Submitted:
08 August 2024
Posted:
09 August 2024
You are already at the latest version
Abstract
Aflatoxin contamination (AC) increases as the severity of drought stress increase in peanut. Identifying drought-tolerant (DT) genotypes with resistance to Aspergillus. flavus in semi-arid tropics may aid in development of peanuts that minimizing aflatoxin contamination. The goal is to identify the DT genotypes and resistant to aflatoxin contamination. The experiments curried out at ICRISAT Sahelian Center on fifty-five genotypes assessed in adjacent intermittent water-stressed (WS) from 60th day after sowing to maturity and well-watered (WW) conditions in a randomized complete block design. The yields and components, incidence of A. flavus colonisation, aflatoxin contamination and the seed coat total pthe olyphenol (SCTPP) were estimated. Water deficit reduced pod yield, seed yield, and haulm yield up to 19.49%, 27.24% and 22.07% respectively and increased the immature pods plant-1, (IMPN) and aflatoxin contamination up to 67.16% and 54.95%. The genotypes ICG 2106, ICG 311, ICG 4684, ICG 4543, and ICG 1415 maintained high yield under WS, less of IMPN and the lowest AC variation between WW and WS. The drought tolerant genotypes showed the relationship with aflatoxin resistance and SCTPP (r2=0.80; r2=0.82) under WW and WS. These findings can be used to select the genotype that combine drought tolerance and minimizing aflatoxin contamination.
Keywords:
peanut
; intermittent drought
; seed coat
; aflatoxin
1. Introduction
Peanut (Arachis hypogaea L) is the second important cash crop in Niger which is produced in five of eight regions of the country. It is called ‘women crop’ because extensive involvement of women in peanut production and processing. Peanut pods grow underground, their development is directly influenced by the water conditions of the surrounding soil [1]. As an underground crop, the pods are subjected to continuous risk of direct contact with populations of aflatoxigenic aspergilli in the soil [2]. Aflatoxin contamination is the most important quality problem in peanuts throughout the world as it is related to serious health problems in human as well as in livestock [3].
Peanut grown in the Sahel often experiences water deficits during the pod-filling phase, which usually coincides with the end of the rainy season [4]. In this zone, peanut production is often affected by an intermittent drought which is an episodic water deficit during plant growth [5]. Previous studies in Niger demonstrated that drought stress for less than ten days was enough to cause significant aflatoxin contamination in the field [6,7]. However, increased duration of terminal drought and temperature are major factors determining the level of aflatoxin contamination [6].
Precisely, at the pod filling stage, drought cause plant stress and produce lower phytoalexin [8] then A. flavus can colonize in peanut pods prior to harvest and the contamination of aflatoxin is more severe under terminal drought [9,10]. Indeed, drought can cause cracking of seed coats, which in turn permits ingress of A. flavus germinating spores and hyphae into embryonic tissues [11]. Recent studies observed that varieties subjected to drought during late growth stages had different levels of resistance to aflatoxin contamination [9]. The different levels of resistance would be linked firstly to drought tolerance mechanisms, either by escape, tolerance or avoidance, may impact the ability of genotypes to minimize aflatoxin production by maintaining kernel water activities allowing phytoalexin production [12]. Secondly, the seed coats and outer shells play vital roles in protecting the seeds from mechanical damage, pest infestation and surviving harsh weather conditions [13]. Then, a combined approach of host resistance followed by pre- and post-harvest management practices is required [14].
Aflatoxin contamination can be minimized by adopting certain cultural, produce handling, and storage practices [12]. However, these practices are not widely adopted particularly by the small farmers in the developing countries, which contribute about 60% to the world peanut production. One of the possible means of reducing aflatoxin contamination of peanut is the use of cultivars resistant to seed invasion by aflatoxin producing fungi or to aflatoxin production. This suggests that drought tolerant genotypes may possess some degree of tolerance to aflatoxin contamination and it has been argued that drought tolerance traits in peanut may have the potential to be used as indirect selection criteria for resistance to pre-harvest aflatoxin contamination [15]. Recent study [16] reported a wide variation between genotypes in incidence, severity, and aflatoxin rate can be biochemical compounds’ diff--erential variability in the tested seeds. It was observed the differences in mycelial growth surface coverage were probably attributed to differences in physical and chemical features of the seed-coat, pod-shell thickness and reticulation [17]. The seed coats and outer shells play vital roles in protecting the seeds from mechanical damage, pest infestation and surviving harsh weather conditions [13]. We have recently demonstrated that seed coat acts as a physical and biochemical barrier against A. flavus infection [18,19]. To develop peanut cultivars resistant to A. flavus infection with reduced aflatoxin contamination, there is a possible defence mechanism at three stages: prevention of fungal invasion in the pericarp, reduction of aflatoxin production the seed coat and cotyledons [20]. Using cultivars resistant to kernel infection by A. flavus is also one of the promising ways to reduce aflatoxin contamination. The objective of this study was to identify the drought tolerant and seed coat biochemical traits to A. flavus resistance and aflatoxin contamination. The specific objectives are to (1) evaluate peanut genotypes under intermittent water deficit to identify the drought tolerants(2) identify specific biochemical traits associated to A. flavus resistance in seed coat and (3) identify the intermittent drought tolerant genotypes with seed coat resistance to A. flavus infection and Aflatoxin contamination.
Materials and Methods
2.1. Plant Materials, Temperature and Relative Humidity
Fifty five lines from ICRISAT peanut mini core collection, contrasting for aflatoxin content [21] were selected and seeds were provided by ICRISAT Niger gene bank. The cultivars included as check, 55-437 and J11 are considered as resistant to pre-harvest aflatoxin contamination while JL24 and Fleur11 are susceptible [22]. Two experiments were conducted (August to December) in year1 and year2 under field conditions at the International Crop Research Institute for the Semi-Arid Tropics, (ICRISAT) Sahelian Centre (ISC) in Sadoré (45km south of Niamey, Niger, 13°N, 2°E). During the crop growing period, the maximum (Max) and minimum (Min) air temperatures and the relative humidity (RH) were recorded daily from a meteorological station located close to the experimental field. The Max and Min temperatures varied from 26.4°C to 41.2°C in year1 and from 29.01°C to 39.30°C in year1 while the RH varied from 29.9% to 68.6% in year2 and from 40.0% to 62.23% in year2. The total water received from rainfall and irrigation were 590 mm3 and 470 mm3 respectively in were well-watered (WW) and intermittent water-stressed (WS) in year1 and 645 mm3 and 505 mm3 in year2.
2.2. Experimental Design and Water Treatment
The experimental design was alpha lattice with two factors, water treatment as main factor, and genotypes (55) as sub-factor randomized in each water treatment with four replications. The spacing between water treatments was 6 m and 2 m between replications. Within each repetition, there were 55 elementary plots with an area of 2 m x 1 m = 2 m2 spaced 1 m apart. The plots subject to WS are suitably watered (30 mm3 per irrigation) like those of the WW treatment until the 60th day after sowing (seed filling stage start) when the water stress was imposed. The WS treatment consisted of stopping irrigation of WS plants until the majority of stressed plants showed clear wilting symptoms before watering and then stopping irrigation again. This cycle continues until the pod’s maturity.
2.3. Crop Management
In each experiment, plot area measured 2 m2 (2m x 1m) including two rows of 2 m length. Rows spacing was 0.5 m and each row had ten hills. Three seeds were sown by hand in each hill, in 3 cm deep after receiving an irrigation of 30 mm using a Linear-move irrigation system (Valmont Irrigation Inc., Valley, Nebraska, USA). Two and three weeks after sowing, the plants were thinned to two and one plants per hill respectively. The plots were subsequently fertilized with 150 kg ha-1 N-P2O5-K2O and irrigated with 30 mm of water. The fields were kept free from weeds by manual cultivation, and regular spraying of decis (deltaméthrin 12 CE), Emacot 050WG (Emamectine benzoate 50g/kg) and Benji controlled insect pests.
2.4. Measurements and Data Collection
From the date of sowing days to flowering was recorded when 50% of plants in each plot reached flowers, day to beginning seeds filling stage and 50% seeds filling stage was monitoring to applied water stress to harvest were recorded when 50% of plants in each plot showed mature pods.
Yield components
After harvesting by hand, the plants of each plot (1m²), the pods number, the immature pods number, the seeds number, the haulm yield, the pods yield, the seeds yield were determined.
2.5. Assessment of Phenotypic Variation to A. Flavus Colonization
The In-Vitro Seed Colonisation (IVSC) was carried out in the laboratory using ten, fifteen, and twenty-five seeds per petri dish. For each genotype, the seeds with seed coat showing no apparent damage were selected. Each petri dish was considered as a replications using a modified method [23]. The A. flavus strains was collected from the sorghum seeds exposed at the Laboratory conditions (25°C-28°C) during seven days and multiplication on Potato Dextrose Agar (PDA) for inoculum preparation [23]. The A. flavus colonies were collected based on morphological characteristics and homogenized by successive culture on PDA medium. The A. flavus conidial suspension was prepared with the spores collected by culture escaping before dilution spores concentration with distilled water and adjusted approximately to 1.9.1×6 spores/millilitre (ml) using Haemocytometer [24]. 500 µl of spore suspension was inoculated in each petri dish before incubation at 28°C for seven days in darkroom. The observation was recorded for percent of colonization after the incubation. Individual seeds were scored for surface colonisation by A. flavus and for colonization severity using the following rating scale. 1=˂5% seeds surface colonized with scanty mycelial growth and no sporulation; 2= 5-25% seeds surface colonized with good mycelial growth and scanty sporulation; 3= 26-50% seeds surface colonized with good mycelial growth and good sporulation ; 4= ˃ 50% seeds surface colonized with heavy sporulation [23]. A. flavus colonization Incidence determination
2.6. Seed Coat Total Polyphenol Extraction and Quantification
Extraction: undamaged and matured seeds of all the studied genotypes were collected after drying. The seeds coats were removed from embryo before grinding to reduce in powder. After grinding, 0,1g of seed coat powder of each genotypes was introduced in the tube and added 10 ml of methanol (50%) before warming at 77°C for 1 hour. Spectrometric Assay: The total polyphenol was determined using FCR (Folin Ciocalteu reagent) [25].The absorbance was estimated at 750 nm using a Spectrophotometer (6715 UV/Vis JENWAY). The calibration curve was plotted using 0,1,2,3,4,5,6 ml of solutions of 0,05mg/ml of tannic acid concentration. The absorbance was measured to determine the content of total polyphenols using the following formula: C= (C1×V)/W with C being the content of total polyphenols expressed in mg equivalent tannic acid/g of dry matter, C1 the concentration of tannic acid established from the calibration curve in mg/L, V the volume of extract in L and W the weight of the plant extracting.
2.7. Genotypes Aflatoxin Content Quantification
Aflatoxin concentration in the seeds of fifty-five genotypes was estimated by an Enzyme Linked Immunosorbent Assay (ELIZA). In each trial (year1 and year2), 100 g of seeds were collected in the two water regimes plot. The 20g of this fine powder was used for the extraction of aflatoxin by dissolving in 70% (v/v) methanol containing 0.5% (w/v) KCl and homogenized and filtered through Whatman No1 filter paper. The filtrate was diluted 1:15 with methanol and used in duplicate to estimate aflatoxin concentration by indirect competitive ELIZA essentially as described by [26].
2.8. Analysis
The data used were the means of the two years experiments. GENSTAT 14th edition (VSN international Ltd. Hemel Hempstead, UK) was used to perform statical analysis. Shapiro-Wilk normality test was done before analysis of variance (ANOVA) which realised to assess the effects of genotype (G), water regime (W), and their interactions for the different traits measured. Microsoft Office Excel 2013 Software (Microsoft Corp., Redmond, WA, USA) was used to establish a relationship between the different yields, Aflatoxin content and the seed coat total polyphenol content. In addition it used to perfom the tables and figures.
Results
3.1. Yields and Yield Components
ANOVA revealed a significant genotype by water treatment interaction G× (Wtrt) for all yields traits (Table1) indicating that these parameters varied depending on the water regime. Water deficit imposed was more severe on seed number per plant and seed yield than on pod number per plant, pod yield and haulm yield (table2). Under well-watered conditions, the highest Pods Number per plant (PNP), and Seeds Number per plant (SNP) were observed on ICG 10950, ICG 11322, ICG 4598, ICG 1415, ICG 1519, ICG 332, ICG 5609, ICG 6407, ICG 3992 and ICG 12235. For pod yield (PY), seed yield (SY) and haulm yield (HY) under WW, the genotypes ICG 12697 (560.6 gm-2 , 376.8 gm-2 and 1022.4 g.m-2 respectively), ICG 12879 (472.2 gm-2, 306.9 gm-2 and 584.7 gm-2 respectively), ICG 2019 (510.7 gm-2, 289.6 gm-2 and 615.1 gm-2 respectively), ICG 4729 (393.3 gm-2, 229 gm-2 and 711.7 gm-2respectively), ICG 4543 (485.5 gm-2, 227.7 gm-2 and 785.4 gm-2 respectively), ICG 4750 (580.1 gm-2, 338 gm-2 and 764.5 gm-2 respectively), ICG 5195 (547.1 gm-2 , 302.3 gm-2 and 850.6 gm-2 respectively), ICG 5609 (524.9 gm-2, 278.9 gm-2 , and 750.7 gm-2 respectively), ICG 6407 (522.4 gm-2 , 290.7 gm-2 and 833.9 gm-2 respectively) and ICG 6703 (546.3 g.m-2, 306 g.m-2 and 932.7 g.m-2 respectively) were the highest performing (Table3).
However, ICG 10950, ICG 12235, ICG 1415, ICG 4598, ICG 5609, and ICG 6407 showed high performance under WW for pods number per plant (PNP), seeds number per plant (SNP) and low immature pods number per plant (IMPN). The latest genotypes revealed also the best in pods and seeds production with less immature pods under WS (Tablee3). In addition, the genotypes ICG 12879, ICG 14523, ICG 2019 and ICG 5195 showed high pods and seeds numbers per plant under WS. Our results showed that among the ten best performing genotypes under WS for pod yield, seed yield and haulm yield, five genotypes were also performing under Well-watered regime. These genotypes were ICG 12697 (363.4 gm-2, 245.3 gm-2, 513.6 gm-2), ICG 12879 (351.7 gm-2g.m-1, 211.7 gm-2, 536.9 gm-2), ICG 2019 (384.1 gm-2 , 197.1 gm-2 , 474.3 gm-2 ), ICG 4729 (329.7 gm-2 , 155.4 gm-2 , 530.2 gm-2 ), and ICG 6703 (377.6 gm-2 , 213 gm-2 g.m-1, 517.2 gm-2 ). In addition, these five drought tolerant genotypes were followed by ICG 4598 (497.8 gm-2 , 197 gm-2 , 803 gm-2), ICG 4684 (445.7 gm-2 , 246.8 gm-2 , 652.1 gm-2 ), ICG 2106 (463.9 gm-2 , 239.7 gm-2 , 586.8 gm-2 ), ICG 12988 (388 gm-2 , 206.5 gm-2 , 533.6 gm-2 ), and ICGIL 11114 (442.2 gm-2, 264.8 gm-2, 627.4 gm-2). However water deficit affected severely all yield components (PNP, SNP). Therefore, the pod and seed yields of the following genotypes are more severely affected by the intermittent water stress during seed filling phase by showing the highest seed yield loss (Table3). They are ICG 12235 (677 gm-2 , 183.4 gm-2 , 756.6 gm-2 ), ICG 1142 (1247.9 gm-2 , 328 gm-2 , 666.6 gm-2 ), ICG 14523 (746.5 gm-2 , 298.8 gm-2 , 780.2 gm-2 ), ICG 14106 (776.9 gm-2 , 252.1 g.m-2 , 513.4 gm-2 ), ICG 14630 (654.8 gm-2 , 150.6 gm-2 , 636.6 gm-2 ), ICGIL 11125 (674.3 gm-2 , 207.8 g.m-2 , 594.2 gm-2 ), and ICGIL17108 (582.3 gm-2 , 104.4 g.m-2 ,808 gm-2).

Table 1.
Results of variance analysis (F value), F (probability), Means, LSD (least significant differences of means 5% level) of yield and its components. Genotype (G), Water Treatment (Wtrt) and genotype-by-water treatment interaction (G×Wtrt) effects were tested. Wtrt= water treatment; PNP=Pods Number per plant-1; SNP= Seeds Number per plant-1; IMPN= Immature Pods Number per plant-1; HY=haulm yield; PY=pods yield; SY=Seeds Yield.
Table 1.
Results of variance analysis (F value), F (probability), Means, LSD (least significant differences of means 5% level) of yield and its components. Genotype (G), Water Treatment (Wtrt) and genotype-by-water treatment interaction (G×Wtrt) effects were tested. Wtrt= water treatment; PNP=Pods Number per plant-1; SNP= Seeds Number per plant-1; IMPN= Immature Pods Number per plant-1; HY=haulm yield; PY=pods yield; SY=Seeds Yield.
| Sources | PNP | IMPN | SNP | HY (gm-2) | PY (gm-2) | SY (gm-2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| of variance | WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | WW | WS |
| F value | 31.62 | 31.75 | 12 | 3.25 | 14.23 | 12 | 8.25 | 4.24 | 14 | 12 | 7.24 | 9.16 |
| F (prob) | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 |
| WTrt | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 | <.001 |
| G×(Wtrt) | <.001 | <.001 | <.001 | 0.015 | 0.027 | 0.020 | ||||||
Significance at 0.05, 0.01 and 0.001 level.
Table 2.
Means variation of Yields and it’s components under well-watered and water stressed treatments and the percentages of decreasing and increasing.
Table 2.
Means variation of Yields and it’s components under well-watered and water stressed treatments and the percentages of decreasing and increasing.
| Traits | Water Traitements | WS negative effect (%) | |
|---|---|---|---|
| WW | WS | ||
| Pods number per plant | 102±31 | 80±25 | 21.92 |
| Immature pods per plant (increasing) | 9±5 | 21±10 | 57.14 |
| Seeds number per plant | 123±31 | 89±27 | 27.36 |
| Pod Yield (gm-2) | 678.54±203.51 | 546.15±163.5 | 19.43 |
| Seed Yield (gm-2) | 316.67±90.26 | 230.37±69.3 | 27.24 |
| Haulm Yield (gm-2) | 828.53±85.30 | 645.57±150.13 | 22.07 |
Table 3.
Means variation of Yields and it’s components under well-watered and water stressed treatments and Least significant differences 5% level (LSD). PNP: Pods number per plant; SNP: Seeds Number per plant; IMPN: Immature Pods Number per plant; HY: Haulm Yield; PY: Pod Yield, and SY: Seed Yield.
Table 3.
Means variation of Yields and it’s components under well-watered and water stressed treatments and Least significant differences 5% level (LSD). PNP: Pods number per plant; SNP: Seeds Number per plant; IMPN: Immature Pods Number per plant; HY: Haulm Yield; PY: Pod Yield, and SY: Seed Yield.
| Well-Watered | Water Stressed | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | PNP | SNP | IMPN | PY (gm-2) | SY (gm-2) | HY (gm-2) | PNP | SNP | IMPN | PY (gm-2) | SY (gm-2) | HY (gm-2) |
| 55-437 | 103±2 | 124±17 | 8±2 | 594.7±96 | 290.6±49 | 736.4±292 | 95±11 | 109±14 | 15±1 | 517±51 | 248.2±46 | 612.8±220 |
| Fleur 11 | 99±12 | 136±13 | 9±1 | 613.5±81 | 338.6±79 | 870.1±75 | 89±5 | 119±16 | 17±2 | 572±138 | 316.2±33 | 658±160 |
| ICG 10950 | 98±5 | 112±19 | 4±1 | 701.6±61 | 376.8±70 | 1022.4±154 | 79±8 | 70±14 | 16±2 | 638.8±30 | 284.3±10 | 740.3±97 |
| ICG 11322 | 76±22 | 72±12 | 4±1 | 567.8±33 | 272.8±39 | 691.6±118 | 62±14 | 65±9 | 26±2 | 529±49 | 242.3±31 | 583.9±127 |
| ICG 1142 | 112±6 | 112±8 | 12±2 | 1664.9±255 | 369.4±78 | 798.6±69 | 84±10 | 78±4 | 23±2 | 1247.9±283 | 328±82 | 666.6±151 |
| ICG 12235 | 207±16 | 116±18 | 34±3 | 766.6±154 | 188.5±32 | 930.1±82 | 141±6 | 56±13 | 76±3 | 677±128 | 183.4±17 | 756.6±148 |
| ICG 12697 | 112±7 | 137±5 | 9±2 | 560.6±76 | 395.4±70 | 637.2±245 | 75±6 | 100±9 | 17±3 | 363.4±50 | 245.3±19 | 513.6±85 |
| ICG 12879 | 101±2 | 129±11 | 8±2 | 472.2±68 | 306.9±27 | 584.7±184 | 86±6 | 64±20 | 16±2 | 351.7±33 | 211.7±48 | 536.9±58 |
| ICG 12988 | 101±17 | 131±21 | 10±2 | 649.7±148 | 280.9±92 | 562.2±80 | 76±13 | 102±11 | 17±2 | 388±38 | 206.5±35 | 533.6±86 |
| ICG 12991 | 115±8 | 153±21 | 4±1 | 525.3±42 | 243.8±36 | 657.5±203 | 79±10 | 86±4 | 17±3 | 443.9±33 | 199.5±25 | 629.5±169 |
| ICG 13099 | 111±3 | 133±9 | 8±1 | 889.1±235 | 370.8±120 | 925.1±86 | 88±10 | 85±20 | 21±2 | 653.8±98 | 286.3±51 | 741.1±133 |
| ICG 13603 | 177±12 | 205±26 | 10±1 | 780.9±83 | 356.2±111 | 775.4±58 | 134±9 | 163±20 | 25±1 | 662.5±131 | 256.6±17 | 589.9±102 |
| ICG 13858 | 76±8 | 112±14 | 8±3 | 475.7±80 | 196.0±57 | 910.5±48 | 48±4 | 67±14 | 21±4 | 397.5±20 | 134.8±32 | 552.6±159 |
| ICG 14106 | 92±8 | 142±17 | 8±2 | 833.5±47 | 321.4±35 | 724±162 | 53±8 | 74±13 | 19±2 | 776.9±145 | 252.1±50 | 513.4±50 |
| ICG 1415 | 100±5 | 98±11 | 13±2 | 655.3±78 | 282.7±28 | 767.3±76 | 86±10 | 71±12 | 22±5 | 624.3±67 | 209.6±34 | 589.7±135 |
| ICG 14523 | 159±13 | 174±12 | 10±2 | 906.0±154 | 378.6±31 | 954.3±58 | 121±12 | 111±3 | 21±3 | 746.5±27 | 298.8±25 | 780.2±125 |
| ICG 14630 | 53±7 | 84±10 | 9±2 | 725.4±107 | 206.9±61 | 893.5±224 | 50±4 | 67±8 | 15±3 | 654.8±68 | 150.6±12 | 636.2±262 |
| ICG 1519 | 86±5 | 79±15 | 8±2 | 590.5±96 | 214±55 | 761.5±185 | 63±11 | 70±16 | 20±3 | 485.6±16 | 180.9±36 | 695.4±208 |
| ICG 156 | 102±14 | 125±11 | 9±1 | 682±84 | 237.9±24 | 1602±468 | 84±2 | 83±9 | 21±3 | 432.6±58 | 193.2±38 | 1074.3±406 |
| ICG 163 | 104±7 | 113±10 | 10±3 | 733.9±204 | 433±83 | 1056.4±194 | 91±2 | 83±9 | 22±3 | 561.6±73 | 338±93 | 769.4±233 |
| ICG 2019 | 69±7 | 105±16 | 11±3 | 510.7±145 | 289.6±82 | 615.1±108 | 37±25 | 49±24 | 20±4 | 384.1±85 | 197.1±73 | 474.3±274 |
| ICG 2106 | 108±4 | 136±11 | 10±3 | 612.9±132 | 329.7±20 | 853.5±83 | 75±10 | 88±18 | 18±3 | 463.9±40 | 239.7±31 | 586.8±318 |
| ICG 3027 | 98±11 | 130±9 | 11±3 | 752.9±50 | 381.7±41 | 892.4±104 | 62±2 | 82±19 | 15±2 | 613.9±81 | 304.3±49 | 718.2±151 |
| ICG 311 | 106±9 | 135±10 | 7±2 | 638.5±106 | 393.6±71 | 676.2±124 | 86±8 | 101±11 | 14±1 | 562.8±102 | 269±20 | 543.1±54 |
| ICG 332 | 63±1 | 83±8 | 5±2 | 658.8±26 | 283.4±34 | 985±60 | 39±5 | 47±11 | 18±2 | 457.5±44 | 162.1±37 | 726.5±144 |
| ICG 334 | 105±9 | 159±20 | 8±1 | 884.7±22 | 444.3±29 | 813.5±172 | 98±7 | 143±15 | 24±2 | 692.3±89 | 286.5±32 | 553.5±138 |
| ICG 36 | 100±13 | 112±15 | 7±2 | 778.8±64 | 222.2±35 | 737.3±111 | 53±7 | 67±11 | 27±3 | 528.2±83 | 148.6±13 | 463.2±110 |
| ICG 3992 | 73±11 | 77±20 | 10±2 | 654.1±130 | 317.7±86 | 958.8±107 | 47±4 | 55±9 | 23±2 | 590.9±73 | 224.1±8 | 831.2±86 |
| ICG 4543 | 89±11 | 126±14 | 4±5 | 485.4±56 | 227.7±19 | 785.4±122 | 73±6 | 87±18 | 18±1 | 385.3±67 | 183.4±52 | 578±141 |
| ICG 4598 | 98±10 | 101±16 | 7±1 | 701.8±143 | 356.4±86 | 1050.8±62 | 76±7 | 89±18 | 19±1 | 497.8±41 | 197±9 | 803±66 |
| ICG 4684 | 87±8 | 121±25 | 10±1 | 633.8±95 | 347.9±41 | 765.3±97 | 71±5 | 98±18 | 19±2 | 445.7±53 | 246.8±49 | 652.1±87 |
| ICG 4729 | 80±2 | 94±6 | 10±2 | 393.3±64 | 229±17 | 711.7±115 | 66±7 | 80±13 | 23±1 | 329.7±47 | 155.4±28 | 530.2±91 |
| ICG 4750 | 89±4 | 116±5 | 9±1 | 580.1±27 | 338±28 | 764.5±86 | 75±9 | 85±10 | 27±4 | 439.8±46 | 186.7±32 | 635±109 |
| ICG 4764 | 84±7 | 125±7 | 8±2 | 727.3±157 | 369.3±68 | 839.8±200 | 79±3 | 108±7 | 25±3 | 549.6±104 | 231.5±24 | 643.7±165 |
| ICG 513 | 100±14 | 138±24 | 12±4 | 861.2±101 | 471.9±98 | 815.3±98 | 85±6 | 110±22 | 29±2 | 688.8±65 | 306±12 | 696.7±150 |
| ICG 5195 | 134±11 | 174±12 | 11±2 | 547.1±21 | 302.3±29 | 850.6±27 | 98±8 | 100±6 | 25±2 | 536±51 | 264.4±82 | 708.2±185 |
| ICG 532 | 90±3 | 114±7 | 10±1 | 729.9±123 | 298.2±55 | 824.7±145 | 71±8 | 104±2 | 18±2 | 532.4±96 | 213.1±60 | 677.5±110 |
| ICG 5609 | 60±6 | 98±21 | 6±2 | 524.9±81 | 278.9±14 | 750.7±171 | 49±10 | 58±9 | 15±3 | 411.5±34 | 146.8±17 | 556.8±147 |
| ICG 5663 | 144±9 | 135±6 | 17±2 | 667.1±78 | 313.9±64 | 735.7±2116 | 91±13 | 85±8 | 26±3 | 541.8±59 | 212.8±51 | 569.4±64 |
| ICG 6263 | 76±11 | 89±14 | 8±2 | 550.2±41 | 152.1±43 | 646.5±131 | 53±7 | 64±15 | 21±3 | 486.8±44 | 86.1±11 | 402±117 |
| ICG 6407 | 62±4 | 79±7 | 7±3 | 522.4±52 | 290.7±17 | 833.9±96 | 52±3 | 67±20 | 19±2 | 500.9±45 | 236.1±34 | 741±110 |
| ICG 6654 | 89±13 | 138±8 | 9±2 | 687.6±94 | 342.7±71 | 697.3±121 | 70±6 | 99±6 | 18±2 | 630.1±77 | 265.3±33 | 596±125 |
| ICG 6703 | 92±7 | 116±16 | 7±2 | 546.3±50 | 306±44 | 932.7±104 | 79±5 | 100±16 | 12±2 | 377.6±40 | 213±23 | 517.2±111 |
| ICG 6813 | 138±9 | 172±14 | 11±2 | 732.6±68 | 377.9±18 | 744.1±429 | 125±14 | 116±3 | 25±2 | 634.7±56 | 343.3±9 | 751.8±98 |
| ICG 6888 | 66±9 | 80±2 | 6±2 | 540.8±64 | 259.9±81 | 830.6±193 | 53±5 | 67±5 | 36±2 | 417.5±26 | 154.4±22 | 657±148 |
| ICG 7181 | 69±5 | 91±15 | 10±2 | 567.4±51 | 310.9±51 | 908.5±89 | 62±10 | 65±5 | 22±3 | 470.5±77 | 220.3±40 | 757±105 |
| ICG 721 | 106±8 | 132±21 | 11±2 | 804.5±75 | 409.4±33 | 1064.5±83 | 96±10 | 123±19 | 15±2 | 638.1±44 | 321.2±46 | 954.4±86 |
| ICG 76 | 149±22 | 162±10 | 9±2 | 933.6±51 | 467.7±69 | 807.4±106 | 132±9 | 131±18 | 19±3 | 727.2±72 | 342.7±41 | 618±155 |
| ICGIL 11102 | 141±31 | 159±29 | 8±2 | 603.7±62 | 282.9±62 | 974.6±139 | 119±8 | 127±21 | 23±2 | 568.6±56 | 248.3±62 | 761.1±177 |
| ICGIL 11110 | 132±13 | 151±8 | 12±2 | 833±102 | 418.1±55 | 848.4±120 | 89±9 | 103±18 | 25±2 | 578.8±132 | 270.8±29 | 567.8±224 |
| ICGIL 11114 | 70±9 | 109±22 | 5±1 | 712±110 | 423.5±85 | 825.2±158 | 65±5 | 91±19 | 21±3 | 442.2±135 | 264.8±32 | 627.4±182 |
| ICGIL 11125 | 121±4 | 132±12 | 11±1 | 724.4±119 | 398.2±56 | 728.5±116 | 105±10 | 108±5 | 19±2 | 674.3±194 | 207.8±61 | 594.2±184 |
| ICGIL 17108 | 135±7 | 131±5 | 15±1 | 765.7±98 | 180.7±35 | 1022.2±111 | 115±6 | 89±9 | 21±2 | 582.3±52 | 104.3±23 | 808±181 |
| J 11 | 116±8 | 127±9 | 8±1 | 497.4±39 | 259.1±39 | 697.3±61 | 79±8 | 87±8 | 17±2 | 443.7±70 | 205.3±13 | 493.5±183 |
| JL24 | 105±9 | 140±19 | 13±2 | 565.8±69 | 280.1±37 | 720.6±33 | 88±3 | 126±26 | 28±2 | 488.5±30 | 245±52 | 539.2±214 |
| Means | 102±31 | 123±31 | 9±4 | 678.5±204 | 316.7±90 | 828.5±279 | 80±25 | 89±27 | 22±8 | 546.2±164 | 230.4±69 | 645.6±215 |
| LSD | 15 | 21 | 5 | 131 | 79 | 145 | 12 | 20 | 9 | 107 | 56 | 140 |
3.2. Intermittent Water Deficit (WS) Effect on Pre-Harvest Aflatoxin Contamination (AC)
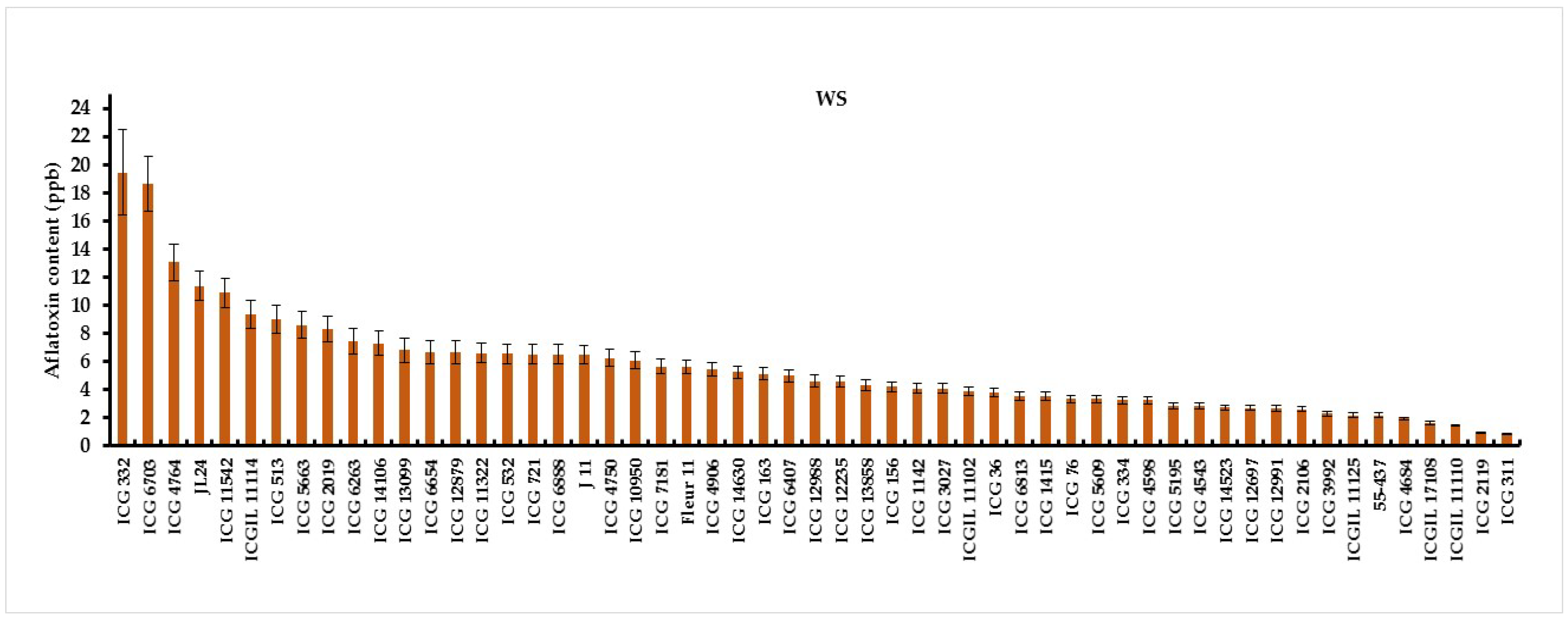
Analysis of variance revealed a significant genotype by water treatment interaction (G×Wtrt) for aflatoxin contamiation under well-watered and water stressed treatments (Table4). Under WW conditions, the lowest aflatoxin contamination were observed on five genotypes, ICG 12988 (0.28 ppb), ICGIL 17108 (0.30 ppb), ICG 5195 (0.40 ppb), ICGIL 11110 (0.46 ppb) and ICG 311(0.48). While ICG 332 (6.80 ppb), ICG 11542 (5.56 ppb), Fleur11 (4.01 ppb) and ICGIL11114 (3.75 ppb) showed the highest aflatoxin contamination under WW treatment (Figure1). Intermittent water stress (WS) imposed during seed filling phase increased aflatoxin contamination up to 67.16% (Figure2). Under water deficit the highest aflatoxin content were observed on ICG 332 (19.44 ppb), ICG 6703 (18.65 ppb), ICG 4764 (13.03 ppb), JL24 (11.37 ppb) and ICG 11542 (10.88 ppb) (Figure2). These genotypes were revealed more sensitive to aflatoxin contamination under drought. While, ICG 311 (0.83 ppb, ICG 2119 (0.90 ppb), ICGIL 17108 (1.6 ppb), ICG 4684 (1.93 ppb) and 55-437 (2.15 ppb) showed the lowest aflatoxin content under drought conditions. These genotypes were resistant to aflatoxin contamination (AC) under WS (Figure2).
Table 4.
Results of variance analysis (F value), F (probability), Means, SLD (least significant differences of means 5% level) of aflatoxin contamination under Well-watered and Stressed treatments.
Table 4.
Results of variance analysis (F value), F (probability), Means, SLD (least significant differences of means 5% level) of aflatoxin contamination under Well-watered and Stressed treatments.
| Aflatoxin B1 | ||
|---|---|---|
| Genotypes | WW | WS |
| Means | 1.81 | 5.51 |
| LSD | 1.14 | 1.49 |
| F value | 9.55 | 56 |
| F (Prob) | <.001 | <.001 |
| G×W(Trt) | <.001 | |
Significance at 0.05, 0.01 and 0.001 level.
Figure 1.
Aflatoxin content (ppb) variation of fifty-five peanut genotypes under well-watered (WW) treatment.
Figure 1.
Aflatoxin content (ppb) variation of fifty-five peanut genotypes under well-watered (WW) treatment.
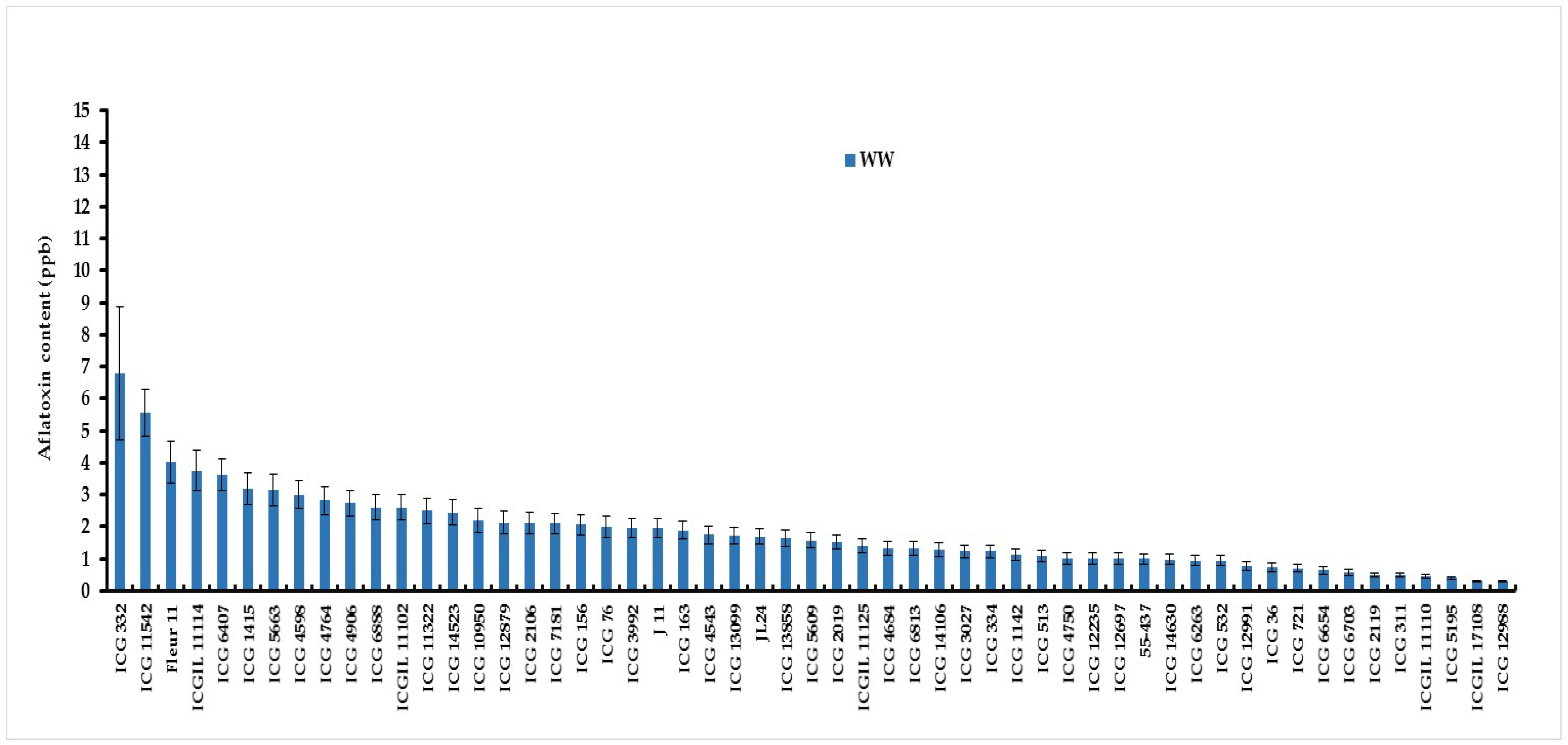
Figure 2.
Aflatoxin content (ppb) variation of fifty-five peanut genotypes under water stressed (WS) treatment.
Figure 2.
Aflatoxin content (ppb) variation of fifty-five peanut genotypes under water stressed (WS) treatment.
3.3. Seeds Colonisation and Total Polyphenol Content of Seed Coat
ANOVA revealed a significant (P˂0.001) genotypic difference for A. flavus infestation rating. The total polyphenol content of seed coat showed also a significant genotypic variation (P˂0.001) (Table5). Tow genotypes ICG 12988 (18%), ICG 311(19%) showed A. flavus infestation as well as the resistant check 55-437(21%). These genotypes with 55-437 were the less infested (rate <25%). This indicated their resistance to A. flavus colonisation. They were followed by ICG 4543 (28.33%), ICG 11542(28.33%), J11 (40%) and ICG 76(42.5%) that showed A. flavus infestation rate less than 50%. The remaining genotypes showed A. flavus infestation rating more than 50% indicating high susceptibility to A. flavus colonization (Table6). The highest total polyphenol content of seed coat was observed on ICGIL 17108 (62.96%), ICG 3027 (57.50%), ICG 14630 (56.95%), ICG 12235 (56.67%) and ICGIL11125 (56%30%). All the tested genotypes showed more than 50% A. flavus infestation rating. While, the lowest seed coat %TPP content was showed by ICG 532 (42.74%), ICG 1415(43.68%), ICG 5609 (43.74%), and ICG 11542 (43.90%) (Table6). Then only ICG 11542 showed less than 50% of A. flavus colonization rating.
Table 5.
: Results of variance analysis (F value), F (probability), Means, LSD (least significant differences of means 5% level) of the Percentage of seed coat Total Polyphenol (%TPP) and the percentage of Aspergillus Flavus Colonisation.
Table 5.
: Results of variance analysis (F value), F (probability), Means, LSD (least significant differences of means 5% level) of the Percentage of seed coat Total Polyphenol (%TPP) and the percentage of Aspergillus Flavus Colonisation.
| Genotypes | %TPP | % AFC |
|---|---|---|
| Means | 50.50 | 85 |
| LSD | 2.54 | 16.63 |
| F value | 19.38 | 4.12 |
| F (Prob) | <.001 | <.001 |
Significance at 0.05, 0.01 and 0.001 level.
Table 6.
Categorisation of fifty-five genotypes according to the percentage of A. flavus colonization incidence and the percentage of seed coat total polyphenol content. CI= Colonization incidence; Percentage of A. flavus colonization = % AFC; percentage of Total Polyphenol =%TPP.
Table 6.
Categorisation of fifty-five genotypes according to the percentage of A. flavus colonization incidence and the percentage of seed coat total polyphenol content. CI= Colonization incidence; Percentage of A. flavus colonization = % AFC; percentage of Total Polyphenol =%TPP.
| Genotypes | % AFC | %TPP | Genotypes | % AFC | %TPP | Genotypes | % AFC | %TPP | |
| CI ≤25% | ICG 311 | 19 | 53.08 | 55-437 | 21 | 49 | ICG 12988 | 18 | 52.1 |
| CI˃25%≤50% | ICG 11322 | 50 | 50.43 | ICG 14523 | 43 | 51.61 | ICG 14523 | 43 | 51.61 |
| ICG 11542 | 28 | 43.90 | ICG 163 | 45 | 47.38 | ICG 3027 | 83 | 57.50 | |
| ICG 4543 | 28 | 47.55 | ICG 76 | 42 | 52.57 | J 11 | 40 | 55.16 | |
| CI ˃50% | Fleur 11 | 86 | 47.90 | ICG 10950 | 91 | 56.05 | ICG 1142 | 100 | 50.32 |
| ICG 12235 | 85 | 56.67 | ICG 12697 | 91 | 51.47 | ICG 12879 | 100 | 45.50 | |
| ICG 12991 | 81 | 48.75 | ICG 13099 | 66 | 50.35 | ICG 4906 | 83 | 46.86 | |
| ICG 13858 | 91 | 53.27 | ICG 14106 | 100 | 48.76 | ICG 1415 | 100 | 43.68 | |
| ICG 14630 | 81 | 56.95 | ICG 14630 | 81 | 56.95 | ICG 156 | 90 | 51.86 | |
| ICG 2019 | 100 | 52.34 | ICG 2106 | 83 | 48.63 | ICG 332 | 100 | 53.22 | |
| ICG 334 | 66 | 50.76 | ICG 36 | 66 | 49.14 | ICG 3992 | 83 | 53.17 | |
| ICG 4598 | 100 | 48.16 | ICG 4684 | 75 | 45.97 | ICG 2119 | 100 | 52.41 | |
| ICG 4750 | 65 | 47.38 | ICG 4764 | 70 | 50.23 | ICG 513 | 100 | 53.69 | |
| ICG 5195 | 100 | 52.36 | ICG 532 | 100 | 42.74 | ICG 5609 | 83 | 43.74 | |
| ICG 5663 | 81 | 49.36 | ICG 6263 | 91 | 54.01 | ICG 6407 | 61 | 52.14 | |
| ICG 6654 | 91 | 48.31 | ICG 6703 | 91 | 46.32 | ICG 6813 | 46 | 46.53 | |
| ICG 6888 | 83 | 46.09 | ICG 7181 | 100 | 45.92 | ICG 721 | 91 | 50.20 | |
| ICGIL 11102 | 100 | 53.09 | ICGIL 11110 | 75 | 51.29 | ICGIL 11114 | 75 | 54.34 | |
| ICGIL 11125 | 61 | 56.30 | ICGIL 17108 | 91 | 62.96 | JL24 | 100 | 47.93 | |
3.4. Relationship between Aflatoxin Contaminations with Total Polyphenol, Pod Yield, Seed Yield, and Haulm Yield
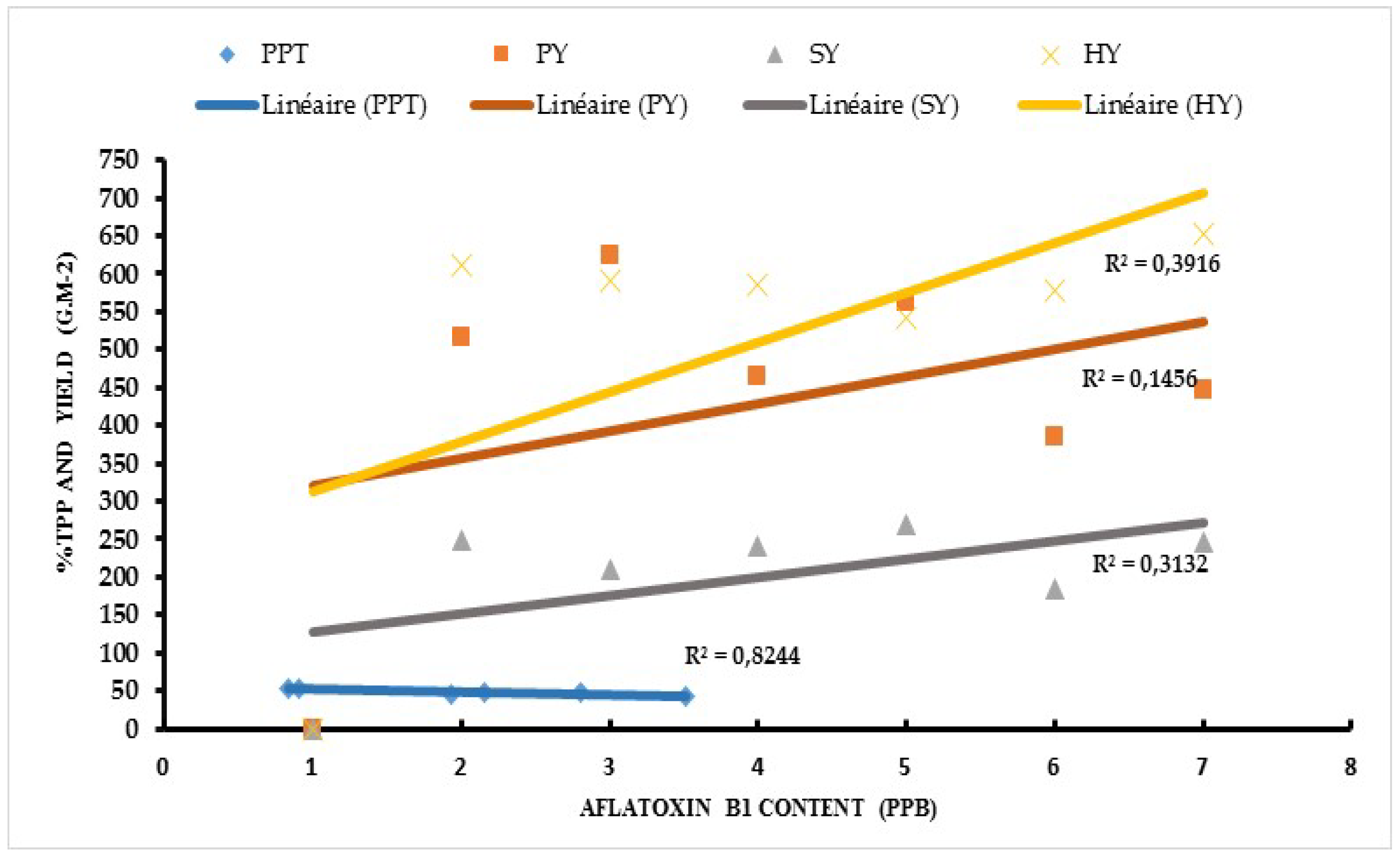
Aflatoxin contamination in resistant genotypes such as ICG 1415, ICG 2106, ICG 311, ICG 4684, ICG 4543 and the resistant check 55-437 showed a strong positive correlation with the seed coat total polyphenol content (r2=0.80; 0.82), the pod yield (r2=0.58; 0.14), seed yield (r2=0.74; 0.31) and haulm yield (r2= 0.47; 0.39) under WW and WS treatments respectively (Figure 3 and Figure 4).
Figure 3.
Relationship between Aflatoxin contamination and Seed Coat total polyphenol content, Pod Yield, Seed Yield, Haulm Yield under Well-Watered conditions (WW).
Figure 3.
Relationship between Aflatoxin contamination and Seed Coat total polyphenol content, Pod Yield, Seed Yield, Haulm Yield under Well-Watered conditions (WW).
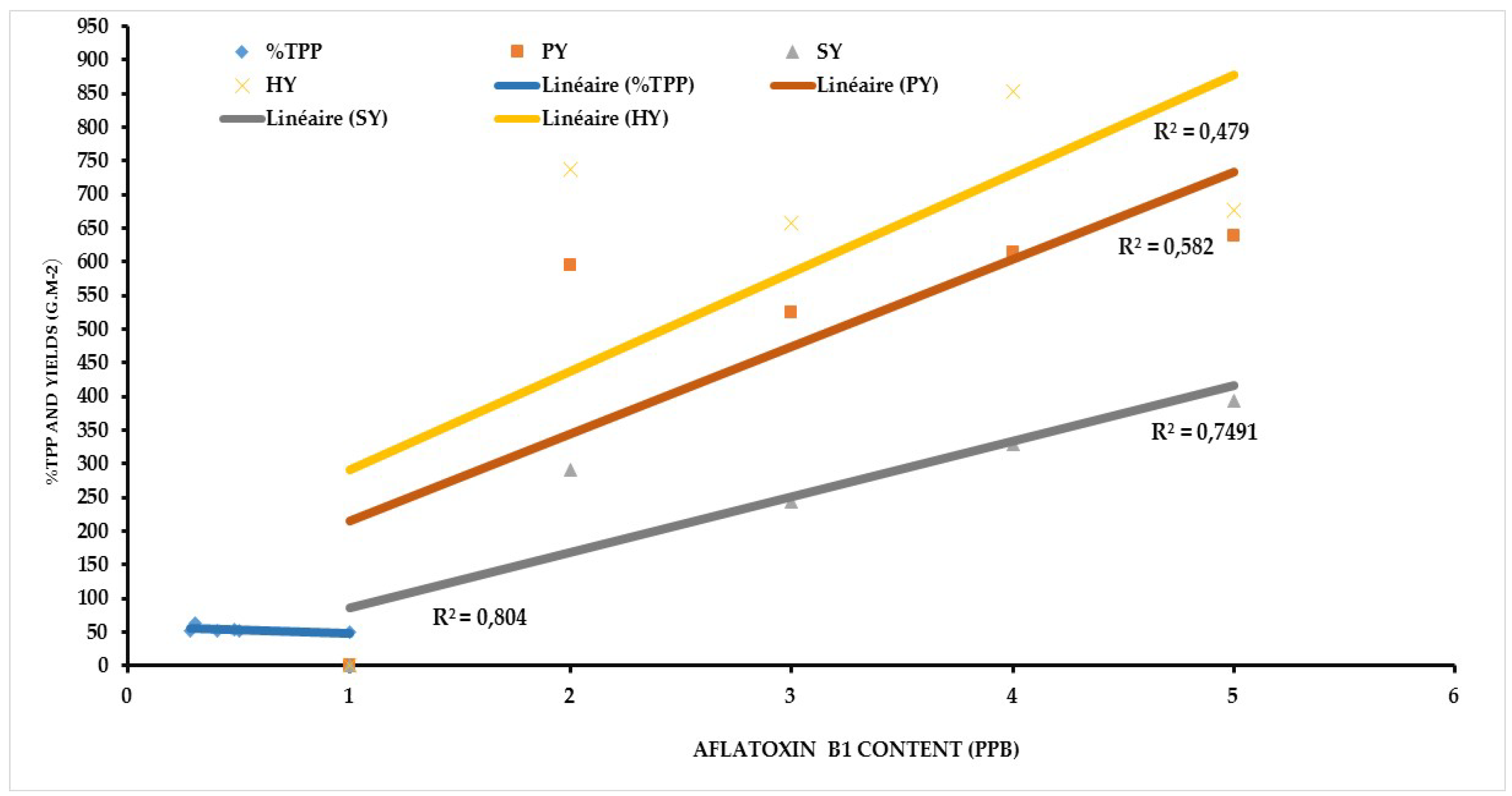
Figure 4.
Relationship between Aflatoxin contamination and Seed Coat total polyphenol content, Pod Yield, Seed Yield, Haulm Yield under Stressed conditions (WS).
Figure 4.
Relationship between Aflatoxin contamination and Seed Coat total polyphenol content, Pod Yield, Seed Yield, Haulm Yield under Stressed conditions (WS).
4. Discussion
This study revealed a wide genotypic variation for all studied traits under two water treatments. The effect of genotype and water treatment interaction (G×Wtrt) showed that genotype‘s performance differed under well-watered treatment (WW) and water stressed (WS) treatments. Indeed, pod yield (PY), seed yield (SY), and haulm yield (HY) were reduced by intermittent water deficit during seed filling up to 19.49%, 27.24% and 22.07% respectively. As for the immature pods number per plant (IMPN) and aflatoxin contamination (AC), they have increased up to 57.14% and 67.16% under WS. This indicated that intermittent drought applied from beginning seed filling to maturity reduced yields, it’s components while increased aflatoxin contamination and immature pods number. This results are similar to the previous studies which reported that water deficit imposed during 60 to 85 DAS lead to 26 % yield loss [27]. It also corroborates with others works that reported 24% of yield reduction during the end of growing season [9,28]. The top ten drought tolerant genotypes revealed in this study are: ICG 12697, ICG 12879, ICG 2019, ICG 2119, ICG 6703, ICG 4598, ICG 4684, ICG 2106, ICG 12988, and ICGIL 11114. These genotypes showed high yields under WS. This tolerance can be explained by their ability to partition dry matter into harvestable yields under limited water supply [29]. Interestingly, five (5) genotypes among these top ten drought tolerants showed low aflatoxin contamination under drought. They are: ICG 12697(2.68 ppb), ICG 2119 (0.90 ppb), ICG 4598 (3.23 ppb), ICG 4684(1.93 ppb), and ICG 2106(2.60 ppb). This indicated that these five genotypes maintained high yields under water deficit and kept intact their seed coat which protected seeds to A. flavus invasion and aflatoxin contamination. Previous studies reported that high aflatoxin levels are usually found in damaged pods compared to pods with intact shells [30]. Thus our findings are similar to the previous results which indicated that high pod yield under drought conditions is related to low seed infection and low aflatoxin contamination [31,32,33].
These last five drought tolerant and aflatoxin resistant genotypes are belonging to Spanish and Valencia botanical types. This suggested that peanut Spanish and Valencia botanical types are more resistant to aflatoxin contamination under intermittent drought during seed filling phase than Virginia type. Because of Virginia type has a long seed filling phase which exposed it more to the risk of water stress that promote pod damage, seed coat crack, A. flavus invasion and aflatoxin contamination while Spanish and Valencia types have a short seed filling phase. In addition to the top ten drought tolerant and low AC under WS, the genotypes ICG 311 (0.3ppb), ICG 14523 (2.71ppb), ICG 4543 (2.80ppb), ICG 5195 (2.83ppb), ICG 6813 (3.51ppb), and ICG 1415 (3.51ppb) showed also low aflatoxin content as the resistant check 55-437 (2.15ppb). Indeed, water deficit allowed identifying the genotypes having a specific tolerance to water deficit and keeping intact their seed coat that limited A. flavus invasion and aflatoxin contamination.
This suggest that water deficit injuries to the pods and testa enables the fungus to enter and infect the kernels [7,30,34]. Recent studies had the same conclusion for the low aflatoxin level of these genotypes under field conditions [21] and under different level of drought and temperature [35]. Furthermore, this study reveal the genotypes which content low aflatoxin content under the limitation of the European Union commission standard of total aflatoxin (4ppb) for human consumption [36]. Then, others authors reported drought tolerant genotypes have been reported to have tolerance to aflatoxin contamination [9,37].
In this study, the singnificant difference of the seed coat total polyphenol (%TPP) and the incidence of A. flavus colonization among genotypes could be due to the diversity of seed coat color, physical and chemical components. These results revealed that seed coat total polyphenol content reduced more A. flavus colonization acording to the seed coat color than to the quantity of total polyphenol in the seed coat. However, the variations of the incidence of A. flavus colonization and the percentage of total polyphenol (%TPP) were observed mainly on the genotypes ICG 311 (19%; 53.07%), ICG 2119 (100%; 52.14%), ICG 4684 (75%; 45.97%), ICG 14523 (43%; 51.61%), ICG 4543 (28%; 47.55%), ICG 5195 (100%; 52.36%), ICG 6813 (46%; 46.53%), and ICG 1415 (100%; 43.68%) respectively. These results suggests that the percentage of total polyphenol did consistently affect aflatoxin contamination. It is in agreement with several studies which reported that other phenolic content and cell wall must have effect with total polyphenol to inhibit pre-harvest aflatoxin contamination [18,19]. Thus, the pink small grain such as 55-437 that showed low incidence of A. Flavus colonization (21%) and low percentage of seed coat total polyphenol (49%) resisted more to A. flavus colonization than the dark big grain such us ICG 12235 that showed 85% of incidence of A.Flavus colonisation and 56.67% of seed coat total polyphenol. Furthermore, aflatoxin contamination under intermittent drought was significantly associated with the seed coat total polyphenol content and seed size. These results corroborate with the previous studies which reported that the difference in mycelial growth surface coverage can be explained by the difference in physical and chemical features of the seed-coat, pod-shell thickness and reticulation [19,38,39,40]. The peanut seed coat is composed of multiple cell wall layers, and peanut varieties differ in composition of flavonoids and tannins which ultimately give different colour to the peanut seed coat [18,19,41].
In this study, the following drought tolerant genotypes ICG 1415, ICG 2106, ICG 311, ICG 4684, ICG 4543 revealed significant relationship between aflatoxin resistance and seed coat total polyphenol under two water treatments. (r2=0.80; r2=0.82). It’s also associated with pod yield (r2=0.58; r2=0.14), seed yield (r2=0.74; r2=0.31), and haulm yield (r2= 0.47; r2=0.39) respectively under WW and WS. This indicated that these drought tolerant genotypes kept their seed coat intact and minimized aflatoxin contamination under intermittent drought. Thereby our findings are similar to the previous studies that reported the existence of seed coat resistance was a logical assumption, considering that seeds with damage testa are more easily and rapidly invaded by fungus than those with intact testa, and coloured testa conferred greater resistance to invasion by A. flavus than white or variegated testa [42,43].
5. Conclusions
Our findings showed that intermittent water deficit during seed filling phase decrease significantly peanut yields and increase aflatoxin contamination. The peanut genotypes with pink color of seed coat belonging to Spanish and Valencia botanical type are more resistants against A. flavus invasion and aflatoxin contamination than Virginia botanical type under intermittent water deficit because of their short seed filling phase. From this study, the genotypes ICG 12697, ICG 2119, ICG 4684, ICG 2106, ICG 311, ICG 4543, ICG 5195 and ICG 1415 showed the best performance for drought tolerance and aflatoxin minimizing. These genotypes can be used in breeding program to select the varieties that combine drought tolerance and minimize aflatoxin contamination in the semi-arid tropic.
Author Contributions
Conceptualization, Venugopal Mendu; Data curation, Maman Aminou; Methodology, Maman Aminou, Falalou Hamidou and Abdou Harou; Project administration, Venugopal Mendu; Supervision, Falalou Hamidou; Writing – original draft, Maman Aminou.
Funding
This research is funded by the United States Agency for International Development (USAID) through Cooperative Agreement No. 7200AA 18CA00003 to the University of Georgia as management entity for U.S. Feed the Future Innovation Lab for Peanut (2018-2023).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors gratefully acknowledge the Technicians of ICRISAT Niamey Crop Physiology Laboratory for field assistance and ICRISAT Genebank Niamey for providing seeds of all genotypes used in this Work.
Conflicts of Interest
Conflicts of Interest: The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Underwood C, Taylor H, Hoveland C. Soil Physical Factors Affecting Peanut Pod Development Agronomy Journal. 1971;63(6):953-4. [CrossRef]
- Horn B. Colonization of wounded peanut seeds by soil fungi: Selectivity for species from Aspergillus section Flavi. Mycologia. . 97 202-17 103852/mycologia971202. (2005). [CrossRef]
- Arunyanark A, Pimratch S, Jogloy S, Wongkaew S, Vorasoot N, Akkasaeng C, et al. Association between aflatoxin contamination and N2 fixationin peanut under drought conditions. 2012.
- Ndunguru B, Ntare B, Williams J, Greenberg D. Assessment of groundnut cultivars for end-of-season drought tolerance in a Sahelian environment. The Journal of Agricultural Science. 1995;125(1):79-85. [CrossRef]
- Hamidou F, Halilou O, Vadez V. Assessment of groundnut under combined heat and drought stress. Journal of Agronomy and Crop Science. 2013;199(1):1-11. [CrossRef]
- Waliyar F, Traore A, Fatondji D, Ntare B. Effect of irrigation interval, planting date, and cultivar on Aspergillus flavus and aflatoxin contamination of peanut in a sandy soil of Niger. Peanut Science. 2003;30(2):79-84. [CrossRef]
- Craufurd P, Prasad P, Waliyar F, Taheri A. Drought, pod yield, pre-harvest Aspergillus infection and aflatoxin contamination on peanut in Niger. Field Crops Research. 2006;98(1):20-9. [CrossRef]
- Dorner JW, Cole RJ, Sanders TH, Blankenship PD. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 1989;105(2):117-28. [CrossRef]
- Girdthai T, Jogloy S, Vorasoot N, Akkasaeng C, Wongkaew S, Holbrook CC, et al. Heritability of, and genotypic correlations between, aflatoxin traits and physiological traits for drought tolerance under end of season drought in peanut (Arachis hypogaea L.). Field Crops Research. 2010;118(2):169-76. [CrossRef]
- Waliyar F, Hassan H, Bonkoungou S. Sources of resistance to Aspergillus flavus and aflatoxin contamination in groundnut genotypes in West Africa. Plant Disease. 1994;78(7):704-8. [CrossRef]
- Keller NP, Butchko RA, Sarr B, Phillips TD. A visual pattern of mycotoxin production in maize kernels by Aspergillus spp. Phytopathology. 1994;84(5):483-8.
- Upadhyaya H, Nigam S, Mehan V, Lenne J. Aflatoxin contamination of groundnut: prospects for a genetic solution through conventional breeding. 1997.
- SOUZA FH, Marcos-Filho J. The seed coat as a modulator of seed-environment relationships in Fabaceae. Brazilian Journal of Botany. 2001;24:365-75. [CrossRef]
- Pandey MK, Kumar R, Pandey AK, Soni P, Gangurde SS, Sudini HK, et al. Mitigating aflatoxin contamination in groundnut through a combination of genetic resistance and post-harvest management practices. Toxins. 2019;11(6):315. [CrossRef]
- Arunyanark A, Jogloy S, Vorasoot N, Akkasaeng C, Kesmala T, Patanothai A. Stability of Relationship Between Chlorophyll Density and Soil Plant Analysis Development Chlorophyll Meter Readings in Peanut Across Different Drought Stress Conditions. Asian Journal of Plant Sciences. 2009;8(2):102-10. [CrossRef]
- Dieme RMA, Faye I, Zoclanclounon YAB, Fonceka D, Ndoye O, Diedhiou PM. Identification of Sources of Resistance for Peanut Aspergillus flavus Colonization and Aflatoxin Contamination. International Journal of Agronomy. 2018;2018:1-7. [CrossRef]
- Jenipher Bisikwa FO. Tolerance Levels of Peanut Varieties against Aspergillus flavus Infection. Journal of Plant Pathology & Microbiology. 2013;04(08). [CrossRef]
- Mendu L, Cobos CJ, Tengey TK, Commey L, Balasubramanian VK, Williams LD, et al. Seed coat mediated resistance against Aspergillus flavus infection in peanut. Plant Gene. 2022;32:100381. [CrossRef]
- Commey L, Tengey TK, Cobos CJ, Dampanaboina L, Dhillon KK, Pandey MK, et al. Peanut Seed Coat Acts as a Physical and Biochemical Barrier against Aspergillus flavus Infection. Journal of Fungi. 2021;7(12):1000. [CrossRef]
- Mixon A. Reducing Aspergillus species infection of peanut seed using resistant genotypes. Wiley Online Library, 1986 0047-2425. [CrossRef]
- Waliyar F, Kumar K, Diallo M, Traore A, Mangala U, Upadhyaya H, et al. Resistance to pre-harvest aflatoxin contamination in ICRISAT’s groundnut mini core collection. European Journal of Plant Pathology. 2016;145(4):901-13. [CrossRef]
- Waliyar F, Bockelee-Morvan A. Resistance of groundnut varieties to Aspergillus flavus in Senegal. 1989.
- Thakur R, Rao V, Reddy S, Ferguson M. Evaluation of wild Arachis germplasm accessions for in vitro seed colonization and anatoxin production by Aspergillus flavus. International Arachis Newsletter. 2000;20:44-6.
- Li Y, Kong W, Li M, Liu H, Zhao X, Yang S, et al. Litsea cubeba essential oil as the potential natural fumigant: Inhibition of Aspergillus flavus and AFB1 production in licorice. Industrial Crops and Products. 2016;80:186-93. [CrossRef]
- Singleton VL, Rossi JA. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. American journal of Enology and Viticulture. 1965;16(3):144-58. [CrossRef]
- Reddy S, Kiran Mayi D, Uma Reddy M, Thirumala-Devi K, Reddy D. Aflatoxins B1 in different grades of chillies (Capsicum annum L.) in India as determined by indirect competitive-ELISA. Food additives & contaminants. 2001;18(6):553-8. [CrossRef]
- Kalariya KA, Singh AL, Goswami N, Mehta D, Mahatma MK, Ajay B, et al. Photosynthetic characteristics of peanut genotypes under excess and deficit irrigation during summer. Physiology and Molecular Biology of Plants. 2015;21(3):317-27. [CrossRef]
- Boontang S, Girdthai T, Jogloy S, Akkasaeng C, Vorasoot N, Patanothai A, et al. Responses of released cultivars of peanut to terminal drought for traits related to drought tolerance. Asian Journal of Plant Sciences. 2010;9(7):423-31. [CrossRef]
- Songsri P, Jogloy S, Vorasoot N, Akkasaeng C, Patanothai A, Holbrook C. Root distribution of drought-resistant peanut genotypes in response to drought. Journal of Agronomy and Crop Science. 2008;194(2):92-103. [CrossRef]
- Sudhakar P, Latha P, Babitha M, Reddy P, Naidu P. Relationship of drought tolerance traits with aflatoxin contamination in groundnut. Indian Journal of Plant Physiology. 2007;12(3):261.
- Holbrook C, Kvien C, Rucker K, Wilson D, Hook J, Matheron M. Preharvest aflatoxin contamination in drought-tolerant and drought-intolerant peanut genotypes. Peanut Science. 2000;27(2):45-8. [CrossRef]
- Arunyanark A, Jogloy S, Wongkaew S, Akkasaeng C, Vorasoot N, Kesmala T, et al. Heritability of aflatoxin resistance traits and correlation with drought tolerance traits in peanut. Field Crops Research. 2010;117(2-3):258-64. [CrossRef]
- Amos M, Sy AT, Blaise K, Sondé ALMK. Assessment of sixteen varieties of groundnut in two agro ecological zones in Burkina Faso for yield and tolerance to aflatoxin. African Journal of Agricultural Research. 2021;17(1):66-78. [CrossRef]
- Okello DK, Kaaya AN, Bisikwa J, Were M, Oloka HK. Management of aflatoxins in groundnuts: A manual for farmers, processors, traders and consumers in Uganda. National Agricultural Research Organization; 2010.
- Hamidou F, Rathore A, Waliyar F, Vadez V. Although drought intensity increases aflatoxin contamination, drought tolerance does not lead to less aflatoxin contamination. Field crops research. 2014;156:103-10. [CrossRef]
- Wilson JS, Otsuki T. Global trade and food safety: winners and losers in a fragmented system: World Bank Publications; 2001.
- Holbrook CC, Stalker HT, Janick J. Peanut breeding and genetic resources. Plant breeding reviews. 2002;22:297-356. [CrossRef]
- LaPrade J, Bartz J, Norden A, Demuynk T. Correlation of peanut seed-coat surface wax accumulations with tolerance to colonization by Aspergillus flavus. J Am Peanut Res Educ Soc. 1973;5:89-94.
- Liang X, Zhou G, Pan R. Study on the relationship of wax and cutin layers in peanut seeds and resistance to invasion and aflatoxin production by Aspergillus flavus. J Trop Subtrop Bot. 2003;11:11-4.
- Dieme RMA, Faye I, Zoclanclounon YAB, Fonceka D, Ndoye O, Diedhiou PM. Identification of Sources of Resistance for Peanut Aspergillus flavus Colonization and Aflatoxin Contamination. International Journal of Agronomy. 2018. [CrossRef]
- Lindsey D, Turner RB. Inhibition of growth of Aspergillus flavus and Trichoderma viride by peanut embryos. Mycopathologia. 1975;55(3):149-52. [CrossRef]
- Mehan V. Screening groundnuts for resistance to seed invasion by Aspergillus flavus and to aflatoxin production. 1989.
- Kasno A, Trustinah T, Purnomo J, Sumartini S. Seed Coat Resistance of Groudnut to Aspergillus Flavus and Their Stability Performance in The Field. AGRIVITA, Journal of Agricultural Science. 2011;33(1):53-62. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



