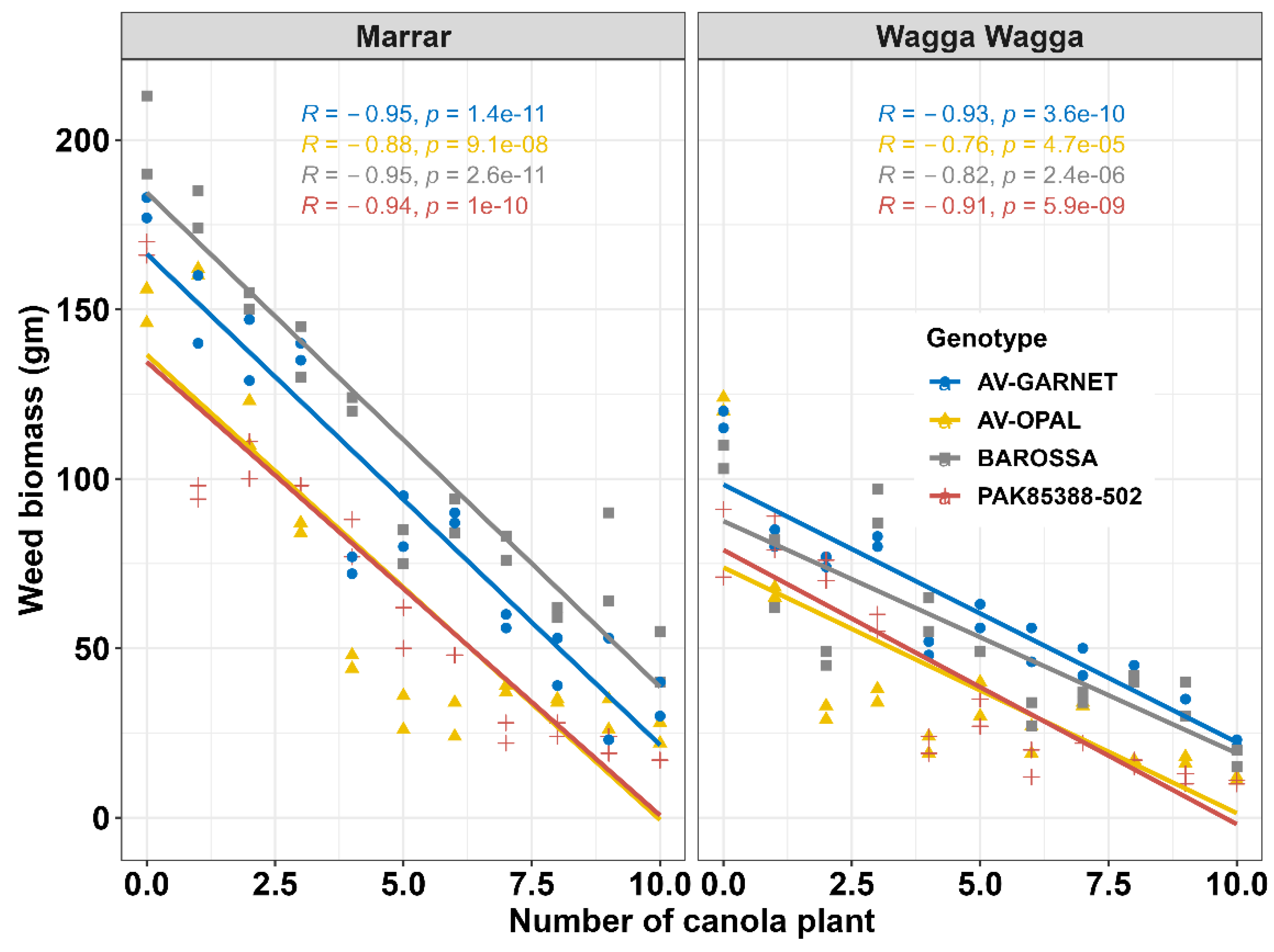
Figure 2.
Correlation matrix among canola traits with weed biomass. Here LR is number of lateral roots/plants, SD is stem density, SL is stem length, SSD is stem specific density, NDVI is normalised differential vegetation index, LAI is leaf area index and Weeds is total weed biomass.
Figure 2.
Correlation matrix among canola traits with weed biomass. Here LR is number of lateral roots/plants, SD is stem density, SL is stem length, SSD is stem specific density, NDVI is normalised differential vegetation index, LAI is leaf area index and Weeds is total weed biomass.
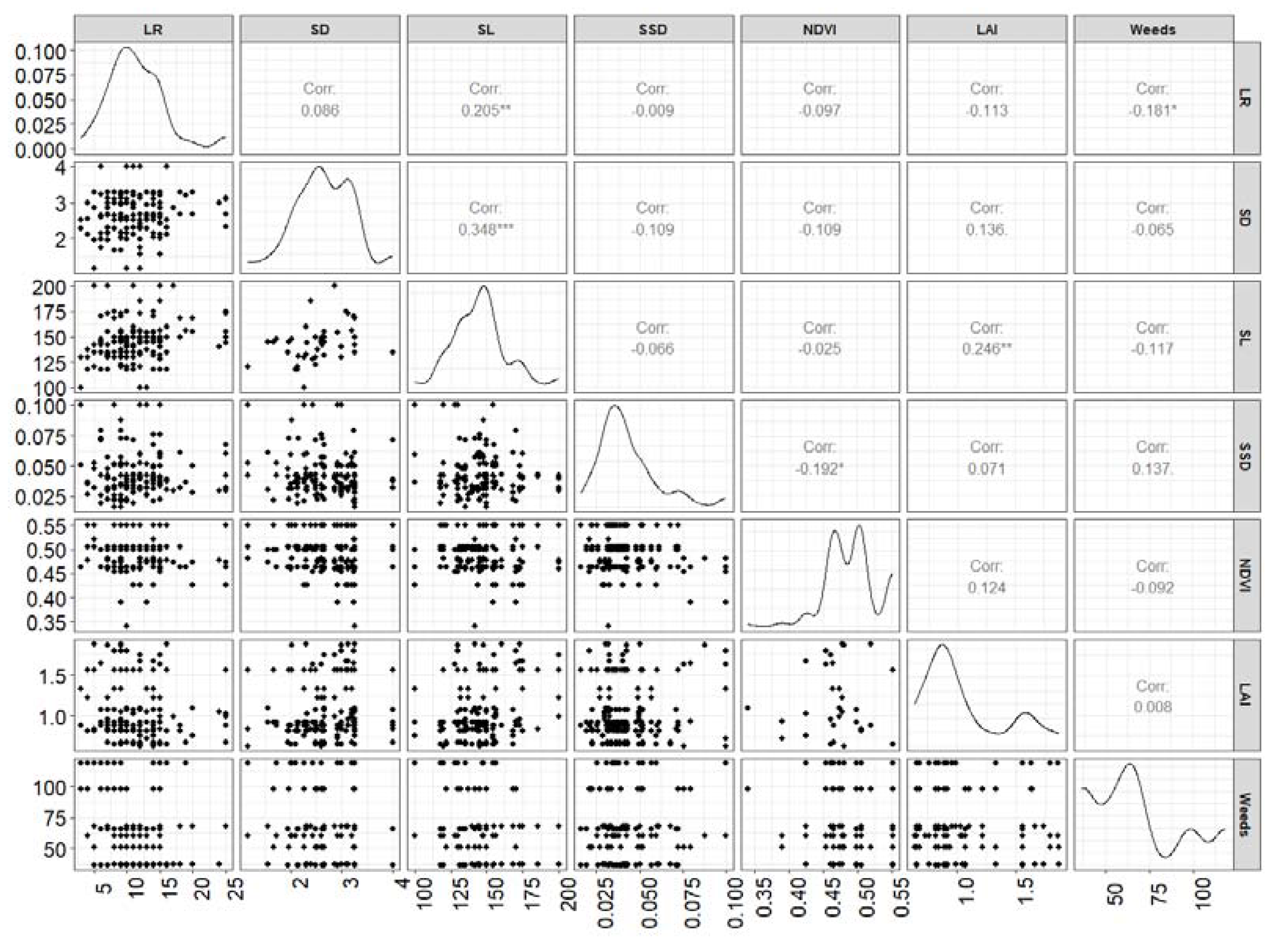
Figure 3.
Performance and adaption of canola genotypes for weed suppression. The black solid line showing the average weed suppression at different locations. .
Figure 3.
Performance and adaption of canola genotypes for weed suppression. The black solid line showing the average weed suppression at different locations. .
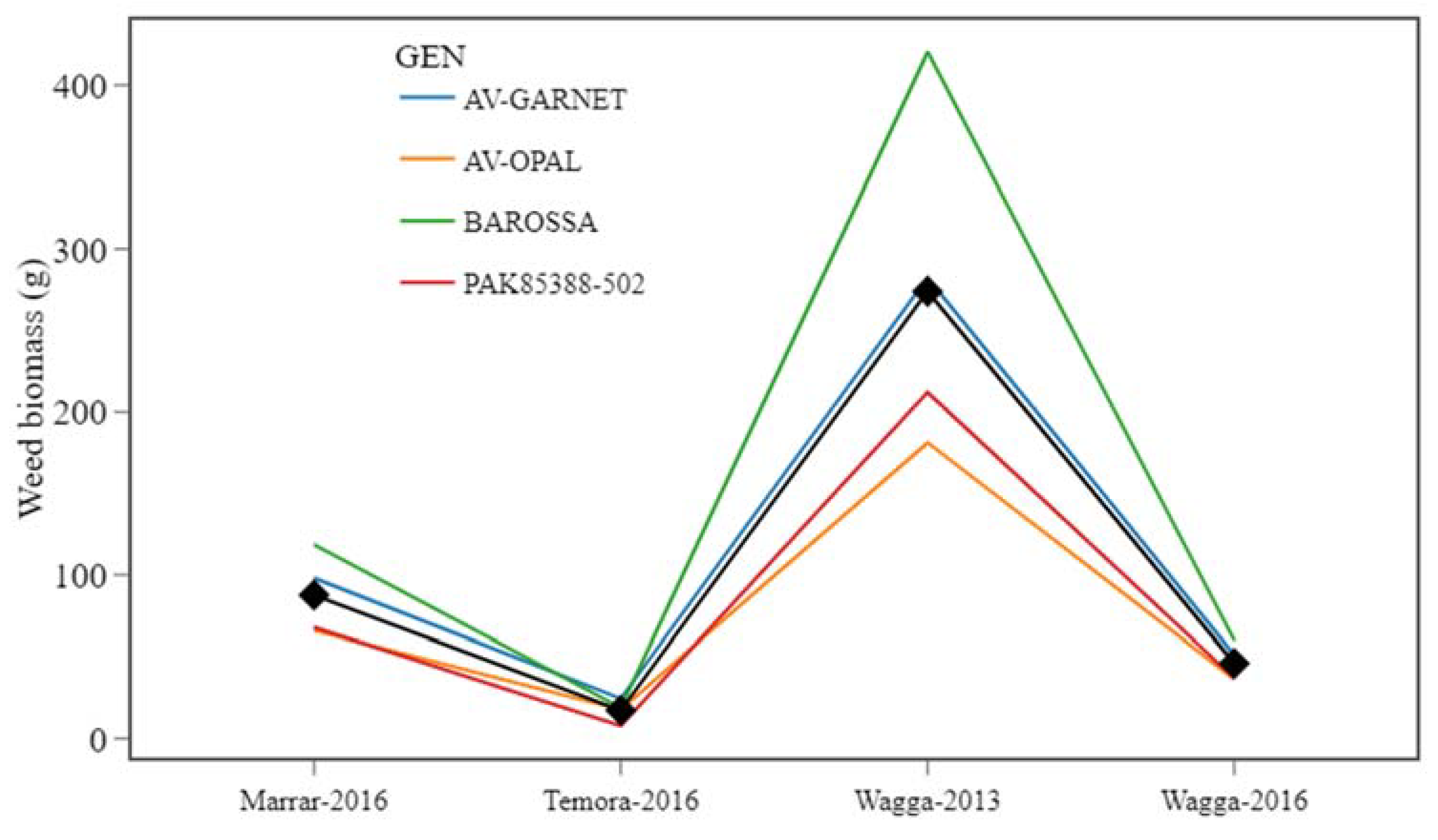
Figure 4.
Additive main effects and multiplicative interaction (AMMI) biplot for weed suppression (weed biomass) of four canola genotypes at different locations.
Figure 4.
Additive main effects and multiplicative interaction (AMMI) biplot for weed suppression (weed biomass) of four canola genotypes at different locations.
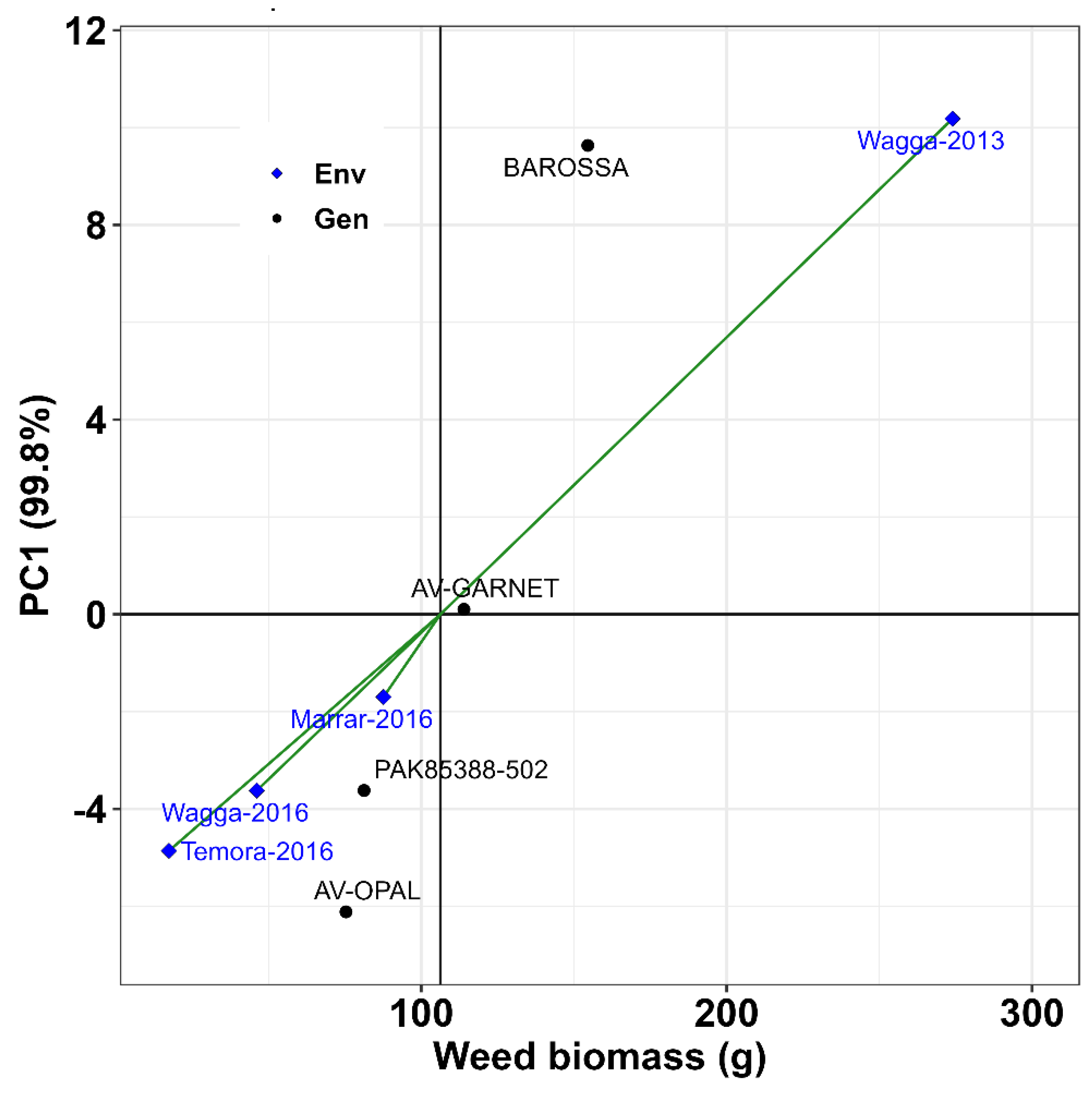

Table 1.
Data measurements for four different field experiments.
| Name | Units | Where measured | Site |
|---|---|---|---|
| Weed number | plants/m2 | 2 x quads (0.5 m x 0.5 m per plot | Wagga Wagga 2016, and 2013, Temora 2016 |
| Weed biomass (drymatter) | g/m2 | 2 x quads (0.5 m x 0.5 m per plot | Wagga Wagga 2016, and 2013, Temora 2016 |
| NDVI (greenness index) | unit less | Whole plot length | Wagga Wagga 2016, Marrar 2016 |
| Ceptometer (LAI and PAR) | unit less | Whole plot length | Wagga Wagga 2016, and Marrar 2016 |
| Canola root length, lateral roots and relative root growth | cm, no/plant | 20 plants/genotype | Wagga Wagga 2016, and Marrar 2016 |
| Canola shoot length, diameter and stem-specific density | cm, g cm–3 | 20 plants/genotype | Wagga Wagga 2016 and Marrar 2016 |
Table 2.
Mean number of lateral roots (LR), relative root growth (RRG), shoot length (SL), stem specific density (SSD), NDVI, LAI and PAR of four canola genotypes at Wagga Wagga (W) and Marrar sites (M) 2016.
Table 2.
Mean number of lateral roots (LR), relative root growth (RRG), shoot length (SL), stem specific density (SSD), NDVI, LAI and PAR of four canola genotypes at Wagga Wagga (W) and Marrar sites (M) 2016.
| Genotype | LR | RRG | SL (cm) | SSD | NDVI | LAI | PAR |
|---|---|---|---|---|---|---|---|
| AV-GARNET | 9.80 (±0.63) | 2.50 (±0.40) M 3.0 (±0.34) W |
142.60 (±3.47) | 0.04 (±0.002) | 0.46 M 0.49 W |
1.10 | 0.54 |
| AV-OPAL | 12.60 (±0.60) | 1.10 (±0.19) M 1.80 (±0.24) W |
137.60 (±4.00) | 0.04 (±0.002) | 0.49 M 0.46 W |
1.00 | 0.53 |
| BAROSSA | 10.55 (±0.63) | 3.18 (±0.49) M 2.00 (±0.21) W |
136.73 (±6.50) | 0.05 (±0.003) | 0.45 M 0.49 W |
1.20 | 0.50 |
| PAK85388-502 | 14.35 (±0.58) | 2.37 (±0.25) M 1.92 (±0.25) W |
164.00 (±4.80) | 0.03(±0.002) | 0.47 M 0.51 W |
1.10 | 0.48 |
Table 3.
Stability and adaptability of four canola genotypes for weed suppression at different environments and two different years.
Table 3.
Stability and adaptability of four canola genotypes for weed suppression at different environments and two different years.
| Genotype | Analysis for all environments | Analysis for unfavourable environments | Analysis for favourable environments | Shukla stability | Regression parameters | ||||||
| Weed biomass (g) | Rank | Weed biomass (g) | Rank | Weed biomass (g) | Rank | Rank | b0 | b1 | R2 | RMSE | |
| AV-GARNET | 114 | 3 | 282 | 3 | 57.8 | 3 | 3 | 114 | 1.01 | 1.00 | 1.87 |
| AV-OPAL | 75.3 | 1 | 181 | 1 | 40.1 | 2 | 2 | 75.3 | 0.63 | 0.99 | 1.45 |
| BAROSSA | 155.0 | 4 | 420 | 4 | 65.9 | 4 | 4 | 155 | 1.58 | 0.99 | 4.15 |
| PAK85388-502 | 81.3 | 2 | 212 | 2 | 37.7 | 1 | 1 | 81.3 | 0.78 | 0.99 | 2.42 |
A low b0 value suggests less variability observed in genotypes for suppression among environments condition, while a low RMSE value indicates regression model fitted very well. The data range for weed biomass is 769 - 2 = 767, and 1% of this range is 7.67, which is considered a threshold for a low RMSE value.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



