Submitted:
07 August 2024
Posted:
11 August 2024
You are already at the latest version
Abstract
Antimicrobial peptides (AMPs) are short, usually cationic peptides with an amphiphilic structure, which allows them to easily bind and interact with the cellular membranes of viruses, bacteria, fungi, and other pathogens. Bacterial AMPs, or bacteriocins, can be produced from Gram-negative and Gram-positive bacteria via ribosomal synthesis to eliminate competing organisms. Bacterial AMPs are vital in addressing the increasing antibiotic resistance of various pathogens, potentially serving as an alternative to ineffective antibiotics. Bacteriocins have a narrow spectrum of action, making them highly specific antibacterial compounds that target particular bacterial pathogens. This review covers the two main groups of bacteriocins produced by Gram-negative and Gram-positive bacteria, their modes of action, classification, sources of positive effects they can play on the human body, and their limitations and future perspectives as an alternative to antibiotics.
Keywords:
bacterial AMPs
; bacteriocins
; antibiotic resistance
; limitations
; perspectives
; modes of action
1. Introduction
Certain bacteria produce bacterial antimicrobial peptides (AMPs) through the ribosomal pathway and are typically effective against closely related bacteria. In 1922, Alexander Fleming identified the first AMP and named it Lysozyme. After its discovery in 1928, Fleming also discovered the first antibiotic - penicillin. Florey, Chain, and Fleming brought the potential of penicillin in medical use into fruition and won the 1945 Nobel Prize in Medicine. In the 1940s and at the beginning of the golden age of antibiotics, the interest in using natural AMPs as therapeutics was lost. In the 1960s, the interest in AMPs as host defense molecules was renewed due to the continuous rise of multidrug-resistant microbial pathogens. In 1981, α-helical AMPs named cecropins, isolated from the Cecropia silk moth, were discovered, followed by magainin from the African clawed frog Xenopus laevis in 1987. In the 1990s, the field of antimicrobial peptides expanded rapidly, reporting over 300 peptides. AMPs have been broadly identified and characterized in all living organisms [1].
Some bacteriocins exhibit a broad inhibitory range. The first bacteriocin, colicin, was identified in 1925 from an Escherichia coli strain. Many bacteriocins have been discovered since then, resulting in a diverse group of proteins in size, target, mode of action, delivery, and immunity mechanisms [2].
In 1939, René Dubos isolated gramicidin, a natural peptide from Bacillus brevis [3]. This peptide was effective against a broad spectrum of Gram-positive and Gram-negative bacteria and was the initial peptide-based topical antibiotic. Nevertheless, the enthusiasm for peptide drugs grew in the early 1980s following the isolation and characterization of cecropins and magainins [4].
Antibiotic-resistant pathogens have become an urgent contemporary problem as many resistant strains continue to emerge [5]. Antimicrobial peptides can be considered alternative therapeutic agents and may be crucial for the fight against antibiotic resistance [6,7]. Figure 1 presents the dynamics of the total number of papers in the field for the last 10 years. The graphic shows the constant interest of researchers in AMPs, which has been accelerating since 2017.
Over 3000 AMPs have been contributed to the Antimicrobial peptide database [8]. AMPs play a crucial role in the innate immune defense of organisms [9]. They are typically small molecules (10-100 amino acids) with an overall positive charge ranging from +2 to +9 and possess amphiphilic properties [10]. Depending on their target, AMPs are classified as antibacterial, antifungal, antiparasitic, and antiviral peptides [11]. The most prevalent AMPs are cationic peptides that exhibit a variety of secondary structures such as α-helices, β-sheets (Figure 2) with two or more disulfide bridges, loops with a single disulfide bond, and extended structures containing specific amino acids like proline, arginine, tryptophan, and glycine [12,13].
Bacteriocins have been identified and isolated from both Gram-positive and Gram-negative bacteria [14]. These peptides play a significant part in supporting bacteria within a bacterial cell community and typically demonstrate antimicrobial solid effects on other bacteria of similar or different genera [15].
2. Classification
Bacteriocins are antimicrobial peptides produced by nearly all prokaryotic lineages via ribosomal synthesis to eliminate competing organisms. Bacteriocins derived from Gram-positive bacteria are generally more prevalent than those from Gram-negative bacteria and Archaea [16]. Bacteriocins are classified based on the cell wall type of producing organisms (Gram-negative and Gram-positive). Several bacteriocins produced by the Archaea domain representatives have also been characterized, such as halocins and sulfolobaceae [17].
2.1. Bacterial AMPs Produced by Gram-Negative Bacteria
Bacteriocins display a wide range of size, structure, and mode of action variations. E. coli and other enterobacteria have been the primary sources of characterized bacteriocins in Gram-negative bacteria [15]. Gram-negative bacteria produce four main classes of bacteriocins: colicins, colicin-like, phage-tail-like bacteriocins, and microcins.
2.1.1. Colicins
Colicins are high-molecular-weight (30–80 kDa) bactericidal proteins sensitive to proteases and heat. They are synthesized by most E. coli strains carrying a single colicinogenic plasmid. These compounds are widely studied and serve as model systems for investigating bacteriocins’ structures, functions, and evolution. Colicin synthesis occurs under stress and is lethal for the producing cells due to co-expression with lysis protein [18].
2.1.2. Phage Tail-Like
Phage tail-like bacteriocins, or tailocins, are substantial protein structures ranging from 20–100 kDa and composed of 8 to 14 distinct polypeptide subunits. These structures exhibit similarities to the bacteriophage tail modules. Tailocins are present in bacterial genomes as a cluster of genes spanning over 40 kbp. This genetic locus encompasses genes responsible for encoding structural proteins, assembly enzymes, chaperones, regulatory genes, and lysis cassettes, all of which function to release bacteriocins into the surrounding environment [19]. Bacteriocins of this category are categorized into two groups—R and F. R-type tailocins are evolutionarily linked to the tails of phages in the Myoviridae family and form a lengthy tube enclosed by a shell, with a complex basal plate featuring receptor-binding proteins (RBPs) at one end. On the other hand, F-type bacteriocins, which belong to the tails of phages in the Siphoviridae family, lack a shell.
2.1.3.М. icrocins
The third type of bacteriocins produced by Gram-negative bacteria is microcins. These are low-molecular-weight (<10 kDa) and highly stable peptides involved in competitive interactions between members of the Enterobacteriaceae family. These bacteriocins are resistant to proteases and extreme pH and temperature values. Clusters of genes encoding microcins are located in plasmids and, less often, in genomic DNA [15]. They include a variable number of genes but demonstrate a conservative organization: open reading frames encoding a microcin precursor, secretion proteins, immune factors, and, in some cases, post-translational modification enzymes. Microcins are divided into two classes based on the presence, nature, and localization of modifications, as well as the gene cluster organization and the sequence of leader peptides [14].
Class I includes peptides with a molecular weight below 5 kDa (microcins B17, C7-C51, and J25) and complex post-translational modifications. Bacteriocins of this group inhibit vital bacterial enzymes, such as DNA gyrases I and II 68 S and RNA polymerases, and can also inhibit the respiratory chain [20]. Class II includes larger (5–10 kDa) plasmid peptides without post-translational modifications (MccL, MccV, and Mcc24) and linear microcins encoded in the chromosome and carrying a C-terminal siderophore (microcins E492, M, H47, I47, and G47). Their toxic effect is forming pores and destroying the target pathogenic bacteria’s cell membrane [21]. The structure of Microcin 7 is shown in Figure 3.
2.2. Bacterial AMPs Produced by Gram-Positive Bacteria
Most bacteriocins of Gram-positive bacteria refer to two main classes. Class I includes lantibiotics and class II—small post-translationally unmodified bacteriocins. Lantibiotics are peptides that undergo extensive post-translational modifications and contain lanthionine residues and methyllanthionine. In the past decade, it has become clear that this class of ribosomally synthesized and post-translationally modified peptides (RiPPs) is more diverse [17]. The third class of bacteriocins previously included large (more than 10 kDa) antibacterial proteins and bacteriolysins, as well as recently identified tailocins of Gram-positive bacteria.
2.2.1. Lantibiotics
Lantibiotics are low-molecular-weight peptides less than 5 kDa in size. They give rigidity to the structure of bacteriocin and resistance to the action of proteases. Based on the biosynthesis specifics, lantipeptides are divided into four classes, two of which include compounds with antibacterial activity. Based on the structure features, we can distinguish three types of lantibiotics—AI, AII, and B corresponding to bacteriocins with linear, globular, and combined conformation [23]. Gram-positive bacteria produce two main types of bacteriocins: lantibiotics (class I) and non-lantibiotics (class II). Lantibiotics are short peptides consisting of 19–38 amino acids and undergo post-translational modifications, including thioether-based ring structures known as lanthionine or beta-methyllanthionine [24]. Some lantibiotics may also contain unique modified amino acids, such as D-alanine in lactocin S. Penisin, isolated from Paenibacillus ehimensis shows activity against methicillin-resistant Staphylococcus aureus (MRSA). Unlike other lantibiotics, it effectively inhibits the growth of Gram-negative bacteria [25]. Due to their diverse structures, it is challenging to classify lantibiotics into subclasses. About 11 subclasses have been proposed for lantibiotics. Nisin, subtilin, lacticin 3147, and thuricin CD belong to class I, with nisin being the most extensively studied [26].
Nisin, epilancin 15X, and microbisporicin from AI-type lantibiotics exert their antibacterial effects by inhibiting cell wall synthesis of the targeted cells through the binding of the bacteriocin N-terminal domain to lipid II, a precursor of peptidoglycan. The structure shown in Figure 4 depicts Nisin. Additionally, the C-terminal domain contributes to the formation of pores, disrupting the membrane potential. Mersacidin, actagardin, and cinnamicin belong to a type-B lantibiotic group characterized by a compact globular tertiary structure [27]. Bovicin HJ50 is assumed to bind to lipid II via its N-terminal fragment, with the C-terminal domain interacting with the membrane and the disulfide-containing region, forming a hairpin-like structure that disrupts the integrity of the phospholipid bilayer. A distinct group of lantibiotics, known as two-component lantibiotics, demonstrate synergistic antibacterial activity. Lacticin 3147, the profoundly studied two-component lantibiotic, is composed of type B α-peptide (LtnA1) and type A1 β-peptide (LtnA2). Its antibacterial effect also involves the formation of pores in the target cell membrane. Lantibiotics are of significant interest due to their structural diversity and potent activity against Gram-positive pathogens. Nisin, for instance, has been utilized as a natural food preservative for the past 50 years, while several new lantibiotics are undergoing clinical trials as antimicrobials.
Lantipeptides, such as lipolantins, are newly discovered and have antibacterial properties. These compounds are identified by the avionin residue and the N-terminal guanidino fatty acid [29]. Microvionin, a bacteriocin from Microbacterium arborescens 5913, is an essential member of this group. It is effective against MRSA and Streptococcus pneumonia, but the exact mechanism of action of lipolantins is not fully understood.
2.2.2. Thiopeptides
Thiopeptides belong to class I and have diverse biological activities, including antibacterial, antiviral, and antiparasitic effects. Firmicutes and Actinomyces species are primary producers of thiopeptides. They disrupt protein synthesis by binding to the 50S ribosome subunit or elongation factors [30]. While thiopeptides are highly potent in the nanomolar range, their poor water solubility and low bioavailability limit their clinical use. GE2270A, a thiopeptide derivative, is currently under clinical trials for treating gastrointestinal infections caused by Clostridium difficile.
The Man-PTS, the main mannose permease in bacteria, serves as a receptor for class IIa bacteriocins (pediocin-like group) and class IId lactococcin A (LcnA) and lactococcin B (LcnB). Class IIa bacteriocins are effective against Clostridium difficile, while LcnA-like bacteriocins only act against Lactococcus lactis strains. Garvicin Q (GarQ), a class IId bacteriocin, shows little resemblance to LcnA-like bacteriocins and has a relatively broad antimicrobial spectrum, including activity against Clostridium difficile and Lactococcus spp, among others.
2.2.3. Modified Thiazole/Oxazole-Microcins-Boromycins
A cluster of bacteriocins with a similar structure to thiopeptides is altered thiazole/oxazole-microcins-boromycins. Their distinct characteristics encompass a macrocyclic amidine, decarboxylated C-terminal thiazole, and various uncommon β-methylated amino acid residues. The bottromycins, which have been discovered so far, are produced by bacteria of the genus Streptomyces and serve as potent agents against multidrug-resistant microorganisms such as MRSA and vancomycin-resistant enterococci (VRE) [31]. Additionally, bottromycins hinder protein synthesis by interacting with the bacterial 50S ribosomal subunit.
The altered thiazole/oxazole-microcins family also encompasses linear azole-containing peptides (LAPs). Currently, two compounds from this category—plantazolicin and goadsporin—have been structurally identified. Plantazolicin is a metabolic byproduct of Bacillus amyloliquefaciens FZB42, exhibiting selective antibacterial activity against closely related strains of the genus Bacillus. Goadsporin, derived from Streptomyces spp. TP-A0584 likewise demonstrates a limited spectrum of action, targeting only members of the genus. The mechanism of action of LAPs has not been investigated.
2.2.4. Sactibiotics
Sactibiotics, produced by the Bacillus genus, comprise unique sulfur-containing peptides. These peptides possess antibacterial properties and can act as spermicides and hemolytics. The initial compound of this kind, subtilosin A, was discovered over 30 years ago. Sactibiotics are characterized by rigid hydrocarbon skeleton structures, with their amino acid residues positioned on the surface of spirals carrying a negative charge at a neutral pH, rendering them hydrophobic [32]. The absence of a common cationic charge suggests that sactibiotics are not drawn to the membrane but instead interact with specific receptors on or within the membrane due to their narrow spectrum of action. Research has observed that subtilosin A uses Trp34 as a membrane anchor, leaving anionic amino acid residues on the surface. Moreover, the peptide can partially integrate into the membrane, disrupting the membrane’s phosphate head groups and methylene groups of lipids. The extent of this disruption depends on the concentration of the bacteriocin, which results in the release of vesicular components. Sactipeptides, also known as sactibiotics, constitute a small subfamily of peptides.
Sactipeptides, sulfur-to-alpha carbon thioether cross-linked peptides, are bacteriocins in the class of RiPPs. They exhibit diverse biological activities, including antibacterial and hemolytic properties [6]. Currently, six sactipeptides have been identified: subtilosin A, the sporulation killing factor SKF, thurincin H, thuricin CD (comprising the sactipeptides Trn-α and Trn-β), huazacin (also known as thuricin Z), and ruminococcin C. Bacillus strains are responsible for producing subtilosin A, thuricins, and SKF. Ruminococcin C is the only sactipeptide identified from a genus other than Bacillus; it was isolated from the human fecal strain Ruminococcus gnavus E1. While initially categorized as sactipeptides, thermocellin (CteA) and Tte1186a were recently reclassified as ranthipeptides, a type of ribosomally synthesized and post-translationally modified peptides distinguished by radical non-α-carbon thioether modifications [33]. Figure 5 presents the classification of bacteriocins produced by Gram-positive and Gram-negative bacteria.
3. Sources
Most bacteriocins currently used are obtained from the secondary metabolism of lactic acid bacteria (LAB). LAB are responsible for the lactic acid fermentation that turn lactose-rich milk into sour yogurt and represent valuable probiotics essential for human health due to synthesis of many bioactive substances. LAB, which are a diverse group of Gram-positive asporogenic heterotrophic bacteria belonging to the phylum Firmicutes, have a long history of safe use and have a Generally Recognized as Safe (GRAS) and Qualified Presumption of Safety (QPS) status [34]. LAB produce substances that fit into all three categories of bacteriocins of Gram-positive bacteria. Additionally, there is a strong emphasis on studying microorganisms that make up the microbiota of humans and animals. The majority of bacteria present in the gut are capable of producing the necessary bacteriocins to maintain the stability of the microbial community. Typically, they harm pathogenic enterobacteria and actinomycetes [35].
Strains within the Bacillus genus generate numerous antimicrobial peptides with diverse chemical compositions. They can produce antimicrobial compounds such as peptides, lipopeptides, and bacteriocins. Similarly, Bacillus species are responsible for the production of primary antibiotics via ribosomal (bacteriocins) or non-ribosomal (polymyxins and iturins) pathways based on their mode of action [36]. B. subtilis produced the most antibiotics, followed by B. brevis, with a few produced by other Bacillus species [37]. Different strains of B. subtilis produce a variety of bacteriocins. For example, B. subtilis, B. subtilis A1/3, B. subtilis 168, B. subtilis strain HILY-85 produces subtilin, ericin S and ericin A, sublancin 168, mersacidin, respectively. Other Bacillus species, like B. licheniformis, B. cereus, B. thuringiensis, and B. pseudomycoides, etc. also produce bacteriocins like bacillocin 490, cerein 8A, thuricin 7, and pseudomycoicidin respectively. A new bacteriocin, amylocyclicin, was recently reported, produced by B. amyloliquefaciens FZB42 [38]. Sonorensin is a new peptide belonging to heterocycloanthracin, a subfamily of bacteriocin isolated from marine bacteria B. sonorensis MT 93 [39]. This peptide showed activity against broad-spectrum bacteria, including B. subtilis, E. coli, Listeria monocytogenes, Pseudomonas aeruginosa, Staphylococcus aureus, and Vibrio vulnificus [27].
The table below (Table 1) presents some of the bacteriocins mentioned in the review with their amino acid sequences.
Figure 6A shows that whereas the amino acid sequences of the aligned peptides exhibit high variability, a general structural pattern is observed. This pattern is characterized by an enrichment of hydrophobic alpha helices at the N-terminus of the AMPs. This is associated with their ability to penetrate the microbial cell membrane and form pores. In contrast, the C-terminus is predominantly composed of beta-sheet structures, which can serve as structural elements and are also involved in membrane interactions, protein-protein interactions, and catalytic activity. More flexible loop regions were also presented and associated with the catalytic activity of the AMPs. Phylogenetically, Figure 6B illustrates the evolutionary relationships between the AMPs, with branch lengths indicating the number of mutations per site. The associated table provides additional information, including the number of amino acids, gene names, and species of origin for each AMP. The phylogenetic analysis reveals the genetic diversity and evolutionary distances among the AMPs, highlighting the mutations that have accumulated over time. Bovicin HJ50 and Thuricine precursor showed the highest divergence and formed two outgroups. Other AMPs showed relatively close relationships forming three distinctive clusters: Nisin, Subtilin, Microbisporocin; Epilancin 15X and Lacticin; and Mersacidin and Microcin.
4. Mechanisms of Action
Ribosomally produced antimicrobial peptides are a varied collection of biologically active bacterial molecules that defend against other microorganisms [40]. Despite differences in their primary structure, their positively charged and amphiphilic properties allow them to attack target bacterial cells by disrupting the cell membrane [41]. Figure 7 summarizes the five known mechanisms of action of AMPs on pathogenic cells.
Colicins are classified into three main groups based on their interaction mechanism with the target cell—pore-forming, nuclease, and peptidoglycan degrading. Receptors responsible for transporting nutrients such as vitamin B12 (cobalamin receptor BtuB), siderophore iron binding FhuA, FepA, Cir, and Fiu, and nucleosides (Tsx receptor) facilitate the uptake of colicins by the target cell. Furthermore, certain colicins utilize porin proteins that regulate the passive diffusion of sugars, phosphates, and amino acids through the outer membrane (OM) [41]. Protein bacteriocins produced by other Gram-negative bacteria are deemed colicin-like due to comparable structural and functional characteristics.
The precise method of tailocins work is not entirely understood, but it likely involves compressing the shell and infiltrating the nucleus through the cell wall. This process creates a channel or pore that impacts the membrane potential of the target cell [19]. The most researched phage tail-like bacteriocins are R-and F-pyocins from P. aeruginosa. Tailocins generally have a limited bactericidal range, affecting specific subgroups of strains within the producing species.
Microcins possess potent antibacterial properties, which rely on intricate mechanisms to penetrate the outer and inner cell membranes as well as the cell wall of Gram-negative bacteria. To bypass the OM, siderophore-microcins attach to receptors that are involved in iron transport. Cyclic microcin J25, distinguished by its N-terminal macrolactam ring, utilizes the hydroxamate receptor and the intracellular membrane protein SbmA. Meanwhile, Microcin C, produced as a heptapeptide adenylate, depends on external membrane porins and ABC membrane transporters. Once in the cytoplasm, it transforms into a non-hydrolyzable aspartyl-adenylate analog [42]. Despite employing different mechanisms to destroy target cells without any structural similarity, microcins adopt a common “Trojan horse” strategy, which could be harnessed in designing and creating new, effective antibiotics.
Nisin, a 34-amino acid lantibiotic, is produced by Gram-positive bacteria such as Lactococcus, Staphylococcus, and Streptococcus species. Various forms of nisin have been identified, including nisin A, - Z, - Q, - U, - F, - H, - O, - J, and - P. Nisin possesses antibacterial properties against a broad range of Gram-positive bacteria, including staphylococci, streptococci, enterococci, bacilli, and listeria. It binds to lipid II, an essential membrane component for peptidoglycan biosynthesis, leading to membrane permeabilization and inhibition of cell wall synthesis in targeted cells [43]. Moreover, in an MRSA model, nisin has been found to induce cell shrinkage and chromosomal DNA condensation, indicating interference with DNA replication or segregation in bacteria.
Epidermin, a 21-amino acid lantibiotic produced by S. epidermidis, demonstrates antimicrobial activity against staphylococci and streptococci. Its mechanism of action involves inhibiting cell wall synthesis through interaction with the cell wall precursor lipid II and sometimes by inducing pore formation [44]. In an in vitro catheter colonization model, epidermin significantly reduced S. epidermidis cells attached to silicone catheters. It also displayed antibacterial activity against over 85% of tested S. aureus (165 strains) responsible for bovine mastitis. In a separate study, epidermin exhibited antibacterial activity against 81.3% of tested S. aureus involved in human infections, including MRSA endemic clones in Brazil. Furthermore, epidermin demonstrated antibacterial effects against S. haemolyticus, S. capitis, S. simulans, S. saprophyticus, S. hominis, and S. epidermidis, although it showed no activity against certain tested S. aureus.
The most thoroughly researched LAB-bacteriocins can possess narrow spectra, acting solely on a limited range of target bacteria, typically within the same species, or have broad spectra targeting other species. There has been extensive research on the mode of action of LAB-bacteriocins against Gram-positive bacteria [45]. The number of LAB-bacteriocins with activity against Gram-negative bacteria is limited, unlike those with activity against Gram-positive bacteria. Although a small number of LAB-bacteriocins active against Gram-negative bacteria has been reported in the past decade, their mode of action is yet to be elucidated. LAB-bacteriocins’ effectiveness against Gram-negative target bacteria can be attributed to the structure of the cell envelope, which comprises three layers. The cytoplasmic membrane of Gram-negative bacteria is surrounded by an OM, consisting of a phospholipid bilayer and a network of lipids and polysaccharides known as lipopolysaccharides (LPSs) [46]. The OM phospholipids are linked to the inner leaflet of the membrane, and LPS is bound to the outer leaflet, known to cause endotoxic shock. It’s worth noting that LPS acts as a barrier to many antibiotics and hydrophobic compounds. However, LPS is the target of colistin, a polycationic antibiotic from the polymyxins group, which are cyclic non-ribosomal polypeptides (NRPs). Colistin is known to bind to LPS and phospholipids in the OM of Gram-negative bacteria, leading to leakage of intracellular contents and bacterial death. The rising number of Gram-negative pathogens resistant to fluoroquinolones, aminoglycosides, and β-lactams (carbapenems, monobactam, cephalosporins, and broad-spectrum penicillins) has led to the revival of colistin as a last-resort therapeutic option for treating infections caused by Gram-negative bacteria that are resistant to the drugs mentioned above [47]. The classification of LAB-bacteriocins is exposed in Figure 8.
AMPs created by Bacillus spp. have become a hopeful substitute for antibiotics because of their wide-ranging ability to fight against resistant pathogens. Despite their potential, the limited production of AMPs under standard laboratory conditions remains a challenge for large-scale production [48]. Along with medium optimization and genetic manipulation, different molecular approaches have been examined to enhance the production of recombinant AMPs. These approaches involve selecting suitable expression systems, modifying expression promoters, and metabolic engineering. AMPs derived from Bacillus show significant promise as alternative antimicrobial agents [49]. Table 2 lists some notable and detailed studies of AMPs derived from bacteria.
5. Effect on Human Health
The primary uses of bacteriocins in terms of human health are essential, with one of the critical applications being the utilization of bacteriocin-producing organisms as probiotics [50]. Probiotics refer to non-pathogenic and non-toxic strains that benefit humans and host animals. They can survive and maintain metabolic activity in the intestinal environment, remaining stable and viable for extended storage periods. Probiotics have shown potential in terms of antimicrobial production, competitive pathogen destruction, competition for nutrients, and modulation of the immune system. Currently, numerous probiotics are utilized daily, including lactic acid bacteria, non-pathogenic strains of E. coli, Bacilli, and yeast [51].
Studies have indicated that either purified bacteriocins or bacteriocin-producing probiotics can reduce pathogen numbers or alter intestinal microbiota composition in mice, chickens, and pigs. For example, a strain of Lactococcus lactis that produces nisin can promote the growth of Bifidobacterium and inhibit the growth of enterococci and streptococci in the intestines of rats across different regions such as the duodenum, ileum, caecum, and colon. Additionally, bacteriocins like Colicin Ib, E1, and microcin C7, which are derived from the E. coli H22 strain, demonstrate the ability to inhibit the growth of both pathogenic and non-pathogenic bacteria, including Enterobacter, Escherichia, Klebsiella, Morganella, Salmonella, Shigella, and Yersinia. Enterocin is a type of bacteriocin synthesized by lactic acid bacteria Enterococcus. Enterocin has excellent antimicrobial effectiveness against foodborne pathogens, such as L. monocytogenes [52].
Furthermore, research has shown that probiotic strains of human origin, such as Lactobacillus salivarius UCC118, produce bacteriocin Abp118 that can eliminate Listeria monocytogenes cells. Pediococcus spp. Lactic acid bacteria are also well-documented as probiotics, and many Pediococcus strains produce pediocin, an effective agent for eradicating Listeria spp. [53]. Ongoing research aims to comprehensively study probiotics for potential use in the pharmaceutical and food industries. MIC and inhibition diameter of some bacterial antimicrobial peptides are depicted in Table 3.
6. Limitations and Perspectives
Bacterial AMPs are vital in addressing the increasing antibiotic resistance of various pathogens, potentially serving as an alternative to antibiotics [54]. Bacteriocins have a narrow spectrum of action, making them highly specific antibacterial compounds that target particular bacterial pathogens [55]. With the wide range of natural bacteriocins available, it is relatively easy to identify effective drugs against specific human pathogens [56]. By developing and utilizing such narrow-spectrum antimicrobials, the number of available medications can be increased, and the lifespan of traditional antibiotics can also be prolonged [57].
Current research primarily focuses on the potential use of Gram-negative bacterial bacteriocins, specifically colicins and pyocins, as antibiotics. Although recent in vivo experiments have shown promising results, there are still unresolved questions about their suitability as therapeutics [58,59]. One key issue is the limited availability of data on the effects of bacteriocins on patients in terms of toxicity or immune response [60]. However, existing information does not indicate any harmful or toxic effects on the body, except for the mortality of chickens treated with pyocins. However, it is unclear whether the preparation was cleared of endotoxins. Furthermore, more research on dosage regimens and the timing of bacteriocin administration is necessary [61]. Addressing this issue requires more comprehensive pharmacokinetic studies [62].
The comparison of the efficacy of bacteriocins and conventional antibiotics has yielded promising results [63]. Piocin S5 has shown to be at least 100 times more effective than tobramycin in treating lung infections in mice, indicating its potential as a replacement for traditional antibiotics used against P. aeruginosa. However, most bacteriocins are known for their low stability and, therefore, need to be administered in higher doses with shorter intervals between doses [64]. This low stability, however, can also decrease the impact of the antibiotic on the body and the environment, ultimately minimizing the development of resistance [65]. Researchers have observed that environmental resistance to bacteriocins can arise by modifying cell surface receptors [66]. It’s important to note that, despite the potential for resistance, many bacteria still respond to specific concentrations of bacteriocins. A new class of antimicrobial agents, termed ‘structurally nanoengineered antimicrobial peptide polymers’ (SNAPPs) exhibit activity against both Gram-positive and Gram-negative bacteria like Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp (ESKAPE) and colistin-resistant and multi-drug resistant (CMDR) pathogens, while demonstrating low toxicity [67].
One disadvantage of using bacteriocins could be the innate resistance of the producing strains [68]. The presence of shared target cell destruction mechanisms and similar cytotoxic domains may lead to cross-immunity between bacterial strains. However, most bacterial strains either don’t produce immune proteins or only make a limited number [69]. As a result, bacteria can only develop resistance to a restricted range of bacteriocins. It is believed that using a combination of bacteriocins can overcome such resistance [70,71].
Author Contributions
Conceptualization, Supervision, Writing - review & editing, Data curation, Project administration, Funding Acquisition – RMG, GS; Writing - original draft, Data curation, Visualization – SD; Data curation, Visualization –YH; Supervision, Writing - review & editing, Data curation – RMG, GS. All authors have read and agreed to the published version of the manuscript.
Funding
APC was funded by Grant NBU-GFSR-27/2024.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used during the current study are available from the corresponding author upon request.
Acknowledgments
This work was conducted at UPIZ Educational and Research Laboratory of Biology-MF-NBU.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mabrouk, D. M. Antimicrobial Peptides: Features, Applications and the Potential Use against Covid-19. Molecular Biology Reports 2022. [Google Scholar] [CrossRef] [PubMed]
- Kassaa, I. A.; Rafei, R.; Moukhtar, M.; Zaylaa, M.; Gharsallaoui, A.; Asehraou, A.; Omari, K. E.; Shahin, A.; Hamze, M.; Chihib, N.-E. LABiocin Database: A New Database Designed Specifically for Lactic Acid Bacteria Bacteriocins. International Journal of Antimicrobial Agents 2019, 54, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Vanzolini, T.; Bruschi, M.; Rinaldi, A.C.; Magnani, M.; Fraternale, A. Multitalented Synthetic Antimicrobial Peptides and Their Antibacterial, Antifungal and Antiviral Mechanisms. Int. J. Mol. Sci. 2022, 23, 545. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, G.; Costantino, G. Antimicrobial Peptides (AMPs): A Patent Review (2015–2020). Expert Opinion on Therapeutic Patents 2020, 30, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M. Antimicrobial Peptides Demonstrate Activity against Resistant Bacterial Pathogens. Infect. Dis. Rep. 2023, 15, 454–469. [Google Scholar] [CrossRef] [PubMed]
- Satchanska, G.; Davidova, S.; Gergova, A. Diversity and Mechanisms of Action of Plant, Animal, and Human Antimicrobial Peptides. Antibiotics 2024, 13, 202. [Google Scholar] [CrossRef] [PubMed]
- Maria Rosa, Loffredo; Nencioni, L.; Maria Luisa, Mangoni; Casciaro, B. Antimicrobial Peptides for Novel Antiviral Strategies in the Current Post-COVID-19 Pandemic. Journal of peptide science 2023, 30. [Google Scholar] [CrossRef]
- Antimicrobial Peptide Database. Unmc.edu. https://aps.unmc.edu/. Accessed on 27.07.2024.
- Freitas, E. D.; Bataglioli, R. A.; Oshodi, J.; Beppu, M. M. Antimicrobial Peptides and Their Potential Application in Antiviral Coating Agents. Colloids and Surfaces B: Biointerfaces 2022, 217, 112693. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, M.; Karimi, A.; Fallah, F.; Akhavan, M. M. Overview of Ribosomal and Non-Ribosomal Antimicrobial Peptides Produced by Gram Positive Bacteria. Cellular and Molecular Biology 2017, 63, 20. [Google Scholar] [CrossRef] [PubMed]
- Gan, B. H.; Gaynord, J.; Rowe, S. M.; Deingruber, T.; Spring, D. R. The Multifaceted Nature of Antimicrobial Peptides: Current Synthetic Chemistry Approaches and Future Directions. Chemical Society Reviews 2021, 50, 7820–7880. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, Antimicrobial Peptides from Bacterial Origin: Overview of Their Biology and Their Impact against Multidrug-Resistant Bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Vrushali, Somase; Desai, S. A.; Patel, V. P.; Patil, V.; Kunal Bhosale. Antimicrobial Peptides: Potential Alternative to Antibiotics and Overcoming Limitations for Future Therapeutic Applications. International Journal of Peptide Research and Therapeutics 2024, 30. [Google Scholar] [CrossRef]
- Ageitos, J. M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T. G. Antimicrobial Peptides (AMPs): Ancient Compounds That Represent Novel Weapons in the Fight against Bacteria. Biochemical pharmacology 2017, 133, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Kjos, M.; Nes, I. F.; Diep, D. B.; Lotfipour, F. Natural Antimicrobial Peptides from Bacteria: Characteristics and Potential Applications to Fight against Antibiotic Resistance. Journal of Applied Microbiology 2012, 113, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Madi-Moussa, D.; Belguesmia, Y.; Charlet, A.; Drider, D.; Coucheney, F. Lacticaseicin 30 and Colistin as a Promising Antibiotic Formulation against Gram-Negative β-Lactamase-Producing Strains and Colistin-Resistant Strains. Antibiotics 2022, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics 2020, 9, 553. [Google Scholar] [CrossRef] [PubMed]
- Cascales, E.; Buchanan, S. K.; Duche, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiology and Molecular Biology Reviews 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed]
- Patz, S.; Becker, Y.; Richert-Pöggeler, K. R.; Berger, B.; Ruppel, S.; Huson, D. H.; Becker, M. Phage Tail-like Particles Are Versatile Bacterial Nanomachines – a Mini-Review. Journal of Advanced Research 2019, 19, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moosazadeh Moghaddam, M.; Mirnejad, R. Antimicrobial Peptides: Features, Action, and Their Resistance Mechanisms in Bacteria. Microbial Drug Resistance (Larchmont, N.Y.) 2018, 24, 747–767. [Google Scholar] [CrossRef]
- Luigui Gallardo-Becerra; Cervantes-Echeverría, M. ; Cornejo-Granados, F.; Vazquez-Morado, L. E.; Ochoa-Leyva, A. Perspectives in Searching Antimicrobial Peptides (AMPs) Produced by the Microbiota. Microbial Ecology 2023, 87. [Google Scholar] [CrossRef]
- Yang, F.; Yang, F.; Huang, J.; Yu, H.; Qiao, S. Microcin C7 as a Potential Antibacterial-Immunomodulatory Agent in the Postantibiotic Era: Overview of Its Bioactivity Aspects and Applications. Int. J. Mol. Sci. 2024, 25, 7213 . [Google Scholar] [CrossRef] [PubMed]
- Martínez, B.; Rodríguez, A.; Suárez, E. Antimicrobial Peptides Produced by Bacteria: The Bacteriocins. New Weapons to Control Bacterial Growth 2016, 15–38. [Google Scholar] [CrossRef]
- King, A.; Zhang, Z.; Glassey, E.; Piro Siuti; Clardy, J. ; Voigt, C. A. Systematic Mining of the Human Microbiome Identifies Antimicrobial Peptides with Diverse Activity Spectra. Nature Microbiology 2023, 8, 2420–2434. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Chaudhry, V.; Mittal, G.; Liao, L. M.; Matos, C. O.; Khatri, N.; Franco, O. L.; Patil, P. B.; Korpole, S. Characterization of the Antimicrobial Peptide Penisin, a Class Ia Novel Lantibiotic from Paenibacillus Sp. Strain A3. Antimicrobial Agents and Chemotherapy 2016, 60, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Newstead, L.L.; Varjonen, K.; Nuttall, T.; Paterson, G.K. Staphylococcal-Produced Bacteriocins and Antimicrobial Peptides: Their Potential as Alternative Treatments for Staphylococcus aureus Infections. Antibiotics 2020, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Basi-Chipalu, S.; Sthapit, P.; Dhital, S. A Review on Characterization, Applications and Structure-Activity Relationships of Bacillus Species-Produced Bacteriocins. Drug Discoveries & Therapeutics 2022, 16, 55–62. [Google Scholar] [CrossRef]
- Anumudu, C.; Hart, A.; Miri, T.; Onyeaka, H. Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods. Molecules 2021, 26, 5552. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Mukai, K.; Kaweewan, I.; Nakagawa, H.; Hosaka, T.; Kodani, S. Heterologous Production and Structure Determination of a New Lanthipeptide Sinosporapeptin Using a Cryptic Gene Cluster in an Actinobacterium Sinosporangium Siamense. Journal of Microbiology (Seoul, Korea) 2023, 61, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, A. A.; Suga, H. Introduction to Thiopeptides: Biological Activity, Biosynthesis, and Strategies for Functional Reprogramming. Cell Chemical Biology 2020, 27, 1032–1051. [Google Scholar] [CrossRef] [PubMed]
- Franz, L.; Kazmaier, U.; Truman, A. W.; Koehnke, J. Bottromycins - Biosynthesis, Synthesis and Activity. Natural Product Reports 2021, 38, 1659–1683. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Rea, M.; Cotter, P.; Hill, C.; Ross, R. The Sactibiotic Subclass of Bacteriocins: An Update. Current Protein & Peptide Science 2015, 16, 549–558. [Google Scholar] [CrossRef]
- Eastman, Karsten A. S.; Kincannon, W. M.; Bandarian, V. Leveraging Substrate Promiscuity of a Radical S-Adenosyl-L-Methionine RiPP Maturase toward Intramolecular Peptide Cross-Linking Applications. ACS Central Science 2022, 8, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Balandin, S. V.; Sheremeteva, E. V.; Ovchinnikova, T. V. Pediocin-like Antimicrobial Peptides of Bacteria. Biochemistry (Moscow) 2019, 84, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Divyashree, M.; Mani, M. K.; Reddy, D.; Kumavath, R.; Ghosh, P.; Azevedo, V.; Barh, D. Clinical Applications of Antimicrobial Peptides (AMPs): Where Do We Stand Now? Protein & Peptide Letters 2020, 27, 120–134. [Google Scholar] [CrossRef]
- Yang, X.; Yousef, A. E. Antimicrobial Peptides Produced by Brevibacillus Spp.: Structure, Classification and Bioactivity: A Mini Review. World Journal of Microbiology and Biotechnology 2018, 34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Military Medical Research 2021, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Lafuente, I.; Sevillano, E.; Nuria Peña; Cuartero, A. ; Hernández, P. E.; Cintas, L. M.; Estefanía Muñoz-Atienza; Borrero, J. Production of Pumilarin and a Novel Circular Bacteriocin, Altitudin A, by Bacillus Altitudinis ECC22, a Soil-Derived Bacteriocin Producer. International Journal of Molecular Sciences 2024, 25, 2020–2020. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V. Structure, Function, and Therapeutic Potential of Marine Bioactive Peptides. Mar. Drugs 2019, 17, 505. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Deber, C. M. Interaction of Designed Cationic Antimicrobial Peptides with the Outer Membrane of Gram-Negative Bacteria. Scientific Reports 2024, 14. [Google Scholar] [CrossRef]
- Cramer, W. A.; Sharma, O.; Zakharov, S. D. On Mechanisms of Colicin Import: The Outer Membrane Quandary. Biochemical Journal 2018, 475, 3903–3915. [Google Scholar] [CrossRef] [PubMed]
- Beis, K.; Rebuffat, S. Multifaceted ABC Transporters Associated to Microcin and Bacteriocin Export. Research in Microbiology 2019, 170, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Götz, F.; Perconti, S.; Popella, P.; Werner, R.; Schlag, M. Epidermin and Gallidermin: Staphylococcal Lantibiotics. International Journal of Medical Microbiology 2014, 304, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, A.; Asadi, A.; Mahdizade Ari, M.; Ohadi, E.; Talebi, M.; Halaj Zadeh, M.; Darb Emamie, A.; Ghanavati, R.; Kakanj, M. Bacteriocins: Properties and Potential Use as Antimicrobials. Journal of Clinical Laboratory Analysis 2021, 36. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R. The Century-Long Journey of Peptide-Based Drugs. Antibiotics 2024, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current Knowledge of the Mode of Action and Immunity Mechanisms of LAB-Bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Meštrović, T.; Juzbašić, M.; Tomas, M.; Erić, S.; Horvat Aleksijević, L.; Bekić, S.; Schwarz, D.; Matić, S.; Neuberg, M.; Škrlec, I. Antimicrobial Peptides—Mechanisms of Action, Antimicrobial Effects and Clinical Applications. Antibiotics 2022, 11, 1417. [Google Scholar] [CrossRef] [PubMed]
- Lewies, A.; Du Plessis, L. H.; Wentzel, J. F. Antimicrobial Peptides: The Achilles’ Heel of Antibiotic Resistance? Probiotics and Antimicrobial Proteins 2018, 11, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Sheau Ling Puan; Pirasannah Erriah; Al-adil, M. ; Normi, Y. M.; Nur, W.; Ali; Siti Aqlima Ahmad; Siti Nurbaya Oslan; Lim, S.; Sabri, S. Antimicrobial Peptides from Bacillus Spp. And Strategies to Enhance Their Yield. Applied Microbiology and Biotechnology 2023, 107, 5569–5593. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Guo, Z.; Xia, B.; Zhang, Y.; Liu, X.; Yu, Y.; Tang, N.; Tong, X.; Wang, M.; Ye, X.; Feng, J.; Chen, Y.; Wang, J. Identification of Antimicrobial Peptides from the Human Gut Microbiome Using Deep Learning. Nature Biotechnology 2022, 40, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Shao, C.; Li, G.; Shan, A.; Chou, S.; Wang, J.; Ma, Q.; Dong, N. Conversion of Broad-Spectrum Antimicrobial Peptides into Species-Specific Antimicrobials Capable of Precisely Targeting Pathogenic Bacteria. Scientific Reports 2020, 10, 944. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Walia, D.; Batra, N. Chapter 6 - Fresh-Cut Fruits: Microbial Degradation and Preservation. ScienceDirect. https://www.sciencedirect.com/science/article/abs/pii/B9780128115152000068.
- Kravchenko, S. V.; Domnin, P. A.; Grishin, S. Y.; Panfilov, A. V.; Azev, V. N.; Mustaeva, L. G.; Gorbunova, E. Y.; Kobyakova, M. I.; Surin, A. K.; Glyakina, A. V.; Fadeev, R. S.; Ermolaeva, S. A.; Galzitskaya, O. V. Multiple Antimicrobial Effects of Hybrid Peptides Synthesized Based on the Sequence of Ribosomal S1 Protein from Staphylococcus Aureus. International Journal of Molecular Sciences 2022, 23, 524–524. [Google Scholar] [CrossRef]
- Ryu, M.; Park, J.; Yeom, J.-H.; Joo, M.; Lee, K. Rediscovery of Antimicrobial Peptides as Therapeutic Agents. Journal of Microbiology 2021, 59, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. H.; Lu, T. K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M. Antimicrobial Peptides Demonstrate Activity against Resistant Bacterial Pathogens. Infect. Dis. Rep. 2023, 15, 454–469 . [Google Scholar] [CrossRef] [PubMed]
- Tonk, M.; Růžek, D.; Vilcinskas, A. Compelling Evidence for the Activity of Antiviral Peptides against SARS-CoV-2. Viruses 2021, 13, 912. [Google Scholar] [CrossRef] [PubMed]
- Mourtada, R.; Herce, H. D.; Yin, D. J.; Moroco, J. A.; Wales, T. E.; Engen, J. R.; Walensky, L. D. Design of Stapled Antimicrobial Peptides That Are Stable, Nontoxic and Kill Antibiotic-Resistant Bacteria in Mice. Nature Biotechnology 2019, 37, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. International Journal of Molecular Sciences 2021, 22, 11401. [Google Scholar] [CrossRef] [PubMed]
- Cresti, L.; Cappello, G.; Pini, A. Antimicrobial Peptides towards Clinical Application—A Long History to Be Concluded. Int. J. Mol. Sci. 2024, 25, 4870 . [Google Scholar] [CrossRef] [PubMed]
- Lourenço, A.L.P.; Rios, T.B.; da Silva, Á.P.; Franco, O.L.; Ramada, M.H.S. Peptide Stapling Applied to Antimicrobial Peptides. Antibiotics 2023, 12, 1400. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-T.; Yang, L.-Y.; Lin, C.-Y.; Wang, C.-T.; Lai, C.-W.; Ko, C.-F.; Shih, Y.-H.; Chen, S.-H. Intelligent De Novo Design of Novel Antimicrobial Peptides against Antibiotic-Resistant Bacteria Strains. Int. J. Mol. Sci. 2023, 24, 6788. [Google Scholar] [CrossRef] [PubMed]
- Almaaytah, A.; Mohammed, G.; Abualhaijaa, A.; Al-Balas, Q. Development of Novel Ultrashort Antimicrobial Peptide Nanoparticles with Potent Antimicrobial and Antibiofilm Activities against Multidrug-Resistant Bacteria. Drug Design, Development and Therapy 2017, 11, 3159–3170. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic Peptides: Current Applications and Future Directions. Signal Transduction and Targeted Therapy 2022, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Lam, S. J.; O’Brien-Simpson, N. M.; Pantarat, N.; Sulistio, A.; Wong, E. H. H.; Chen, Y.-Y.; Lenzo, J. C.; Holden, J. A.; Blencowe, A.; Reynolds, E. C.; Qiao, G. G. Combating Multidrug-Resistant Gram-Negative Bacteria with Structurally Nanoengineered Antimicrobial Peptide Polymers. Nature Microbiology 2016, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, T.; Chetia, M.; Chatterjee, S. Antimicrobial Peptides and Proteins: From Nature’s Reservoir to the Laboratory and Beyond. Frontiers in Chemistry 2021, 9, 691532. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Yang, P.; Lei, J.; Zhao, J. Biological Function of Antimicrobial Peptides on Suppressing Pathogens and Improving Host Immunity. Antibiotics 2023, 12, 1037. [Google Scholar] [CrossRef] [PubMed]
- Bull, T.J.; Munshi, T.; Lopez-Perez, P.M.; Tran, A.C.; Cosgrove, C.; Bartolf, A.; Menichini, M.; Rindi, L.; Parigger, L.; Malanovic, N.; et al. Specific Cationic Antimicrobial Peptides Enhance the Recovery of Low-Load Quiescent Mycobacterium tuberculosis in Routine Diagnostics. Int. J. Mol. Sci. 2023, 24, 17555. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C. D.; Hiss, J. A.; Hancock, R. E. W.; Schneider, G. Designing Antimicrobial Peptides: Form Follows Function. Nature Reviews Drug Discovery 2011, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graph showing the number of publications over ten years (2014-2024) using the keywords “bacterial antimicrobial peptides.” The search was done in July 2024, reaching 800 papers. They are expected to reach close to 1,500 by the end of the year.
Figure 1.
Graph showing the number of publications over ten years (2014-2024) using the keywords “bacterial antimicrobial peptides.” The search was done in July 2024, reaching 800 papers. They are expected to reach close to 1,500 by the end of the year.
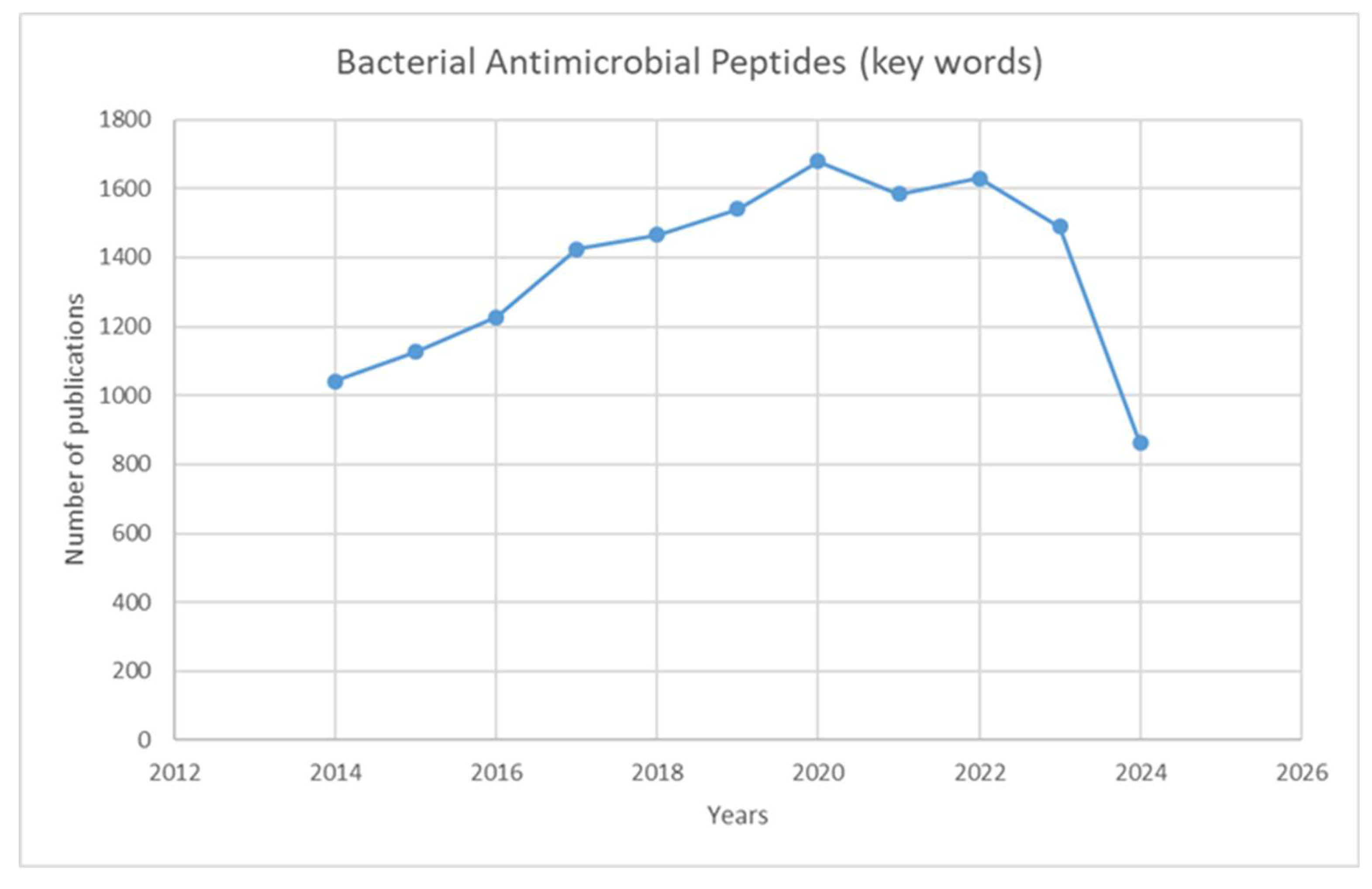
Figure 2.
Simplified models of alpha helix, β-sheet, and extended coil structures [6].
Figure 2.
Simplified models of alpha helix, β-sheet, and extended coil structures [6].
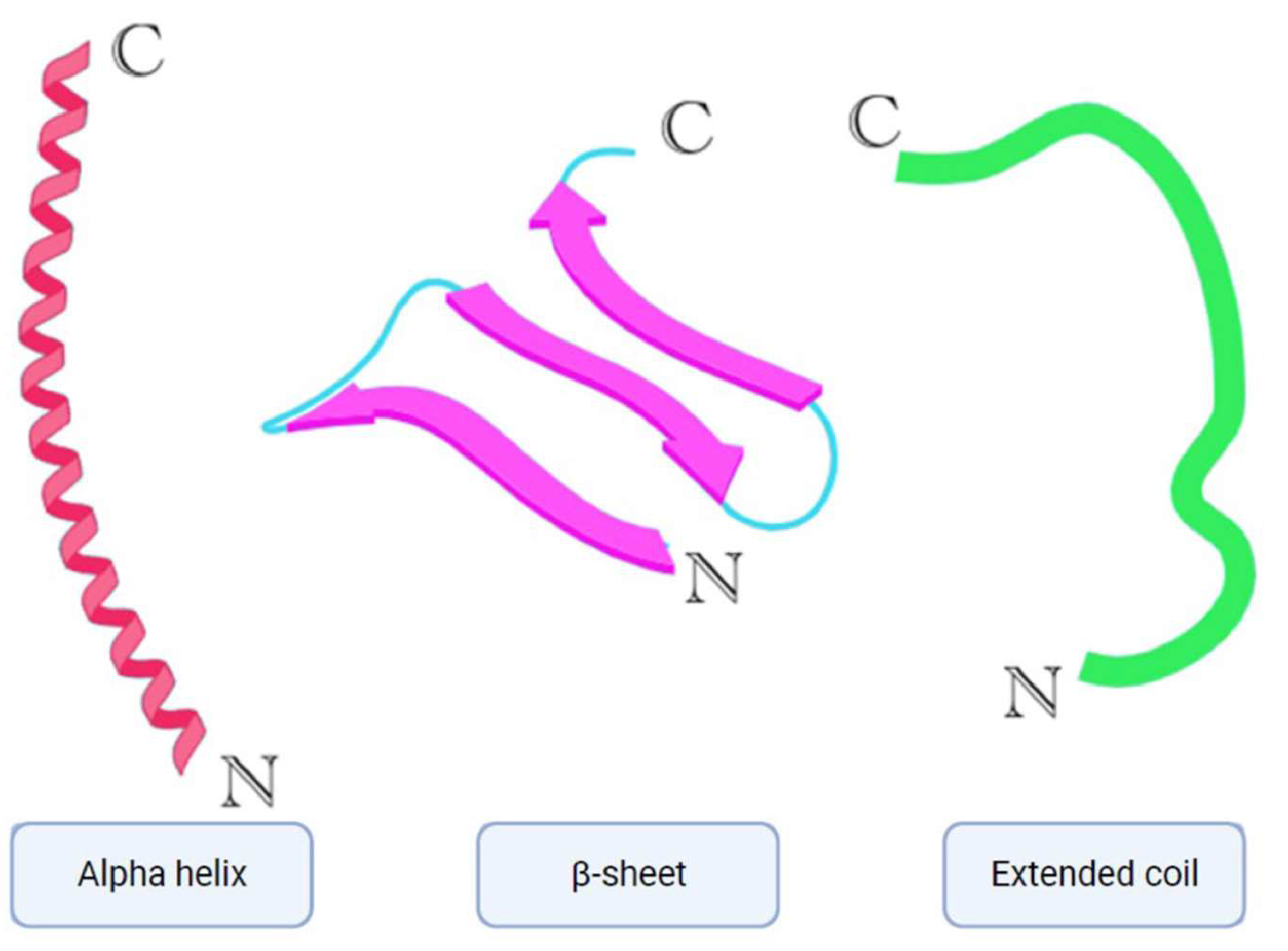
Figure 3.
The structure of Microcin 7 (McC7) [22].
Figure 3.
The structure of Microcin 7 (McC7) [22].
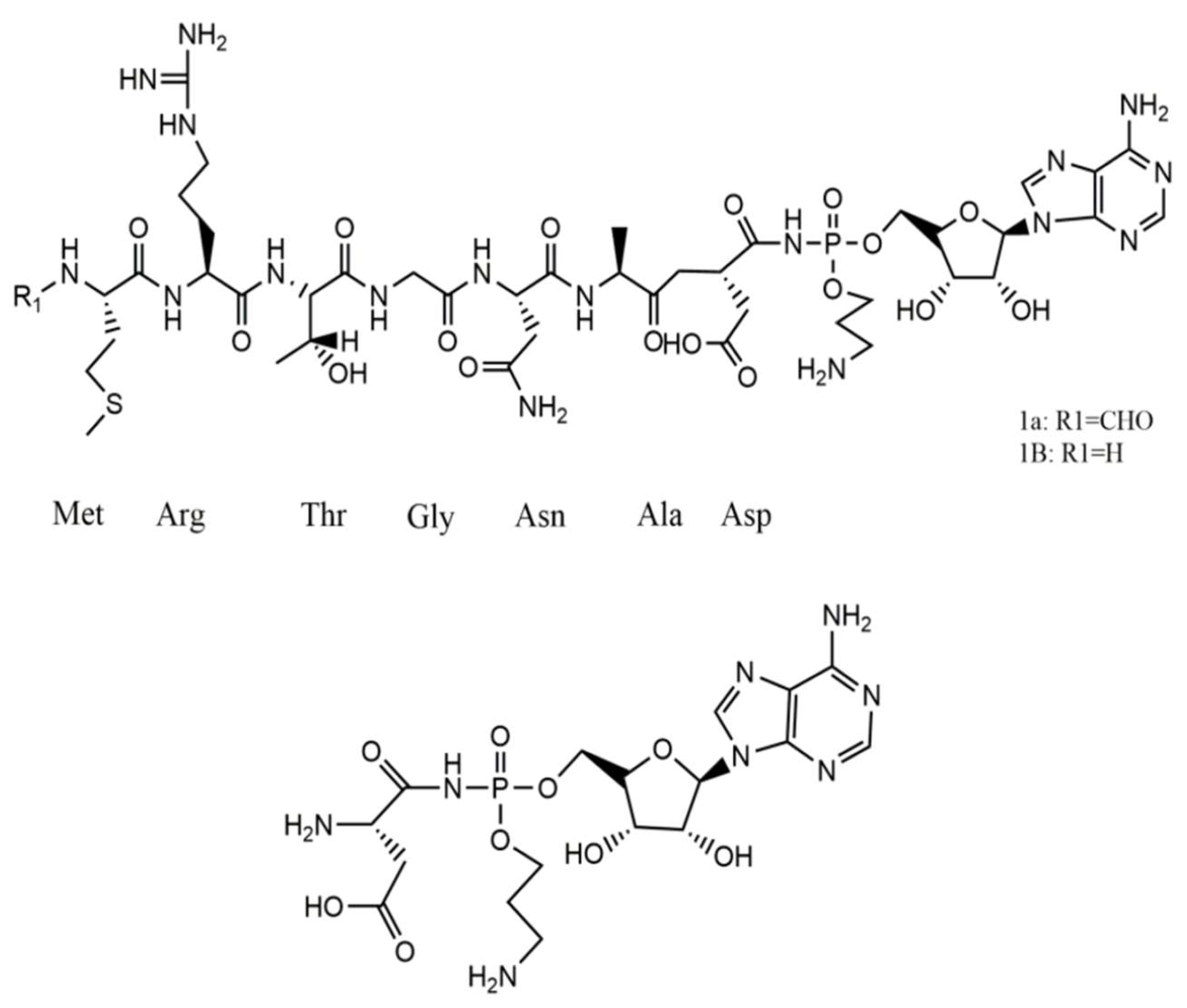
Figure 4.
Structure of Nisin [28].
Figure 4.
Structure of Nisin [28].
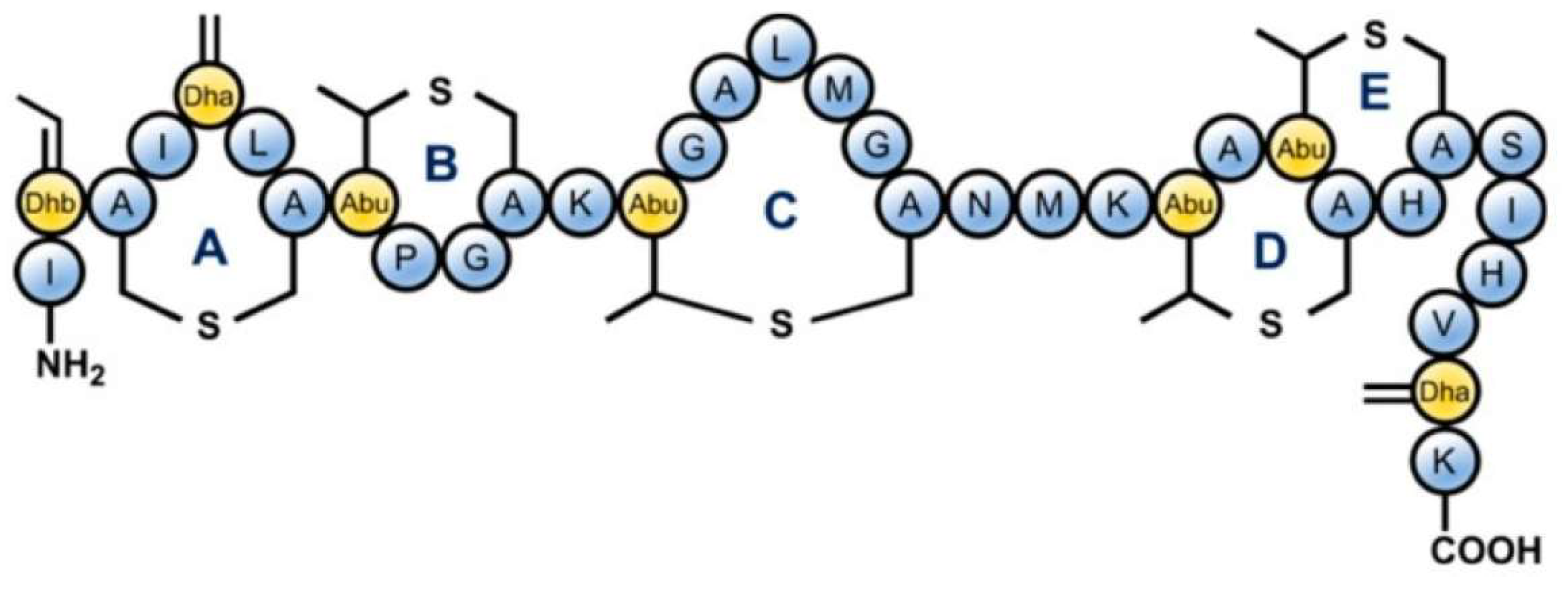
Figure 5.
Classification of bacteriocins produced by Gram-positive (A) and Gram-negative (B) bacteria [12].
Figure 5.
Classification of bacteriocins produced by Gram-positive (A) and Gram-negative (B) bacteria [12].
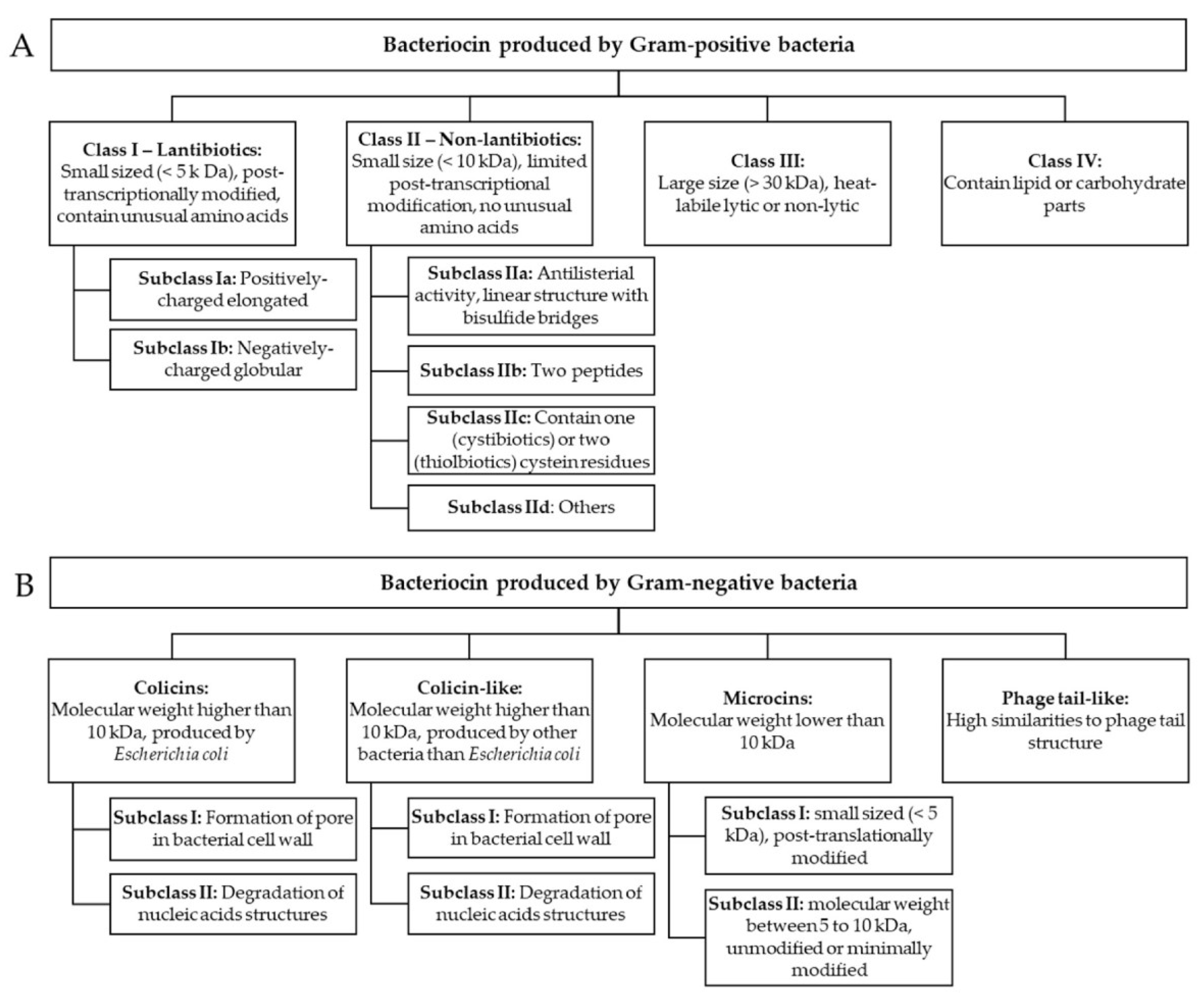
Figure 6.
[A] Alignment of nine bacterial antimicrobial peptides (AMPs). The alignment was performed using Geneious Prime 2023.1, a Geneious algorithm. Annotation revealed the distribution of alpha-helical domains, represented by grey cylinders above each peptide sequence and beta-sheets depicted by black arrows. [B] Phylogenetic tree of the AMPs (Geneious Prime 2023.1). The tree includes the number of mutations per site, the length of the sequences, the corresponding genes, and their species of origin; the number of residue changes per site represents distance.
Figure 6.
[A] Alignment of nine bacterial antimicrobial peptides (AMPs). The alignment was performed using Geneious Prime 2023.1, a Geneious algorithm. Annotation revealed the distribution of alpha-helical domains, represented by grey cylinders above each peptide sequence and beta-sheets depicted by black arrows. [B] Phylogenetic tree of the AMPs (Geneious Prime 2023.1). The tree includes the number of mutations per site, the length of the sequences, the corresponding genes, and their species of origin; the number of residue changes per site represents distance.
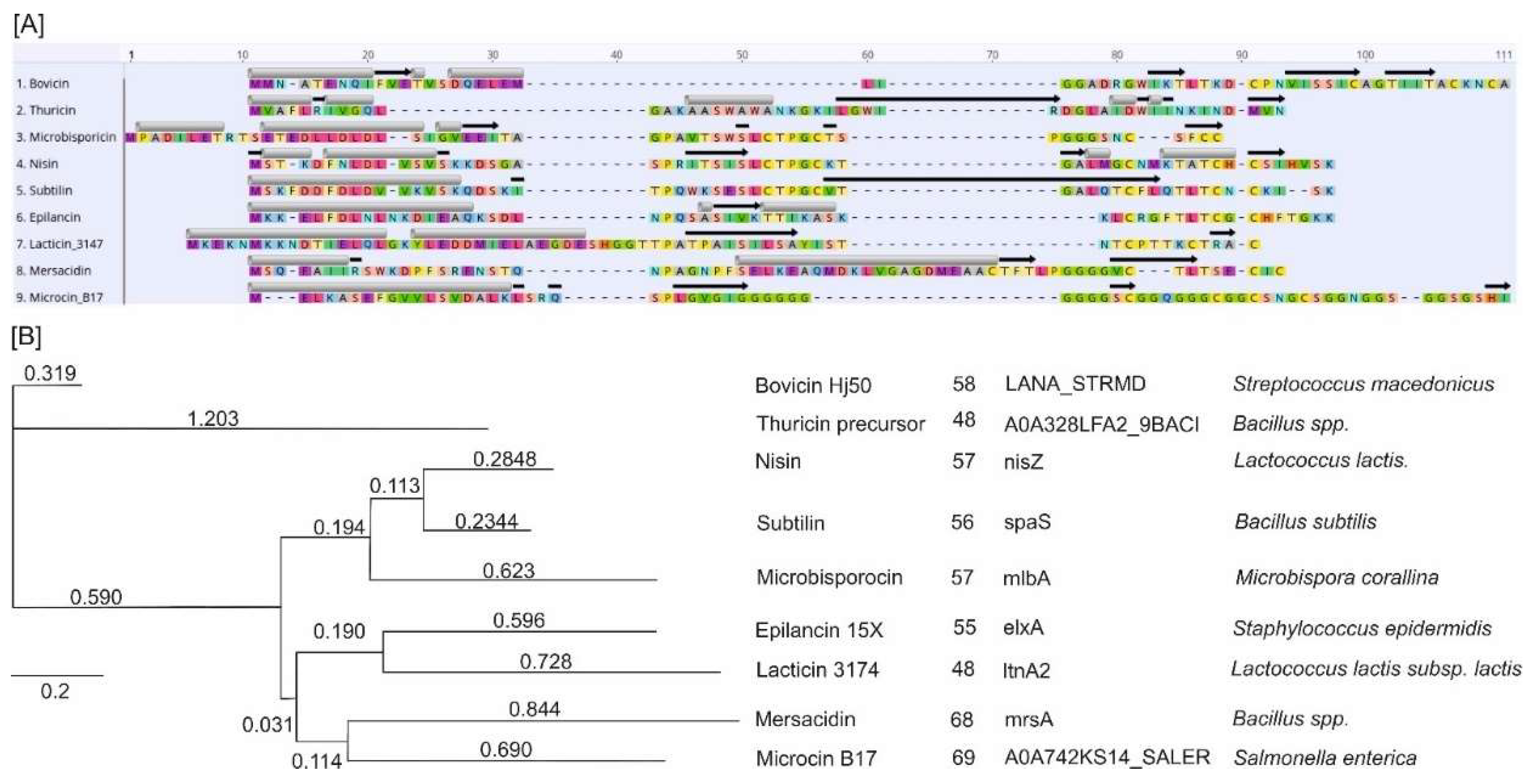
Figure 7.
The mechanisms of action of AMPs on a pathogenic cell. The figure shows two significant types of peptides—membrane-bound and intracellular active peptides. The outer layer, “1”, is the cytoplasmic membrane of a pathogen cell, and the AMPs are marked as “2”. Membrane-bound peptides can act by five mechanisms: the barrel-stave, toroidal, carpet, floodgate, and aggregate channel models. The intracellular AMPs are represented by numbers “3” to “7” and show the inhibition of enzymes required for the binding of cell wall structural proteins, DNA and RNA synthesis, ribosomal functions and chaperone protein synthesis, and cellular respiration via ROS (Reactive Oxygen Species) formation, and ATP (Adenosine Triphosphate) and NADH (Nicotinamide Adenine Dinucleotide) [6].
Figure 7.
The mechanisms of action of AMPs on a pathogenic cell. The figure shows two significant types of peptides—membrane-bound and intracellular active peptides. The outer layer, “1”, is the cytoplasmic membrane of a pathogen cell, and the AMPs are marked as “2”. Membrane-bound peptides can act by five mechanisms: the barrel-stave, toroidal, carpet, floodgate, and aggregate channel models. The intracellular AMPs are represented by numbers “3” to “7” and show the inhibition of enzymes required for the binding of cell wall structural proteins, DNA and RNA synthesis, ribosomal functions and chaperone protein synthesis, and cellular respiration via ROS (Reactive Oxygen Species) formation, and ATP (Adenosine Triphosphate) and NADH (Nicotinamide Adenine Dinucleotide) [6].
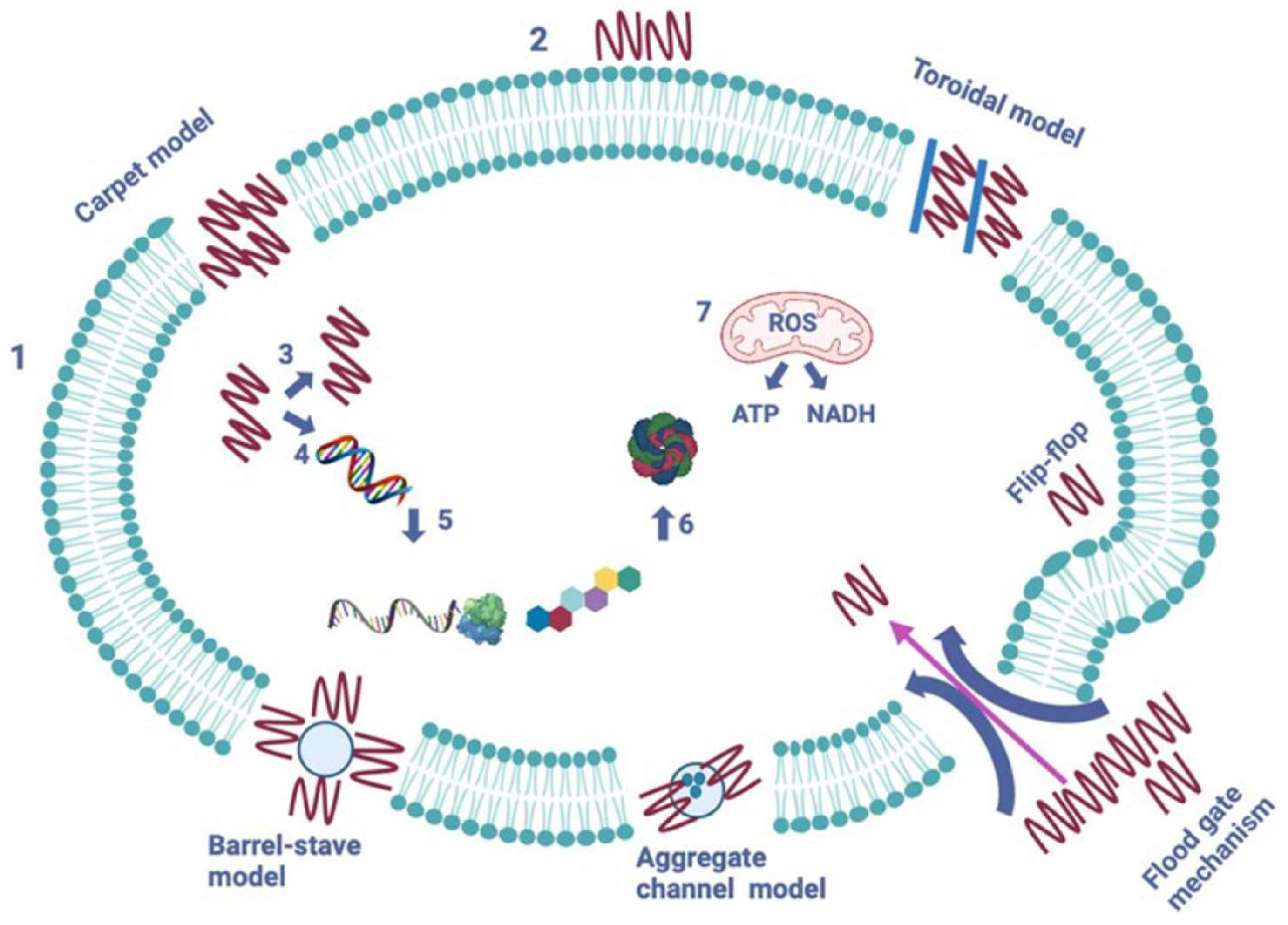
Figure 8.
Suggested classification scheme for LAB-bacteriocins [47].
Figure 8.
Suggested classification scheme for LAB-bacteriocins [47].
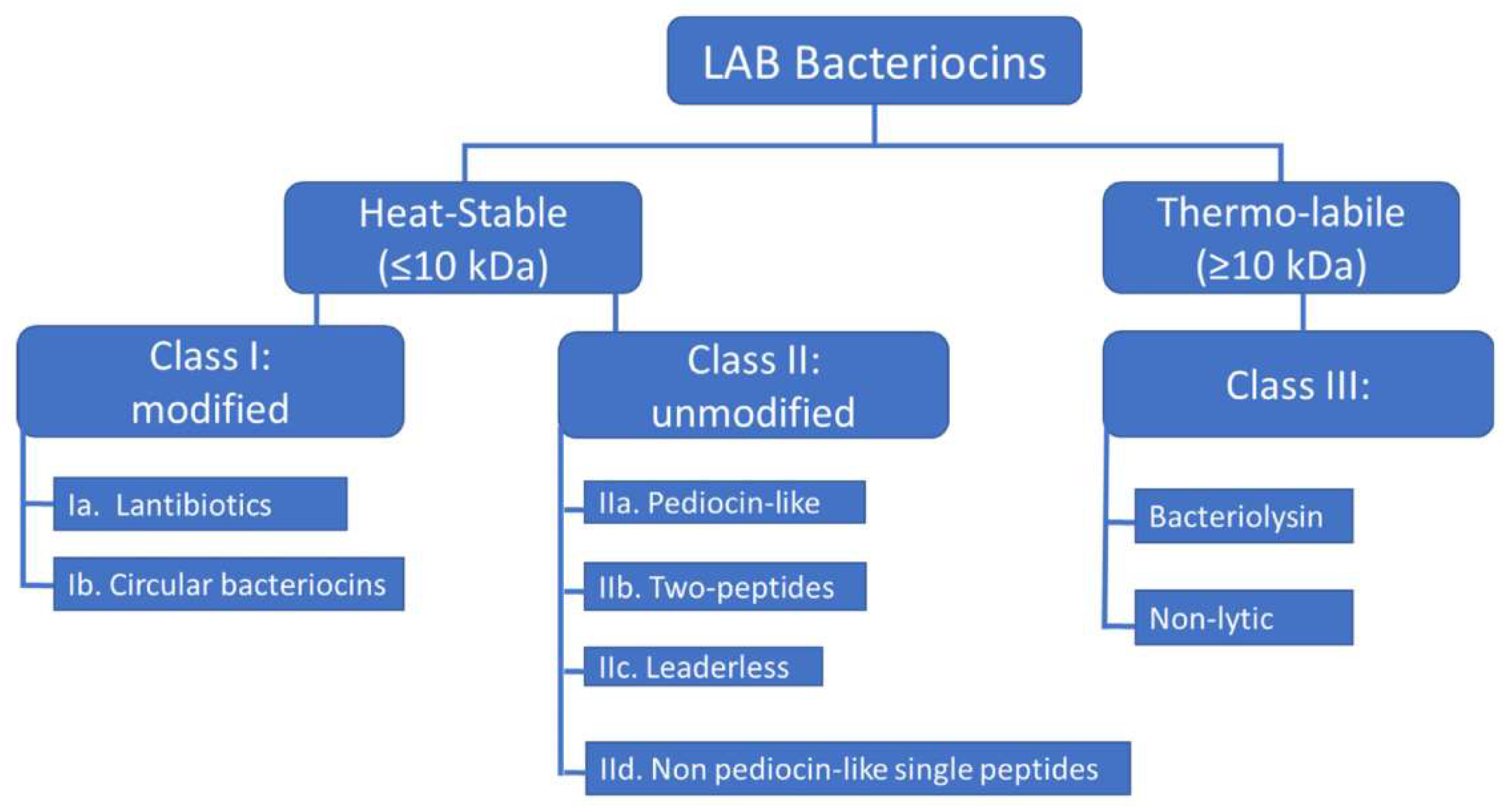

Table 1.
Amino acid sequences of some bacteriocins and their respective UniProt identification numbers.
Table 1.
Amino acid sequences of some bacteriocins and their respective UniProt identification numbers.
| Bacteriocin | UniProt ID | Amino acid sequence |
|---|---|---|
| Nisin | P29559 | MSTKDFNLDLVSVSKKDSGASPRITSISLCTPGCKTGALMGCNMKTATCHCSIHVSK |
| Colicin M | A0A761KWA3 | METLTVHAPSPSTNLPSYGNGAFSLSAPHVPGAGPLLVQVVYSFFQSPNMCLQALTQLEDYIKKHGASNPLTLQIISTNIGYFCNAERNLVLHPGISVYDAYHFAKPAPSQYDYRSMNMKQMSGNVTTPIVALAHYLWGNGAERSVNIANIGLKISPMKINQIKDIIKSGVVGTFPVSTKFTHATGDYNVITGAYLGNITLKTEGTLTISANGSWTYNGVVRSYDDKYDFNASTHRGVIGESLTRLGAMFSGKEYQILLPGEIHIKESGKR |
| Microcin B17 | A0A742KS14 | MELKASEFGVVLSVDALKLSRQSPLGVGIGGGGGGGGGGSCGGQGGGCGGCSNGCSGGNGGSGGSGSHI |
| Subtilin | P10946 | MSKFDDFDLDVVKVSKQDSKITPQWKSESLCTPGCVTGALQTCFLQTLTCNCKISK |
| Lacticin 3147 | O87237 | MKEKNMKKNDTIELQLGKYLEDDMIELAEGDESHGGTTPATPAISILSAYISTNTCPTTKCTRAC |
| Thuricin precursor | A0A328LFA2 | MVAFLRIVGQLGAKAASWAWANKGKILGWIRDGLAIDWIINKINDMVN |
| Epilancin 15X | P86047 | MKKELFDLNLNKDIEAQKSDLNPQSASIVKTTIKASKKLCRGFTLTCGCHFTGKK |
| Microbisporicin | W2EQT3 | MPADILETRTSETEDLLDLDLSIGVEEITAGPAVTSWSLCTPGCTSPGGGSNCSFCC |
| Mersacidin | A0A2H4RAU1 | MSQEAIIRSWKDPFSRENSTQNPAGNPFSELKEAQMDKLVGAGDMEAACTFTLPGGGGVCTLTSECIC |
| Bovicin HJ50 | H2A7G5 | MMNATENQIFVETVSDQELEMLIGGADRGWIKTLTKDCPNVISSICAGTIITACKNCA |
Table 2.
Table summarizing some of the bacterial AMPs with their source and activity.
| Bacterial AMP | Source | Active against |
|---|---|---|
| Colicin | E. coli | Enterobacter, Escherichia, Klebsiella, Morganella, Salmonella, Shigella, and Yersinia |
| Gramicidin | B. brevis | Gram-positive; Gram-negative |
| Microvionin | Microbacterium arborescens | MRSA and Streptococcus pneumonia |
| Plantazolicin | Bacillus amyloliquefaciens | Closely related strains of the genus Bacillus |
| Goadsporin | Streptomyces spp | Closely related strains of the genus Streptomyces |
| Sonorensin | B. sonorensis | B. subtilis, E. coli, Listeria monocytogenes, Pseudomonas aeruginosa, Staphylococcus aureus, and Vibrio vulnificus |
| Nisin | Lactococcus, Staphylococcus, and Streptococcus spp. | Staphylococci, streptococci, enterococci, bacilli, and listeria |
| Epidermin | S. epidermidis | S. haemolyticus, S. capitis, S. simulans, S. saprophyticus, S. hominis, S. epidermidis, S. aureus |
| Microcin C7 | E. coli | Enterobacter, Escherichia, Klebsiella, Morganella, Salmonella, Shigella, and Yersinia |
| Microcin L | E. coli | Escherichia coli, Salmonella enterica, Shigella spp., Pseudomonas aeruginosa |
| Abp118 | Lactobacillus salivarius | Listeria monocytogenes |
| Pediocin | Pediococcus spp | Listeria spp. |
| Bovicin HC5 | Streptococcus bovis | Listeria monocytogenes |
| Bottromycin A2 | Streptomyces bottropensis | MRSA, VRE |
| Enterocin A | Enterococcus faecium | Listeria monocytogenes |
Table 3.
Antibacterial activities of various bacteriocins produced by Gram-positive and Gram-negative bacteria [12].
Table 3.
Antibacterial activities of various bacteriocins produced by Gram-positive and Gram-negative bacteria [12].
| Bacteriocin | Producer strain | Active against | MIC (mg/L) |
Inhibition Diameter (mm) |
|---|---|---|---|---|
| Nisin A | Lactococcus lactis | MRSA Vancomycin-intermediate S. aureus (VISA) VRE |
0.5–4.1 2–>8.3 2–>8.3 |
|
| Epidermin | Staphylococcus epidermidis |
Staphylococcus aureus Streptococcus agalactiae |
>14 >14 |
|
| Bovicin HC5 | Streptococcus bovis | Listeria monocytogenes | >16 | |
| Bottromycin A2 | Streptomyces bottropensis | MRSA VRE |
1 0.5 |
|
| Pediocin PA-1 | Pediococcus acidilactici | Listeria monocytogenes | 0.0013–0.0062 | |
| Enterocin A | Enterococcus faecium | Listeria monocytogenes | 0.0002–0.0011 | |
| Microcin L | Escherichia coli |
Escherichia coli Salmonella enterica Shigella spp. Pseudomonas aeruginosa |
12–18 12–18 12–18 8–12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



