
Submitted:
07 August 2024
Posted:
08 August 2024
You are already at the latest version
Abstract
Soil formation results from the weathering and alteration of bedrock, influenced by climatic conditions and biotic factors, including living organisms. The mineralogical composition of the parent material, along with plant species and soil microorganisms, forms the foundational elements of an ecosystem's energy cycle. Afforestation in arid-semi arid regions plays a crucial role in preventing erosion and enhancing soil quality, offering significant economic and ecological benefits. This study assessed the impact of parent material diversity on soil physicochemical properties, microbial basal respiration (MR), and microbiological characteristics, as well as their subsequent effects on plant growth, 15 years post-afforestation. For this purpose, various soil physicochemical parameters, MR, soil microbial biomass carbon (Cmic), stoichiometry (microbial quotient= Cmic/Corg = qMic and metabolic quotient = MR/Cmic = qCO2), and tree growth metrics such as height and diameter were measured. The results of the one-way ANOVA indicated that afforested soils on limestone bedrock exhibited significantly better physicochemical and microbiological properties, as well as tree growth parameters, compared to those on andesite. Notably, sensitive microbial properties such as Cmic, MR, and qMic were positively influenced by afforestation. The highest Cmic (323 g C kg−1) and MR (1.3 CO2–C g−1 h−1) values were recorded in limestone-derived soils. Currently, the measured properties of soils can be ranked as LS>AS>LC>AC. Conversely, the highest qCO2 values were observed in the control plots of andesite parent material (7.22). This study highlights the importance of selecting appropriate parent material and understanding soil characteristics to optimize afforestation efforts in bare areas. As a result, the higher plant growth capacity and carbon retention of limestone soil indicate that it has high microbial biomass and microbial activity (qMic).
Keywords:
Parent material
; afforestation
; Scots pine
; microbial biomass C
; microbial respiration
; plant growth
1. Introduction
Soil is formed under the influence of bedrock, climate, vegetation, and topographic factors. However, the dominance of one factor over the others can lead to variations in soil properties and land productivity [1]. Semiarid and arid ecosystems, for example, are primarily influenced by parent material and the continental climate, with summer drought being a critical stress factor [2,3]. The physical properties, mineral composition, and texture of the parent material significantly affect the soil formation rate and the characteristics of the resulting soil [4]. The hardness of rocks is also crucial; soils formed on hard rocks have lower formation rates than the rate of soil loss through erosion, leading to poor habitat characteristics. Birkeland [5] ranked bedrock stability against soil formation factors as follows: quartzite, chert > granite, basalt > sandstone, siltstone > dolomite, limestone.
The parent material contributes to the nutrient cycle by releasing nutrients through weathering and maintaining a balance between nutrient loss and storage [1,4]. After climate, plant and microorganism activities play the most important roles in soil formation. Trees, in particular, influence parent material through biomechanical, chemical, and biological effects [1,3]. Traditionally, studies have focused on the biochemical, ecological, and edaphic influences on soils, but the significance of biomechanical effects is increasingly recognized [6,7,8]. Most research on the biomechanical effects of vegetation has centered on tree root growth, but trees also impact the physical displacement of regolith materials and the formation of root debris and exudates [9]. Plant roots mechanically break down rocks during growth, accelerating soil formation processes. Additionally, root exudates and mechanical root activities help reduce erosion, stabilizing soil ecosystems [10]. Organic acids and exudates produced by plants and microorganisms enhance the chemical weathering of minerals and the release of nutrients [11].
Over the last 20 years, the Turkish Ministry of Agriculture and Forestry has conducted afforestation activities to increase forest cover to 30% of Turkey’s surface area. Scots pine and black locust are commonly used for erosion control due to their high tolerance to drought and poor soil conditions [12]. Planting native tree species has accelerated natural succession [13] and carbon sequestration [14]. Plants, known as the organisms with the largest biomass in terrestrial ecosystems, contribute greatly to the formation and development of soils through litter residues, leaves, branches, roots, and exudates of their aboveground biomass [15,16]. In addition, vegetation affects the organic carbon (OC) concentration added to soils and the diversity, quality, and quantity of soil microorganisms [17]. For example, the biomass of microbial organisms in soils converted from forest to agriculture decreases significantly [18,19]. Similarly, agricultural land contains less soil organic C (SOC) and Cmic compared to native forests [20]. Vegetation type strongly influences soil-forming processes, contributing significantly to organic matter and microbial community development and activity [21,22]. Microbial stoichiometric indices such as microbial basal respiration (MR), microbial quotient (qMic), metabolic quotient (qCO2), and C mineralization are related to the SOC cycle in the environment. With the help of these stoichiometric indices, the effects of changes in land use or ecosystem on soil quality, microbial populations, and microbial activity rates can be estimated [23,24].
While many studies suggest that microbial biomass is controlled by soil physicochemical properties [25,26], others indicate that aboveground vegetation type has a stronger influence on microbial biomass and activity [27,28]. Kara et al. [10] found positive effects of ecosystem changes on microbial biomass and activity in afforested areas in both the short and long term. Research on afforested ecosystems shows that vegetation enhances soil-forming processes, leading to organic matter accumulation and microbial community development [29,30,31]. Trees provide nutrients to the soil through dead plant residues and root secretions, which microorganisms break down and mineralize, forming a usable nutrient source for plants [32]. Microbial organisms decompose this litter, returning essential organic nutrients to the soil. This process enriches the growing environment, enhancing the productivity of the soil. As a result, plant growth parameters, such as tree height (TH) and diameter at breast height (DBH), improve significantly. The upper soil horizon, a reservoir for soil microbial biomass, is crucial for decomposition and nutrient cycling [33]. This symbiotic relationship underpins the nutrient cycle, essential for ecosystem health.
Soil microorganisms are the most rapidly affected by environmental conditions, various forestry management practices, and sudden changes on or within the soil surface. Consequently, measuring microbial activities and properties following forestry management activities is essential for monitoring changes in soil fertility. Changes in microbial properties can provide early indications of potential ecosystem disruptions [32,34,35,36,37]. Assessing microbial biomass in soils is crucial in many soil monitoring studies and programs [35], but it should be considered alongside other habitat parameters for a comprehensive ecological assessment. Although recent studies focus on the effects of soil microbiological properties on soil fertility, there is still limited information on their relationship with tree growth. This research aimed to new perspectives on the effects of physicochemical, microbial properties, and stoichiometry of soils derived from different bedrocks on seedling growth parameters, highlighting the importance of evaluating soil quality parameters in forests.
2. Materials and Methods
2.1. Study Sites
The study was conducted in Gümüşhane Province, located in the Northeast Black Sea mountainous region of Turkey (40°19′–40°23′ N and 39°30′–39°38′ E). The elevation ranges from 1,400 to 1,600 meters, with steep slopes (45–50%) covered by shallow soil (typically 50 cm or less) on a west-southwest aspect. The region has a “Warm, dry-summer continental climate” (Dsb) according to the Köppen-Geiger climate classification (www.koeppen-geiger.vu-wien.ac.at/). Weather data, including long-term averages (1990–2018) for monthly air temperature and precipitation in Gümüşhane Province, were obtained from the Turkish State Meteorological Service (www.mgm.gov.tr). The annual mean temperature is 9.4°C, with a high of 39.5°C in July and a low of −19.0°C in February. The topsoil temperature drops below 0°C and freezes during the winter. Annual precipitation is 461 mm, with about 54% occurring during the growing season between April and September. The main soil type in the area is Entisol, characterized by minimal horizon development. The rugged topography consists of andesitic and limestone bedrock. Table 1 provides a detailed characterization of the study site.
2.2. Experimental Design, Soil Sampling, and Pretreatment
This study was conducted in the spring of 2019 at Scotch pine (Pinus sylvestris L.) afforestation sites, planted over 30 hectares in 2005 by the General Directorate of Afforestation of Türkiye. The sites were initially prepared using an excavator, and irrigation and fertilization were applied during the first five years after planting. The control area was significantly degraded grassland with less than 5% vegetation cover, exposed to erosion and not fertilized. A grid-based sampling design was used to select three 10x10 m² experimental areas from different bedrocks, representing a total of 30 hectares. Tree growth parameters, such as tree height (TH) and diameter of breast height (DBH), were directly measured within each 100 m² area using a Blume-Leiss and compass, respectively. Soil samples were taken at the beginning of the vegetation period in the Scots pine forest, with the bare land used as a control. A total of 40 sample points were systematically selected from two different parent materials afforested and control areas (LS= limestone sample; LC= limestone control; AS= Andesite sample; AC= Andesite control). Before collecting disturbed mineral soil samples, the litter, root debris, and stones were removed, and stored in plastic bags. In the field study, microbial soil samples were first sieved using <2 mm sieve, then distilled water was added until 50-60% moisture content was reached and quickly transferred to the laboratory for storage in a 4°C refrigerator before analysis. The disturbed and microbial soil samples were collected from the topsoil layer at 0-20 cm depth. Other soil samples were dried by spreading them in drying cabinets to make them air-dried, after drying the soils were ground and sieved (<2 mm) to determine the physicochemical parameters of the soil.
2.3. Physical and Chemical Properties of Soils
Soil moisture content was determined gravimetrically by oven-drying samples at 105°C until a stable weight was achieved. Soil particle size (PS) was measured using the hydrometer method [38]. Soil pH and electrical conductivity (EC) were assessed using a 1:2.5 soil/water suspension for pH and a 1:5 soil/water suspension for EC, measured with appropriate meters. Soil organic carbon (SOC) was estimated using the potassium dichromate oxidation method [39], total nitrogen by the Kjeldahl method [40], while total lime (CaCO3) content was determined using the Scheibler calcimeter method [41].
2.4. Microbial Biomass C (Cmic)
The chloroform-fumigation-extraction method was performed to determine Soil microbial biomass carbon (Cmic) [42]. Each soil samples were divided into 2 portions and the first half of the soils stored for microbial analyses were fumigated with ethanol-free chloroform at 25°C for 24 h in a vacuumed desiccator, while the second portion of samples was not fumigated. Both fumigated and non-fumigated soil samples were mixed in 0.5 M K2SO4 (1:4 w/v), followed by shaking the suspension for 30 minutes at 200 rpm in an oscillator. The extracts were filtered through the Whatman-42 filter paper and stored before titration. Total organic carbon in both fumigated and non-fumigated extracts was estimated as specified by the Walkley Black method [43,44]. Cmic was calculated from the difference in extractable organic C between fumigated and unfumigated soil samples using the formula: biomass C = 2.64 *EC, where EC refers to the difference in extractable organic C between the treatments; 2.64 is the proportionality factor for biomass C released by fumigation extraction [42,44]. The microbial quotient (Cmic/Corg or qMic) was calculated by expressing microbial C as a percentage of total soil organic C.
2.5. Microbial Respiration
Microbial respiration was determined using the absorption method to quantify carbon dioxide (CO2) [45]. Moist soil samples (50 g dry weight equivalent) were placed in 60-mL beakers and incubated in the dark at 25°C in 500 mL airtight, sealed jars along with 10 mL 0.1 M NaOH. After 7-day incubation, the CO2 generated was measured by titration of the excess NaOH with 0.05 M HCl [46]. An empty jar without soil was used as the control. The difference in the volume of HCl consumed between the treatment and the control was used to calculate CO2 emission from soil microorganisms. The metabolic quotient qCO2 is an important parameter that can be used to estimate soil quality and stress response. It was calculated as the microbial respiration rate (μg CO2-C h−1) per mg of Cmic [45].
2.6. Statistical Analyses
The data were tested for normality and homogeneity of distribution using skewness, kurtosis, and Shapiro–Wilk tests. Parameters that deviated from normal distribution were log-transformed (log10) before analyzing variance (ANOVA). After confirming that the data met the assumptions for parametric tests, one-way ANOVA was applied to assess the effects of parent material on all parameters, followed by Duncan’s multiple range tests (α = 0.05). Pearson’s linear correlation analysis was also conducted to assess the relationships between soil properties and plant growth characteristics. Discriminant analysis (DA) was employed to analyze the differences between groups of multivariate data using one or more discriminant functions to maximally separate the identified groups. DA was specifically used to differentiate between the tree growths based on soil variables. All statistical analyses were carried out using the Statistical Package for the Social Sciences (SPSS Inc., Chicago, IL, USA) and figures were generated by using the XLSTAT Windows software package.
3. Results
Under similar ecological conditions, soils formed from different bedrocks exhibited significant differences in physical and chemical properties, including sand, silt, clay content, pH, EC, SOC, TN, and C/N ratio. These variations also extended to microbial activity indicators such as MR, Cmic, qMic, and qCO2. Consequently, these differences in soil properties and microbial activity led to significant variations in the growth characteristics of the seedlings planted in the area (Table 2).
3.1. Physicochemical Properties of the Soil
The results indicated significant differences in the analyzed properties of soils developed on different parent materials, with notable improvements in soils from post-afforestation environments. The ANOVA test revealed that the physical, chemical, and microbiological properties of these soils were significantly different from each other and the control areas. These differences in soil properties influenced soil fertility, resulting in varying development rates among the seedlings planted in the different areas. These findings are consistent with other studies [1,3,47]. The mean values of some physicochemical properties and plant growth parameters are presented in Table 3, while the microbial indices and microbial basal respiration are depicted in Figure 1 and Figure 2.
According to international soil texture classifications, the soils at the research sites displayed varying textures based on parent material. In the afforestation site, soils derived from limestone were classified as clayey, while those in the limestone control site were also clayey. Soils from basaltic andesite were identified as sandy clayey in the afforestation sites and sandy loam in the control area. The percentages of sand, silt, and clay differed significantly depending on the parent material. Soils from basaltic andesite had significantly higher sand content compared to those from limestone (Table 3). Conversely, the average silt and clay contents were significantly higher in limestone-derived soils (Table 3). In other words, andesite produced coarse-grained soils with significantly higher sand and lower silt and clay percentages. Notably, the afforestation sites exhibited a decrease in sand and dust percentages compared to the control sites, while the clay percentages increased (Table 3).
The mean soil reaction (pH) values and electrical conductivity (EC) of the limestone soils statistically differed from the andesite soils (P < 0.001) (Table 2). The mean pH and EC values in all soil samples including controls were found to be the highest in the control sites on limestone bedrock (Table 3). However, the lowest pH and EC values were consistently observed in the soils of the areas with significant andesite bedrock. Moreover, the mean pH (8.08 and 8.18) and EC (0.17 and 0.18 dSm−1) in the limestone bedrock sampling site were higher than andesite afforestation and control sites with 7% and 8%; 30%, and 42%, respectively. The average pH and EC values in all soil samples, including controls, were found to be the highest in afforestation sites on limestone bedrock (Table 3). However, the lowest pH and EC values were consistently observed in soils from areas with significant amounts of andesite bedrock. The pH of limestone bedrock was moderately alkaline (8.08–8.18) in afforestation and control sites and slightly alkaline (7.59–7.57) in andesite bedrock afforestation and control sites. Again, the EC values, like the pH in limestone bedrock control and afforestation sites, were higher than those observed in andesite bedrock. EC values were found below the salinity threshold level (0.20 dS cm−1).
As expected, SOC and TN contents differed significantly among the parent materials (p < 0.0001). The average SOC contents of the afforestation sites were 1.65% in andesite and 2.40% in limestone, while TN content was 0.10% in andesite and 0.19% in limestone. SOC and TN amounts in afforestation sites increased by 300% in limestone and 333% in andesite compared to control sites. These results indicate that SOC and TN levels were highest in afforestation sites constructed on limestone parent material (Table 3). The highest annual mean C/N ratio was observed in the andesite control site (22.74), followed by the limestone control site (18.76), the andesite afforestation site, and finally, the lowest in the limestone afforestation site (12.61). The mean C/N ratio of the andesite control site was 17%, 29%, and 44% higher than the limestone control, andesite afforestation, and limestone afforestation sites, respectively (Table 3). Additionally, the soil CaCO3 percentage was significantly higher in soils derived from limestone compared to andesite. After afforestation, this value decreased by 30% (from 32.90% to 23.09%) in limestone parent material and by 36% (from 4.85% to 3.08%) in andesite parent material compared to the control sites.
3.2. Soil Microbiological Properties
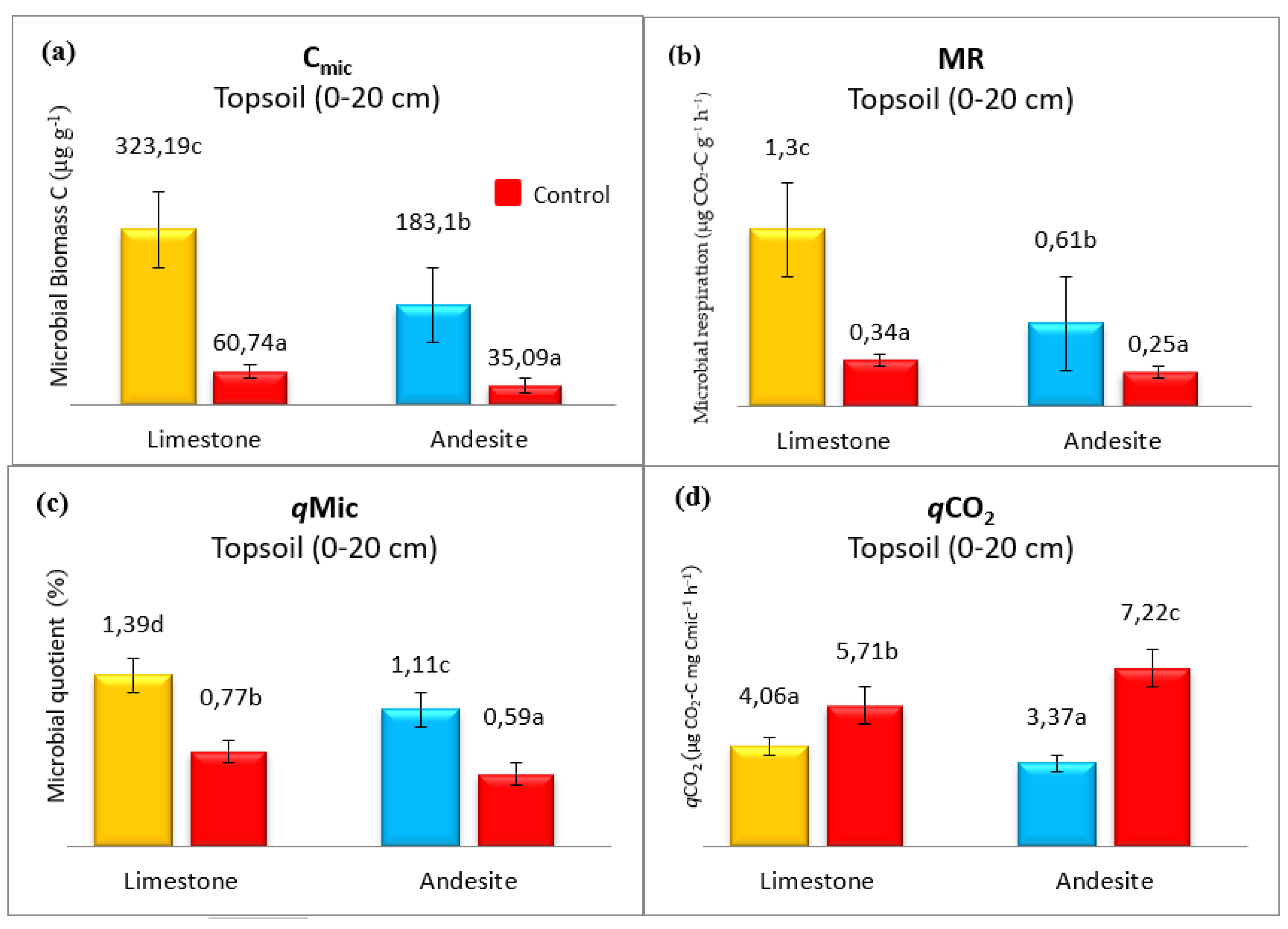
The significant changes were determined in the Cmic values of the areas after afforestation. The highest average Cmic values in soil samples were found in the LS site (323.19 g C kg−1), while the lowest was found in the AC site (35.09 g C kg−1) (Figure 2a). Again, The highest average microbial respiration rate (1.3 μg CO2–C g−1 h−1) from different soil samples was found in LS soil, followed by AS (0.61 μg CO2–C g−1 h−1), LC (0.34 μg CO2–C g−1 h−1) and AC (0.25 μg CO2–C g−1 h−1), respectively (Figure 2b). MR and Cmic values of soils increased by approximately 530% with afforestation on both bedrocks. The highest mean value of the Cmic/Corg (qMic) ratio was found in the LS site at 1.39, while the lowest value was observed in AC sites at 0.59 (Figure 2c). MR, Cmic, and qMic values were ranked from largest to smallest as LS > AS > LC > AC. The highest and lowest qCO2 values of soil samples were recorded in AC (7.22) and AS (3.37) sites, respectively (Figure 2d). The qCO2 values of control sites were found to be significantly higher than afforested sites. Unlike other microbial indices, the qCO2 index order is AC> LC> LS>AS.
Figure 2.
Changes in microbial biomass C (Cmic) (a), microbial respiration (MR) (b), Microbial quotient (Cmic/Corg= qMic) (c), and metabolic quotient (qCO2) (d) with afforestation and difference of parent material. Different letters above the bars indicate significant differences at p < 0.05 among the land use types.
Figure 2.
Changes in microbial biomass C (Cmic) (a), microbial respiration (MR) (b), Microbial quotient (Cmic/Corg= qMic) (c), and metabolic quotient (qCO2) (d) with afforestation and difference of parent material. Different letters above the bars indicate significant differences at p < 0.05 among the land use types.
3.3. Plant Growth Characteristics
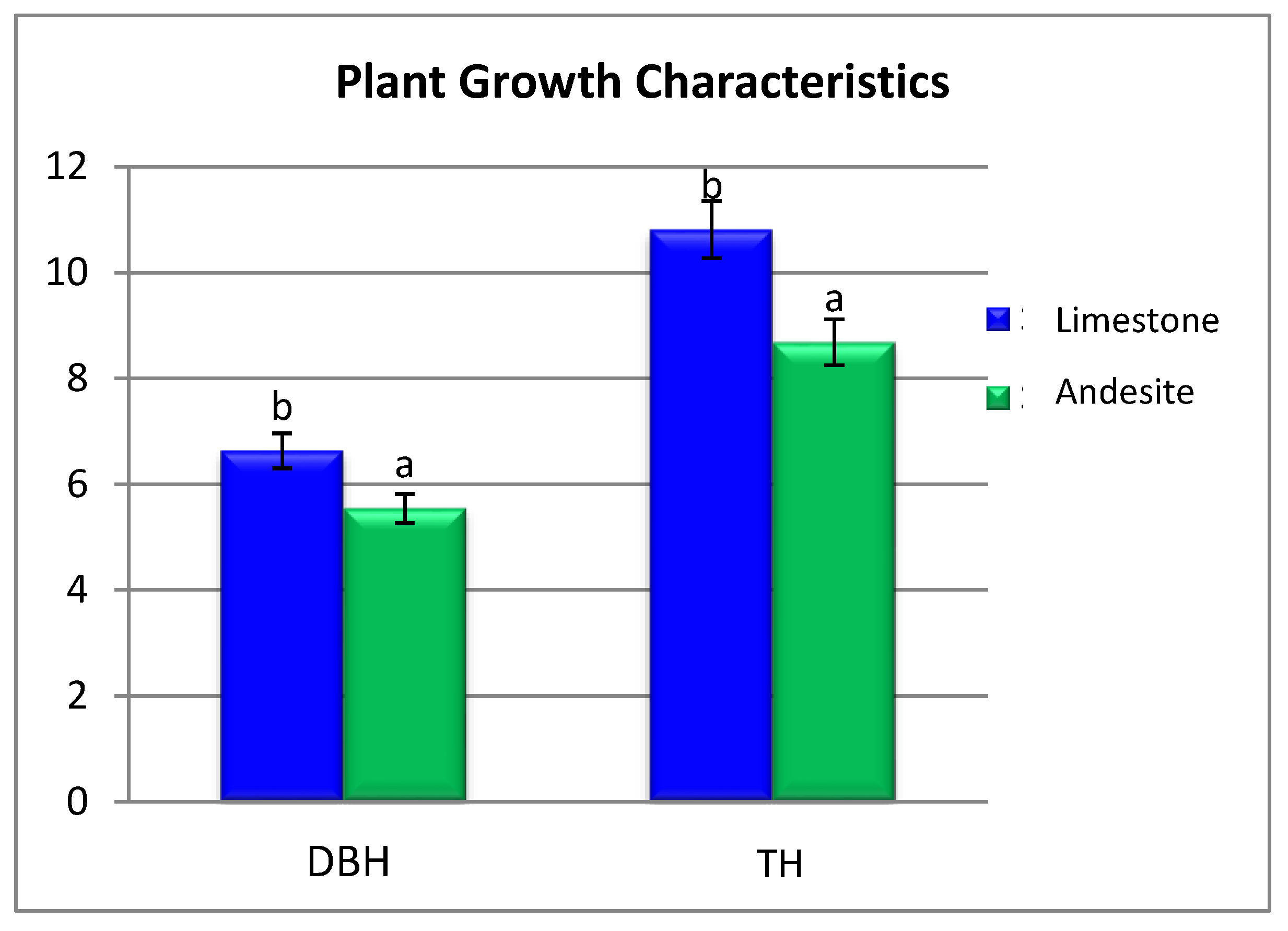
The average height of the saplings on limestone bedrock was 10.81 m, compared to 8.68 m on andesite parent material. Similarly, the average diameter at breast height (DBH) of saplings on limestone parent material was 6.63 m, while on andesite parent material; it was 5.54 m (Figure 2). These results indicate that, following afforestation, the average height and diameter of saplings on limestone parent material were 20% and 25% higher, respectively, than those on andesite parent material, despite the presence of the same tree species and ecological conditions.
Figure 2.
Changes in plant growth characteristics in different parent material sites. Different letters above the bars indicate significant differences at p < 0.05 among the land use types. DBH: Breast High Diameter and TH: Tree High.
Figure 2.
Changes in plant growth characteristics in different parent material sites. Different letters above the bars indicate significant differences at p < 0.05 among the land use types. DBH: Breast High Diameter and TH: Tree High.
The Pearson correlation coefficients between TH, DBH, Cmic, MR, qMic, qCO2, SOC, TN, pH, EC, CaCO3, sand, silt, and clay were calculated for both afforested and control areas (Table 3). It was determined that there were positive correlations between TH and dust, clay, pH, EC, SOC, TN, CaCO3, Cmic, and DBH (p ≤ 0.001). The highest positive correlations with TH were shown by clay, CaCO3, DBH, and Cmic, respectively. Again, the highest positive correlation values were found between DBH and CaCO3, Cmic, MR, and Clay, from largest to smallest, respectively. Here, it was determined that microbial parameters had a positive effect on plant growth parameters at least at the significance level of 0.05. In addition, all of the physicochemical properties of the soils, except sand and C/N ratio, showed positive correlations with plant growth parameters. Other interesting positive correlation values were found between Cmic, SOC and TN. On the other hand, the highest negative correlations were found between plant growth parameters and sand and C/N (Figure 3a).
In addition, when the relationships of soil properties in control areas belonging to different bedrocks were examined according to Pearson correlation, it was determined that sand, C/N, and qCO2 values had a significant negative relationship with clay, silt, pH, EC, CaCO3, Cmic, MR, qMic. Except for qCO2, the microbial indices of the soil showed the highest positive correlation with clay, dust, and CaCO3 among the physicochemical properties (p<0.001) (Figure 3b).
Discriminant analysis (DA) was able to distinguish between soils formed from limestone and andesite bedrocks in terms of physicochemical, microbiological properties, and plant growth parameters (Figure 4). The soil variables contributing to the classification and the values of the three canonical functions are shown in Table 4. Evaluations were made by accepting significant values above 0.3 [48]. Eigenvalues for the first 2 axes in all soil samples were 1.027 and 0.841, respectively; and explained 61.53% and 24.25% of the total variance (Figure 4a). Correlation coefficients show that DA axis 1 has the strongest correlation with qMic, C/N, qCO2, TH, DBH, and clay, respectively. DA axis 2 is seen to be mainly related to SOC, TN, Cmic, and MR, respectively, with the strongest correlation (Figure 4).
4. Discussion
In this study, it was revealed that physicochemical and microbiological soil properties as well as plant growth parameters were significantly affected by the type of parent material. Limestone soils formed fine-grained soils with significantly lower sand and higher silt and clay contents. Some studies indicated that soils developed from limestone bedrock had lower sand and higher clay and silt contents than other bedrock types such as granite and acrostic [4], and andesite [1]. Similarly, [3] determined that the grain sizes of soils from granite, granodiorite, and schist bedrocks were different.
The parent material difference also influenced the chemical properties of soils, such as pH, EC, SOC, TN, and CaCO3, in both control and afforestation sites. Previous studies have reported significant variations in soil pH values due to differences in parent material [1,3,4]. The EC values of soils from both parent materials at the study sites were below the critical salinity level of 0.20 dS cm−1. SOC and TN content differed significantly depending on the parent material and were higher in afforestation sites compared to control sites. The increase in carbon and nitrogen sources in soils after afforestation affects the quality and quantity of organic material, including litter production [49], root exudates [50], and microbial community structure and activity [15]. Plant residues such as leaves, cones, and branches contribute to increase SOC and TN levels in the ecosystem [32,51,52]. Moreover, factors like higher clay content, tree presence [53,54], root exudates, and plant residues [15] promote soil formation. While the effect of bedrock on soil properties was observed in control sites, afforestation, particularly with Scots pine, appears to have significantly contributed to the increases in SOC and TN. The concentration of SOC and TN in the study area may be lower than in mature forests, as only fifteen years have passed since afforestation. It may take a longer period for litter to accumulate on the soil surface and integrate into the soil. Typically, SOC and TN concentrations are higher in the topsoil layer (0-20 cm) due to litter accumulation [4,32]. As expected, CaCO3 content was significantly higher in soils with limestone parent material, attributed to the inherent lime content of limestone. There was a notable decrease in CaCO3 content after afforestation, likely due to plant influence on soil formation. It has been reported that afforestation with different species can reduce the amount of lime in the soil, particularly in the topsoil (Babur, 2019). Limestone soils are characterized by alkaline properties and texture variations between sand (sandstone) and clay (shale) [55].
The chemical properties of soils such as pH, SOC, and TN and microbial properties of Cmic, Nmic, MR, and other indices are important sensitive indicators affected by several factors such as seasonality, tree species, bedrock properties, and soil properties [16,56]. The available Cmic in soils directly indicates the microbial biomass pool in the soil. The study results illustrated that the parent material significantly affects the Cmic concentrations in soils. Mahia et al. [47] stated that schist was significantly different from granite parent material in Cmic, while Babur (2019) reported that soils developed from limestone parent material had higher Cmic than andesite. In addition, afforestation of bare land contributes to the nutrient cycle of the ecosystem, ecosystem productivity, soil quality, and the increase in the amount of soluble and storable organic C in the soil with the input of organic matter from litter and dead roots [57,58]. Increased organic carbon in soil also affects the biological properties of soils [59]. Soil Cmic is generally dependent on soil organic matter as a substrate [24]. In addition, exudates brought to the soil by plant roots can increase Cmic accumulation, especially in topsoil [60]. In this article, it was confirmed that the increased organic matter input after afforestation also significantly increased the Cmic amount.
Microbial respiration is the most important microbial factor in monitoring decomposition [35,46]. Soil MR indicates the oxidation status of the soil by microbes and is considered one of the most important and sensitive indicators of the carbon cycle in soils [61,62]. Babur et al. [32], in their study conducted in four different seasons, determined that m respiration in winter was 8.96 µg CO2–C g−1 h−1, which was higher than in summer (6.19 µg CO2–C g−1 h−1) and autumn (6.85 µg CO2–C g−1 h−1). In addition, in the same study, it was determined that there were significant differences in Cmic and MR values of different tree species. Similarly, it was stated that the optimum soil temperature and moisture content, which accelerate microbial activities in different tree plantations, were in rainy seasons [63].
The qMic values of all soil samples were found to be within the range suggested by Anderson and Domsch [64]. qMic can be used to quickly estimate the change in organic C [65] and to compare soil quality parameters in ecosystems with different organic matter [59]. The qMic ratio is determined by using the relationship between microbial biomass carbon and soil organic carbon [66]. The qMic ratio of andesite soils of the study areas was lower than that of limestone. This provided more suitable conditions for the growth of soil microorganisms in the limestone bedrock. Although the average qMic values increased after afforestation, this value remained low for fertile lands and did not reach equilibrium 15 years after afforestation. The qMic value is estimated by microbial activity, i.e., soil basal respiration and organic carbon cycling within an ecosystem [35,67]. Higher MR and Cmic amounts in limestone soils indicated that it had more microbial activity, proving faster decomposition.
The qCO2 is often used to describe the soil stress response of an ecosystem [67]. Environmental stresses, nutrient scarcity, and substrate resistance to decomposition can increase qCO2 [68]. On the other hand, the productivity of the area can be increased by using natural vegetation in stressed areas [69]. Although Anderson [67] reported that qCO2 values in neutral soils ranged from 0.5 to 2.0 μg CO2-C g−1Cmic h−1, in this study, the average qCO2 values in areas afforested with andesite and limestone bedrocks were found to be 3.37 to 4.06 μg CO2-C g−1Cmic h−1, respectively. This may cause microbial respiration to increase, decomposition to slow down and energy to be consumed in it. Low qCO2 rate in soils provides positive information about efficient carbon use for that soil, low environmental stress levels, usable nutrients being free in the area, microbial activity, and soil quality [22].
The qCO2 and OC mineralization rates in soil constitute the basis of the carbon cycle in terrestrial ecosystems [10,70]. SOC content negatively correlated with the metabolic coefficient in both parent materials. Some studies have noted that OC and qCO2 in different forest soils are closely related [1,60]. As a result of the correlation analysis, it was determined that basal respiration was positively correlated with soil organic carbon in both parent materials. Similar results were found with Cheng and Xia [71].
Differences in parent material and soil properties showed significant differences in plant growth parameters such as tree height and diameter at breast height. In particular, Scots pines on limestone bedrock were 20% and 25% larger in height and diameter than on andesite bedrock, respectively. There are many studies on the importance of the effects of habitat properties on plant yield and growth characteristics [72,73,74]. Microbiological, physical, and chemical soil properties can improve as the age of the afforested area. Afforestation and rehabilitation practices in forest management contribute to the health and productivity of soils and support the restoration of soil microbial activities.
Available C fractions are one of the most important environmental factors affecting soil microbial activity [22,75]. Analyses performed on soil samples, particularly microbial biomass, MR, and other indices, may be responsible for the separation of different parent materials and control areas with DA because the parent material is an important factor in soil formation, and tall plants also provide more living space and substrate for microbial communities in soils with their aboveground and belowground biomass and waste. Improvement of microorganisms’ living environments can also be understood by the increase in qMic value and decrease in qCO2 value [10,76]. In fact, from our research, it was determined that there was an increase in qMic amount and a decrease in qCO2 amount in soils on both bedrocks after afforestation. These results provide strong evidence that afforestation on different bedrocks results in different soil properties and microbial communities compared to bare lands.
5. Conclusion
In this study, it was determined that the physicochemical and microbiological properties of soils formed from different parent materials were different and this significantly affected the growth parameters of the planted seedlings. It was determined that the fertility of soils in terrestrial ecosystems was significantly affected primarily by the type of parent material and then by the presence of vegetation on it. The fact that limestone parent rock can be easily decomposed compared to andesite and that it creates deep and fertile soil allowed Scots pine trees to grow better on limestone parent rocks. It was shown that afforestation contributed to the increase in organic C and N, especially in the upper soils, in bare lands, and this contributed to the increase in microbial biomass and activity in the soils. Soils from limestone parent rocks afforested with Scots pine were energetically more efficient (had a lower qCO2) with more microbial biomass and a higher qMic (Cmic/Corg) compared to andesite soils. This study emphasized how important parent material is in terrestrial ecosystems and that it directly affects soil microbial activity. In particular, afforestation provides substrate and C to the ecosystem via tree litter, thus increasing soil microbial communities and activities. Since the measured microbial parameters are sensitive to changes in the ecosystem, they may have the potential to be used as indicators of forest management on soil organic matter quality. In order to ensure data continuity in such areas and to verify the data of afforestation sites in arid and semiarid areas, more comprehensive studies are required. In afforestation to be made on andesite bedrocks, more care should be taken, and maintenance, fertilization, and even irrigation during periods of water stress can contribute to the growth of seedlings and environmental fertility. In the afforestation of arid and semiarid areas, attention should be paid to the selection of local species and close origins. In addition, it would be more appropriate to make afforestation using mixed species with leaves and conifers to increase the ecological sensitivity limit. Thus, ecosystem diversity, nutrient cycle, and soil C input will be supported.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, E.B. and E.B.; methodology, E.B.; software, E.B.; validation, E.B.; formal analysis, E.B.; investigation, E.B.; resources, E.B.; data curation, E.B.; writing—original draft preparation, E.B.; writing—review and editing, E.B.; visualization, E.B.; supervision, E.B.; project administration, E.B.; funding acquisition, E.B. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Acknowledgments
Thank Mr. Muhammed PALA for helping me in my field study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Babur, E. Effects of parent material on soil microbial biomass carbon and basal respiration within young afforested areas. Scand. J. For. Res. 2019, 34, 94–101. [Google Scholar] [CrossRef]
- Lavee, H.; Imeson, A.C.; Sarah, P. The impact of climate change on geomorphology and desertification along a Mediterranean-arid transect. Land Degrad Dev. 1998, 9, 407–424. [Google Scholar] [CrossRef]
- Kooijman, A.M.; Jongejans, J.; Sevink, J. Parent material effects on Mediterranean woodland ecosystem in NE Spain. Catena. 2005, 59, 55–68. [Google Scholar] [CrossRef]
- Gökbulak, F.; Özcan, M. Hydro-physical properties of soils developed from different parent materials. Geoderma. 2008, 145, 376–380. [Google Scholar] [CrossRef]
- Birkeland, P.W. Pedology, Weathering and Geomorphological Research; Oxford University Press: New York, 1974. [Google Scholar]
- Johnson, D.L. Darwin would be proud: Bioturbation, dynamic denudation, and the power of theory in science. Geoarchaeology, 2002, 17, 7–40. [Google Scholar] [CrossRef]
- Gabet, E.J.; Reichmann, O.J.; Seabloom, E.W. The effects of bioturbation on soil processes and sediment transport. Annu. Rev. Earth Planet. Sci. 2003, 31, 249–273. [Google Scholar] [CrossRef]
- Phillips, J.D.; Marion, D.A. Biomechanical effects, litholological variability, and pedodiversity in some forest soils of Arkansas. Geoderma 2004, 124, 73–89. [Google Scholar] [CrossRef]
- Phillips, J.D.; Marion, D.A. Biomechanical Effects of Trees on Soil and Regolith: Beyond Treethrow, Ann. Assoc. Am. Geogr., 2006, 96, 233–247. [Google Scholar] [CrossRef]
- Kara, O.; Babur, E.; Altun, L.; Seyis, M. Effects of afforestation on microbial biomass C and respiration in eroded soils of Turkey. J. Sustain. For. 2016, 35, 385–396. [Google Scholar] [CrossRef]
- Anderson, D.W. The effect of parent material and soil development on nutrient cycling in temperate ecosystems. Biogeochem. 1988, 5, 71–97. [Google Scholar] [CrossRef]
- Turkish Ministry of Agriculture and Forestry, The afforestation and erosion control national mobilization action plan (2008-2012) and combating with erosion action plan (2013- 2017) Ankara, Turkey, 2008, (accesed in 2024).
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science, 2005, 310, 1628–1632. [Google Scholar] [CrossRef]
- Silver, W.L.; Kueppers, L.M.; Lugo, A.E.; Ostertag, R.; Matzek, V. Carbon sequestration and plant community dynamics following reforestation of tropical pasture. Ecol Appl 2004, 14, 1115–1127. [Google Scholar] [CrossRef]
- Grayston, S.J.; Vaughan, D.; Jones, D. Rhizosphere carbon flow in trees, in comparison with annual plants: The importance of root exudation and its impact on microbial activity and nutrient availability. Appl Soil Ecol. 1996, 5, 29–56. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroglu, T.; Danish, S.; Haggblom, M.M.; Ozlu, E.; Gozukara, G.; Uslu, O.S. Spatial responses of soil carbon stocks, total nitrogen, and microbial indices to post-wildfire in the Mediterranean red pine forest. J. Environ. Manag. 2022, 320, 115939. [Google Scholar] [CrossRef]
- Bauhus, J.D.; Pare, D.; Cote, L. Effects of tree species, stand age, and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol Biochem, 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Kara, O.; Bolat, I. The effect of different land uses on soil microbial biomass carbon and nitrogen in Bartin Province. Turk. J. Agric. For. 2008, 32, 281–288. [Google Scholar]
- Yang, Y.; Guo, J.; Chen, G.; Yin, Y.; Gao, R.; Lin, C. Effects of forest conversion on soil labile organic carbon fractions and aggregate stability in subtropical China. Plant Soil. 2009, 323, 153–162. [Google Scholar] [CrossRef]
- Pandey, C.B.; Singh, G.B.; Singh, S.K.; Singh, R.K. Soil nitrogen and microbial biomass carbon dynamics in native forests and derived agricultural land uses in a humid tropical climate of India. Plant Soil. 2010, 333, 453–467. [Google Scholar] [CrossRef]
- Korkanç, S.Y. Effects of afforestation on soil organic carbon and other soil properties. Catena. 2014, 123, 62–69. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroglu, T.; Solaiman, Z.M.; Battaglia, M.L. Microbial Respiration, Microbial Biomass and Activity Are Highly Sensitive to Forest Tree Species and Seasonal Patterns in the Eastern Mediterranean Karst Ecosystems. Sci Total Environ. 2021, 775, 145868. [Google Scholar] [CrossRef]
- Wang, B.; Jiang, Y.; Wei, X.; Zhao, G.; Guo, H.; Bai, X. Effects of forest type, stand age, and altitude on soil respiration in subtropical forests of China. Scand J Forest Res. 2011, 26, 40–47. [Google Scholar] [CrossRef]
- Babur, E.; Kara, O.; Fathi, R.; Susam, Y.; Riaz, M.; Arif, M.; Akhtar, K. Wattle fencing improved soil aggregate stability, organic carbon stocks and biochemical quality by restoring highly eroded mountain region soil. J. Environ. Manag. 2021, 288, 112489. [Google Scholar] [CrossRef]
- Singh, B.K.; Munro, S.; Potts, J.M.; Millard, P. Influence of grass species and soil type on rhizosphere microbial community structure in grassland soils. Appl Soil Ecol, 2007, 36, 147–155. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Macdonald, L.M.; Rogers, S.L.; Gregg, A.L.; Bolger, T.P.; Baldock, J.A. Habitat selective factors influencing the structural composition and functional capacity of microbial communities in agricultural soils. Soil Biol Biochem, 2008, 40, 803–813. [Google Scholar] [CrossRef]
- Bezemer, T.M.; Lawson, C.S.; Hedlund, K.; Edwards, A.R.; Brook, A.J.; Igual, J.M.; Van Der Putten, W.H. Plant species and functional group effects on abiotic and microbial soil properties and plant-soil feedback responses in two grasslands. J Ecol, 2006, 94, 893–904. [Google Scholar] [CrossRef]
- Lamb, E.G.; Kennedy, N.; Siciliano, S.D. Effects of plant species richness and evenness on soil microbial community diversity and function. Plant Soil, 2011, 338, 483–495. [Google Scholar] [CrossRef]
- Fernández-Ondoño, E.; Rojo Serrano, L.; Jiménez, M.N.; Navarro, F.B.; Díez, M.; Martín, F.; Aguilar, J. Afforestation improves soil fertility in south-eastern Spain. Eur J For Res, 2010, 129, 707–717. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For Ecol Manag, 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Korkanç, S.Y. Effects of afforestation on soil organic carbon and other soil properties. Catena. 2014, 123, 62–69. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroğlu, T.; Riaz, M.; Uslu, O.S. Seasonal Variations in Litter Layers’ Characteristics Control Microbial Respiration and Microbial Carbon Utilization Under Mature Pine, Cedar, and Beech Forest Stands in the Eastern Mediterranean Karstic Ecosystems. Microb. Ecol. 2022, 84, 153–167. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Tüfekçioglu, A.; Küçük, M. Comparison of Decomposition Rates of Beech (Fagus orientalis Lipsky) and Spruce (Picea orientalis (L.) Link) Litter in Pure and Mixed Stands of Both Species in Artvin, Turkey. Turk J Agric For 2005, 29, 429–438. [Google Scholar]
- Babur, E.; Dindaroğlu, T.; Roy, R.; Seleiman, M.F.; Ozlu, E.; Battaglia, M.L.; Uslu, Ö.S. Relationship between organic matter and microbial biomass in different vegetation types. In Microbial Syntrophy-Mediated Eco-Enterprising; Elsevier: Amsterdam, The Netherlands, 2022; pp. 225–245. [Google Scholar]
- Winding, A.; Hund-Rinke, K.; Rutgers, M. The use of microorganisms in ecological soil classification and assessment concepts. Ectoxicol Environ Saf 2005, 62, 230–248. [Google Scholar] [CrossRef] [PubMed]
- Akburak, S.; Son, Y.; Makineci, E.; Çakir, M. Impacts of Low-Intensity Prescribed Fire on Microbial and Chemical Soil Properties in a Quercus Frainetto Forest, J. For. Res. 2018. 29, 687–696. [CrossRef]
- Bolat, I. Microbial Biomass, Basal Respiration, and Microbial Indices of Soil in Diverse Croplands in a Region of Northwestern Turkey (Bartın), Environ. Monit. Assess. 2019. 191, 695.
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Walkley, A.; Black, A.I. An examination of the Degtjareff method for determining soil organic matter, and proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-total. In Methods of soil analysis, Part 2 Chemical and Microbiological Properties (Ed. A.L. Page). SSSA Book series No: 9, Madison, 1982; pp. 595-622.
- Rowell, D.L. Soil science; methods and applications; Longman Publishers (Pte) Ltd.: Singapore, 1994; p. 350. [Google Scholar]
- Horwath, W.R.; Paul, E.A. Microbial biomass. In methods of soil analysis, part 2. microbiological and biochemical properties-SSSA book series, no. 5; Soil Science Society of America: Madison, WI, USA, 1994; pp. 753–773. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform Fumigation and the Release of Soil Nitrogen: A Rapid Direct Extraction Method to Measure Microbial Biomass Nitrogen in Soil, Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Dilly, O.; Munch, J.C. Ratios between estimates of microbial biomass content and microbial activity in soils. Biol Fertil Soils. 1998, 27, 374–379. [Google Scholar] [CrossRef]
- Alef, K. Soil respiration. In Methods in applied soil microbiology and biochemistry; Alef, K., Nannipieri, P., Eds.; Academic: London, 1995; pp. 214–219. [Google Scholar]
- Mahía, J.; Pérez-Ventura, L.; Cabaneiro, A.; Díaz-Raviña, M. Soil microbial biomass under pine forests in the north-western Spain: influence of stand age, site index and parent material. Sist Rec. Forest. 2006, 15, 152–159. [Google Scholar] [CrossRef]
- Hair, J.F., Jr.; Anderson, R.E.; Tatham, R.L.; Black, W.C. Multivariate data analysis: With readings; Macmillan: New York, NY, 1992. [Google Scholar]
- Paul, K.I.; Polglase, P.J.; Nyakuengama, J.G.; Khanna, P.K. Change in soil carbon following afforestation. Forest Ecol Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroglu, T. Seasonal changes of soil organic carbon and microbial biomass carbon in different forest ecosystems. Environmental Factors Affecting Human Health, Ivan Uher, IntechOpen 2020.
- Rothe, A.; Cromack, K., Jr.; Resh, S.C.; Makineci, E.; Son, Y. Soil carbon and nitrogen changes under Douglas-fir with and without red alder. Soil Sci Soc Am J 2002, 66, 1988–1995. [Google Scholar] [CrossRef]
- Abdalmoula, M.M.; Makineci, E.; Özturna, A.G.; Pehlivan, S.; Şahin, A.; Tolunay, D. Soil organic carbon accumulation and several physicochemical soil properties under stone pine and maritime pine plantations in coastal dune, Durusu-Istanbul. Environ Monit Assess 2019, 191, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Oostra, S.; Majdi, H.; Olsson, M. Impact of tree species on soil carbon stocks and soil acidity in southern Sweden. Scand J Forest Res. 2006, 21, 364–371. [Google Scholar] [CrossRef]
- Yildiz, O.; Esen, D.; Karaoz, O.M.; Sarginci, M.; Toprak, B.; Soysal, Y. Effects of different site preparation methods on soil carbon and nutrient removal from Eastern beech regeneration sites in Turkey’s Black Sea region. Applied Soil Ecology 2010, 45, 49–55. [Google Scholar] [CrossRef]
- Kantarcı, D. Soil Science Lessons Book; Istanbul University, Department of Soil Science and Ecology, Publications of Istanbul University, No. 4261, Faculty of Forestry No. 462: Istanbul, 2000; p. 420. [Google Scholar]
- Das, S.; Deb, S.; Sahoo, S.S.; Sahoo, U.K. Soil Microbial Biomass Carbon Stock and Its Relation with Climatic and Other Environmental Factors in Forest Ecosystems: A Review. Acta Ecologica Sinica 2023, 933–945. [Google Scholar] [CrossRef]
- Yüksek, T. The restoration effects of black locust (Robinia pseudoacacia L) plantation on surface soil properties and carbon sequestration on lower hillslopes in the semi-humid region of Coruh Drainage Basin in Turkey. Catena 2012, 90, 18–25. [Google Scholar] [CrossRef]
- Uslu, O.S.; Babur, E.; Battaglia, M.L.; et al. Effects of gyttja applications on hay yield and quality of a rangeland in the Mediterranean region. Int. J. Environ. Sci. Technol. 2023, 20, 4139–4150. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L. Soil organic matter under different forest types in Southern China. Geoderma. 2007, 142, 349–356. [Google Scholar] [CrossRef]
- Tian, J.; Fan, M.; Guo, J.; Marschner, P.; Li, X.; Kuzyakov, Y. Effects of land use intensity on dissolved organic carbon properties and microbial community structure. Euro J Soil Biol. 2012, 52, 67–72. [Google Scholar] [CrossRef]
- Soong, J.L.; Fuchslueger, L.; Marañon-Jimenez, S.; Torn, M.S.; Janssens, I.A.; Penuelas, J.; Richter, A. Microbial Carbon Limitation: The Need for Integrating Microorganisms into Our Understanding of Ecosystem Carbon Cycling, Glob. Chang. Biol. 2020, 26, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Tomar, U.; Baishya, R. Seasonality and Moisture Regime Control Soil Respiration, Enzyme Activities, and Soil Microbial Biomass Carbon in a Semi-Arid Forest of Delhi, India. Ecol Process 2020, 9, 50. [Google Scholar] [CrossRef]
- Edwards, C.A.; Arancon, N.Q. The Role of Earthworms in Organic Matter and Nutrient Cycles. In Biology and Ecology of Earthworms; Springer US: New York, NY, 2022; pp. 233–274. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. Ratios of microbial biomass carbon to total organic carbon in arable soils. Soil Biol Biochem. 1989, 21, 471–479. [Google Scholar] [CrossRef]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- Insam, H.; Domsch, K.H. Relationship between soil organic carbon and microbial biomass on chronosequences of reclamation sites. Microb Ecol. 1988, 15, 177–188. [Google Scholar] [CrossRef]
- Anderson, T.H. Microbial eco-physiological indicators to assess soil quality. Agr Ecosyst Environ. 2003, 98, 285–293. [Google Scholar] [CrossRef]
- Spohn, M. Microbial respiration per unit microbial biomass depends on litter layer carbon-to-nitrogen ratio. Biogeoscience. 2015, 12, 817–823. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A.; Elser, J. Carbon use efficiency of microbial communities: stoichiometry, methodology and modeling. Ecol Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef]
- Hartman, W.H.; Ye, R.; Horwath, W.R.; Tringe, S.G. A genomic perspective on stoichiometric regulation of soil carbon cycling. ISME J. 2017, 11, 2652–2665. [Google Scholar] [CrossRef]
- Cheng, Y.B.; Xia, Y.D. Soil microbial and enzymatic activities across a chronosequence of Chinese Pine plantation development on the Loess Plateau of China. Pedosphere. 2012, 22, 1–12. [Google Scholar]
- Khalil, H.A.; Hossain, M.S.; Rosamah, E.; Azli, N.A.; Saddon, N.; Davoud Poura, Y.; Islam, M.N.; Dungani, R. The role of soil properties and it’s interaction towards quality plant fiber: a review. Renew. Sustain. Energy Rev. 2015, 43, 1006–1015. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Rossi, E.; Milbau, A.; Nijs, I. Habitat properties and plant traits interact as drivers of non-native plant species’ seed production at the local scale. Ecol. Evol. 2018. 8, 4209–4223. [CrossRef]
- Martins-Noguerol, R.; Matías, L.; Pérez-Ramos, I.M.; Moreira, X.; Francisco, M.; Pedroche, J.; DeAndrés-Gil, C.; Gutiérrez, E.; Salas, J.J.; Moreno-Pérez, A.J.; et al. Soil Physicochemical Properties Associated with the Yield and Phytochemical Composition of the Edible Halophyte Crithmum Maritimum. Sci. Total Environ. 2023, 869, 161806. [Google Scholar] [CrossRef] [PubMed]
- Yüksek, T. The restoration effects of black locust (Robinia pseudoacacia L) plantation on surface soil properties and carbon sequestration on lower hillslopes in the semi-humid region of Coruh Drainage Basin in Turkey. Catena 2012, 90, 18–25. [Google Scholar] [CrossRef]
- Sarkara, S.; Dasb, D.K.; Singhc, A.; Laikd, R.; et al. Seasonal variations in soil characteristics control microbial respiration and carbon use under tree plantations in the Middle Gangetic region. Heliyon. 2024. [Google Scholar] [CrossRef]
Figure 3.
Correlation maps of soil samples of different parent material planted (a) and control (b) sites according to correlation coefficient matrix (R-values) for physical, chemical, and microbiological characteristics of soils in different land uses.
Figure 3.
Correlation maps of soil samples of different parent material planted (a) and control (b) sites according to correlation coefficient matrix (R-values) for physical, chemical, and microbiological characteristics of soils in different land uses.
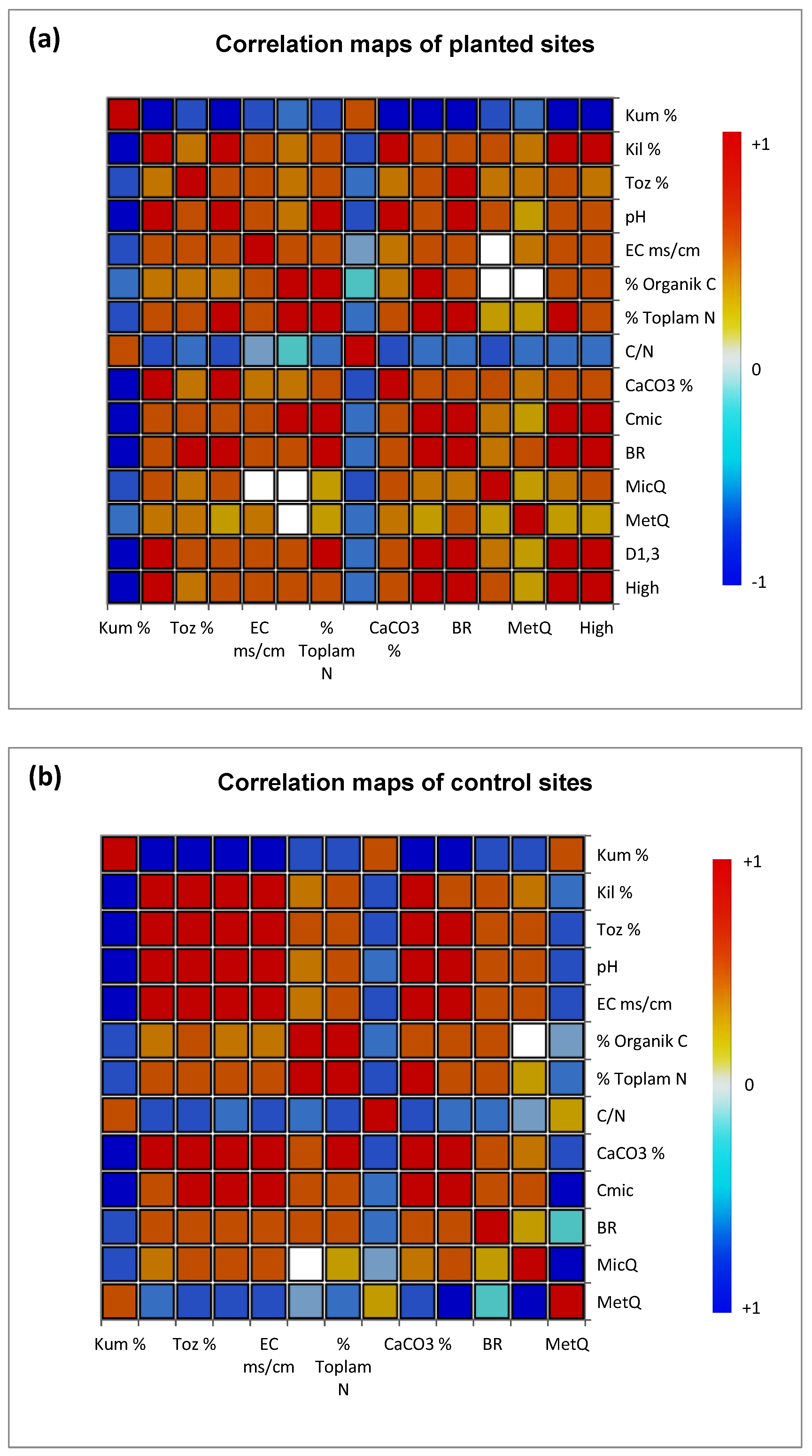
Figure 4.
Scatter plot of the first two discriminant function values in soil samples.
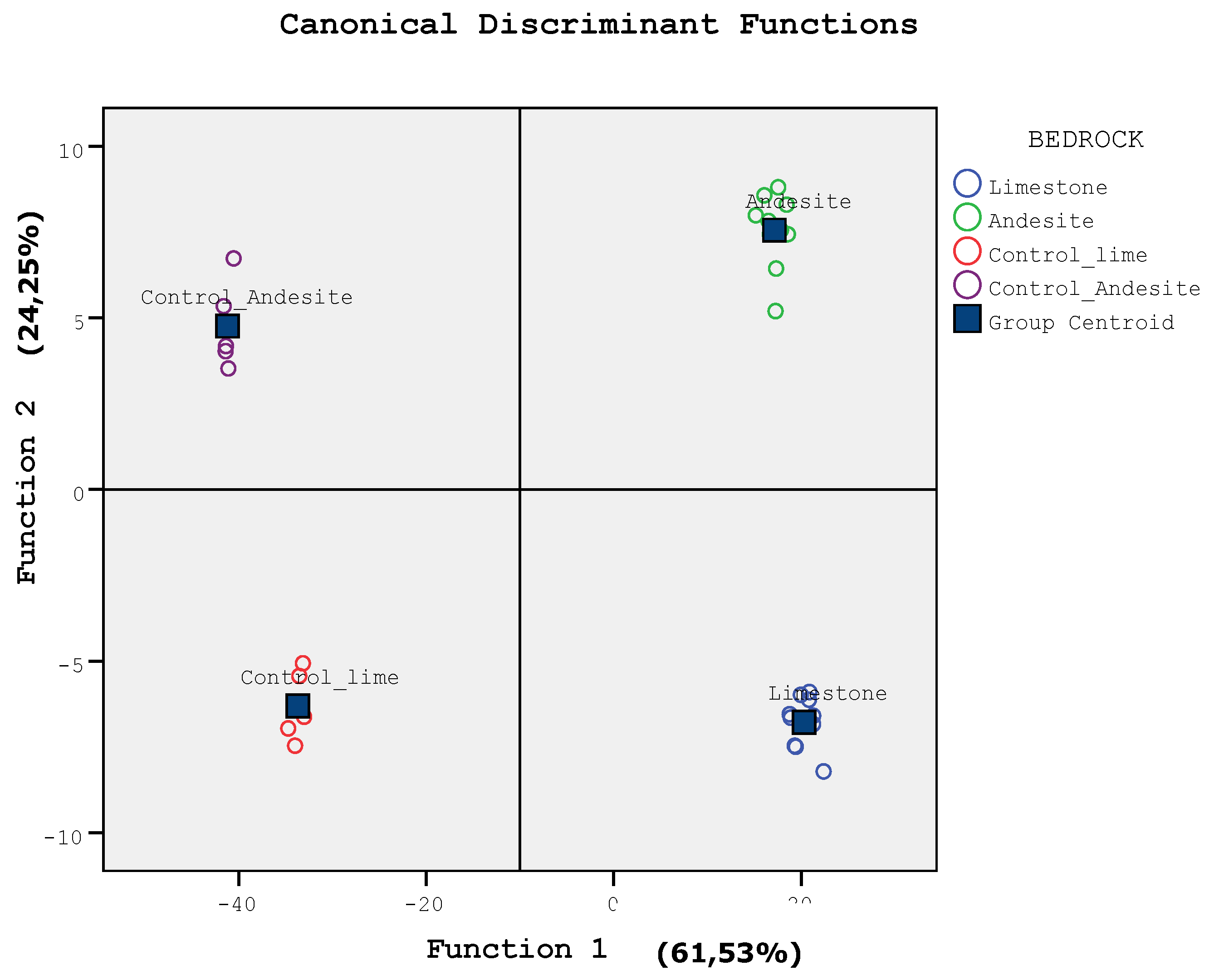

Table 1.
Study site characteristics.
| Environmental Factors | Limestone | Andesite | ||
|---|---|---|---|---|
| Sample | Control | Sample | Control | |
| Longitude | 39o 33′ 19′’ | 39o 33′ 46′’ | 39o 33′ 43′’ | 39o 33′ 23′’ |
| Latitude | 40o 21′ 08′’ | 40o 21′ 23′’ | 40o 21′ 20′’ | 40o 21′ 07′’ |
| Altitude mean (m) | 1480 | 1520 | 1500 | 1450 |
| Slope degree mean (%) | 50 | 40 | 45 | 50 |
| Afforestation date | 2005 | - | 2005 | - |
| Vegetation type | Scotch Pine | Bare space | Scotch Pine | Bare space |
| Stand canopy (%) | 70-100 | - | 70-100 | - |
| Soil texture | Loamy clay | Clay loam | Sandy clay loam | Sandy loam |
| pH class | Moderately Alkaline | Slightly Alkaline | Moderately Alkaline | Slightly Alkaline |
Table 2.
F statistics of the one-way ANOVA for the effects of parent material on soil physicochemical, microbial properties, and plant growth characteristics.
Table 2.
F statistics of the one-way ANOVA for the effects of parent material on soil physicochemical, microbial properties, and plant growth characteristics.
| Sand | Silt | Clay | pH | EC | SOC | TN | C/N | CaCO3 | Cmic | MR | qMic | qCO2 | DBH | TH | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parent Material | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| F-values | 355.8 | 42.6 | 117,4 | 60,5 | 9.8 | 21.5 | 48.8 | 24.3 | 123.2 | 84.3 | 62.6 | 44.5 | 27.9 | 56.0 | 85.6 |
Abbreviations: EC=electrical conductivity (dSm−1); TN=total soil N (%); SOC=soil organic carbon (%); Cmic=carbon in the microbial biomass (μg g−1); MR= microbial basal respiration (μg CO2–C g−1 soil h−1); qMic = microbial quotient; qCO2 = metabolic quotient; DBH = diameter at breast high and TH= tree high. a No significant differences are indicated by ns. * Significant at P ≤ 0.05. ** Significant at P ≤ 0.01. *** Significant at P ≤ 0.001.
Table 3.
Selected soil characteristics developed from different parent materials at the topsoil depths.
Table 3.
Selected soil characteristics developed from different parent materials at the topsoil depths.
| Parent Material | |||||
|---|---|---|---|---|---|
| Limestone (n = 30) | Andesite (n = 30) | ||||
| Sample | Control | Sample | Control | ||
| SOIL PHYSICOCHEMICHAL PROPERTIES | Sand (%) | 53,50±0,94 a | 59,74±1,00 b | 72,58±,13 c | 75,28±1,55 d |
| Silt (%) | 15,40±2,89 b | 22,08±0,91 c | 10,60±1,06 a | 13,66±0,87 b | |
| Clay (%) | 31,10±2,65 c | 18,18±1,68 b | 16,82±2,15 b | 11,02±1,71 a | |
| pH (H2O) | 8,05±0,1b | 8,18±0,09 b | 7,59±0,09 a | 7,57±0,15 a | |
| EC (mS cm−1) | 0,17±0,04 b | 0,18±0,02 b | 0,12±0,02 a | 0,13±0,01 a | |
| SOC (%) | 2,40±0,8 c | 0,80±0,11 a | 1,65±0,05 b | 0,61±0,12 a | |
| TN (%) | 0,19±0,05 c | 0,04±0,00 a | 0,10±0,02 b | 0,03±0,01 a | |
| C/N | 12,61±1,88 a | 18,76±1,20 b | 16,22±2,39 b | 22,74±3,31 c | |
| CaCO3 (%) | 23,09±3,42 b | 32,90±6,31 b | 3,08±1,24 a | 4,85±2,04 a | |
| SOIL MICROBIAL PROPERTIES | Cmic | 323,19±64,96 c | 60,74±9,03 a | 183,10±9,31 b | 35,09±3,65 a |
| MR | 1,3±0,24 c | 0,34±0,04 a | 0,61±0,15 b | 0,25±0,024 a | |
| qMic | 1,39±0,20 d | 0,77±0,12 b | 1,11±0,08 c | 0,59±0,09 a | |
| qCO2 | 4,06±0,65 a | 5,71±1,23 b | 3,37±0,85 a | 7,22±0,67 c | |
| GROWTH PROPERTIES | DBH | 6,63±0,26 b | - | 5,54±0,38 a | - |
| TH | 10,81±0,70 b | - | 8,68±0,19 a | - | |
Table 4.
Standardized Canonical Discriminant Function Coefficients of soil properties and plant growth characteristics.
Table 4.
Standardized Canonical Discriminant Function Coefficients of soil properties and plant growth characteristics.
| Function | |||
| Soil variables | 1 | 2 | 3 |
| TH | 1,195 | 0,330 | 0,384 |
| DBH | 1,233 | -0,133 | -0,510 |
| Sand | -0,613 | 0,900 | -0,258 |
| Clay | -0,906 | 0,000 | -0,483 |
| pH | 0,262 | 0,103 | 0,779 |
| EC | -1,798 | -0,646 | -1,695 |
| SOC | 17,718 | 2,887 | 17,085 |
| TN | 0,936 | -0,066 | -2,746 |
| C/N | -0,512 | -0,197 | -1,344 |
| CaCO3 | -0,030 | -0,258 | 0,436 |
| Cmic | -14,303 | -2,517 | -11,114 |
| MR | -0,434 | 0,412 | 0,607 |
| qMic(Cmic/Corg) | 7,158 | 0,590 | 5,012 |
| qCO2 | 0,958 | 0,069 | 0,004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



