
Submitted:
07 August 2024
Posted:
08 August 2024
You are already at the latest version
Abstract
Ewing sarcoma (ES) is an aggressive bone and soft-tissue pediatric cancer. High vitronectin (VN) expression has been associated with poor prognosis in other cancers, and we aimed to determine the utility of this extracellular matrix glycoprotein as a biomarker of aggressiveness in ES. Silk-fibroin plus gelatin-tyramine hydrogels (HGs) were fabricated with and without cross-linked VN and cultivated with A673 and PDX73 ES cell lines for two and three weeks. VN secretion to culture media was assessed using ELISA. Morphometric analysis was applied for phenotypic characterization. VN release to culture media was higher in 3D-models than in monolayer cultures, and intracellular, intercellular and pericluster presence was also observed. A673-HGs showed lower density of clusters but a proportion of larger clusters than PDX73-HGs, which presented low cluster circularity. The cluster density of A673-HGs without added VN was higher than with added VN, and slightly lower in the case of PDX73-HGs. Furthermore, a culture time of three weeks provided no benefits in cluster growth compared to two weeks, especially in A673 -HGs. These advances in 3D modeling and digital quantification pave the way for future studies in ES and other cancers to deepen understanding about intra- and intercellular heterogeneity and anti-adhesion VN therapies.
Keywords:
extracellular matrix
; hydrogels
; digital quantification
; childhood cancer
1. Introduction
Ewing sarcoma (ES) is the second most frequent bone or soft-tissue cancer in children, adolescents, and young adults [1,2]. The exact origin of ES is still under debate, but mesenchymal stem cells (MSC) are currently considered the most likely candidates [3,4,5]. Histologically, ES is characterized by sheets of small, round, and uniformly blue tumor cells with round nuclei and limited cytoplasm. These cells display morphological characteristics indicative of neural differentiation, such as rosette formation [5,6]. Standard diagnostic practice consists of immunohistochemical staining for the membrane markers CD99 and NKX2.2, and evaluation for loss of neural differentiation [2].
ES is well-characterized at the molecular level and is mainly defined by recurring translocations that generate oncogenic gene fusions involving the FET gene family (EWSR1, FUS, TAF15) as 5’ gene fusion partners, with EWSR1 being the common partner in most cases. It also involves members of the ETS gene family of transcription factors as 3’ partners, particularly FLI1 (in about 85% of cases), ERG (around 10%), and occasionally other variants such as ETV1, ETV4, or FEV (less than 1% of cases) [2]. Concurrent chromosomal copy number alterations are also observed, including gains on chromosomes 1q, 8, 12, or 20 and losses on 16q [2,3,7].
The current standard treatment for ES is a multimodal approach consisting of conventional chemotherapy and surgery established through collaborative international trials, which has led to an improved overall survival (OS) rate of approximately 85% for localized disease [6,8]. In contrast, patients with metastatic disease at diagnosis present survival rates of less than 30% [2,6]. A fuller understanding of the prognostic factors that influence treatment outcomes poses a significant clinical challenge, and this raises a need for more in-depth exploration of novel diagnostic and therapeutic targets [5].
The tumor microenvironment (TME) plays a critical role in the effectiveness of cancer treatments, particularly immunotherapy. It can influence tumor behavior and aggressiveness due to the remarkable adaptability of tumor cells and intratumor heterogeneity [9]. Within the TME, the extracellular matrix (ECM) forms a complex network of fibrillar proteins, glycoproteins, and proteoglycans, which act as the scaffold for cellular embedding [10,11]. Cells connect their cytoskeleton to the ECM through receptors such as integrins, thereby generating biophysical forces (traction and compression) known as biotensegrity [12]. This leads to the transmission of biochemical signals via mechanotransduction processes, and ultimately affects gene expression, and cell survival, progression, and migration [13,14,15]. Moreover, mechanotransduction also induces feedback mechanisms that reciprocally influence ECM composition and architecture. This results in clonal selection of the tumor cells best adapted to the niche [16,17,18].
In the context of ES, it is plausible that changes in the mechanical properties of the ECM contribute to the abnormal growth and ability to spread of these cells. Previous studies with ES have shown increased synthesis of ECM proteins in response to activation of the Wnt pathway [19], and TAZ/YAP1 proteins, which are involved in sensing mechanical signals, have also been observed to play a role in ES progression [20,21]. These findings have raised the possibility of developing new therapeutic strategies targeting different players in the TME of bone ES [22].
Among the components of the ECM, vitronectin (VN) is a glycoprotein also present in plasma which is secreted as a monomer or multimer. It contains multiple binding sites to cells (including integrins and other membrane receptors such as uPAR and PAI-1 [23], and to ECM components (fibers and proteoglycans), creating temporary connections and facilitating cell adhesion and migration [23,24,25]. VN has been implicated in tumor progression and metastasis in various types of cancer, including osteosarcoma [26], melanoma [27], hepatocarcinoma [28], breast cancer [29], lung cancer [30] and ovarian cancer [31]. In the childhood cancer neuroblastoma (NB), the presence of VN in biopsy samples has been associated with poor prognosis [32]. VN expression and its relation to aggressiveness have been poorly studied in ES. In a preliminary investigation, our group described VN expression in a subgroup of ES, observing a branching in VN expression reminiscent of track formation, which could facilitate migration and metastasis [33].
Given the rarity of childhood tumors, and in line with the ethical commitment to reducing animal experimentation, other material source options need to be explored for ES research. Three-dimensional (3D) tumor models have become valuable tools offering promising advantages for cancer research [16,34,35]. These biomaterial-based models surpass traditional 2D cultures by reproducing several features of the complex ECM, allowing controlled studies of biotensegrity and intratumor heterogeneity [36]. Moreover, 3D models have served as testing platforms for new drugs [34,35,37]. Despite this, the use of these 3D models, including free- and hydrogel- (HG) based spheroids, scaffold constructs, and flow perfusion systems [38,39] is not yet widespread in ES research.
In previous studies, we used innovative 3D models of NB such as bio-printed HGs with methacrylated gelatin and methacrylated alginate with varying degrees of stiffness, polyethylene glycol HGs with cross-linked VN [10,12,16,22,40], and, more recently, silk fibroin with tyramine-gelatin (sf-GTA)-based HGs, also with and without cross-linked VN [41].
Our aim in this study was to determine whether two different ES cell lines would be able to adapt and grow in sf-GTA 3D models. We also explored the effect of cross-linking VN in 3D models on cell growth, morphology and VN expression and secretion to the ECM of the ES cell lines and compared these results with those in previous 2D cultures. Finally, we assessed potential applications of these HG models for future research.
2. Results
2.1. VN cell Expression and Secretion to Culture Media in 2D Models
Anti-VN staining revealed different VN expression patterns between the A673 and PDX73 cell lines cultured in monolayer in the cytospin area (approx. 2 cm²). We quantified 71,776 cells in the cytospin of the A673 cell line, finding a 12.31 µm cell diameter and 0.68 nuclei/cell ratio. VN expression was low (5.13% of positive cells) and was characterized by a heterogenous and granular staining pattern with localization in cell membranes in some cells (Figure 1A). In contrast, we quantified 131,821 cells with a 13.01 µm cell diameter and 0.73 nuclei/cell ratio in the cytospin of the PDX73 cell line. These cytospin showed positivity in many cells (76.67%), with weak intensity and more homogeneous staining, around almost the entire cell membrane (Figure 1B).
VN release to culture media of the two Ewing sarcoma cell lines was detected by applying ELISA. In correlation with our immunohistochemical cell expression findings, the A673 cell line cultured in monolayer showed a lower VN release to culture media per million of cells (1.2 ng/mL) compared to the PDX73 cell line (1.9 ng/mL, Figure 1C), although the cell count in 2D cultures was higher for A673 than for PDX73 (50 million and 20 million, respectively).
2.2. VN Cell Expression and Secretion to the ECM in 3D Models
As already noted in 2D, in 3D cultures different VN expression patterns were observed between the two ES cell lines. Subjective evaluation of VN immunostainings revealed that in A673 HGs without added VN at 2 weeks of culture, cells exhibited high expression intensity (+++), and that VN was primarily stored intracellularly and to a lesser extent in the pericellular region [territorial VN] (Figure 2 IA). At 3 weeks with the same scaffold composition and in HGs with added VN (Figure 2 IB, C and D), the intensity was moderate (++) and more prevalent in the intercellular zone (interterritorial VN), and adjacent to the edge between the cluster cells and the scaffold.
PDX73 cells at 2 and 3 weeks without added VN (Figure 2 IE and 2 IG) exhibited moderate (++) interterritorial staining intensity and particularly pericluster expression, where a marked VN pattern could be observed close to the cluster contour (as a spiked crown), especially at 2 weeks. In HGs with added VN, there was low-intensity interterritorial VN (+) without a clearly discernible crown at 2 weeks (Figure 2 IF). Curiously, at 3 weeks (Figure 2 IH), in HGs of the same composition there was almost no intra- or interterritorial VN expression; however, a pale halo was visible around the clusters, lighter than the rest of the HG stain, with irregular shape and sometimes much thicker on one side of the cluster than the other.
VN release to 3D culture media was null in control HGs (without cells) with added VN, after both 2 and 3 weeks of culture. However, culture media of both ES cell lines grown in 3D showed positive VN detection (Figure 2 II). Overall, VN was higher in 3D cultures than in 2D of both cell lines (p = 0.044, Figure S1).
VN crosslinking in HGs did not substantially affect VN secretion by ES cells, which remained elevated despite the large quantities present in the scaffold. Longer culture times (3 weeks) in HGs with VN cross-linking decreased cell growth in the A673 cell line but increased VN release, coinciding with the lower amount of intracellularly retained VN observed in immunostaining. Longer culture time slightly increased PDX73 cell numbers, and VN secretion to culture media remained elevated under all the 3D conditions studied.
2.3. Digital Analisis and Morphometric Parameters of Clusters and Cells of 3D Models
Cluster density, cell density by cluster size, cluster area and circularity of clusters were measured by digital analysis and validated subjectively by experts.
Subjective observation revealed that at 2 weeks of culture, in approximately 95% of clusters of A673 cell line grown in HGs without and with added VN, the cells displayed large nuclei with varying hematoxylin intensity and evident cytoplasm (medium to high eosinophilic), with reduced intercellular spaces (Figure 3A and B). In contrast, after 3 weeks of culture, in HGs without and with added VN (Figure 3C and D) approximately 80% of clusters contained fewer cells, each with small nuclei, reduced cytoplasm, and wide intercellular spaces. All PDX73 clusters showed similar cell morphology regardless of culture time or scaffold composition. These cells had bigger nuclei than in the A673 line, with great variability in hematoxylin intensity, creating the so-called dark and light cells. The cytoplasm was generally thin, with limited intercellular spaces, and minimal eosinophilic material (Figure 3E to 3H).
We used data from a total of 8665 clusters and 17,5227 cells from the 23 H&E images (Table S1) for digital analysis. Note that there was variability in the number and size of clusters between HGs replicates (Figure S3 A and S3 B). The median area of the HGs sections was 8.2 cm2 (between 3.99 cm2 and 20.85 cm2). Irrespective of scaffold composition and time of culture, the A673 cell line exhibited significant heterogeneity in the cellular features in clusters and a larger cluster size compared to the PDX73 cell line, which showed uniform cellular characteristics with smaller cluster sizes. Specifically, in the A673 cultures, we detected 727 clusters in the three HGs replicates without added VN at 2 weeks (median cluster area= 1358.7 µm2), 524 clusters in the three HGs with added VN at 2 weeks (median cluster area= 1385.7 µm2), 269 clusters in HGs without added VN at 3 weeks (median cluster area= 719.21 µm2) and 483 clusters in HGs with added VN at 3 weeks (median cluster area= 925.085 µm2). In the case of PDX73 cell line, 1709 clusters were found in HGs without added VN at 2 weeks (median cluster area= 595.43 µm2), 1708 clusters in HGs with added VN at 2 weeks (median cluster area= 437.12 µm2), 1505 clusters in HGs with added VN at 3 weeks (median cluster area= 457.49 µm2) and 1740 clusters in HGs without added VN at 3 weeks (median cluster area= 488.44 µm2).
2.3.1. Cluster Density
The A673 showed lower cluster density (median=0.028 cluster/nm2) than the PDX73 cell line (median=0.051 cluster/nm2, p < 0.0001, Figure S2). Specifically, cluster density of the A673 cell line (Figure 4A) was higher when cultured in HGs without added VN than with added VN (median=0.0302 cluster/nm2 vs median =0.0263 cluster/nm2, respectively). Moreover, we detected a slight increase in cluster density with time of culture (median = 0.028 clusters/nm² at 2 weeks vs. median = 0.0325 clusters/nm² at 3 weeks in HGs without added VN, and median = 0.0215 clusters/nm² at 2 weeks vs. median = 0.0234 clusters/nm² at 3 weeks in HGs with added VN).
The PDX73 cell line (Figure 4B) also showed notably higher cluster density in HGs without added VN than in HGs with VN cross-linking at 2 weeks (median=0.060 cluster/nm2 vs. median=0.046 cluster/nm2, respectively). At the longest culture time, however, the reverse situation was observed. Cluster density was lower in HGs without added VN than in the HGs with added VN, reflecting a better adaptation to these scaffolds over time (median=0.048 cluster/nm2 at vs. median=0.051 cluster/nm, respectively).
2.3.1.1. Cell Density by Cluster Size
Cluster density was also calculated after classifying clusters by cell count following the established criteria (Table 1, Material and Methods section).
A673 showed higher density in the largest clusters (big, giant and huge) than PDX73, which exhibited more density for the smallest (small and medium) ones (Figure 5). Specifically, cell line A673 (Figure 5, top graphs) showed no remarkable differences in any size category with respect to VN cross-linking in the scaffolds. Regarding time, we observed higher cluster density in the largest clusters at 2 than at 3 weeks of culture. However, small clusters increased their density with time. Classifying the clusters of PDX73 HGs by size (Figure 5, bottom graphs), we found only subtle differences between the two scaffold compositions and the two culture times studied.
2.3.2. Cluster Areas
The high variability in A673 cell line cluster areas (Figure 6, top graphs), despite previous classification by cell count, is partly due to cell size and intercellular spaces (Figure 3). Briefly, the small and medium clusters of A673 in HGs without added VN at 3 weeks showed a significantly lower area than the scaffolds with added VN (p < 0.0001 for small and p = 0.017 for medium clusters), the latter ones also showing an increase in small cluster areas with time (p = 0.0010). Among the big clusters, the HGs with added VN showed a significant area decrease with time (p= 0.0051). Finally, the giant clusters, as opposed to the rest of the cluster sizes, had a bigger area in HGs without than with added VN at 3 weeks (p = 0.021).
Validated subjectively, the PDX73 cell line (Figure 6, bottom graphs) showed more stable cluster areas between conditions and slightly smaller cluster area sizes than the A673 cell line (Figure 3). However, the small clusters showed a decrease in area in the non-VN-added scaffolds over time (p = 0.0008), while in the medium clusters, the area was significantly lower at 2 than at 3 weeks within VN-added HGs (p = 0.0124). Data regarding the cluster area of the two ES cell lines is summarized in Tables S2 and S3.
2.3.3. Circularity of Clusters
The A673 cell line (Figure 7, top graphs), showed higher circularity than PDX73 for all cluster size categories. The circularity of the small clusters was lower in HGs without added VN than in VN-added scaffolds, at both timepoints, 2 weeks (p < 0.001) and 3 weeks (p < 0.001). The medium clusters showed an irregular shape at 3 weeks compared to 2 weeks in non-added VN HGs (p-value = 0.009) and when compared with the VN-added HGs 3 weeks (p-value < 0.001). The largest clusters showed circularities closer to 1 in all the A673 culture conditions studied (Tables S4 and S5).
High irregularity was detected, mainly in all cluster sizes in 3D models of PDX73 (Figure 7, bottom graphs). Small, medium and large clusters showed a similar pattern. They presented a similar or lower irregularity at 3 weeks of culture than at 2 weeks, which varied depending on the composition of HGs (Figure 7). The largest clusters showed more reduced circularity than the smaller ones. Particularly notable was a higher circularity of giant clusters in non-VN-added scaffolds at 3 weeks compared to the other conditions. Tables S6 and S7 show comparison of circularity of PDX73 clusters between cluster size categories in each 3D culture condition.
3. Discussion
ES is considered a cancer type with poor prognosis. Its clinical characteristics often result in late diagnosis after spread to metastatic sites, thus limiting effective treatment [42]. Chemotherapy resistance is closely linked to specific TME–cell interactions in ES [43], and the influence of biotensegrity and mechanical alterations of the ECM facilitates cancer cell proliferation and dissemination [9]. Previous studies have shown that primary tumor stiffness is greater than in normal tissues and has been linked to cancer progression and metastasis [32,44,45]. This has prompted researchers to develop alternatives for new therapies, including mechanotherapies, aimed at modulating mechanical forces; for example, by disrupting communication between tumor cells and ECM components, and thus inhibiting migration[46]. Simplified replication of the TME through 3D models is essential to improve our understanding of tumor aggressiveness, progression and migration, and can potentially be used to develop new diagnostic biomarkers and test therapeutic strategies. However, 3D models are as yet underutilized in ES research; the complexity, cost, and technical challenges of developing these systems represent obstacles to scalability and broader adoption [38,39].
Our research is focused on alternative 3D models that are simple and quick to manufacture in terms of composition and structure. We designed HG models comprising sf-GTA, with or without VN cross-linking. This glycoprotein was chosen for its overexpression and implication in several cancer types associated with tumor aggressiveness [26,28,29]. The constructed HGs allowed us to simulate 3D tumors with two mechanical forces, induced by the scaffolds and by the cell dynamic related to the VN amount added and/or secreted by cells. Since VN cross-linking increases matrix stiffness, promoting migration and proliferation [12,47], we hypothesized that cross-linking VN to the HGs would activate more mechanotransduction pathways in cells through ECM–cell interaction [13,49].
Cell growth and morphology in the 2D and 3D cultures were monitored and evaluated using digital pathology analyses. The integration of artificial intelligence (AI) applied to digital pathology has proven crucial in oncology research [48], providing new approaches for preclinical studies which enable us to evaluate not only cell features, but also the composition and architecture of the ECM [49], through which new biomarkers can be assessed. This integration optimizes analysis and reduces the time pathologists need to perform more precise evaluations [50], thereby enhancing diagnostic, prognostic, and predictive clinical decision-making in ES and other tumors [40].
When we scaled from the 2D cultures, we observed that the biocompatibility and tensegrity of the constructed ECM provided a suitable environment for the two ES cell lines, A673 and PDX73, to grow and lead cluster formation and VN secretion.
Positive detection of VN secreted to 2D and 3D culture media indicated that both cell lines synthesized this glycoprotein in all assayed conditions, even in the presence of cross-linked VN in the HG [51,52]. Nonetheless, VN release was higher in 3D than 2D cultures, reflecting the importance of cell growth in a more biomimetic 3D environment.
The lower amount of VN secreted by A673 than PDX73 cells in 2D correlates with less adaptation to 3D. A673 is an established cell line (1973) that has undergone numerous 2D culture passages; the 2D cultured cells express VN with high heterogeneity, which may be due to the usual lack of ECM experiments in those culture conditions. The PDX73 cell line, a short-term culture (established in 2015) with no more than 13 passages in 2D culture, may have better retained the characteristics of the original tumor. In this cell line, we validated previously described evidence that VN is more uniformly distributed on the cell membrane due to a more complex environment in which focal adhesions are continuously formed [17,34]. How this VN secretion by 3D cultured ES cells may be reflected in the blood plasma of patients with these tumors is an interesting future line of study. Elevated VN levels have previously been described in plasma (and other fluids) of poor prognosis patients with glioma[53], melanoma[27], breast[54], ovarian and endometrial cancer[55], and in children with Hodgkin’s lymphoma[56], acute lymphoblastic leukemia[57] and NB[58]. The use of circulating biomarkers in liquid biopsy is becoming increasingly widespread due to its simplicity and speed, and they play an especially important role in cancers such as ES to avoid invasive biopsies in children and because of the high intratumoral heterogeneity of these tumors, as is clearly reflected in the 3D models presented here.
Although both cell lines exhibited the phenotypic trait of small, round and blue cells of bone and soft tissue sarcomas[2], 2D and 3D cultures analysis with anti-VN and H&E stains revealed differences in their morphology and biological behavior. Taking all data together, A673 cell line showed lower cluster density but larger cluster sizes than PDX73 cells. However, our results revealed that VN presence in scaffolds had a differential effect on cluster proliferation in both A673 and PDX73 cell lines. The decreased cluster density of A673 cells in the presence of VN could indicate that it exerts an inhibitory influence on cell adaptation and growth [40]. However, the slight increase in cluster density observed in the HG with added VN cultured with the PDX73 cell line during the third week, contrary to what happens without added VN, suggests that this cell line may have specific adaptive or resistant mechanisms allowing a rebound in proliferation in VN presence [16,33]. These findings reflect a high variability in cell adaptation and behavior among cell lines. They can help to optimize culture conditions and better understand cellular interactions in these 3D environments.
Regarding the culture times studied, we observed that extending the time to 3 weeks produced smaller clusters of A673 cells in HGs, with large spaces between them and pre-apoptotic like features, leading to loss of cell–cell and cell–HG adhesion. When evaluating density and area by cluster size, the A673 cell line showed differential behavior in the proliferation of large versus small clusters over time, as well as a marked reduction in cluster areas at 3 weeks independent of VN addition that could also be attributed to cell stress and death. Increased VN secretion has been associated with the bodily response to tissue damage or inflammation in studies with hepatic tissues, indicating that cells are trying to survive [59]. The higher VN release to culture media at 3 weeks compared to 2 weeks could be associated with this cell response to stress or to cell lysis. On the other hand, the relative stability in cluster density and area of the PDX73 cells along time, and its morphological observation in H&E indicate a better adaptation to the 3D models.
Cluster circularity analysis yielded interesting data regarding cellular migration. In the A673 cell line, clusters were round with few cell protrusions (circularity close to 1), especially in HGs with cross-linked VN, suggesting that this glycoprotein probably does not facilitate intercellular adhesion for easy migration of this cell line [60]. Evaluating PDX73 clusters, their low circularity, together with the VN detected in culture media over time, and the pericluster crown formation with interterritorial VN in HGs without added VN, may be related to cell migration [59,61]. Moreover, when PDX73 was cultivated in HGs with added VN, low circularity and VN secretion was maintained, but we observed irregular pale halo formations around the clusters, lighter in anti-VN stain than the rest of the VN-HGs, which was particularly notable at 3 weeks. This finding could be linked to scaffold degradation by ES cells, suggesting that they are preparing genetic and epigenetic pathways to control their environment and promote cell migration [40,61]. This also supports the previously described role of VN in cell migration, as it leads to cell attachment to and detachment from the ECM through its multiple binding sites. In other cancers, such as lung adenocarcinoma or ovarian carcinoma, VN has been shown to become a potent pro-migratory factor in plasma and other fluids, which when released from its inhibitory complex with fibrinogen facilitates cancer cell escape from the tumor, spread through the blood stream and lymphatic system and subsequent metastasis to body cavities with low levels of fibrinogen [52,62]. Interesting future lines of study include further analysis of cluster and cell movement in our 3D models with and without added VN, also compared with cell VN secretion, to elucidate whether this pro-migratory capacity also affects ES cells.
4. Conclusions
In this study, we successfully cultivated two ES cell lines in simple, economical and quick-to-manufacture 3D models. The differing response between A673 and PDX73 cell lines in 3D sf-GTA models with and without VN cross-linking highlights the importance of considering the specific characteristics of each cell line, as a reflection of inter- and intratumor heterogeneity in patient samples. Both cell lines secreted VN, and cross-linking VN in HGs appeared to have a positive effect on cell growth in the PDX73 cell line. However, culture time longer than 3 weeks seems to have been detrimental, particularly in the case of the A673 cell line. In this regard, studies with more cell lines (especially PDX cells) are ongoing to analyze whether added VN could influence tumor aggressiveness and thus be proposed as a biomarker. The 3D models presented in this study will allow significant progress in our understanding of ES at both molecular and clinical levels. Moreover, VN detection in liquid biopsies should be explored as a potential prognosis biomarker in ES patients.
Our findings could also have significant implications for designing therapeutic strategies (mechanotherapies) that modulate ECM–cell interactions; i.e., by blocking components such as VN in ES treatment [63,64,65]. The well-known A673 cell line could be ideal for studies of basic molecular mechanisms and experiments that require stability and easy manipulation. In contrast, the PDX73 cell line would be more suitable for preclinical studies, due to its closer fidelity to the patient’s original tumor, making it valuable for assessing treatment efficacy and developing personalized therapeutic strategies. A broader spectrum of cellular behaviors in different scaffolds composition along time would be useful to test and predict possible responses to therapies, especially mechanotherapies, in these 3D models.
We have provided a robust platform for investigating how ECM impacts tumor cell survival, progression and migration. We have modulated its stiffness to study a specific element of the tumor ECM, VN, opening the possibility of its use to study other proteins of interest and other tumor types. As a new drug-testing platform, this approach paves the way for developing more effective and personalized therapeutic strategies.
5. Materials and Methods
5.1. Monolayer Cultures (2D)
The cell lines used were commercial ES cell line A673 (derived from the muscle cancer cells of a 15-year-old girl in 1973, ATCC, Masassas, VA, USA) and ES cell line PDX73 (patient-derived xenograft taken from the metastatic costal tumor of a 16-year-old girl in 2015, Hospital del Rocio, Sevilla). Gibco™ DMEM medium (Gibco, Life Technologies, Waltham, MA, USA) and Ham’s F-10 Nutrient Mix, GlutaMAX medium (Gibco, Life Technologies, Waltham, MA, USA) were used for A673 and PDX73 cultures, respectively, supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. Cell cultures were maintained at 37°C with 5% CO2 in T75 cell culture flasks (plus 0.1% gelatin coated in the case of PDX73 to facilitate cell adhesion). The media was changed every 2–3 days, and cultures were trypsinized when confluence was reached.
5.2. Hydrogel Cultures (3D)
Following the previously described protocol [58,66] 3D HGs were prepared from a solution of lyophilized GTA (Sigma Aldrich, Merck, USA) and sf (Sigma Aldrich, Merck, USA) at a ratio of 25:75, respectively, dissolved in IMDM (4% w/v). VN (PrepoTech, USA) was resuspended in dPBS and combined with the sf-GTA mix with a final concentration of 0.4 mg/mL for the HGs with added VN; the same amount of dPBS alone was added for the HGs without added VN. Horseradish peroxidase (20 U/mL) was also added to the mix. Cells were trypsinized from the previous 2D culture, quantified with trypan blue using a BioRad TC20 Automated Cell Counter, and resuspended in the HG mix to obtain an amount of 1.25x105 commercial cells or 2.5x105 PDX cells per HG. To start the polymerization process, 2 µL of hydrogen peroxide (0.01%) was placed in the center of each well in a 24-well plate and mixed with aliquots of 60 μL of the mix solution. HGs were maintained in the incubator for 1 hour until complete polymerization, then 2 mL of supplemented culture media (same that for the 2D cultures) was added over each HG. The HG culture media was replaced every 3 days during the 2 and 3 weeks that the 3D models were maintained. Three HG replicates of each composition and culture time were cultivated with each cell line. The waste medium was frozen (-80 °C) and preserved for ELISA.
5.3.2. D and 3D Sample Preparation for Digital Analysis
Once the monolayer culture of each line reached cell confluence, a 250 μL aliquot containing approximately 100,000 cells was taken to perform the cytospin procedure. Slides were kept in ethanol until staining. HGs were formalin-fixed and paraffin-embedded (FFPE) after 2 and 3 weeks of culture, then cut into 3 µm sections. Both cytospin and HGs section samples were stained with H&E and immunostained with anti-VN rabbit monoclonal antibody (EP873Y, Clone; ab45139, Abcam, Cambridge, MA, USA) diluted at 1:100, using OptiView Amplification Kit (Ventana Medical Systems Inc., Tucson, EE.UU.) in the BenchMark XT automated slide staining system (Ventana Medical Systems Inc., Tucson, USA). The mounted plates were scanned with Ventana iScanHT (Roche) at 20X and the resulting images were saved in TIF format. Cell and cluster quantity and morphology in H&E and anti-VN stainings from 2D and 3D models were subjectively evaluated by expert observers, including the reference pathologist of the group (SN), who also validated the objective digital analyses. As previously mentioned, the nomenclature for VN expression was adopted from Burgos-Panadero’s research on NB [32]. It was classified into two distinct categories: strong VN intensity inside cells or immediately adjacent to the cell membrane was termed territorial VN, and weak to moderate VN intensity in intercellular location was interterritorial VN. A third category, named pericluster VN, was established to designate the strong VN intensity detected at the edge of the clusters, just between the cells and the scaffold, as well as in the scaffold itself close to the clusters forming as a crown. Some clusters of HGs with added VN showed reduced VN intensity in the scaffold around them (a weak stain) that we refer to as a pale halo.
5.4. Digital Microscopic Analyses
QuPhatTH and its extensions was used for all digital analyses [67]. A StarDist script was adjusted to detect VN positivity in cytospin of 2D cultures, based on the diaminobenzidine mean intensity threshold in cytoplasm. For digital analyses of 3D models, a semi-automatic method was developed to detect HG area, clusters and cell counts in the scanned H&E-stained slides (Supplementary data. Appendix A). In total, 11 H&E stain images were analyzed for A673 (one image was excluded from the study because the HG was broken into multiple fragments during FFPE processing and the holes and folds precluded correct analysis) and 12 for PDX73. To evaluate cluster density, the number of clusters divided by HG area of each replicate was analyzed. We used quartile analysis of the cell count in each cluster of the ES cell lines, and a consensus was reached for cluster size classification (Table 1). Cluster area and circularity were parameters provided by QuPathTH. Clusters with a more circular shape have a circularity value closer to 1, meaning that less cell protrusions are present. Therefore, an irregular shape will have reduced circularity.
5.5. Detection of VN in Culture Media
For 2D cultures, media (8 mL) was collected from the confluent culture in a T75 cell culture flask after quantifying the culture cells with BioRad TC20. The media from 2D cultures was concentrated 16-fold with SpeedVac™. In the 3D cultures, culture media from two HG replicates (4 mL) of each composition and time of culture was mixed and concentrated 8-fold. With this step, we reduced the variability intrinsic to HG replicates and achieved the minimum VN level for ELISA detection (assay sensitivity of 15.19 pg/mL). As culture media was collected with each change (every 3 days), we analyzed VN levels with ELISA at several time-points. However, the cell count was estimated in both HGs at 2 weeks and 3 weeks time-points (when HGs were FFPE) by digital analysis in 3 µm sections, then was scaled to the approximate volume of HGs (28 mm3) and summed. VN level detected by ELISA in the 2D and 3D culture media was divided by concentration factor and by cell count to determine VN secretion in ng/mL per million cells. The amount of VN in culture media was determined by sandwich colorimetric enzyme-linked immunosorbent assay (ELISA, Novus Biologicals, USA) according to the manufacturer’s instructions. Antibody reactions, absorbance reading and results calculation steps were performed using a Triturus automated analyzer (Grífols, Barcelona).
5.6. Statistical Analysis
The database used was obtained from QuPathTH. Statistical analyses were performed with R studio version 4.2.1 (Foundation for Statistical Computing, Vienna, Austria), using the ggplot2 package version 3.5.1 (Wickham, 2016) to generate figures, and GraphPad Prism 8 (Graphpad Software, Boston, USA). Data were analyzed for normality by Shapiro-Wilk test, then ANOVA (Kruskal-Wallis method with Dun´s multiple comparison) was used for nonparametric comparisons of cluster density, area, and cluster circularity between cell lines, culture times and HG compositions. Differences were considered if the p-value was <0.05.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author’s Note:
Readers who wish to obtain additional data or details related to this study are encouraged to contact the corresponding authors.
Author Contributions
AL-C and KP-H carried out the in vitro experiments and culture media collection and preparation, and wrote the original draft of the manuscript. DA-L performed the ELISAs. JD, CS-A and EdA provided A673 and PDX73 cells. KP-H did the digital analysis, IV-M and SG-A helped in scripts elaboration. SN validated the morphometrical analyses, reviewed the paper and suggested draft changes. RN elaborated the concept, designed the study, acquired funding, reviewed the paper, and suggested draft changes. All authors read and approved the final manuscript.
Funding
This study was supported by Fundación CRIS Contra el Cáncer (2023/188), Institute of Health Carlos III, Madrid/ERDF (PI20/01107) and CIBERONC (CB16/12/00484 and CB16/12/00361). IV-M was supported by Ministry of Science, Innovation and Universities of Spain (FPU20/05344).
Institutional Review Board Statement
The study “An in vivo platform for translational drug development in cancer” was conducted according to the guidelines of the Biomedical Research Law 14/2007 and approved by the Ethics Research Committee of Hospital Universitario Virgen del Rocio (20 January 2014).
Acknowledgments
The authors would like to thank Elisa Alonso for technical support, and Kathryn for English correction.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Newman, E.A.; Abdessalam, S.; Aldrink, J.H.; Austin, M.; Heaton, T.E.; Bruny, J.; Ehrlich, P.; Dasgupta, R.; Baertschiger, R.M.; Lautz, T.B.; et al. Update on Neuroblastoma. J Pediatr Surg 2019, 54, 383–389. [CrossRef]
- Grünewald, T.G.P.; Cidre-Aranaz, F.; Surdez, D.; Tomazou, E.M.; de Álava, E.; Kovar, H.; Sorensen, P.H.; Delattre, O.; Dirksen, U. Ewing Sarcoma. Nat Rev Dis Primers 2018, 4, 5. [CrossRef]
- Gargallo, P.; Yáñez, Y.; Juan, A.; Segura, V.; Balaguer, J.; Torres, B.; Oltra, S.; Castel, V.; Cañete, A. Review: Ewing Sarcoma Predisposition. Pathology & Oncology Research 2020, 26, 2057–2066. [CrossRef]
- Hameed, M. Small Round Cell Tumors of Bone. Arch Pathol Lab Med 2007, 131, 192-204. [CrossRef]
- Ozaki, T. Diagnosis and Treatment of Ewing Sarcoma of the Bone: A Review Article. Journal of Orthopaedic Science 2015, 20, 250–263. [CrossRef]
- Zöllner, S.K.; Amatruda, J.F.; Bauer, S.; Collaud, S.; de Álava, E.; DuBois, S.G.; Hardes, J.; Hartmann, W.; Kovar, H.; Metzler, M.; et al. Ewing Sarcoma—Diagnosis, Treatment, Clinical Challenges and Future Perspectives. J Clin Med 2021, 10, 1685. [CrossRef]
- Eaton, B.R.; Claude, L.; Indelicato, D.J.; Vatner, R.; Yeh, B.; Schwarz, R.; Laack, N. Ewing Sarcoma. Pediatr Blood Cancer 2021, 68. [CrossRef]
- Setty, B.A.; Gikandi, A.; DuBois, S.G. Ewing Sarcoma Drug Therapy: Current Standard of Care and Emerging Agents. Pediatric Drugs 2023, 25, 389–397. [CrossRef]
- Sanegre, S.; Lucantoni, F.; Burgos-Panadero, R.; de La Cruz-Merino, L.; Noguera, R.; Álvaro Naranjo, T. Integrating the Tumor Microenvironment into Cancer Therapy. Cancers (Basel) 2020, 12, 1677. [CrossRef]
- Burgos-Panadero, R.; Lucantoni, F.; Gamero-Sandemetrio, E.; Cruz-Merino, L. de la; Álvaro, T.; Noguera, R. The Tumour Microenvironment as an Integrated Framework to Understand Cancer Biology. Cancer Lett 2019, 461, 112–122. [CrossRef]
- Chen, F.; Zhuang, X.; Lin, L.; Yu, P.; Wang, Y.; Shi, Y.; Hu, G.; Sun, Y. New Horizons in Tumor Microenvironment Biology: Challenges and Opportunities. BMC Med 2015, 13, 45. [CrossRef]
- Noguera, R.; Nieto, O.A.; Tadeo, I.; Fariñas, F.; Álvaro, T. Extracellular Matrix, Biotensegrity and Tumor Microenvironment. An Update and Overview. Histol Histopathol 2012, 27, 693–705. [CrossRef]
- Pickup, M.W.; Mouw, J.K.; Weaver, V.M. The Extracellular Matrix Modulates the Hallmarks of Cancer. EMBO Rep 2014, 15, 1243–1253. [CrossRef]
- Elgundi, Z.; Papanicolaou, M.; Major, G.; Cox, T.R.; Melrose, J.; Whitelock, J.M.; Farrugia, B.L. Cancer Metastasis: The Role of the Extracellular Matrix and the Heparan Sulfate Proteoglycan Perlecan. Front Oncol 2020, 9. [CrossRef]
- Discher, D.E.; Janmey, P.; Wang, Y.L. Tissue Cells Feel and Respond to the Stiffness of Their Substrate. Science 2005, 310, 1139–1143. [CrossRef]
- López-Carrasco, A.; Martín-Vañó, S.; Burgos-Panadero, R.; Monferrer, E.; Berbegall, A.P.; Fernández-Blanco, B.; Navarro, S.; Noguera, R. Impact of Extracellular Matrix Stiffness on Genomic Heterogeneity in MYCN-Amplified Neuroblastoma Cell Line. Journal of Experimental & Clinical Cancer Research 2020, 39, 226. [CrossRef]
- Ben-David, U.; Ha, G.; Tseng, Y.-Y.; Greenwald, N.F.; Oh, C.; Shih, J.; McFarland, J.M.; Wong, B.; Boehm, J.S.; Beroukhim, R.; et al. Patient-Derived Xenografts Undergo Mouse-Specific Tumor Evolution. Nat Genet 2017, 49, 1567–1575. [CrossRef]
- Yuan, Z.; Li, Y.; Zhang, S.; Wang, X.; Dou, H.; Yu, X.; Zhang, Z.; Yang, S.; Xiao, M. Extracellular Matrix Remodeling in Tumor Progression and Immune Escape: From Mechanisms to Treatments. Mol Cancer 2023, 22, 48. [CrossRef]
- Hawkins, A.G.; Julian, C.M.; Konzen, S.; Treichel, S.; Lawlor, E.R.; Bailey, K.M. Microenvironmental Factors Drive Tenascin C and Src Cooperation to Promote Invadopodia Formation in Ewing Sarcoma. Neoplasia 2019, 21, 1063–1072. [CrossRef]
- Rodríguez-Núñez, P.; Romero-Pérez, L.; Amaral, A.T.; Puerto-Camacho, P.; Jordán, C.; Marcilla, D.; Grünewald, T.G.; Alonso, J.; de Alava, E.; Díaz-Martín, J. Hippo Pathway Effectors YAP1/TAZ Induce an EWS–FLI1 -opposing Gene Signature and Associate with Disease Progression in Ewing Sarcoma. J Pathol 2020, 250, 374–386. [CrossRef]
- Bierbaumer, L.; Katschnig, A.M.; Radic-Sarikas, B.; Kauer, M.O.; Petro, J.A.; Högler, S.; Gurnhofer, E.; Pedot, G.; Schäfer, B.W.; Schwentner, R.; et al. YAP/TAZ Inhibition Reduces Metastatic Potential of Ewing Sarcoma Cells. Oncogenesis 2021, 10. [CrossRef]
- Vasileva, E.; Warren, M.; Triche, T.J.; Amatruda, J.F. Dysregulated Heparan Sulfate Proteoglycan Metabolism Promotes Ewing Sarcoma Tumor Growth. Elife 2022, 11. [CrossRef]
- Wullkopf, L.; West, A.-K. V.; Leijnse, N.; Cox, T.R.; Madsen, C.D.; Oddershede, L.B.; Erler, J.T. Cancer Cells’ Ability to Mechanically Adjust to Extracellular Matrix Stiffness Correlates with Their Invasive Potential. Mol Biol Cell 2018, 29, 2378–2385. [CrossRef]
- Aveic, S.; Davtalab, R.; Vogt, M.; Weber, M.; Buttler, P.; Tonini, G.P.; Fischer, H. Calcium Phosphate Scaffolds with Defined Interconnecting Channel Structure Provide a Mimetic 3D Niche for Bone Marrow Metastasized Tumor Cell Growth. Acta Biomater 2019, 88, 527–539. [CrossRef]
- DeClerck, Y.A.; Mercurio, A.M.; Stack, M.S.; Chapman, H.A.; Zutter, M.M.; Muschel, R.J.; Raz, A.; Matrisian, L.M.; Sloane, B.F.; Noel, A.; et al. Proteases, Extracellular Matrix, and Cancer. Am J Pathol 2004, 164, 1131–1139. [CrossRef]
- Shi, K.; Lan, R.-L.; Tao, X.; Wu, C.-Y.; Hong, H.-F.; Lin, J.-H. Vitronectin Significantly Influences Prognosis in Osteosarcoma. Int J Clin Exp Pathol 2015, 8, 11364–11371.
- Tas, F.; Karabulut, S.; Bilgin, E.; Tastekin, D.; Duranyildiz, D. Clinical Significance of Serum Fibronectin and Vitronectin Levels in Melanoma Patients. Melanoma Res 2014, 24, 475–479. [CrossRef]
- Zhu, W.; Li, W.; Yang, G.; Fu, C.; Jiang, G.; Hu, Q. Vitronectin Silencing Inhibits Hepatocellular Carcinoma in Vitro and in Vivo. Future Oncology 2015, 11, 251–258. [CrossRef]
- Bera, A.; Subramanian, M.; Karaian, J.; Eklund, M.; Radhakrishnan, S.; Gana, N.; Rothwell, S.; Pollard, H.; Hu, H.; Shriver, C.D.; et al. Functional Role of Vitronectin in Breast Cancer. PLoS One 2020, 15, e0242141. [CrossRef]
- Ciereszko, A.; Dietrich, M.A.; Słowińska, M.; Nynca, J.; Ciborowski, M.; Kisluk, J.; Michalska-Falkowska, A.; Reszec, J.; Sierko, E.; Nikliński, J. Identification of Protein Changes in the Blood Plasma of Lung Cancer Patients Subjected to Chemotherapy Using a 2D-DIGE Approach. PLoS One 2019, 14, e0223840. [CrossRef]
- Schneider, G.; Suszynska, M.; Kakar, S.; Ratajczak, M.Z. Vitronectin in the Ascites of Human Ovarian Carcinoma Acts as a Potent Chemoattractant for Ovarian Carcinoma: Implication for Metastasis by Cancer Stem Cells. J Cancer Stem Cell Res 2016, 4, 1. [CrossRef]
- Burgos-Panadero, R.; Noguera, I.; Cañete, A.; Navarro, S.; Noguera, R. Vitronectin as a Molecular Player of the Tumor Microenvironment in Neuroblastoma. BMC Cancer 2019, 19, 479. [CrossRef]
- Lopez-Carrasco, A.; Diaz-Martin, J.; Machado, I.; Navarro, S.; Alava, E.; Noguera, R. The Morphology of the Extracellular Matrix as a Classifier of Types and Subtypes of Ewing and Ewing-like Sarcomas: A Morpho-Molecular Study. Histology and Histopathology 2021, 37, supp. 1, p. 109.
- Guerrieri, A.N.; Bellotti, C.; Penzo, M.; Columbaro, M.; Pannella, M.; De Vita, A.; Gambarotti, M.; Mercatali, L.; Laranga, R.; Dozza, B.; et al. A Novel Patient-Derived Immortalised Cell Line of Myxofibrosarcoma: A Tool for Preclinical Drugs Testing and the Generation of near-Patient Models. BMC Cancer 2023, 23, 1194. [CrossRef]
- Kopper, O.; de Witte, C.J.; Lõhmussaar, K.; Valle-Inclan, J.E.; Hami, N.; Kester, L.; Balgobind, A.V.; Korving, J.; Proost, N.; Begthel, H.; et al. An Organoid Platform for Ovarian Cancer Captures Intra- and Interpatient Heterogeneity. Nat Med 2019, 25, 838–849. [CrossRef]
- Braekeveldt, N.; Wigerup, C.; Gisselsson, D.; Mohlin, S.; Merselius, M.; Beckman, S.; Jonson, T.; Börjesson, A.; Backman, T.; Tadeo, I.; et al. Neuroblastoma Patient-derived Orthotopic Xenografts Retain Metastatic Patterns and Geno- and Phenotypes of Patient Tumours. Int J Cancer 2015, 136. [CrossRef]
- Rae, C.; Amato, F.; Braconi, C. Patient-Derived Organoids as a Model for Cancer Drug Discovery. Int J Mol Sci 2021, 22, 3483. [CrossRef]
- Munoz-Garcia, J.; Jubelin, C.; Loussouarn, A.; Goumard, M.; Griscom, L.; Renodon-Cornière, A.; Heymann, M.-F.; Heymann, D. In Vitro Three-Dimensional Cell Cultures for Bone Sarcomas. J Bone Oncol 2021, 30, 100379. [CrossRef]
- Santoro, M.; Lamhamedi-Cherradi, S.-E.; Menegaz, B.A.; Ludwig, J.A.; Mikos, A.G. Flow Perfusion Effects on Three-Dimensional Culture and Drug Sensitivity of Ewing Sarcoma. Proceedings of the National Academy of Sciences 2015, 112, 10304–10309. [CrossRef]
- Monferrer, E.; Sanegre, S.; Martín-Vañó, S.; García-Lizarribar, A.; Burgos-Panadero, R.; López-Carrasco, A.; Navarro, S.; Samitier, J.; Noguera, R. Digital Image Analysis Applied to Tumor Cell Proliferation, Aggressiveness, and Migration-Related Protein Synthesis in Neuroblastoma 3D Models. Int J Mol Sci 2020, 21, 8676. [CrossRef]
- Vieco-Martí, I.; Monferrer, E.; López-Carrasco, A.; Granados-Aparici, S.; Navarro, S.; Noguera, R. Building Silk-Fibroin 3D Hydrogels with Enzymatically Cross-Linked Vitronectin to Study Neuroblastoma Aggressiveness. Histology and Histopathology 2021, 37, supp. 1, p. 152.
- Choi, E.Y.K.; Gardner, J.M.; Lucas, D.R.; McHugh, J.B.; Patel, R.M. Ewing Sarcoma. Semin Diagn Pathol 2014, 31, 39–47. [CrossRef]
- Lamhamedi-Cherradi, S.-E.; Santoro, M.; Ramammoorthy, V.; Menegaz, B.A.; Bartholomeusz, G.; Iles, L.R.; Amin, H.M.; Livingston, J.A.; Mikos, A.G.; Ludwig, J.A. 3D Tissue-Engineered Model of Ewing’s Sarcoma. Adv Drug Deliv Rev 2014, 79–80, 155–171. [CrossRef]
- Cacho-Díaz, B.; García-Botello, D.R.; Wegman-Ostrosky, T.; Reyes-Soto, G.; Ortiz-Sánchez, E.; Herrera-Montalvo, L.A. Tumor Microenvironment Differences between Primary Tumor and Brain Metastases. J Transl Med 2020, 18, 1. [CrossRef]
- Handorf, A.M.; Zhou, Y.; Halanski, M.A.; Li, W.-J. Tissue Stiffness Dictates Development, Homeostasis, and Disease Progression. Organogenesis 2015, 11, 1–15. [CrossRef]
- Abdollahiyan, P.; Oroojalian, F.; Baradaran, B.; de la Guardia, M.; Mokhtarzadeh, A. Advanced Mechanotherapy: Biotensegrity for Governing Metastatic Tumor Cell Fate via Modulating the Extracellular Matrix. Journal of Controlled Release 2021, 335, 596–618. [CrossRef]
- Tadeo, I.; Berbegall, A.P.; Escudero, L.M.; Ãlvaro, T.; Noguera, R. Biotensegrity of the Extracellular Matrix: Physiology, Dynamic Mechanical Balance, and Implications in Oncology and Mechanotherapy. Front Oncol 2014, 4. [CrossRef]
- Yang, Y.; Zhang, Y.; Li, Y. Artificial Intelligence Applications in Pediatric Oncology Diagnosis. Explor Target Antitumor Ther 2023, 157–169. [CrossRef]
- Baxi, V.; Edwards, R.; Montalto, M.; Saha, S. Digital Pathology and Artificial Intelligence in Translational Medicine and Clinical Practice. Modern Pathology 2022, 35, 23–32. [CrossRef]
- Hanna, M.G.; Ardon, O.; Reuter, V.E.; Sirintrapun, S.J.; England, C.; Klimstra, D.S.; Hameed, M.R. Integrating Digital Pathology into Clinical Practice. Modern Pathology 2022, 35, 152–164. [CrossRef]
- Thomas, D.; Rathinavel, A.K.; Radhakrishnan, P. Altered Glycosylation in Cancer: A Promising Target for Biomarkers and Therapeutics. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 2021, 1875, 188464. [CrossRef]
- Schneider, G.; Bryndza, E.; Poniewierska-Baran, A.; Serwin, K.; Suszynska, M.; Sellers, Z.P.; Merchant, M.L.; Kaliappan, A.; Ratajczak, J.; Kucia, M.; et al. Evidence That Vitronectin Is a Potent Migration-Enhancing Factor for Cancer Cells Chaperoned by Fibrinogen: A Novel View of the Metastasis of Cancer Cells to Low-Fibrinogen Lymphatics and Body Cavities. Oncotarget 2016, 7, 69829–69843. [CrossRef]
- Chen, M.H.; Lu, C.; Sun, J.; Chen, X.D.; Dai, J.X.; Cai, J.Y.; Chen, X.L. Diagnostic and Prognostic Value of Serum Vitronectin Levels in Human Glioma. J Neurol Sci 2016, 371, 54–59. [CrossRef]
- Kadowaki, M.; Sangai, T.; Nagashima, T.; Sakakibara, M.; Yoshitomi, H.; Takano, S.; Sogawa, K.; Umemura, H.; Fushimi, K.; Nakatani, Y.; et al. Identification of Vitronectin as a Novel Serum Marker for Early Breast Cancer Detection Using a New Proteomic Approach. J Cancer Res Clin Oncol 2011, 137, 1105–1115. [CrossRef]
- Turan, T.; Torun, M.; Atalay, F.; Gönenç, A. Assessment of Vitronectin, Soluble Epithelial-Cadherin and TGF-Β1 as a Serum Biomarker with Predictive Value for Endometrial and Ovarian Cancers. Turk J Pharm Sci 2017, 14, 141–147. [CrossRef]
- Repetto, O.; Caggiari, L.; De Zorzi, M.; Elia, C.; Mussolin, L.; Buffardi, S.; Pillon, M.; Muggeo, P.; Casini, T.; Steffan, A.; et al. Quantitative Plasma Proteomics to Identify Candidate Biomarkers of Relapse in Pediatric/Adolescent Hodgkin Lymphoma. Int J Mol Sci 2022, 23, 1–15. [CrossRef]
- Braoudaki, M.; Lambrou, G.I.; Vougas, K.; Karamolegou, K.; Tsangaris, G.T.; Tzortzatou-Stathopoulou, F. Protein Biomarkers Distinguish between High- and Low-Risk Pediatric Acute Lymphoblastic Leukemia in a Tissue Specific Manner. J Hematol Oncol 2013, 6. [CrossRef]
- López-Carrasco, A.; Vieco-Martí, I.; Granados-Aparici, S.; Acevedo-León, D.; Estañ-Capell, N.; Portugal, R.; Huerta-Aragonés, J.; Cañete, A.; Navarro, S.; Noguera, R. Vitronectin Levels in Plasma of Neuroblastoma Patients and Culture Media of 3D Models: A Prognostic Circulating Biomarker?. Preprints 2024, 2024070957. [CrossRef]
- Stepanek, O.; Brdicka, T.; Angelisova, P.; Horvath, O.; Spicka, J.; Stockbauer, P.; Man, P.; Horejsi, V. Interaction of Late Apoptotic and Necrotic Cells with Vitronectin. PLoS One 2011, 6, e19243. [CrossRef]
- Goyal, U.; Ta, M. A Novel Role of Vitronectin in Promoting Survival of Mesenchymal Stem Cells under Serum Deprivation Stress. Stem Cell Res Ther 2020, 11, 181. [CrossRef]
- Leavesley, D.I.; Kashyap, A.S.; Croll, T.; Sivaramakrishnan, M.; Shokoohmand, A.; Hollier, B.G.; Upton, Z. Vitronectin—Master Controller or Micromanager? IUBMB Life 2013, 65, 807–818. [CrossRef]
- Heyman, L.; Leroy-Dudal, J.; Fernandes, J.; Seyer, D.; Dutoit, S.; Carreiras, F. Mesothelial Vitronectin Stimulates Migration of Ovarian Cancer Cells. Cell Biol Int 2010, 34, 493–502. [CrossRef]
- Kurozumi K.; Ichikawa T.; Onishi M.; Fujii K.; Date I. Cilengitide treatment for malignant glioma: current status and future direction. Neurol Med Chir (Tokyo) 2012, 52, 539-47. [CrossRef] [PubMed]
- Stoks, M.; Vieco-Martí, I.; Noguera, I.; Sánchez-Sánchez, M.; Burgos-Panadero, R.; Navarro, S.; Noguera, R. Digital Image Analysis Workflows for Evaluation of Cell Behavior and Tumor Microenvironment to Aid Therapeutic Assessment in High-Risk Neuroblastoma. Comput Biol Med 2023, 164. [CrossRef]
- Burgos-Panadero, R.; El Moukhtari, S.H.; Noguera, I.; Rodríguez-Nogales, C.; Martín-Vañó, S.; Vicente-Munuera, P.; Cañete, A.; Navarro, S.; Blanco-Prieto, M.J.; Noguera, R. Unraveling the Extracellular Matrix-Tumor Cell Interactions to Aid Better Targeted Therapies for Neuroblastoma. Int J Pharm 2021, 608. [CrossRef]
- Hasturk, O.; Jordan, K.E.; Choi, J.; Kaplan, D.L. Enzymatically Crosslinked Silk and Silk-Gelatin Hydrogels with Tunable Gelation Kinetics, Mechanical Properties and Bioactivity for Cell Culture and Encapsulation. 2019. [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open Source Software for Digital Pathology Image Analysis. Sci Rep 2017, 7, 16878. [CrossRef]
Figure 1.
Vitronectin (VN) expression and secretion of Ewing sarcoma (ES) cell lines in 2D cell cultures. (A-B) Cytospins from 2D cultures of A673 (A) and PDX73 (B) cell lines, immunostained with anti-vitronectin antibody, are shown in 20µm of viewer scale. (C) Detection of vitronectin secreted and number of cells. Green bars represent concentration of VN secreted to the culture media measured with the scale of ng/mL per million of cells (left Y-axis), and dots represent the number of cells quantified in the monolayer cultures measured with scale of millions of cells, as shown in the right Y-axis.
Figure 1.
Vitronectin (VN) expression and secretion of Ewing sarcoma (ES) cell lines in 2D cell cultures. (A-B) Cytospins from 2D cultures of A673 (A) and PDX73 (B) cell lines, immunostained with anti-vitronectin antibody, are shown in 20µm of viewer scale. (C) Detection of vitronectin secreted and number of cells. Green bars represent concentration of VN secreted to the culture media measured with the scale of ng/mL per million of cells (left Y-axis), and dots represent the number of cells quantified in the monolayer cultures measured with scale of millions of cells, as shown in the right Y-axis.

Figure 2.
Vitronectin (VN) expression and secretion of Ewing sarcoma (ES) cell lines in 3D cell cultures. (I) Digital images of VN expression of ES cell lines A673 (A-D) and PDX73 (E-H) grown in hydrogels (HGs) without and with added VN (50µm of viewer scale) and zoom zones in a 10 µm viewer scale (Top left squares). (A and D) Non-added-VN HGs at 2 weeks (2w). (B and E) Added-VN HGs at 2w. (C and F) Non-added-VN HGs at 3 weeks (3w). (G) Added-VN HGs at 3w culture. (II) Detection of vitronectin (VN) secreted to culture media by the cell lines grown in hydrogels (HGs). Orange and blue bars represent concentration of VN secreted to the culture media measured with scale of the left Y-axis in ng/mL per million of cells and dots represent number of cells quantified in the 3D cultures measured in millions of cells (right Y-axis). Blue and orange bars represent HGs without (no VN) and with added VN (VN), respectively. Light colors refer to 2w culture and dark colors to 3w.
Figure 2.
Vitronectin (VN) expression and secretion of Ewing sarcoma (ES) cell lines in 3D cell cultures. (I) Digital images of VN expression of ES cell lines A673 (A-D) and PDX73 (E-H) grown in hydrogels (HGs) without and with added VN (50µm of viewer scale) and zoom zones in a 10 µm viewer scale (Top left squares). (A and D) Non-added-VN HGs at 2 weeks (2w). (B and E) Added-VN HGs at 2w. (C and F) Non-added-VN HGs at 3 weeks (3w). (G) Added-VN HGs at 3w culture. (II) Detection of vitronectin (VN) secreted to culture media by the cell lines grown in hydrogels (HGs). Orange and blue bars represent concentration of VN secreted to the culture media measured with scale of the left Y-axis in ng/mL per million of cells and dots represent number of cells quantified in the 3D cultures measured in millions of cells (right Y-axis). Blue and orange bars represent HGs without (no VN) and with added VN (VN), respectively. Light colors refer to 2w culture and dark colors to 3w.
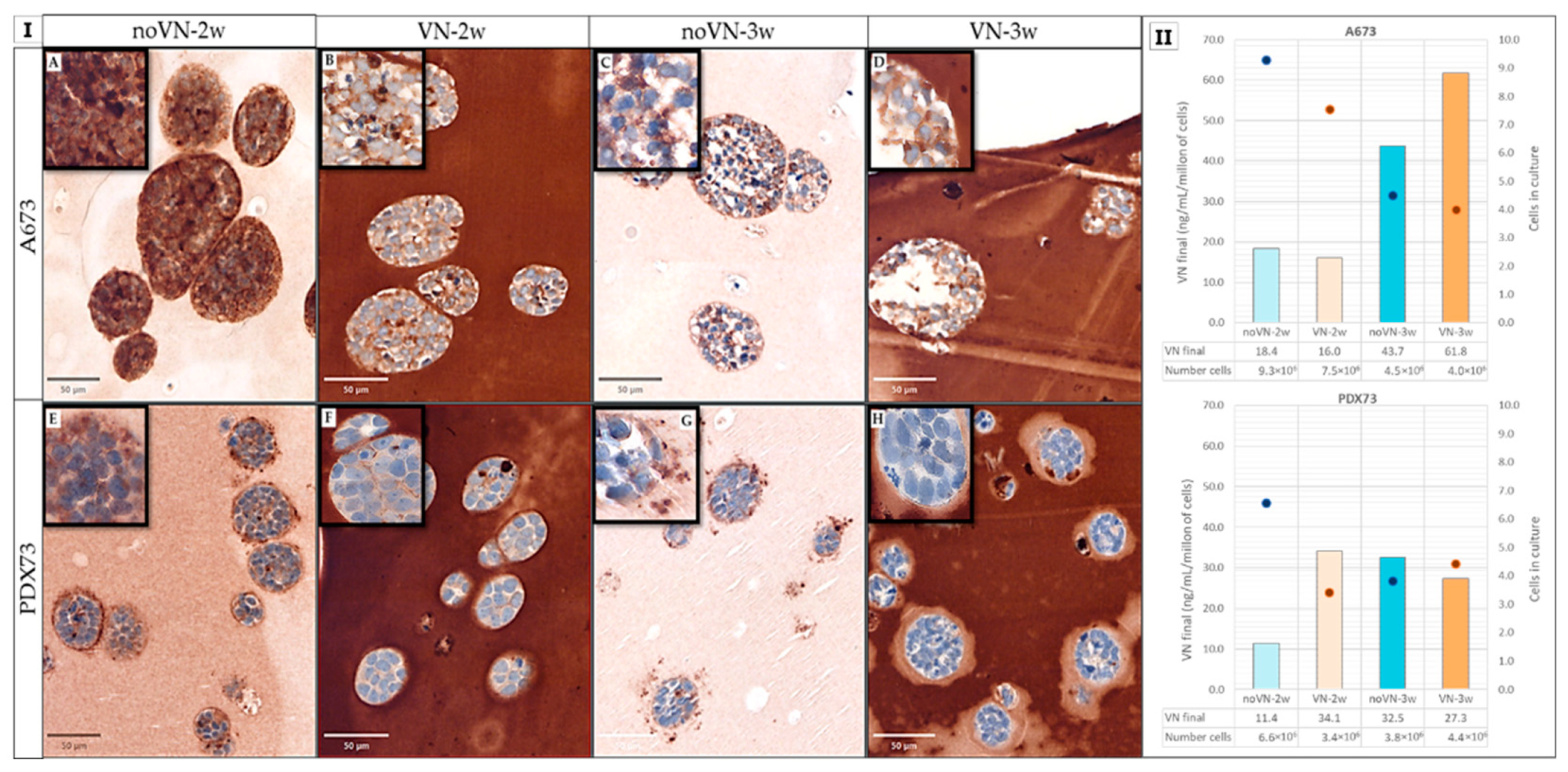
Figure 3.
Digital imagen of examples of clusters in hydrogels (HGs) staining with hematoxylin and eosin. (A-D) HGs of cell line A673, (E-H) HGs of cell line PDX73, (A and E) non-added-VN at 2 weeks (2w), (B and F) added-VN at 2w, (C and G) non-added-VN at 3w, (D and H) added-VN at 3w of culture. In 50µm of viewer scale and zoom zones in a 10 µm viewer scale (Top squares).
Figure 3.
Digital imagen of examples of clusters in hydrogels (HGs) staining with hematoxylin and eosin. (A-D) HGs of cell line A673, (E-H) HGs of cell line PDX73, (A and E) non-added-VN at 2 weeks (2w), (B and F) added-VN at 2w, (C and G) non-added-VN at 3w, (D and H) added-VN at 3w of culture. In 50µm of viewer scale and zoom zones in a 10 µm viewer scale (Top squares).
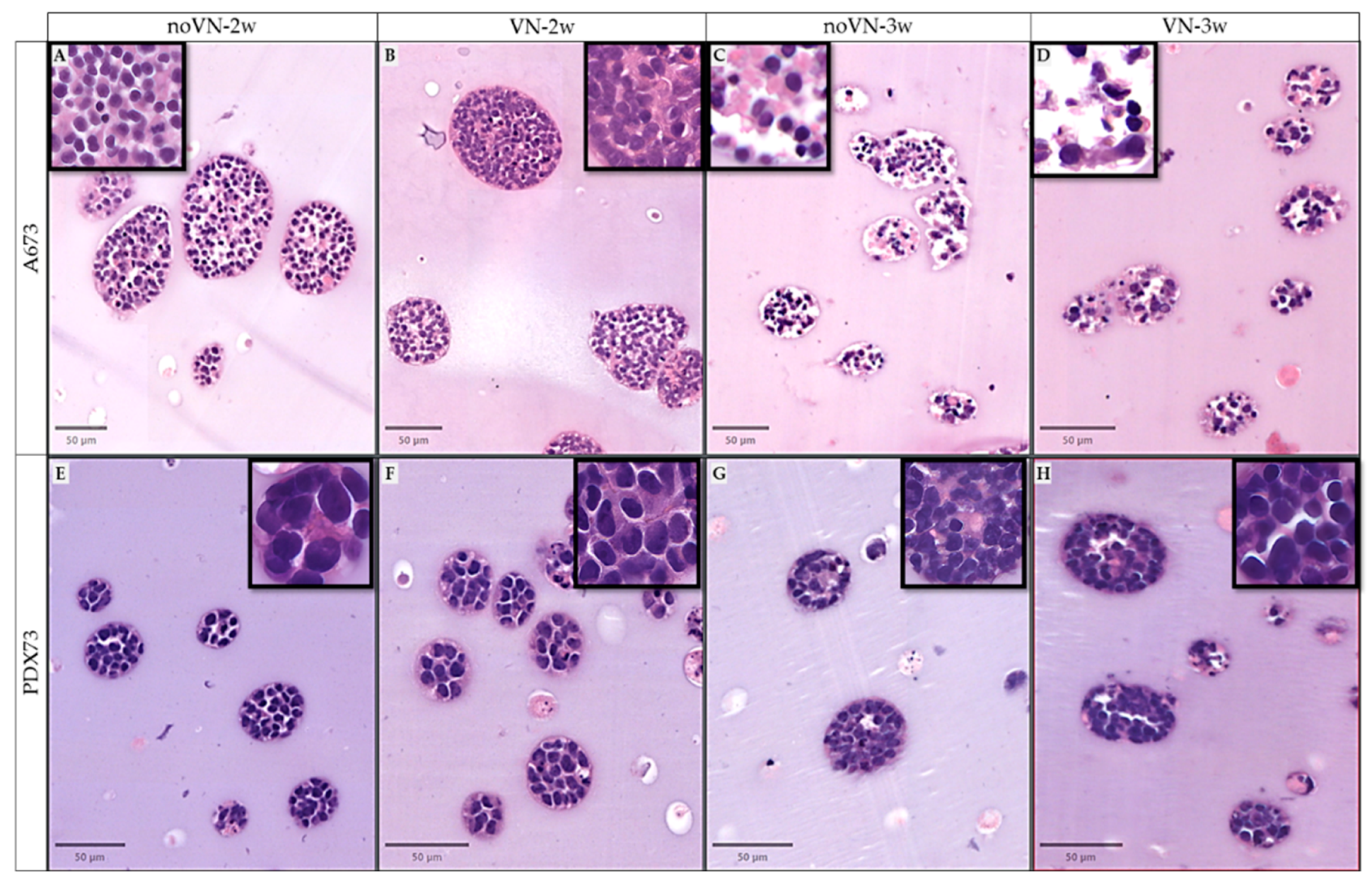
Figure 4.
Cluster density (clusters/nm²) in 3D cultures evaluated by hydrogel composition and time of culture. (A) A673 and (B) PDX73 cell cultures. Boxes in cyan represent non-added-VN in the scaffold (noVN), boxes in orange represent added-VN in the scaffold (VN), light colors represent 2-week culture (2w) and dark colors represent 3-week culture (3w).
Figure 4.
Cluster density (clusters/nm²) in 3D cultures evaluated by hydrogel composition and time of culture. (A) A673 and (B) PDX73 cell cultures. Boxes in cyan represent non-added-VN in the scaffold (noVN), boxes in orange represent added-VN in the scaffold (VN), light colors represent 2-week culture (2w) and dark colors represent 3-week culture (3w).
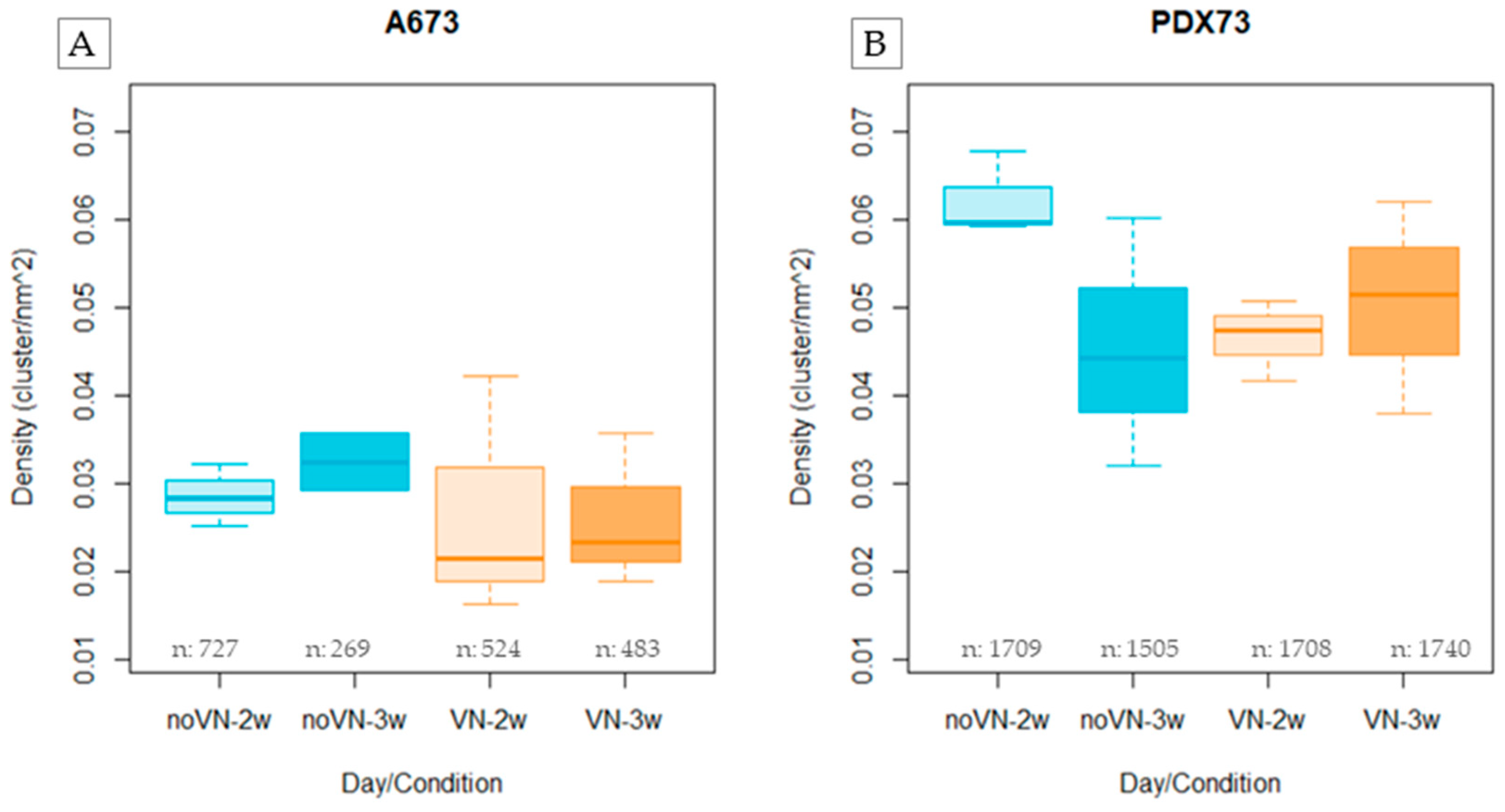
Figure 5.
Cluster density (clusters/nm²) in 3D cultures classified by cluster size. The top graphs indicate the A673 cell line and bottom graphs the PDX73 cell line. Boxes in cyan represent non-added-VN in the scaffold (noVN), boxes in orange represent added-VN in the scaffold (VN), light color shading represents 2-week cultures (2w) and dark colors represent 3-week cultures (3w). Note the changes in density scale between cluster sizes, as indicated in the Y-axis of the graphs.
Figure 5.
Cluster density (clusters/nm²) in 3D cultures classified by cluster size. The top graphs indicate the A673 cell line and bottom graphs the PDX73 cell line. Boxes in cyan represent non-added-VN in the scaffold (noVN), boxes in orange represent added-VN in the scaffold (VN), light color shading represents 2-week cultures (2w) and dark colors represent 3-week cultures (3w). Note the changes in density scale between cluster sizes, as indicated in the Y-axis of the graphs.

Figure 6.
Cluster area (μm²) in 3D cultures classified by cluster size. Top graphs refer to A673 cell line and bottom graphs to PDX73 cell line. Boxes in cyan represent non-added-VN in the scaffold (noVN), boxes in orange represent added-VN in the scaffold (VN), light colors represent 2 weeks cultures (2w) and dark colors represent 3 weeks cultures (3w). One-way ANOVA with Dun´s multiple comparison test revealed significant differences: * = p < 0.05, ** = p < 0.01, *** = p < 0.001, **** = p < 0.0001. Note the changes in area scale between cluster sizes, as indicated in the Y-axis of the graphs.
Figure 6.
Cluster area (μm²) in 3D cultures classified by cluster size. Top graphs refer to A673 cell line and bottom graphs to PDX73 cell line. Boxes in cyan represent non-added-VN in the scaffold (noVN), boxes in orange represent added-VN in the scaffold (VN), light colors represent 2 weeks cultures (2w) and dark colors represent 3 weeks cultures (3w). One-way ANOVA with Dun´s multiple comparison test revealed significant differences: * = p < 0.05, ** = p < 0.01, *** = p < 0.001, **** = p < 0.0001. Note the changes in area scale between cluster sizes, as indicated in the Y-axis of the graphs.
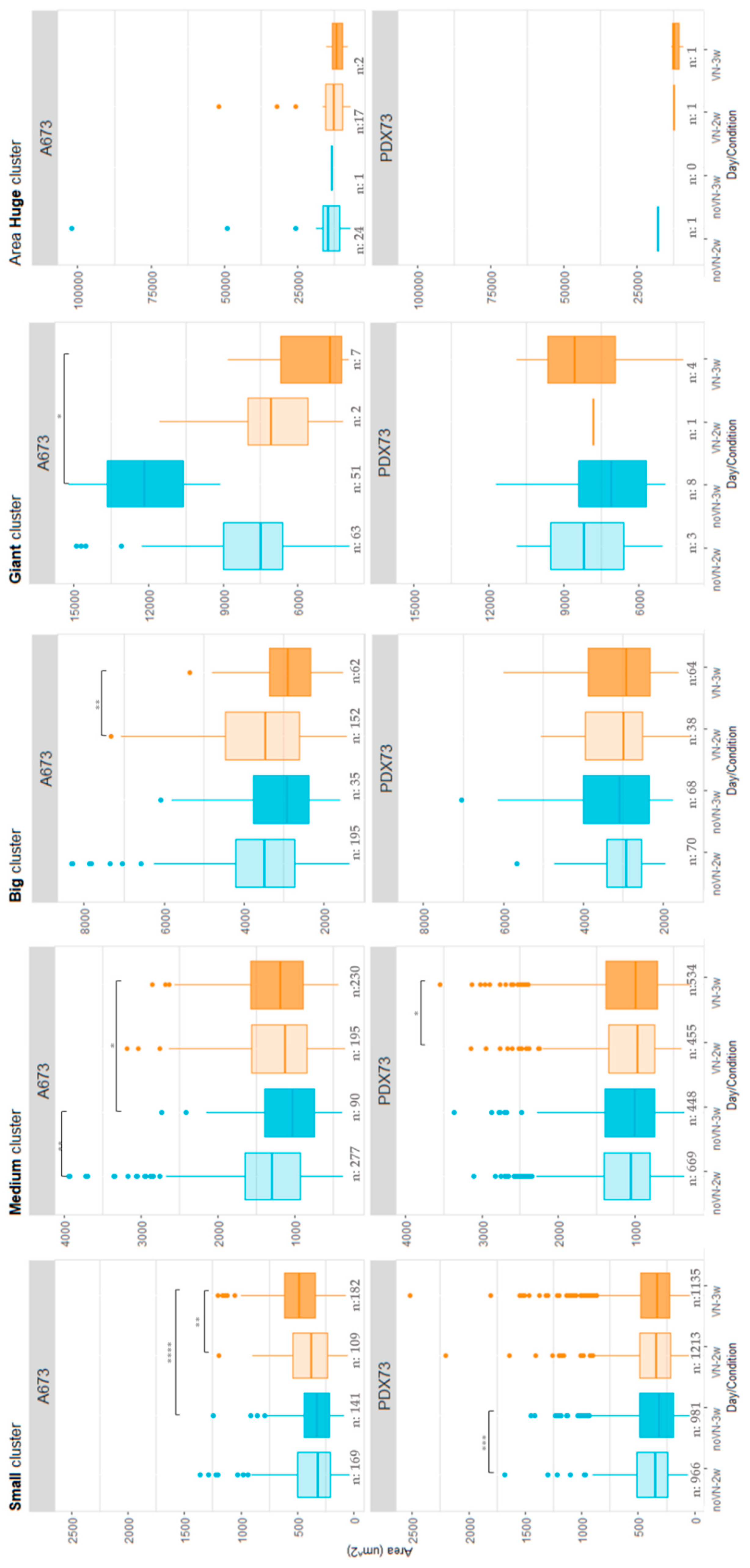
Figure 6.
Cluster circularity in 3D cultures classified by cluster size. The top graphs show the A673 cell line and bottom graphs the PDX73 cell line. Boxes in cyan represent no added VN in the scaffold (noVN), boxes in orange represent added VN in the scaffold (VN), the lighter shading of each color representing culture for 2 weeks (2w) and the darker shading culture for 3 weeks (3w). One-way ANOVA with Dun´s multiple comparison test revealed significant difference * = p < 0.05, ** = p < 0.01, *** = p < 0.001. **** = p<0.0001. Clusters with a more circular shape have a circularity value closer to 1.
Figure 6.
Cluster circularity in 3D cultures classified by cluster size. The top graphs show the A673 cell line and bottom graphs the PDX73 cell line. Boxes in cyan represent no added VN in the scaffold (noVN), boxes in orange represent added VN in the scaffold (VN), the lighter shading of each color representing culture for 2 weeks (2w) and the darker shading culture for 3 weeks (3w). One-way ANOVA with Dun´s multiple comparison test revealed significant difference * = p < 0.05, ** = p < 0.01, *** = p < 0.001. **** = p<0.0001. Clusters with a more circular shape have a circularity value closer to 1.
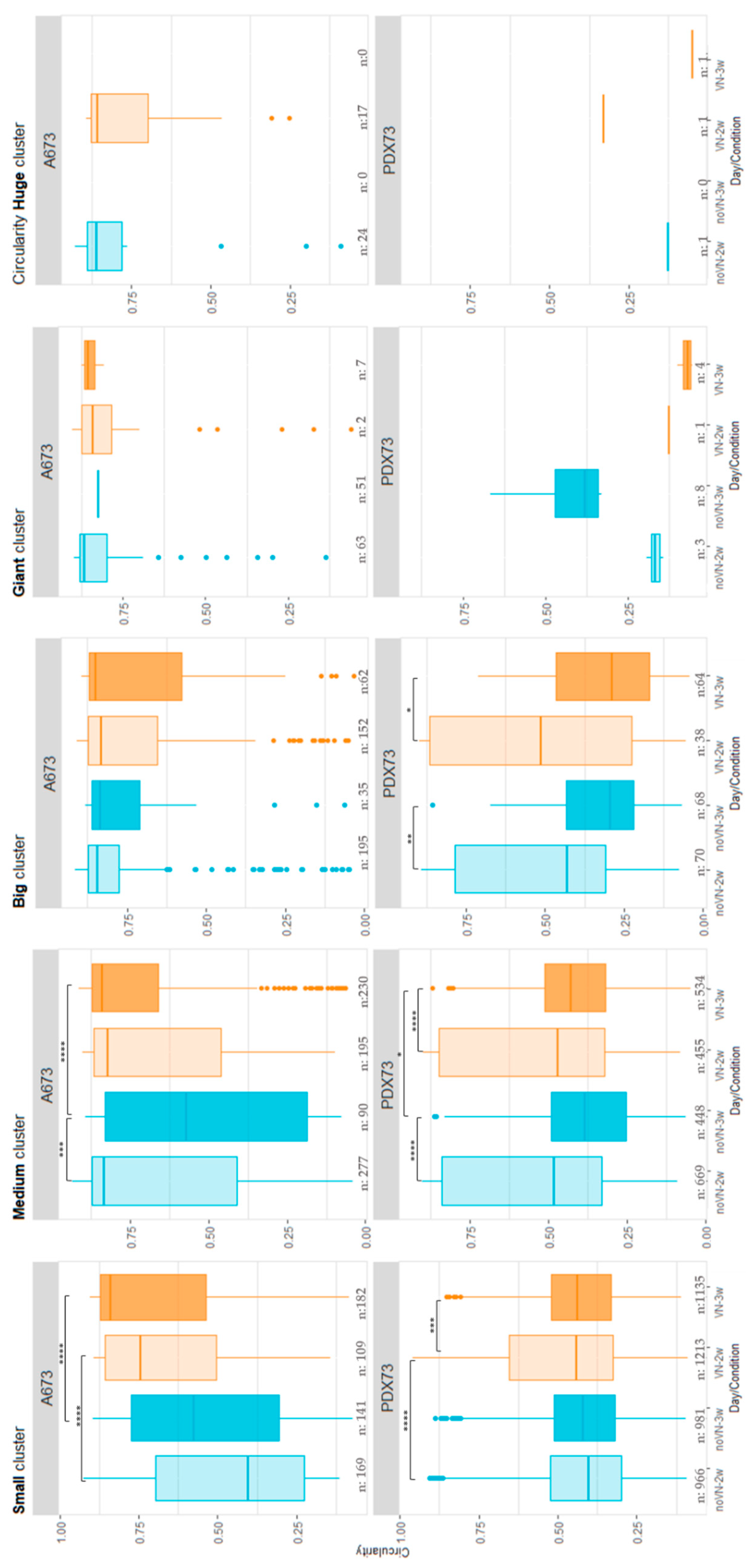

Table 1.
Cluster Classification Parameters.
| Number of Cells | Classification | Color Code |
|---|---|---|
| <9 >10 and 38< |
Small Medium |
Cyan Green |
| >39 and 95< >96 and 169< >170 |
Big Giant Huge |
Yellow Magenta Blue |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



