
Submitted:
06 August 2024
Posted:
07 August 2024
You are already at the latest version
Abstract
Cys is one of the least abundant amino acids in proteins. However, it is often highly conserved and usually found in important structural and functional regions of proteins. Its unique chemical properties allow it to undergo several post-translational modifications, many of which mediated by reactive oxygen, nitrogen, sulfur or carbonyl species. Thus, in addition to their role in catalysis, protein stability and metal binding, Cys residues are crucial for redox regulation of metabolism and signal transduction. In this review, we discuss Cys post-translational modifications (PTMs) and their role in plant metabolism and signal transduction. These modifications include oxidation of the thiol group (S-sulfenylation, S-sulfinylation and S-sulfonylation), the formation of disulfide bridges, S-glutathionylation, persulfidation, S-cyanylation S-nitrosation, S-carbonylation, S-acylation, prenylation, CoAlation as well as the formation of thiohemiacetal. For each of these PTMs, we discuss the origin of the modifier, the mechanisms involved in PTM, as well as their reversibility. Examples of the involvement of Cys PTMs in the modulation of protein structure, function, stability, and localization are presented to highlight their importance in the regulation of plant metabolic and signaling pathways.
Keywords:
post-translational modification
; cysteine
; regulation
; signal transduction
; metabolism
; thiol
; redox modification
1. Introduction
Protein post-translational modifications (PTMs) are chemical or enzymatic modifications of proteins that can affect various aspects of protein activity through changes in structure, function, regulation, localization, interactions, or stability to name a few examples [1]. A large number of PTMs have been described in plants, similarly to other living organisms [1,2,3,4,5]. Due to their sessile lifestyle, PTMs are especially important for plants in which they serve as efficient regulatory mechanisms, allowing rapid and often reversible cellular responses to changes in homeostasis, as well as adjustments to alterations in metabolism, physiology or external stimuli [2,5,6]. In plants, one of the most widespread consequence resulting from modifications in their biotic or abiotic environment is an alteration in the cell redox status [7,8,9,10]. Such changes govern a variety of redox signaling events that are involved in adjusting plant metabolism or signaling pathways [7,9,11,12]. Redox signaling occurs notably via the oxidative modification of Cys residues in proteins [13].
Cys is thought to be a comparatively recent addition to the genetic code [14] and is involved in important structural and functional regions of proteins [15,16]. Since two codons translate to Cys, the latter should theoretically represent 3.3% of the amino acids in proteins [17]. However, it is underrepresented in organisms from all kingdoms of life [17]. Interestingly, Cys content in proteins increases with the complexity of the organisms, ranging from 0.4-0.5% in Archae to around 2.3% in mammals [17,18]. When it comes to plants, analyses show a similar trend, with a Cys content ranging from ∼1.5% in green algae to ∼1.9% in land plants [18,19]. The vast majority of proteins contain Cys. For instance, more than 92% of total plant protein sequences contain at least one Cys, with a median of six residues/protein [18]. In most organisms, Cys distribution in protein sequences is also peculiar, with a preference for the CXXC sequence pattern [17]. The CXXC motif is usually found in oxidoreductase and metal-binding domains [17]. However, in contrast to other organisms, plant proteins do not exhibit a high representation of this specific motif [17]. Cys also displays a distinctive conservation pattern. In most proteins, Cys is either more than 90% conserved, or less than 10% conserved, indicating a strong selective pressure to both maintain important functional Cys and remove other ones [20]. The negative selection of Cys residues appears to be stronger for isolated Cys on protein surfaces [20].
Cys has unique physico-chemical properties among all protein amino acids. Its side chain carries a thiol group (-SH) which can deprotonate as a consequence of various interactions with its environment. This loss of a proton generates a thiolate (-S-), increasing the nucleophilicity of the side chain [16]. This is a key determinant in Cys reactivity. The average pKa of protein Cys residues exposed to the solvent is around 7.5, a much lower value than that of buried Cys (around 9.5). This low pKa contributes to the fact that surface Cys are much more reactive [20]. Indeed, all protein Cys residues are not equally reactive. With a pKa value of 8.45 found for free Cys thiols, most Cys would be expected to exist in their protonated form [21]. However, the protein microenvironment can significantly affect Cys thiols pKa. For instance, basic amino acids, such as His, Arg or Lys, as well as metal ions adjacent to a Cys residue tend to lower the pKa of Cys thiol thereby stabilizing its thiolate form and promoting its reactivity [22,23]. The end-positioning of Cys residues on α-helices is another factor that affects their pKa. It has long been known that α-helices behave as dipoles [24]. Studies of the thioredoxin superfamily have shown that the dipole properties of an α-helix, together with the localization of a Cys at the N-terminus of the helix contributes to lowering the thiol pKa, thereby enhancing its reactivity [25,26]. Another way by which the protein microenvironment might increase the reactivity of a thiol group is by decreasing the activation energy during the transition state of the reaction involving the thiol. This has been demonstrated in the case of mechanistic studies of thiol/disulfide exchange [27,28]. In enzyme catalyzed reactions, contrary to a polar solvent, the hydrophobic environment provided by the protein does not stabilize the reactants in relation to the reaction transition state. This process reduces the activation energy required for the reaction to proceed allowing a faster reaction rate [28].
The reactivity of thiols is a crucial factor responsible for the involvement of Cys residues in multiple facets of protein function, including folding, catalysis, proteostasis and signal transduction [29]. The Cys sulfur atom possess oxidation states ranging from − 2 to + 4 [30,31], that allow a variety of redox post-translational modifications (PTMs). Disulfide-base reversible modifications of Cys residues, such as disulfide bridge or mixed disulfide with low-molecular weight thiols is often referred to as redox or thiol switches [29,32]. These play an important role in the modulation of protein activity, function, and localization in response to external stimuli. Redox signaling can involve reactive oxygen species (ROS), reactive nitrogen species (RNS), reactive carbonyl species (RCS) and reactive sulfur species (RSS) [33]. PTMs on Cys residues include S-sulfenylation, S-sulfinylation and S-sulfonylation, disulfide bridge (S-S) formation, S-glutathionylation, persulfidation, S-cyanylation, S-nitrosation, S-carbonylation, S-acylation, prenylation, CoAlation and thiohemiacetal formation. An analysis of the literature shows the increasing complexity of Cys modifications in plants and reveals evidence of a growing recognition of the importance of these PTMs in the regulation of metabolism and signal transduction. For reference, a searchable database catalogs modified proteins and PTM sites in several model plants (Plant PTM Viewer 2.0, https://www.psb.ugent.be/webtools/ptm-viewer/) [4]. Another database, based on deep learning framework was recently established to help facilitate protein Cys modifications in eukaryotes (pCysMod, http://pcysmod.omicsbio.info/) [34]. Unsurprisingly, many plant proteins are identified as targets for multiple PTMs. Indeed, since multiple molecules present in the same cell compartment at the same time are able to interact with reactive thiols, multiple Cys modifications can compete with each other in vivo, adding a layer of complexity in the understanding of Cys-mediated signaling in plant cells [33,35,36,37].
The aim of this review is to provide a comprehensive survey of the various PTMs of Cys residues identified in plants. For each modification, we evaluate the current state of knowledge on mechanisms facilitating PTM and its reversion, whether spontaneous or enzymatic. For each PTM, examples of targets involved in metabolism and/or signal transduction are identified and the implication of the modification in the regulation of these targets are critically discussed in relation to their function. In some cases, gaps in knowledge and possible future avenues of research on Cys PTMs in plants are also identified.
2. S-sulfenylation
Under normal conditions, aerobic metabolic processes, such as photosynthesis and respiration, constantly produce low levels of ROS as by-products. ROS are present in the cell in different forms, such as O2●− (superoxide radical), H2O2 (hydrogen peroxide) and ●OH (hydroxyl radical) [38]. Various biotic and abiotic stresses can disrupt redox homeostasis by promoting significant increases in ROS, leading to oxidative stress. Basal and stress-induced ROS production and detoxification in plants has been extensively reviewed and the reader is therefore invited to consult relevant publications and references within [39,40,41,42,43,44,45,46,47,48]. ROS greatly differ in their reactivity, diffusion rate and concentration in cells [38]. They can also cause a range of reversible and irreversible damage to lipids, DNA and proteins, altering their function in the cells [41,42]. Although they were initially thought to be exclusively toxic molecules leading to oxidative distress, it is now widely recognized that ROS can also generate oxidative eustress, or ‘good stress’ [49] by having essential signaling functions [33,37,38,50].
2.1. S-Sulfenylation is Promoted by Oxidative Conditions and is a Stepping-Stone Towards Other Cys Redox PTMs
Among ROS, H2O2 has the longest half-life and the highest capacity for diffusion [41], which makes it highly suitable for redox signaling. Indeed, H2O2 acts as a second messenger in plants by diffusing in cells and across membranes via aquaporins, thereby allowing both autocrine and paracrine signaling [45]. H2O2 is relatively stable and its reaction with reduced free Cys or glutathione (GSH) is slow compared to some other ROS and RCS [49]. H2O2 generally reacts more easily with protein Cys but its reactivity for thiol oxidation is highly dependent on a favorable protein microenvironment that reduces activation energy [49]. This dependency on protein structure for reactivity allows specificity of H2O2 mediated redox signal [49]. Thiolates, which are more reactive than thiols towards H2O2, can perform a nucleophilic attack on H2O2, causing a reversible two electrons oxidation to a sulfenic acid (-SOH) [51], potentially altering enzymes function and activity (Figure 1) [42]. It is noteworthy that in addition to H2O2, natural or artificial hydroperoxides and peroxynitrite can also cause thiol oxidation to sulfenic acid [31,52]. The latter is usually considered highly unstable and acts as an intermediate towards several Cys redox PTMs (Figure 1A), including, as discussed below, S-sulfinylation, S-sulfonylation, S-glutathionylation, S-S formation, or persulfidation [22]. As discussed in the section below, there is also an enzymatic pathway responsible for the generation of S-sulfenylated Cys. This occurs in instances where a S-sulfinylated Cys can be reduced by a sulfiredoxin (SRX) [53]. The stability of the S-sulfenylated Cys is mainly determined by its molecular environment. Improvement of sulfenic acid stability is determined by decreased solvent accessibility, absence of a proximal Cys that could induce the formation of a S-S and stabilization of the sulfenate by an H-bond network with adjacent amino acids [22]. In addition, the sulfenic acid has a unique reactivity since it can act both as a nucleophile and an electrophile [31]. For instance, the nucleophilic reactions of sulfenic acid include its overoxidation to sulfinic acid [31]. For this, the sulfenic acid performs a nucleophilic attack on H2O2, leading to irreversible sulfinic and sulfonic forms of oxidation discussed below. Electrophilic sulfenic acid reactions lead for example to Cys persulfidation which involves a reaction with H2S and cannot occur with a non-oxidized thiolate (see section below, [54]). As well, sulfenic acid can react with a thiol to create an intramolecular or intermolecular S-S or a mixed disulfide [31] (see S-S and S-glutathionylation sections below). The ability of sulfenic acid to act as an electrophile has also been exploited by using its reactivity towards 5,5-dimethyl-1,3-cyclohexanedione (dimedone). This highly selective reaction has been used to develop chemoselective dimedone-based probes enabling the detection of sulfenylated proteins in cells [55]. More recently, a more reactive benzo[c][1,2]thiazine-based (BTD) probe [56] was used to identify the Arabidopsis thaliana (Arabidopsis) sulfenome [57]. Chemoselective methods for surveying and identifying the different levels of Cys thiol oxidation have been recently reviewed [58].
2.2. S-Sulfenylation is a Reversible Primary Cys Oxidation
The reversion of Cys thiol oxidation from its sulfenic acid back to the thiol form is possible (Figure 1). In vitro, it can be achieved using a variety of reducing agents such as dithiothreitol (DTT), arsenite [31] or ascorbate in the case of 1-Cys peroxiredoxins (PRXs) [59,60]. The pool of ascorbate in normally highly reduced in plants [61]. Thus, the in vitro activity of ascorbate for the reduction of Cys sulfenic acid may have some relevance in vivo, although this remains to be seen. Nevertheless, the role of ascorbate in this process has been known for some time in animals [62]. The reduction of sulfenic acid can also occur using other mechanisms [63]. Methionine sulfoxide reductase B1 (MSRB1) is a 1-Cys reductase involved in the reduction of Met sulfoxide back to Met, allowing proteins that carry oxidized Met to return to their basic state. In the process, the catalytic Cys of MSRB1 becomes oxidized to a sulfenic acid. The reaction of the S-sulfenylated catalytic Cys with GSH leads to its S-glutathionylation (see section below for mechanistic details). The S-glutathionylated Cys can then be reduced back to its thiol form by a glutaredoxin (GRX) [63]. The study demonstrated that GRXC4 and GRXS12 could carry out the reaction in vitro. In addition to GSH, some TRX can act as an electron donor for the reduction of S-sulfenylated Cys. While the latter work showed a lack of capacity for TRXh1 to reduce S-sulfenylated MSRB1 [63], MSRB2 was a substrate. A subsequent study provided strong evidence that S-sulfenylated MSRB1 could be reduced by TRX CDSP32 under physiological conditions in a process that involved the formation of a S-S [64]. More recently, the enzymatic reduction of protein sulfenic acid has also been demonstrated in vitro for EF-Tu, a redox sensitive chloroplastic translation elongation factor of Arabidopsis [65]. The study identified that TRX f1 mediated the reduction of S-sulfenylated Cys149. Thus, various pathways may contribute to the conversion of Cys sulfenic acid to its thiol form, and much remains to be investigated concerning the possible differences in efficiency of the various actors in the process.
2.3. S-sulfenylation as a Redox-Control Mechanism in Plant Primary Metabolism
ROS-mediated oxidation modulates the activity of many enzymes involved in metabolism and stress response. Cellular oxidative conditions, as discussed above, are conducive to protein S-sulfenylation. A survey of Arabidopsis cell culture sulfenylome revealed that more than 1,000 proteins were S-sulfenylated in response to treatments with H2O2 (up to 400 µM), with an average of 1.5 modification/protein [57]. The study noted a particular enrichment in protein classes belonging to important metabolic pathways, while several S-sulfenylated sites were predicted or proved to have a functional importance. Among these in vivo targets, cytosolic glyceraldehyde-3-phosphate dehydrogenase (GAPDH) had been previously shown to be strongly inhibited by H2O2 [66]. In the case of GAPDH, S-sulfenylation can also lead to inhibitory S-glutathionylation (see below). A decrease in cytosolic GAPDH activity, such as documented in an Arabidopsis knockout line leads to decreased ATP and tricarboxylic cycle intermediates pools [67]. This is consistent with the fact that oxidative stress generally inhibits several aspects of respiratory metabolism [6]. Moreover, in plants and other systems, oxidative inhibition of triose phosphate metabolism enzymes in glycolysis has been linked to a redirection of C flux to the oxidative pentose phosphate pathway (OPPP) to serve for NADPH generation. [6,68]. This can be used by enzymes in the redoxin family in order to restore redox homeostasis. The evolutionary conservation of this redox-controlled metabolic switch between glycolysis and the OPPP may indicate an ancient origin for the strategy.
Lately, there has been other examples illustrating the potential for Cys S-sulfenylation as a regulatory mechanism for metabolic enzymes in the chloroplast. A first investigation has shown that the plastidial triosephosphate isomerase in Arabidopsis is inhibited by Cys74 S-sulfenylation resulting from H2O2 accumulation induced by salt stress (Figure 1B) [69]. A consequence of this inhibition was an accumulation of methylglyoxal [69], which is known to promote H2O2 formation [70], thus creating a feedback loop. More recently, examination of the redox properties of plastidial NAD-dependent malate dehydrogenase revealed four redox-active Cys [71]. It was determined that, following in vitro oxidative treatment, a great proportion of Cys129 was S-sulfenylated. This residue was further found responsible for the reversible oxidative inhibition of the enzyme using a directed mutagenesis study. Interestingly, reduced or oxidized nicotinamide adenine dinucleotide cofactors offered a relatively high level of protection against plastidial NAD-dependent malate dehydrogenase inhibition [71]. This proposed S-sulfenylation regulatory mechanism could impact stromal compartment dicarboxylate metabolism in a redox dependent manner. However, this remains to be further established using in vivo approaches.
2.4. Involvement of Protein S-Sulfenylation in Stress Signal Transduction
In the past years, several proteomic surveys have illustrated the importance of S-sulfenylation-dependent mechanisms in plant stress signaling [57,72,73,74]. Current approaches to study this topic take advantage of above-mentioned dimedone-based sulfenate probes in vivo. A recent publication illustrates the role played by Respiratory Burst Oxidase Homolog (RBOH) in protein S-sulfenylation during the response to pathogen [74]. RBOH is a plasma membrane localized NADPH oxidase that generates O2●− in the apoplast [75]. H2O2 is then formed upon detoxification of O2●− by a superoxide dismutase present in the extracellular space [76]. RBOH is an important player involved in the recognition of Pathogen-Associated Molecular Patterns (PAMPs) [77]. Upon recognition by pathogens-derived signals, plasma membrane receptors trigger a phospho-relay signaling cascade that leads to the activating phosphorylation of RBOH by Ca2+-dependent protein kinases and Botrytis-Induced Kinase 1 (BIK1) [77,78]. Mitogen Activated Protein kinases (MAPKs) cascades are also activated in this process, resulting in the transcriptional activation of PAMP-related genes [79]. ROS production occurring during pathogen-triggered RBOH activation plays a role in setting off cellular oxidative conditions that govern plant pathogen responses. This can lead to hormonal signaling, metabolic reprograming, protein redox-PTMs or cell death related to hypersensitive response (HR) [74,80]. In Nicotiana benthamiana, pathogen derived signals induced a dramatic increase in protein S-sulfenylation, which was attenuated by silencing of RBOHB [74]. This strongly supports an important role for RBOH in the control over S-sulfenylation in response to pathogen stress. The study further demonstrated that pathogen signals induced HR in leaves, which was sensitive to dimedone. Since, as stated earlier, dimedone reacts with S-sulfenylated residues, it was concluded that signaling via S-sulfenylation is key for HR in response to pathogens.
MAPK cascades mediate signal transduction in response to pathogens and a variety of other stresses in plants. Several MAPKs are activated as a result of an increase in cellular or extracellular H2O2 or other oxidative conditions [75]. In most cases however, the underlying mechanistic details remain to be established. Arabidopsis proteomic surveys aiming at identifying in vivo targets of S-sulfenylation revealed that several MAPKs, and in particular MAPK4, are modified in response to H2O2 [57,72]. Following Arabidopsis cell culture exposure to H2O2, MAPK4 was shown to be modified on Cys181 which resides in the Common Docking (CD) motif, a signature MAPK protein interaction domain (Figure 1C) [57]. Using a site directed mutagenesis approach, Cys181 was demonstrated to be critical for in vitro maximal kinase activity [57]. Further analyses were conducted using transgenic plants transformed with MAPK4 variants where Cys181 was replaced by Ser (oxidation insensitive) or Asp (putative S-sulfenylation-mimic) [81]. This study demonstrated that the Cys181 to Asp mutation disrupted the proper function of the CD domain and that Cys181 is essential for adequate signal transduction in vivo. MAPK4 is involved in mediating plant response to cold, salt and pathogens as well as cell division [82,83,84]. In particular, MAPK4 is a known suppressor of resistance to Pseudomonas syringae [82]. A transgenic plant carrying the Cys181 to Asp variant phenocopied an mapk4 insertion line, while the one carrying Cys181 to Ser variant behaved as the WT [81]. The study also further documented the importance of MAPK4 Cys181 in plant growth and development. The studies discussed above offer evidence that S-sulfenylation is an important mode of signal transduction in plants and the approaches used in these works, including the use of S-sulfenylation-mimic variants offer a potent tool to study the in vivo relevance of this redox PTM. Nevertheless, while significant progress has been made, component(s) involved in the reduction of Cys181 remain to be identified.
3. S-Sulfinylation and S-Sulfonylation
Sustained oxidative conditions can lead to the spontaneous overoxidation of the sulfenic acid to sulfinic and sulfonic acid forms, which are typically associated to oxidative distress (Figure 2A). Sulfonic acid, which is the most highly oxidized species of thiol, is completely irreversible. The sulfinate level of oxidation is generally irreversible, except in the specific case of PRXs. The latter are thiol-dependent enzymes that decompose peroxides using a peroxidatic (catalytic) Cys (CysP) (Figure 2B) [53]. During the catalytic cycle of a PRX, its CysP becomes oxidized to the sulfenic acid form, which is normally reduced by a resolving Cys (CysR) [85]. This leads to the formation of a S-S between CysP and CysR, which is further reduced by a thioredoxin (TRX) to complete the catalytic cycle [85,86]. Interestingly, under oxidative conditions, the sulfenic CysP can become further oxidized to a sulfinic acid [53,85]. Such overoxidized CysP is catalytically inactive, but can be reduced by an ATP-dependent SRX [87,88] which returns the CysP to its sulfenic form [53,85]. However upon overoxidation, plant PRX oligomerizes and acquires a novel function as a chaperone (Figure 2B) [86,89], as documented in other systems [90]. This chaperone activity in overoxidized, high molecular weight PRX has been documented in vitro using insulin and citrate synthase as substrates [91,92]. The physiological relevance of PRX overoxidation and oligomerization has been studied. In most physiological stress condition, there is little change in PRX status, whereas treatment with the herbicide methyl viologen, an efficient ROS inducer [93], can effectively increase PRX molecular weight [94]. In addition, PRX can be modified by other PTMs besides overoxidation [53,95]. Thus, further research will be needed to understand the interplay between PRX PTMs, stress conditions and PRX functions.
In addition to the above-described non-enzymatic oxidation of sulfenic acid, plant cysteine oxidases (PCOs), can catalyze the addition of two oxygen atoms on a thiol group to form a sulfinic acid [96]. PCOs can be classified into two groups [97]. Group A PCOs are ubiquitous in plants and are not regulated by O2 tension, whereas PCOs of Group B are specific to spermatophytes and induced by O2 deficiency [97]. In Arabidopsis, a family of five PCOs (AtPCO1 to AtPCO5) have been described, with a higher affinity for Cys residues localized at the N-termini of proteins [96]. Recent progress has been made in the resolution of the structure of PCOs and the elucidation of their catalytic mechanism [98,99]. So far, evidence suggests that PCOs from Group A (e.g. AtPCO4 and AtPCO5) and Group B (e.g. AtPCO2) have significant similarities in the structure of their catalytic site with a cupin-like double-stranded β-helix containing a triad of His residues coordinating a metal cofactor [98,99]. PCOs are involved in the oxidation of N-terminal Cys residues of specific proteins leading to N-degron pathway dependent proteolysis. Oxidation of N-terminal Cys to the sulfinic acid form can lead to protein destabilization and degradation through the N-end rule pathway, which is conserved in mammals, bacteria and plants [100]. Indeed, overoxidation of N-terminal cysteine is required for its arginylation which then induces proteosomal degradation [100,101]. The role of Cys oxidation in the N-end rule pathway in plants was recently reviewed [102].
The implication of Cys oxidation by PCOs in the signal transduction of oxygen deficiency is supported by studies on RAP2.12, a member of group VII ETHYLENE RESPONSE FACTORs (ERF-VIIs) (Figure 2C). ERF-VIIs are important transcription factors that promote the response to low O2 stress (hypoxia) [103]. They contain a highly conserved Cys residue at the N-terminal [104]. In Arabidopsis, when O2 becomes limiting for the maintenance of aerobic metabolism, AtPCO1 and AtPCO2 become less efficient for the oxidation of RAP2.12. The kinetic properties of AtPCOs make them less active under the physiological conditions prevailing in the hypoxic cell (pH, O2 concentrations), making them excellent candidates to act as plant O2 sensors [105]. Thus, under normoxia, PCOs are active and oxidize the N-terminal Cys residue of ERF-VIIs, thereby promoting their arginylation by tRNA-ARGINYL-TRANSFERASE and their degradation by the proteasome [98,106,107]. Low O2 availability lowers PCO activity, resulting in an increased stability of ERF-VIIs, which can then fulfill their function as transcriptional activators of the hypoxic response [107]. Furthermore, its was recently demonstrated that this PCO/ERF-VII pathway is under the control the metabolic energy sensor Target Of Rapamycin through a mechanism that allows the coordination of ERF-VII-mediated response to hypoxia with the energy status of the cell [108].
Oxygen sensing and signaling via PCO and the Cys/Arg branch of the N-degron pathway also play important roles in plant development [109]. Indeed, due to lack of internal O2 transport, plant tissues display diffusion-dependent O2 gradients and some tissues, such as the shoot apical meristem (SAM), are normally in an hypoxic state [109]. This situation may allow O2 concentration-dependent stabilization of transcription factors in different cell types and thus, regulation of plant development [110], as documented in the case of LITTLE ZIPPER 2 (ZPR2) [109]. ZPR2 has a conserved Cys at position 2 and is a substrate of PCO after removal of the N-terminal Met by a Met-aminopeptidase [109]. ZPR2 functions as an activator of leaf initiation that acts by regulating class-III homeodomain-leucine zippers (HD-ZIP III), which are necessary to initiate new primordia from SAM. Thus, hypoxic conditions prevailing in the SAM stabilize ZPR2 and regulate SAM activity [109].
Another important plant process regulated by overoxidation of Cys and the N-end rule pathway is vernalization. Recombinant AtPCO1-5 can catalyze VERNALIZATION 2 (VRN2) oxidation to sulfinic acid on its N-terminal Cys2 residue in vitro [111], leading to its destabilisation and degradation. VRN2 is a major regulator of vernalization in Arabidopsis [112]. It is constitutively expressed and stays enclosed in the meristems in aerobic conditions and warm temperature [111]. Low temperatures reduce O2 diffusion and therefore its availability for both respiration and the enzymatic activity of PCOs. Cold also inhibits PCO activity and VRN2 oxidation [111]. These conditions lead to VRN2 stabilization and its subsequent accumulation in plant organs in response to hypoxia and long-term cold exposure [111]. PCOs therefore contribute to stress response signaling during hypoxia and cold exposure. Hence, the regulation of transcription factor stability by catalyzed overoxidation provides an example of how, despite being usually associated to oxidative distress, S-sulfinylation can play essential roles in oxidative eustress.
4. Disulfide Bridge Formation
Two protein Cys residues can form a covalent bond called S-S (Figure 3). The importance of S-S as a key element in protein structure and function has long been recognized. A number of kinetically distinct pathways involving inter- or intramolecular reactions can lead to their generation [113]. These include (i) a two-electron Cys oxidation; (ii) a one-electron Cys oxidation involving the formation of a thiyl radical and (iii) a thiol disulfide exchange with a preformed S-S. In order for such covalent bonding to occur, the two intervening Cys residues must come in close proximity (within a few nm) [114]. Once formed, the S-S is a stable covalent bond between two sulfur atoms. Interestingly, the distance between the sulfur atoms is significantly lower for stable structural disulfides compared to reversible, usually regulatory ones (respectively 2.05 Å vs 2.18 Å) [115]. The generation of the S-S can be facilitated by thiolation of one of the Cys residues [28,113]. It can also be promoted by S-glutathionylation of one Cys, which can promote S-S formation with an adjacent Cys [116] or by S-nitrosation of an adjacent Cys (see S-glutathionylation and S-nitrosation sections below). As well, an oxidative environment, such as the conditions prevailing in the lumen of the eukaryotic endoplasmic reticulum (ER), favors S-S formation and proper protein folding [113,117]. Pathways and mechanisms of oxidative protein folding have recently been thoroughly reviewed for plants [118]. In the ER lumen, oxidative protein folding of nascent proteins is catalyzed by protein disulfide isomerase (PDI). This process consists in a disulfide relay system in which PDI is first oxidized by an oxidized ER oxidoreductin (ERO) [119]. ERO uses O2 as an electron acceptor, producing H2O2 in the process. Reduced PDI can also catalyze the reduction of S-S on misfolded proteins (Figure 3A) [120].
S-S notably contribute to maintain protein structure and stability [15], but also serve vital regulatory purposes [121]. An important aspect of the latter function is the dynamic and reversible nature of S-S formation. Disulfide stability in proteins varies depending on a few parameters, one of which being the dihedral angle of the bond [28]. This feature is influenced by the tertiary structure of the protein. The most stable angle is found at 90°, and there is increasing strain on bonding as the angle diverges from this value [28]. However, the most important impact on disulfides in proteins is due to redox mechanisms responsible for S-S formation by oxidation and their elimination by reduction (Figure 3A). Since the early works on plant enzyme redox modifications, it has been recognized that S-S reduction is mainly catalyzed by thioredoxins (TRXs) [122]. The Arabidopsis genome contains 41 TRX genes [123]. The plant TRX system and its role in metabolism and signaling has been a major area of research and the subject of several extensive reviews in recent years, therefore the reader is directed to these resources for a more complete overview of the topic [124,125,126,127]. In addition to TRXs, there is now evidence that GRXs can reduce protein S-S [128]. Several reaction mechanisms, all using GSH as reductant, have been proposed [128].
4.1. Disulfide Bridge Reduction is an Important Regulatory Mechanism that Links Light Harvesting and CO2 Fixation in the Chloroplast
Redox regulation of metabolic enzymes by reversible S-S generation provides sensing of environmental conditions and is especially important in the chloroplast stroma (Figure 3B). This compartment contains the assimilatory enzymes of the Calvin-Benson-Bassham (CBB) cycle, as well as enzymes of the glycolysis and the pentose phosphate pathway (PPP), which use the products of photosynthesis. Plants thus need a strict control over these enzymes in order to quickly tune CO2 fixation to changes in excitation pressure and to the light/dark cycle [129,130]. Four CBB cycle enzymes are redox-regulated through reversible S-S formation: phosphoribulokinase (PRK) [131,132], heterotetrameric GAPDH [133], fructose-1,6-bisphosphatase (FBPase) [134], and sedoheptulose-1,7-bisphosphatase [135]. In all cases, enzyme activity is inhibited by S-S formation in dark conditions. In addition, homotetrameric GAPDH and PRK are also regulated by interaction with the redox-sensitive CP12 scaffold protein [136]. In this instance, S-S formation on CP12 serves to initiate an interaction between GAPDH bound to NAD+ and CP12, and results in a small decrease in GAPDH activity [137]. The formation of the a ternary PRK/GAPDH/CP12 complex is then possible with oxidized PRK, resulting in a much larger decrease in activity for both enzymes [137]. Upon illumination, the chloroplast electron transport chain provides electrons for the subsequent reduction of ferredoxin (Fd) used by Fd-dependent TRX reductase (FTR) to reduce TRX [126,138]. Through a disulfide exchange mechanism, the TRX system mediates the reduction of S-S on target enzymes [126]. A comparative study of the reduction of different targets by the FTR/TRX system suggests that the final electron transfer from TRX to the target enzyme is a rate limiting step in this redox regulatory process [139]. As noted above, CBB- cycle enzymes are activated by S-S reduction. In contrast, the first enzyme in the PPP, plastidic glucose-6-phosphate dehydrogenase is active upon formation of a S-S between Cys149 and Cys157 [140]. This oxidation promotes a change in conformation that improves enzyme efficiency. Conversely, the reduction of S-S mediated by reduced TRX f1 deactivates the enzyme.
4.2. Disulfide Bridge Formation in the Chloroplast under Dark Conditions
As illustrated above, the reduction of chloroplasts enzymes upon illumination is relatively well characterized. However, oxidation mechanisms upon a switch to dark conditions remain poorly understood. Nonetheless, recent studies are providing information that help identification of players responsible for oxidation of reduced targets in the dark (Figure 3B). As seen above, chloroplast redox regulation is highly dependent on TRXs, which must be reduced to catalyze target disulfide reduction in the light. In chloroplasts, there is a second redox pathway, which uses NADPH-dependent TRX reductase C (NTRC). NTRC is implicated in the antioxidant capacity of the chloroplast by reducing 2-Cys peroxiredoxins (2CPs), which gets oxidized while scavenging H2O2 [141]. 2CPs can also be reduced, although less efficiently, by other plastidic TRX [142]. In the Arabidopsis ntrc mutant, reduction of 2CPs is therefore mediated by the Fd/FTR/TRX system, which causes a depletion in reduced TRX, indirectly affecting the regulation of TRX targets [142]. Decreasing the level of 2CPs in the ntrc mutant background enabled recovery of the WT phenotype, indicating the important role of NTRC in chloroplast redox homeostasis by regulating 2CPs [142]. Furthermore, a study using genetically encoded redox probes provided further support for a key role of 2CPs in the oxidative inhibition of CBB cycle function [143]. Thus, the Fd/FTR/TRX system for regulation of CBB cycle enzymes and the NTRC/2CPs for H2O2 detoxification are linked by the redox status of 2CPs [142]. By draining electrons from TRXs, 2CPs allow fast oxidation of TRXs in the dark enabling inactivation of CBC enzymes within 15 min of darkness [144].
5. S-Glutathionylation
Glutathione synthesis and degradation in plants was recently extensively reviewed [47]. Briefly, the Glu-Cys ligase (GSH1) conjugates the γ-carboxyl group of Glu and the amino group of Cys. Glutathione synthase (GSH2) then uses the resulting γ-glutamylcysteine and Gly to produce glutathione. GSH2 activity is present in the cytosol and the plastid whereas the step catalyzed by GSH1 is solely localized in the chloroplast and redox regulated [47]. Thus, glutathione synthesis is linked to the plastid redox state. This sensitivity to redox is mediated by the formation of intramolecular S-S between Cys178 and Cys398 , which activates Arabidopsis GSH1 and has been proposed to act as a redox switch for glutathione synthesis [145].
Glutathione is usually present in mM concentrations in plants, mostly in its monomeric reduced form (GSH) [47]. Oxidative stress promotes the accumulation of its dimeric oxidized form (GSSG). GSSG can be recycled to its reduced form by glutathione reductase (GR), using the reducing power of NADPH [47]. GSH is involved in the cellular redox buffer and the provision of electrons to the Foyer-Halliwell–Asada cycle during H2O2 detoxification [48]. The value of the GSH/GSSG ratio is therefore linked to the removal of H2O2. In absence of stress, this ratio is normally very high [48]. The maintenance of an appropriate GSH/GSSG ratio is dependent on GRs. Indeed, lack of this activity in the cytosol or organellar compartments leads to accumulation of GSSG that can be documented using genetically encoded redox sensors [146,147].
A low cellular GSH/GSSG ratio promotes Cys S-glutathionylation [48,51]. This formation of a mixed S-S between glutathione and an accessible protein Cys residue can occur spontaneously [6]. However, with a pKa of 8.8, GSH is highly protonated and thus weakly reactive in the physiological pH range, specially towards thiols [148]. Thus, the S-glutathionylation reaction (Figure 4A) can involve GSH and a sulfenic acid, or result from a disulfide exchange between GSSG and a thiolate residue [6,148,149]. Nitrosoglutathione (GSNO) has also been shown to act as a mediator of protein S-glutathionylation [150]. When tested as a S-glutathionylation agent, GSNO was differently effective on various targets [150]. In animals, S-glutathionylation appears to be at least partially catalyzed. A study on human glyoxalase II revealed that this enzyme could mediate in vitro S-glutathionylation of specific targets [151]. The involvement of glyoxalase II in S-glutathionylation has not been explored in plants so far. In addition, animal glutathione S-transferase Pi (GST Pi) also promotes protein S-glutathionylation in vivo and in vitro [152]. In contrast to animals, plants lack GST Pi [153]. There is nevertheless a study that has documented the catalysis of S-glutathionylation in plants. In this research, plant GRXC2 stimulated S-glutathionylation of the Leu-rich receptor Ser/Thr protein kinase BAK1 using GSSG as substrate [154]. By this means, GRXC2 inhibited BAK1 kinase activity [154]. This mechanism could potentially allow a redox regulation of the brassinosteroid signaling pathway, in which BAK1 is active [154,155]. However, the in vivo significance of BAK1 S-glutathionylation remains to be established. There are several examples of a regulatory role of protein S-glutathionylation in glycolytic and respiratory metabolism, as reviewed lately [6].
S-glutathionylation is fully reversible (Figure 4A). In vitro, strong reductants such as DTT are commonly used to induce non-enzymatic protein deglutathionylation [66,156]. In vivo, reducing conditions such as high GSH/GSSG ratio promote removal of glutathione (deglutathionylation) [157]. A study on human PDIs showed a limited capacity for deglutathionylation in vitro [158], however, this has not been explored in plants. Some evidence for protein deglutathionylation by cyanide has also been provided in mammalian cells [159] but, so far, this possibility does not appear to have been reported in plants. Deglutathionylation is most likely catalyzed by GRXs in vivo [48,160,161]. Plant genomes encode a large GRX gene family, from approximatively 30 genes in Arabidopsis, rice and poplar to 85 in wheat [162,163]. Two catalytic mechanisms have been described for the removal of the glutathione moiety on proteins by GRXs [160]. The reduction of the mixed S-S between glutathione and a protein first involves a nucleophilic attack of the modified Cys by a thiolated GRX Cys active site. In the monothiol mechanism, the resulting S-glutathionylated GRX is subsequently reduced by GSH, generating GSSG in the process. In the dithiol mechanism, a second Cys attacks the mixed S-S between the GRX and the glutathione, resulting in the formation of a S-S between the two Cys of the GRX and the liberation of GSH. A later reduction of the S-S on the GRX allows it to become active for a new catalytic cycle. In addition to GRXs, TRXs have also been implicated in the deglutathionylation of plant proteins in vitro [66,161]. TRXs and GRXs are related proteins involved in thiol-disulfide exchange [164]. Their substrate specificity is considered to be broad and they may exhibit some limited overlap. The precise determinants of GRX and TRX substrate specificity remain poorly understood. Nevertheless, a recent modeling study has shown that electrostatic complementarity could play an important role in determining interactions between the different redoxins isoforms and their interaction partners [165]. Studies conducted in vitro on two Arabidopsis cytosolic GAPDH isoforms show that TRX can catalyze deglutathionylation of GAPDH in a GSH independent manner, although less efficiently than GRX [66].
5.1. S-Glutathionylation as a Means of Protecting Metabolic Enzymes against Irreversible Oxidation
Because of its reversibility, and the fact that deglutathionylation restores an intact thiol, S-glutathionylation has long been recognized as a means to protect protein Cys against irreversible oxidation of thiols due to S-sulfonylation (Figure 4A) [149,166]. This protective function has been documented for plant metabolic enzymes. Between a few tens and a few hundreds of proteins have been identified as S-glutathionylation targets in various plant proteomic surveys [167,168,169,170,171]. Among these, metabolic enzymes are usually abundantly represented. An example of S-glutathionylation serving as a protective mechanism against irreversible oxidation comes from a study of ascorbate peroxidases (APXs) in the red alga Galdieria partita and in tobacco [172]. APX is responsible for H2O2 detoxification in the Foyer-Halliwell–Asada cycle, but may become inactive in absence of ascorbate due to irreversibly oxidized Cys residues [172]. S-glutathionylation of several APX Cys residues was demonstrated in vitro in the presence of H2O2 and GSH, and a protective role of S-glutathionylation under oxidative stress was suggested to occur in vivo [172]. S-glutathionylation was also shown to protect Arabidopsis chloroplastic α-amylase 3 (AMY3) activity from overoxidation in vitro [161]. It is thought that this mechanism could allow the recovery of AMY3 function (stress-induced starch degradation) after exposure to oxidative conditions generated under stress [161]. Excess H2O2 can also cause the irreversible inactivation of GAPDH, a key glycolytic enzyme which is also involved in signaling. This enzyme possesses a catalytic Cys that is highly sensitive to inactivation by H2O2. In this case also, S-glutathionylation was shown to mitigate the effects of oxidative distress in vitro [66].
5.2. Metabolic Enzymes Targeted by Regulatory S-Glutathionylation under Oxidative Conditions
There are numerous examples of S-glutathionylated enzymes in plant primary metabolism. As seen above, there are multiple cases of redox regulation in chloroplast metabolism. Enzyme sensitivity to reducing power in the organelle is also mediated by S-glutathionylation [51]. An analysis of the conservation of S-glutathionylated sites in chloroplast proteins has provided evidence for evolutionary conservation for some target proteins [171]. This indicates an ancient origin for the implication of S-glutathionylation in chloroplastic stress response. As an example, S-glutathionylation of at least three chloroplastic AMY3 Cys residues has been described in Arabidopsis (Figure 4A) [161]. Among these, Cys499 and Cys587 were previously shown to be involved in a regulatory S-S reversible by TRX, in a process similar to redox-modified CBB cycle enzymes [173]. In a proposed model, S-glutathionylation of one Cys in the pair led to the formation of the S-S, resulting in spontaneous deglutathionylation of the other [161]. AMY3 deglutathionylation and S-S reduction was respectively promoted by GRX and TRX [161].
Implications of reversible S-glutathionylation in the regulation of glycolytic and respiratory metabolism was reviewed a short while ago [6]. More recently, the cytosolic NADP-dependent isocitrate dehydrogenase (cICDH) was shown to be subject to regulatory S-glutathionylation (Figure 4A) in a study that provides an example of GSNO as S-glutathionylation agent [174]. By generating 2-oxoglutarate used as a carbon skeleton in N assimilation, cICDH plays a key function at the interface between C and N metabolisms [175]. The sensitivity of cICDH was demonstrated by decreases in extractable cICDH activity from leaves of the Arabidopsis mutants impacted in H2O2 detoxification or GSSG reduction [174]. Furthermore, in vitro cICDH activity was inhibited in the presence of GSSG, GSNO, or treatment with H2O2 plus GSH [174]. In these assays, GSNO appeared to be particularly effective. Detailed analyses revealed that GSNO induced S-glutathionylation of cICDH on Cys363. Following treatments with GSNO, there was also evidence for S-nitrosation of the protein, although the targeted Cys residue(s) could not be identified. ICDH activity could be restored by GRXC1 and GRXC2 and, less efficiently, by TRXs [174].
5.3. Involvement of S-Glutathionylation in Signaling
3′-phosphoadenosine 5′-phosphate (PAP) is a product of sulfotransferases [176,177]. In some instances, PAP has been described as a potent retro-inhibitor for these enzymes [178]. It is also a product and an inhibitor of the stromal acyl carrier protein synthase [179]. PAP also partakes in retrograde signaling between the chloroplast and the nucleus, as it regulates Plastid Redox Associated Nuclear Genes (PRANGs) [180]. PAP is degraded to AMP by the chloroplastic PAP phosphatase SAL1 [181,182]. Under normal conditions, low levels of PAP are therefore controlled by SAL1 activity in a process that involves S-glutathionylation (Figure 4B). Under stress-induced oxidative conditions, Arabidopsis SAL1 activity decreases in conjunction with its dimerization, S-glutathionylation and the formation of an intramolecular S-S between evolutionary conserved Cys167 and Cys190 [183]. Treatment of SAL1 with GSSG in vitro promoted S-glutathionylation of Cys119 and Cys190 and down-regulated monomeric and dimeric SAL1 activity. These experiments also revealed the existence of a mechanism in which a prior Cys S-glutathionylation induced the formation of a S-S between Cys167 and Cys190 by means of a thiol disulfide exchange, leading to the down regulation of SAL1 [183]. This inhibition of SAL1 leads in turn to an increased steady state level of PAP which acts as a chloroplast-to-nucleus retrograde signal [181]. Later studies have led to the development of a model where PAP accumulation allows it to bind and inhibit 5′–3′ exoribonucleases involved in PRANGs expression by degradation of uncapped RNAs, interference with RNA polymerase II function and/or silencing [182].
6. Protein Persulfidation
Hydrogen sulfide (H2S) is an important intermediate in the plant sulfate assimilatory pathway [184] as well as a gaseous pollutant that can be absorbed by plants [184,185,186]. It has recently emerged as a signaling molecule in plants [35,187,188]. The modification of a protein thiol by H2S is called persulfidation. In the literature, it is also sometimes referred to as protein persulfuration or protein sulfhydration. However, this latter terminology is considered incorrect since no hydration reaction is involved [54]. This PTM allows H2S-based signal transduction. The production of H2S occurs in several subcellular compartments. During sulfate assimilation, it is generated in plastids by sulfite reductase [184] and transferred on O-acetylserine (OAS) by OAS(thiol)lyases (OSTLs) to generate Cys (Figure 5A) [184,189,190]. Apart from the sulfur assimilation pathway, the production of intracellular H2S by several other plant pathways has been recently reviewed [189,190]. These mainly involve the catabolism of Cys by L-Cys desulfhydrases (LCD), D-Cys desulfhydrases (DCD) and the cytosolic OSTL homolog DES1 Figure 5A). LCDs and DCDs produce H2S, ammonia and pyruvate in the cytosol while DES1 breaks down Cys into H2S and OAS. Other enzymes are involved, such as the mitochondrial β-cyanoalanine synthase (βCS), which catalyzes the conversion of Cys and hydrogen cyanide (HCN) to H2S and β-cyanoalanine. In addition, a small family of 3-mercaptopyruvate sulfurtransferases (MSTs) characterized in Arabidopsis [191] also generate H2S using 3-mercaptopyruvate as sulfur donor and reduced TRX or GRX as electron donors (Figure 5A). Furthermore, upon reaction with reduced Cys or GSH, they respectively generate Cys persulfide (Cys-SSH) and glutathione persulfide (GSSH) [191].
6.1. Addition of Sulfide on Cys Results from Direct Persulfidation or Transpersulfidation
Several mechanisms can lead to the spontaneous persulfidation of Cys (Figure 5A). In aqueous solution, H2S dissociates in hydrosulfide (HS-, pKa = 7.0 at 25°) and sulfide anions (S2-, pKa = 17-19 at 25C°)[54]. Thus, even if we use the term H2S in this review, the more nucleophilic HS- is probably the most abundant form at physiological pH [54]. HS- can perform a nucleophilic attack on an oxidized thiol, such as a sulfenic acid or a disulfide, but cannot react with a reduced thiol [192]. Other means of protein persulfidation have been discussed [54], such as those involving radical sulfur species (RSSH•–) or inorganic polysulfide (-S-Sn-S-) however, the relevance of these reactions in plants still remains to be clearly established. Experimentally, p-methoxyphenyl(morpholino)phosphinodithioic acid (GYY4137) and NaHS are used as sulfide donors for protein modification and physiological studies [185,193,194,195].
In Arabidopsis, the ability of MSTs to catalyze a transpersulfidation reaction (transfer of a sulfide from one protein to another, Figure 5A) has been demonstrated in vitro using roGFP2 as a model protein substrate [191]. In this reaction, MST becomes persulfidated on its catalytic Cys following interaction with its substrate 3-mercaptopyruvate and in absence of TRX or GRX. The transfer of sulfide occurs from MST to a thiolate residue on roGFP2. Following a rearrangement, the persufidated Cys forms a disulfide bridge with a nearby thiol, resulting in the generation of H2S [191]. Interestingly, the catalytic Cys of MST is also subject to inhibitory oxidation by H2O2. The persulfidation of this residue also has a protective role against irreversible oxidation of the MST [191].
Cys modification by persulfidation is reversible in vitro with artificial reducing agents such as DTT and tris(2-carboxyethyl)phosphine (TCEP) [194,196]. In animals, redoxins can reduce protein persulfides and their levels are controlled by the thioredoxin system [54,197]. It is quite possible that this process also takes place in plants, however, it remains to be formally demonstrated.
6.2. Metabolic Targets of Cys Persulfidation
Over the past decade, high-throughput proteomic methods have been developed and used to survey the extent of protein persulfidation in plants [198,199,200,201]. Based on a biotin tag switch method, a large number of persulfidated proteins have been identified in Arabidopsis showing widespread occurrence of this PTM in leaves and roots as well as its regulation by environmental conditions [199,200]. Comparison of various proteomic studies revealed that persulfidation seems far more abundant in Arabidopsis than S-nitrosylation or S-glutathionylation [202].
Persulfidation has been implicated in the regulation of key enzymes in the metabolic pathways such as Gln synthetase in N assimilation, G6PDH in the OPPP, GAPDH in glycolysis, cytosolic ascorbate peroxidase (APX) in the ascorbate-glutathione cycle and aminocyclopropane-carboxylic acid oxidase (ACO) in ethylene synthesis (Figure 5B) [194,203]. In a study of APX, in vitro activity was shown to be modestly stimulated by persulfidation, whereas Gln synthetase was inhibited by a treatment with NaHS at nM concentration [194]. For ACO, incubation of recombinant protein with NaHS led to the reversible inhibition by persulfidation of Cys60, and treatments of Solanum lycopersicum (tomato) plants with the H2S donor further supported in vivo enzyme activity inhibition by persulfidation [203]. In a recent study on Arabidopsis and tomato G6PDHs, persulfidation was shown to play an important role in the regulation of the activity of this enzyme, which catalyzes C entry in the OPPP [204]. G6PDH modification was detected on Cys155 in Arabidopsis G6PDH6 and Cys159 in tomato G6PDHC. The same residues was also shown to be sensitive to oxidation by H2O2. G6PDHs are structurally relatively well conserved between plants and animals. The fact that the persulfide-modified residues are only found in cytosolic isoforms suggests a plant- and isoform-specific modification [204]. In vitro and in vivo treatments with NaHS inducing persulfidation of G6PDH6 and G6PDHC enhanced enzyme activity. This effect was reversed in the presence of DTT. Further analyses indicated that G6PDH persulfidation increased affinity of NADP used as substrate and promoted enzyme oligomerization towards the formation of tetramers. In addition, exposure of Arabidopsis seedlings to salt stress caused an oxidation of Cys155 and resulted in decreased enzyme activity. However, in the presence of NaSH, a competition between oxidation and persulfidation occurred, highlighting the potential of Cys155 persulfidation in the protection of G6PDH activity under oxidative conditions.
Persulfidation of Arabidopsis cytosolic GAPDH C1 was shown to reversibly increase enzyme activity in vitro [194]. Proteomic surveys have shown that cytosolic and chloroplastic GAPDH isoforms are modified in vivo by persulfidation [199]. The relative localization to the nucleus and the cytoplasm for GAPDH isoforms C1 and C2 was compared in WT and des1 mutant of Arabidopsis [205]. Decreased localization to the nucleus was reported in the des1 background whereas treatment of the mutant with NaHS increased nuclear localization. These results support the hypothesis of a preferential nuclear localization upon GAPDH persulfidation [205]. Moonlighting functions and nuclear localization have been reported before for animal and plant GAPDHs under various stress conditions [206]. In Arabidopsis, persulfidation of GAPDH appears likely to promote its migration to the nucleus. However, its nuclear function still remains to be deciphered [205,206].
6.3. Cys Persulfidation Involvement in ABA-Mediated Stomatal Movement
A wide variety of signaling functions have been shown to be impacted by persulfidation in plants, including in abiotic stress tolerance [35,188,207]. In particular, it has been implicated in drought stress signaling by regulating ABA-mediated stomatal movement (Figure 5C) [208,209,210]. ABA regulation of guard cell function implicates a complex network of signals comprising protein kinases, H2O2, NO and H2S-regulated steps [208,211]. Initial investigations showed that in the guard cell, ABA induces a production of ROS which was linked to the activity of NADPH oxidase RBOHD and F [212]. The synthetic H2S donor GYY4137 was then shown to inhibit the activity of Nicotiana tabacum inward-rectifying K+ channel [193]. Among the protein kinases involved in ABA signal transduction, Open Stomata 1 /Sucrose nonfermenting 1-RELATED PROTEIN KINASE2.6 (OST1/SnRK2.6) is responsible for mediating the phosphorylation of Thr306 on the inward K+ channel KAT1, thereby reducing K+ uptake by the guard cell and promoting stomatal closure in Arabidopsis [213]. Recently, it has been demonstrated that ABA signaling induces the persulfidation of Cys44 and Cys205 on DES1, which is recognized as a major source of H2S in the cytosol [209]. This PTM leads to the enhancement of DES1 activity in an autoactivating mechanism. The production of H2S by DES1 leads to the persulfidation of RBOHD on Cys825 and Cys890, thereby stimulating its activity and promoting the production of ROS [209]. In turn, oxidation of the persulfidated Cys residues on DES1 due to the rise of ROS provides a negative feedback mechanism leading to a decrease in DES1 activity [209]. DES1 mediated H2S production in guard cells also contributes to the mediation of ABA signaling by promoting the persulfidation of OST1/SnRK2.6 on Cys131 and Cys137 [208]. Interestingly, persulfidation of this key protein kinase increases its activity [208] while S-nitrosylation on Cys137, which is close to the catalytic site, is inhibitory [214]. This complex cross-talk between H2S and NO signals acting as second messengers in various aspects of plant physiology has been recently reviewed [215]. So far, there is strong evidence that, in guard cells, the two molecules collaborate in the fine regulation of components of ABA signaling [187].
7. S-Cyanylation
S-cyanylation is a PTM resulting from the reaction of HCN on a protein Cys residue (Figure 6). HCN is a pollutant naturally present at low levels in the environment where it often is the result of human activity [216]. It is volatile and can dissociate to H+ and CN- when dissolved in aqueous solutions (pKa = 9.2). Therefore, in the physiological pH range, HCN mainly occurs in its undissociated form. HCN can be formed enzymatically or non-enzymatically in a variety of living systems, from bacteria to mammals [217,218,219]. It is widely recognized as a poisonous compound due to its enzyme inhibitory effects, most importantly on cytochrome c oxidase [220], although a physiological function as a gasotransmitter is also currently debated [219].
In plants, a number of pathways, many of which related to stress defense, are implicated in the generation of HCN (Figure 6). The latter is produced during the synthesis of ethylene by 1-aminocyclopropane-1-carboxylate (ACC) oxidase [221]. Therefore, biotic and abiotic stresses, as well as developmental situations that promote ethylene synthesis [222] result in HCN formation. The hydrolysis of cyanogenic glycosides by β-glycosidase is another source of HCN in a relatively large number of plant families [217,220]. This so-called cyanogenesis mechanism is thought to be an effective deterrence strategy upon wounding or attack by herbivores [220]. HCN is also formed during the synthesis of the phytoalexin camalexin [223] and in a reaction that uses glyoxylate and hydroxylamine [224]. However, the enzymatic mechanism responsible for this reaction remains elusive. Plant cellular HCN can be detoxified by β-cyanoalanine synthase (CAS) [225,226]. However, this reaction also produces HS-, which, at high concentration can inhibit cytochrome c oxidase [227]. In addition, as seen above, HS- is also a protein Cys modifier.
In animal and plant systems, HCN has been reported to form protein adducts in vivo by S-cyanylation [228,229]. HCN nucleophilic properties allow a non-enzymatic attack of S-S, including those present in proteins [219,230,231]. As a result, the bond is broken and S-cyanylation most likely occurs on the Cys residue which is the farthest from an electrophilic group [230,232]. It is thought that HCN can also attack the mixed disulfide bridge between GSH and Cys, resulting in the deglutathionylation of the Cys. In vitro, protein S-cyanylation is promoted by the use of an oxidative treatment (e.g. H2O2), presumed to induce S-S formation [229]. Although the CN adduct can be eliminated in vitro [233], S-cyanylation is regarded as an irreversible PTM in living systems [229].
There is still limited information available on the occurrence and the physiological relevance of protein S-cyanylation in plants. Nevertheless, in a ground-breaking study, feeding of an Arabidopsis CAS null mutant with ACC was used to increase endogenous HCN levels in order to detect S-cyanylated proteins [229]. This strategy allowed the identification of 163 targets. Among the modified proteins, there was an enrichment in metabolic enzymes involved in non-photosynthetic and photosynthetic carbon metabolism. One of the identified targets was Enolase 2 (ENO2), which catalyzes the penultimate step of the cytosolic glycolytic pathway. ENO2 was shown to be activated by S-cyanylation on Cys346, hinting to the possible involvement of HCN in the regulation of glycolytic flux (Figure 6). Interestingly, the locus LOS2/ENO2, which encodes for ENO2 also produces a truncated form of the protein, C-MYC BINDING PROTEIN1 (LOS2) which serves as a transcriptional regulator. The effect of HCN on this protein is however unknown. Among the other S-cyanylation targets identified in this study were Met synthase 1 and 2, as well as S-adenosyl-homocysteine hydrolase 1 (Figure 6). These enzymes are involved in Met and S-adenosyl Met metabolism. Their modification by S-cyanylation could therefore impact methylation reactions, gene silencing or ethylene synthesis [229]. However, this still remains to be established. More recently, another proteomic study using the CAS null mutant confirmed that enzymes involved in Met and S-adenosyl Met metabolism are preferred targets of S-cyanylation in plant roots [234].
Obviously, much remains to be done to further understand the physiological relevance of these findings. This will require a careful examination of the function of S-cyanylation targets. In addition, future research efforts will need to consider the fact that S-cyanylation only affects oxidized Cys residues (involved in S-S or mixed disulfide bonds).
8. S-Nitrosation
Nitric oxide (NO) is a gaseous free radical that can easily diffuse across membranes. In plants, NO synthesis can be achieved through several pathways, most of which involve the reduction of nitrite [235]. Several details in plant NO production are not yet fully understood. Although sequences with homology to animal nitric oxide synthase (NOS) have been found in plants, the role of NOS in plants NO synthesis remains elusive [235]. It appears that metabolic routes involving the reduction of NO2- contribute more likely to the production of plant NO (Figure 7A). A reaction involving nitrate reductase (NR), which catalyzes the reduction of NO3- to NO2-, has been implicated in the process [235]. To produce NO, electrons from NR are supplied to NO-forming nitrite reductase (NOFNiR) leading to the reduction of NO2- to NO (Astier et al., 2018). Hence, NR mutants (e.g. nia1 and nia2 in Arabidopsis) are often used to decrease NO production in planta [236]. Another pathway, in which the reduction of NO2- to NO by the mitochondrial electron transport chain under anaerobic conditions has been documented [237]. Following its formation, NO can be scavenged by plant hemoglobins (Phytoglobins, Pgbs), which are induced by O2 deficiency. Evidence for this was initially provided by studies in transgenic Medicago sativa (alfalfa) roots [238] and Zea mays (maize) cell cultures [239] with different Pgb expression levels. Under O2 deficiency conditions, NO levels were inversely correlated to Pgb expression level in both cases.
8.1. Mechanisms Involved in Protein S-Nitrosation
NO can react directly with protein Cys to cause a PTM called S-nitrosation. This denomination is used here to describe the modification of a Cys residue by NO instead of the commonly used S-nitrosylation, as the latter refers to the modification of a metal by NO [240]. NO plays an important role in plants, animals and bacteria as a second messenger and controls a wide variety of biological functions [241]. Direct protein S-nitrosation (Figure 7A) occurs via different reaction mechanisms of NO with protein Cys and has been reviewed in some detail [240,242]. These reactions are complex and probably facilitated by additional redox reactions under physiological conditions [242]. Briefly, direct S-nitrosation can be the result of (i) a thiolate reaction with the nitrosonium cation (NO+) formed by NO oxidation by transition metals, (ii) the reaction of NO with a protein thyil radical or (iii) a Cys thiol reaction with NO, in which the participation of a second NO molecule leads to the production of a nitrosated Cys and HNO [243]. A transnitrosation reaction can also lead to S-nitrosation [241]. The latter can be associated to the denitrosation of the NO donor [244]. Although all these mechanisms promote protein S-nitrosation, it is important to note that S-nitrosation induced by different reactions (e.g. radical reaction and transnitrosation) do not necessarily have the same protein target specificity [241,245]. However, this issue is understudied in plants.
In plant tissues GSNO, acts as a stable NO reservoir for spontaneous protein S-nitrosation [246]. GSNO is recognized as the main NO donor for transnitrosation reactions (Figure 7B), although some enzymes, such as human GAPDH, may also possess a trans-nitrosase activity [247]. Mechanisms affecting cellular GSNO will therefore indirectly impact S-nitrosation. The spontaneous reaction of the glutathione thiyl radical with NO has been described as a pathway leading to the formation of GSNO [247]. In plants, it is assumed that the main enzyme regulating levels of GSNO is S-nitrosoglutathione reductase (GSNOR), a cytosolic class III alcohol dehydrogenase implicated in GSNO catabolism [246,248,249]. GSNOR catalyzes the NADH-dependent reduction of GSNO to GSSG and NH3 [248]. This enzyme has three conserved solvent-accessible Cys that are sensitive to redox modifications. A study on Lotus japonicus revealed that LjGSNOR1 and LjGSNOR2 can be modestly activated by persulfidation and inhibited by S-glutathionylation [250]. However, the latter reaction was slow (several h) and required mM concentrations of GSSG. It is thus unclear if S-glutathionylation of GSNOR is relevant in vivo. Arabidopsis GSNOR is also inhibited by H2O2 in vitro and by conditions promoting oxidative stress in vivo [251]. Interestingly, tomato GSNOR is reversibly inhibited by S-nitrosation of solvent-accessible Cys271 [252,253]. This inhibition has been proposed to be involved in a mechanism that would allow proper NO signaling during nitrosative burst by allowing initial GSNO accumulation [252]. More recently, enzymes belonging to the aldo-keto reductase (AKR) family were identified as NADPH dependent GSNO reductase in mammals [254] and Arabidopsis [255]. Two Arabidopsis AKR4C are significantly upregulated in a GSNOR null mutant, which displays a higher NADPH-dependent GSNO reduction than the WT, suggesting that AKR4Cs are involved in GSNO homeostasis and compensate for the loss of GSNOR [255].
8.2. Protein Denitrosation
Current knowledge on plant denitrosation was recently extensively reviewed [256]. Briefly, protein denitrosation can involve enzymatic or non-enzymatic reactions (Figures 7B, 7C, 7D). In Arabidopsis, cytosolic TRXh5 can catalyze denitrosation of specific proteins [257]. Implication of other TRXs in plant protein denitrosation remains to be tested, however the high amino acid sequence similarity between TRX isoforms suggest that it is likely [256,257]. Despite their sequence similarities, TRX isoform differ in their subcellular localization and specific interactions, which could influence their substrate specificity [256]. PRX IIE is involved in a trans-denitrosation mechanism for the bZIP67 transcription factor [244]. Protein denitrosation activity was documented for animal SRX [258] and GRX [259] but the relevance of these enzymes in plant protein denitrosation remains to be investigated. Some enzymes, such as GAPDH [260] and GSNOR [261] can be denitrosated in an enzyme-independent way by direct reaction between GSH and the S-nitrosated thiol, resulting in GSNO production (see example below).
8.3. Targets of Protein S-Nitrosation in Plant Metabolism
Plant protein S-nitrosation has now been studied for over 20 years with proteomic surveys employing the biotin switch technique [262,263,264]. Several hundreds of S-nitrosated proteins, together with S-nitrosation sites have been identified. Among these, the Arabidopis GAPC1 isoform of cytosolic GAPDH is known to be strongly inhibited by GSNO mediated S-nitrosation on its catalytic Cys149 [260]. GAPC1 denitrosation is promoted in vitro by GSH, but not by the TRX system [260]. Furthermore, the GSH-dependent denitrosation is influenced by the GSH/GSNO ratio, but unaffected by the GSH/GSSG ratio [260]. These results contrast with the mechanism taking place with animal GAPDH, which is effectively denitrosated by TRX and resistant to GSH-mediated denitrosation [265,266]. Although S-nitrosation is currently more studied than denitrosation, the latter certainly deserves to be more thoroughly investigated, as there appears to be some protein and/or organism specific mechanisms at work.
S-nitrosation can also be involved in cooperative Cys modification for metabolic regulation. This remarkable mechanism has been documented with pea FBPase. This CBB cycle enzyme can be S-nitrosated by GSNO in light conditions (Figure 7C), when the enzyme is normally present in its reduced form (see Figure 3B) [267]. When fully reduced, FBPase is active. A high GSNO/GSH ratio promotes S-nitrosation of FBPase Cys153. This S-nitrosation can be reversed if GSH concentration increases (i.e. under low GSNO/GSH), leading to the formation of GSNO. If conditions do not favor GSH-dependent denitrosation, the presence S-nitrosated Cys153 induces a rapid S-S between Cys153 and adjacent Cys173 with the concomitant loss of NO. The newly formed S-S effectively inactivates the enzyme. In absence of TRX activity, such as in dark conditions, the enzyme is kept in this oxidized and inactive form. Under conditions where light favor TRX reduction, S-S can then be reduced and the enzyme becomes active [267]. FBPase, which has long been known to be regulated by reversible S-S formation. The effects of its S-nitrosation provide additional sensitivity to environmental factors that may affect NO status and the chloroplastic GSNO/GSH ratio to fine-tune the function of the CBB cycle.
8.4. Involvement of S-Nitrosation in Signal Transduction Pathways
S-nitrosation also plays an important role in signal transduction by affecting other major PTMs (e.g. sumoylation, phosphorylation or acetylation) that are involved in biochemical regulation. This topic was recently reviewed [268]. At the physiological level, it is now recognized that S-nitrosation plays a role in important aspects of plant hormone signaling [269]. It is for example the case for the ABA signaling during stomatal closure. As discussed above, the activity of protein kinase OST1/SnRK2.6 which is involved in the regulation of the KAT1 inward K+ channel is stimulated by persulfidation (Figure xC). Investigations into the regulation of this protein kinase demonstrated a negative regulation of ABA signaling by NO [270]. In particular, gsnor1-3, an Arabidopis mutant with increased NO accumulation, was insensitive to the closure of stomata induced by ABA. Further investigations demonstrated that OST1/SnRK2.6 activity was inhibited in vitro in the presence of the NO donor GSNO. This inhibition was caused by S-nitrosation of conserved Cys137, which was stimulated in vivo as a result of ABA treatment. It is possible that this work has even deeper significance. Indeed, S-nitrosation of Cys137 could be relevant to the regulations of other evolutionary-related protein kinases [270].
A second example of the importance of S-nitrosation in signaling is found in SA signal transduction. In this process, NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 (NPR1) is regulated by reversible S-nitrosation (Figure 7D). NPR1 is a key regulator of SA signaling and Systemic Acquired Resistance (SAR), was first identified using a genetic screen that aimed to identify genetic lesions causing a lack of systemic response normally induced by Pseudomonas syringae [271]. SA, which is involved in the positive regulation of SAR [272], binds to NPR1 [273]. Under normal conditions, NPR1 can be found as a high molecular weight oligomer with inter-subunit S-S (Figure 7D) [274]. NPR1 is S-nitrosated by GSNO on Cys156, which promotes its oligomerization and sequestration in the cytoplasm [275]. Upon pathogen attack, there is an increase in endogenous SA and SA binding to NPR1. Upon SA accumulation, TRXh5 catalyzes the conversion of NPR1 oligomers to monomers, which are translocated to the nucleus to activate the expression of pathogenesis-related genes [274]. The monomerisation of NPR1 involves TRXh5 in two ways: it reduces the inter-subunit S-S [275] and catalyzes its denitrosation [257].
9. S-Carbonylation by Reactive Carbonyl Species
Membrane lipids are a major target of oxidation, especially under stress, due to their abundance and proximity with ROS and free radical production sites, such as RBOH, mitochondrial and chloroplastic electron transport chains [41,276,277,278]. This leads to an oxidative degradation of polyunsaturated fatty acids (PUFA) and subsequent oxylipin production (Figure 8A). Oxylipins are also produced enzymatically, mainly by lipoxygenases (LOX) that act on linoleic acid and α-linolenic acid (Figure 8A) [276,279]. This enzymatic pathway can lead to the production of important stress signaling molecules such as cis-(+)-12-oxophytodienoic acid (OPDA) and jasmonic acid (JA) [276,279]. Oxylipins also comprise more than a dozen species which contain a reactive electrophilic α,β-unsaturated carbonyl moiety that are collectively termed reactive carbonyl species (RCSs) [280]. RCSs include, but not exclusively, compounds such as acrolein, OPDA, 4-hydroxy-2-nonenal (HNE) and malondialdehyde (MDA) [280]. In this review, we follow a restrictive definition of RCSs given in the review by Mano [280]. Nevertheless, it is important to note that RCS definition appears to vary between authors and that species such as methylglyoxal or some aldehydes are regularly referred to as RCSs, although they are not included in the previous definition [280]. The latter publication also specifies the terms reactive electrophile species and oxylipins, which are sometimes interchanged with RCSs in the literature [280].
Since lipid composition and abundance of PUFAs varies between subcellular membranes, some, such as the chloroplast appear to be a major RCS production site [281,282]. Furthermore, the hypothesis of stress- or compartment-specific RCS signatures (usually referred to as oxylipin signature) is increasingly supported by the literature [283,284,279,285]. RCS production, effect, target specificity and detoxification vary between members of the RCS group and were recently reviewed [281,286].
In plants, RCS can be detoxified enzymatically. Various enzymes are involved and they display some specificity towards the structure of their RCS substrate(s). Enzyme activities such as aldehyde dehydrogenase, aldehyde reductase, aldo-keto reductase, 2-alkenal reductase, alkenal/one oxidoreductase and glutathione transferase Tau (GST τ) have been implicated in RCS detoxification [287]. In a survey of the in vitro activity of 23 GST τ isoforms with different RCSs, acrolein and HNE were the preferred substrate of 11 isoforms [287]. Non-enzymatic RCS scavenging also occurs and involves the formation of a conjugate between RCS and polyphenols [288] or GSH. RCS detoxification by the GSH pool (Figure 8A) under conditions of high GSH/GSSG ratio is illustrated in a study of Arabidopsis overexpressing GR. Overexpression of GR led to higher levels of GSH and GSH/GSSG ratio, which was associated with enhanced RCS detoxification capacity compared to the WT [289].
9.1. Interrelations between RCS and ROS Signaling
RCS are produced downstream of ROS and increase during oxidative stress [281]. Like ROS, RCS can cause both eustress (signaling) and distress (damage) [286,290]. In a study on Chlamydomonas reinhardtii, acrolein treatment at low dose significantly upregulated genes involved in GSH, S and ascorbate metabolism in addition to redox homeostasis enzymes leading to acclimation to ROS [290]. However, RCS distress, caused by higher doses of acrolein, led to loss of cell viability [290]. RCSs could thus act downstream of ROS as signal in plant stress response [282,286,291,292,293,294]. However, it remains to be seen whether any of these effects involve Cys carbonylation.
Nevertheless, there is evidence that RCSs impact the redox network. RCSs may affect cellular glutathione contents and redox state, as illustrated with high acrolein treatments [290,294,295]. As discussed above, RCS enzymatic or non-enzymatic detoxification that consumes GSH could deplete the GSH pool and increase the GSSG/GSH ratio. This could indirectly impact redox-sensitive Cys PTMs, for instance, by altering protein S-glutathionylation [295]. However, a formal demonstration of this effect has yet to be provided.
RCSs have a longer half-life than ROS and their shared characteristics with lipid allows them to diffuse through membranes [296]. The greater diversity of RCS compared to ROS could contribute to allow for a greater specificity of downstream ROS signaling [291]. Thus, RCS should be considered in the pursuit of understanding how stress-specific and compartment specific signaling pathways are generated [282,297].
9.2. Protein Thiols Modification by RCSs and S-OPDAylation
RCS electrophilic β carbon allows covalent modification of targets such as nucleic acids and proteins [282]. Because of its reactivity, Cys is a prominent RCS target, although other protein amino acids (e.g. His, Lys) can also be modified [298,299]. RCS can modify Cys residues via a Michael reaction mechanism, resulting in a primary protein carbonylation [279]. The reactivity of the RCS modifier, as well as its availability which is controlled by its formation/scavenging rates thought to control the nature of protein carbonyls [281,282]. Furthermore, protein carbonyls can undergo secondary reactions leading to irreversible and complex protein modifications [282]. Thus, protein carbonylation is traditionally considered as being irreversible. However, some reports in animals show some support for thiol-dependent decarbonylation [300,301]. In plants, protein decarbonylation still appears poorly characterized. In a report, MDA-induced protein carbonylation was partially reversed, but the mechanism remains elusive [302]. It is possible that the reversibility of Cys carbonylation varies in function of the target protein and/or the primary and secondary modifications induced by RCSs.
The effect of RCS modifications on specific targets of the redox network in plants was recently reviewed [279] and Cys modifications of specific targets are beginning to be identified [303,304]. Nevertheless, progress in this area is still limited, perhaps due to the complexity of the detection of primary and secondary modifications. The cytotoxic and reactive species acrolein and HNE are among the most studied RCSs [278,295]. There are reports that these compounds may cause specific enzyme inhibition in plants [295,305]. So far, no direct evidence links these inhibitions to Cys modification. However, this hypothesis certainly deserves consideration since the enzyme activity of several reported RCS targets [295] is known to be sensitive to Cys oxidation.
OPDA is structurally related to cyclopentenone prostaglandin hormones found in animals [304]. Prostaglandins are known to exert some of their physiological functions through oxidation of Cys thiols [306]. In plants, recent studies demonstrated that OPDA can react with protein thiols, resulting in S-OPDAylation [304]. In vitro TRXf1 modification by OPDA was detected and Cys modification was documented by mass spectrometry. This S-OPDAylation led to an inhibition of TRXf1 to reductively activate fructose-1,6-bisphosphatase (Figure 8B) [304]. Subsequent inhibitory S-OPDAylation of chloroplast TRXm4 and cytosolic TRXh3 and h5 was also found [307]. The data suggest a unified mode of action for OPDA on TRXs, However, individual S-OPDAylated residues were not confirmed by mass spectrometry in the latter study.
ODPA is a precursor of jasmonic acid, a phytohormone implicated in response to biotic and abiotic stress responses [308]. Similarly to OPDA, which is reported to modulate key elements in plants redox regulation [307], cyclopentenone prostaglandins target essential components of the redox regulation network in animal cells under stress [306,309]. The fact that prostaglandin regulatory mechanisms by protein redox PTM provides a perspective and a possible working model worth exploring for the action mode of OPDA in plants.
10. S-Acylation
In plants, the chloroplast is the main site for de novo fatty acids production [310]. S-acylation (also known as S-palmitoylation) consists of the reversible modification of a Cys residue by a fatty acid, usually palmitic acid (C16) or stearic acid (C18), through a thioester bond [311]. Among the known lipid modifications of proteins, S-acylation is unique because of its reversibility, thus making it a dynamic means of regulation [311,312]. S-acylation is not a trivial PTM in plants. A recent study in different Arabidopsis tissues identified 1849 Cys residues modified by S-acylation on 1094 different proteins using an acyl-resin-assisted capture (acyl-RAC) strategy [313]. The meristem and the silique contained the most acylated proteins. While about half of protein acylation were contributed by a single tissue, only 6.5% were found in all tissues studied, indicative of a certain tissue specificity. S-acylation is catalyzed by a family of enzymes called Protein S-acyl transferases (PATs, also called DHHCs), that contain a Asp-His-His-Cys (DHHC) motif shown to be necessary for activity [314]. This motif is conserved in PATs found in microorganisms, animals and plants [315,316,317]. PATs are membrane proteins that are distributed in several different cellular membrane compartment (Figure 9A) [318]. The Arabidopsis genome contains 24 PATs, with half of them being targeted to the plasma membrane [318] while the rest localize to different endomembrane systems. This situation contrasts with yeast and mammalians PATs, which are predominantly localized to the ER and Golgi [318]. Another unique feature of S-acylation is that it does not occur on conserved sites or consensus sequence motifs [319]. It is rather thought that it is influenced by the ability of the catalytic site of PATs to access their target Cys [320]. It is currently unknown if PATs are redox regulated, but possible modification of the conserved Cys in the DHHC motif could have an impact on S-acylation activity. Recently, a proteomic survey identified PAT24 as a persulfidation target in Arabidopsis roots [200]. PAT24 mutants display an altered root hair phenotype, indicating that S-acylation, catalyzed by PAT24, plays an important role in root hair growth [321].
10.1. S-Acylation Appears to be Critical for Target Proteins Membrane Localization
The effect of S-acylation on proteins that lack transmembrane helices (TMHs) is an increase in their hydrophobicity, thereby promoting their association with membranes (Figure 9A). For example, among all S-acylation target proteins identified in Arabidopsis, the vast majority (73%) lacked TMH [313]. Accordingly, this suggests that S-acylation may significantly influence cellular membrane distribution for target proteins. However, many proteins already associated with membranes can also be S-acylated [322]. It is thus unlikely that the function of protein S-acylation would be limited to membrane anchoring. Early studies suggested that S-acylation of a Cys residue in close proximity to a transmembrane domain (TMD) may serve to influence the conformation of the TMD within the membrane [323]. However, recent investigations of this effect in plants suggest that a careful validation of the epitope-tag methods used to study these phenomenon is warranted before drawing strong conclusions [311]. Nevertheless, several studies document the impacts of S-acylation lie in the regulation of signal transduction and cell wall metabolism.
The cellulose synthase complex (CSC) is responsible for the synthesis of the most abundant plant polymer. The CSC is a large transmembrane protein complex which consists of at least 18 cellulose synthase (CESA) subunits [324]. It is located at the plasma membrane where it consumes UDP-Glucose on the cytoplasmic side and generates nascent cellulose microfibrils on the apoplastic side. Multiple CESA subunits were found to be S-acylated [325], with an estimated 100 S-acylated Cys per CSC. Out of the twenty-six Cys in Arabidopsis CESA7, four Cys of the Variable Region 2 (VR2) and two others, found at the C-terminal end (CT) are S-acylated and were studied using a site directed mutagenesis approach in transgenic lines. This indicated that VR2 region Cys residues were important for plant cellulose contents [325]. And that, while VR2 and CT Cys are not essential for CSC assembly at the endomembrane level up to the Golgi vesicles, they are required for the correct localization to the plasma membrane [325]. Thus, S-acylation of the CSC subunits may control cellulose production by allowing the correct intracellular trafficking of the complex to the proper site for cellulose production after its assembly on endomembranes.
The importance of protein S-acylation can further be illustrated by its impact on several signaling pathways in plants, as previously reviewed [319]. In recent years, there has been mounting evidence that Ca2+ sensing and signaling proteins are S-acylated. It was shown that S-acylation of several Ca2+ sensors Calcineurin B-Like proteins (CBLs) is required for their membrane positioning and their signaling function in ion homeostasis. For example, the proper localization of Arabidopsis CBL6 was shown to depend on S-acylation of several Cys residues at the N-terminus of the protein [326]. This also impacted the correct targeting of CBL6-interacting protein kinase in planta, implying that modification of CBL6 is responsible for the targeting of the protein complex [326].
In another study, CBL5 and the Ca2+-regulated protein kinase CPK6 were shown to be subject to a double lipid modification (N-myristoylation and S-acylation), which were required for proper localization of the two proteins to the plasma membrane (Figure 9B) [327]. The dual modification allowed the recruitment of Calcineurin B-like protein-interacting protein kinase 11 (CIPK11) by CBL5 in a process that is involved in the regulation of guard cell Slow anion channel 1 (SLAC1) (Figure 9B). SLAC1 is a key element in abscisic acid-dependent stomatal closure. Further studies using heterologous expression in Xenopus oocytes indicate that the dual lipid modification of CBL5 is critical for the activation of SLAC1 by phosphorylation by the CBL5/CIPK11 complex [327]. Thus, this example illustrates the impact of N-myristoylation and S-acylation in the regulation of stomatal function.
S-acylation is also involved in plant response to phosphate (Pi) deficiency (Figure 9C). Arabidopsis non-specific phospholipase C4 (NPC4) is a promiscuous enzymes that hydrolyzes membrane phospholipids, generating a free head group and diacylglycerol which is involved in signaling [328]. The importance of NPCs in plant signaling for growth development and stress has recently been reviewed [329]. In NPC4, S-acylation of Cys533, located at the C-terminal extremity the protein was shown to be necessary for its plasma membrane association [330]. Palmitate was identified as the main modifier. Using a mutation analysis strategy, S-acylation of Cys533 was shown to be responsible for successfully restoring NPC4 function in a npc4 mutant [330]. These data strongly support the involvement of a single S-acylation site in the proper localization of NPC4, thereby allowing it to access its substrate(s) and fulfill its signaling functions.
10.2. De-S-Acylation Players and Evidence of Their Involvement in Plant Signal Transduction
In animals, two distinct types of proteins, acyl protein thioesterases (APT) [331] and ABHD17-family depalmitoylases [332], have been implicated in the de-S-acylation of proteins by cleaving the Cys-fatty acid thioester bond. A group of 11 Arabidopsis hydrolases, named ABHD17-like acyl protein thioesterases, (ABAPTs) was recently identified. They share a conserved ABHD region with mammalian ABHD17 proteins and were shown to catalyse de-S-acylation in plants and act using a similar catalytic mechanism (Figure 9A) [333].
Plants seemingly share poor sequence homology with mammalian APT1 and APT2 [311]. Nonetheless, a few studies support a role for APT1 in plant de-S-acylation (Figure 9A, 9D) [334,335]. In Medicago species, NACsa is a transcription factor localized at the plasma membrane through Cys26 S-acylation (Figure 9D) [334]. Under drought stress condition, APT1 catalyzes NACsa de-S-acylation, thereby promoting its translocation to the nucleus and the transcription of glyoxalase I (GLXI). GLXI activation increases the GSH/GSSG ratio [334]. These data indicate that APT1 is a mediator of cellular redox potential under drought. Interestingly, APT1 is itself redox-regulated. Its sequence contains three redox sensitive Cys; Cys20, Cys22 and Cys23, that can be S-glutathionylated in control conditions [335]. S-glutathionylation of APT1 promotes its inactive, monomeric form. Drought increases intracellular H2O2, leading to oxidative stress and promoting the formation of a tetrameric APT1 complex with de-S-acylase activity. The de-S-acylation of NACsa leads increased GLXI and a higher GSH/GSSG ratio, contributing to the restoration of redox homeostasis and promoting S-glutathionylation and inactivation of APT1 [335]. As a result, APT1 behaves as an effective redox sensor during drought.
Recently, de-S-acylation was identified as a key mechanism in hormone cross talk. Brassinosteroids (BRs) are important plant hormones mainly involved in plant growth and development [336], while salicylic acid (SA) is associated with plant defense and stress response [337]. BRs signaling involves the interaction of brassinosteroid insensitive 1 (BRI1) receptor and brassinosteroids signaling kinase (BSKs) at the plasma membrane [336]. BRs binding to BRI1 leads to its activation and phosphorylation of BSKs, initiating the BRs signaling cascade [336]. In normal conditions, BSKs are maintained at the plasma membrane by S-acylation, allowing their interaction with BRI1 [338]. Stresses, such as pathogen infection, induce an important increase in SA biosynthesis and signaling [337]. SA increases the expression of ABAPT11 [338]. This promotes the de-S-acylation of BSKs, reducing their membrane localisation, thus impairing their function BRs signalisation [338].
11. Prenylation
Protein prenylation has been known to take place in plants for quite some time (see the review by [339]). Protein prenylation is the irreversible covalent modification of a Cys residue by isoprenoids. The known modifiers are 15- (farnesyl moiety) or 20- (geranylgeranyl moiety) carbon chains. These isoprenoids are formed by condensation of 5-carbon units (isopentenyl pyrophosphate and its isomer dimethylallyl pyrophosphate) that are produced by two plant isoprenoid biosynthetic routes, the mevalonate pathway in the cytoplasm and the methylerythritol 4P pathway in the chloroplast [340]. Protein modification by isoprenoids is catalyzed by protein prenyltransferases. These enzymes are classified as farnesyl transferases (FT), geranylgeranyl transferases (GGT-1) and Rab-geranylgeranyl transferases (RGGT, also called GGT-2), depending on their specificity for various isoprenoids [341]. Protein prenylation is widely considered as a means of anchoring proteins in membranes, in particular in the secretory pathway [342,343]. Prenylation occurs at the C-terminal end of proteins on a consensus site. In the case of FT and CCT-1, they modify the consensus Cys-a-a-X motif, where a represents an aliphatic residue and X any amino acid [344]. This prenylation step directs the modified protein to the endoplasmic reticulum (ER) where it is followed by the proteolytic removal of the aaX sequence is catalyzed by STE24 or RCE1 endoproteases [343]. The resulting free carboxyl residue of the prenylated Cys then becomes available for methylation by a protein-S-isoprenylcysteine O-methyltransferase (ICMT) [343]. The subcellular localization of the post-prenylation processing machinery supports an ER membrane localization for proteins with the Cys-a-a-X motif, however, they can also follow the secretory pathway [343].
The RGGT/CCT2 class recognizes targets that are bound to the Rab Escort Protein and carry consensus sequences such as Cys-Cys, Cys-X-Cys, Cys-Cys-X, Cys-Cys-X-X and Cys-Cys-X-X-X, in which both Cys present in the motif can be modified [345]. Despite the above described classification of protein prenyltransferases in two categories distinguished by consensus sequences, protein substrate recognition may not be strictly exclusive [346]. An extensive searchable protein prenylation database that includes plants is available to facilitate research on this topic (https://mendel.imp.ac.at/PrePS/index.html) [347].
Figure 10.
Protein prenylation in plants. A: Pathways responsible for prenyl group synthesis and mechanism of protein prenylation. Prenyl groups (blue colored structure representing isoprenoid C15 or C20 chains) are synthesized by the mevalonate or the methylerythritol 4P pathways. Farnesyl transferases (FT), geranylgeranyl transferases (GGT-1) and Rab-geranylgeranyl transferases (RGGT/GGT-2) modify a Cys (yellow dot) present in a Cys-A-A-X (A = aliphatic and X = any amino acid) motif located at the C terminus, directing the protein to the ER membrane. A proteolytic removal of C-terminal the AAX sequence is catalyzed by STE24 or RCE1 endo-proteases, and a methylation of the prenylated Cys is done by a protein-S-isoprenylcysteine O-methyltransferase (ICMT). B: Involvement of prenylation in the formation of an oxidative pentose phosphate pathway (OPPP) in the ER. Prenylated 6-phosphogluconolactonase (PGL2) is localized to the ER membrane and interacts with ER S-acylated (red jagged line) phosphogluconate dehydrogenases (PGD). PGL2 recruits glucose-6P dehydrogenase 5.4 (G6PD5.4) to the membrane. The result constitutes a metabolon containing the first three steps of the OPPP in ER the lumen. Such organization could improve C flux in the OPPP. See text for additional details.
Figure 10.
Protein prenylation in plants. A: Pathways responsible for prenyl group synthesis and mechanism of protein prenylation. Prenyl groups (blue colored structure representing isoprenoid C15 or C20 chains) are synthesized by the mevalonate or the methylerythritol 4P pathways. Farnesyl transferases (FT), geranylgeranyl transferases (GGT-1) and Rab-geranylgeranyl transferases (RGGT/GGT-2) modify a Cys (yellow dot) present in a Cys-A-A-X (A = aliphatic and X = any amino acid) motif located at the C terminus, directing the protein to the ER membrane. A proteolytic removal of C-terminal the AAX sequence is catalyzed by STE24 or RCE1 endo-proteases, and a methylation of the prenylated Cys is done by a protein-S-isoprenylcysteine O-methyltransferase (ICMT). B: Involvement of prenylation in the formation of an oxidative pentose phosphate pathway (OPPP) in the ER. Prenylated 6-phosphogluconolactonase (PGL2) is localized to the ER membrane and interacts with ER S-acylated (red jagged line) phosphogluconate dehydrogenases (PGD). PGL2 recruits glucose-6P dehydrogenase 5.4 (G6PD5.4) to the membrane. The result constitutes a metabolon containing the first three steps of the OPPP in ER the lumen. Such organization could improve C flux in the OPPP. See text for additional details.
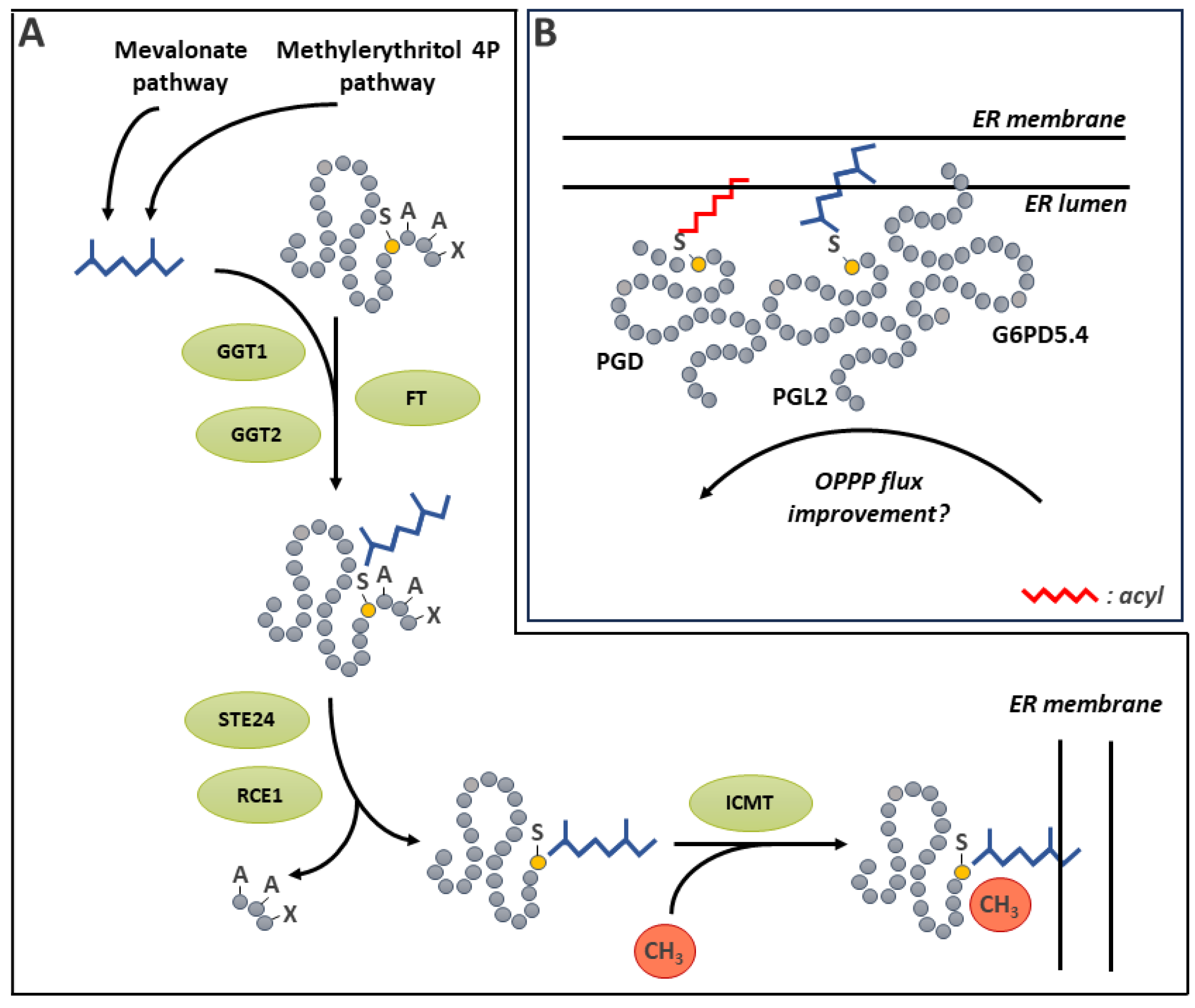
Plant protein prenylation has been the subject of numerous reviews, including a recent one on its involvement in plant stress responses [344] and another on signaling aspects [348]. We chose to only focus on a single recent study exemplifying the impact of prenylation in metabolism. This concerns the organization of the oxidative pentose phosphate pathway (OPPP) enzymes. OPPP commits carbon the for the generation of NADPH and carbon skeletons used in redox homeostasis and anabolic pathways [349]. Traditionally, this pathway has been thought to operate in the cytosol and the plastid stroma [349], while some steps have also been found in the peroxisome [350]. In Arabidopsis, one of the isoforms of the second OPPP enzyme (6-phosphogluconolactonase, PGL2) contains a C-terminal Cys-a-a-X motif [351] indicating its possible prenylation. Additionally, all three phosphogluconate dehydrogenases (PGD, the third step of the OPPP) can be S-acylated[313,351]. In an intriguing study, the presence of PGL2 at the ER was documented. Moreover, this subcellular localization was shown to be dependent on the Cys residue of the Cys-a-a-X motif. Furthermore, there is evidence that PGL2 recruits G6PD5.4 in specific ER subdomains. G6PD5.4 is a functional isoform of glucose-6P dehydrogenase arising by alternative splicing of G6PD5 and carrying an N-terminal extension serving as ER membrane anchor. Additional evidence suggests that S-acylated PGD2 may interact with the first two OPPP enzymes [351]. Thus, a combination of G6PD5.4N-terminal ER membrane targeting, together with PGL2 prenylation and PGD2 S-acylation allows the constitution of a metabolon regrouping the first three steps of the OPPP on the cytosolic side of the ER membrane. It is hypothesized that such organization should improve metabolic flux in the OPPP. Since the significance of this organization also depends on the proportion of OPPP enzymes involved in the ER metabolon relative to other subcellular localizations, the study by Linnenbrügger [351] opens up exciting future research opportunities.
12. CoAlation
Coenzyme A (CoA) is an ubiquitous cofactor in prokaryotes and eukaryotes [352]. Its five steps biosynthetic pathway from panthothenate has been elucidated for Arabidopsis [353,354]. Protein modification by CoA (CoAlation) was first described in animal [355] cells and shortly after in bacteria and Amoebozoa [356,357]. These studies were greatly facilitated with the use of an anti-CoA antibody able to detect protein CoAlation. This PTM can occur spontaneously. It was promoted by oxidative stress conditions in vivo and fully reversible in vitro using DTT [355,356,357]. A large number of CoAlation targets were identified, many of which being involved in metabolic processes [355,356]. Such widespread, and evolutionary ancient occurrence of protein CoAlation suggests it could also occur in plants. However, evidence for this is still extremely limited. A study on soybean protein Tyr phosphatase by mass spectrometry suggested the possible CoAlation of the recombinant enzyme produced in E. coli, in addition to S-glutathionylation [167]. The characterization of this PTM does not appear to have been pursued. It is unknown so far if this finding are only a reflection of the experimental conditions [167].
13. Thiohemiacetal Formation
Thiohemiacetal formation is the result of a Cys nucleophilic attack on an acyl group. It is a reversible reaction for example involved in the catalytic mechanism of a number of dehydrogenases [358,359]. In this case, the thiohemiacetal is a reaction intermediate involving a covalent bond between an enzyme’s catalytic Cys and its substrate [359]. In less common instances, however, thiohemiacetal formation may have regulatory functions. For instance, in plants, thiohemiacetal formation has been involved in the regulation of betaine aldehyde dehydrogenase (BADH). BADH catalyzes the oxidation of betaine aldehyde (BAL), which constitutes the last step in the biosynthetic pathway of the major plant osmoprotectant glycine betaine (GB) [360]. BADH catalytic Cys291 forms a thiohemiacetal as part of its catalytic mechanism. Interestingly, in a structural study of Spinacia oleracea BADH, it was shown that Cys450, a non-catalytic residue also undergoes thiohemiacetal formation with BAL [361]. This reaction hinders substrate binding, effectively inhibiting BADH activity. This mechanism was discussed as a possible regulatory mechanism that would prevent excessive NAD+ consumption during GB synthesis [361].
Studies on plant mitochondrial alternative oxidase (AOX) provide other evidence for the possible involvement of thiohemiacetal formation in regulatory mechanisms. AOX is an inner mitochondrial membrane dimeric terminal oxidase. It diverts electrons from the cytochrome pathway, bypassing complexes III and cytochrome c oxidase, thereby reducing respiratory ATP yield [362]. The reversible formation of an inter-subunit S-S is involved in AOX regulation. AOX carrying the S-S is inactive. When NAD(P)H levels are high in the mitochondrial matrix, there is reduction of the disulfide bridge, leading to an AOX with basal activity. Pyruvate, as well as various matrix α-ketoacids such as glyoxylate, oxaloacetate or α-ketoglutarate then interact with the reduced thiols and fully activate AOX. The formation of a thiohemiacetal between the reduced Cys residues and the α-keto acid has long been suspected to be involved in this regulation [363,364]. A site directed mutagenesis study has led to the proposition of a model for the activation of Arabidopsis AOX by pyruvate [364]. In this model, the S-S involving Cys78 on both subunits would block access of pyruvate to a reaction with this residue, whereas reduction of the bond would allow the Cys78 thiol to interact with pyruvate. This is consistent with the increased AOX activity obtained after substitution of Cys78 by Glu, which possess a side chain able to mimic a thiohemiacetal [364,365]. This mutant is also insensitive to pyruvate [364]. However, the detection of a thiohemiacetal by mass spectrometry remains elusive in AOX and its crystal structure is not available [365]. Thus, although converging results suggest that AOX regulation by pyruvate occurs via the formation of a thiohemiacetal, the exact mechanism still remains to be elucidated.
14. Conclusion
The chemical properties of Cys residues make them reactive to a wide variety of PTMs. With this review, we wanted to cover the range of Cys PTMs present in plants. In some cases, for example with the reversible formation of S-S, substantial research advances have been made over the years and now provide an already broad overview of the involvement this process in plant biochemistry. For several other PTMs, such as, persulfidation, S-cyanilation, S-glutathionylation, S-nitrosation, S-carbonylation or CoAlation, we are still very much in the early steps in our exploration of their implication in the molecular physiology of plants. Future progress on these issues is undoubtedly going to benefit from current advances in specialized proteomic and chemistry techniques, the setting up of resources like specialized PTM databases and the development of analytical and predictive bioinformatics tools. A point of particular interest is the fact that certain Cys residues can be modified by multiple PTMs under a variety of conditions (see, for example the GAPDH catalytic Cys mentioned in this review). This exemplifies the sometimes-complex interactions that can take place between different PTMs for a given Cys residue. We believe that we are currently only scratching the surface of this complexity.
Author Contributions
Conceptualization, J.R.; investigation, C.B; C.C. and J.R.; resources, J.R.; writing—original draft preparation, C.B and J.R.; writing—review and editing, C.B; C.C.; and J.R.; visualization, C.B; C.C.; and J.R.; supervision, J.R.; project administration, J.R.; funding acquisition, J.R. All authors have read and agreed to the published version of the manuscript.
Funding
C.B. was supported by a graduate fellowship from the Fonds de recherche du Québec – Nature et Technologies. This research was funded by the Natural Science and Engineering Research Council (Discovery grant 2019-05955 to J.R.).
Acknowledgments
C.B. and C.C. acknowledge financial support from the faculty of graduate studies of the Université de Montréal. The authors wish to tank Dr. Sonia Dorion for reviewing this manuscript and suggesting improvements.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beltrao, P.; Bork, P.; Krogan, N.J.; van Noort, V. Evolution and functional cross-talk of protein post-translational modifications. Mol. Syst. Biol. 2013, 9, 714. [Google Scholar] [CrossRef]
- Spoel, S.H. Orchestrating the proteome with post-translational modifications. J. Exp. Bot. 2018, 69, 4499–4503. [Google Scholar] [CrossRef]
- Stührwohldt, N.; Schaller, A. Regulation of plant peptide hormones and growth factors by post-translational modification. Plant Biol. 2019, 21, 49–63. [Google Scholar] [CrossRef]
- Willems, P.; Horne, A.; Van Parys, T.; Goormachtig, S.; De Smet, I.; Botzki, A.; Van Breusegem, F.; Gevaert, K. The Plant PTM Viewer, a central resource for exploring plant protein modifications. The Plant Journal 2019, 99, 752–762. [Google Scholar] [CrossRef]
- O’Leary, B.; Plaxton, W.C. Multifaceted functions of post-translational enzyme modifications in the control of plant glycolysis. Curr. Opin. Plant Biol. 2020, 55, 28–37. [Google Scholar] [CrossRef]
- Dumont, S.; Rivoal, J. Consequences of oxidative stress on plant glycolytic and respiratory metabolism. Front. Plant Sci. 2019, 10, 166. [Google Scholar] [CrossRef]
- Farooq, M.A.; Niazi, A.K.; Akhtar, J.; Saifullah; Farooq, M. ; Souri, Z.; Karimi, N.; Rengel, Z. Acquiring control: The evolution of ROS-Induced oxidative stress and redox signaling pathways in plant stress responses. Plant Physiology and Biochemistry 2019, 141, 353–369. [Google Scholar] [CrossRef]
- Mathe, C.; Garda, T.; Freytag, C.; M-Hamvas, M. The Role of Serine-Threonine Protein Phosphatase PP2A in Plant Oxidative Stress Signaling-Facts and Hypotheses. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10. [Google Scholar] [CrossRef]
- Fortunato, S.; Lasorella, C.; Dipierro, N.; Vita, F.; de Pinto, M.C. Redox Signaling in Plant Heat Stress Response. Antioxidants 2023, 12. [Google Scholar] [CrossRef]
- Chaput, V.; Martin, A.; Lejay, L. Redox metabolism: the hidden player in carbon and nitrogen signaling? J. Exp. Bot. 2020, 71, 3816–3826. [Google Scholar] [CrossRef]
- Hendrix, S.; Dard, A.; Meyer, A.J.; Reichheld, J.-P. Redox-mediated responses to high temperature in plants. J. Exp. Bot. 2023, 74, 2489–2507. [Google Scholar] [CrossRef]
- Leichert, L.I.; Dick, T.P. Incidence and physiological relevance of protein thiol switches. Biol. Chem. 2015, 396, 389–399. [Google Scholar] [CrossRef]
- Trifonov, E.N. The triplet code from first principles. J. Biomol. Struct. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef]
- Fass, D.; Thorpe, C. Chemistry and Enzymology of Disulfide Cross-Linking in Proteins. Chem. Rev. 2018, 118, 293–322. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radical Biology and Medicine 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Miseta, A.; Csutora, P. Relationship between the occurrence of cysteine in proteins and the complexity of organisms. Molecular Biology and Evolution 2000, 17, 1232–1239. [Google Scholar] [CrossRef]
- Wiedemann, C.; Kumar, A.; Lang, A.; Ohlenschläger, O. Cysteines and Disulfide Bonds as Structure-Forming Units: Insights From Different Domains of Life and the Potential for Characterization by NMR. Frontiers in Chemistry 2020, 8. [Google Scholar] [CrossRef]
- Willems, P.; Huang, J.J.; Messens, J.; Van Breusegem, F. Functionally annotating cysteine disulfides and metal binding sites in the plant kingdom using AlphaFold2 predicted structures. Free Radical Biology and Medicine 2023, 194, 220–229. [Google Scholar] [CrossRef]
- Marino, S.M.; Gladyshev, V.N. Cysteine Function Governs Its Conservation and Degeneration and Restricts Its Utilization on Protein Surfaces. J. Mol. Biol. 2010, 404, 902–916. [Google Scholar] [CrossRef]
- Clement, G.E.; Hartz, T.P. Determination of the microscopic ionization constants. J. Chem. Educ. 1971, 48, 395. [Google Scholar] [CrossRef]
- Trost, P.; Fermani, S.; Calvaresi, M.; Zaffagnini, M. Biochemical basis of sulphenomics: how protein sulphenic acids may be stabilized by the protein microenvironment. Plant Cell and Environment 2017, 40, 483–490. [Google Scholar] [CrossRef]
- Habjanič, J.; Chesnov, S.; Zerbe, O.; Freisinger, E. Impact of naturally occurring serine/cysteine variations on the structure and function of Pseudomonas metallothioneins†. Metallomics 2019, 12, 23–33. [Google Scholar] [CrossRef]
- Hol, W.G.J.; van Duijnen, P.T.; Berendsen, H.J.C. The α-helix dipole and the properties of proteins. Nature 1978, 273, 443–446. [Google Scholar] [CrossRef]
- Kortemme, T.; Creighton, T.E. Ionisation of Cysteine Residues at the Termini of Model α-Helical Peptides. Relevance to Unusual Thiol pKaValues in Proteins of the Thioredoxin Family. J. Mol. Biol. 1995, 253, 799–812. [Google Scholar] [CrossRef]
- Miranda, J.J.L. Position-dependent interactions between cysteine residues and the helix dipole. Protein Sci. 2003, 12, 73–81. [Google Scholar] [CrossRef]
- Fernandes, P.A.; Ramos, M.J. Theoretical insights into the mechanism for thiol/disulfide exchange. Chemistry-a European Journal 2004, 10, 257–266. [Google Scholar] [CrossRef]
- Netto, L.E.S.; de Oliveira, M.A.; Tairum, C.A.; Neto, J.F.D. Conferring specificity in redox pathways by enzymatic thiol/disulfide exchange reactions. Free Radic. Res. 2016, 50, 206–245. [Google Scholar] [CrossRef]
- Radzinski, M.; Oppenheim, T.; Metanis, N.; Reichmann, D. The Cys Sense: Thiol Redox Switches Mediate Life Cycles of Cellular Proteins. Biomolecules 2021, 11, 469. [Google Scholar] [CrossRef] [PubMed]
- Reddie, K.G.; Carroll, K.S. Expanding the functional diversity of proteins through cysteine oxidation. Curr. Opin. Chem. Biol. 2008, 12, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Carroll, K.S. Sulfenic acid chemistry, detection and cellular lifetime. Biochimica Et Biophysica Acta-General Subjects 2014, 1840, 847–875. [Google Scholar] [CrossRef]
- Huang, J.J.; Willems, P.; Van Breusegem, F.; Messens, J. Pathways crossing mammalian and plant sulfenomic landscapes. Free Radical Biology and Medicine 2018, 122, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.K.; Kaushik, P. Free Radicals Mediated Redox Signaling in Plant Stress Tolerance. Life-Basel 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Yu, K.; Wu, G.D.; Zhang, Q.F.; Wang, P.Q.; Zheng, J.; Liu, Z.X.; Wang, J.C.; Gao, X.J.; Cheng, H. pCysMod: Prediction of Multiple Cysteine Modifications Based on Deep Learning Framework. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Corpas, F.J.; Gonzalez-Gordo, S.; Palma, J.M. Protein nitration: A connecting bridge between nitric oxide (NO) and plant stress. Plant Stress 2021, 2. [Google Scholar] [CrossRef]
- Mukherjee, S.; Corpas, F.J. H2O2, NO, and H2S networks during root development and signalling under physiological and challenging environments: Beneficial or toxic? Plant Cell and Environment 2023, 46, 688–717. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annual Review of Plant Physiology and Plant Molecular Biology 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef]
- Khedia, J.; Agarwal, P.; Agarwal, P.K. Deciphering hydrogen peroxide-induced signalling towards stress tolerance in plants. 3 Biotech 2019, 9, 395. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Dorion, S.; Ouellet, J.C.; Rivoal, J. Glutathione metabolism in plants under stress: Beyond reactive oxygen species detoxification. Metabolites 2021, 11, 641. [Google Scholar] [CrossRef]
- Foyer, C.H.; Kunert, K. The ascorbate–glutathione cycle coming of age. J. Exp. Bot. 2024, 75, 2682–2699. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nature Reviews Molecular Cell Biology 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Cejudo, F.J.; Sandalio, L.M.; Van Breusegem, F. Understanding plant responses to stress conditions: redox-based strategies. J. Exp. Bot. 2021, 72, 5785–5788. [Google Scholar] [CrossRef]
- Vogelsang, L.; Dietz, K.J. Regulatory thiol oxidation in chloroplast metabolism, oxidative stress response and environmental signaling in plants. Biochem. J. 2020, 477, 1865–1878. [Google Scholar] [CrossRef]
- Dietz, K.J.; Jacob, S.; Oelze, M.L.; Laxa, M.; Tognetti, V.; de Miranda, S.M.; Baier, M.; Finkemeier, I. The function of peroxiredoxins in plant organelle redox metabolism. J.Exp.Bot. 2006, 57, 1697–1709. [Google Scholar] [CrossRef]
- Liebthal, M.; Maynard, D.; Dietz, K.J. Peroxiredoxins and Redox Signaling in Plants. Antioxidants & Redox Signaling 2018, 28, 609–624. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Paulsen, C.E.; Truong, T.H.; Garcia, F.J.; Homann, A.; Gupta, V.; Leonard, S.E.; Carroll, K.S. Peroxide-dependent sulfenylation of the EGFR catalytic site enhances kinase activity. Nat. Chem. Biol. 2011, 8, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Yang, J.; Liebler, D.C.; Carroll, K.S. Diverse Redoxome Reactivity Profiles of Carbon Nucleophiles. J. Am. Chem. Soc. 2017, 139, 5588–5595. [Google Scholar] [CrossRef]
- Huang, J.; Willems, P.; Wei, B.; Tian, C.; Ferreira, R.B.; Bodra, N.; Martínez Gache, S.A.; Wahni, K.; Liu, K.; Vertommen, D. Mining for protein S-sulfenylation in Arabidopsis uncovers redox-sensitive sites. Proceedings of the National Academy of Sciences 2019, 116, 21256–21261. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Carroll, K.S. Activity-Based Sensing for Site-Specific Proteomic Analysis of Cysteine Oxidation. Acc. Chem. Res. 2020, 53, 20–31. [Google Scholar] [CrossRef]
- Monteiro, G.; Horta, B.B.; Pimenta, D.C.; Augusto, O.; Netto, L.E.S. Reduction of 1-Cys peroxiredoxins by ascorbate changes the thiol-specific antioxidant paradigm, revealing another function of vitamin C. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 4886–4891. [Google Scholar] [CrossRef]
- Pulido, P.; Cazalis, R.; Cejudo, F.J. An antioxidant redox system in the nucleus of wheat seed cells suffering oxidative stress. The Plant Journal 2009, 57, 132–145. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Zito, E.; Hansen, Henning G. ; Yeo, Giles S.H.; Fujii, J.; Ron, D. Endoplasmic Reticulum Thiol Oxidase Deficiency Leads to Ascorbic Acid Depletion and Noncanonical Scurvy in Mice. Mol. Cell 2012, 48, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Tarrago, L.; Laugier, E.; Zaffagnini, M.; Marchand, C.; Le Marechal, P.; Rouhier, N.; Lemaire, S.D.; Rey, P. Regeneration mechanisms of Arabidopsis thaliana methionine sulfoxide reductases B by glutaredoxins and thioredoxins. J. Biol. Chem. 2009, 284, 18963–18971. [Google Scholar] [CrossRef] [PubMed]
- Tarrago, L.; Laugier, E.; Zaffagnini, M.; Marchand, C.H.; Le Maréchal, P.; Lemaire, S.D.; Rey, P. Plant Thioredoxin CDSP32 Regenerates 1-Cys Methionine Sulfoxide Reductase B Activity through the Direct Reduction of Sulfenic Acid *<sup></sup>. J. Biol. Chem. 2010, 285, 14964–14972. [Google Scholar] [CrossRef] [PubMed]
- Toriu, M.; Horie, M.; Kumaki, Y.; Yoneyama, T.; Kore-eda, S.; Mitsuyama, S.; Yoshida, K.; Hisabori, T.; Nishiyama, Y. Chloroplast translation factor EF-Tu of Arabidopsis thaliana can be inactivated via oxidation of a specific cysteine residue. Biochem. J. 2023, 480, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Bedhomme, M.; Adamo, M.; Marchand, C.H.; Couturier, J.; Rouhier, N.; Lemaire, S.D.; Zaffagnini, M.; Trost, P. Glutathionylation of cytosolic glyceraldehyde-3-phosphate dehydrogenase from the model plant Arabidopsis thaliana is reversed by both glutaredoxins and thioredoxins in vitro. Biochem. J. 2012, 445, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ma, F.; Wei, F.; Fanella, B.; Allen, D.K.; Wang, X. Cytosolic Phosphorylating Glyceraldehyde-3-Phosphate Dehydrogenases Affect <em>Arabidopsis</em> Cellular Metabolism and Promote Seed Oil Accumulation. Plant Cell 2014, 26, 3023–3035. [Google Scholar] [CrossRef] [PubMed]
- Talwar, D.; Miller, C.G.; Grossmann, J.; Szyrwiel, L.; Schwecke, T.; Demichev, V.; Mikecin Drazic, A.-M.; Mayakonda, A.; Lutsik, P.; Veith, C.; et al. The GAPDH redox switch safeguards reductive capacity and enables survival of stressed tumour cells. Nature Metabolism 2023, 5, 660–676. [Google Scholar] [CrossRef]
- Fu, Z.-W.; Feng, Y.-R.; Gao, X.; Ding, F.; Li, J.-H.; Yuan, T.-T.; Lu, Y.-T. Salt stress-induced chloroplastic hydrogen peroxide stimulates pdTPI sulfenylation and methylglyoxal accumulation. Plant Cell 2023, 35, 1593–1616. [Google Scholar] [CrossRef]
- Hoque, T.S.; Uraji, M.; Ye, W.; Hossain, M.A.; Nakamura, Y.; Murata, Y. Methylglyoxal-induced stomatal closure accompanied by peroxidase-mediated ROS production in Arabidopsis. J. Plant Physiol. 2012, 169, 979–986. [Google Scholar] [CrossRef]
- Cosse, M.; Rehders, T.; Eirich, J.; Finkemeier, I.; Selinski, J. Cysteine oxidation as a regulatory mechanism of Arabidopsis plastidial NAD-dependent malate dehydrogenase. Physiol. Plant. 2024, 176, e14340. [Google Scholar] [CrossRef]
- Waszczak, C.; Akter, S.; Eeckhout, D.; Persiau, G.; Wahni, K.; Bodra, N.; Van Molle, I.; De Smet, B.; Vertommen, D.; Gevaert, K. Sulfenome mining in Arabidopsis thaliana. Proceedings of the National Academy of Sciences 2014, 111, 11545–11550. [Google Scholar] [CrossRef]
- Yu, L.Q.; Dai, Z.Z.; Zhang, Y.T.; Iqbal, S.; Lu, S.P.; Guo, L.; Yao, X. Proteome-wide identification of S-sulfenylated cysteines reveals metabolic response to freezing stress after cold acclimation in <i>Brassica napus</i>. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Hino, Y.; Inada, T.; Yoshioka, M.; Yoshioka, H. NADPH oxidase-mediated sulfenylation of cysteine derivatives regulates plant immunity. J. Exp. Bot. 2024. [Google Scholar] [CrossRef]
- Liu, Y.; He, C. A review of redox signaling and the control of MAP kinase pathway in plants. Redox Biology 2017, 11, 192–204. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Kadota, Y.; Sklenar, J.; Derbyshire, P.; Stransfeld, L.; Asai, S.; Ntoukakis, V.; Jones, Jonathan D. ; Shirasu, K.; Menke, F.; Jones, A.; et al. Direct Regulation of the NADPH Oxidase RBOHD by the PRR-Associated Kinase BIK1 during Plant Immunity. Mol. Cell 2014, 54, 43–55. [Google Scholar] [CrossRef]
- Kobayashi, M.; Ohura, I.; Kawakita, K.; Yokota, N.; Fujiwara, M.; Shimamoto, K.; Doke, N.; Yoshioka, H. Calcium-Dependent Protein Kinases Regulate the Production of Reactive Oxygen Species by Potato NADPH Oxidase. Plant Cell 2007, 19, 1065–1080. [Google Scholar] [CrossRef]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nature Reviews Immunology 2016, 16, 537–552. [Google Scholar] [CrossRef]
- Chaouch, S.; Queval, G.; Noctor, G. AtRbohF is a crucial modulator of defence-associated metabolism and a key actor in the interplay between intracellular oxidative stress and pathogenesis responses in Arabidopsis. The Plant Journal 2012, 69, 613–627. [Google Scholar] [CrossRef]
- Siodmak, A.; Shahul Hameed, U.F.; Rayapuram, N.; Völz, R.; Boudsocq, M.; Alharbi, S.; Alhoraibi, H.; Lee, Y.-H.; Blilou, I.; Arold, S.T.; et al. Essential role of the CD docking motif of MPK4 in plant immunity, growth, and development. New Phytol. 2023, 239, 1112–1126. [Google Scholar] [CrossRef]
- Petersen, M.; Brodersen, P.; Naested, H.; Andreasson, E.; Lindhart, U.; Johansen, B.; Nielsen, H.B.; Lacy, M.; Austin, M.J.; Parker, J.E.; et al. Arabidopsis map kinase 4 negatively regulates systemic acquired resistance. Cell 2000, 103, 1111–1120. [Google Scholar] [CrossRef]
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol.Cell 2004, 15, 141–152. [Google Scholar] [CrossRef]
- Kosetsu, K.; Matsunaga, S.; Nakagami, H.; Colcombet, J.; Sasabe, M.; Soyano, T.; Takahashi, Y.; Hirt, H.; Machida, Y. The MAP kinase MPK4 is required for cytokinesis in Arabidopsis thaliana. Plant Cell 2010, 22, 3778–3790. [Google Scholar] [CrossRef]
- Puerto-Galán, L.; Pérez-Ruiz, J.M.; Guinea, M.; Cejudo, F.J. The contribution of NADPH thioredoxin reductase C (NTRC) and sulfiredoxin to 2-Cys peroxiredoxin overoxidation in Arabidopsis thaliana chloroplasts. J. Exp. Bot. 2015, 66, 2957–2966. [Google Scholar] [CrossRef]
- Lee, E.S.; Kang, C.H.; Park, J.H.; Lee, S.Y. Physiological Significance of Plant Peroxiredoxins and the Structure-Related and Multifunctional Biochemistry of Peroxiredoxin 1. Antioxidants & Redox Signaling 2018, 28, 625–639. [Google Scholar] [CrossRef]
- Liu, X.P.; Liu, X.Y.; Zhang, J.; Xia, Z.L.; Liu, X.; Qin, H.J.; Wang, D.W. Molecular and functional characterization of sulfiredoxin homologs from higher plants. Cell Res. 2006, 16, 287–296. [Google Scholar] [CrossRef]
- Iglesias-Baena, I.; Barranco-Medina, S.; Lazaro-Payo, A.; Lopez-Jaramillo, F.J.; Sevilla, F.; Lazaro, J.J. Characterization of plant sulfiredoxin and role of sulphinic form of 2-Cys peroxiredoxin. J. Exp. Bot. 2010, 61, 1509–1521. [Google Scholar] [CrossRef]
- Barranco-Medina, S.; Lazaro, J.J.; Dietz, K.J. The oligomeric conformation of peroxiredoxins links redox state to function. FEBS Lett. 2009, 583, 1809–1816. [Google Scholar] [CrossRef]
- Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two Enzymes in One: Two Yeast Peroxiredoxins Display Oxidative Stress-Dependent Switching from a Peroxidase to a Molecular Chaperone Function. Cell 2004, 117, 625–635. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jang, H.H.; Lee, J.R.; Sung, N.R.; Lee, H.B.; Lee, D.H.; Park, D.-J.; Kang, C.H.; Chung, W.S.; Lim, C.O.; et al. Oligomerization and chaperone activity of a plant 2-Cys peroxiredoxin in response to oxidative stress. Plant Sci. 2009, 177, 227–232. [Google Scholar] [CrossRef]
- Muthuramalingam, M.; Seidel, T.; Laxa, M.; Nunes de Miranda, S.M.; Gärtner, F.; Ströher, E.; Kandlbinder, A.; Dietz, K.-J. Multiple Redox and Non-Redox Interactions Define 2-Cys Peroxiredoxin as a Regulatory Hub in the Chloroplast. Mol. Plant 2009, 2, 1273–1288. [Google Scholar] [CrossRef]
- Hassan, H.M. Exacerbation of superoxide radical formation by Paraquat. In Methods Enzymol; Academic Press, 1984; Volume 105, pp. 523–532. [Google Scholar]
- Cerveau, D.; Ouahrani, D.; Marok, M.A.; Blanchard, L.; Rey, P. Physiological relevance of plant 2-Cys peroxiredoxin overoxidation level and oligomerization status. Plant, Cell & Environment 2016, 39, 103–119. [Google Scholar] [CrossRef]
- Aran, M.; Caporaletti, D.; Senn, A.M.; Tellez de Iñon, M.T.; Girotti, M.R.; Llera, A.S.; Wolosiuk, R.A. ATP-dependent modulation and autophosphorylation of rapeseed 2-Cys peroxiredoxin. The FEBS Journal 2008, 275, 1450–1463. [Google Scholar] [CrossRef]
- Weits, D.A.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Hubberten, H.M.; Riegler, H.; Hoefgen, R.; Perata, P.; van Dongen, J.T.; Licausi, F. Plant cysteine oxidases control the oxygen-dependent branch of the N-end-rule pathway. Nature Communications 2014, 5. [Google Scholar] [CrossRef]
- Weits, D.A.; Zhou, L.; Giuntoli, B.; Carbonare, L.D.; Iacopino, S.; Piccinini, L.; Lombardi, L.; Shukla, V.; Bui, L.T.; Novi, G.; et al. Acquisition of hypoxia inducibility by oxygen sensing N-terminal cysteine oxidase in spermatophytes. Plant, Cell & Environment 2023, 46, 322–338. [Google Scholar] [CrossRef]
- White, M.D.; Carbonare, L.D.; Puerta, M.L.; Lacopino, S.; Edwards, M.; Dunne, K.; Pires, E.; Levy, C.; McDonough, M.A.; Licausi, F.; et al. Structures of Arabidopsis thaliana oxygen-sensing plant cysteine oxidases 4 and 5 enable targeted manipulation of their activity. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 23140–23147. [Google Scholar] [CrossRef]
- Chen, Z.; Guo, Q.; Wu, G.; Wen, J.; Liao, S.; Xu, C. Molecular basis for cysteine oxidation by plant cysteine oxidases from Arabidopsis thaliana. J. Struct. Biol. 2021, 213, 107663. [Google Scholar] [CrossRef]
- Sriram, S.M.; Kim, B.Y.; Kwon, Y.T. The N-end rule pathway: emerging functions and molecular principles of substrate recognition. Nature Reviews Molecular Cell Biology 2011, 12, 735–747. [Google Scholar] [CrossRef]
- Hu, R.-G.; Sheng, J.; Qi, X.; Xu, Z.; Takahashi, T.T.; Varshavsky, A. The N-end rule pathway as a nitric oxide sensor controlling the levels of multiple regulators. Nature 2005, 437, 981–986. [Google Scholar] [CrossRef]
- Dissmeyer, N. Conditional Protein Function via N-Degron Pathway-Mediated Proteostasis in Stress Physiology. In Annual Review of Plant Biology, Vol 70; Merchant, S.S., Ed.; Annual Review of Plant Biology; 2019; Volume 70, pp. 83–117. [Google Scholar]
- Giuntoli, B.; Perata, P. Group VII Ethylene Response Factors in Arabidopsis: Regulation and Physiological Roles. Plant Physiol. 2017, 176, 1143–1155. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.; Perata, P.; van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–U177. [Google Scholar] [CrossRef]
- White, M.D.; Kamps, J.J.A.G.; East, S.; Taylor Kearney, L.J.; Flashman, E. The plant cysteine oxidases from Arabidopsis thaliana are kinetically tailored to act as oxygen sensors. J. Biol. Chem. 2018, 293, 11786–11795. [Google Scholar] [CrossRef]
- White, M.D.; Klecker, M.; Hopkinson, R.J.; Weits, D.A.; Mueller, C.; Naumann, C.; O’Neill, R.; Wickens, J.; Yang, J.; Brooks-Bartlett, J.C.; et al. Plant cysteine oxidases are dioxygenases that directly enable arginyl transferase-catalysed arginylation of N-end rule targets. Nature Communications 2017, 8, 14690. [Google Scholar] [CrossRef]
- Loreti, E.; Perata, P. The Many Facets of Hypoxia in Plants. Plants 2020, 9, 745. [Google Scholar] [CrossRef]
- Kunkowska, A.B.; Fontana, F.; Betti, F.; Soeur, R.; Beckers, G.J.M.; Meyer, C.; De Jaeger, G.; Weits, D.A.; Loreti, E.; Perata, P. Target of rapamycin signaling couples energy to oxygen sensing to modulate hypoxic gene expression in <i>Arabidopsis</i>. Proceedings of the National Academy of Sciences 2023, 120, e2212474120. [Google Scholar] [CrossRef]
- Weits, D.A.; Kunkowska, A.B.; Kamps, N.C.W.; Portz, K.M.S.; Packbier, N.K.; Nemec Venza, Z.; Gaillochet, C.; Lohmann, J.U.; Pedersen, O.; van Dongen, J.T.; et al. An apical hypoxic niche sets the pace of shoot meristem activity. Nature 2019, 569, 714–717. [Google Scholar] [CrossRef]
- Weits, D.A.; van Dongen, J.T.; Licausi, F. Molecular oxygen as a signaling component in plant development. New Phytologist 2021, 229, 24–35. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Tedds, H.M.; Labandera, A.M.; Bailey, M.; White, M.D.; Hartman, S.; Sprigg, C.; Mogg, S.L.; Osborne, R.; Dambire, C.; et al. Oxygen-dependent proteolysis regulates the stability of angiosperm polycomb repressive complex 2 subunit VERNALIZATION 2. Nature Communications 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Labandera, A.-M.; Tedds, H.M.; Bailey, M.; Sprigg, C.; Etherington, R.D.; Akintewe, O.; Kalleechurn, G.; Holdsworth, M.J.; Gibbs, D.J. The PRT6 N-degron pathway restricts VERNALIZATION 2 to endogenous hypoxic niches to modulate plant development. New Phytol. 2021, 229, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P. Kinetics and mechanisms of thiol-disulfide exchange covering direct substitution and thiol oxidation-mediated pathways. Antioxid. Redox Signal. 2013, 18, 1623–1641. [Google Scholar] [CrossRef]
- Kolšek, K.; Aponte-Santamaría, C.; Gräter, F. Accessibility explains preferred thiol-disulfide isomerization in a protein domain. Sci. Rep. 2017, 7, 9858. [Google Scholar] [CrossRef]
- Sun, M.A.; Wang, Y.; Zhang, Q.; Xia, Y.; Ge, W.; Guo, D. Prediction of reversible disulfide based on features from local structural signatures. BMC Genomics 2017, 18, 279. [Google Scholar] [CrossRef]
- Barinova, K.V.; Serebryakova, M.V.; Eldarov, M.A.; Kulikova, A.A.; Mitkevich, V.A.; Muronetz, V.I.; Schmalhausen, E.V. S-glutathionylation of human glyceraldehyde-3-phosphate dehydrogenase and possible role of Cys152-Cys156 disulfide bridge in the active site of the protein. Biochimica et Biophysica Acta (BBA) - General Subjects 2020, 1864, 129560. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.-c. Oxidative protein folding fidelity and redoxtasis in the endoplasmic reticulum. Trends Biochem. Sci. 2023, 48, 40–52. [Google Scholar] [CrossRef]
- Meyer, A.J.; Riemer, J.; Rouhier, N. Oxidative protein folding: state-of-the-art and current avenues of research in plants. New Phytol. 2019, 221, 1230–1246. [Google Scholar] [CrossRef]
- Fan, F.; Zhang, Q.; Zhang, Y.; Huang, G.; Liang, X.; Wang, C.-c.; Wang, L.; Lu, D. Two protein disulfide isomerase subgroups work synergistically in catalyzing oxidative protein folding. Plant Physiol. 2021, 188, 241–254. [Google Scholar] [CrossRef]
- Wilkinson, B.; Gilbert, H.F. Protein disulfide isomerase. Biochim. Biophys. Acta 2004, 1699, 35–44. [Google Scholar] [CrossRef]
- Couturier, J.; Chibani, K.; Jacquot, J.P.; Rouhier, N. Cysteine-based redox regulation and signaling in plants. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Wolosiuk, R.A.; Schürmann, P. Thioredoxin and enzyme regulation. Trends Biochem. Sci. 1979, 4, 93–96. [Google Scholar] [CrossRef]
- Chibani, K.; Pucker, B.; Dietz, K.-J.; Cavanagh, A. Genome-wide analysis and transcriptional regulation of the typical and atypical thioredoxins in Arabidopsis thaliana. FEBS Lett. 2021, 595, 2715–2730. [Google Scholar] [CrossRef]
- Mata-Pérez, C.; Spoel, S.H. Thioredoxin-mediated redox signalling in plant immunity. Plant Sci. 2019, 279, 27–33. [Google Scholar] [CrossRef]
- Balsera, M.; Buchanan, B.B. Evolution of the thioredoxin system as a step enabling adaptation to oxidative stress. Free Radical Biology and Medicine 2019, 140, 28–35. [Google Scholar] [CrossRef]
- Cejudo, F.J.; González, M.-C.; Pérez-Ruiz, J.M. Redox regulation of chloroplast metabolism. Plant Physiol. 2020, 186, 9–21. [Google Scholar] [CrossRef]
- da Fonseca-Pereira, P.; Souza, P.V.L.; Fernie, A.R.; Timm, S.; Daloso, D.M.; Araújo, W.L. Thioredoxin-mediated regulation of (photo)respiration and central metabolism. J. Exp. Bot. 2021, 72, 5987–6002. [Google Scholar] [CrossRef]
- Geissel, F.; Lang, L.; Husemann, B.; Morgan, B.; Deponte, M. Deciphering the mechanism of glutaredoxin-catalyzed roGFP2 redox sensing reveals a ternary complex with glutathione for protein disulfide reduction. Nature Communications 2024, 15, 1733. [Google Scholar] [CrossRef]
- Lemaire, S.D.; Michelet, L.; Zaffagnini, M.; Massot, V.; Issakidis-Bourguet, E. Thioredoxins in chloroplasts. Curr. Genet. 2007, 51, 343–365. [Google Scholar] [CrossRef]
- Dziubek, D.; Poeker, L.; Siemitkowska, B.; Graf, A.; Marino, G.; Alseekh, S.; Arrivault, S.; Fernie, A.R.; Armbruster, U.; Geigenberger, P. NTRC and thioredoxins m1/m2 underpin the light acclimation of plants on proteome and metabolome levels. Plant Physiol. 2023, 194, 982–1005. [Google Scholar] [CrossRef]
- Wolosiuk, R.A.; Buchanan, B.B. Regulation of chloroplast phosphoribulokinase by the ferredoxin/thioredoxin system. Arch. Biochem. Biophys. 1978, 189, 97–101. [Google Scholar] [CrossRef]
- Brandes, H.K.; Larimer, F.W.; Hartman, F.C. The molecular pathway for the regulation of phosphoribulokinase by thioredoxin f. J. Biol. Chem. 1996, 271, 3333–3335. [Google Scholar] [CrossRef]
- Sparla, F.; Pupillo, P.; Trost, P. The C-terminal Extension of Glyceraldehyde-3-phosphate Dehydrogenase Subunit B Acts as an Autoinhibitory Domain Regulated by Thioredoxins and Nicotinamide Adenine Dinucleotide*. J. Biol. Chem. 2002, 277, 44946–44952. [Google Scholar] [CrossRef]
- Chiadmi, M.; Navaza, A.; Miginiac-Maslow, M.; Jacquot, J.P.; Cherfils, J. Redox signalling in the chloroplast: structure of oxidized pea fructose-1,6-bisphosphate phosphatase. The EMBO Journal 1999, 18, 6809–6815. [Google Scholar] [CrossRef]
- Dunford, R.P.; Durrant, M.C.; Catley, M.A.; Dyer, T.A. Location of the redox-active cysteines in chloroplast sedoheptulose-1,7-bisphosphatase indicates that its allosteric regulation is similar but not identical to that of fructose-1,6-bisphosphatase. Photosynthesis Res. 1998, 58, 221–230. [Google Scholar] [CrossRef]
- Wedel, N.; Soll, J.; Paap, B.K. CP12 provides a new mode of light regulation of Calvin cycle activity in higher plants. Proc.Natl.Acad.Sci.U.S.A 1997, 94, 10479–10484. [Google Scholar] [CrossRef]
- Marri, L.; Trost, P.; Pupillo, P.; Sparla, F. Reconstitution and Properties of the Recombinant Glyceraldehyde-3-Phosphate Dehydrogenase/CP12/Phosphoribulokinase Supramolecular Complex of Arabidopsis. Plant Physiol. 2005, 139, 1433–1443. [Google Scholar] [CrossRef]
- Wolosiuk, R.A.; Buchanan, B.B. Thioredoxin and glutathione regulate photosynthesis in chloroplasts. Nature 1977, 266, 565–567. [Google Scholar] [CrossRef]
- Yoshida, K.; Hisabori, T. Determining the Rate-Limiting Step for Light-Responsive Redox Regulation in Chloroplasts. Antioxidants 2018, 7, 153. [Google Scholar] [CrossRef]
- Née, G.; Aumont-Nicaise, M.; Zaffagnini, M.; Nessler, S.; Valerio-Lepiniec, M.; Issakidis-Bourguet, E. Redox regulation of chloroplastic G6PDH activity by thioredoxin occurs through structural changes modifying substrate accessibility and cofactor binding. Biochem. J. 2013, 457, 117–125. [Google Scholar] [CrossRef]
- Moon, J.C.; Jang, H.H.; Chae, H.B.; Lee, J.R.; Lee, S.Y.; Jung, Y.J.; Shin, M.R.; Lim, H.S.; Chung, W.S.; Yun, D.-J.; et al. The C-type Arabidopsis thioredoxin reductase ANTR-C acts as an electron donor to 2-Cys peroxiredoxins in chloroplasts. Biochemical and Biophysical Research Communications 2006, 348, 478–484. [Google Scholar] [CrossRef]
- Pérez-Ruiz, J.M.; Naranjo, B.; Ojeda, V.; Guinea, M.; Cejudo, F.J. NTRC-dependent redox balance of 2-Cys peroxiredoxins is needed for optimal function of the photosynthetic apparatus. Proceedings of the National Academy of Sciences 2017, 114, 12069–12074. [Google Scholar] [CrossRef]
- Lampl, N.; Lev, R.; Nissan, I.; Gilad, G.; Hipsch, M.; Rosenwasser, S. Systematic monitoring of 2-Cys peroxiredoxin-derived redox signals unveiled its role in attenuating carbon assimilation rate. Proceedings of the National Academy of Sciences 2022, 119, e2119719119. [Google Scholar] [CrossRef]
- Ojeda, V.; Pérez-Ruiz, J.M.; Cejudo, F.J. 2-Cys Peroxiredoxins Participate in the Oxidation of Chloroplast Enzymes in the Dark. Mol. Plant 2018, 11, 1377–1388. [Google Scholar] [CrossRef]
- Gromes, R.; Hothorn, M.; Lenherr, E.D.; Rybin, V.; Scheffzek, K.; Rausch, T. The redox switch of gamma-glutamylcysteine ligase via a reversible monomer-dimer transition is a mechanism unique to plants. Plant J. 2008, 54, 1063–1075. [Google Scholar] [CrossRef]
- Marty, L.; Siala, W.; Schwarzländer, M.; Fricker, M.D.; Wirtz, M.; Sweetlove, L.J.; Meyer, Y.; Meyer, A.J.; Reichheld, J.-P.; Hell, R. The NADPH-dependent thioredoxin system constitutes a functional backup for cytosolic glutathione reductase in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 9109–9114. [Google Scholar] [CrossRef]
- Marty, L.; Bausewein, D.; Muller, C.; Bangash, S.A.K.; Moseler, A.; Schwarzlander, M.; Muller-Schussele, S.J.; Zechmann, B.; Riondet, C.; Balk, J.; et al. Arabidopsis glutathione reductase 2 is indispensable in plastids, while mitochondrial glutathione is safeguarded by additional reduction and transport systems. New Phytol. 2019, 224, 1569–1584. [Google Scholar] [CrossRef]
- Paulsen, C.E.; Carroll, K.S. Cysteine-Mediated Redox Signaling: Chemistry, Biology, and Tools for Discovery. Chem. Rev. 2013, 113, 4633–4679. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Colombo, G.; Giustarini, D.; Milzani, A. Protein S-glutathionylation: a regulatory device from bacteria to humans. Trends Biochem.Sci. 2009, 34, 85–96. [Google Scholar] [CrossRef]
- Giustarini, D.; Milzani, A.; Aldini, G.; Carini, M.; Rossi, R.; Dalle-Donne, I. S-nitrosation versus S-glutathionylation of protein sulfhydryl groups by S-nitrosoglutathione. Antioxidants & redox signaling 2005, 7, 930–939. [Google Scholar]
- Ercolani, L.; Scirè, A.; Galeazzi, R.; Massaccesi, L.; Cianfruglia, L.; Amici, A.; Piva, F.; Urbanelli, L.; Emiliani, C.; Principato, G.; et al. A possible S-glutathionylation of specific proteins by glyoxalase II: An in vitro and in silico study. Cell Biochemistry and Function 2016, 34, 620–627. [Google Scholar] [CrossRef]
- Townsend, D.M.; Manevich, Y.; He, L.; Hutchens, S.; Pazoles, C.J.; Tew, K.D. Novel role for glutathione S-transferase Pi regulator of protein s-glutathionylation following oxidative and nitrosative stress. J. Biol. Chem. 2009, 284, 436–445. [Google Scholar] [CrossRef]
- Mo, Z.; Huang, Y.; Pu, T.; Duan, L.; Pi, K.; Luo, J.; Long, B.; Lu, A.; Liu, R. Genome-wide identification and characterization of Glutathione S-Transferases (GSTs) and their expression profile under abiotic stresses in tobacco (Nicotiana tabacum L.). BMC Genomics 2023, 24, 341. [Google Scholar] [CrossRef]
- Bender, Kyle W. ; Wang, X.; Cheng, George B.; Kim, Hyoung S.; Zielinski, Raymond E.; Huber, Steven C. Glutaredoxin AtGRXC2 catalyses inhibitory glutathionylation of Arabidopsis BRI1-associated receptor-like kinase 1 (BAK1) in vitro. Biochem. J. 2015, 467, 399–413. [Google Scholar] [CrossRef]
- Mao, J.; Li, J. Regulation of Three Key Kinases of Brassinosteroid Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 4340. [Google Scholar] [CrossRef]
- van der Linde, K.; Gutsche, N.; Leffers, H.-M.; Lindermayr, C.; Müller, B.; Holtgrefe, S.; Scheibe, R. Regulation of plant cytosolic aldolase functions by redox modifications. Plant Physiology and Biochemistry 2011, 49, 946–957. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Bedhomme, M.; Marchand, C.H.; Couturier, J.; Gao, X.H.; Rouhier, N.; Trost, P.; Lemaire, S.D. Glutaredoxin S12: Unique Properties for Redox Signaling. Antioxidants & Redox Signaling 2012, 16, 17–32. [Google Scholar] [CrossRef]
- Peltoniemi, M.J.; Karala, A.-R.; Jurvansuu, J.K.; Kinnula, V.L.; Ruddock, L.W. Insights into deglutathionylation reactions: Different intermediates in the glutaredoxin and protein disulfide isomerase catalyzed reactions are defined by the γ-linkage present in glutathione. J. Biol. Chem. 2006, 281, 33107–33114. [Google Scholar] [CrossRef]
- Randi, E.B.; Zuhra, K.; Pecze, L.; Panagaki, T.; Szabo, C. Physiological concentrations of cyanide stimulate mitochondrial Complex IV and enhance cellular bioenergetics. Proc. Natl. Acad. Sci. U. S. A. 2021, 118. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Bedhomme, M.; Marchand, C.H.; Morisse, S.; Trost, P.; Lemaire, S.D. Redox Regulation in Photosynthetic Organisms: Focus on Glutathionylation. Antioxidants & Redox Signaling 2012, 16, 567–586. [Google Scholar] [CrossRef]
- Gurrieri, L.; Distefano, L.; Pirone, C.; Horrer, D.; Seung, D.; Zaffagnini, M.; Rouhier, N.; Trost, P.; Santelia, D.; Sparla, F. The thioredoxin-regulated α-amylase 3 of Arabidopsis thaliana is a target of S-glutathionylation. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Rouhier, N.; Couturier, J.; Jacquot, J.P. Genome-wide analysis of plant glutaredoxin systems. J. Exp. Bot. 2006, 57, 1685–1696. [Google Scholar] [CrossRef]
- He, X.Y.; Chen, W.Y.; Sun, X.C.; Gao, Y.; He, Y.R.; Xu, X.T.; Su, C.J.; Lv, Y.F.; Ren, B.Y.; Yin, H.Y.; et al. Genome-Wide Identification and Characterization of Glutaredoxin Family Genes in Common Wheat. Agronomy-Basel 2023, 13. [Google Scholar] [CrossRef]
- Mondal, S.; Singh, S.P. New insights on thioredoxins (Trxs) and glutaredoxins (Grxs) by in silico amino acid sequence, phylogenetic and comparative structural analyses in organisms of three domains of life. Heliyon 2022, 8, e10776. [Google Scholar] [CrossRef]
- Bodnar, Y.; Gellert, M.; Hossain, F.M.; Lillig, C.H. Breakdown of Arabidopsis thaliana thioredoxins and glutaredoxins based on electrostatic similarity-Leads to common and unique interaction partners and functions. PLoS One 2023, 18. [Google Scholar] [CrossRef]
- Giustarini, D.; Rossi, R.; Milzani, A.; Colombo, R.; Dalle-Donne, I. S-Glutathionylation: from redox regulation of protein functions to human diseases. Journal of Cellular and Molecular Medicine 2004, 8, 201–212. [Google Scholar] [CrossRef]
- Dixon, D.P.; Skipsey, M.; Grundy, N.M.; Edwards, R. Stress-Induced Protein S-glutathionylation in Arabidopsis. Plant Physiol. 2005, 138, 2233–2244. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Vanacker, H.; Le Marechal, P.; Marchand, C.; Schroda, M.; Lemaire, S.D.; Decottignies, P. In vivo targets of S-thiolation in Chlamydomonas reinhardtii. J. Biol. Chem. 2008, 283, 21571–21578. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Bedhomme, M.; Groni, H.; Marchand, C.H.; Puppo, C.; Gontero, B.; Cassier-Chauvat, C.; Decottignies, P.; Lemaire, S.D. Glutathionylation in the photosynthetic model organism Chlamydomonas reinhardtii: A proteomic survey. Mol. Cell. Proteomics 2012, 11. [Google Scholar] [CrossRef]
- Gietler, M.; Nykiel, M.; Orzechowski, S.; Fettke, J.; Zagdańska, B. Proteomic analysis of S-nitrosylated and S-glutathionylated proteins in wheat seedlings with different dehydration tolerances. Plant Physiology and Biochemistry 2016, 108, 507–518. [Google Scholar] [CrossRef]
- Müller-Schüssele, S.J.; Bohle, F.; Rossi, J.; Trost, P.; Meyer, A.J.; Zaffagnini, M. Plasticity in plastid redox networks: evolution of glutathione-dependent redox cascades and glutathionylation sites. BMC Plant Biol. 2021, 21, 322. [Google Scholar] [CrossRef]
- Kitajima, S.; Kurioka, M.; Yoshimoto, T.; Shindo, M.; Kanaori, K.; Tajima, K.; Oda, K. A cysteine residue near the propionate side chain of heme is the radical site in ascorbate peroxidase. FEBS J. 2008, 275, 470–480. [Google Scholar] [CrossRef]
- Seung, D.; Thalmann, M.; Sparla, F.; Abou Hachem, M.; Lee, S.K.; Issakidis-Bourguet, E.; Svensson, B.; Zeeman, S.C.; Santelia, D. Arabidopsis thaliana AMY3 is a unique redox-regulated chloroplastic α-amylase. J. Biol. Chem. 2013, 288, 33620–33633. [Google Scholar] [CrossRef] [PubMed]
- Niazi, A.K.; Bariat, L.; Riondet, C.; Carapito, C.; Mhamdi, A.; Noctor, G.; Reichheld, J.-P. Cytosolic Isocitrate Dehydrogenase from Arabidopsis thaliana Is Regulated by Glutathionylation. Antioxidants (Basel) 2019, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Hodges, M. Enzyme redundancy and the importance of 2-oxoglutarate in plant ammonium assimilation. J. Exp. Bot. 2002, 53, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Rivoal, J.; Hanson, A.D. Choline-O-sulfate biosynthesis in plants: Identification and partial characterization of a salinity-inducible choline sulfotransferase from species of Limonium (Plumbaginaceae). Plant-Physiol. 1994, 106, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Varin, L.; Marsolais, F.; Richard, M.; Rouleau, M. Sulfation and sulfotransferases 6: Biochemistry and molecular biology of plant sulfotransferases. FASEB J. 1997, 11, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Jain, J.C.; Grootwassink, J.W.D.; Kolenovsky, A.D.; Underhill, E.W. Purification and properties of 3'-phosphoadenosine-5'-phosphosulfate desulfoglucosinolate sulfotransferase from Brassica juncea cell cultures. Phytochemistry 1990, 29, 1425–1428. [Google Scholar] [CrossRef]
- Yang, L.M.; Fernandez, M.D.; Lamppa, G.K. ACYL CARRIER PROTEIN (ACP) IMPORT INTO CHLOROPLASTS - COVALENT MODIFICATION BY A STROMAL HOLOACP SYNTHASE IS STIMULATED BY EXOGENOUSLY ADDED COA AND INHIBITED BY ADENOSINE 3',5'-BISPHOSPHATE. Eur. J. Biochem. 1994, 224, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.X.; Phua, S.Y.; Crisp, P.; McQuinn, R.; Pogson, B.J. Learning the Languages of the Chloroplast: Retrograde Signaling and Beyond. Annu. Rev. Plant Biol. 2016, 67, 25–53. [Google Scholar] [CrossRef]
- Estavillo, G.M.; Crisp, P.A.; Pornsiriwong, W.; Wirtz, M.; Collinge, D.; Carrie, C.; Giraud, E.; Whelan, J.; David, P.; Javot, H.; et al. Evidence for a SAL1-PAP chloroplast retrograde pathway that functions in drought and high light signaling in Arabidopsis. Plant Cell 2011, 23, 3992–4012. [Google Scholar] [CrossRef]
- Chan, K.X.; Phua, S.Y.; Van Breusegem, F. Secondary sulfur metabolism in cellular signalling and oxidative stress responses. J. Exp. Bot. 2019, 70, 4237–4250. [Google Scholar] [CrossRef]
- Chan, K.X.; Mabbitt, P.D.; Phua, S.Y.; Mueller, J.W.; Nisar, N.; Gigolashvili, T.; Stroeher, E.; Grassl, J.; Arlt, W.; Estavillo, G.M.; et al. Sensing and signaling of oxidative stress in chloroplasts by inactivation of the SAL1 phosphoadenosine phosphatase. Proc. Natl. Acad. Sci. U. S. A. 2016, 113, E4567–E4576. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gao, Y.; Yang, A. Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef]
- Lisjak, M.; Teklic, T.; Wilson, I.D.; Whiteman, M.; Hancock, J.T. Hydrogen sulfide: environmental factor or signalling molecule? Plant, Cell & Environment 2013, 36, 1607–1616. [Google Scholar] [CrossRef]
- Ausma, T.; De Kok, L.J. Atmospheric H2S: Impact on Plant Functioning. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; González-Gordo, S.; Cañas, A.; Palma, J.M. Nitric oxide and hydrogen sulfide in plants: which comes first? J. Exp. Bot. 2019, 70, 4391–4404. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, M.; Zhou, H.; Zhao, D.; Gotor, C.; Romero, L.C.; Shen, J.; Ge, Z.; Zhang, Z.; Shen, W.; et al. Hydrogen sulfide, a signaling molecule in plant stress responses. J. Integr. Plant Biol. 2021, 63, 146–160. [Google Scholar] [CrossRef]
- González-Gordo, S.; Palma, J.M.; Corpas, F.J. Appraisal of H2S metabolism in Arabidopsis thaliana: In silico analysis at the subcellular level. Plant Physiology and Biochemistry 2020, 155, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xie, Y. Hydrogen Sulfide Signaling in Plants. Antioxidants & Redox Signaling 2023, 39, 40–58. [Google Scholar] [CrossRef]
- Moseler, A.; Dhalleine, T.; Rouhier, N.; Couturier, J. Arabidopsis thaliana 3-mercaptopyruvate sulfurtransferases interact with and are protected by reducing systems. J. Biol. Chem. 2021, 296, 100429. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Redox biochemistry of hydrogen sulfide. J. Biol. Chem. 2010, 285, 21903–21907. [Google Scholar] [CrossRef]
- Papanatsiou, M.; Scuffi, D.; Blatt, M.R.; García-Mata, C. Hydrogen Sulfide Regulates Inward-Rectifying K+ Channels in Conjunction with Stomatal Closure. Plant Physiol. 2015, 168, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Aroca, Á.; Serna, A.; Gotor, C.; Romero, L.C. S-Sulfhydration: A Cysteine Posttranslational Modification in Plant Systems. Plant Physiol. 2015, 168, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhang, J.; Zhou, H.; Zhao, D.; Duan, T.; Wang, S.; Yuan, X.; Xie, Y. Hydrogen Sulfide-Linked Persulfidation Maintains Protein Stability of ABSCISIC ACID-INSENSITIVE 4 and Delays Seed Germination. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Yruela, I.; Gotor, C.; Bassham, D.C. Persulfidation of ATG18a regulates autophagy under ER stress in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Dóka, É.; Ida, T.; Dagnell, M.; Abiko, Y.; Luong, N.C.; Balog, N.; Takata, T.; Espinosa, B.; Nishimura, A.; Cheng, Q.; et al. Control of protein function through oxidation and reduction of persulfidated states. Science Advances 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Macinkovic, I.; Devarie-Baez, N.O.; Pan, J.; Park, C.-M.; Carroll, K.S.; Filipovic, M.R.; Xian, M. Detection of Protein S-Sulfhydration by a Tag-Switch Technique. Angewandte Chemie International Edition 2014, 53, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Flores, A.; Romero, L.C.; Gotor, C. Label-Free Quantitative Proteomic Analysis of Nitrogen Starvation in Arabidopsis Root Reveals New Aspects of H2S Signaling by Protein Persulfidation. Antioxidants 2021, 10, 508. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Jurado-Flores, A.; Filipovic, M.R.; Gotor, C.; Romero, L.C. Detection of protein persulfidation in plants by the dimedone switch method. In Methods Enzymol.; Jez, J., Ed.; Academic Press, 2022; Volume 676, pp. 385–402. [Google Scholar]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Nitric oxide and hydrogen sulfide modulate the NADPH-generating enzymatic system in higher plants. J. Exp. Bot. 2020, 72, 830–847. [Google Scholar] [CrossRef]
- Jia, H.; Chen, S.; Liu, D.; Liesche, J.; Shi, C.; Wang, J.; Ren, M.; Wang, X.; Yang, J.; Shi, W.; et al. Ethylene-Induced Hydrogen Sulfide Negatively Regulates Ethylene Biosynthesis by Persulfidation of ACO in Tomato Under Osmotic Stress. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Wang, X.; Shi, C.; Hu, Y.; Ma, Y.; Yi, Y.; Jia, H.; Li, F.; Sun, H.; Li, T.; Wang, X.; et al. Persulfidation maintains cytosolic G6PDs activity through changing tetrameric structure and competing cysteine sulfur oxidation under salt stress in Arabidopsis and tomato. New Phytol. 2023, 240, 626–643. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Schneider, M.; Scheibe, R.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Regulates the Cytosolic/Nuclear Partitioning of Glyceraldehyde-3-Phosphate Dehydrogenase by Enhancing its Nuclear Localization. Plant Cell Physiol. 2017, 58, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.; Knuesting, J.; Berndt, C.; Morgan, B.; Scheibe, R. Cytosolic thiol switches regulating basic cellular functions: GAPDH as an information hub? Biol. Chem. 2015, 396, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, X.; Feng, J.; Zhu, S. Biological Functions of Hydrogen Sulfide in Plants. Int. J. Mol. Sci. 2022, 23, 15107. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jia, H.; Wang, X.; Shi, C.; Wang, X.; Ma, P.; Wang, J.; Ren, M.; Li, J. Hydrogen Sulfide Positively Regulates Abscisic Acid Signaling through Persulfidation of SnRK2.6 in Guard Cells. Mol. Plant 2020, 13, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, J.; Zhou, M.; Zhou, H.; Cui, B.; Gotor, C.; Romero, L.C.; Fu, L.; Yang, J.; Foyer, C.H.; et al. Persulfidation-based Modification of Cysteine Desulfhydrase and the NADPH Oxidase RBOHD Controls Guard Cell Abscisic Acid Signaling. Plant Cell 2020, 32, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Flores, A.; Aroca, A.; Romero, L.C.; Gotor, C. Sulfide promotes tolerance to drought through protein persulfidation in Arabidopsis. J. Exp. Bot. 2023, 74, 4654–4669. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.-P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.-K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.M.; Mori, I.C.; Pei, Z.-M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. The EMBO Journal 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Sato, A.; Sato, Y.; Fukao, Y.; Fujiwara, M.; Umezawa, T.; Shinozaki, K.; Hibi, T.; Taniguchi, M.; Miyake, H.; Goto, Derek B. ; et al. Threonine at position 306 of the KAT1 potassium channel is essential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6 protein kinase. Biochem. J. 2009, 424, 439–448. [Google Scholar] [CrossRef]
- Wang, P.; Du, Y.; Hou, Y.-J.; Zhao, Y.; Hsu, C.-C.; Yuan, F.; Zhu, X.; Tao, W.A.; Song, C.-P.; Zhu, J.-K. Nitric oxide negatively regulates abscisic acid signaling in guard cells by S-nitrosylation of OST1. Proceedings of the National Academy of Sciences 2015, 112, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Singh, P.; Tripathi, D.K.; Corpas, F.J.; Singh, V.P. Nitric oxide and hydrogen sulfide: an indispensable combination for plant functioning. Trends Plant Sci. 2021, 26, 1270–1285. [Google Scholar] [CrossRef] [PubMed]
- Jaszczak, E.; Polkowska, Ż.; Narkowicz, S.; Namieśnik, J. Cyanides in the environment-analysis-problems and challenges. Environ. Sci. Pollut. Res. Int. 2017, 24, 15929–15948. [Google Scholar] [CrossRef] [PubMed]
- Yulvianti, M.; Zidorn, C. Chemical Diversity of Plant Cyanogenic Glycosides: An Overview of Reported Natural Products. Molecules 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Hydrogen cyanide production by soil bacteria: Biological control of pests and promotion of plant growth in sustainable agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Zuhra, K.; Szabo, C. The two faces of cyanide: an environmental toxin and a potential novel mammalian gasotransmitter. FEBS J. 2022, 289, 2481–2515. [Google Scholar] [CrossRef] [PubMed]
- Gleadow, R.M.; Moller, B.L. Cyanogenic Glycosides: Synthesis, Physiology, and Phenotypic Plasticity. In Annual Review of Plant Biology, Vol 65; Merchant, S.S., Ed.; Annual Review of Plant Biology; 2014; Volume 65, pp. 155–185. [Google Scholar]
- Dong, J.G.; Fernández-Maculet, J.C.; Yang, S.F. Purification and characterization of 1-aminocyclopropane-1-carboxylate oxidase from apple fruit. Proceedings of the National Academy of Sciences 1992, 89, 9789–9793. [Google Scholar] [CrossRef] [PubMed]
- Fatma, M.; Asgher, M.; Iqbal, N.; Rasheed, F.; Sehar, Z.; Sofo, A.; Khan, N.A. Ethylene Signaling under Stressful Environments: Analyzing Collaborative Knowledge. Plants 2022, 11, 2211. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, C.; Westphal, L.; Schmotz, C.; Prade, E.; Scheel, D.; Glawischnig, E. The Multifunctional Enzyme CYP71B15 (PHYTOALEXIN DEFICIENT3) Converts Cysteine-Indole-3-Acetonitrile to Camalexin in the Indole-3-Acetonitrile Metabolic Network of <i>Arabidopsis thaliana</i>. Plant Cell 2009, 21, 1830–1845. [Google Scholar] [CrossRef]
- Hucklesby, D.P.; Dowling, M.J.; Hewitt, E.J. Cyanide formation from glyoxylate and hydroxylamine catalysed by extracts of higher-plant leaves. Planta 1982, 156, 487–491. [Google Scholar] [CrossRef]
- Álvarez, C.; García, I.; Romero, L.C.; Gotor, C. Mitochondrial Sulfide Detoxification Requires a Functional Isoform O-Acetylserine(thiol)lyase C in Arabidopsis thaliana. Mol. Plant 2012, 5, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Zidenga, T.; Siritunga, D.; Sayre, R.T. Cyanogen Metabolism in Cassava Roots: Impact on Protein Synthesis and Root Development. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Borisov, V.B.; Forte, E. Impact of Hydrogen Sulfide on Mitochondrial and Bacterial Bioenergetics. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Fasco, M.J.; Hauer, C.R.; Stack, R.F.; O'Hehir, C.; Barr, J.R.; Eadon, G.A. Cyanide Adducts with Human Plasma Proteins: Albumin as a Potential Exposure Surrogate. Chem. Res. Toxicol. 2007, 20, 677–684. [Google Scholar] [CrossRef] [PubMed]
- García, I.; Arenas-Alfonseca, L.; Moreno, I.; Gotor, C.; Romero, L.C. HCN Regulates Cellular Processes through Posttranslational Modification of Proteins by S-cyanylation. Plant Physiol. 2018, 179, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.J.; Kharasch, N. Derivatives of Sulfenic Acids. XXXVI. The Ionic Scission of the Sulfur-Sulfur Bond. 1 Part 1. J. Am. Chem. Soc. 1960, 82, 3071–3075. [Google Scholar] [CrossRef]
- Gawron, O.; Fernando, J. Kinetics of the cyanide-cystine reaction. J. Am. Chem. Soc. 1961, 83, 2906–2908. [Google Scholar] [CrossRef]
- Catsimpoolas, N.; Wood, J.L. Specific Cleavage of Cystine Peptides by Cyanide. J. Biol. Chem. 1966, 241, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Yu, G.; Leeuwon, S.Z.; Liu, W.R. Site-Specific Conversion of Cysteine in a Protein to Dehydroalanine Using 2-Nitro-5-thiocyanatobenzoic Acid. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Arenas-Alfonseca, L.; Yamada, M.; Romero, L.C.; García, I. New Insights on the Role of ß-Cyanoalanine Synthase CAS-C1 in Root Hair Elongation through Single-Cell Proteomics. Plants 2023, 12, 4055. [Google Scholar] [CrossRef]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: an update. J. Exp. Bot. 2017, 69, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Stoimenova, M.; Igamberdiev, A.U.; Gupta, K.J.; Hill, R.D. Nitrite-driven anaerobic ATP synthesis in barley and rice root mitochondria. Planta 2007, 226, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C.; Hasinoff, B.B.; Igamberdiev, A.U.; Manac'h, N.; Rivoal, J.; Hill, R.D. Expression of a stress-induced hemoglobin affects NO levels produced by alfalfa root cultures under hypoxic stress. Plant J. 2003, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C.; Hasinoff, B.B.; Rivoal, J.; Hill, R.D. Class-1 hemoglobins, nitrate and NO levels in anoxic maize cell-suspension cultures. Planta 2004, 219, 66–72. [Google Scholar] [PubMed]
- Corpas, F.J.; González-Gordo, S.; Palma, J.M. Nitric oxide: A radical molecule with potential biotechnological applications in fruit ripening. J. Biotechnol. 2020, 324, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, I.; Lindermayr, C. Nitric oxide-based protein modification: formation and site-specificity of protein <i>S</i>-nitrosylation. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Dent, M.R.; DeMartino, A.W. Nitric oxide and thiols: Chemical biology, signalling paradigms and vascular therapeutic potential. Br. J. Pharmacol. n/a. [CrossRef]
- Suarez, S.A.; Muñoz, M.; Alvarez, L.; Venâncio, M.F.; Rocha, W.R.; Bikiel, D.E.; Marti, M.A.; Doctorovich, F. HNO Is Produced by the Reaction of NO with Thiols. J. Am. Chem. Soc. 2017, 139, 14483–14487. [Google Scholar] [CrossRef]
- Sánchez-Vicente, I.; Albertos, P.; Sanz, C.; Wybouw, B.; De Rybel, B.; Begara-Morales, J.C.; Chaki, M.; Mata-Pérez, C.; Barroso, J.B.; Lorenzo, O. Reversible S-nitrosylation of bZIP67 by peroxiredoxin IIE activity and nitro-fatty acids regulates the plant lipid profile. Cell Rep. 2024, 43, 114091. [Google Scholar] [CrossRef]
- Frungillo, L.; Skelly, M.J.; Loake, G.J.; Spoel, S.H.; Salgado, I. <i>S</i>-nitrosothiols regulate nitric oxide production and storage in plants through the nitrogen assimilation pathway. Nature Communications 2014, 5. [Google Scholar] [CrossRef]
- Jahnová, J.; Luhová, L.; Petrivalsky, M. S-Nitrosoglutathione Reductase-The Master Regulator of Protein S-Nitrosation in Plant NO Signaling. Plants-Basel 2019, 8. [Google Scholar] [CrossRef]
- Kalinina, E.; Novichkova, M. Glutathione in Protein Redox Modulation through S-Glutathionylation and S-Nitrosylation. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Sakamoto, A.; Ueda, M.; Morikawa, H. Arabidopsis glutathione-dependent formaldehyde dehydrogenase is an S-nitrosoglutathione reductase. FEBS Lett. 2002, 515, 20–24. [Google Scholar] [CrossRef]
- Xu, S.; Guerra, D.; Lee, U.; Vierling, E. S-nitrosoglutathione reductases are low-copy number, cysteine-rich proteins in plants that control multiple developmental and defense responses in Arabidopsis. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Cutrona, M.C.; Wienkoop, S.; Begara-Morales, J.C.; Sandal, N.; Orera, I.; Barroso, J.B.; Stougaard, J.; Becana, M. Altered Plant and Nodule Development and Protein <i>S</i>-Nitrosylation in <i>Lotus japonicus</i> Mutants Deficient in <i>S</i>-Nitrosoglutathione Reductases. Plant Cell Physiol. 2020, 61, 105–117. [Google Scholar] [CrossRef]
- Kovacs, I.; Holzmeister, C.; Wirtz, M.; Geerlof, A.; Frohlich, T.; Romling, G.; Kuruthukulangarakoola, G.T.; Linster, E.; Hell, R.; Arnold, G.J.; et al. ROS-mediated inhibition of S-nitrosoglutathione reductase contributes to the activation of anti-oxidative mechanisms. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Guerra, D.; Ballard, K.; Truebridge, I.; Vierling, E. S-Nitrosation of Conserved Cysteines Modulates Activity and Stability of <i>S</i>-Nitrosoglutathione Reductase (GSNOR). Biochemistry 2016, 55, 2452–2464. [Google Scholar] [CrossRef]
- Tichá, T.; Lochman, J.; Činčalová, L.; Luhová, L.; Petřivalský, M. Redox regulation of plant S-nitrosoglutathione reductase activity through post-translational modifications of cysteine residues. Biochemical and Biophysical Research Communications 2017, 494, 27–33. [Google Scholar] [CrossRef]
- Stomberski, C.T.; Anand, P.; Venetos, N.M.; Hausladen, A.; Zhou, H.L.; Premont, R.T.; Stamler, J.S. AKR1A1 is a novel mammalian <i>S</i>-nitroso-glutathione reductase. J. Biol. Chem. 2019, 294, 18285–18293. [Google Scholar] [CrossRef]
- Treffon, P.; Rossi, J.; Gabellini, G.; Trost, P.; Zaffagnini, M.; Vierling, E. Quantitative Proteome Profiling of a S-Nitrosoglutathione Reductase (GSNOR) Null Mutant Reveals a New Class of Enzymes Involved in Nitric Oxide Homeostasis in Plants. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Treffon, P.; Vierling, E. Focus on Nitric Oxide Homeostasis: Direct and Indirect Enzymatic Regulation of Protein Denitrosation Reactions in Plants. Antioxidants 2022, 11, 1411. [Google Scholar] [CrossRef]
- Kneeshaw, S.; Gelineau, S.; Tada, Y.; Loake, G.J.; Spoel, S.H. Selective Protein Denitrosylation Activity of Thioredoxin-<i>h</i>5 Modulates Plant Immunity. Mol. Cell 2014, 56, 153–162. [Google Scholar] [CrossRef]
- Sunico, C.R.; Sultan, A.; Nakamura, T.; Dolatabadi, N.; Parker, J.; Shan, B.; Han, X.M.; Yates, J.R.; Masliah, E.; Ambasudhan, R.; et al. Role of sulfiredoxin as a peroxiredoxin-2 denitrosylase in human iPSC-derived dopaminergic neurons. Proc. Natl. Acad. Sci. U. S. A. 2016, 113, E7564–E7571. [Google Scholar] [CrossRef]
- Ren, X.; Sengupta, R.; Lu, J.; Lundberg, J.O.; Holmgren, A. Characterization of mammalian glutaredoxin isoforms as S-denitrosylases. FEBS Lett. 2019, 593, 1799–1806. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Morisse, S.; Bedhomme, M.; Marchand, C.H.; Festa, M.; Rouhier, N.; Lemaire, S.D.; Trost, P. Mechanisms of Nitrosylation and Denitrosylation of Cytoplasmic Glyceraldehyde-3-phosphate Dehydrogenase from Arabidopsis thaliana*. J. Biol. Chem. 2013, 288, 22777–22789. [Google Scholar] [CrossRef]
- Zhang, T.; Ma, M.; Chen, T.; Zhang, L.; Fan, L.; Zhang, W.; Wei, B.; Li, S.; Xuan, W.; Noctor, G.; et al. Glutathione-dependent denitrosation of GSNOR1 promotes oxidative signalling downstream of H2O2. Plant, Cell & Environment 2020, 43, 1175–1191. [Google Scholar] [CrossRef]
- Jaffrey, S.R.; Erdjument-Bromage, H.; Ferris, C.D.; Tempst, P.; Snyder, S.H. Protein S-nitrosylation: a physiological signal for neuronal nitric oxide. Nat. Cell Biol. 2001, 3, 193–197. [Google Scholar] [CrossRef]
- Lindermayr, C.; Saalbach, G.; Durner, J.r. Proteomic Identification of S-Nitrosylated Proteins in Arabidopsis. Plant Physiol. 2005, 137, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Huang, X.; Chen, L.; Sun, X.; Lu, C.; Zhang, L.; Wang, Y.; Zuo, J. Site-Specific Nitrosoproteomic Identification of Endogenously S-Nitrosylated Proteins in Arabidopsis. Plant Physiol. 2015, 167, 1731–1746. [Google Scholar] [CrossRef] [PubMed]
- Paige, J.S.; Xu, G.; Stancevic, B.; Jaffrey, S.R. Nitrosothiol Reactivity Profiling Identifies S-Nitrosylated Proteins with Unexpected Stability. Chem. Biol. 2008, 15, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, R.; Stuehr, D.J. Thioredoxin-1 Regulates Cellular Heme Insertion by Controlling S-Nitrosation of Glyceraldehyde-3-phosphate Dehydrogenase*. J. Biol. Chem. 2012, 287, 16179–16186. [Google Scholar] [CrossRef] [PubMed]
- Serrato, A.J.; Romero-Puertas, M.C.; Lázaro-Payo, A.; Sahrawy, M. Regulation by S-nitrosylation of the Calvin-Benson cycle fructose-1,6-bisphosphatase in Pisum sativum. Redox Biology 2018, 14, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Kolbert, Z.; Durner, J.; Lindermayr, C.; Corpas, F.J.; Brouquisse, R.; Barroso, J.B.; Umbreen, S.; Palma, J.M.; Hancock, J.T.; et al. Regulating the regulator: nitric oxide control of post-translational modifications. New Phytol. 2020, 227, 1319–1325. [Google Scholar] [CrossRef]
- Pande, A.; Mun, B.G.; Rahim, W.; Khan, M.; Lee, D.S.; Lee, G.M.; Al Azzawi, T.N.I.; Hussain, A.; Kim, C.K.; Yun, B.W. Phytohormonal Regulation Through Protein S-Nitrosylation Under Stress. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.C.; Dua, Y.Y.; Hou, Y.J.; Zhao, Y.; Hsu, C.C.; Yuan, F.J.; Zhu, X.H.; Tao, W.A.; Song, C.P.; Zhu, J.K. Nitric oxide negatively regulates abscisic acid signaling in guard cells by S-nitrosylation of OST1. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Bowling, S.A.; Gordon, A.S.; Dong, X. Characterization of an Arabidopsis Mutant That Is Nonresponsive to Inducers of Systemic Acquired Resistance. Plant Cell 1994, 6, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Dempsey, D.; apos; Amick, M. ; Klessig, D.F. Salicylic Acid, a Multifaceted Hormone to Combat Disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, D.; Chu, J.Y.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Despres, C. The Arabidopsis NPR1 Protein Is a Receptor for the Plant Defense Hormone Salicylic Acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef]
- Mou, Z.; Fan, W.H.; Dong, X.N. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Zuo, J.; Dong, X. Plant immunity requires conformational changes [corrected] of NPR1 via S-nitrosylation and thioredoxins. Science (New York, N.Y.) 2008, 321, 952–956. [Google Scholar] [CrossRef]
- Mosblech, A.; Feussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiology and Biochemistry 2009, 47, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Chmielowska-Bąk, J.; Izbiańska, K.; Deckert, J. Products of lipid, protein and RNA oxidation as signals and regulators of gene expression in plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Alché, J.d.D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biology 2019, 23, 101136. [Google Scholar] [CrossRef]
- Knieper, M.; Viehhauser, A.; Dietz, K.J. Oxylipins and Reactive Carbonyls as Regulators of the Plant Redox and Reactive Oxygen Species Network under Stress. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Mano, J. Reactive carbonyl species: Their production from lipid peroxides, action in environmental stress, and the detoxification mechanism. Plant Physiology and Biochemistry 2012, 59, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I.; Mano, J.i. Lipid peroxidation-derived reactive carbonyl species (RCS): Their interaction with ROS and cellular redox during environmental stresses. Environmental and Experimental Botany 2019, 165, 139–149. [Google Scholar] [CrossRef]
- Mano, J.; Biswas, M.S.; Sugimoto, K. Reactive Carbonyl Species: A Missing Link in ROS Signaling. Plants (Basel) 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Vick, B.A.; Farmer, E.E. Dinor-oxo-phytodienoic acid: a new hexadecanoid signal in the jasmonate family. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 10473–10478. [Google Scholar] [CrossRef]
- Gosset, V.; Harmel, N.; Göbel, C.; Francis, F.; Haubruge, E.; Wathelet, J.P.; du Jardin, P.; Feussner, I.; Fauconnier, M.L. Attacks by a piercing-sucking insect (Myzus persicae Sultzer) or a chewing insect (Leptinotarsa decemlineata Say) on potato plants (Solanum tuberosum L.) induce differential changes in volatile compound release and oxylipin synthesis. J. Exp. Bot. 2009, 60, 1231–1240. [Google Scholar] [CrossRef]
- Gorshkov, V.Y.; Toporkova, Y.Y.; Tsers, I.D.; Smirnova, E.O.; Ogorodnikova, A.V.; Gogoleva, N.E.; Parfirova, O.I.; Petrova, O.E.; Gogolev, Y.V. Differential modulation of the lipoxygenase cascade during typical and latent Pectobacterium atrosepticum infections. Ann. Bot. 2022, 129, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.S.; Mano, J. Lipid Peroxide-Derived Reactive Carbonyl Species as Mediators of Oxidative Stress and Signaling. Front. Plant Sci. 2021, 12, 720867. [Google Scholar] [CrossRef]
- Mano, J.i.; Kanameda, S.; Kuramitsu, R.; Matsuura, N.; Yamauchi, Y. Detoxification of Reactive Carbonyl Species by Glutathione Transferase Tau Isozymes. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Sugimoto, K.; Matsuoka, Y.; Sakai, K.; Fujiya, N.; Fujii, H.; Mano, J.i. Catechins in green tea powder (matcha) are heat-stable scavengers of acrolein, a lipid peroxide-derived reactive carbonyl species. Food Chem. 2021, 355, 129403. [Google Scholar] [CrossRef]
- Yin, L.; Mano, J.i.; Tanaka, K.; Wang, S.; Zhang, M.; Deng, X.; Zhang, S. High level of reduced glutathione contributes to detoxification of lipid peroxide-derived reactive carbonyl species in transgenic Arabidopsis overexpressing glutathione reductase under aluminum stress. Physiol. Plant. 2017, 161, 211–223. [Google Scholar] [CrossRef]
- Roach, T.; Stöggl, W.; Baur, T.; Kranner, I. Distress and eustress of reactive electrophiles and relevance to light stress acclimation via stimulation of thiol/disulphide-based redox defences. Free Radical Biology and Medicine 2018, 122, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.S.; Mano, J. Lipid Peroxide-Derived Short-Chain Carbonyls Mediate Hydrogen Peroxide-Induced and Salt-Induced Programmed Cell Death in Plants. Plant Physiol. 2015, 168, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Ye, W.; Matsushima, D.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, S.; Mano, J.; Murata, Y. Reactive Carbonyl Species Mediate ABA Signaling in Guard Cells. Plant Cell Physiol 2016, 57, 2552–2563. [Google Scholar] [CrossRef]
- Islam, M.M.; Ye, W.; Matsushima, D.; Rhaman, M.S.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, M.S.; Mano, J.; Murata, Y. Reactive Carbonyl Species Function as Signal Mediators Downstream of H2O2 Production and Regulate [Ca2+]cyt Elevation in ABA Signal Pathway in Arabidopsis Guard Cells. Plant Cell Physiol 2019, 60, 1146–1159. [Google Scholar] [CrossRef]
- Biswas, M.S.; Terada, R.; Mano, J. Inactivation of carbonyl-detoxifying enzymes by H2O2 is a trigger to increase carbonyl load for initiating programmed cell death in plants. Antioxidants (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Mano, J.i.; Miyatake, F.; Hiraoka, E.; Tamoi, M. Evaluation of the toxicity of stress-related aldehydes to photosynthesis in chloroplasts. Planta 2009, 230, 639–648. [Google Scholar] [CrossRef]
- Parvez, S.; Long, M.J.C.; Poganik, J.R.; Aye, Y. Redox Signaling by Reactive Electrophiles and Oxidants. Chem. Rev. 2018, 118, 8798–8888. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.J.; Berger, S. Reactive electrophilic oxylipins: Pattern recognition and signalling. Phytochemistry 2009, 70, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Tola, A.J.; Jaballi, A.; Missihoun, T.D. Protein Carbonylation: Emerging Roles in Plant Redox Biology and Future Prospects. Plants 2021, 10, 1451. [Google Scholar] [CrossRef]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Petersen, D.R. Cysteine Modification by Lipid Peroxidation Products Inhibits Protein Disulfide Isomerase. Chem. Res. Toxicol. 2005, 18, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Marcocci, L.; Das, D.; Wang, X.; Luo, H.; Zungu-Edmondson, M.; Suzuki, Y.J. Mechanism of protein decarbonylation. Free Radic. Biol. Med. 2013, 65, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Chételat, A.; Reymond, P.; Farmer, E.E. Selective and powerful stress gene expression in Arabidopsis in response to malondialdehyde. The Plant Journal 2004, 37, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Tola, A.J.; Missihoun, T.D. Iron Availability Influences Protein Carbonylation in Arabidopsis thaliana Plants. Int. J. Mol. Sci. 2023, 24, 9732. [Google Scholar] [CrossRef]
- Maynard, D.; Viehhauser, A.; Knieper, M.; Dreyer, A.; Manea, G.; Telman, W.; Butter, F.; Chibani, K.; Scheibe, R.; Dietz, K.-J. The In Vitro Interaction of 12-Oxophytodienoic Acid and Related Conjugated Carbonyl Compounds with Thiol Antioxidants. Biomolecules 2021, 11, 457. [Google Scholar] [CrossRef]
- Millar, A.H.; Leaver, C.J. The cytotoxic lipid peroxidation product, 4-hydroxy-2-nonenal, specifically inhibits decarboxylating dehydrogenases in the matrix of plant mitochondria. FEBS Lett. 2000, 481, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Oeste, C.L.; Pérez-Sala, D. Modification of cysteine residues by cyclopentenone prostaglandins: Interplay with redox regulation of protein function. Mass Spectrom. Rev. 2014, 33, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Knieper, M.; Vogelsang, L.; Guntelmann, T.; Sproß, J.; Gröger, H.; Viehhauser, A.; Dietz, K.-J. OPDAylation of Thiols of the Redox Regulatory Network In Vitro. Antioxidants 2022, 11, 855. [Google Scholar] [CrossRef] [PubMed]
- Kaur, D.; Schedl, A.; Lafleur, C.; Martinez Henao, J.; van Dam, N.M.; Rivoal, J.; Bede, J.C. Arabidopsis Transcriptomics Reveals the Role of Lipoxygenase2 (AtLOX2) in Wound-Induced Responses. Int. J. Mol. Sci. 2024, 25, 5898. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Li, L.; Iwamoto, N.; Nakajima-Takagi, Y.; Kaneko, H.; Nakayama, Y.; Eguchi, M.; Wada, Y.; Kumagai, Y.; Yamamoto, M. The Antioxidant Defense System Keap1-Nrf2 Comprises a Multiple Sensing Mechanism for Responding to a Wide Range of Chemical Compounds. Molecular and Cellular Biology 2009, 29, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Hölzl, G.; Dörmann, P. Chloroplast Lipids and Their Biosynthesis. Annu. Rev. Plant Biol. 2019, 70, 51–81. [Google Scholar] [CrossRef] [PubMed]
- Hemsley, P.A. S-acylation in plants: an expanding field. Biochem. Soc. Trans. 2020, 48, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Smotrys, J.E.; Linder, M.E. Palmitoylation of Intracellular Signaling Proteins: Regulation and Function. Annu. Rev. Biochem. 2004, 73, 559–587. [Google Scholar] [CrossRef]
- Kumar, M.; Carr, P.; Turner, S.R. An atlas of Arabidopsis protein S-acylation reveals its widespread role in plant cell organization and function. Nat. Plants 2022, 8, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.F.; Feng, Y.; Chen, L.; Davis, N.G. The yeast DHHC cysteine-rich domain protein Akr1p is a palmitoyl transferase. J. Cell Biol. 2002, 159, 23–28. [Google Scholar] [CrossRef]
- Hou, H.; John Peter, A.T.; Meiringer, C.; Subramanian, K.; Ungermann, C. Analysis of DHHC Acyltransferases Implies Overlapping Substrate Specificity and a Two-Step Reaction Mechanism. Traffic 2009, 10, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Abdulrahman, D.A.; Meng, X.; Veit, M. S-Acylation of Proteins of Coronavirus and Influenza Virus: Conservation of Acylation Sites in Animal Viruses and DHHC Acyltransferases in Their Animal Reservoirs. Pathogens 2021, 10, 669. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zeng, H.; Wu, J.; Huang, J.; Gao, Q.; Tang, D.; Cai, L.; Liao, Z.; Wang, Y.; Liu, X.; et al. Screening DHHCs of S-acylated proteins using an OsDHHC cDNA library and bimolecular fluorescence complementation in rice. The Plant Journal 2022, 110, 1763–1780. [Google Scholar] [CrossRef] [PubMed]
- Batistič, O. Genomics and Localization of the Arabidopsis DHHC-Cysteine-Rich Domain S-Acyltransferase Protein Family. Plant Physiol. 2012, 160, 1597–1612. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, D.; Hemsley, P.A. Fats and function: protein lipid modifications in plant cell signalling. Curr. Opin. Plant Biol. 2017, 40, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, R.N.P.; Snijder, J.; van de Waterbeemd, M.; Schouten, A.; Granneman, J.; Heck, A.J.R.; Gros, P. Stochastic palmitoylation of accessible cysteines in membrane proteins revealed by native mass spectrometry. Nature Communications 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Li, E.; Feng, Q.N.; Zhao, X.Y.; Ge, F.R.; Zhang, Y.; Li, S. Protein palmitoylation is critical for the polar growth of root hairs in Arabidopsis. BMC Plant Biol. 2015, 15. [Google Scholar] [CrossRef]
- Blaskovic, S.; Blanc, M.; van der Goot, F.G. What does S-palmitoylation do to membrane proteins? FEBS J. 2013, 280, 2766–2774. [Google Scholar] [CrossRef] [PubMed]
- Joseph, M.; Nagaraj, R. Interaction of Peptides Corresponding to Fatty Acylation Sites in Proteins with Model Membranes (∗). J. Biol. Chem. 1995, 270, 16749–16755. [Google Scholar] [CrossRef]
- Purushotham, P.; Ho, R.; Zimmer, J. Architecture of a catalytically active homotrimeric plant cellulose synthase complex. Science 2020, 369, 1089–1094. [Google Scholar] [CrossRef]
- Kumar, M.; Wightman, R.; Atanassov, I.; Gupta, A.; Hurst, C.H.; Hemsley, P.A.; Turner, S. S-Acylation of the cellulose synthase complex is essential for its plasma membrane localization. Science 2016, 353, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.X.; Beckmann, L.; Kudla, J.; Batistic, O. N-terminal S-acylation facilitates tonoplast targeting of the calcium sensor CBL6. FEBS Lett. 2017, 591, 3745–3756. [Google Scholar] [CrossRef]
- Saito, S.; Hamamoto, S.; Moriya, K.; Matsuura, A.; Sato, Y.; Muto, J.; Noguchi, H.; Yamauchi, S.; Tozawa, Y.; Ueda, M.; et al. N-myristoylation and S-acylation are common modifications of Ca2+-regulated Arabidopsis kinases and are required for activation of the SLAC1 anion channel. New Phytol. 2018, 218, 1504–1521. [Google Scholar] [CrossRef]
- Fan, R.; Zhao, F.; Gong, Z.; Chen, Y.; Yang, B.; Zhou, C.; Zhang, J.; Du, Z.; Wang, X.; Yin, P.; et al. Insights into the mechanism of phospholipid hydrolysis by plant non-specific phospholipase C. Nature Communications 2023, 14, 194. [Google Scholar] [CrossRef]
- Ali, U.; Lu, S.; Fadlalla, T.; Iqbal, S.; Yue, H.; Yang, B.; Hong, Y.; Wang, X.; Guo, L. The functions of phospholipases and their hydrolysis products in plant growth, development and stress responses. Prog. Lipid Res. 2022, 86, 101158. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, K.; Jin, X.; Yan, J.; Lu, S.; Shen, Q.; Guo, L.; Hong, Y.; Wang, X.; Guo, L. Acylation of non-specific phospholipase C4 determines its function in plant response to phosphate deficiency. The Plant Journal 2021, 106, 1647–1659. [Google Scholar] [CrossRef] [PubMed]
- Dekker, F.J.; Rocks, O.; Vartak, N.; Menninger, S.; Hedberg, C.; Balamurugan, R.; Wetzel, S.; Renner, S.; Gerauer, M.; Schölermann, B.; et al. Small-molecule inhibition of APT1 affects Ras localization and signaling. Nat. Chem. Biol. 2010, 6, 449–456. [Google Scholar] [CrossRef]
- Lin, D.T.S.; Conibear, E. ABHD17 proteins are novel protein depalmitoylases that regulate N-Ras palmitate turnover and subcellular localization. eLife 2015, 4, e11306. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, M.; Li, Y.; Chen, Z.; Zhuge, C.; Ouyang, Y.; Zhao, Y.; Lin, Y.; Xie, Q.; Yang, C.; et al. An ABHD17-like hydrolase screening system to identify de-S-acylation enzymes of protein substrates in plant cells. Plant Cell 2021, 33, 3235–3249. [Google Scholar] [CrossRef]
- Duan, M.; Zhang, R.; Zhu, F.; Zhang, Z.; Gou, L.; Wen, J.; Dong, J.; Wang, T. A Lipid-Anchored NAC Transcription Factor Is Translocated into the Nucleus and Activates Glyoxalase I Expression during Drought Stress. Plant Cell 2017, 29, 1748–1772. [Google Scholar] [CrossRef]
- Ji, T.; Zheng, L.H.; Wu, J.L.; Duan, M.; Liu, Q.W.; Liu, P.; Shen, C.; Liu, J.L.; Ye, Q.Y.; Wen, J.Q.; et al. The thioesterase APT1 is a bidirectional-adjustment redox sensor. Nature Communications 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional Regulators of Plant Growth, Development, and Stress Responses[OPEN]. Plant Cell 2019, 32, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic Acid: Biosynthesis and Signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Chen, Z.; Huang, L.T.; Ouyang, Y.W.; Wang, Z.Y.; Wu, S.; Ye, W.X.; Yu, B.Y.; Zhang, Y.H.; Yang, C.W.; et al. Salicylic acid attenuates brassinosteroid signaling via protein de-S-acylation. EMBO J. 2023, 42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.L.; Casey, P.J. Protein prenylation: Molecular mechanisms and functional consequences. Annu. Rev. Biochem. 1996, 65, 241–269. [Google Scholar] [CrossRef] [PubMed]
- Rohmer, M. The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria, algae and higher plants. Nat.Prod.Rep. 1999, 16, 565–574. [Google Scholar] [CrossRef]
- Yalovsky, S. Protein prenylation CaaX processing in plants. In The Enzymes; Tamanoi, F., Hrycyna, C.A., Bergo, M.O., Eds.; Academic Press, 2011; Volume 29, pp. 163–182. [Google Scholar]
- Downes, B.P.; Saracco, S.A.; Lee, S.S.; Crowell, D.N.; Vierstra, R.D. MUBs, a Family of Ubiquitin-fold Proteins That Are Plasma Membrane-anchored by Prenylation*. J. Biol. Chem. 2006, 281, 27145–27157. [Google Scholar] [CrossRef]
- Bracha-Drori, K.; Shichrur, K.; Lubetzky, T.C.; Yalovsky, S. Functional analysis of Arabidopsis postprenylation CaaX processing enzymes and their function in subcellular protein targeting. Plant Physiol. 2008, 148, 119–131. [Google Scholar] [CrossRef]
- Hála, M.; Žárský, V. Protein prenylation in plant stress responses. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Running, M. The role of lipid post–translational modification in plant developmental processes. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef]
- Wilson, A.L.; Erdman, R.A.; Castellano, F.; Maltese, W.A. Prenylation of Rab8 GTPase by type I and type II geranylgeranyl transferases. Biochem. J. 1998, 333, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Maurer-Stroh, S.; Eisenhaber, F. Refinement and prediction of protein prenylation motifs. Genome Biol. 2005, 6, R55. [Google Scholar] [CrossRef] [PubMed]
- Quinn, O.; Kumar, M.; Turner, S. The role of lipid-modified proteins in cell wall synthesis and signaling. Plant Physiol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D. Pentose Phosphate Pathway Reactions in Photosynthesizing Cells. Cells 2021, 10, 1547. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.; Hölscher, C.; Schwöppe, C.; von Schaewen, A. Alternative targeting of Arabidopsis plastidic glucose-6-phosphate dehydrogenase G6PD1 involves cysteine-dependent interaction with G6PD4 in the cytosol. The Plant Journal 2011, 66, 745–758. [Google Scholar] [CrossRef]
- Linnenbrügger, L.; Doering, L.; Lansing, H.; Fischer, K.; Eirich, J.; Finkemeier, I.; von Schaewen, A. Alternative splicing of Arabidopsis G6PD5 recruits NADPH-producing OPPP reactions to the endoplasmic reticulum. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, R.; Zhang, Y.-M.; Rock, C.O.; Jackowski, S. Coenzyme A: Back in action. Prog. Lipid Res. 2005, 44, 125–153. [Google Scholar] [CrossRef]
- Kupke, T.; Hernández-Acosta, P.; Steinbacher, S.; Culiáñez-Macià, F.A. Arabidopsis thaliana Flavoprotein AtHAL3a Catalyzes the Decarboxylation of 4′-Phosphopantothenoylcysteine to 4′-Phosphopantetheine, a Key Step in Coenzyme A Biosynthesis*. J. Biol. Chem. 2001, 276, 19190–19196. [Google Scholar] [CrossRef]
- Kupke, T.; Hernández-Acosta, P.; Culiáñez-Macià, F.A. 4′-Phosphopantetheine and Coenzyme A Biosynthesis in Plants*. J. Biol. Chem. 2003, 278, 38229–38237. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Peak-Chew, S.Y.; Newell, C.; Miller-Aidoo, S.; Mangal, S.; Zhyvoloup, A.; Bakovic´, J.; Malanchuk, O.; Pereira, G.C.; Kotiadis, V.; et al. Protein CoAlation: a redox-regulated protein modification by coenzyme A in mammalian cells. Biochem. J. 2017, 474, 2489–2508. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Zhyvoloup, A.; Bakovic, J.; Thomas, N.; Yu, B.Y.K.; Das, S.; Orengo, C.; Newell, C.; Ward, J.; Saladino, G.; et al. Protein CoAlation and antioxidant function of coenzyme A in prokaryotic cells. Biochem. J. 2018, 475, 1909–1937. [Google Scholar] [CrossRef] [PubMed]
- Aloum, L.; Brimson, C.A.; Zhyvoloup, A.; Baines, R.; Baković, J.; Filonenko, V.; Thompson, C.R.L.; Gout, I. Coenzyme A and protein CoAlation levels are regulated in response to oxidative stress and during morphogenesis in Dictyostelium discoideum. Biochemical and Biophysical Research Communications 2019, 511, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Clares, R.A.; González-Segura, L.; Díaz-Sánchez, Á.G. Crystallographic evidence for active-site dynamics in the hydrolytic aldehyde dehydrogenases. Implications for the deacylation step of the catalyzed reaction. Chemico-Biological Interactions 2011, 191, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.N.; Haines, B.E.; Stauffacher, C.V.; Helquist, P.; Wiest, O. Computational Study of Base-Catalyzed Thiohemiacetal Decomposition in Pseudomonas mevalonii HMG-CoA Reductase. The Journal of Physical Chemistry B 2023, 127, 4931–4938. [Google Scholar] [CrossRef] [PubMed]
- Hanson, A.D.; Rathinasabapathi, B.; Rivoal, J.; Burnet, M.; Dillon, M.O.; Gage, D.A. Osmoprotective compounds in the Plumbaginaceae: a natural experiment in metabolic engineering of stress tolerance. Proc.Natl.Acad.Sci.U.S.A 1994, 91, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Zárate-Romero, A.; Murillo-Melo, Darío S. ; Mújica-Jiménez, C.; Montiel, C.; Muñoz-Clares, Rosario A. Reversible, partial inactivation of plant betaine aldehyde dehydrogenase by betaine aldehyde: mechanism and possible physiological implications. Biochem. J. 2016, 473, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, G.C.; Dahal, K.; Alber, N.A.; Chadee, A. Photosynthesis, respiration and growth: A carbon and energy balancing act for alternative oxidase. Mitochondrion 2020, 52, 197–211. [Google Scholar] [CrossRef]
- Umbach, A.L.; Siedow, J.N. The reaction of the soybean cotyledon mitochondrial cyanide-resistant oxidase with sulfhydryl reagents suggests that alpha-keto acid activation involves the formation of a thiohemiacetal. J. Biol. Chem. 1996, 271, 25019–25026. [Google Scholar] [CrossRef]
- Rhoads, D.M.; Umbach, A.L.; Sweet, C.R.; Lennon, A.M.; Rauch, G.S.; Siedow, J.N. Regulation of the cyanide-resistant alternative oxidase of plant mitochondria -: Identification of the cysteine residue involved in α-keto acid stimulation and intersubunit disulfide bond formation. J. Biol. Chem. 1998, 273, 30750–30756. [Google Scholar] [CrossRef]
- Selinski, J.; Hartmann, A.; Kordes, A.; Deckers-Hebestreit, G.; Whelan, J.; Scheibe, R. Analysis of Posttranslational Activation of Alternative Oxidase Isoforms. Plant Physiol. 2017, 174, 2113–2127. [Google Scholar] [CrossRef]
Figure 1.
Protein S-sulfenylation in plants. A: The transition of Cys residues from thiolate to S-sulfenylated state. A Cys residue (yellow dot) in deprotonated state (thiolate, S-) (top left corner) undergoes oxidation to its sulfenic acid form (S-OH) upon exposure to oxidative conditions caused by reactive oxygen species (ROS). Reductants such as dithiothreitol (DTT), ascorbate (ASC) or thioredoxin (TRX) have been shown to reverse this reaction. The S-sulfenylated form allows further Cys modifications, such as generation of a reversible mixed disulfide with glutathione (GSH) to yield an S-glutathionylation (S-SG); a reaction with a protein Cys in thiolate form, to generate a reversible disulfide bridge (S-S); a reaction with H2S to generate a persulfidation (S-SH); and a higher degree of oxidation towards the sulfinic acid form (SO2H). The latter reaction can be reversed with an ATP-dependent sulfiredoxin (SRX). B: Inhibition of plastidial triose phosphate isomerase (pTPI) by S-sulfenylation. Under high H2O2 conditions created by salt stress, pTPI becomes inactive due to S-sulfenylation. The inability of pTPI to process triose phosphates leads to the formation of methylglyoxal, which acts as a promoter of H2O2 formation. C: Regulation of MAPK4 by S-sulfenylation. MAPK4 is a negative regulator of resistance to Pseudomonas syringae. Under normal conditions, the protein is active, and suppresses resistance to the pathogen. Under oxidative stress conditions, a Cys residue of the protein’s Common Docking (CD) motif becomes S-sulfenylated, thereby disrupting MAPK4 function and enabling resistance to the pathogen. See text for additional details.
Figure 1.
Protein S-sulfenylation in plants. A: The transition of Cys residues from thiolate to S-sulfenylated state. A Cys residue (yellow dot) in deprotonated state (thiolate, S-) (top left corner) undergoes oxidation to its sulfenic acid form (S-OH) upon exposure to oxidative conditions caused by reactive oxygen species (ROS). Reductants such as dithiothreitol (DTT), ascorbate (ASC) or thioredoxin (TRX) have been shown to reverse this reaction. The S-sulfenylated form allows further Cys modifications, such as generation of a reversible mixed disulfide with glutathione (GSH) to yield an S-glutathionylation (S-SG); a reaction with a protein Cys in thiolate form, to generate a reversible disulfide bridge (S-S); a reaction with H2S to generate a persulfidation (S-SH); and a higher degree of oxidation towards the sulfinic acid form (SO2H). The latter reaction can be reversed with an ATP-dependent sulfiredoxin (SRX). B: Inhibition of plastidial triose phosphate isomerase (pTPI) by S-sulfenylation. Under high H2O2 conditions created by salt stress, pTPI becomes inactive due to S-sulfenylation. The inability of pTPI to process triose phosphates leads to the formation of methylglyoxal, which acts as a promoter of H2O2 formation. C: Regulation of MAPK4 by S-sulfenylation. MAPK4 is a negative regulator of resistance to Pseudomonas syringae. Under normal conditions, the protein is active, and suppresses resistance to the pathogen. Under oxidative stress conditions, a Cys residue of the protein’s Common Docking (CD) motif becomes S-sulfenylated, thereby disrupting MAPK4 function and enabling resistance to the pathogen. See text for additional details.
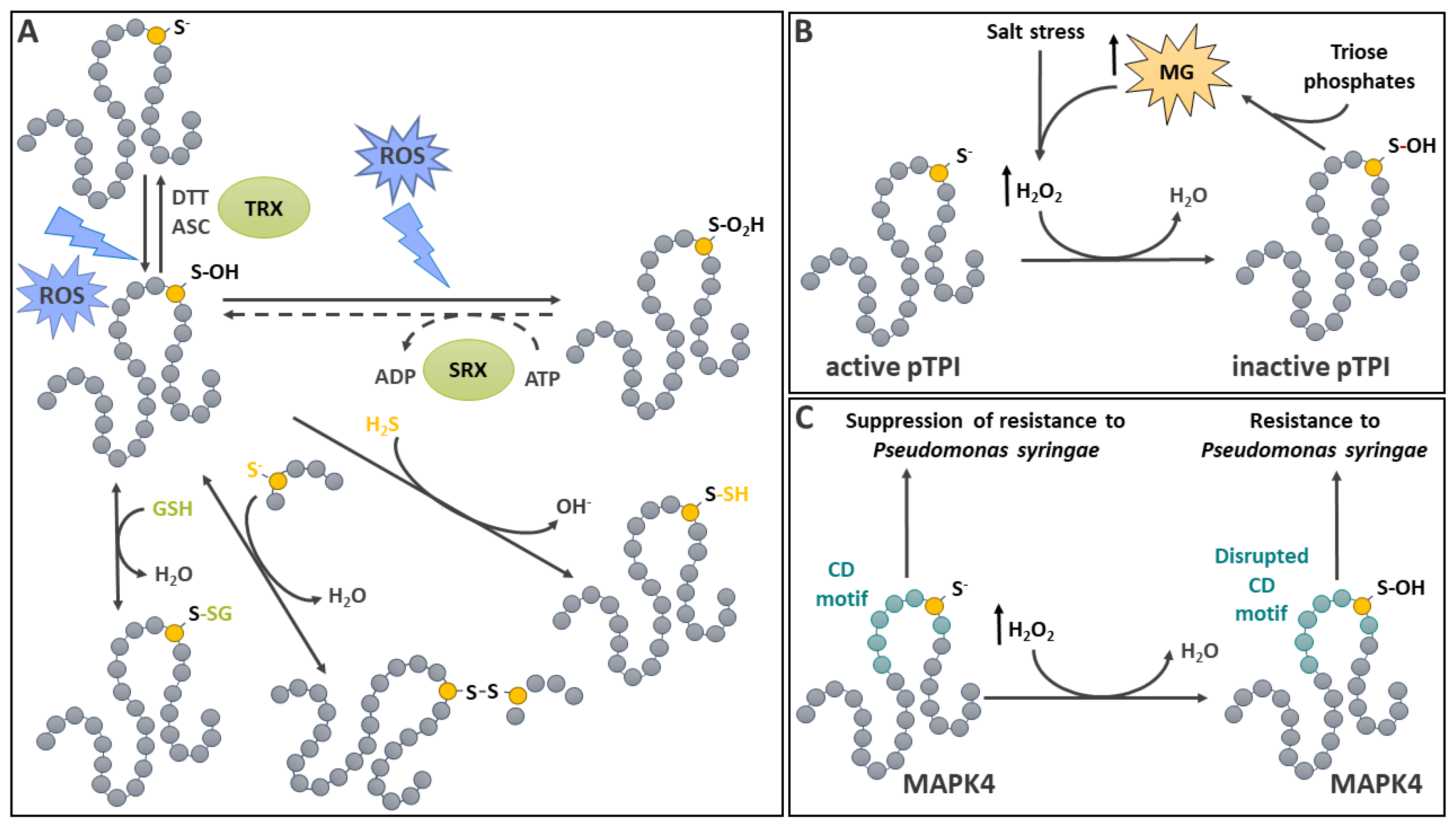
Figure 2.
Protein S-sulfinylation and S-sulfonylation in plants. A: The transition of Cys residues from thiol to S-sulfonylated state. When fully protonated (left), Cys residues (yellow dots) are in the thiol state (SH). Depending on conditions prevailing in the environment, the thiol can deprotonate to form a nucleophilic thiolate (S-) sensitive to oxidation by H2O2. Sequential oxidations of the thiolate by H2O2 (left to right) lead to its S-sulfenylation (SOH), S-sulfinylation (SO2H) and S-sulfonylation (SO3H). The last step is considered irreversible. B: Modulation of peroxiredoxin functions by oxidation status. Peroxiredoxin (PRX) detoxifies H2O2 to H2O using a peroxidatic (catalytic) Cys (CysP). The reaction oxidizes CysP to its sulfenic acid form, which can be reduced back with the help of a thioredoxin (TRX). In the presence of H2O2, the sulfenylated CysP can be further oxidized to a sulfinic acid. This step is reversible in a reaction catalyzed by sulfiredoxin (SRX). Sulfinylated PRX can lead to the formation of an oligomeric, catalytically inactive PRX displaying in vitro chaperone function. C: Involvement of plant cysteine oxidases in O2 status signal transduction. The transcription factor RAP2.12 positively regulates plant gene expression in response to hypoxia. Under normoxic conditions, RAP2.12 has low stability because it is oxidized by a plant cysteine oxidase (PCO), leading to sulfinylation of an N-terminal Cys, which is then arginylated by a tRNA-ARGINYL-TRANSFERASE (t-RNA AT). This process leads RAP2.12 to its degradation by the proteasome. Under hypoxia, PCO is inactivated, leading to the stabilization of RAP2.12, allowing the promotion of hypoxic-responsive genes expression. See text for additional details.
Figure 2.
Protein S-sulfinylation and S-sulfonylation in plants. A: The transition of Cys residues from thiol to S-sulfonylated state. When fully protonated (left), Cys residues (yellow dots) are in the thiol state (SH). Depending on conditions prevailing in the environment, the thiol can deprotonate to form a nucleophilic thiolate (S-) sensitive to oxidation by H2O2. Sequential oxidations of the thiolate by H2O2 (left to right) lead to its S-sulfenylation (SOH), S-sulfinylation (SO2H) and S-sulfonylation (SO3H). The last step is considered irreversible. B: Modulation of peroxiredoxin functions by oxidation status. Peroxiredoxin (PRX) detoxifies H2O2 to H2O using a peroxidatic (catalytic) Cys (CysP). The reaction oxidizes CysP to its sulfenic acid form, which can be reduced back with the help of a thioredoxin (TRX). In the presence of H2O2, the sulfenylated CysP can be further oxidized to a sulfinic acid. This step is reversible in a reaction catalyzed by sulfiredoxin (SRX). Sulfinylated PRX can lead to the formation of an oligomeric, catalytically inactive PRX displaying in vitro chaperone function. C: Involvement of plant cysteine oxidases in O2 status signal transduction. The transcription factor RAP2.12 positively regulates plant gene expression in response to hypoxia. Under normoxic conditions, RAP2.12 has low stability because it is oxidized by a plant cysteine oxidase (PCO), leading to sulfinylation of an N-terminal Cys, which is then arginylated by a tRNA-ARGINYL-TRANSFERASE (t-RNA AT). This process leads RAP2.12 to its degradation by the proteasome. Under hypoxia, PCO is inactivated, leading to the stabilization of RAP2.12, allowing the promotion of hypoxic-responsive genes expression. See text for additional details.
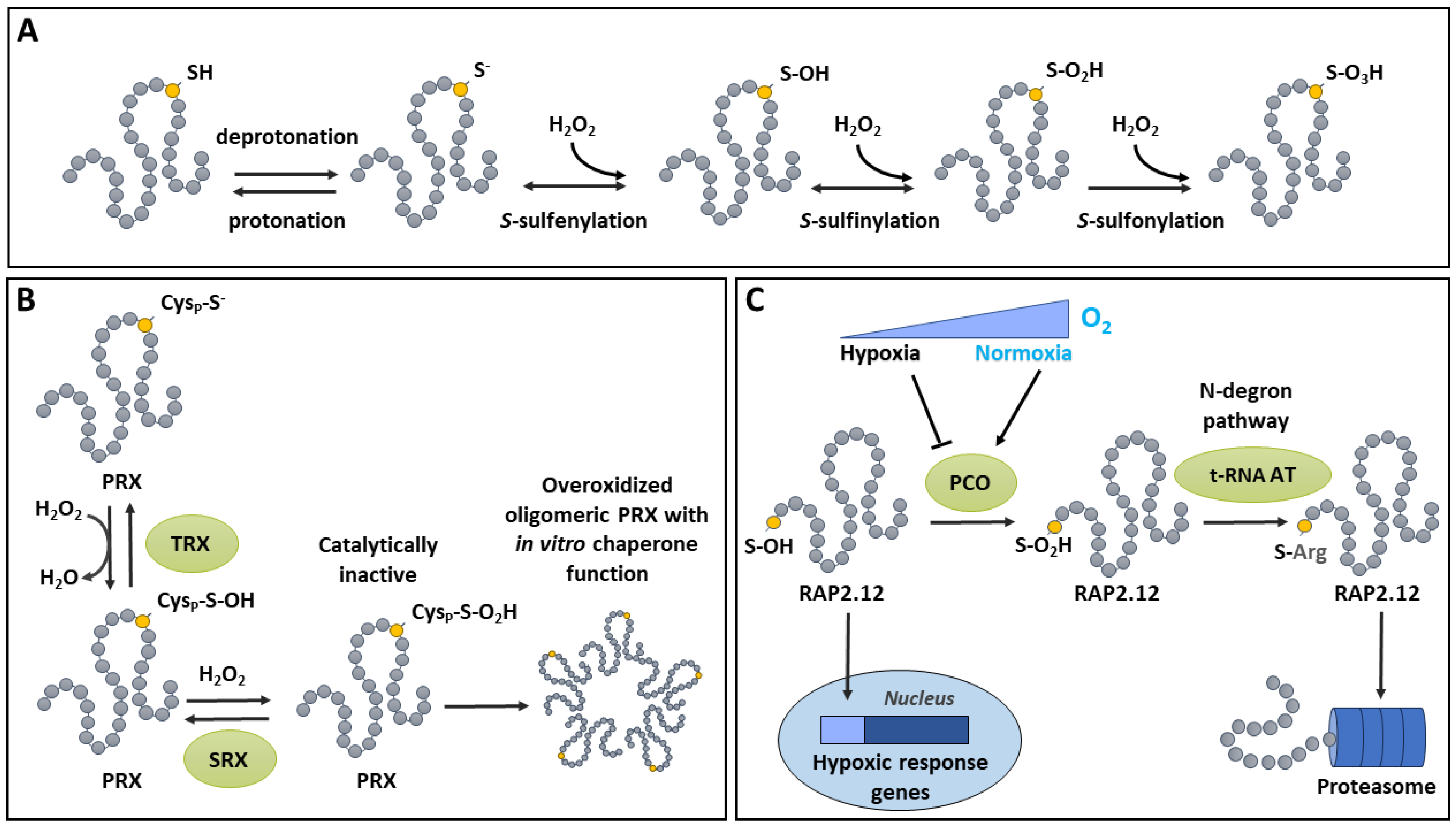
Figure 3.
Reversible disulfide bridge formation in plants. A: Reversible formation of disulfide bridge (S-S). Oxidative conditions promoted by ROS and oxidized protein disulfide isomerase (PDIox) in the endoplasmic reticulum favour S-S formation between two adjacent Cys (yellow dots). Thioredoxin (TRX) and glutaredoxins (GRX) reduce S-S, concomitantly with consumption of reducing power (e-). In the ER, reduced PDI (PDIred) can reduce S-S on misfolded proteins. B: Importance of disulfide bridge formation for metabolic regulation in the light and the dark. In the light (white background), the photosynthetic electron transport chain (ETC) generates reducing power for the reduction of thioredoxins (TRX) via ferredoxin (Fd) and ferredoxin thioredoxin reductase (FTR). Reduced TRX reduces disulfide bridges on targets in the chloroplast stroma. Activated targets (pointed arrow) include enzymes of the Calvin-Benson-Bassham (CBB) cycle: fructose-1,6-bisphosphatase (FBPase); sedoheptulose-1,7-bisphosphatase (SBPase), phosphoribulo kinase (PRK) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The oxidative pentose phosphate pathway enzyme glucose-6-phosphate dehydrogenase (G6PDH) is inactivated (blunted arrow). At the onset of dark conditions (grey background), targets become oxidized (S-S). In these conditions, reduced TRX becomes inactive by transferring electrons to 2-Cys peroxiredoxin (2CP), thereby terminating activation of CBB cycle enzymes. Note that, in the light, 2CP detoxifies H2O2 generated at the level of ETC and is maintained reduced by NADP-thioredoxin reductase C (NTRC) using photosynthetic NADPH. Subscripts red and ox respectively symbolize reduced and oxidized state. See text for additional details.
Figure 3.
Reversible disulfide bridge formation in plants. A: Reversible formation of disulfide bridge (S-S). Oxidative conditions promoted by ROS and oxidized protein disulfide isomerase (PDIox) in the endoplasmic reticulum favour S-S formation between two adjacent Cys (yellow dots). Thioredoxin (TRX) and glutaredoxins (GRX) reduce S-S, concomitantly with consumption of reducing power (e-). In the ER, reduced PDI (PDIred) can reduce S-S on misfolded proteins. B: Importance of disulfide bridge formation for metabolic regulation in the light and the dark. In the light (white background), the photosynthetic electron transport chain (ETC) generates reducing power for the reduction of thioredoxins (TRX) via ferredoxin (Fd) and ferredoxin thioredoxin reductase (FTR). Reduced TRX reduces disulfide bridges on targets in the chloroplast stroma. Activated targets (pointed arrow) include enzymes of the Calvin-Benson-Bassham (CBB) cycle: fructose-1,6-bisphosphatase (FBPase); sedoheptulose-1,7-bisphosphatase (SBPase), phosphoribulo kinase (PRK) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The oxidative pentose phosphate pathway enzyme glucose-6-phosphate dehydrogenase (G6PDH) is inactivated (blunted arrow). At the onset of dark conditions (grey background), targets become oxidized (S-S). In these conditions, reduced TRX becomes inactive by transferring electrons to 2-Cys peroxiredoxin (2CP), thereby terminating activation of CBB cycle enzymes. Note that, in the light, 2CP detoxifies H2O2 generated at the level of ETC and is maintained reduced by NADP-thioredoxin reductase C (NTRC) using photosynthetic NADPH. Subscripts red and ox respectively symbolize reduced and oxidized state. See text for additional details.
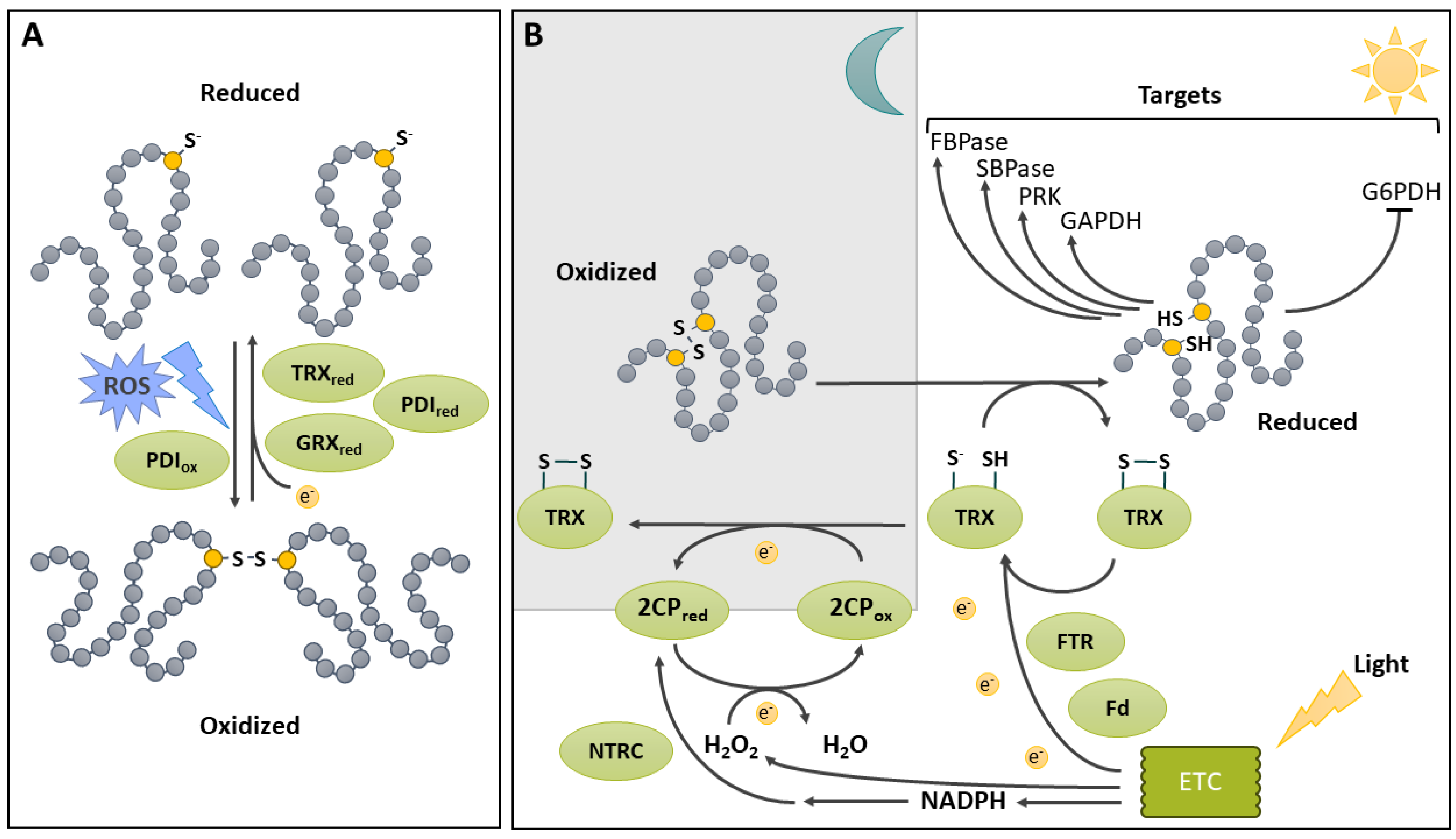
Figure 4.
Protein S-glutathionylation in plants. A: Mechanisms of S-glutathionylation/deglutathionylation and regulation by S-glutathionylation. S-glutathionylation (S-SG) occurs on deprotonated (S-) Cys (yellow dot) in a glutaredoxin (GRX) catalyzed reaction with oxidized glutathione (GSSG), or spontaneously in a reaction involving nitrosoglutathione (GSNO). It can also occur spontaneously between a sulfenylated Cys (S-OH) and reduced glutathione (GSH). Deglutathionylation is catalyzed by GSH-dependent GRX or by thioredoxin (TRX). S-glutahionylation of a Cys residue protects the protein from ROS-dependent oxidation towards the S-sulfinylated form (-SO2H, top of the panel). S-glutathionylation protects amylase 3 (AMY3) from overoxidation and inhibits its activity towards starch degradation. AMY3 activity is recovered after stress It also has an inhibitory effect on cytosolic NADP-dependent isocitrate dehydrogenase (cICDH), which is active in nitrogen assimilation. B: Involvement of S-glutathionylation of SAL1 in expression of plastid redox-associated nuclear genes. SAL1 is a 3′-phosphoadenosine 5′-phosphate (PAP) phosphatase that controls the degradation of PAP to AMP in the chloroplast. In oxidative stress conditions, S-glutathionylation of SAL1 induces the formation of an intramolecular disulfide bridge (S-S) which participates in increasing PAP levels. According to a model, PAP is exported to the nucleus, where it binds 5′–3′ exoribonucleases (XRN) involved in inhibition of RNA polymerase II (RNA polII) transcription of plastid redox-associated nuclear genes (PRANGs). PAP binding to XRN relieves inhibition of RNA polII, allowing PRANGs transcription. See text for additional details.
Figure 4.
Protein S-glutathionylation in plants. A: Mechanisms of S-glutathionylation/deglutathionylation and regulation by S-glutathionylation. S-glutathionylation (S-SG) occurs on deprotonated (S-) Cys (yellow dot) in a glutaredoxin (GRX) catalyzed reaction with oxidized glutathione (GSSG), or spontaneously in a reaction involving nitrosoglutathione (GSNO). It can also occur spontaneously between a sulfenylated Cys (S-OH) and reduced glutathione (GSH). Deglutathionylation is catalyzed by GSH-dependent GRX or by thioredoxin (TRX). S-glutahionylation of a Cys residue protects the protein from ROS-dependent oxidation towards the S-sulfinylated form (-SO2H, top of the panel). S-glutathionylation protects amylase 3 (AMY3) from overoxidation and inhibits its activity towards starch degradation. AMY3 activity is recovered after stress It also has an inhibitory effect on cytosolic NADP-dependent isocitrate dehydrogenase (cICDH), which is active in nitrogen assimilation. B: Involvement of S-glutathionylation of SAL1 in expression of plastid redox-associated nuclear genes. SAL1 is a 3′-phosphoadenosine 5′-phosphate (PAP) phosphatase that controls the degradation of PAP to AMP in the chloroplast. In oxidative stress conditions, S-glutathionylation of SAL1 induces the formation of an intramolecular disulfide bridge (S-S) which participates in increasing PAP levels. According to a model, PAP is exported to the nucleus, where it binds 5′–3′ exoribonucleases (XRN) involved in inhibition of RNA polymerase II (RNA polII) transcription of plastid redox-associated nuclear genes (PRANGs). PAP binding to XRN relieves inhibition of RNA polII, allowing PRANGs transcription. See text for additional details.
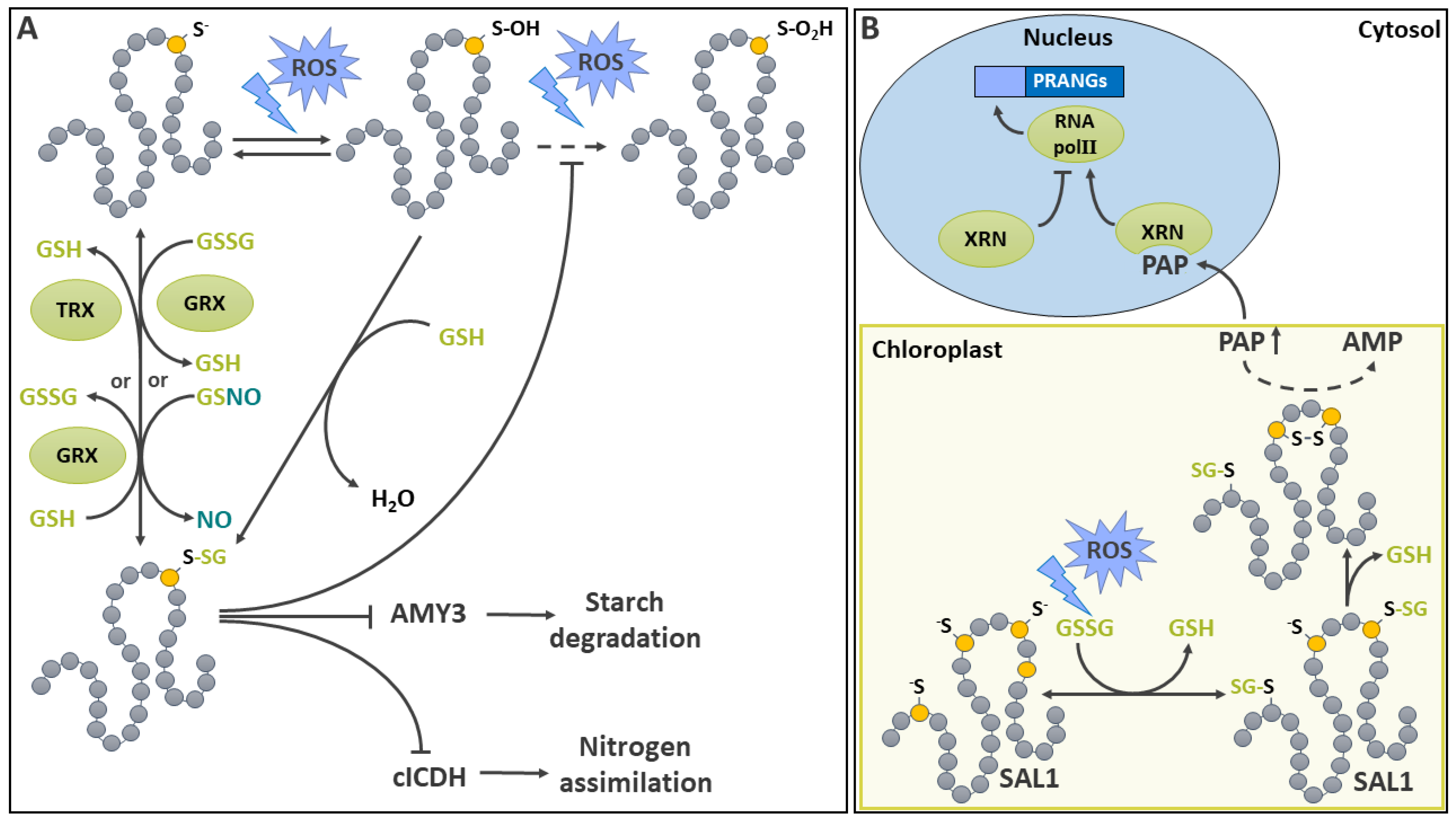
Figure 5.
Protein persulfidation in plants. A: Pathways controlling cellular H2S together with mechanisms of persulfidation. In the top of the panel, pathways that promote cellular H2S levels are represented. These include uptake from the environment, SO42- assimilation, 3-mercaptopyruvate sulfurtransferases (MSTs), and the catabolism of free Cys by L-Cys desulfhydrases (LCD), D-Cys desulfhydrases (DCD) and the cytosolic O-acetylserine (thiol)lyase (OSTL) homolog DES1, while OSTL consumes H2S for Cys synthesis. Note that H2S is used, although HS- is probably present in physiological conditions. The bottom of the panel represents the two pathways implicated in protein persulfidation. The direct persulfidation pathway modifies Cys residues (yellow dots) in the S-sulfenylated form (S-SH)) using free H2S. Other mechanisms of direct persulfidation (dotted arrow) do not have strong experimental support. The transpersulfidation pathway allows the transfer of a persulfide group from MST to a thiolated Cys (S-). B: Metabolic targets of persulfidation in plants. The effect of persulfidation is represented (pointed arrow: increased activity, blunt arrow: inhibition). Some target names are abbreviated: ACO, aminocyclopropane-carboxylic acid oxidase; APX: cytosolic ascorbate peroxidase; G6PDH, glucose-6-phosphate dehydrogenase; GAPDH: cytosolic glyceraldehyde 3P dehydrogenase. Increased nuclear localization has also been reported as a result of GAPDH persulfidation. C: Involvement of persulfidation in signal transduction. H2S serves to activate DES1 by persulfidation. Upon abscisic acid (ABA) stimulation, DES1-derived H2S modifies Open Stomata 1/Sucrose nonfermenting 1-RELATED PROTEIN KINASE2.6 (OST1/SnRK2-6), which is inhibited by NO signaling and responsible for the inhibitory phosphorylation of the KAT1 potassium channel in the guard cell membrane. Phosphorylation of KAT1 inhibits potassium transport, leading to stomatal closure. In a regulatory negative feedback loop, modification of Respiratory Burst Oxidase Homolog D (RBOHD) by H2S activates it, leading to ROS formation and inhibition of DES1. See text for additional details.
Figure 5.
Protein persulfidation in plants. A: Pathways controlling cellular H2S together with mechanisms of persulfidation. In the top of the panel, pathways that promote cellular H2S levels are represented. These include uptake from the environment, SO42- assimilation, 3-mercaptopyruvate sulfurtransferases (MSTs), and the catabolism of free Cys by L-Cys desulfhydrases (LCD), D-Cys desulfhydrases (DCD) and the cytosolic O-acetylserine (thiol)lyase (OSTL) homolog DES1, while OSTL consumes H2S for Cys synthesis. Note that H2S is used, although HS- is probably present in physiological conditions. The bottom of the panel represents the two pathways implicated in protein persulfidation. The direct persulfidation pathway modifies Cys residues (yellow dots) in the S-sulfenylated form (S-SH)) using free H2S. Other mechanisms of direct persulfidation (dotted arrow) do not have strong experimental support. The transpersulfidation pathway allows the transfer of a persulfide group from MST to a thiolated Cys (S-). B: Metabolic targets of persulfidation in plants. The effect of persulfidation is represented (pointed arrow: increased activity, blunt arrow: inhibition). Some target names are abbreviated: ACO, aminocyclopropane-carboxylic acid oxidase; APX: cytosolic ascorbate peroxidase; G6PDH, glucose-6-phosphate dehydrogenase; GAPDH: cytosolic glyceraldehyde 3P dehydrogenase. Increased nuclear localization has also been reported as a result of GAPDH persulfidation. C: Involvement of persulfidation in signal transduction. H2S serves to activate DES1 by persulfidation. Upon abscisic acid (ABA) stimulation, DES1-derived H2S modifies Open Stomata 1/Sucrose nonfermenting 1-RELATED PROTEIN KINASE2.6 (OST1/SnRK2-6), which is inhibited by NO signaling and responsible for the inhibitory phosphorylation of the KAT1 potassium channel in the guard cell membrane. Phosphorylation of KAT1 inhibits potassium transport, leading to stomatal closure. In a regulatory negative feedback loop, modification of Respiratory Burst Oxidase Homolog D (RBOHD) by H2S activates it, leading to ROS formation and inhibition of DES1. See text for additional details.
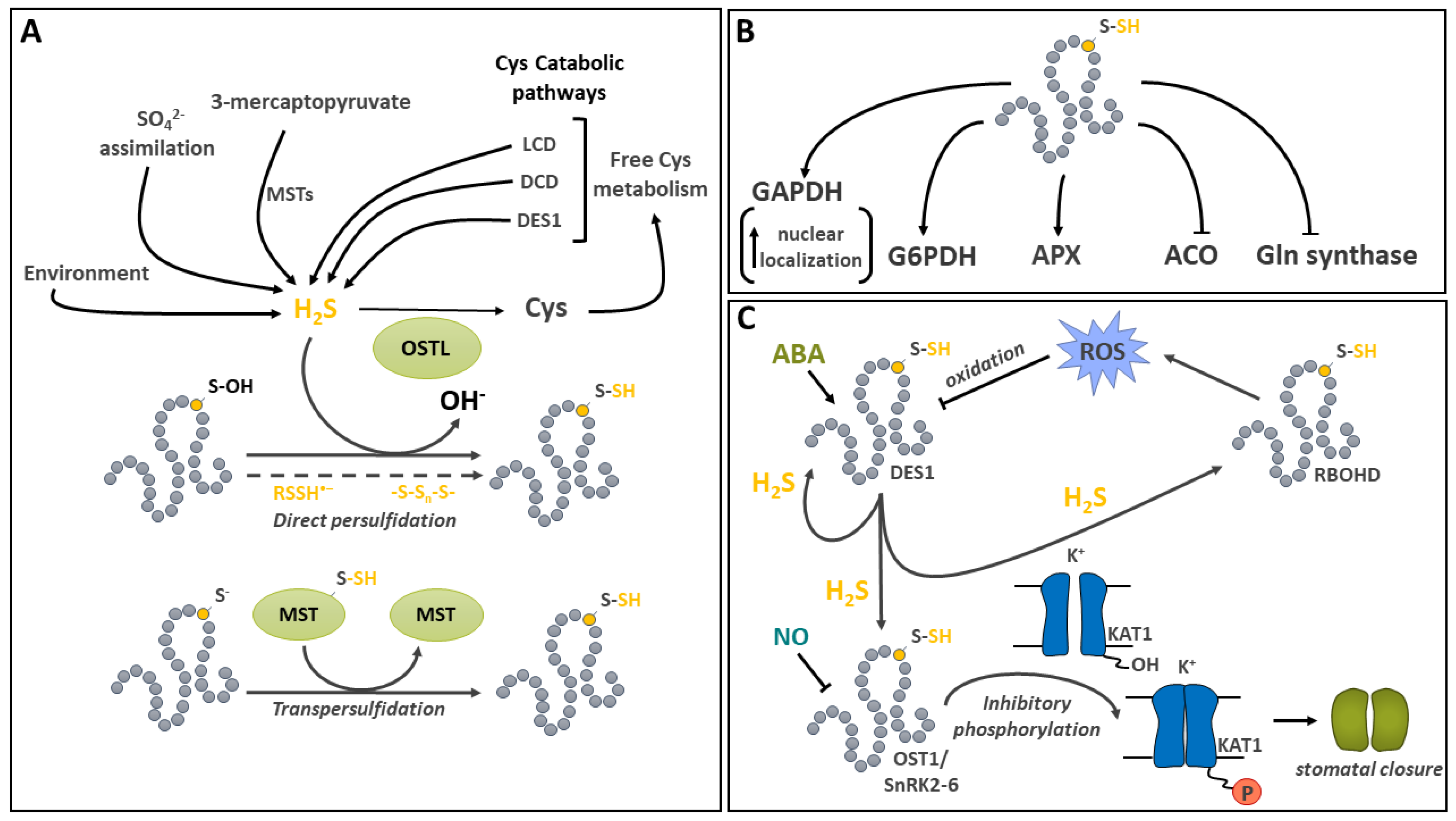
Figure 6.
Protein S-cyanylation in plants. The main pathways involved in the generation of HCN are represented. HCN can also be taken from the environment. The reaction of β-cyanoalanine synthase (CAS) consumes HCN in plants. S-cyanylation (purple symbols) occurs mainly on Cys residues (yellow dots) engaged in disulfide bonds. Among the few targets of S-cyanylation identified in plants, the glycolytic enzyme Enolase 2 (ENO2) was shown to be activated by the modification. Other targets were identified, and some are involved in pathways leading to ethylene synthesis. As indicated by question marks, many effects of S-cyanylation are still largely unknown. See text for additional details.
Figure 6.
Protein S-cyanylation in plants. The main pathways involved in the generation of HCN are represented. HCN can also be taken from the environment. The reaction of β-cyanoalanine synthase (CAS) consumes HCN in plants. S-cyanylation (purple symbols) occurs mainly on Cys residues (yellow dots) engaged in disulfide bonds. Among the few targets of S-cyanylation identified in plants, the glycolytic enzyme Enolase 2 (ENO2) was shown to be activated by the modification. Other targets were identified, and some are involved in pathways leading to ethylene synthesis. As indicated by question marks, many effects of S-cyanylation are still largely unknown. See text for additional details.
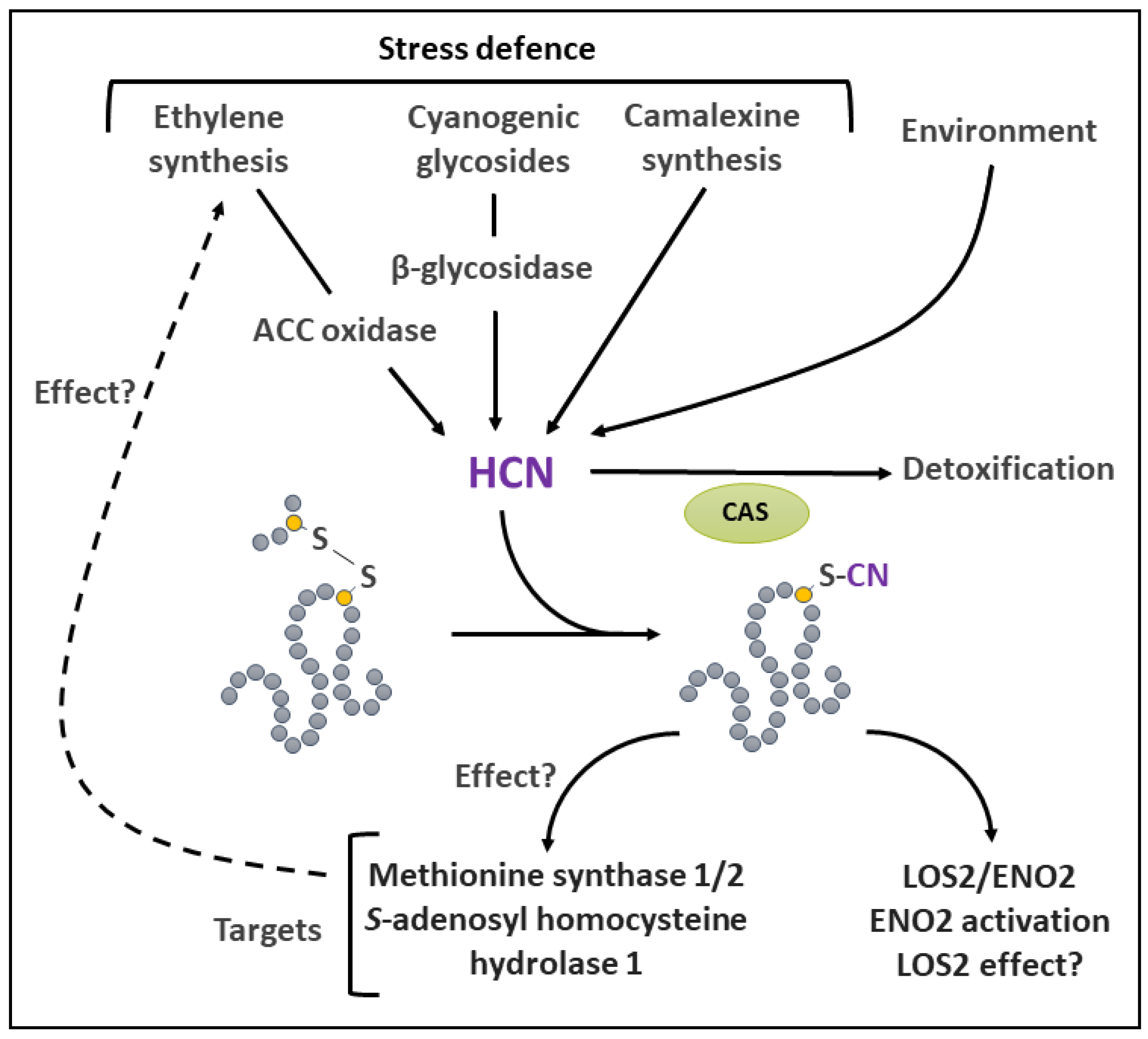
Figure 7.
Protein S-nitrosation in plants. A: The generation of NO in plants and the modification of Cys by direct S-nitrosation. NO can be formed from NO2- by a pathway that involves nitrate reductase (NR) and NO forming nitrite reductase (NOFNiR). It can also be formed by reduction of NO2- by the mitochondrial electron transport chain. NO can be scavenged by phytoglobin. Direct reactions of NO with Cys (yellow dots) thiolate (S-), thyil (●S) or thiol (SH) can lead to S-nitrosation (SNO). B: Protein transnitrosation by S-nitrosoglutahione. S-nitrosoglutahione (GSNO) is a stable form of NO with can be used in transnitrosation of proteins. GSNO is catabolized by GSNO reductase (GSNOR), which is susceptible to inhibition by GSNO and oxidized glutathione (GSSG) and stimulated by H2S. C: Regulation of fructose-1,6-bisphosphatase (FBPase) by reversible S-nitrosation. In its reduced form (SH, top left), FBPase is active. High GSNO promotes the S-nitrosation of FBPase (top right), whereas high glutathione (GSH) reverts it. The S-nitrosated FBPase can undergo the formation of a disulfide bridge (S-S, bottom) that inhibits FBPase activity. S-S reduction by thioredoxin (TRX) occurs in the light and restores FBPase activity. D: Regulation of NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 (NPR1) by S-nitrosation. NPR1 is a receptor for the phytohormone salicylic acid (SA), which transcriptionally regulates systemic acquire resistance through the induction of pathogenesis-related (PR) genes. In absence of pathogen stress, NPR1 is S-nitrosated by GSNO and sequestered in the cytosol in the form of large oligomers with intersubunit S-S. Upon pathogen attack, TRXh5 promotes monomerization of NPR1 by reducing inter-subunit S-S and catalyzing its denitrosation. In turn, reduced monomeric NPR1 can enter the nucleus where it promotes PR genes transcription. See text for additional details.
Figure 7.
Protein S-nitrosation in plants. A: The generation of NO in plants and the modification of Cys by direct S-nitrosation. NO can be formed from NO2- by a pathway that involves nitrate reductase (NR) and NO forming nitrite reductase (NOFNiR). It can also be formed by reduction of NO2- by the mitochondrial electron transport chain. NO can be scavenged by phytoglobin. Direct reactions of NO with Cys (yellow dots) thiolate (S-), thyil (●S) or thiol (SH) can lead to S-nitrosation (SNO). B: Protein transnitrosation by S-nitrosoglutahione. S-nitrosoglutahione (GSNO) is a stable form of NO with can be used in transnitrosation of proteins. GSNO is catabolized by GSNO reductase (GSNOR), which is susceptible to inhibition by GSNO and oxidized glutathione (GSSG) and stimulated by H2S. C: Regulation of fructose-1,6-bisphosphatase (FBPase) by reversible S-nitrosation. In its reduced form (SH, top left), FBPase is active. High GSNO promotes the S-nitrosation of FBPase (top right), whereas high glutathione (GSH) reverts it. The S-nitrosated FBPase can undergo the formation of a disulfide bridge (S-S, bottom) that inhibits FBPase activity. S-S reduction by thioredoxin (TRX) occurs in the light and restores FBPase activity. D: Regulation of NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 (NPR1) by S-nitrosation. NPR1 is a receptor for the phytohormone salicylic acid (SA), which transcriptionally regulates systemic acquire resistance through the induction of pathogenesis-related (PR) genes. In absence of pathogen stress, NPR1 is S-nitrosated by GSNO and sequestered in the cytosol in the form of large oligomers with intersubunit S-S. Upon pathogen attack, TRXh5 promotes monomerization of NPR1 by reducing inter-subunit S-S and catalyzing its denitrosation. In turn, reduced monomeric NPR1 can enter the nucleus where it promotes PR genes transcription. See text for additional details.
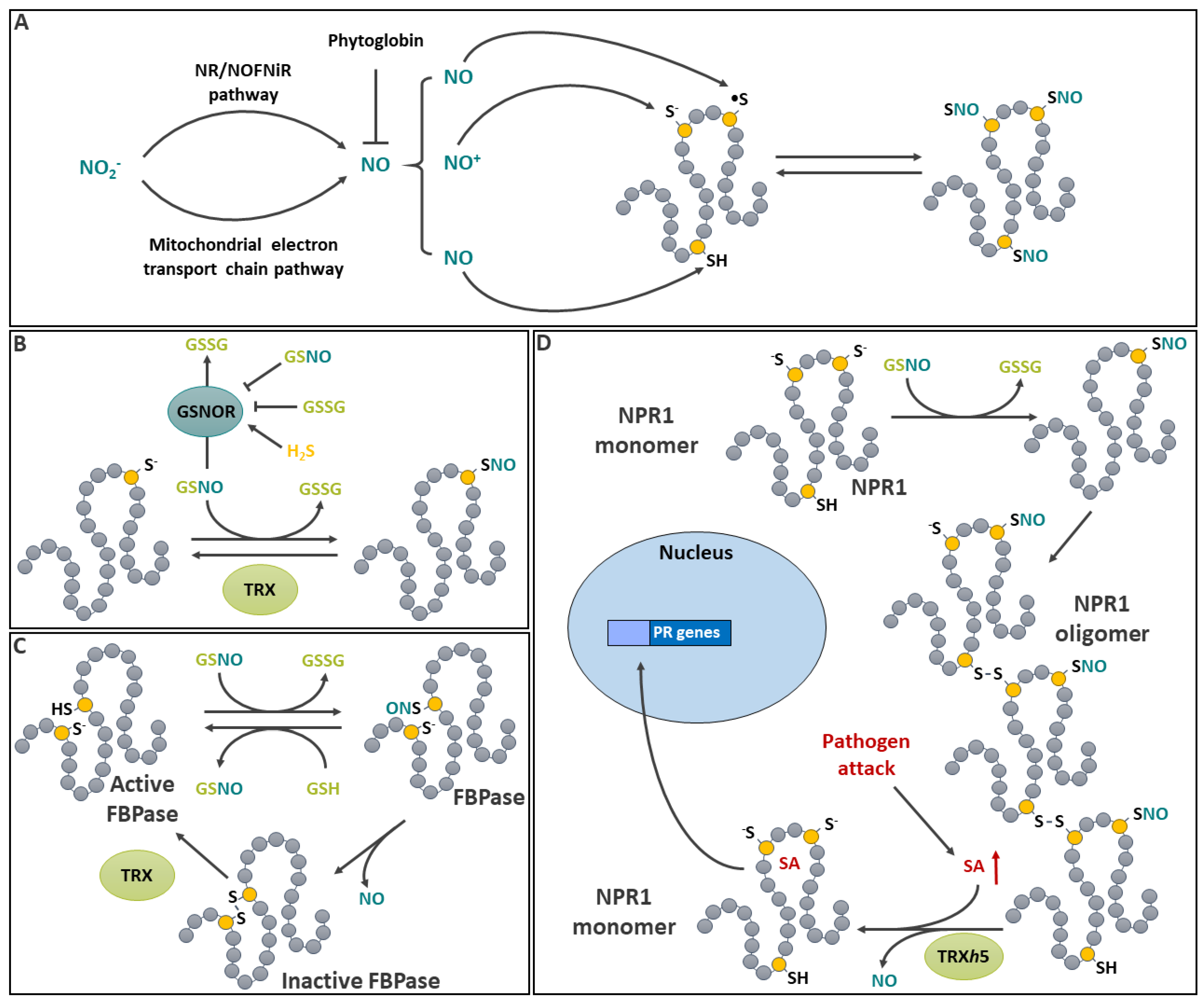
Figure 8.
Protein S-carbonylation in plants. A: generation of reactive carbonyl species (RCS) and their reaction with protein Cys residues. Two main pathways are involved in the generation of RCS. The non-enzymatic pathway results from the attack of membrane-derived unsaturated fatty acids (marked with blue dots) by reactive oxygen species. The enzymatic pathway proceeds via lipoxygenase (LOX). Both pathways can produce a variety of reactive electrophilic α,β-unsaturated carbonyl compounds collectively termed RCSs. Detoxification of RCS can occur and often associated with reduced glutathione (GSH consumption. RCS can modify Cys residues (yellow dots) in thiolate form (-S-), leading to protein S-carbonylation. The modification is widely considered irreversible as the decarbonylation reaction, indicated with a question mark, is still hypothetical in plants. B: Regulation of thioredoxin by S-carbonylation. The RCS cis-(+)-12-oxophytodienoic acid (OPDA) is formed enzymatically in the LOX pathway. Thioredoxin f1 (TRX) is responsible for the reduction of a disulfide bridge on fructose-1,6-bisphosphatase (FBPase) thereby activating the enzyme. In vitro modification of TRX with OPDA (S-OPDAylation) renders TRX unable to activate FBPase. See text for additional details.
Figure 8.
Protein S-carbonylation in plants. A: generation of reactive carbonyl species (RCS) and their reaction with protein Cys residues. Two main pathways are involved in the generation of RCS. The non-enzymatic pathway results from the attack of membrane-derived unsaturated fatty acids (marked with blue dots) by reactive oxygen species. The enzymatic pathway proceeds via lipoxygenase (LOX). Both pathways can produce a variety of reactive electrophilic α,β-unsaturated carbonyl compounds collectively termed RCSs. Detoxification of RCS can occur and often associated with reduced glutathione (GSH consumption. RCS can modify Cys residues (yellow dots) in thiolate form (-S-), leading to protein S-carbonylation. The modification is widely considered irreversible as the decarbonylation reaction, indicated with a question mark, is still hypothetical in plants. B: Regulation of thioredoxin by S-carbonylation. The RCS cis-(+)-12-oxophytodienoic acid (OPDA) is formed enzymatically in the LOX pathway. Thioredoxin f1 (TRX) is responsible for the reduction of a disulfide bridge on fructose-1,6-bisphosphatase (FBPase) thereby activating the enzyme. In vitro modification of TRX with OPDA (S-OPDAylation) renders TRX unable to activate FBPase. See text for additional details.
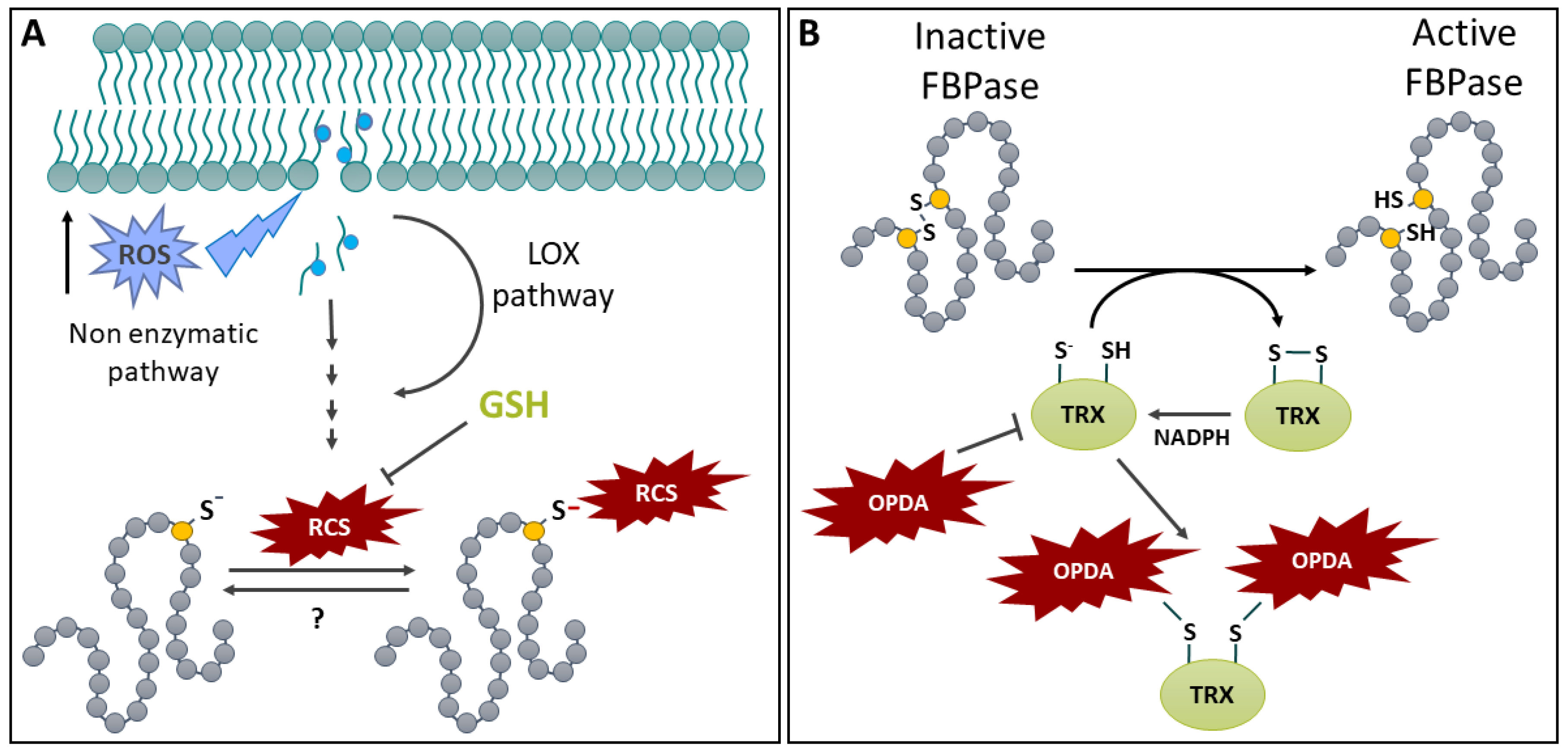
Figure 9.
Protein S-acylation in plants. A: S-acylation and deacylation. Membrane-localized protein S-acyl transferase (PAT) catalyzes the addition of an acyl group (red jagged line) on a Cys (yellow dot) thiol present on a soluble protein. The addition allows the S-acylated protein to become membrane-associated. The reverse reaction (deacylation) is catalyzed by ABHD17-like acyl protein thioesterases (ABAPT) or acyl protein thioesterases (APT), which release the acyl group and solubilize the protein. B: Involvement of S-acylation in the regulation of SLAC1. In the guard cell plasma membrane, the Slow anion channel 1 (SLAC1) is a key element in abscisic acid-dependent stomatal closure. The channel is activated by phosphorylation. A double lipid modification (N-myristoylation on a Gly (green dot) and S-acylation) allow the membrane localization of otherwise soluble Calcineurin B-Like protein 5 (CBL5) and Ca2+-regulated protein kinase CPK6. This allows the recruitment of Calcineurin B-like protein-interacting protein kinase 11 (CIPK11) by CBL5 and the phosphorylation (activation) of SLAC1. C: Involvement of S-acylation in Pi deficiency signalization. The non-specific phospholipase C4 (NPC4) is S-acylated on a C-terminal Cys. Its phospholipase activity allows it to generate diacylglycerol that acts in Pi deficiency signalization. D: Involvement of de-S-acylation of transcription factor NACsa in signal transduction. Under normal conditions, the transcription factor NACsa is sequestered to the plasma membrane by S-acylation. Upon drought, an increase in H2O2 activates monomeric acyl protein thioesterase 1 (APT1) to its tetrameric form. Tetrameric APT1 deacylates NACsa and releases it from the membrane, allowing its translocation to the nucleus where it activates the transcription of Glyoxalase 1 (GLX1), which promotes glutathione build-up acting to inhibit H2O2 accumulation. See text for additional details.
Figure 9.
Protein S-acylation in plants. A: S-acylation and deacylation. Membrane-localized protein S-acyl transferase (PAT) catalyzes the addition of an acyl group (red jagged line) on a Cys (yellow dot) thiol present on a soluble protein. The addition allows the S-acylated protein to become membrane-associated. The reverse reaction (deacylation) is catalyzed by ABHD17-like acyl protein thioesterases (ABAPT) or acyl protein thioesterases (APT), which release the acyl group and solubilize the protein. B: Involvement of S-acylation in the regulation of SLAC1. In the guard cell plasma membrane, the Slow anion channel 1 (SLAC1) is a key element in abscisic acid-dependent stomatal closure. The channel is activated by phosphorylation. A double lipid modification (N-myristoylation on a Gly (green dot) and S-acylation) allow the membrane localization of otherwise soluble Calcineurin B-Like protein 5 (CBL5) and Ca2+-regulated protein kinase CPK6. This allows the recruitment of Calcineurin B-like protein-interacting protein kinase 11 (CIPK11) by CBL5 and the phosphorylation (activation) of SLAC1. C: Involvement of S-acylation in Pi deficiency signalization. The non-specific phospholipase C4 (NPC4) is S-acylated on a C-terminal Cys. Its phospholipase activity allows it to generate diacylglycerol that acts in Pi deficiency signalization. D: Involvement of de-S-acylation of transcription factor NACsa in signal transduction. Under normal conditions, the transcription factor NACsa is sequestered to the plasma membrane by S-acylation. Upon drought, an increase in H2O2 activates monomeric acyl protein thioesterase 1 (APT1) to its tetrameric form. Tetrameric APT1 deacylates NACsa and releases it from the membrane, allowing its translocation to the nucleus where it activates the transcription of Glyoxalase 1 (GLX1), which promotes glutathione build-up acting to inhibit H2O2 accumulation. See text for additional details.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



