
Submitted:
10 August 2024
Posted:
13 August 2024
You are already at the latest version
Abstract
This review provides a comprehensive analysis of the pelvic plexus and its regulation across various mammalian species, including rats, cats, dogs, and pigs. The pelvic and hypogastric nerves play crucial roles in regulating pelvic functions such as micturition, defecation, and erection. The anatomical organization of these nerves varies, forming either well-defined ganglia or complex plexuses. Despite these variations, the neurons within these structures are consistently regulated by key neurotransmitters—norepinephrine and acetylcholine. These neurons also possess receptors for testosterone and prolactin, particularly in rats, indicating the significant role of these hormones in neuronal function and development. Moreover, neuropeptides such as vasoactive intestinal peptide (VIP), substance P, neuropeptide Y (NPY), somatostatin (SOM), galanin (GAL), and calcitonin gene-related peptide (CGRP) are co-released with neurotransmitters to modulate pelvic functions. This review highlights the complex interplay between neurotransmitters, neuropeptides, and hormones in regulating pelvic physiology and emphasizes the importance of hormonal regulation in maintaining the functionality and health of the pelvic plexus across different species.
Keywords:
major pelvic ganglion
; pelvic area
; parasympathetic and sympathetic innervation
1. Introduction
The major pelvic ganglion (MPG) is a key structure within the pelvic plexus, a complex network of nerve fibers and neuronal bodies strategically located among the pelvic organs. This ganglion plays a critical role in regulating various physiological functions, including urination, defecation, erection, ejaculation, and glandular secretion, by transmitting nerve signals from the central nervous system to the pelvic organs, such as the urinary tract, lower intestine, and reproductive organs [1,2,3].
In male rats, the MPG constitutes the principal component of the pelvic plexus, complemented by accessory ganglia also known as peripheral ganglia. The nomenclature and function of these homologous structures may vary among different mammalian species. For instance, in females, it is referred to as the paracervical ganglion or Frankenhauser's ganglion, while in rabbits and guinea pigs, the term "pelvic plexus" is used due to its anatomical complexity. In cats, this structure is known as the "inferior hypogastric plexus," and in dogs, it is termed the "periprostatic plexus," reflecting anatomical and functional differences between species [2,3,4,5].
Therefore, this review aims to compare the anatomical, histological, and functional characteristics of the major pelvic ganglion across various mammalian species, including dogs, guinea pigs, cats, pigs, and male rats. Aspects such as anatomy, nerve connections, histology, neurochemistry, and physiology will be addressed to enhance the understanding of similarities and differences in pelvic function regulation among these species.
General Overview and Anatomical Organization
The MPG, also known as the inferior hypogastric ganglion, is a crucial structure in the autonomic nervous system of mammals. It is part of the pelvic plexus and plays a vital role in regulating the visceral functions of the pelvic region, including the bladder, rectum, and reproductive organs. It is located in the lower part of the pelvic cavity, adjacent to the hypogastric arteries (also known as internal iliac arteries) and near the rectum and bladder. It is bilaterally symmetrical, attached to the serous cavity of the dorsal lobe of the prostate, and connected by ipsilateral and contralateral commissural nerves. The ganglion measures approximately 2x4 mm (Figure 1), has a pyramidal shape, and exhibits characteristics of sexual dimorphism [6,7,8,9,10,11,12,13,14].
The pelvic plexus in male rats comprises three pairs of accessory ganglia (AG) and one pair of major pelvic ganglia (MPG). The AG, also known as peripheral ganglia or hypogastric ganglia, are located near the base of the deferent ducts. Historically, they have been considered identical to the MPG, although with lesser innervation to certain reproductive organs. These ganglia are situated close to the midline and communicate with the larger ganglia located laterally on the opposite side(Figure 1A), allowing preganglionic and postganglionic axons to innervate contralateral neural structures [13].
The pelvic plexus varies among species. For example, in guinea pigs, there are an anterior major pelvic ganglion (AMPG), located at the entrance of the deferent ducts into the bladder, which resembles the MPG of the male rat, and a posterior major pelvic ganglion (PMPG). These ganglia are responsible for innervating the urinary bladder, the distal portion of the urethra, the deferent ducts, the seminal vesicle, a portion of the prostate, the penis, and the rectum [15,16,17,18,19,20,21]. In pigs, the pelvic plexus is composed of a network of filaments and several small ganglia surrounded by connective tissue, located in the medial part of the iliac vessels and extending bilaterally to the lower part of the peritoneum and the lobulated external surface of the seminal vesicles [22]. They also have an AMPG located between the proximal end of the deferent duct and the caudal part of the seminal vesicle, in addition to the PMPG. In contrast, cats have a more complex pelvic plexus, where the pelvic nerves converge and intertwine with the ganglionic plexus at the base of the bladder, forming fine filaments that extend to the lateral external surface of the detrusor muscle. This structure comprises between 40 and 60 ganglia, each containing approximately 10 to 20 neurons with extensive dendritic branching [1,23,24]. In dogs, the pelvic plexus (PP) is located 5 to 10 mm lateral to the prostate, just beneath the parietal pelvic peritoneum and ventrolateral to the rectum. It typically appears as a well-defined, rhomboid, and flat structure, with dimensions ranging from 5 to 15 mm [5].
Although there are notable differences in the anatomy of pelvic ganglia among laboratory animals, the physiological importance, advantages, and disadvantages of neuronal aggregation in these ganglia, as opposed to dispersion in a large plexus, have not yet been reported. This underscores the importance of understanding anatomical differences among species for clinical and research applications.
Regarding its organization, in most mammals, this ganglion consists of sympathetic and parasympathetic neurons that synthesize adrenaline or acetylcholine, respectively. The acetylcholine-synthesizing neurons are divided into two types, differing in size and staining intensity. The most prevalent cell type in the ganglion ranges from 30 to 45 micrometers in size and shows weak to moderate immunostaining. The other cell type measures between 15 and 25 micrometers and exhibits very intense immunostaining. These neurons are located at the elongated end of the ganglion, mainly grouped at the dorsal end of the pelvic ganglion and around the pelvic nerve as it enters the major pelvic ganglion [25]. Additionally, the ganglion contains a large population of small, intensely fluorescent cells (SIF) with coarse granules in their cytoplasm, and many ganglion cells are closely juxtaposed by SIF cells [25]. In rats, two types of SIF cells have been identified based on the size and morphology of their granules. The most prevalent SIF cells contain numerous vesicles with diameters ranging from 1500 to 2700 Å. The second type of SIF cells is characterized by small vesicles, between 500 and 1200 Å, located peripherally. These cells can receive afferent innervation from a ganglion cell but also innervate ganglion cells, possibly making contact with satellite cells responsible for generating a regulatory microenvironment. It has been reported that in this type of efferent innervation, the nerve fibers contain numerous smaller clear vesicles compared to those observed in SIF cells. Furthermore, SIF cells may appear singly or in groups of varying sizes, dispersed among both cholinergic and adrenergic ganglion neurons. Although much is still unknown about the function of these cells in the ganglion, it is suggested that they contain norepinephrine, without ruling out the possibility of synthesizing other neurotransmitters such as catecholamines. A notable feature of these cells is their proximity to blood vessels, which allows them to be hormonally modulated [25,26].
Within the ganglion, as in all autonomic ganglia, there are also satellite cells characterized by relatively simple morphology. These cells typically make contact with a neuron, are flat, and contain little cytoplasm. As they encase the ganglion cell, it is proposed that they protect the neuron from exposure to molecules from the circulation, modulating neuronal function by allowing the passage of some proteins and macrophages, although this is not yet fully understood. Structurally, it has been observed that the surface in contact with the connective tissue is flatter compared to the internal surface, increasing the contact area with the neuron [27]. The neuron-satellite cell unit is commonly observed, but there may be clusters where a single satellite cell surrounds two or three neurons [28]. It is also possible for a neuron to be surrounded by several satellite cells, depending on the neuron’s size. For example, in geckos, the ratio is 3.8:1, in mice it is 5.5:1, in rats and mice it is 8.2:1, and in rabbits it is 10.2:1 [29]. It characteristic of satellite cells is that they form a functional syncytium because they are interconnected by gap junctions assembled by the Cx43 protein (Procacci et al., 2008), and they have immune properties as they express proteins such as CD40, 54, 14, and 68 [30,31,32].
The other cell type present in the ganglion is Schwann cells, the most abundant glial cells in the peripheral nervous system. Their primary function is to cover the axon with myelin sheaths composed of 70-85% lipids and 15-30% proteins, increasing the conduction velocity of nerve impulses through saltatory conduction. These cells influence axon structure and diameter and also protect the axon from mechanical damage. Additionally, they can modulate synaptic activity and play a key role in synaptic competition [33,34,35,36].
In summary, this ganglion shares characteristics with other autonomic ganglia in terms of cell types. However, its distinction lies in being a mixed ganglion, as it contains both sympathetic and parasympathetic fibers from the hypogastric and pelvic nerves (Figure 2).
Preganglionic and Postganglionic Connections
The MPG is an autonomic nerve ganglion located adjacent to the dorsolateral lobe of the prostate and is responsible for innervating the entire pelvic floor. Postganglionic fibers emerging from this ganglion regulate functions such as urination, defecation, erection, and secretion of the accessory reproductive glands [37,38]. However, its function is also regulated by preganglionic fibers originating from the hypogastric nerve (NHg), typically classified as sympathetic, and the pelvic nerve (NPv), which is parasympathetic. This dual innervation classifies the ganglion as mixed, as it contains both sympathetic and parasympathetic fibers [38,39,40].
Sympathetic fibers originate from the intermediolateral cell column of thoracolumbar segments T10-L2. The axons of these neurons form the lumbar splanchnic nerves (LSN), which project to the inferior mesenteric ganglion (IMG). From the caudal area of the IMG, the NHg descends, joining the inferior hypogastric plexus, travels behind the ureter, and branches into hypogastric accessory nerves and the main NHg, which terminates in the rostral region of the major pelvic ganglion [7,41,42,43]. In rats, preganglionic fibers composing this nerve originate from 81% of neurons located in the dorsal gray commissure of the L1-L2 spinal segments, with diameters between 24 and 21 µm, respectively [44]. This nerve comprises 223 unmyelinated fibers and 1,363 myelinated fibers, constituting 85% of the total fibers present in the nerve. Few afferent or sensory fibers are present, as 96% of these fibers remain in the nerve following dorsal root ganglectomy and 65% remain after ventral rhizotomy. In cats, these neurons are located in segments L2-L5, predominantly in segments L3-L5 in the intermediolateral horn, where preganglionic sympathetic neurons are found. The number of neurons composing this nerve is approximately 20,000, with diameters of 300-400 µm, closely approximating the 21,000 fibers in the composite nerve, which includes 1,300 afferent fibers (6.5%), 1,700 preganglionic fibers (8.5%), and the remainder as postganglionic fibers (85%) [45]. Compared to the fibers in rats, this represents a 13-fold increase [46]. Thus, nerves emerging from the major pelvic ganglion and directed toward the ventral lobe originate from the ventral area, while those innervating the dorsolateral lobe originate from the rostral-dorsal area of the major pelvic ganglion [3,47].
In rats, preganglionic neurons originating from the pelvic nerve are located in the intermediolateral gray substance of the L6-S1 spinal segments [48,49]. Approximately 550 neurons, measuring 10 x 20 µm, were in the intermediolateral gray with dendrites extending toward the lateral follicle, and an average of 1,500 sensory neurons were also reported [50]. In monkeys, an average of 820 preganglionic neurons are found in the sacral parasympathetic nucleus in the intermediate gray of S1-S3 segments, a pattern similar to that in cats [51]. Regardless of the spinal segments involved, it is evident that the sacral parasympathetic system exhibits a similar organization across various mammalian species [49,52]. In rats, nerve fibers emerge from the ventral horn and branch into the somatomotor branch, innervating the iliococcygeus and pubococcygeus muscles, and the viscerocutaneous branch, which enters the dorsal region of the major pelvic ganglion [48]. It is important to note that information exiting these nerve terminals is regulated by interneurons in contact with the neurons forming this nerve. Thus, preganglionic neurons receive excitatory glutamatergic inputs from two distinct populations of interneurons located in the dorsal and medial regions of the sacral parasympathetic nucleus [53,54]. This partially explains why this nerve, along with the hypogastric nerve, can regulate various functions of the pelvic area.
As previously mentioned, the major pelvic ganglion regulates different pelvic functions through postganglionic nerve fibers emerging from it. In dogs, a prominent efferent branch emerges from the caudal end of the plexus and extends toward the prostatic apex as the main cavernous nerve (CN). After traveling 15 to 20 mm, it branches to send a small branch to the rectum, while the main trunk continues to the prostate apex and along the urethra to the pelvic floor. This main CN measures up to 5 mm in width and 40 to 60 mm in length. In some cases, an additional efferent nerve emerges from the medial edge of the plexus and extends toward the middle prostate, branching widely into the periprostatic fascia and extending to the posterolateral and lateral parts of the prostate, eventually connecting with the main CN [5]. Additionally, this nerve also innervates the bladder, urethra, and ureters along with the hypogastric ganglion [11,55,56,57].
In pigs, the hypogastric nerve, derived from the caudal mesenteric ganglion, descends from the posterior pole of this ganglion on both sides and, as it progresses, runs parallel to the lumbar column before entering the pelvic cavity and joining the pelvic plexus. Unlike in humans, in pigs, this nerve rarely forms an elongated plexus but appears as long branches that anastomose without forming a compact structure, as observed in rats. This nerve has a diameter ranging from 22 to 320 micrometers and consists of four to ten nerve bundles with few myelinated fibers, measuring 1 to 2-3 micrometers in diameter, distributed irregularly or forming clusters among unmyelinated fibers. On the other hand, pelvic nerves in pigs are thinner than the hypogastric nerve and originate from branches emerging from the S1 segment immediately after exiting the sacral foramen [22]. The pelvic nerve in pigs has a single branch composed of 6-12 nerve bundles, each with a diameter ranging from 30 to 360 micrometers, wrapped in loose connective tissue. These nerves contain myelinated fibers, with a few measuring between 6 and 7 micrometers in diameter, but predominantly fibers ranging from 1.7 to 27 micrometers in diameter [22]. Of the nerves emerging from the pelvic plexus and targeting the pelvic organs in pigs, the pudendal nerve descends along the external surface of the ligament to re-enter the pelvis through the lesser sciatic foramen, receiving a branch from the sciatic nerve upon entry, usually just after exiting the lesser sciatic foramen. This nerve sends branches to the border between the rectum and the bulbourethral gland, one of which extends forward to innervate the bulbourethral gland and the urethra. This branch divides into thinner branches, accompanied by blood vessels, penetrating the urethral muscular wall [22]. The myelinated nerve fibers originating from the pudendal nerve form nerve bundles approximately 150-180 micrometers in diameter. Within these bundles, nerve fibers exhibit diameters ranging from 5 to 5.5 micrometers, with some between 8.5 and 13.5 micrometers [22,58,59,60,61]. In guinea pigs, there are two mesenteric ganglia, anterior and posterior to the inferior mesenteric artery, approximately 3 mm from the aorta. These ganglia are connected by fine nerves, which in turn receive nerve fibers from the third, fourth, and fifth lumbar ganglia of the ganglionic chain [15], directing fibers toward the inferior mesenteric ganglion from which the colonic and hypogastric nerves emerge. The colonic nerves consist of a dozen strands accompanying the inferior mesenteric artery toward the colon, eventually innervating the rectum. There are two main hypogastric nerves and one accessory hypogastric nerve emerging from the inferior mesenteric ganglia. The main hypogastric nerves extend left and right to terminate in the deferent ducts and seminal vesicles [15]. These ganglia, also known as anterior pelvic ganglia, receive connections from pelvic nerves and send fibers to the deferent ducts, seminal vesicles, bladder, prostate, and ureters in males. Other dispersed cell groups along the main hypogastric nerves are considered additional hypogastric ganglia. The posterior pelvic nerves, located deeper in the pelvic cavity, form a plexus connected to the anterior ganglia, contributing to the innervation of organs such as the rectum and urethra. These nerves, primarily originating from the second, third, and fourth sacral nerves, also give rise to branches innervating the perineal and anal areas. Additionally, there are two pudendal nerves, which provide innervation to structures such as the urethra, penis, anal sphincter, and perineal skin [15]. Notably, neuronal activity from the urethra, rectum, and genitals ascends to the spinal cord via the pelvic nerve and is subsequently transmitted through the spinal cord to the cerebral cortex in both dogs and cats [11,62]. Furthermore, it has been reported that seminal emission control has two sympathetic pathways; one originating from the hypogastric nerve and the other from the lumbosacral sympathetic trunks, which appears to function as a compensatory system for seminal emission in the event of hypogastric nerve damage [63,64,65]. In the guinea pig pelvic ganglion, the enzymes tyrosine hydroxylase (TH) and dopamine β-hydroxylase (DBH) are present, along with the enzyme for acetylcholine synthesis, choline acetyltransferase (ChAT). Additionally, two main types of neurons have been reported: a) those containing NOS located in the caudal part of the major pelvic ganglion (APG) and b) TH-positive neurons lacking NOS located in the cranial part [66].
Neurotransmitters and Receptors in the Pelvic Plexus
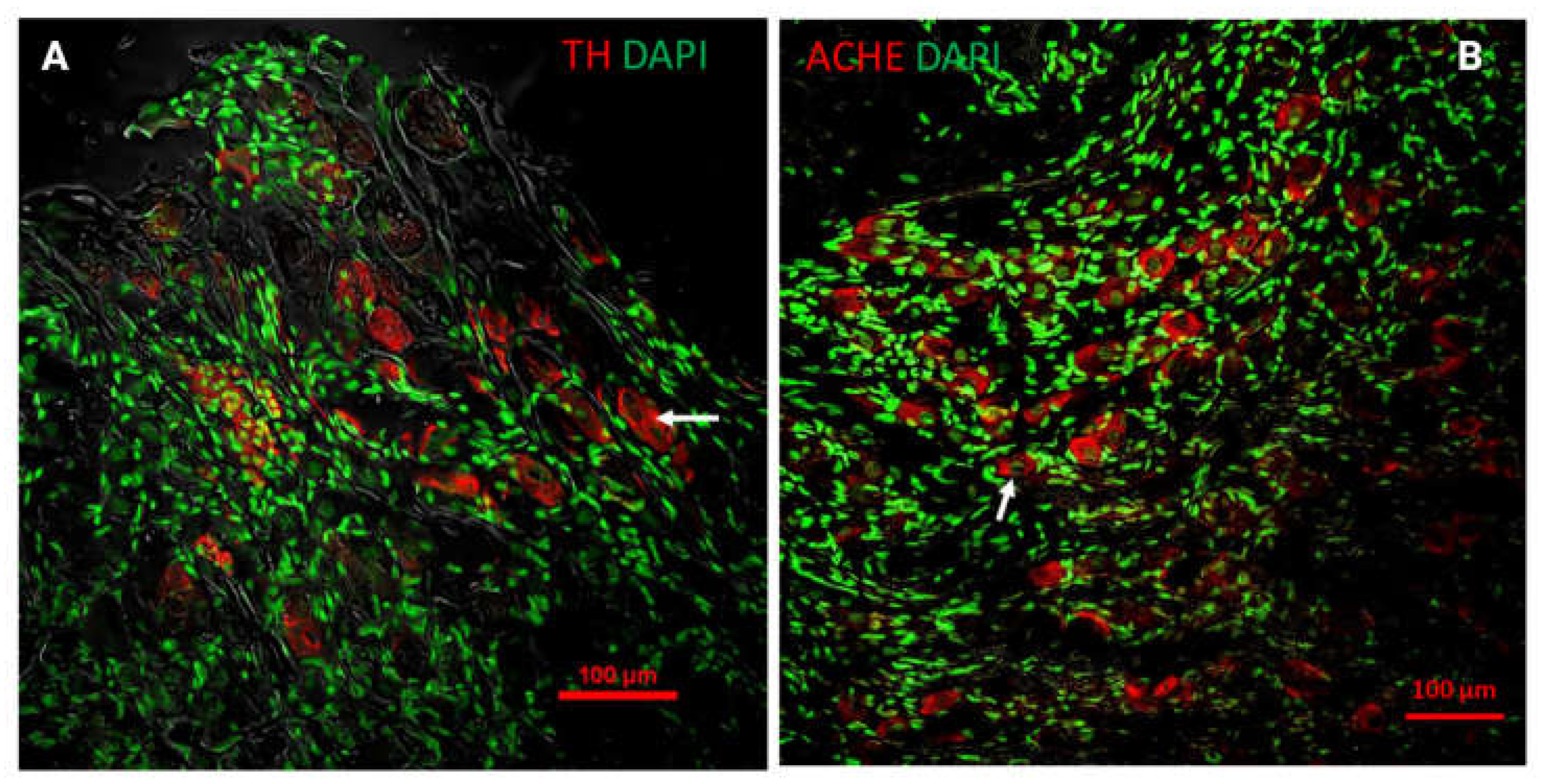
A notable feature of the MPG, at least in rats, is its mixed nature; it contains both sympathetic and parasympathetic fibers, with neurons synthesizing either norepinephrine (adrenergic neurons) or acetylcholine (cholinergic neurons) (Figure 3A,B). It has been described that all neurons in the MPG are either cholinergic or adrenergic, and they do not synthesize both neurotransmitters, with fewer than 5% of neurons lacking either neurotransmitter [10,40].
These neurons exhibit distinct topography within the MPG. Adrenergic neurons, with areas ranging from 600 to 1400 μm², are located near the entry point of the hypogastric nerve (NHg). Only a few are found where the NHg integrates with the pelvic nerve (NPv), and none are present where the cavernous nerve originates [10,25,67,68]. Sympathetic neurons have been reported to express neuropeptide Y (NPY), which is involved in norepinephrine-mediated vasoconstriction of pelvic organs [69].
In contrast, cholinergic neurons range from 300 to 700 μm². Dail, Evan, & Eason (1975) reported that there is a cluster of small cholinergic neurons that show intense acetylcholine staining and are located near the NPv, while another population of small neurons with weak acetylcholine staining is found near small branches from the hypogastric nerve [3,70] (see Figure 3B). Additionally, cholinergic neurons co-express neuropeptides such as somatostatin (SOM), cholecystokinin (CCK), galanin (GAL), calcitonin gene-related peptide (CGRP), neuropeptide Y (NPY), and vasoactive intestinal peptide (VIP), and also express the cocaine- and amphetamine-regulated transcript (CART) peptide [3,23,70]. It is important to note that afferent fibers innervating the colon are polymodal, responding to temperature and chemical changes [71].
The relative distribution of sympathetic and parasympathetic neurons in the pelvic ganglia varies across species, gender, and strain, highlighting interspecies variations in the neurochemical regulation of pelvic organ function, which may have significant implications for physiology and pathology in these animals.
In the guinea pig's pelvic ganglion, particularly in the caudal region near the rectal and penile nerves, primarily NOS-immunoreactive neurons are found, and almost all neurons contain choline acetyltransferase (ChAT), indicating cholinergic neurons that are also immunoreactive to neuropeptides such as VIP, NPY, SOM, or dopamine β-hydroxylase (DBH). Conversely, in the cranial region, neurons are abundant in TH-immunoreactivity, suggesting adrenergic neurons, with almost all co-expressing NPY [18]. In the pig's pelvic ganglion, 40% of the neurons show immunoreactivity to acetylcholine (ChAT) or the vesicular acetylcholine transporter (VAChT), and 39% also exhibit immunoreactivity to NOS, VIP, NPY, or SOM, suggesting co-expression of neurotransmitters and peptides, highlighting the functional and neurochemical diversity of neurons in the pelvic ganglion [17,72,73]. In dogs, it has been reported that penile erection and bladder control mediated by the pelvic nerve are likely influenced by vasoactive intestinal peptide [62,65,74,75].
Hormonal Regulation in the Pelvic Ganglia
The anatomical and functional organization of the major pelvic ganglion (MPG) and its neurotransmitters and neurohormones that regulate various pelvic functions across different animal species are well understood. Given that reproduction is one of these functions, it is reasonable to investigate whether the development of this ganglion is subject to hormonal regulation, particularly by steroid hormones, with androgens being of primary interest.
Steroid hormones play a crucial role in the development and function of the autonomic nervous system, including the MPG, which is present in various mammalian species. The MPG is a key structure in the innervation of pelvic organs and is influenced by several steroid hormones, especially sex hormones such as androgens and estrogens. In rats, for example, it has been shown that the MPG is sensitive to androgens. Castration performed at twelve hours postnatally affects the activity of enzymes responsible for synthesizing acetylcholine and norepinephrine, an effect that is reversed by administering testosterone within ten days post-castration. Estrogens can also reverse the effects of castration on acetylcholine transferase activity but do not affect tyrosine hydroxylase activity. This study indicates a critical postnatal period dependent on the presence of androgens [76,77,78,79].
Furthermore, it is known that neurons in the MPG are affected by the absence of androgens, as maintained castration for two to seven weeks results in adrenergic neurons reaching only about 60% of their normal size, without affecting cholinergic neurons. This suggests that adrenergic neurons are more sensitive to androgen absence and that ganglion development depends on the presence of this androgen [80]. In vitro studies have revealed that both androgens and estrogens, derived from testosterone due to the presence of aromatase, can stimulate the growth of more complex neurites in both adrenergic and cholinergic neurons expressing nitric oxide synthase (NOS) [81,82]. It is noteworthy that approximately 40% of neurons innervating the penile corpus cavernosum contain androgen receptors; however, only 20% of neurons expressing nitric oxide and 40% of those expressing vasoactive intestinal peptide (VIP) contain androgen receptors [81]. Although the precise mechanisms are not fully understood, it is known that testosterone modifies the membrane potential of these ganglionic neurons by inducing significant depolarizations relative to their resting potential [83], reinforcing the notion that androgen regulation affects the electrophysiological properties of these neurons [84,85].
Considering that the MPG controls reproductive functions and is sensitive to testosterone, it has also been shown that sexual behavior impacts not only the prostate, causing plastic changes in the gland [86], but also affects the ganglion. Experimental conditions reveal that the ganglion area is larger compared to sexually naïve individuals, likely due to an increase in neuronal area from 0-300 μm² to 600-900 μm². However, when preganglionic innervation is interrupted, these effects are not easily reversed; although there is a slight reduction in neuronal area and size, the effects are not as drastic as in sexually inexperienced animals. This plasticity observed in the MPG appears to be a consequence of elevated blood levels of testosterone and prolactin, both of which have receptors in this ganglion. However, the effects of testosterone and prolactin do not seem to require increased receptor expression, as receptor levels remain basal under these conditions. This suggests that basal receptor expression is sufficient to mediate behavioral effects, and these effects are not due to changes in α1-adrenergic and M3 muscarinic receptors analyzed in the ganglion [38,87]. Conversely, the absence of preganglionic neural control or axotomy increases the expression of muscarinic and androgen receptors, implying that these receptors are downregulated under physiological conditions and play a role in better control of reproductive functions under these conditions [87].
Few studies have investigated the effects of castration on the development or consequences of testosterone absence on the MPG in other species, such as cats, dogs, and pigs. Studies in pigs indicate that androgens are also important for neuron survival. Castration significantly affects the survival of pelvic ganglion neurons, with a loss of 74% and 90% after 3 and 6 months, respectively. These conditions also show an increase in caspase 3 expression, suggesting that neurons die via androgen-induced apoptosis [17]. Thus, the decrease or increase in certain neurotransmitters or neuropeptides affected by castration may also be a result of testosterone deprivation, enhancing neuronal responses without significant effects when the hormone is reintroduced, at least in boars [72].
2. Conclusions
As observed throughout this review, pelvic functions in various species, including rats, cats, dogs, and pigs, are regulated by the pelvic and hypogastric nerves. The anatomical arrangement may vary slightly, forming either a well-developed ganglion or a plexus. Regardless of the specific structure, the neurons within these systems are regulated and contain two classic neurotransmitters: norepinephrine and acetylcholine. These neurons possess receptors for both neurotransmitters, as well as receptors for testosterone and prolactin, with the latter notably in rats. The presence of testosterone is crucial for the proper functioning of these neurons. Additionally, it is noteworthy that neurons in different species also contain neuropeptides such as VIP, substance P, NPY, SOM, GAL, and CGRP. These neuropeptides can be co-released with neurotransmitters to regulate various pelvic functions, including micturition, defecation, erection, and the control of accessory glands related to reproduction.
Acknowledgments
This study was funded by Conahcyt Fellowship 1036378 to J.N.L.G., 1101339 M.P.P.A. and by the Neuroscience (UV-CA-28) and Neurochemical (UV-CA-304) Groups of the Brain Research Institute.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bradley WE, Teague CT. The Pelvic Ganglia. J Urol. 1968;100(5):649-652. [CrossRef]
- Keast JR. Plasticity of Pelvic Autonomic Ganglia and Urogenital Innervation. Int Rev Cytol. 2006;248:141-208. [CrossRef]
- Keast JR. Visualization and immunohistochemical characterization of sympathetic and parasympathetic neurons in the male rat major pelvic ganglion. Neuroscience. 1995;66(3):655-662. [CrossRef]
- Crowcroft PJ, Szurszewski JH. A study of the inferior mesenteric and pelvic ganglia of guinea-pigs with intracellular electrodes. J Physiol. 1971;219(2):421-441. [CrossRef]
- Gianduzzo TRJ, Colombo JR, El-Gabry E, Haber GP, Gill IS. Anatomical and Electrophysiological Assessment of the Canine Periprostatic Neurovascular Anatomy: Perspectives as a Nerve Sparing Radical Prostatectomy Model. J Urol. 2008;179(5):2025-2029. [CrossRef]
- Paxinos G. The Rat Nervous System. The Rat Nervous System. Published online May 4, 2004:1-1309. [CrossRef]
- Dowling P, Ranson RN, Santer RM. Age-associated changes in distribution of the P2X2 receptor in the major pelvic ganglion of the male rat. Neurosci Lett. 2006;404(3):320-323. [CrossRef]
- Keast JR, de Groat WC. Immunohistochemical characterization of pelvic neurons which project to the bladder, colon, or penis in rats. Journal of Comparative Neurology. 1989;288(3):387-400. [CrossRef]
- Keast JR, Booth AM, de Groat WC. Distribution of neurons in the major pelvic ganglion of the rat which supply the bladder, colon or penis. Cell Tissue Res. 1989;256(1):105-112. [CrossRef]
- Wanigasekara Y, Kepper ME, Keast JR. Immunohistochemical characterisation of pelvic autonomic ganglia in male mice. Cell Tissue Res. 2003;311(2):175-185. [CrossRef]
- Purinton PT, Oliver JE. Spinal cord origin of innervation to the bladder and urethra of the dog. Exp Neurol. 1979;65(2):422-434. [CrossRef]
- Langworthy OR. Commissures of autonomic fibers in the pelvic organs of rats. Anat Rec. 1965;151(4):583-587. [CrossRef]
- Arellano J, Xelhuantzi N, Mirto N, Hernández ME, Cruz Y. Neural interrelationships of autonomic ganglia from the pelvic region of male rats. Auton Neurosci. 2019;217:26-34. [CrossRef]
- Dail WG, Harji F, Gonzales J, Galindo R. Multiple vasodilator pathways from the pelvic plexus to the penis of the rat. International Journal of Impotence Research 1999 11:5. 1999;11(5):277-285. [CrossRef]
- Costa M, Furness JB. The origins of the adrenergic fibres which innervate the internal anal sphincter, the rectum, and other tissues of the pelvic region in the guinea-pig. Z Anat Entwicklungsgesch. 1973;140(2):129-142. [CrossRef]
- Pidsudko Z. Immunohistochemical characteristics and distribution of neurons in the paravertebral, prevertebral and pelvic ganglia supplying the urinary bladder in the male pig. J Mol Neurosci. 2014;52(1):56-70. [CrossRef]
- Kaleczyc J, Kasica-Jarosz N, Pidsudko Z, Dudek A, Klimczuk M, Sienkiewicz W. Effect of castration on pelvic neurons in the male pig. Histochem Cell Biol. 2020;153(3):135-151. [CrossRef]
- Elfvin LG, Holmberg K, Emson P, Schemann M, Hökfelt T. Nitric oxide synthase, choline acetyltransferase, catecholamine enzymes and neuropeptides and their colocalization in the anterior pelvic ganglion, the inferior mesenteric ganglion and the hypogastric nerve of the male guinea pig. J Chem Neuroanat. 1997;14(1):33-49. [CrossRef]
- Luckensmeyer GB, Keast JR. Projections from the prevertebral and major pelvic ganglia to the ileum and large intestine of the male rat. J Auton Nerv Syst. 1994;49(3):247-259. [CrossRef]
- G DW. The Pelvic Plexus: Innervation of Pelvic and Extrapelvic Visceral Tissues. Microsc Res Tech. 1996;35(2):95-106. Accessed June 29, 2024. https://jglobal.jst.go.jp/en/detail?JGLOBAL_ID=200902195749010476.
- Janig W, McLachlan EM. Organization of lumbar spinal outflow to distal colon and pelvic organs. 1987;67(4):1332-1404.
- TSAKNAKIS A. Morphological Studies of the Pelvic Plexus of the Pig. Zentralbl Veterinarmed A. 1971;18(4):310-324. [CrossRef]
- Keast JR. Visualization and immunohistochemical characterization of sympathetic and parasympathetic neurons in the male rat major pelvic ganglion. Neuroscience. 1995;66(3):655-662. [CrossRef]
- CARLSTEDT A, NORDGREN S, FASTH S, HULTEN L. The influence of the pelvic nerves on. anorectal motility in the cat. Acta Physiol Scand. 1989;135(1):57-64. [CrossRef]
- Dail WG, Evan AP, Eason HR. The major ganglion in the pelvic plexus of the male rat - A histochemical and ultrastructural study. Cell Tissue Res. 1975;159(1):49-62. [CrossRef]
- Evan AP, Dail WG, Dammrose D, Palmer C. Scanning electron microscopy of cell surfaces following removal of extracellular material. Anat Rec. 1976;185(4):433-445. [CrossRef]
- Pannese E. Cuantitativo, cambios estructurales y moleculares en la neuroglia de mamíferos en envejecimiento: Una revisión. Revista europea de histoquímica. 2021;65(s1). [CrossRef]
- Pannese E, Ledda M, Arcidiacono G, Rigamonti L. Clusters of nerve cell bodies enclosed within a common connective tissue envelope in the spinal ganglia of the lizard and rat. Cell Tissue Res. 1991;264(2):209-214. [CrossRef]
- Ledda M, De Palo S, Pannese E. Ratios between number of neuroglial cells and number and volume of nerve cells in the spinal ganglia of two species of reptiles and three species of mammals. Tissue Cell. 2004;36(1):55-62. [CrossRef]
- van Velzen M, Laman JD, KleinJan A, Poot A, Osterhaus ADME, Verjans GMGM. Neuron-Interacting Satellite Glial Cells in Human Trigeminal Ganglia Have an APC Phenotype. The Journal of Immunology. 2009;183(4):2456-2461. [CrossRef]
- Hanani M, Verkhratsky A. Satellite Glial Cells and Astrocytes, a Comparative Review. Neurochemical Research 2021 46:10. 2021;46(10):2525-2537. [CrossRef]
- Andreeva D, Murashova L, Burzak N, Dyachuk V. Satellite Glial Cells: Morphology, functional heterogeneity, and role in pain. Front Cell Neurosci. 2022;16:1019449. [CrossRef]
- Auld DS, Robitaille R. Glial cells and neurotransmission: An inclusive view of synaptic function. Neuron. 2003;40(2):389-400. [CrossRef]
- Darabid H, Arbour D, Robitaille R. Glial Cells Decipher Synaptic Competition at the Mammalian Neuromuscular Junction. Journal of Neuroscience. 2013;33(4):1297-1313. [CrossRef]
- Negro S, Pirazzini M, Rigoni M. Models and methods to study Schwann cells. J Anat. 2022;241(5):1235. [CrossRef]
- Salzer J, Feltri ML, Jacob C. Schwann Cell Development and Myelination. Cold Spring Harb Perspect Biol. Published online March 19, 2024:a041360. [CrossRef]
- Espinosa-Medina I, Saha O, Boismoreau F, Brunet JF. The “sacral parasympathetic”: ontogeny and anatomy of a myth. Clinical Autonomic Research. 2018;28(1):13-21. [CrossRef]
- Sánchez Zavaleta V. Efecto de la conducta sexual y la denervación preganglionar sobre las características histológicas de la próstata y del ganglio pélvico mayor en la rata. Published online 2021. Accessed June 29, 2024. https://repositorioslatinoamericanos.uchile.cl/handle/2250/4146189.
- Kepper M, Keast J. Immunohistochemical properties and spinal connections of pelvic autonomic neurons that innervate the rat prostate gland. Cell Tissue Res. 1995;281(3):533-542. [CrossRef]
- Bertrand MM, Keast JR. Dissection of pelvic autonomic ganglia and associated nerves in male and female rats. Journal of Visualized Experiments. 2020;2020(157). [CrossRef]
- McVary KT, Razzaq A, Lee C, Venegas MF, Rademaker A, McKenna KE. Growth of the Rat Prostate Gland is Facilitated by the Autonomic Nervous System. Biol Reprod. 1994;51(1):99-107. [CrossRef]
- Manzo J, Garcia LI, Hernandez ME, Carrillo P, Pacheco P. Neuroendocrine control of urine-marking behavior in male rats. Physiol Behav. 2002;75(1-2):25-32. [CrossRef]
- Forrest SL, Payne SC, Keast JR, Osborne PB. Peripheral injury of pelvic visceral sensory nerves alters GFRα (GDNF family receptor alpha) localization in sensory and autonomic pathways of the sacral spinal cord. Front Neuroanat. 2015;9(APR). [CrossRef]
- Hancock MB, Peveto CA. A preganglionic autonomic nucleus in the dorsal gray commissure of the lumbar spinal cord of the rat. Journal of Comparative Neurology. 1979;183(1):65-72. [CrossRef]
- Baron R, Jänig W, McLachlan EM. The afferent and sympathetic components of the lumbar spinal outflow to the colon and pelvic organs in the cat. I. The hypogastric nerve. Journal of Comparative Neurology. 1985;238(2):135-146. [CrossRef]
- Hulsebosch CE, Coggeshall RE. An analysis of the axon populations in the nerves to the pelvic viscera in the rat. Journal of Comparative Neurology. 1982;211(1):1-10. [CrossRef]
- Jessica Natalia Landa García. Caracterización Anatómica y Molecular de Las Neuronas Del Ganglio Pélvico Mayor Que Inervan a La Próstata Ventral y Dorsolateral de La Rata Macho Adulta de La Cepa Wistar . Doctorado. INSTITUTO DE INVESTIGACIONES CEREBRALES, Universidad Veracruzana; 2024.
- Pacheco P, Martinez-Gomez M, Whipple B, Beyer C, Komisaruk BR. Somato-motor components of the pelvic and pudendal nerves of the female rat. Brain Res. 1989;490(1):85-94. [CrossRef]
- Nadelhaft I, Roppolo J, Morgan C, de Groat WC. Parasympathetic preganglionic neurons and visceral primary afferents in monkey sacral spinal cord revealed following application of horseradish peroxidase to pelvic nerve. Journal of Comparative Neurology. 1983;216(1):36-52. [CrossRef]
- Nadelhaft I, Booth AM. The location and morphology of preganglionic neurons and the distribution of visceral afferents from the rat pelvic nerve: A horseradish peroxidase study. Journal of Comparative Neurology. 1984;226(2):238-245. [CrossRef]
- Morgan CW, De Groat WC, Felkins LA, Zhang S -J. Intracellular injection of neurobiotin or horseradish peroxidase reveals separate types of preganglionic neurons in the sacral parasympathetic nucleus of the cat. Journal of Comparative Neurology. 1993;331(2):161-182. [CrossRef]
- Mawe GM, Bresnahan JC, Beattie MS. A light and electron microscopic analysis of the sacral parasympathetic nucleus after labelling primary afferent and efferent elements with HRP. Journal of Comparative Neurology. 1986;250(1):33-57. [CrossRef]
- Araki I, De Groat WC. Unitary excitatory synaptic currents in preganglionic neurons mediated by two distinct groups of interneurons in neonatal rat sacral parasympathetic nucleus. https://doi.org/101152/jn1996761215. 1996;76(1):215-226. [CrossRef]
- Banrezes B, Andrey P, Maschino E, et al. Spatial segregation within the sacral parasympathetic nucleus of neurons innervating the bladder or the penis of the rat as revealed by three-dimensional reconstruction. Neuroscience. 2002;115(1):97-109. [CrossRef]
- TORBEY K, LEADBETTER WF. Innervation of the Bladder and Lower Ureter: Studies on Pelvic Nerve Section and Stimulation in the Dog. J Urol. 1963;90(4):395-404. [CrossRef]
- CREED KE, TULLOCH AGS. The Effect of Pelvic Nerve Stimulation and Some Drugs on the Urethra and Bladder of the Dog. Br J Urol. 1978;50(6):398-405. [CrossRef]
- CREED KE. THE ROLE OF THE HYPOGASTRIC NERVE IN BLADDER AND URETHRAL ACTIVITY OF THE DOG. Br J Pharmacol. 1979;65(3):367-375. [CrossRef]
- Kaleczyc J, Kasica-Jarosz N, Pidsudko Z, Przyborowska A, Sienkiewicz W. The expression of androgen receptor in neurons of the anterior pelvic ganglion and celiac-superior mesenteric ganglion in the male pig. Pol J Vet Sci. 2019;22(1):151-155. [CrossRef]
- Pidsudko Z. Immunohistochemical characteristics and distribution of sensory dorsal root ganglia neurons supplying the urinary bladder in the male pig. Journal of Molecular Neuroscience. 2014;52(1):71-81. [CrossRef]
- Janikiewicz P, Wasilewska B, Mazur U, Franke-Radowiecka A, Majewski M, Bossowska A. The Influence of an Adrenergic Antagonist Guanethidine (GUA) on the Distribution Pattern and Chemical Coding of Dorsal Root Ganglia (DRG) Neurons Supplying the Porcine Urinary Bladder. International Journal of Molecular Sciences 2021, Vol 22, Page 13399. 2021;22(24):13399. [CrossRef]
- Kaleczyc J, Scheuermann DW, Pidsudko Z, Majewski M, Lakomy M, Timmermans JP. Distribution, immunohistochemical characteristics and nerve pathways of primary sensory neurons supplying the porcine vas deferens. Cell Tissue Res. 2002;310(1):9-17. [CrossRef]
- Andersson PO, Sjogren C, Uvnas B, Uvnas-Moberg K. Urinary bladder and urethral responses to pelvic and hypogastric nerve stimulation and their relation to vasoactive intestinal polypeptide in the anaesthetized dog. Acta Physiol Scand. 1990;138(3):409-416. [CrossRef]
- Kihara K, Sato K, Ando M, Sato T, Oshima H. Lumbosacral Sympathetic Trunk as a Compensatory Pathway for Seminal Emission After Bilateral Hypogastric Nerve Transections in the Dog. J Urol. 1991;145(3):640-643. [CrossRef]
- Kihara K, Sato K, Oshima H. Sympathetic Efferent Pathways Projecting to the Vas Deferens. Microsc Res Tech. 1998;42:398-408.
- Aoki H, Matsuzaka J, Banya Y, et al. Effects of Hypogastric Nerve and Sympathetic Chain Stimulation on the Pelvic Nerve Induced Penile Erection in the Dog. Urol Int. 1991;47(1):25-34. [CrossRef]
- Elfvin LG, Holmberg K, Emson P, Schemann M, Hökfelt T. Nitric oxide synthase, choline acetyltransferase, catecholamine enzymes and neuropeptides and their colocalization in the anterior pelvic ganglion, the inferior mesenteric ganglion and the hypogastric nerve of the male guinea pig. J Chem Neuroanat. 1997;14(1):33-49. [CrossRef]
- Dail WG, Moll MA, Weber K. Localization of vasoactive intestinal polypeptide in penile erectile tissue and in the major pelvic ganglion of the rat. Neuroscience. 1983;10(4):1379-1386. [CrossRef]
- Black IB, Geen SC. Trans-synaptic regulation of adrenergic neuron development: Inhibition by ganglionic blockade. Brain Res. 1973;63(C):291-302. [CrossRef]
- Zukowska Z, Pons J, Lee EW, Li L. Neuropeptide Y: a new mediator linking sympathetic nerves, blood vessels and immune system? 2011;81(2):89-94. [CrossRef]
- Dail WG, Evan AP, Eason HR. The major ganglion in the pelvic plexus of the male rat - A histochemical and ultrastructural study. Cell Tissue Res. 1975;159(1):49-62. [CrossRef]
- Su X, Gebhart GF. Effects of tricyclic antidepressants on mechanosensitive pelvic nerve afferent fibers innervating the rat colon. Pain. 1998;76(1-2):105-114. [CrossRef]
- Sienkiewicz W. Sources of the porcine testis innervation. Andrologia. 2010;42(6):395-403. [CrossRef]
- Klimczuk M, Kaleczyc J, Franke-Radowiecka A, Czaja K, Podlasz P, Lakomy M. Immunohistochemical characterisation of cholinergic nerve fibres supplying accessory genital glands in the pig. https://vetmed.agriculturejournals.cz/doi/1017221/5604-VETMED.html. 2005;50(3):119-130. [CrossRef]
- Carati CJ, Creed KE, Keogh EJ. Autonomic control of penile erection in the dog. J Physiol. 1987;384(1):525-538. [CrossRef]
- Andersson PO, Bloom SR, Mellander S. Haemodynamics of pelvic nerve induced penile erection in the dog: possible mediation by vasoactive intestinal polypeptide. J Physiol. 1984;350(1):209-224. [CrossRef]
- Partanen M, Hervonen A. The effect of long-term castration on the histochemically demonstrable catecholamines in the hypogastric ganglion of the rat. J Auton Nerv Syst. 1979;1(2):139-147. [CrossRef]
- Partanen M, Hervonen A. The formaldehyde-induced fluorescence of the developing hypogastric (main belvic) ganglion of the rat - Short adrenergic neurons and the effect of testosterone. Histochemistry. 1979;62(3):249-258. [CrossRef]
- Melvin JE, Hamill RW. Androgen-specific critical periods for the organization of the major pelvic ganglion. Journal of Neuroscience. 1989;9(2):736-742. [CrossRef]
- Melvin JE, Hamill RW. The major pelvic ganglion: androgen control of postnatal development. J Neurosci. 1987;7(6):1607-1612. [CrossRef]
- Keast JR, Saunders RJ. Testosterone has potent, selective effects on the morphology of pelvic autonomic neurons which control the bladder, lower bowel and internal reproductive organs of the male rat. Neuroscience. 1998;85(2):543-556. [CrossRef]
- Schirar A, Chang C, Rousseau JP. Localization of Androgen Receptor in Nitric Oxide Synthase- and Vasoactive Intestinal Peptide-Containing Neurons of the Major Pelvic Ganglion Innervating the Rat Penis. J Neuroendocrinol. 1997;9(2):141-150. [CrossRef]
- Purves-Tyson TD, Arshi MS, Handelsman DJ, Cheng Y, Keast JR. Androgen and estrogen receptor-mediated mechanisms of testosterone action in male rat pelvic autonomic ganglia. Neuroscience. 2007;148(1):92-104. [CrossRef]
- Félix B, Catalin D, Miolan JP, Niel JP. Effects of Testosterone on the Electrical Properties and Nicotinic Transmission of the Major Pelvic and Coeliac Ganglion Neurones. J Neuroendocrinol. 2001;13(2):193-198. [CrossRef]
- Kanjhan R, Osborne PB, Ouyang M, Keast JR. Postnatal maturational changes in rat pelvic autonomic ganglion cells: A mixture of steroid-dependent and -independent effects. J Neurophysiol. 2003;89(1):315-323. [CrossRef]
- Hervonen A, Kanerva L. Adrenergic and Nonadrenergic Axons of the Rabbit Uterus and Oviduct. Acta Physiol Scand. 1972;85(1):139-141. [CrossRef]
- Víctor Hugo Cruz Rivas. Repercusión de La Denervación Prostática y La Conducta Sexual Sobre Los Niveles Séricos de Prolactina y Testosterona y La Expresión de Sus Receptores En Ratas Sexualmente Expertas. Doctorado. Instituto de Investigaciones Cerebrales, Universidad Veracruzana; 2024.
- Mateos Moreno. Efecto de la conducta sexual y la denervación pélvica y/o hipogástrica sobre la expresión de receptores adrenérgicos, colinérgicos, andrógenos y prolactina en el ganglio pélvico mayor de la rata macho. Tesis. Published 2021. Accessed June 29, 2024. https://cdigital.uv.mx/.
Figure 1.
Schematic Representation of the Pelvic Plexus. (A) Pelvic plexus of the male rat. (B) Photomicrograph of the right pelvic ganglion of the adult male rat. VP, ventral prostate; AN, accessory nerves; DP, dorsal prostate; CN, cavernous nerve; MPG, major pelvic ganglion; PN, pelvic nerve; HN, hypogastric nerve; DNP, dorsal nerve of the penis; IMG, inferior mesenteric ganglia; VB, viscerocutaneous branch of the pelvic nerve; SBPN, Sensory branch of pudendal nerve; VP, ventral prostate; DP, dorsal prostate.
Figure 1.
Schematic Representation of the Pelvic Plexus. (A) Pelvic plexus of the male rat. (B) Photomicrograph of the right pelvic ganglion of the adult male rat. VP, ventral prostate; AN, accessory nerves; DP, dorsal prostate; CN, cavernous nerve; MPG, major pelvic ganglion; PN, pelvic nerve; HN, hypogastric nerve; DNP, dorsal nerve of the penis; IMG, inferior mesenteric ganglia; VB, viscerocutaneous branch of the pelvic nerve; SBPN, Sensory branch of pudendal nerve; VP, ventral prostate; DP, dorsal prostate.
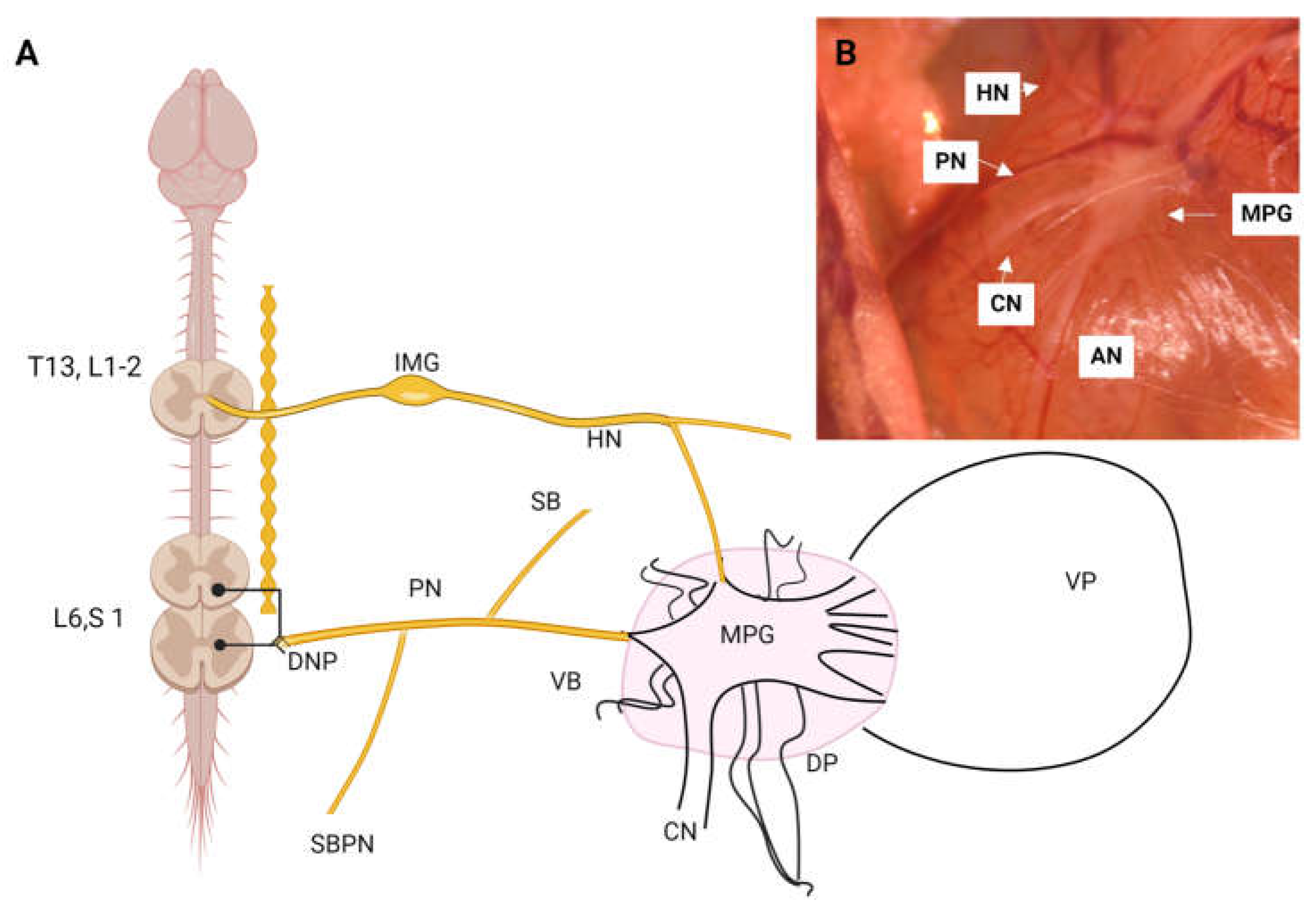
Figure 2.
Hematoxylin and eosin staining of the major pelvic ganglion in the male rat. AN, accessory nerves; CN, cavernous nerve; PN, pelvic nerve; HN, hypogastric nerve.
Figure 2.
Hematoxylin and eosin staining of the major pelvic ganglion in the male rat. AN, accessory nerves; CN, cavernous nerve; PN, pelvic nerve; HN, hypogastric nerve.
Figure 3.
Neurotransmitters in the pelvic plexus. (A) Immunofluorescence for tyrosine hydroxylase (TH) in the MPG of an adult male rat; (B) Immunofluorescence for acetylcholinesterase (ACHE) in the MPG of an adult male rat.
Figure 3.
Neurotransmitters in the pelvic plexus. (A) Immunofluorescence for tyrosine hydroxylase (TH) in the MPG of an adult male rat; (B) Immunofluorescence for acetylcholinesterase (ACHE) in the MPG of an adult male rat.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



