
Submitted:
09 August 2024
Posted:
12 August 2024
You are already at the latest version
Abstract
The ovary is the major determinant of female reproductive health. Ovarian functions are mainly related to the primordial follicle pool, which is gradually loss with aging. Ovarian aging and reproductive dysfunctions share oxidative stress as common underlying mechanism. ROS signaling is essential for normal ovarian processes yet can contribute to various ovarian disorders when disrupted. Therefore, balance in the redox system is crucial for proper ovarian functions. In the present study, by focusing on mRNAs and ncRNAs described in the ovary and taking into account only validated ncRNA interactions, we built up an ovarian antioxidant ceRNA network, namely OvAnOx ceRNA, composed of 5 mRNAs (SOD1, SOD2, CAT, PRDX3, GR), 10 miRNAs and 5 lncRNAs (XIST, FGD5-AS1, MALAT1, NEAT1, SNHG1). Our bioinformatic analysis indicated that the components of the OvAnOx ceRNA not only contribute to antioxidant defence, but are also involved in other ovarian functions. Indeed, antioxidant enzymes encoded by mRNA of OvAnOx ceRNA operate within a regulatory network that impacts ovarian reserve, follicle dynamics, and oocyte maturation in normal and pathological conditions. The OvAnOx ceRNA represents a promising tool to unravel the complex dialogue between redox potential and ovarian signalling pathways involved in reproductive health, aging and diseases.
Keywords:
ceRNA network
; oxidative stress
; female fertility
; antioxidant cell defence
; non-coding RNAs
1. Introduction
The ovary is the main regulator of female reproductive function. It provides oocytes for fertilization and synthesizes reproductive hormones needed for proper fertility. During fetal development, oocytes establish associations with somatic cells to form primordial follicles that create a finite reserve of primary oocytes (the primordial follicle pool). Regulatory pathways that govern the subsequent activation of follicle growth must ensure that the reserve of oocytes in humans lasts for up to 50 years. During the fourth decade of life, the ovarian follicle pool undergoes a sustained decline and provides mature oocytes with reduced competence [1]. This process is known as ovarian aging and is responsible for the early decline of the reproductive function in women [2]. Apart from this physiological process, ovarian functions can be hampered by pathological conditions such as premature-ovarian insufficiency (POI) and polycystic ovarian syndrome (PCOS) [3]. Ovarian dysfunctions related to aging and pathological reduced functionality share the oxidative stress as one of the main causative mechanisms [2,4,5,6]. This emphasizes the need for investigation to create effective strategies based on selective targeting of specific redox-modulating mechanisms, also considering that there is limited evidence in support of supplemental oral antioxidants for sub-fertile women [6,7].
Recent evidence has shown the involvement of non-coding RNAs (ncRNA) in the antioxidant systems that scavenge free radicals for maintaining a healthy level of reactive oxygen species (ROS). ROS are by-products of cellular oxidative metabolism and play a pivotal role in many cellular functions. Gene expression, cell signaling, and redox homeostasis all depend on the equilibrium between the creation and removal of ROS known as “redox homeostasis” [8,9]. This is maintained by a highly responsive dynamic system that detects changes in redox status and realigns metabolic activities to restore stability [10]. Either an increase in ROS concentration or a decrease in scavenging capacity causes an imbalance in the redox environment leading to ROS accumulation and oxidative damage to lipids, proteins, and DNA [8,9].
Non-coding RNAs constitute most of the human transcriptome and perform essential regulatory functions in every step of gene expression [11]. They are classified into two major classes, based on their length: small non-coding RNAs (e.g. microRNAs), smaller than 200 nucleotides, and long non-coding RNAs (lncRNAs) ranging from 200 nucleotides to 100 kilobases or more [11]. LncRNAs,the most heterogeneous class of ncRNAs, are involved in a wide spectrum of molecular mechanisms regulating genome functions generating complex networks of RNA-RNA competitive interactions [12]. Different studies have demonstrated interactions among lncRNAs and miRNAs, miRNAs and mRNAs, and lncRNAs and mRNAs [12,13]. These RNA molecules collaborate to create dynamic regulatory networks, with lncRNAs acting as competing endogenous RNAs (ceRNAs) [14];[15]. ceRNA-networks are complex and redundant, as a single mRNA can be targeted by different miRNAs, which in turn can interact with various lncRNAs, and a single lncRNA can sponge several miRNAs, regulating different mRNAs. ceRNAs that regulate miRNAs- mRNAs interactions are also strong proponents of many diseased conditions [16,17,18]. Some miRNAs/lncRNAs can worsen disease progression by impacting ROS-related processes, while others can effectively protect cells from ROS-induced damage [19]. These RNAs have tissue-specific influences, and can interact within ceRNA-networks to modulate gene expression [12,13]. They play a crucial role in maintaining redox balance by affecting key antioxidant enzymes [20,21].
In this challenging new context, we investigated the potential regulation of antioxidant enzymes by ceRNA networks in the ovary. Using a bioinformatics approach, we established a prediction model for interactions among mRNAs encoding antioxidant enzymes, miRNAs, and lncRNAs considering RNAs known to be expressed in the human ovary. Based on these data, we built up a potential ovarian antioxidant ceRNA-network, here referred to as OvAnOx ceRNA, comprising miRNAs targeting antioxidant enzyme mRNAs and the lncRNAs targeting these miRNAs.
2. Materials and Methods
Gene Ontology and Pathways Analysis, Localization and Expression of the 21 Antioxidant Enzymes
This study focused on 21 antioxidant genes [22]. The Gene Ontology for molecular functions and pathway analysis of the 21 antioxidant enzymes were investigated by Panther 19.0 (http://www.pantherdb.org). To verify their expression in the human ovary, we queried the Ovarian Kaleidoscope Database (https://appliedbioinfo.com/). For cellular localization, we consulted scientific papers annotated in common databases. Mitochondrial localization was investigated using Human Mitocarta3.0 (https://www.broadinstitute.org/mitocarta/mitocarta30-inventory-mammalian-mitochondrial-proteins-and-pathways) and MitoProteome (http://www.mitoproteome.org). Exosome localization was assessed using Exocarta (http://www.exocarta.org) and exoRBase 2.0 (http://www.exorbase.org) databases. Transcript expression in the human ovary was explored by querying the human protein atlas (HPA) – tissue dataset section (https://www.proteinatlas.org)
LncRNA-miRNA-mRNA Competing Endogenous RNA Networks
The miRNA-mRNA interaction analyses were performed by using miRTarbase (https://mirtarbase.cuhk.edu.cn) database, selecting only interactions validated by functional experimental evidence. To investigate the interactions between lncRNAs and the miRNAs targeting antioxidant enzymes mRNAs, we consulted Starbase (https://starbase.sysu.edu.cn/) selecting lncRNAs with at least two target binding sites for miRNAs.
We designed the competing endogenous RNA networks (ceRNA-network) considering the miRNAs targeting antioxidant enzymes mRNAs and the lncRNAs targeting these miRNAs. Interaction network miRNA-mRNA, lncRNA-miRNA and lncRNA-miRNA-mRNA were designed by Cytoscape 3.8.2.
Cellular Localization of miRNAs and lncRNAs Regulating the Antioxidant Genes
In order to investigate if the previously identified miRNAs and the lncRNAs are expressed in the ovary, we used miRDB (http://www.mirdb.org) and lncBase v.3-DIANA tool (https://diana.e-ce.uth.gr/lncbasev3), respectively.
3. Results
Antioxidant Genes Control Significant Molecular Functions and Biological Pathways
The Gene Ontology (GO) analysis revealed a significant enrichment of the 21 genes selected in this study across 10 molecular functions (Figure 1A). Molecular functions are listed hierarchically from top to bottom, with each gene transcript potentially associated with multiple functions. Catalytic activity (GO:0003824) emerged as the predominant GO category, with a high number of genes contributing (18), and encompassing functions such as oxidoreductase activity (GO:0016491) and transferase activity (GO:0016740); (Figure 1A). Notably, one of the most significant GO predictions included antioxidant activity (GO:0016209). The most significant molecular pathways involving a larger number of genes include the detoxification of ROS and cellular responses to chemical stress (Figure 1 B), glutathione conjugation, phase II - Conjugation of compounds, biological oxidations, and Cellular responses to stress. Remarkably, 3 genes play a key role in the FOXO-mediated transcription pathway (Figure 1B), underscoring their specific role in these cellular processes.
Expression and Intraovarian Localization of the Antioxidant Enzymes
Computational analysis by the Ovarian Kaleidoscope database revealed that all the selected antioxidant enzymes are expressed in the human ovary at different levels and 15 mRNAs had been found inside the exosomes (Table 1).
Concerning their intraovarian localization, GCLC, GCLM, GLRX2, GSR, SOD1, SOD2 and TXNRD1 were found in the oocyte. Only SOD1 and SOD2 show ubiquitous intraovarian expression. The localization of TXRND2, PRDX3, MGST1, GSPT1, GSTM1 remains undetermined. Unique localization is shown by GPX1, reported in LCs, GSTA4, present in the T compartment, GSTT1 and TNX2 in the GCs, and TXNRD1 in the oocyte (Table 2).
Transcriptome data analysis revealed a higher expression level of GSTP1, SOD1, and GPX3 with 340.3, 305.2 and 160.3 normalized Transcripts per Million (NTPM) in the human ovary, respectively (Figure 2).
LncRNA-miRNA-mRNA Competing Endogenous rna Networks
The search for miRNAs targeting the antioxidant enzyme mRNAs produced results for some of the initially selected enzymes. Specifically, we found 10 mRNAs targeted by 22 miRNAs (Table 3).
For the protein-encoding genes not listed in the table, no data on miRNAs targeting them is currently available in public databases or the literature. As shown in Table 3 and Figure 3, different target mRNAs may be regulated by the same miRNAs and multiple miRNAs may regulate a single mRNA (Figure 3).
The search for the lncRNAs, with at least two target binding sites for the 22 miRNAs, returned only 10 miRNAs sponged by the 22 lncRNAs (Table 4).
As reported in Figure 4, a single lncRNA can sponge different miRNAs and a single miRNA can interact with different lncRNAs. Among the identified lncRNAs, XIST and MALAT1 show the highest number of interactors (Table 4 and Figure 4).
As reported in Materials and Methods, the competing endogenous RNA networks (ceRNA-network) were designed considering the miRNAs targeting antioxidant enzymes’ mRNAs and the lncRNAs targeting these miRNAs. The resulting ceRNA-network, namely “antioxidant ceRNA-network“, showed that PRDX3, SOD1, SOD2, GSR and CAT transcripts can take part in different regulatory loops involving lncRNAs and miRNAs (Figure 5A)
To identify the ovarian “antioxidant ceRNA network”, we focused on lncRNAs and miRNAs expressed in the ovary. We found 10 miRNAs and 5 lncRNAs interacting with our 5 mRNAs inside the ovary. The network depicted in Figure 5B represents the identified lncRNAs that are part of different redundant networks. Through the regulation of 4 miRNAs, XIST may control GSR, CAT, SOD2, SOD1 and PRDX3. By sponging 3 miRNAs, MALAT1 may control three of them, SOD2, SOD1 and PRDX3. The action of NEAT and SNHG1 seems to specifically target the superoxide activity, whereas FGD5-AS1 is involved only in PRDX3 regulation (Figure 5B).
4. Discussion
The ovarian function relies on a fine regulation of redox balance that governs follicular development by activating specific pathways and preventing oxidative damage to germ cells. ROS signaling is a double-edged sword, playing essential roles in normal ovarian function and contributing to various ovarian pathologies when dysregulated [2,4,5]. During follicular development, moderate levels of ROS act as signaling molecules crucial for follicular maturation, oocyte meiosis, and ovulation. Controlled ROS levels ensure the atresia of non-dominant follicles, allowing only the healthiest to mature. The LH surge increases ROS production, facilitating follicular wall breakdown and oocyte release [23]. ROS also play a key role in the inflammatory response essential for ovulation and regulate genes involved in proteolysis and tissue remodeling [23,24]. Additionally, ROS impact luteal cell survival and function, affecting progesterone production, luteal phase duration, and angiogenesis for corpus luteum maintenance [6,25,26,27]. Accumulating evidence demonstrates that ROS are key signals in the initiation of apoptosis in antral follicles and granulosa cells of antral follicles by diverse stimuli, such as gonadotropin withdrawal, exposure to exogenous toxicants, and exposure to ionizing radiation, and that antioxidants protect against these stimuli [28].
In the present study, by focusing on mRNAs and ncRNAs present in the ovary and taking into account only validated ncRNA interactions, we built up an ovarian antioxidant ceRNA network, namely OvAnOx ceRNA, that is composed of 5 mRNAs (SOD1, SOD2, CAT, PRDX3, GR), 10 miRNAs and 5 lncRNAs (XIST, FGD5-AS1, MALAT1, NEAT1, SNHG1). Following the discussion of results regarding the antioxidant enzymes under study, the main components of OvAnOx ceRNA are here considered with reference to their role in the regulation of ovarian antioxidant activity and cellular processes.
Antioxidant Genes in the Human Ovary
According to the results, the genes included in our analysis are representative of all the catalytic reactions involved in ROS detoxification in the human ovary. When we focused on functional pathways, it emerged that our genes of interest are involved in the cellular response to stressing conditions, detoxification of ROS, biological oxidation, phase-II detoxification, GSH conjugation and FOXO-mediated transcription. FOXO transcription factors work together with Nrf2 to upregulate the expression of antioxidant enzymes, providing a coordinated defence against oxidative stress [29,30]. In accordance with our bioinformatics analysis, the 21 enzymes under study cover antioxidant activities in different intracellular and extracellular compartments. Indeed, most of them have been described as exosome cargo. A peculiar distribution in the oocyte, granulosa and theca cells and extracellular environment is also reported. Of note, only one paper described the presence of antioxidant enzymes in exosomes released in the culture media of mammalian granulosa cells [31]. The observation that TXNRD1 is uniquely expressed in the oocyte, GSTT1 and TXN2 in GCs, and GPX1 and GSTM2 in LCs, might deserve attention in the attempt to characterize the role of antioxidant enzymes in the ovary.
Among the selected enzymes, the most expressed gene is GSTP1 followed by SOD1, and GPX3, suggesting a prominent role of these genes in the ovarian antioxidant defence.
SODs are involved in the initial and most important step for controlling the redox state by catalyzing the transformation of anion superoxide O2• into molecular oxygen (O2) and hydrogen peroxide (H2O2) [32]. Superoxide anion is one of the first ROS formed during the reduction of molecular oxygen during metabolism and plays a key role in redox signaling pathways. Catalase (CAT), PRDX (peroxiredoxin), and GPX catalyze the conversion of hydrogen peroxide into water after the dismutation event [33,34,35,36]. In addition to GPX, many enzymes included in this study use glutathione (GSH) as an electron donor. Many reductive cellular enzyme systems depend upon the use of the tripeptide glutathione. Reduced GSH is oxidized to GSSG (oxidized glutathione) by GPX. The conversion of GSSG to GSH via glutathione reductase (GSR) with NADPH consumption is a common enzymatic method for sustaining GSH in most tissues [37,38]. Thus, the ability of cells to scavenge oxidants is fundamentally dependent on this entire process, known as "GSH recycling" [39]. By catalyzing the conjugation of reactive metabolites with GSH, glutathione transferase (GST) is essential in the detoxification process [40]. Glutamate cysteine ligase (GCL) and glutathione synthetase (GS) can catalyze the de novo synthesis of GSH from glutamate, cysteine, and glycine [41]. Glutaredoxin (GRX) (also known as thioltransferase) catalyzes the reduction of protein disulfides and mixed disulfides between proteins and GSH [42]. An important reductive system is represented by Thioredoxin Reductase (TXNRD) and Thioredoxin (TRX) [43,44]. TRX reduces oxidized proteins by donating electrons, which are replenished by TRXRD using NADPH [43,44].
The OvAnOx ceRNA Network
mRNA Components
The mRNA components of the OvAnOx ceRNA network, SOD1, SOD2, CAT, GSR and PRDX3, form a critical network for defence against oxidative stress and maintenance of a redox state suitable for proper ovarian function. Numerous knock-out mouse models have been used to explore the role of the enzymes included in the OvAnOx ceRNA network. SOD1 deficient mice show reduced fertility, with a reduction in preovulatory follicles and corpora lutea [45]. By contrast, in SOD2-deficient mice all follicular phases were detected, and viable pups were produced when their ovaries are transplanted into wild-type mice, indicating that SOD2 plays a less significant role than SOD1 [45]. Unchanged is the fertility of mice with an inactivating mutation in the GSR gene [46,47] or in CAT gene [48].
SOD1 and SOD2 are absent in primordial and primary follicles. SOD2 appears in secondary follicles, while SOD1 is first seen in theca cells after antral cavity formation and in granulosa cells at the dominant follicle stage [49]. Both isoforms are found in follicular fluid, with increased amounts and activity during antral development [49]. In luteinized granulosa and theca cells, SOD1 and SOD2 are highly expressed. Their enzymatic activity decreases with follicular growth, potentially inhibiting estrogen synthesis by suppressing FSH-induced aromatase in granulosa cells. SOD activity peaks at pro-oestrus with reduced superoxide radicals compared to the estrous stage [49]). During corpus luteum regression, increased ROS levels coincide with reduced SOD1 and increased SOD2, addressing mitochondrial ROS from cytokines and inflammation [50]. Aging is linked to decreased SODs and catalase in granulosa cells, contributing to reproductive decline [51]. Oxidative stress from SOD2 deficiency inhibits progestin and estradiol production in granulosa cells by affecting key steroidogenic enzymes, and SOD1 activity varies in PCOS women [52,53].
Oocytes experience increased ROS levels due to active metabolism in the preovulatory follicle and ovulation [23,54]. They express all three SOD isoforms, with SOD1 and SOD3 in the nucleus protecting DNA and regulating redox-sensitive gene transcription [55,56]. Age-related oxidative damage causes meiotic segregation errors, mitigated by extra SOD1 or SOD2 [57]. Oocytes have lower catalase expression compared to other cells, but catalase protects DNA during meiotic maturation and is involved in follicle development, the estrous cycle, and ovarian steroidogenesis [56]. Catalase activity increases in granulosa cells during ovarian growth and luteinization, aiding follicle selection and preventing ROS-mediated apoptosis in dominant follicles [51,58,59].
GSH synthesized in oocytes regulates the sulfur-oxygen reduction state, promotes cytoplasmic maturation, and protects against oxidative stress, improving spindle function and embryo development [60,61]. GSR expression decreases in aging oocytes, leading to oxidative damage and ovarian decline [62,63], but is highest during metaestrus, crucial for reproduction. GSH is essential for oocyte competence, influenced by gonadotropin signaling [64]. Oocytes have the highest GSR activity in the ovary, with GSH levels in cumulus cells increasing during maturation [65]. FSH therapy promotes GSH synthesis and prevents apoptosis in antral follicles, but its antiapoptotic effect is reduced if GSH synthesis is inhibited [66,67].
PRDX3 expression decreases during luteinization of preovulatory follicles in pigs and is stimulated by gonadotropins in theca cells, aiding the antioxidant system during ovulation. In aged mouse oocytes, Prdx3 mRNA expression is reduced, increasing oxidative stress sensitivity [68]. Mitochondrial antioxidants Prdx3 decrease with age in mice ovaries [22].
lncRNAs Components
In recent years, the role of lncRNA in oxidative stress has emerged specifically in oxidative stress-related diseases such as neurodegenerative pathologies, atherosclerosis and diabetes [69,70]. ceRNA has made little progress in some female reproductive diseases, particularly in PCOS disease [17,71,72,73,74,75,76,77,78,79,80]), indicating that the effect of ceRNAs in female reproduction is poorly understood and needs to be further explored. At the best of our knowledge, no paper investigated their role in the regulation of ovarian OS, a condition known to be involved in female reproductive dysfunctions [2,4,5,6].
The lncRNA XIST triggers X chromosome inactivation [81] and regulates oocyte loss by suppressing miR-23b-3p/miR-29a-3p and upregulating STX17 in perinatal mouse [2]ovaries [82]. Highly expressed in fetal ovaries, XIST is downregulated after birth as the primordial follicle pool forms. XIST accelerates oocyte autophagy during perinatal oocyte loss [82]. XIST is expressed early in unfertilized oocytes and pronuclei-stage zygotes [83]. A ceRNA network incorporating XIST was constructed to predict revealed differences in GCs from patients with EM [84]. Downregulated in PCOS patients' serum, it correlates with adverse pregnancy outcomes [85].
MALAT1 influences oxidative stress response, acting as an antioxidant by lowering Keap1 levels, thereby activating and stabilizing Nrf2 in H2O2-induced human umbilical vein endothelial cells (HUVECs). This enhances antioxidant capacity and reduces oxidative damage. MALAT1 also regulates Nrf2 and, in addition, can activate the p38MAPK pathway to modulate apoptosis and oxidative stress [86,87]. In ovarian function, MALAT1 knockdown increases apoptosis and reduces proliferation in granulosa cells by promoting P53 degradation [88]. MALAT1 regulates ovarian follicular atresia, apoptosis, and steroid synthesis, and is upregulated in KGN cells after AMH stimulation [89,90]. PCOS patients show lower MALAT1 levels, suggesting its potential role in PCOS pathogenesis and targeted therapy [73].
NEAT1, a highly conserved lncRNA, is highly expressed in PCOS patients, promoting the expression of Androgen Receptor (AR), follistatin (FST), and IRS-2, potentially involved in PCOS pathogenesis [73]. NEAT1 exacerbates metabolic disorders in PCOS mice by downregulating miR-324-3p and upregulating BRD3 [91]. In Neat1 knockout mice, corpus luteum formation is impaired, leading to decreased fertility, which can be partially rescued by progesterone [92]. NEAT1 is downregulated in premature ovarian failure (POF) mice, where it modulates the STC2/MAPK pathway to reduce apoptosis and autophagy [93].
The miRNAs Components
Over the last decade, research has highlighted the regulatory interplay between miRNAs and redox signaling. Oxidative stress can regulate miRNAs, and miRNAs can influence cellular redox status [94]. ROS exposure can inhibit Dicer activity, delaying miRNA maturation, and affect pri-miRNA structures and promoter methylation [95]. Many ROS-responsive miRNAs, in turn, influence the Nrf2 system [69,95,96].
Specific miRNAs play crucial roles in ovarian function and oxidative stress regulation [97,98,99,100,101,102]. miR-214 offers protection against oxidative damage by targeting GSR and cytochrome P450 oxidoreductase (POR), and is involved in cell survival, embryonic development, and ovarian cancer resistance [103]. miR-23b-3p promotes oocyte autophagy by reducing mature miR-23b-3p levels, which is crucial for oocyte death regulation [82]. miR-377-3p is proposed as a marker of oocyte quality, aiding in predicting ovarian superovulation potential [104]. miR-206 is linked to PCOS, regulates granulosa cell viability and apoptosis via the PI3K/AKT pathway, and is a potential biomarker for superovulation response [105,106,107,108].
Additionally, miR-206 regulates oocyte maturation and granulosa cell development by targeting AURKA [105,109]. RNAseq analysis in goat ovary showed miR-206 upregulated in ovarian stroma, indicating roles in ovarian organogenesis and hormone secretion by oocyte meiosis [109].
miR-26a-5p is upregulated in PCOS, involved in corpus luteum development, and key in reproductive span regulation. miR-383-5p decreases in PCOS patients, suppresses the PI3K/AKT pathway, and enhances KGN cell apoptosis [110,111,112,113,114]. These miRNAs modulate redox status and are crucial for ovarian health, influencing processes from oocyte maturation to hormone secretion and disease resistance [115].
5. Conclusions
Many reproductive disorders, such as polycystic ovarian syndrome (PCOS), endometriosis, and unexplained infertility are pathological effects of decreased antioxidant defence systems. Decreased antioxidant systems have also been linked to age-related reproductive reduction. Considering the importance of redox balance in ovarian functions and the controversial use of antioxidant therapies in the treatment of female fertility [7], the results of the present bioinformatic study represent a valuable contribution to the knowledge on selectively targeting redox-modulating systems.
Overall, based on the literature, all the components of the OvAnOx ceRNA are known to play a role in ovarian physiology. From our bioinformatic analysis, an antioxidant action based on superoxide and hydrogen peroxide scavenging and glutathione recycling is under the control of ncRNAs involved in numerous ovarian functions other than redox modulation. Indeed, we have predicted that antioxidant enzymes (i.e. SOD1, SOD2, CAT, GRS and PRDX3) can act within a complex regulatory network that receives and transmits signals from a variety of intracellular processes including regulation of ovarian reserve, follicle dynamics, apoptosis, oocyte maturation under physiological and pathological conditions. Based on these observations, the OvAnOx ceRNA represents a promising tool to investigate the complex dialogue between redox potential and ovarian signalling pathways involved in reproductive health, aging and diseases.
Author Contributions
For “Conceptualization, T.C. and D.P.C.; methodology, D.E.G and B.R.; data curation B.R. and D.E.G; writing—original draft preparation, T.C., D.P.C., D.E.G and B.R.; writing—review and editing, T.C, D.P.C., D.E.G and B.R.; visualization, B.R. and D.E.G.; supervision, T.C and D.P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gershon, E.; Dekel, N. Newly Identified Regulators of Ovarian Folliculogenesis and Ovulation. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Amicarelli, F.; Carbone, M.C.; Monteleone, P.; Caserta, D.; Marci, R.; Artini, P.G.; Piomboni, P.; Focarelli, R. Cellular and molecular aspects of ovarian follicle ageing. Hum Reprod Update 2008, 14, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Tatone, C.; Di Emidio, G.; Placidi, M.; Rossi, G.; Ruggieri, S.; Taccaliti, C.; D'Alfonso, A.; Amicarelli, F.; Guido, M. AGEs-related dysfunctions in PCOS: evidence from animal and clinical research. J Endocrinol 2021, 251, R1–R9. [Google Scholar] [CrossRef] [PubMed]
- Ruder, E.H.; Hartman, T.J.; Goldman, M.B. Impact of oxidative stress on female fertility. Curr Opin Obstet Gynecol 2009, 21, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhao, Q.; Li, Y.; Zheng, Z.; Kong, X.; Shu, C.; Liu, Y.; Shi, Y. The role of oxidative stress in ovarian aging: a review. J Ovarian Res 2022, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Gao, Y.; Feng, Z.; Zhang, B.; Na, Z.; Li, D. Reactive oxygen species and ovarian diseases: Antioxidant strategies. Redox Biol 2023, 62, 102659. [Google Scholar] [CrossRef] [PubMed]
- Showell, M.G.; Mackenzie-Proctor, R.; Jordan, V.; Hart, R.J. Antioxidants for female subfertility. Cochrane Database Syst Rev 2020, 8, CD007807. [Google Scholar] [CrossRef]
- de Almeida, A.; de Oliveira, J.; da Silva Pontes, L.V.; de Souza Junior, J.F.; Goncalves, T.A.F.; Dantas, S.H.; de Almeida Feitosa, M.S.; Silva, A.O.; de Medeiros, I.A. ROS: Basic Concepts, Sources, Cellular Signaling, and its Implications in Aging Pathways. Oxid Med Cell Longev 2022, 2022, 1225578. [Google Scholar] [CrossRef]
- Sies, H.; Mailloux, R.J.; Jakob, U. Fundamentals of redox regulation in biology. Nat Rev Mol Cell Biol 2024. [CrossRef]
- Lei, X.G.; Zhu, J.H.; Cheng, W.H.; Bao, Y.; Ho, Y.S.; Reddi, A.R.; Holmgren, A.; Arner, E.S. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol Rev 2016, 96, 307–364. [Google Scholar] [CrossRef]
- Nemeth, K.; Bayraktar, R.; Ferracin, M.; Calin, G.A. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat Rev Genet 2024, 25, 211–232. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat Rev Mol Cell Biol 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Gou, L.T.; Zhu, Q.; Liu, M.F. Small RNAs: An expanding world with therapeutic promises. Fundam Res 2023, 3, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J Integr Bioinform 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Caponnetto, A.; Ferrara, C.; Fazzio, A.; Agosta, N.; Scribano, M.; Vento, M.E.; Borzì, P.; Barbagallo, C.; Stella, M.; Ragusa, M.; et al. A Circular RNA Derived from the Pumilio 1 Gene Could Regulate PTEN in Human Cumulus Cells. Genes 2024, 15, 124. [Google Scholar] [CrossRef] [PubMed]
- Caponnetto, A.; Battaglia, R.; Ferrara, C.; Vento, M.E.; Borzì, P.; Paradiso, M.; Scollo, P.; Purrello, M.; Longobardi, S.; D'Hooghe, T.; et al. Down-regulation of long non-coding RNAs in reproductive aging and analysis of the lncRNA-miRNA-mRNA networks in human cumulus cells. J Assist Reprod Genet 2022, 39, 919–931. [Google Scholar] [CrossRef]
- Barbagallo, D.; Palermo, CI.; Barbagallo, C.; Battaglia, R.; Caponnetto, A.; Spina, V.; Ragusa, M.; Di Pietro, C.; Scalia, G.; Purrello, M. ; Competing endogenous RNA network mediated by circ_3205 in SARS-CoV-2 infected cells. Cell Mol Life Sci 2022, 79, 75. [Google Scholar] [CrossRef] [PubMed]
- Nejadi Orang, F.; Abdoli Shadbad, M. Competing endogenous RNA networks and ferroptosis in cancer: novel therapeutic targets. Cell Death Dis 2024, 15, 357. [Google Scholar] [CrossRef]
- Ciesielska, S.; Slezak-Prochazka, I.; Bil, P.; Rzeszowska-Wolny, J. Micro RNAs in Regulation of Cellular Redox Homeostasis. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Kinoshita, C.; Aoyama, K. The Role of Non-Coding RNAs in the Neuroprotective Effects of Glutathione. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol Reprod 2011, 84, 775–782. [Google Scholar] [CrossRef]
- Shkolnik, K.; Tadmor, A.; Ben-Dor, S.; Nevo, N.; Galiani, D.; Dekel, N. Reactive oxygen species are indispensable in ovulation. Proc Natl Acad Sci U S A 2011, 108, 1462–1467. [Google Scholar] [CrossRef] [PubMed]
- Kala, M.; Shaikh, M.V.; Nivsarkar, M. Equilibrium between anti-oxidants and reactive oxygen species: a requisite for oocyte development and maturation. Reprod Med Biol 2017, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Jamil, M.; Debbarh, H.; Aboulmaouahib, S.; Aniq Filali, O.; Mounaji, K.; Zarqaoui, M.; Saadani, B.; Louanjli, N.; Cadi, R. Reactive oxygen species in reproduction: harmful, essential or both? Zygote 2020, 28, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Liu, X.; Pei, X.; Gao, H.; Pan, P.; Yang, Y. Mechanism of Mitochondrial Homeostasis Controlling Ovarian Physiology. Endocrinology 2022, 164. [Google Scholar] [CrossRef]
- Wang, L.; Tang, J.; Wang, L.; Tan, F.; Song, H.; Zhou, J.; Li, F. Oxidative stress in oocyte aging and female reproduction. J Cell Physiol 2021, 236, 7966–7983. [Google Scholar] [CrossRef]
- Devine, P.J.; Perreault, S.D.; Luderer, U. Roles of reactive oxygen species and antioxidants in ovarian toxicity. Biol Reprod 2012, 86, 27. [Google Scholar] [CrossRef]
- Klotz, L.O.; Sanchez-Ramos, C.; Prieto-Arroyo, I.; Urbanek, P.; Steinbrenner, H.; Monsalve, M. Redox regulation of FoxO transcription factors. Redox Biol 2015, 6, 51–72. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid Redox Signal 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Saeed-Zidane, M.; Linden, L.; Salilew-Wondim, D.; Held, E.; Neuhoff, C.; Tholen, E.; Hoelker, M.; Schellander, K.; Tesfaye, D. Cellular and exosome mediated molecular defense mechanism in bovine granulosa cells exposed to oxidative stress. PLoS One 2017, 12, e0187569. [Google Scholar] [CrossRef] [PubMed]
- Case, A.J. On the Origin of Superoxide Dismutase: An Evolutionary Perspective of Superoxide-Mediated Redox Signaling. Antioxidants (Basel) 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Bhella, D.; Lindsay, J.G. Reconstitution of the mitochondrial PrxIII antioxidant defence pathway: general properties and factors affecting PrxIII activity and oligomeric state. J Mol Biol 2007, 372, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Yewdall, N.A.; Peskin, A.V.; Hampton, M.B.; Goldstone, D.C.; Pearce, F.G.; Gerrard, J.A. Quaternary structure influences the peroxidase activity of peroxiredoxin 3. Biochem Biophys Res Commun 2018, 497, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Alrumaihi, F.; Sarwar, T.; Babiker, A.Y.; Khan, A.A.; Prabhu, S.V.; Rahmani, A.H. Exploring Therapeutic Potential of Catalase: Strategies in Disease Prevention and Management. Biomolecules 2024, 14. [Google Scholar] [CrossRef]
- Trenz, T.S.; Delaix, C.L.; Turchetto-Zolet, A.C.; Zamocky, M.; Lazzarotto, F.; Margis-Pinheiro, M. Going Forward and Back: The Complex Evolutionary History of the GPx. Biology (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Chen, T.H.; Wang, H.C.; Chang, C.J.; Lee, S.Y. Mitochondrial Glutathione in Cellular Redox Homeostasis and Disease Manifestation. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic Biol Med 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic Res 1999, 31, 273–300. [Google Scholar] [CrossRef]
- Mazari, A.M.A.; Zhang, L.; Ye, Z.W.; Zhang, J.; Tew, K.D.; Townsend, D.M. The Multifaceted Role of Glutathione S-Transferases in Health and Disease. Biomolecules 2023, 13. [Google Scholar] [CrossRef]
- Zhang, H.; Forman, H.J. Glutathione synthesis and its role in redox signaling. Semin Cell Dev Biol 2012, 23, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.M.; Mieyal, J.J. Protein-thiol oxidation and cell death: regulatory role of glutaredoxins. Antioxid Redox Signal 2012, 17, 1748–1763. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Fan, C.; Zhao, J.; Wang, L.; Duan, D.; Shen, T.; Li, X. Fluorescent Probes for Mammalian Thioredoxin Reductase: Mechanistic Analysis, Construction Strategies, and Future Perspectives. Biosensors (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Yang, B.; Lin, Y.; Huang, Y.; Shen, Y.Q.; Chen, Q. Thioredoxin (Trx): A redox target and modulator of cellular senescence and aging-related diseases. Redox Biol 2024, 70, 103032. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Dionne, L.; Guo, Q.; Kumar, T.R.; Lebovitz, R.M. Ovarian function in superoxide dismutase 1 and 2 knockout mice. Endocrinology 1998, 139, 4008–4011. [Google Scholar] [CrossRef] [PubMed]
- Pretsch, W. Glutathione reductase activity deficiency in homozygous Gr1a1Neu mice does not cause haemolytic anaemia. Genet Res 1999, 73, 1–5. [Google Scholar] [CrossRef]
- Rogers, L.K.; Bates, C.M.; Welty, S.E.; Smith, C.V. Diquat induces renal proximal tubule injury in glutathione reductase-deficient mice. Toxicol Appl Pharmacol 2006, 217, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; Xiong, Y.; Ma, W.; Spector, A.; Ho, D.S. Mice lacking catalase develop normally but show differential sensitivity to oxidant tissue injury. J Biol Chem 2004, 279, 32804–32812. [Google Scholar] [CrossRef]
- Wang, S.; He, G.; Chen, M.; Zuo, T.; Xu, W.; Liu, X. The Role of Antioxidant Enzymes in the Ovaries. Oxid Med Cell Longev 2017, 2017, 4371714. [Google Scholar] [CrossRef]
- Vu, H.V.; Lee, S.; Acosta, T.J.; Yoshioka, S.; Abe, H.; Okuda, K. Roles of prostaglandin F2alpha and hydrogen peroxide in the regulation of Copper/Zinc superoxide dismutase in bovine corpus luteum and luteal endothelial cells. Reprod Biol Endocrinol 2012, 10, 87. [Google Scholar] [CrossRef]
- Tatone, C.; Carbone, M.C.; Falone, S.; Aimola, P.; Giardinelli, A.; Caserta, D.; Marci, R.; Pandolfi, A.; Ragnelli, A.M.; Amicarelli, F. Age-dependent changes in the expression of superoxide dismutases and catalase are associated with ultrastructural modifications in human granulosa cells. Mol Hum Reprod 2006, 12, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Bizon, A.; Tchorz, A.; Madej, P.; Lesniewski, M.; Wojtowicz, M.; Piwowar, A.; Franik, G. The Activity of Superoxide Dismutase, Its Relationship with the Concentration of Zinc and Copper and the Prevalence of rs2070424 Superoxide Dismutase Gene in Women with Polycystic Ovary Syndrome-Preliminary Study. J Clin Med 2022, 11. [Google Scholar] [CrossRef]
- Seleem, A.K.; El Refaeey, A.A.; Shaalan, D.; Sherbiny, Y.; Badawy, A. Superoxide dismutase in polycystic ovary syndrome patients undergoing intracytoplasmic sperm injection. J Assist Reprod Genet 2014, 31, 499–504. [Google Scholar] [CrossRef]
- Pandey, A.N.; Chaube, S.K. A moderate increase of hydrogen peroxide level is beneficial for spontaneous resumption of meiosis from diplotene arrest in rat oocytes cultured in vitro. Biores Open Access 2014, 3, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Matos, L.; Stevenson, D.; Gomes, F.; Silva-Carvalho, J.L.; Almeida, H. Superoxide dismutase expression in human cumulus oophorus cells. Mol Hum Reprod 2009, 15, 411–419. [Google Scholar] [CrossRef]
- Cetica, P.D.; Pintos, L.N.; Dalvit, G.C.; Beconi, M.T. Antioxidant enzyme activity and oxidative stress in bovine oocyte in vitro maturation. IUBMB Life 2001, 51, 57–64. [Google Scholar] [CrossRef]
- Perkins, A.T.; Greig, M.M.; Sontakke, A.A.; Peloquin, A.S.; McPeek, M.A.; Bickel, S.E. Increased levels of superoxide dismutase suppress meiotic segregation errors in aging oocytes. Chromosoma 2019, 128, 215–222. [Google Scholar] [CrossRef]
- Behl, R.; Pandey, R.S. FSH induced stimulation of catalase activity in goat granulosa cells in vitro. Anim Reprod Sci 2002, 70, 215–221. [Google Scholar] [CrossRef]
- Serke, H.; Bausenwein, J.; Hirrlinger, J.; Nowicki, M.; Vilser, C.; Jogschies, P.; Hmeidan, F.A.; Blumenauer, V.; Spanel-Borowski, K. Granulosa cell subtypes vary in response to oxidized low-density lipoprotein as regards specific lipoprotein receptors and antioxidant enzyme activity. J Clin Endocrinol Metab 2010, 95, 3480–3490. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.D.A.; Adona, P.R.; Guemra, S.; Damiao, B.C.M. Oxidative homeostasis in oocyte competence for in vitro embryo development. Anim Sci J 2019, 90, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Luciano, A.M.; Goudet, G.; Perazzoli, F.; Lahuec, C.; Gerard, N. Glutathione content and glutathione peroxidase expression in in vivo and in vitro matured equine oocytes. Mol Reprod Dev 2006, 73, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Katz-Jaffe, M.G.; Lane, S.L.; Parks, J.C.; McCallie, B.R.; Makloski, R.; Schoolcraft, W.B. Antioxidant Intervention Attenuates Aging-Related Changes in the Murine Ovary and Oocyte. Life (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Li, J.; Yu, Y.; Zhang, W.; Song, M.; Liu, Z.; Min, Z.; Hu, H.; Jing, Y.; et al. Single-Cell Transcriptomic Atlas of Primate Ovarian Aging. Cell 2020, 180, 585–600 e519. [Google Scholar] [CrossRef]
- Dumollard, R.; Ward, Z.; Carroll, J.; Duchen, M.R. Regulation of redox metabolism in the mouse oocyte and embryo. Development 2007, 134, 455–465. [Google Scholar] [CrossRef]
- Kaneko, T.; Iuchi, Y.; Kawachiya, S.; Fujii, T.; Saito, H.; Kurachi, H.; Fujii, J. Alteration of glutathione reductase expression in the female reproductive organs during the estrous cycle. Biol Reprod 2001, 65, 1410–1416. [Google Scholar] [CrossRef]
- Tsai-Turton, M.; Luderer, U. Opposing effects of glutathione depletion and follicle-stimulating hormone on reactive oxygen species and apoptosis in cultured preovulatory rat follicles. Endocrinology 2006, 147, 1224–1236. [Google Scholar] [CrossRef]
- Hoang, Y.D.; Nakamura, B.N.; Luderer, U. Follicle-stimulating hormone and estradiol interact to stimulate glutathione synthesis in rat ovarian follicles and granulosa cells. Biol Reprod 2009, 81, 636–646. [Google Scholar] [CrossRef]
- Park, J.I.; Jeon, H.J.; Jung, N.K.; Jang, Y.J.; Kim, J.S.; Seo, Y.W.; Jeong, M.; Chae, H.Z.; Chun, S.Y. Periovulatory expression of hydrogen peroxide-induced sulfiredoxin and peroxiredoxin 2 in the rat ovary: gonadotropin regulation and potential modification. Endocrinology 2012, 153, 5512–5521. [Google Scholar] [CrossRef]
- Hernández-Cruz, E.Y.; Arancibia-Hernández, Y.L.; Loyola-Mondragón, D.Y.; Pedraza-Chaverri, J. Oxidative Stress and Its Role in Cd-Induced Epigenetic Modifications: Use of Antioxidants as a Possible Preventive Strategy. Oxygen 2022, 2, 177–212. [Google Scholar] [CrossRef]
- Infante-Menendez, J.; Gonzalez-Lopez, P.; Huertas-Larez, R.; Gomez-Hernandez, A.; Escribano, O. Oxidative Stress Modulation by ncRNAs and Their Emerging Role as Therapeutic Targets in Atherosclerosis and Non-Alcoholic Fatty Liver Disease. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Fan, H.; Zhou, D.; Zhang, X.; Jiang, M.; Kong, X.; Xue, T.; Gao, L.; Lu, D.; Tao, C.; Wang, L. hsa_circRNA_BECN1 acts as a ceRNA to promote polycystic ovary syndrome progression by sponging the miR-619-5p/Rab5b axis. Mol Hum Reprod 2023, 29. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Gong, J.; Guo, Y.; Li, Y.; Huang, H.; Liu, X. Construction of a ceRNA network in polycystic ovary syndrome (PCOS) driven by exosomal lncRNA. Front Genet 2022, 13, 979924. [Google Scholar] [CrossRef] [PubMed]
- ElMonier, A.A.; El-Boghdady, N.A.; Fahim, S.A.; Sabry, D.; Elsetohy, K.A.; Shaheen, A.A. LncRNA NEAT1 and MALAT1 are involved in polycystic ovary syndrome pathogenesis by functioning as competing endogenous RNAs to control the expression of PCOS-related target genes. Noncoding RNA Res 2023, 8, 263–271. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, L.; Cao, Y.; Zhai, J. Construction of a ceRNA-based lncRNA-mRNA network to identify functional lncRNAs in polycystic ovarian syndrome. Aging (Albany NY) 2021, 13, 8481–8496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhou, Z.; Wang, P.; He, X.; Liu, Y.; Chu, M. The SLC19A1-AS/miR-1343/WNT11 axis is a novel positive regulatory ceRNA network governing goat granulosa cell proliferation. Int J Biol Macromol 2024, 264, 130658. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, Y.; Yang, L.; Du, X.; Li, Q. Nuclear lncRNA NORSF reduces E2 release in granulosa cells by sponging the endogenous small activating RNA miR-339. BMC Biol 2023, 21, 221. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, L.; Sulaiman, Z.; Wang, C.; Wang, L.; Zhu, J.; Liu, S.; Cheng, Z. Comprehensive analysis of lncRNA-miRNA-mRNA ceRNA network and key genes in granulosa cells of patients with biochemical primary ovarian insufficiency. J Assist Reprod Genet 2024, 41, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Gupta, P.; Sahoo, S.; Mallick, B. Non-coding RNAs and their cross-talks impacting reproductive health of women. Wiley Interdiscip Rev RNA 2022, 13, e1695. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Lin, J.Z. Identification of long non-coding RNA biomarkers and signature scoring, with competing endogenous RNA networks- targeted drug candidates for recurrent implantation failure. Hum Fertil (Camb) 2022, 25, 983–992. [Google Scholar] [CrossRef]
- Hu, H.; Jia, Q.; Xi, J.; Zhou, B.; Li, Z. Integrated analysis of lncRNA, miRNA and mRNA reveals novel insights into the fertility regulation of large white sows. BMC Genomics 2020, 21, 636. [Google Scholar] [CrossRef]
- Alfeghaly, C.; Castel, G.; Cazottes, E.; Moscatelli, M.; Moinard, E.; Casanova, M.; Boni, J.; Mahadik, K.; Lammers, J.; Freour, T.; et al. XIST dampens X chromosome activity in a SPEN-dependent manner during early human development. Nat Struct Mol Biol 2024. [CrossRef]
- Zhou, M.; Liu, X.; Qiukai, E.; Shang, Y.; Zhang, X.; Liu, S.; Zhang, X. Long non-coding RNA Xist regulates oocyte loss via suppressing miR-23b-3p/miR-29a-3p maturation and upregulating STX17 in perinatal mouse ovaries. Cell Death Dis 2021, 12, 540. [Google Scholar] [CrossRef] [PubMed]
- Avner, R.; Wahrman, J.; Richler, C.; Ayoub, N.; Friedmann, A.; Laufer, N.; Mitrani-Rosenbaum, S. X inactivation-specific transcript expression in mouse oocytes and zygotes. Mol Hum Reprod 2000, 6, 591–594. [Google Scholar] [CrossRef]
- Wu, R.; Li, J.; Li, J.; Zhang, N.; Zhou, W.; Ren, L.; Chen, Q.; Li, Y. Construction of Competing Endogenous RNA Networks Incorporating Transcription Factors to Reveal Differences in Granulosa Cells from Patients with Endometriosis. Genet Test Mol Biomarkers 2021, 25, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhu, H.; Li, Y.; Zhuang, J.; Cao, T.; Wang, Y. Expression of serum lncRNA-Xist in patients with polycystic ovary syndrome and its relationship with pregnancy outcome. Taiwan J Obstet Gynecol 2020, 59, 372–376. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Kowluru, R.A. Long Noncoding RNA MALAT1 and Regulation of the Antioxidant Defense System in Diabetic Retinopathy. Diabetes 2021, 70, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Zhang, R.; Song, X.; Ni, L.; Lai, Z.; Liu, C.; Ye, W. The long non-coding RNA MALAT1 activates Nrf2 signaling to protect human umbilical vein endothelial cells from hydrogen peroxide. Biochem Biophys Res Commun 2018, 495, 2532–2538. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiang, Y.; Song, Y.; Zhang, D.; Tan, L. MALAT1 downregulation is associated with polycystic ovary syndrome via binding with MDM2 and repressing P53 degradation. Mol Cell Endocrinol 2022, 543, 111528. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, P.; Lu, W. lncRNA MALAT1 Regulates Mouse Granulosa Cell Apoptosis and 17beta-Estradiol Synthesis via Regulating miR-205/CREB1 Axis. Biomed Res Int 2021, 2021, 6671814. [Google Scholar] [CrossRef]
- Tu, M.; Wu, Y.; Wang, F.; Huang, Y.; Qian, Y.; Li, J.; Lv, P.; Ying, Y.; Liu, J.; Liu, Y.; et al. Effect of lncRNA MALAT1 on the Granulosa Cell Proliferation and Pregnancy Outcome in Patients With PCOS. Front Endocrinol (Lausanne) 2022, 13, 825431. [Google Scholar] [CrossRef]
- Wu, L.; Tu, Z.; Bao, Y.; Zhai, Q.; Jin, L. Long noncoding RNA NEAT1 decreases polycystic ovary syndrome progression via the modulation of the microRNA-324-3p and BRD3 axis. Cell Biol Int 2022, 46, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Shimada, M.; Yanaka, K.; Mito, M.; Arai, T.; Takahashi, E.; Fujita, Y.; Fujimori, T.; Standaert, L.; Marine, J.C.; et al. The lncRNA Neat1 is required for corpus luteum formation and the establishment of pregnancy in a subpopulation of mice. Development 2014, 141, 4618–4627. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Ke, Y.; Qiu, P.; Gao, J.; Deng, G.P. LncRNA NEAT1 inhibits apoptosis and autophagy of ovarian granulosa cells through miR-654/STC2-mediated MAPK signaling pathway. Exp Cell Res 2023, 424, 113473. [Google Scholar] [CrossRef]
- Carbonell, T.; Gomes, A.V. MicroRNAs in the regulation of cellular redox status and its implications in myocardial ischemia-reperfusion injury. Redox Biol 2020, 36, 101607. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Wedel, S.; Cavinato, M.; Jansen-Durr, P. MicroRNA Regulation of Oxidative Stress-Induced Cellular Senescence. Oxid Med Cell Longev 2017, 2017, 2398696. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Ahmadi, Z.; Samarghandian, S.; Mohammadinejad, R.; Yaribeygi, H.; Sathyapalan, T.; Sahebkar, A. MicroRNA-mediated regulation of Nrf2 signaling pathway: Implications in disease therapy and protection against oxidative stress. Life Sci 2020, 244, 117329. [Google Scholar] [CrossRef]
- Santonocito, M.; Vento, M.; Guglielmino, M.R.; Battaglia, R.; Wahlgren, J.; Ragusa, M.; Barbagallo, D.; Borzì, P.; Rizzari, S.; Maugeri, M.; et al. Molecular characterization of exosomes and their microRNA cargo in human follicular fluid: bioinformatic analysis reveals that exosomal microRNAs control pathways involved in follicular maturation. Fertil Steril 2014, 102, 1751–1761.e1. [Google Scholar] [CrossRef]
- Battaglia, R.; Vento, M.E.; Ragusa, M.; Barbagallo, D.; La Ferlita, A.; Di Emidio, G.; Borzí, P.; Artini, P.G.; Scollo, P.; Tatone, C.; et al. MicroRNAs Are Stored in Human MII Oocyte and Their Expression Profile Changes in Reproductive Aging. Biol Reprod 2016, 95, 131. [Google Scholar] [CrossRef]
- Battaglia, R.; Vento, M.E.; Borzì, P.; Ragusa, M.; Barbagallo, D.; Arena, D.; Purrello, M.; Di Pietro, C. Non-coding RNAs in the Ovarian Follicle. Front Genet 2017, 8, 57. [Google Scholar] [CrossRef]
- Battaglia, R.; Musumeci, P.; Ragusa, M.; Barbagallo, D.; Scalia, M.; Zimbone, M.; Lo Faro, J.M.; Borzì, P.; Scollo, P.; Purrello, M.; et al. Ovarian aging increases small extracellular vesicle CD81+ release in human follicular fluid and influences miRNA profiles. Aging (Albany NY) 2020, 12, 12324–12341. [Google Scholar] [CrossRef]
- Amin, M.M.J.; Trevelyan, C.J.; Turner, N.A. MicroRNA-214 in Health and Disease. Cells 2021, 10. [Google Scholar] [CrossRef]
- Battaglia, R.; Caponnetto, A.; Caringella, A.M.; Cortone, A.; Ferrara, C.; Smirni, S.; Iannitti, R.; Purrello, M.; D'Amato, G.; Fioretti, B.; et al. Resveratrol Treatment Induces Mito-miRNome Modification in Follicular Fluid from Aged Women with a Poor Prognosis for In Vitro Fertilization Cycles. Antioxidants (Basel) 2022, 11, 1019. [Google Scholar] [CrossRef]
- Chang, W.; Wang, J.; Tao, D.; Zhang, Y.; He, J.; Shi, C. Identification of a novel miRNA from the ovine ovary by a combinatorial approach of bioinformatics and experiments. J Vet Med Sci 2016, 77, 1617–1624. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, C.; Yang, J.; Li, Y.; Feng, J.; Wang, B.; Zhang, J.; Guo, X. The Roles of the miRNAome and Transcriptome in the Ovine Ovary Reveal Poor Efficiency in Juvenile Superovulation. Animals (Basel) 2021, 11. [Google Scholar] [CrossRef]
- Zhou, J.; Jin, X.; Sheng, Z.; Zhang, Z. miR-206 serves an important role in polycystic ovary syndrome through modulating ovarian granulosa cell proliferation and apoptosis. Exp Ther Med 2021, 21, 179. [Google Scholar] [CrossRef]
- Gad, A.; Sanchez, J.M.; Browne, J.A.; Nemcova, L.; Laurincik, J.; Prochazka, R.; Lonergan, P. Plasma extracellular vesicle miRNAs as potential biomarkers of superstimulatory response in cattle. Sci Rep 2020, 10, 19130. [Google Scholar] [CrossRef]
- Zhang, Z.; Sang, M.; Liu, S.; Shao, J.; Cai, Y. Differential expression of long non-coding RNA Regulator of reprogramming and its molecular mechanisms in polycystic ovary syndrome. J Ovarian Res 2021, 14, 79. [Google Scholar] [CrossRef]
- Diaz, M.; Bassols, J.; Lopez-Bermejo, A.; de Zegher, F.; Ibanez, L. Low Circulating Levels of miR-451a in Girls with Polycystic Ovary Syndrome: Different Effects of Randomized Treatments. J Clin Endocrinol Metab 2020, 105. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Zou, X.; Liu, G.; Deng, M.; Sun, B.; Guo, Y.; Liu, D.; Li, Y. A Preliminary Study on the Characteristics of microRNAs in Ovarian Stroma and Follicles of Chuanzhong Black Goat during Estrus. Genes (Basel) 2020, 11. [Google Scholar] [CrossRef]
- De Nardo Maffazioli, G.; Baracat, E.C.; Soares, J.M.; Carvalho, K.C.; Maciel, G.A.R. Evaluation of circulating microRNA profiles in Brazilian women with polycystic ovary syndrome: A preliminary study. PLoS One 2022, 17, e0275031. [Google Scholar] [CrossRef]
- Ibrahim, S.; Taqi, M.O.; Sosa, A.S.A.; El-Naby, A.A.H.; Mahmoud, K.G.M.; Darwish, H.R.H.; Abd El Hameed, A.R.; Nawito, M.F. Spatiotemporal expression pattern of miR-205, miR-26a-5p, miR-17-5p, let-7b-5p, and their target genes during different stages of corpus luteum in Egyptian buffaloes. J Genet Eng Biotechnol 2022, 20, 37. [Google Scholar] [CrossRef]
- Kim, Y.Y.; Kim, K.S.; Kim, Y.J.; Kim, S.W.; Kim, H.; Ku, S.Y. Transcriptome Analyses Identify Potential Key microRNAs and Their Target Genes Contributing to Ovarian Reserve. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Fan, F.; Liang, C.; Zhou, Y.; Qiao, X.; Sun, Y.; Jiang, Y.; Kang, L. Variants of pri-miR-26a-5p polymorphisms are associated with values for chicken egg production variables and affects abundance of mature miRNA. Anim Reprod Sci 2019, 201, 93–101. [Google Scholar] [CrossRef]
- Kang, L.; Yang, C.; Wu, H.; Chen, Q.; Huang, L.; Li, X.; Tang, H.; Jiang, Y. miR-26a-5p Regulates TNRC6A Expression and Facilitates Theca Cell Proliferation in Chicken Ovarian Follicles. DNA Cell Biol 2017, 36, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, X.; Miao, S.; Cao, Q. MiR-383-5p promotes apoptosis of ovarian granulosa cells by targeting CIRP through the PI3K/AKT signaling pathway. Arch Gynecol Obstet 2022, 306, 501–512. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular functions (GO) (A) and associated biological pathways (B) of the 21 antioxidant enzymes-encoding genes selected for this study. The significance is reported as −log10 P-value for both panels (P < 0.05).
Figure 1.
Molecular functions (GO) (A) and associated biological pathways (B) of the 21 antioxidant enzymes-encoding genes selected for this study. The significance is reported as −log10 P-value for both panels (P < 0.05).
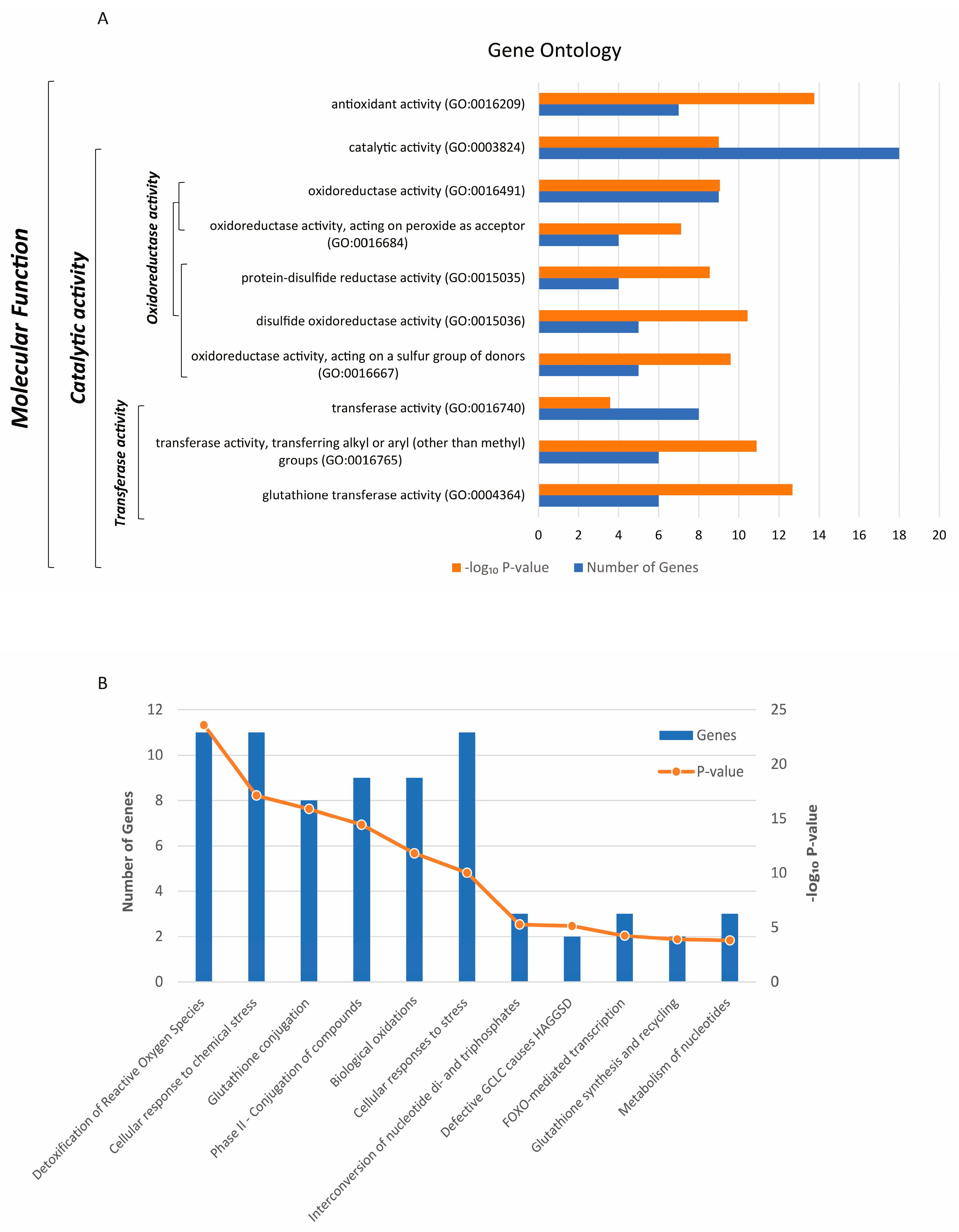
Figure 2.
Expression of the selected antioxidant enzymes, in terms of Transcripts, within the ovarian tissue.
Figure 2.
Expression of the selected antioxidant enzymes, in terms of Transcripts, within the ovarian tissue.
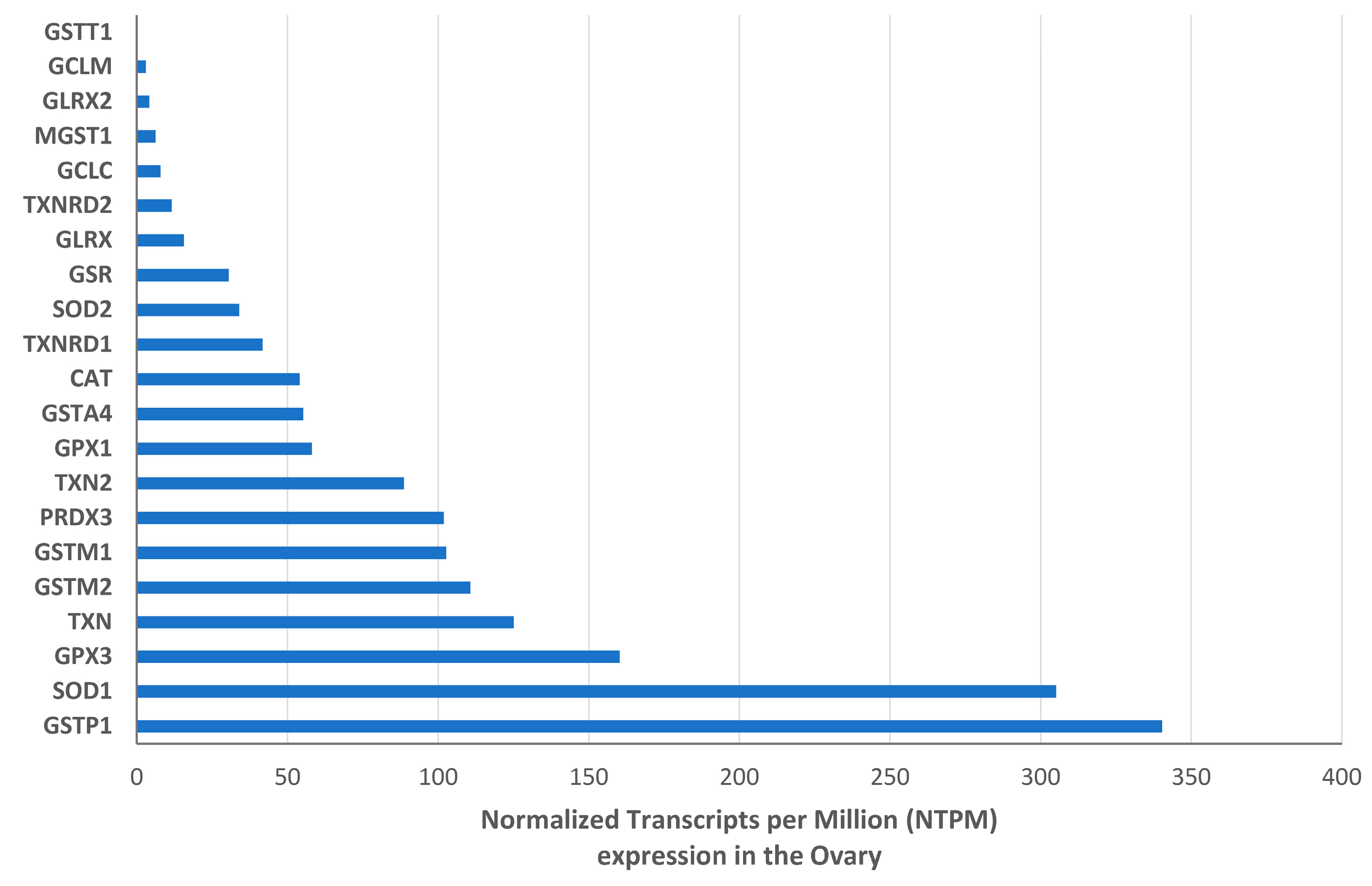
Figure 3.
Network showing the interactions between miRNAs and antioxidant enzyme mRNAs.
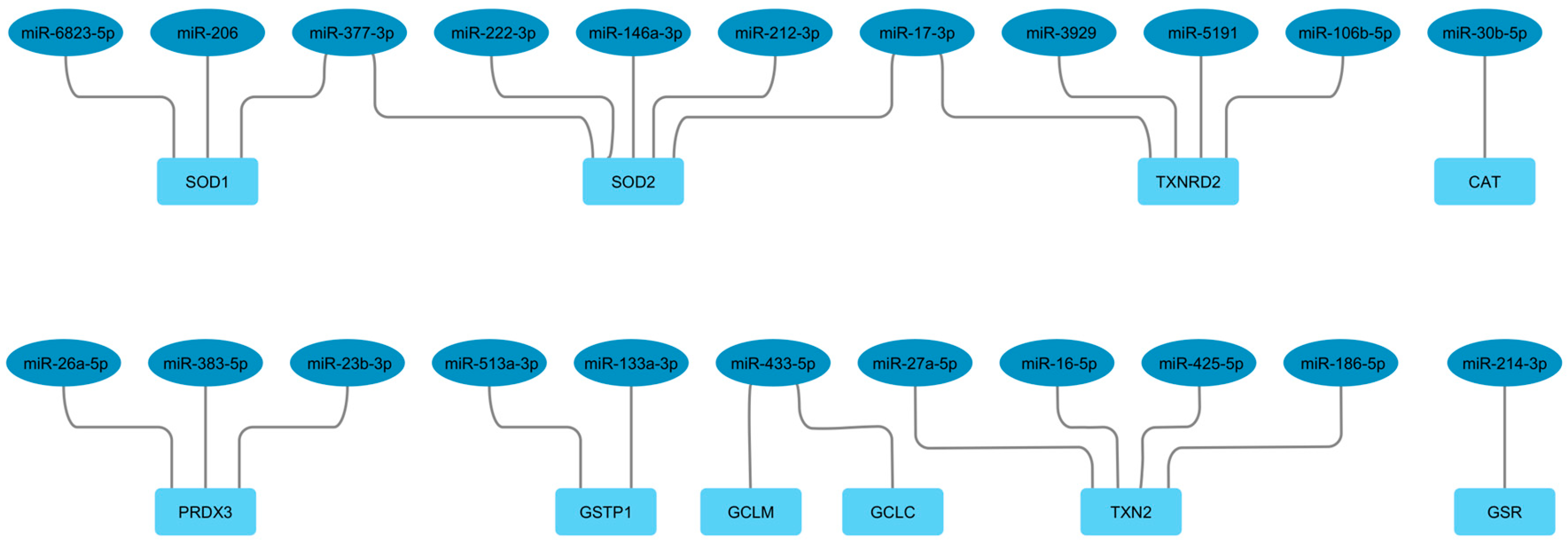
Figure 4.
Network showing the interaction between the miRNAs targeting antioxidant enzymes and lncRNAs. The nodes are ranked according to the degree scoring method, with a color scheme from highly central (red color) to central (yellow).
Figure 4.
Network showing the interaction between the miRNAs targeting antioxidant enzymes and lncRNAs. The nodes are ranked according to the degree scoring method, with a color scheme from highly central (red color) to central (yellow).
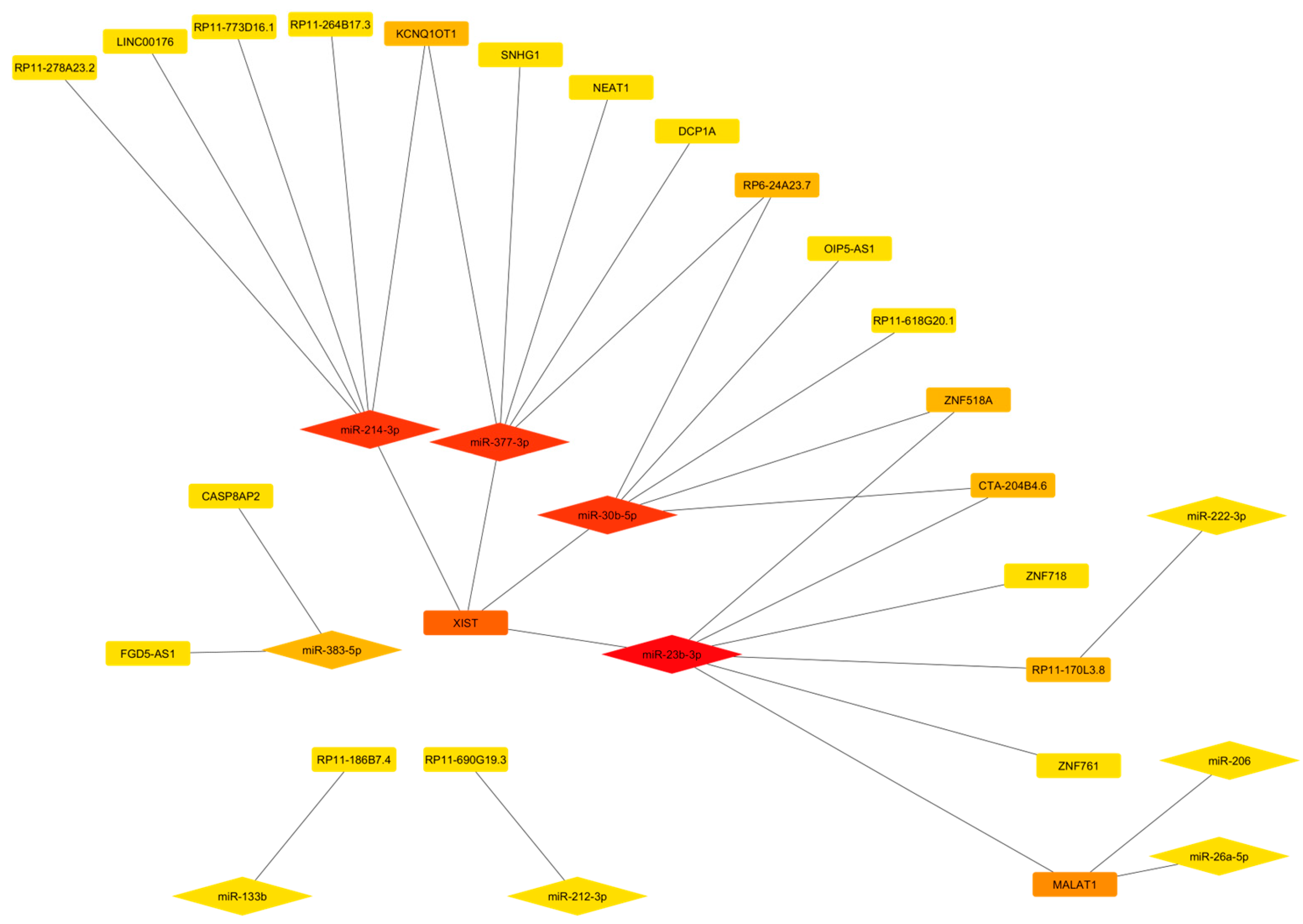
Figure 5.
mRNA-miRNA-lncRNA ceRNA-network. A) Antioxidant ceRNA network B) Ovarian antioxidant ceRNA-network, OvAnOx ceRNA network. miRNAs are red-colored, mRNAs are yellow-colored and lncRNAs are blue-colored.
Figure 5.
mRNA-miRNA-lncRNA ceRNA-network. A) Antioxidant ceRNA network B) Ovarian antioxidant ceRNA-network, OvAnOx ceRNA network. miRNAs are red-colored, mRNAs are yellow-colored and lncRNAs are blue-colored.
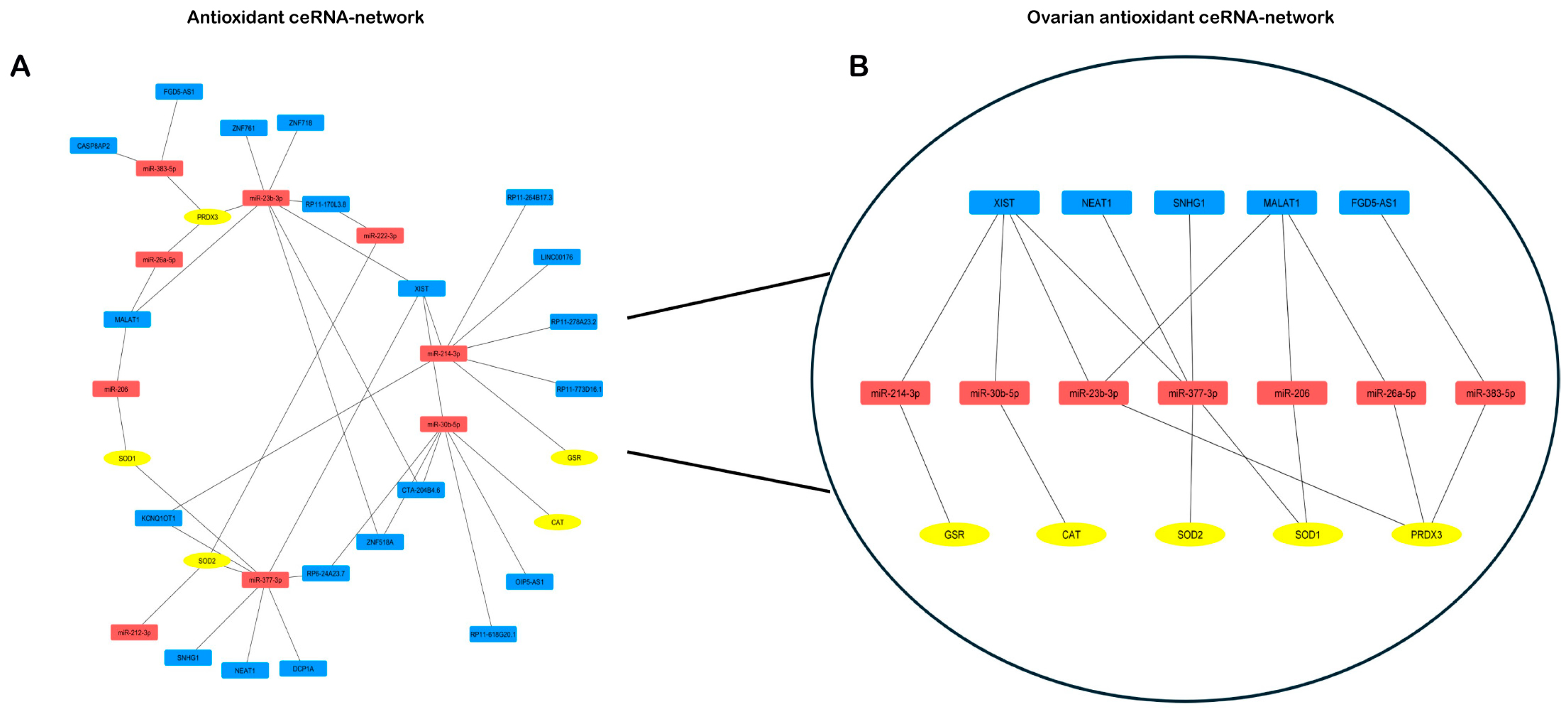

Table 1.
Localization of the 21 antioxidant enzymes.
| Cellular Localization | Extracellular Localization | ||
|---|---|---|---|
| Gene Name | Cytoplasm | Mitochondria | Exosomes |
| CAT | |||
| GCLC | |||
| GCLM | |||
| GLRX | |||
| GLRX2 | |||
| GPX1 | |||
| GPX3 | |||
| GSR | |||
| GSTA4 | |||
| GSTM1 | |||
| GSTM2 | |||
| GSTP1 | |||
| GSTT1 | |||
| MGST1 | |||
| PRDX3 | |||
| SOD1 | |||
| SOD2 | |||
| TXN | |||
| TXN2 | |||
| TXNRD1 | |||
| TXNRD2 | |||
Abbreviations: CAT: Catalase; GCLC: Glutamate-Cysteine Ligase Catalytic Subunit; GCLM: Glutamate-Cysteine Ligase Modifier Subunit; GLRX: Glutaredoxin; GLRX2: Glutaredoxin 2; GPX1: Glutathione Peroxidase 1; GPX3: Glutathione Peroxidase 3; GSR: Glutathione Reductase; GSTA4: Glutathione S-Transferase Alpha 4; GSTM1: Glutathione S-Transferase Mu 1; GSTM2: Glutathione S-Transferase Mu 2; GSTP1: Glutathione S-Transferase Pi 1; GSTT1: Glutathione S-Transferase Theta 1; MGST1: Microsomal Glutathione S-Transferase 1; PRDX3: Peroxiredoxin 3; SOD1: Superoxide Dismutase 1; SOD2: Superoxide Dismutase 2; TXN: Thioredoxin; TXN2: Thioredoxin 2; TXNRD1: Thioredoxin Reductase 1; TXNRD2: Thioredoxin Reductase 2.
Table 2.
Ovarian Localization of the 21 antioxidant enzymes.
| Gene Name | FF | O | CC | GC | TC | LC | SC | ND |
|---|---|---|---|---|---|---|---|---|
| CAT | ||||||||
| GCLC | ||||||||
| GCLM | ||||||||
| GLRX | ||||||||
| GLRX2 | ||||||||
| GPX1 | ||||||||
| GPX3 | ||||||||
| GSR | ||||||||
| GSTA4 | ||||||||
| GSTM1 | ||||||||
| GSTM2 | ||||||||
| GSTP1 | ||||||||
| GSTT1 | ||||||||
| MGST1 | ||||||||
| PRDX3 | ||||||||
| SOD1 | ||||||||
| SOD2 | ||||||||
| TXN | ||||||||
| TXN2 | ||||||||
| TXNRD1 | ||||||||
| TXNRD2 |
Follicular Fluid (FF); Oocyte (O); Cumulus Cells (CC); Granulosa Cells (GC); Theca Cells (TC); Luteal Cells (LC); Stromal Cells (SC); Not Determined (ND).
Table 3.
miRNAs regulating the 21 antioxidant enzyme mRNAs.
| CAT | GCLC | GCLM | GSR | GSTP1 | PRDX3 | SOD1 | SOD2 | TXN2 | TXNRD2 | |
|---|---|---|---|---|---|---|---|---|---|---|
| miR-16-5p | ||||||||||
| miR-17-3p | ||||||||||
| miR-23b-3p | ||||||||||
| miR-26a-5p | ||||||||||
| miR-27a-5p | ||||||||||
| miR-30b-5p | ||||||||||
| miR-106b | ||||||||||
| miR-133a | ||||||||||
| mir-146a | ||||||||||
| miR-186-5p | ||||||||||
| miR-206 | ||||||||||
| miR-212-3p | ||||||||||
| miR-214-3p | ||||||||||
| miR-222-3p | ||||||||||
| miR-377-3p | ||||||||||
| miR-383-3p | ||||||||||
| miR-425-5p | ||||||||||
| miR-433-5p | ||||||||||
| miR-513a-3p | ||||||||||
| miR-3929 | ||||||||||
| miR-5191 | ||||||||||
| miR-6823-5p |
Table 4.
miRNAs regulating 21 antioxidant enzyme mRNAs are sponged by lncRNAs.
| miR-23b-3p | miR-26a-5p | miR-30b-5p | miR-133b | miR-206 | miR-212-3p | miR-214-3p | miR-222-3p | miR-377-3p | miR-383-5p | |
| CASP8AP2 | ||||||||||
| CTA-204B4.6 | ||||||||||
| DCP1A | ||||||||||
| FGD5-AS1 | ||||||||||
| KCNQ1OT1 | ||||||||||
| LINC00176 | ||||||||||
| MALAT1 | ||||||||||
| NEAT1 | ||||||||||
| OIP5-AS1 | ||||||||||
| RP11-170L3.8 | ||||||||||
| RP11-186B7.4 | ||||||||||
| RP11-264B17.3 | ||||||||||
| RP11-278A23.2 | ||||||||||
| RP11-618G20.1 | ||||||||||
| RP11-690G19.3 | ||||||||||
| RP11-773D16.1 | ||||||||||
| RP6-24A23.7 | ||||||||||
| SNHG1 | ||||||||||
| XIST | ||||||||||
| ZNF518A | ||||||||||
| ZNF718 | ||||||||||
| ZNF761 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



