
Submitted:
13 August 2024
Posted:
13 August 2024
You are already at the latest version
Abstract
To investigate the effects of predation of the paddle crab Charybdis japonica on the culture and survival of scallops (Mizuhopecten yessoensis) during bottom culture, we investigated the behavioral characteristics of three sizes (small, medium, and large) of scallops in response to exposure to crabs. We also measured the activities of superoxide dismutase, catalase, arginine kinase, and octopine dehydrogenase in the gill, adductor muscle, and mantle of scallops before and after exposure to predation. Tissues that showed significant differences between control and test specimens were selected for deep sequencing of the transcriptome to identify and validate the key genes that were sensitive to predation. We found when M. yessoensis is stimulated by the presence of predators, its behavioral characteristics and related physiological and ecological indexes undergo significant changes. The results are relevant for developing specifications for M. yessoensis seedling casting during bottom culture.
Keywords:
Charybdis japonica
; Mizuhopecten yessoensis
; dynamometer method
; behavioral traits
; transcriptome
1. Introduction
The scallop Mizuhopecten yessoensis is a large cold-water filter-feeding bivalve with a fan-shaped shell [1]. It is native to Japan and Korea and mostly inhabits inner gulf regions with high salinity and no freshwater injection. Due to its high nutritional value, delicious flavor, large size, and high economic importance, M. yessoensis is popular among consumers. M. yessoensis was introduced into China in the 1980s and has become one of the most important farmed shellfish species in the northern part of China through large-scale aquaculture [2].
In China, M. yessoensis currently is cultured in two main modes: raft culture and bottom culture, and the latter is the most common mode [3]. As the scale of scallop bottom culture increased, problems such as low recapture rate and high mortality rate emerged, which have seriously affected the development of the industry [4]. Consumption by predatory organisms is one of the main causes of high mortality of scallops. In natural waters, predators of scallops are mainly sea stars, snails, and crabs. According to [5], their study of sea scallops (Placopecten magellanicus) in the Atlantic Ocean Bay found that the predatory effects of sea stars (Astro pecten americanus) and crabs (Cancer irroratus) predation on (P. magellanicus) larvae affected scallop aggregation and that this predation affected scallop survival and varied with age. According to [6], Sea stars (Asterias vulgaris Verrill) and crabs (Cancer irroratus Say) on the predatory behavior of juvenile scallops at different densities, and the results of the study showed that crabs and sea stars have high predation rates on juvenile individuals. The paddle crab Charybdis japonica is a natural predator of shellfish and the main predator of bottom-cultured shellfish in the Yellow Sea and Bohai Sea area. Thus, predation on scallops by C. japonica has caused great economic losses to fishermen [7], as it has very important impacts on scallop culture and survival. Quantifying predator-prey interactions and gaining insight into predator behaviour is critical to optimising recovery strategies. Understanding the dynamics of predator-prey size relationships can improve the success of recovery efforts [8].
Motor behavior is an essential function of animals, as it affects their survival, growth, and reproduction success [9]. The Motor behavior of scallops plays a contributing role in their targeted culture, healthy development, and improvement of economic benefits for farmers. Strong escape responses should enhance survival in the face of predation [10]. When encountering a predator, scallops can close the shell, swim, and jump to avoid being eaten [11]. The blood (body cavity fluid) is an important tissue used by aquatic animals to respond to external stimuli, and the activities of various immune-related enzymes in the blood can be used as a measure of the health status of aquatic animals. The strength of an aquatic organism’s immune system is an important physiological indicator of its ability to adapt to the external environment [12].
It is important to understand whether and how scallops can escape from predators to develop the aquaculture industry and produce healthy scallops, but little is known about the mechanisms involved in the response of M. yessoensis to predatory stimuli. In this laboratory study, we assessed the motor behavior and ability of different sized scallops to avoid predation by crabs. We also compared enzyme activities of tissues between control scallops and those exposed to continuous predatory stimulation. Finally, we conducted transcriptome sequencing of adductor muscle samples of M. yessoensis to identify genes involved in the physiological and biochemical response of scallops to stimulation by predators. Our results may be applied to improve the bottom-seeding technology of scallops in terms of prevention and control of predators.
2. Materials and Methods
2.1. Experimental Materials
M. yessoensis were caught in Longwangtang Bay (Lvshunkou District, Dalian, China) in November 2022 and transported back to the Key Laboratory of Mariculture and Stock Enhancement in North China’s Sea (Ministry of Agriculture, Dalian Ocean University, Dalian, P.R. China) in a wet state using a holding tank. Scallops were rinsed clean and temporarily cultured in tanks. They were fed with Spirulina powder at regular intervals every day. After 7 days of culture, scallops that could close naturally and were healthy and undamaged were selected for use in the experiment.
C. japonica were also caught in Longwangtang Bay in November 2022. Healthy crabs with complete appendages and cephalothorax width of 89.81–111.21 mm were cultured without feeding for 7 days. Individuals without obvious trauma, with intact chelicerae, and able to feed normally were selected for use in the experiment.
The shell length, shell height, and shell width of scallops were measured using Vernier calipers (Marr Precision Measuring Instruments Co., Ltd., Suzhou, China) with an accuracy of 0.01 mm. The wet weight of each scallop was measured using electronic scales (Changshu Shuangjie Testing Instrument Factory, Jiangsu, China) with an accuracy of 0.01 g. Scallops were classified into three size groups according to shell length, height, and width. Table 1 shows the basic biological indexes for the specimens.
2.2. Experimental Methods
2.2.1. Behavioral Observations of Scallops
For each experiment described below, we filmed for 72 h using a webcam (Model: KS-X6-QG4, Police Vision Guard, Guangdong, China). The camera was installed 3 m directly above the tank, which allowed filming under light-free conditions. For each group, a 1 h window was randomly selected to count the number of swimming events (shell opening and closing > 3 times consecutively) and jumping events (shell opening and closing ≤ 3 times consecutively).
2.2.2. Crab-Scallop Experiments
In order to gain insight into the motor behavior and escape mechanism of scallops in response to crab predation, a series of experiments were conducted, testing three different sizes of scallops. In the crab-scallop experiment, the experimental groups consisted of five small (X-sB), five medium (X-mB), or five large (X-1B) scallops in a tank, to which a crab was added for predation stimulation. The control groups for each scallop size group (sB, mB, and lB) were devoid of the presence of a crab. Each group was replicated three times.
2.2.3 Force of Clap Measurement
- Experimental design
The scallop shell closure force was measured following a previously described force gauge method [9,13]. The experimental setup included a digital force gauge (Nscing SH-III-100N, digital force gauge, Suce, Nanjing, Jiangsu, China), water tank, and stand (Figure 1). The lower valve of a scallop was fixed on the experimental table in the water tank, and the upper valve was able to open and close freely. The force meter was fixed with a bracket. One end consisted of a hooked rod fixed between the upper and lower valve, which could accurately capture each time the scallop closed its valve, and the other end was a data port connected to a personal computer using a USB port.
- Force recordings
The scallops were fixed to the test bench and the force measuring device was installed to assess the movement state of different sizes of scallops in the natural state and during continuous stimulation by the presence of a crab. The contraction of the adductor muscles is mainly divided into two stages. Phasic contractions (i.e., the rapid opening and closing of the shell) are generated by the contraction of the transverse striated muscle in the adductor muscles; in the force output diagram, every peak value represents one clap (one cycle of opening and closing) of the scallop. The force generated by the process is the force of the clap (Fclap), and the tense contraction (i.e., sustained for > 0.5 s) is the contraction of the shell. The second stage is tonic contractions (i.e., sustained for > 0.5 s), which result in slow opening and closing of the shell. They are generated by the contraction of the smooth muscles in the adductor muscles, and they show an obvious and sustained value in the output graph of the force measurement. The force generated by this process is the tonic contraction force (Ftonic). Typically, a scallop undergoes a cycle of motion that consists of rapidly clapping n times within a short period of time, followed by one more sustained slow contractions. The force generated by this process is called the phasic contractile force (Fphasic), which is the sum of Fclap.
According to Zhang et al. [9] and the results of a pre-test, M. yessoensis stops moving after 3 min of continuous stimulation by a predator, and a period of recovery time is needed before it can attempt to evade again. Therefore, the measurement time for each scallop in this experiment was set at 180.2 ± 0.2 s. First, the movement of each scallop in each size class was measured in the natural state (i.e., no crab). After 48 h, the shell closure of each scallop in each size class under continuous stimulation by the presence of C. japonica was measured. The data indexes were the maximum force of clap (Fmax), number of phasic contractions (Tphasic), number of tonic contractions (Ttonic), and frequency of shell closure (the number of times the scallop closes its shell per minute).
Because the initial response and strength of the scallop is important for its survival when it encounters a predator, the number of shell closures in the first 30 s as a percentage of the total number of shell closures (P30s) was also calculated. In order to minimize inter-individual errors, 15 parallels were set up for each set of experiments.
2.2.4 Enzyme Activity Assay
The adductor muscles, gills, and mantle tissues were removed from three scallops in the unstimulated control group and three scallops from each size group after exposure to crabs, frozen in liquid nitrogen, and preserved at −80°C for subsequent enzyme activity assays. Activities of superoxide dismutase (SOD), catalase (CAT), octopine dehydrogenase (ODH), and arginine kinase (AK) activities in each tissue from each scallop were measured using the FANKEW ELISA kit (Bio-Techne China Co. Ltd, Shanghai, China).
2.3 Transcriptome Analysis
2.3.1 RNA Extraction and Transcriptome Sequencing
Immunological measurements showed significant differences in enzyme activities of the adductor muscles of medium-sized scallops. Therefore, we selected the medium-sized adductor muscles of scallops in the natural state (control group) and after continuous stimulation by C. japonica for 3 min (experimental group) for transcriptome sequencing. We referred to them as mBJ (medium-sized scallop muscle tissue) and X-mBJ (stimulation by crab medium-sized scallop muscle tissue). Total RNA was extracted using the TruSeq Stranded mRNA LTSample Prep Kit (Illumina, USA) following the manufacturer’s protocol. Each RNA sample was subjected to RNase-free DNase I digestion (Promega, Madison, USA) to eliminate genomic DNA contamination.
The purity of the extracted RNA was detected by UV absorption and agar-agar gel electrophoresis. When OD260/OD280 was between 1.8 and 2.1 and the 28S and 18S bands were bright and clear, the RNA purity was considered to be high and could be used for subsequent experiments. First strand and second strand cDNA was synthesized using the interrupted mRNA fragments as a template. The cDNA was purified using a kit (Agencourt AMPure XP, Beverly, USA). The short cDNA fragments were subjected to end repair and adapter ligation. Then, the suitable fragments were selected and enriched by PCR amplification. A total amount of 4μg RNA per sample was used for library construction. The libraries were constructed using the TruSeq Stranded mRNA LT Sample Prep Kit (Illumina, USA) according to the manufacturer’s instructions. These library qualities was evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, USA). Then, the libraries were sequenced on the Illumina sequencing platform (HiSeq™ 2500), and 125/150-bp paired-end reads were generated. The RNA prepara-tion, libraries construction, and sequencing procedure were performed by Beijing Novozymes Technology Co., Ltd (Tianjin, China)
2.3.2 Bioinformatics Analysis
The quality of the raw data was assessed using FastQC. Raw data (raw reads) were processed using Trimmomatic [14] to remove adapters, reads containing ploy-N, and low-quality reads to obtain the clean reads. Then the clean reads were mapped to the M. yessoensis genome (https://www.ncbi.nlm.nih.gov/genome/12193) using HISAT2 [15]. The FPKM (Frag-ments Per Kilobase of exon model per Million mapped fragments) value of each gene was calculated using cufflinks [16], and the read counts of each gene were obtained by htseq-count [17]. DESeq [18] was used to compare RNA-seq data between mBJ group and X-mBJ group. Differentially expressed genes (DEGs) in mBJ versus X-mBJ were identified using the DESeq functions estimateSizeFactors and nbinom Test. The false discovery rate (FDR) control method was used to identify the threshold of the p-value in multiple tests in order to compute the significance of the differences. Threshold values were set as p < 0.05 and fold change >2. Gene ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) [19] athway enrichment analysis were performed. The stress-related genes were selected, and the sequences of selected DEGs and annotated genes were verified and compared by BLAST to ensure their similarity. The bioinformatic analysis was conducted by Beijing Novozymes Technology Co.
2.3.3 qRT-PCR
To examine the reliability of the RNA-Seq results, 9 DEGs were randomly selected for qRT-PCR validation, They included KIF13B, CYP2C8, ZCCHC8, TRXL, RAD17, OTOF, C25B8.10, CHRNA2, and PROM1A. The housekeeping gene Gapdh was used as the reference gene. The primers were designed by Primer Premier 6, with lengths of 18-27 bp, GC (ratio of two bases, guanine and cytosine, to the total base) content of 45%–55%, and synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). The RNA was reverse transcribed to cDNA using TIANGEN® FastKing gDNA Dispelling RT SuperMix (Tiangen Biotech, China). Suitable primers were designed using Primer 6 (Table 2) and synthesized by Sangon Biotech Co.,Ltd. (Shanghai, China). qRT-PCR was performed with TIANGEN® Talent qPCR PreMix (SYBR Green) (Tiangen Biotech, China) on a Roche LightCycler®96 (Roche Diagnostics, Switzerland) real-time PCR system according to the manufacturer’s protocol. Samples from the same batch experiment of the RNA-seq were used for confirmation experiment. All reactions were performed in triplicate. The qRT-PCR conditions were as follows: 3 min at 95℃, followed by 45 cycles of 5 s at 95℃, and 15s at 60℃. Dissociation curve analysis was carried out to determine the target specificity. The relative expression of target genes was calculated using the 2-ΔΔCT method [19].
2.4 Statistical Analysis
The experimental data were expressed as Mean ± Standard deviation (mean ± S.D.). SPSS25.0 software was used to conduct one-way ANOVA on the motor behavior indicators of the same state (normal state or crab stimulated state) with different specifications or the same specifications with different states, and LSD was used to make a two-by-two comparison of the indicators with significant differences; two-way ANOVA was conducted to analyze the interactions between the specifications and the states, with P<0.05 regarded as the difference being significant, and P<0.01 regarded as the difference being extremely significant.
3. Results
3.1. Scallop Behavior
Video recordings showed that it was difficult for crabs to prey on scallops and that scallops avoided crabs through continuous closed-shell behavior or movement behavior. In the crab-scallop experiment, the number of jumping and swimming behaviors of each scallop increased significantly in the presence of the crab. These behaviors differed among scallop sizes, as the number of jumps was greater than the number of swims in medium- and large-sized scallops. The number of swimming episodes of scallops of all sizes in the experimental groups stimulated by the presence of crabs was significantly greater than that of the control group (P < 0.05). Additionally, the number of movement behaviors of small- and medium-sized scallops in the experimental group was significantly higher than that of the control group, but there was little difference in this measurement for large-sized scallops in the treatment and control groups (Figure 2).
3.2 Force of Clap
Continuous stimulation by exposure to crabs resulted in different response patterns in different sizes of scallops. Specifically, Tphasic and Ttonic of small- and medium-sized scallops increased significantly relative to the control (P < 0.05), whereas the increases observed for large-sized scallops was not significant (Figure 3). Pressure transducer-dynamometer recordings showed that in the presence of the predator, scallops alternated between phasic and tonic contractions, for large-sized scallops, the phasic contraction force lasted throughout the 3 min experimental period. Under predation stimulation, the number of claps was intensive and the phasic contraction force of small- and medium-sized scallops increased (Figure 3). The force measurements showed that the larger scallops had a larger clap force than the smaller scallops. Additionally, within a size class, the force of the clap was greater when the predator was present than when it was absent. Over time, the shell-closing ability of scallops of all sizes decreased (Figure 3 and Figure 4).
Figure 4 shows the Fmax value and average shell closure frequency of different sizes of scallops. In the presence of the predator, the Fmax of small-sized scallops was only 7.82 N, whereas that of large-sized scallops was 15.45 N. The one-way analysis of variance (ANOVA) revealed a significant difference among the Fmax of different sized scallops (P < 0.05). Significant differences among small, medium, and large scallops both with and without the predator were also detected for the closure frequency (the number of times that the cycle of locomotion was completed within a unit of time). Small- and medium-sized scallops closed their shells more frequently than large-sized scallops in both the control and crab-stimulated state. Compared with the control, crab-stimulation resulted in increased frequency of shell closure of scallops of all sizes, and the increase in frequency of small- and medium-sized scallops was statistically significant (P < 0.05). Additionally, the difference in the frequency of shell closure of scallops before and after stimulation was significant within all size classes, but the value was highest for the medium-sizes scallops.
Figure 5 shows the Ftotal, Fphasic, and Ftonic data. Overall, the Ftonic of scallops was greater than that of Fphasic in the control group for all scallop size classes, and the Ftonic between the control and experimental group was significant when the scallops were stimulated by the presence of the predator. The one-way ANOVA results showed no significant difference in Fphasic and Ftonic between small- and medium-sized scallops in the absence of crabs, but the Fphasic and Ftonic values of these size groups decreased after exposure to the predator (P < 0.05). Fphasic and Ftonic of large-sized scallops were lower than those of small- and medium-sized scallops. A significant interaction between scallop size and the presence or absence of crabs was also detected (two-way ANOVA, P < 0.05). Both size and the presence or absence of the predator also had significant effects (P < 0.05) on Ftonic and Ftotal, and an interaction effect was detected.
3.3 Changes in Enzyme Activity of Scallops due to Crab Predation
Exposure to the predator significantly affected the activities of SOD, CAT, ODH, and AK in the gill, adductor muscles, and mantle of scallops. Fluctuations in these activities occurred in the tissues after sustained stimulation (Figure 6). The enzyme activities in gill tissues of different-sized scallops changed to different degrees after exposure to crabs (Figure 6A). Compared to the controls, the activities of SOD, CAT, and ODH in the gills of medium- and large-sized scallops exposed to the predator were significantly different (P < 0.01), and the decline of SOD in medium-sized scallops was the most prominent. The activity of AK in each size group showed a significant decrease (P < 0.01).
In the mantle, crab stimulation led to an extreme decrease of SOD activity (P < 0.01) in medium-sized scallops and an extreme increase of CAT activity but a decrease in AK activity in large-sized scallops (P < 0.01). However, changes in enzyme activities in small scallops were not obvious (Figure 6B).
In the adductor muscles, the SOD activity increased significantly (P < 0.01) and the ODH activity decreased significantly (P < 0.01) in large-sized scallops exposed to the predator. Medium-sized scallops showed the most pronounced changes in enzyme activities after exposure to crabs. Compared with the control group, the activities of SOD and ODH increased very significantly (P < 0.01), and the activities of CAT and AK decreased significantly (P < 0.05). No difference was observed for the activities of CAT in small-sized scallops, but the differences in SOD, ODH, and AK activities were highly significant.
3.4 Transcriptome Results
3.4.1 Transcriptome Sequencing
The adductor muscle tissue RNA concentration of mBJs and X-mBJs (X-mBJ1, X-mBJ2, and X-mBJ3) samples ranged from 70.0000 to 289.0000 ng/μL, and the total amount of RNA was 2.4500–9.2480 μg. The RNA integrity value was > 6.0, and the quality of the samples met the quality requirements for library sequencing.
A total of 58.41–61.74×106 raw reads were obtained from transcriptome sequencing, and 55.62–60.62×106 clean reads were obtained after removing the junctions and low-quality reads, with a data volume of 54.68 G. The GC percentage was 41.05%–43.85%, and the Q20 and Q30 values were over 94.92% and 87.6%, respectively. The highest error rate of a single base was only 0.07%, which showed that our data were highly credible and could be used for subsequent analysis.
3.4.2 Analysis of Differentially Expressed Genes (DEGs)
The correlation coefficients within and between groups were calculated and plotted as a heatmap, according to the Fragments Per Kilobase per Million mapped fragments (FPKM) values of all of the genes in each sample. As shown in the left part of Figure 8, there was a difference in the sample correlations between the mBJ and X-mBJ groups.
DESeq(https://bioconductor.org/packages/release/bioc/html/DESeq2.html) was used to compare the RNA-seq data of the X-mBJ and mBJ groups and draw a volcano plot to quickly and intuitively identify the regulation and distribution of the differentially expressed genes (DEGs) in the adductor muscle of scallops after crab stimulation (right part of Figure 7).
Transcriptome sequencing resulted in 780 significant DEGs, including 322 up-regulated genes and 458 down-regulated genes. The expression ranges of up- and down-regulated genes were comparable, and the overall significance differences were relatively obvious.
3.4.3 Gene Ontology (GO) Functional Enrichment Analysis of DEGs
Based on GO database analysis and functional enrichment classification of the 780 DEGs in the X-mBJ vs. mBJ comparison, 198, 98, and 331 DEGs were annotated to the biological process, cellular component, and molecular function categories, respectively. Figure 8 shows the 10 GO entries with the most DEGs enriched for each category. DEGs in the biological process were mainly enriched in the cellular nitrogen compound biosynthetic process, phosphate-containing compound metabolic process, and phosphorus metabolic process. In the cellular component category, DEGs were mainly enriched in protein-containing complex, cytoplasm, and membrane-bounded organelle processes. In the molecular function category, DEGs were enriched the processes of transporter activity, transmembrane transporter activity, and transition metal ion binding. These results showed that the gene expression of scallops of different sizes changed to different degrees after exposure to predation.
3.4.4 Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of DEGs
KEGG is a comprehensive database that integrates genomic, chemical, and systemic-functional information, and KEGG analysis improved our understanding of the biological functions of DEGs in scallops stimulated by crab. A total of 159 DEGs were enriched in 86 KEGG pathways, and Figure 9 lists the 20 most enriched pathways. Among them, ribosome, oxidative phosphorylation, autophagy-animal, purine metabolism, ribosome biogenesis in eukaryotes, and nucleocytoplasmic transport were the most significantly enriched.
3.4.5 qRT-PCR Validation Analysis
Nine DEGs were subjected to qRT-PCR validation analysis, and the expression patterns of the selected genes were identical and highly correlated with the RNA-Seq results (R2 = 0.91). These results confirm the reliability of the transcriptome analysis results (Figure 10).
4. Discussion
4.1 Effects of Crab Predation on the Closed-Shell Force of Scallops
The shell opening and closing behavior and mantle state of a bivalve can visually reflect the physiological condition of the shellfish, which is an effective behavioral indicator of the organism’s response to environmental changes [21]. For example, a scallop can produce slow or fast movements to move away from predators through the contraction or diastole of the adductor muscles [22]. In this study, the Fmax of large, medium, and small scallops was measured using a dynamometer, and the Tphasic, Ttonic, Fmax, and frequency of shell closure data were collected and analyzed. We found that the Fmax of scallops both with and without the presence of the predator increased with increasing scallop size and that the Fmax of scallops in the same size group produced a greater shell closure force in the presence of crabs, Zhang et al [9] reported similar results. Besides, and the experiment used different specifications of crabs to stimulate scallops, the data showed that with the increase of C. japonica specifications, the number of jumps of scallops increased significantly, although increased swimming behavior was not obvious. Based on all of these findings, we speculate that scallops first choose the bitemporal contraction response when stimulated by predators (i.e., shells open and close quickly to produce jumping behaviors to escape from being eaten). The results were consistent with the findings of Zhang et al [9] who used starfish (Asterias amurensis) to stimulate M. yessoensis.
Scallops have two strategies to cope with predators: either they close their shells tightly or they jump quickly to escape. A thick, hard shell provides protection against predators, but conversely a heavy shell reduces the scallop’s ability to swim. Therefore, there is a trade-off between mechanical protection and swimming ability when scallops face threats from predators, and their escape behavior in the face of predatory stimuli may change over time during individual development [11]. We found that the Tphasic and Ttonic values of crab-stimulated scallops were higher than those of the control group for both small- and medium-sized scallops. Additionally, the frequency of shell closure increased significantly upon stimulation, and small-sized scallops closed their shells frequently in the first 30s after stimulation. In contrast, no significant changes in the Tphasic and Ttonic values of large-sized scallops were detected after stimulation. The duration of shell closure in this experimental group was more than twice as long as that of the control group, and the increase in the frequency of shell closure was not significant. These results suggest that small- and medium-sized scallops opted for locomotor behavior to escape from predators, whereas large-sized scallops preferentially used shell closure to escape from predators.
As scallops grow, the shell thickens and the ability to cope with predator attacks improves, but swimming ability may decline [11]. Changes in swimming behavior and ability with age and size vary among species. The locomotor ability (number of shell closures and magnitude of shell closure force) of all sizes of scallops tends to decrease with age. According to Tremblay et al [13], the swimming behavior of Amusium balloti, Placopecten magellanicus, Pecten fumatus, Mimachlamys asperrima, and Crassadoma gigantea and found that some scallops could avoid predation by staying still due to the advantage of their shell shape, whereas the more active scallops avoided predators by swimming. In a study of Aequipecten opercularis, shell closure during the first and second escape responses was more frequent in smaller individuals than in larger ones [23]; which is consistent with the results of our study of M. yessoensis. Therefore, we hypothesized that smaller-sized M. yessoensis were more active and avoided predation by C. japonica through rapid shell closure, whereas larger scallops avoided predators by decreasing the frequency and increasing the duration of shell closure.
4.2 Effects of Crab Predation on the Enzyme Activities of Scallops
Enzyme activity is the fundamental driver of all chemical changes in living organisms, so the regulation of enzyme activity is one of the most important ways to realize the regulation of biological metabolism [23]. The immunity of scallops is affected by exposure to stressors, including predators. SOD and CAT are two important immunoenzymes in the shellfish immune system, and they play an important role in maintaining the balance of the antioxidant system in shellfish [4,25]. CAT and SOD scavenge and balance intracellular reactive oxygen radicals; thus they are important indicators of the immune defense ability of shellfish [11]. SOD is also involved in defense against aging and biomolecular damage [26].
In this study, we compared the SOD, CAT, ODH, and AK activities in the adductor muscles of control scallops and scallops exposed to crabs. The SOD and CAT activities of different sized scallops were affected crab stimulation. The SOD activity in the adductor muscles of large- and medium-sized scallops increased significantly, whereas it decreased significantly in the adductor muscles of small-sized scallops. Thus, the acute stress response depended on scallop size. Previous studies has also reported that the acute stress response varies according to size and age. According to Yang et al [27], the immune response of age II Chinese softshell turtles (Trionyx sinensis) was positively correlated with body weight compared to age I turtles and that an increase in body weight resulted in an increase in the immune response. Mourente et al [28] found the antioxidant enzyme activities of male red prawns (Aristeus antennatus) were closely related to body length and age, and the activity of SOD increased with increasing body length. Hao et al [29] also showed that the antioxidant capacity and high temperature tolerance of older M. yessoensis were significantly higher than those of smaller and younger scallops.
Generally, stronger CAT and peroxidase activities are related to greater resistance and ability to eliminate free radicals. When the living environment of organism changes, thereby putting them under stress, the activities of these two enzymes undergo changes to allow the organism to adapt to the new environment [30]. In our study, the CAT activity of adductor muscles of different sized scallops exposed to crabs decreased to different degrees, and it decreased most obviously in medium-sized scallops. In contrast, the SOD enzyme activity of medium-sized scallops increased significantly. We speculate that CAT was preferentially involved in the scavenging of reactive oxygen radicals when scallops perceived the threat of predation, and the specific reasons need to be further investigated.
Putative ODHs (OpDHs) play an important role in the anaerobic metabolism of marine invertebrates, especially shellfish. The first OpDH was discovered in 1959, when Van Thoai and Robin found that ODH was produced by an enzymatic reaction in a variety of marine invertebrates [31]. In 1969 Van Thoai et al isolated and purified ODH for the first time from scallop adductor muscles [32]. Other researchers reported that the production of octopus enzyme by ODH is analogous to the production of lactic acid by lactate dehydrogenase in vertebrate muscles. In their studies, the elevated ODH levels correlated with an increase in swimming ability, as adenosine triphosphate (ATP) production in this activity was supported by hydrolysis of phosphoarginine, which was subsequently followed by the continuation of oxidation through ODH-catalyzed NADH and L-arginine to produce octopus alkali, NAD+, and water to ensure the intracellular redox balance [32,33].
Our analysis of the adductor muscle activity of scallops revealed a highly significant difference in the ODH activity between control and crab-stimulated scallops of all three sizes. In particular, we detected a highly significant increase in the ODH activity of medium- and small-sized scallops in the groups exposed to predation. Therefore, we hypothesized that the up-regulation of ODH synthesis may be related to acute stress or sudden swimming activity in M. yessoensis. According to [33], the escape and swimming behaviors produced by bivalves in response to a predator are supported by the trans-phosphorylation of phospho-L-arginine and by anaerobic glycolysis to obtain ATP to support this intense muscle activity. During the escape response or subsequent recovery, bivalves restore phospho-L-arginine via anaerobic glycolysis and ultimately produce octopine. Octopine plays different roles in different tissues, and its level was higher in swimming scallops. The adductor muscle mass and ODH content Argopecten ventricosus, which are chronically exposed to predators, are high [35], which is consistent with the results of our study of M. yessoensis.
AK is one of the most important enzymes regulating energy metabolism in invertebrates, and it plays a role in the reversible transfer of phosphate groups between ATP via the enzyme-specific guanidinium receptor to keep ATP at relatively stable levels [36]. It also can be directly or indirectly involved in related immune responses [37]. AKs may be involved in the maintenance of normal life activities and in an organism’s defense against adverse external environments and when an organism is subjected to environmental threats that result in locomotor behaviors [38]. In our study, we found that AK activity in different tissues of scallops of all sizes was down-regulated after crab stimulation. AK expression in the adductor muscles of medium- and small-sized scallops was significantly down-regulated, indicating that scallops experience muscle stress and undergo accelerated energy metabolism when is exposed to predators. This result suggests that AK plays an important role in the muscle movements of stressed scallops [39]. reported similar findings. Predation stress causes scallops to produce explosive locomotor behavior, which requires a large amount of energy. According to Smits et al. [33], AK plays a role in vigorous muscle movements after analyzing the enzyme activity of scallop adductor muscles following escape or locomotor behavior. According to Gäde et al [40], AK decreased significantly in bivalves after jumping locomotor movements, which is consistent with our findings for M. yessoensis. The decrease in AK expression under predation stress also reflects the increase in stress-related energy demand, and the elevated AK content accelerates the energy metabolism of the organism. This process provides energy for the organism to cope with predation stress, which is conducive to the scallop’s swimming ability and survival under predator attack.
4.3 Effects of Crab Predation on the Transcriptome of Scallops
Predator-prey relationships develop gradually in nature, with the predator producing a stress response in its prey [41]. Scallops have a typical escape response that involves a series of rapid valvular internalizations or claps that allow them to jump or swim to escape from the predator [11]. Physiological responses of organisms are mediated by gene expression, and predation pressure leads to stress responses in animals, and differential genes may be focused on stress, energy metabolism, immunity and other related components. In this study, transcriptome sequencing of adductor muscles of scallops after predation stimulation by crabs yielded 780 DEGs, of which 623 were annotated. GO annotation showed that more differential genes were enriched for cellular nitrogen compound biosynthetic process, phosphate-containing compound metabolic process, transporter activity, and transmembrane transporter activity than for other processes. These enriched genes indicated that predation stress activated a series of physiological activities in the scallops, including significant up-regulation of complement C1q like 4 (C1QL4).
C1QL4 is a member of the C1q/TNF superfamily, for which the presence of a globular C1q domain is the hallmark [42–45]. The members of this superfamily are involved in various important physiological functions, such as the innate immune response [46], insulin metabolism [47], and synapse homeostasis [48,49]. C1q is part of complement C1 and participates in the classical activation pathway, in which it binds to antibodies in antigen-antibody complexes and activates C1r and C1s. Therefore, C1q is an important bridge connecting acquired and innate immunity, and C1q-like proteins have been found in numerous taxa (e.g., Lampetra japonica, Branchiostoma, Pyrosomella verticilliata, and Echinocardium) [50]. In addition to playing a role in classical activation pathways, C1q is involved in many immune processes [51], including removal of regulatory cells to maintain immune tolerance [52,53], B cells [54,55], T cells [56], and fibroblasts [57]. C1q also plays a role in development [58], trauma healing [59], and other processes. In studies on rodents, C1QL4 expression was found to be regulated by developmental and hormonal factors, and it activated steroids to produce an acute response [60]. In the present study, scallops showed significant up-regulation of C1QL4 after exposure to the predator. We hypothesize that this predatory stress induced changes in the energy metabolism and cellular function of scallops that were associated with the expression of C1QL4. Therefore, C1QL4 may play a role in the response of scallops to predatory stimuli.
Transcriptome sequencing of scallops after predator stimulation revealed significant changes in the expression of kinesin family member 13 B (KIF13B), adenylate kinase 9 (AK9), and hemicentin (HMCN) genes. KIF is a superfamily of microtubule-associated motor proteins that serve a variety of functions in cells, such as mediating intracellular vesicle and organelle transport as well as cytokinesis [61], that utilize the energy generated by ATP hydrolysis to transport cellular material along microtubules [62]. KIFs also play a central role in the regulation of synaptic function. According to Willemsen et al [63], KIF4A and KIF5C are important in regulating both excitatory and inhibitory synaptic transmission in rat hippocampal neurons. In vivo, KIF13B binds to vascular endothelial growth factor to act together in cells to promote endothelial cell growth, which is crucial in the growth of blood vessels [64]. We found that KIF13B expression was significantly up-regulated in scallop adductor muscles after stimulation by crabs. We hypothesize that predatory stimulation caused stress in scallops, leading to synaptic dysfunction or dysregulation.
AK is a monomeric enzyme found throughout plants, animals, and microorganisms that plays an important role in energy metabolism as well as a variety of biological processes within the cell. AK9 is the ninth isoform of AK [65]. The main function of AKs is to catalyze the interconversion between adenosine diphosphate (ADP) and ATP: ATP + AMP ↔ 2ADP. When the levels of ATP and ADP change, so do the levels of AMP, thus enabling AMP-influenced enzymes and compounds to receive stress signals and respond metabolically [66]. AK and the downstream AMP signaling system are an integrated metabolic monitoring system in the body. This system senses and regulates the cellular energy state and transmits signals to metabolic sensors to maintain the energy balance of the organism and respond to various stresses by altering the cellular energy metabolism through growth factors and hormones [66]. According to Yang [67], an AK4 knockdown experiment was conducted in male zebrafish, revealing alterations in cellular energy status and an increase in cellular death within the spermatheca. In our experiment, AK9 expression in adductor muscles of scallops was down-regulated after predation stimulation. We propose that scallops respond to predators by accelerating their metabolism under predatory conditions, thus enhancing their ability to escape and reducing stress-induced damage.
HMCN1, member of the hemicentin family of proteins, is an extracellular matrix protein that encodes immunoglobulins [68]. It is involved in the formation of the dynamic base of the cell and has important roles in cell organization, migration, invasion of the basement membrane, and formation of stable cell-cell contacts [69]. Chowdhury et al [70] found that HMCN1 can direct fibroblast differentiation, regulate the formation of stress fibers during differentiation, and induce transforming growth factor-β-mediated effects. Carney et al [71] studied zebrafish and found that HMCN1 mutation defects led to developmental defects such as blistering of fins. Additionally, HMCN1 in mammals plays an important role in tissue development and injury response [72]. We found that HMCN1 expression was down-regulated in scallops after they were stimulated by predation. We hypothesize that it is also involved in regulating the stress or escape behavior of scallops, but the specific regulatory mechanism needs to be further investigated.
In marine fish exposed to predators, expression of genes such as prominin 1A (PROM1A) were down-regulated and members of the zinc finger protein family and kinesin family were up-regulated [73]. In our study, expression levels of DEGs such as ZCCHC8, a member of the zinc finger protein family, and KIF13B, C1ql4, and OTOF, members of the kinesin family, were up-regulated, and DEGs such as CHRNA2, HMCN1, and PROM1A were down-regulated. Therefore, we hypothesize that these genes could play a regulatory role when scallops are stimulated by predators, but the specific regulatory mechanism needs to be further verified.
5. Conclusions
In summary, we found that the predation by C. japonica causes stress and related physiological responses in M. yessoensis. Small- and medium-sized scallops tend to escape from predation by rapid swimming and other escape behaviors, whereas large-sized scallops tend to close their shells and remain motionless. During this time period, physiological activities such as enzyme activity, substance metabolism, and cell proliferation are regulated to cope with predation stress.
Author Contributions
Z.H. designed the study. D.Z., Y.T. performed the study. Z.H. and X.L. analyzed the data. X.L. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funds from Marine Economy Development Special Project of Liaoning Province Department of Natural Resources, the Central Government Subsidy Project for Liaoning Fisheries (2023) and the Science and Technology Foundation of Dalian (2021JB11SN035).
Institutional Review Board Statement
Since Mizuhopecten yessoensis and Charybdis japonica are invertebrates, ethical review and approval were waived for this study. The content of this article does not involve human or animal research in its institutional review board statement.
Data Availability Statement
All data generated or analyzed during this study are included in this published article. The Transcriptome raw sequence reads obtained in this study have been submitted to the NCBI SRA database under accession numbers PRJNA1035918. https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1035918.
Acknowledgments
The authors wish to express thanks to the staff of Key Laboratory of Mariculture and Stock Enhancement in North China’s Sea, Ministry of Agriculture, China, for their help with the experiment. In addition, the author would like to thank the International Science Editing Company for helping to improve the language of this article.
Conflicts of Interest
The authors declare that they have no known competing financial or non-financial, professional, or personal conflicts that could have appeared to influence the work reported in this paper.
References
- Zhang, Z.X., Zhang, J.H., & Wu, W.G. Ecological carrying capacity assessment of bottom-culture Yesso scallops, Patinopecten yessoensis, in Zhangzi Island. Journal of Fishery Sciences of China. 2021,28(07):878-887.
- Cong, H.H., Geng, W.H., & Zhu, J.W. Patinopecten yessoensis mantle polypeptides enhanced the structural stability of myofibrillar proteins from silver carp. Meat Research. 2022, 36(09): 13-19.
- Wang, Y., & Zhou, L. Bottom sowing of proliferation of Patinopecten yessoensis yield research; case in Zhangzidao. Chinese Fisheries Economics. 2014, 32(01): 104-109.
- Gao, Z. K. Effects of environmental stresses on physiological, immunological parameters and behavioral characteristics of Patinopecten yessoensis. M.A. Thesis. Shanghai Ocean University. Shanghai. 2016.
- Shank, B.V., Hart, D.R., & Friedland, K.D. Post-settlement predation by sea stars and crabs on the sea scallop in the mid-Atlantic bight. Marine Ecology Progress. 2012, 468, 161–177. [CrossRef]
- Barbeau, M.A., Scheibling, R.E. Behavioral mechanisms of prey size selection by sea stars (Asterias vulgaris verrill) and crabs (Cancer irroratus say) preying on juvenile sea scallops (placopecten magellanicus (Gmelin)). Journal of Experimental Marine Biology and Ecology. 1994, 180(1), 103–136. [CrossRef]
- Yu, Z.H., Yang, H.S., & Liu, B.Z. Predation of scallop Chlamys farreri by crab Charybdis japonica. Marine Science. 2010,34(12), 62-66.
- Sclafani, M., Bopp, J., & Havelin, J. Predation on planted and wild bay scallops (Argopecten Irradians Irradians) by busyconine whelks: studies of behavior incorporating acoustic telemetry.2021, 169(66). [CrossRef]
- Zhang, J.H., Xia, Y.Y., & Gao, Z.K. Force production during shell clap of scallop Pationopecten yessoensis and its response to predator starfish. Journal of Fishery Sciences of China. 2021,28(07), 871–877.
- Guderley, H.E., Himmelman, J.H., & Nadeau, M. Effect of different predators on the escape response of placopecten magellanicus. Marine Biology: International Journal on Life in Oceans and Coastal Waters. 2015, 162(7), 1407–1415. [CrossRef]
- Xia, Y.Y. Effects of Environmental Stress on the Survival, Behavior Metabolism and Immunity of Scallops. M.A. Thesis. Shanghai Ocean University. Shanghai. 2010.
- Duan L Z., Huang X Y., & Hu W B. Comparison of three coelomic fluid enzymes activities in scallop Patinopecten yesoensis. Chinese Agricultural Science Bulletin. 2015, 31(02), 112–117.
- Tremblay, I., Guderley, H.E., & Himmelman, J.H. Swimming away or clamming up: The use of phasic and tonic adductor muscles during escape responses varies with shell morphology in scallops. The Jounrnal of Experimental Biology. 2012, 215(Pt23), 4131–4143. [CrossRef]
- Bolger, A.M., Lohse, M., & Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics. 2014, 30(15), 2114–2120. [CrossRef]
- Kim, D., Langmead, B., & Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nature Methods. 2015,12(4), 357–360. [CrossRef]
- Trapnell, C., Williams, B.A., & Pertea, G. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature Biotechnology. 2010,28(5), 511–515. [CrossRef]
- Anders, S., Pyl, P.T., & Huber, W. HTSeq—A python framework to work with high-throughput sequencing data. Bioinformatics. 2015,31(2),166–169. [CrossRef]
- Anders, S., & Huber, W. Differential expression analysis for sequence count data. Nature Precedings. 2010, 1–1. [CrossRef]
- Kanehisa, M., Araki, M., & Goto, S., Hattori, M. KEGG for linking genomes to life and the environment. Nucleic Acids Research, 2007, 36(suppl-1), D480–D484. [CrossRef]
- Livak, K. J., & Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods. 2001, 25(4), 402–408.
- Xia, Y.Y., Zhang, J.H., & Liu, Y. Behavioral characteristics and physiological responses to hypoxic stress in Patinopecten yessoensis. Journal of Fishery Sciences of China. 2021,28(10), 1319–1328.
- Schalkhausser, B., Bock, C., & Partner, H-O. Escape performance of temperate king scallop, Pecten maximus under ocean warming and acidification. Marine Biology. 2014, 161, 2819–2829. [CrossRef]
- Schmidt, M., Philipp, E.E.R., & Abele, D. Size and age-dependent changes of escape response to predator attack in the Queen scallop Aequipecten opercularis. Marine Biology Research. 2008, 4(6), 442–450. [CrossRef]
- Greenway, S.C., & Storey, K.B. The effect of prolonged anoxia on enzyme activities in oysters (Crassostrea virginica) at different seasons. Journal of Experimental Marine Biology and Ecology. 1999, 242(2), 259–272. [CrossRef]
- Liu, Z.H., Mou, H.J., & Wang, Q.Y. Research progress of immune related enzymes in Mollusca. Marine Fisheries Research. 2003, 024(003), 86–90.
- Yao, C.L., Wang, W.N., & Wang, A.L. Progess of studies on superoxide dismutase in the body of aquatic animals. Marine Sciences. 2003, 27(10), 18–21.
- Yang, X.L., & Zhou, J.G. Influence of age size and nutrition of trionyx sinensis on the immune response. Journal of Fishery Sciences of China. 1999, 23(4), 5.
- Mourente, G., & Daz-Salvago, E. Characterization of antioxidant systems, oxidation status and lipids in brain of wild-caught size-class distributed Aristeus antennatus (Risso,1816) Crustacea, Decapoda. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology. 1999, 124(4), 405–416.
- Hao, Z.L., Tang, X.J., & Ding, J. Survival rate,oxygen consumption rate and immune enzymetic activity of Mizuhopecten yessoensis at high temperature. Chinese Journal of Ecology. 2014, 33(6), 7.
- Ding, R.X., Huang, X.M., & Zhao, W. Effects of pH acute stress on the behavior and immune enzyme activity of Babylonia Areolata. Fishery modernization. 2022, 49(6), 7.
- Thoai, N.V., Huc, C., & Pho, D.B. Octopine déshydrogénase Purification et propriétés catalytiques. Biochimica et Biophysica Acta (BBA)-Enzymology. 1969, 191(1), 46–57.
- Zheng Y. Octopine dehydrogenase in the adductor muscle of live scallop and its changes during postharvest. M.A. Thesis. Dalian Ocean University. Dalian. 2018.
- Smits, S.H.J., Meyer, T., & Mueller, A. Insights into the Mechanism of Ligand Binding to Octopine Dehydrogenase from Pecten maximus by NMR and Crystallography. Plos One. 2010, 5(8), e12312. [CrossRef]
- Strahl, J., Dringen, R., & Schmidt, M. M. Metabolic and physiological responses in tissues of the long-lived bivalve Arctica islandica to oxygen deficiency. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology. 2011, 158(4), 513–519. [CrossRef]
- Guerra, C., Zenteno-Savín, T., & Maeda-Martínez, A.N. The effect of predator exposure and reproduction on oxidative stress parameters in the Catarina scallop Argopecten ventricosus. Comparative Biochemistry and physiology. Part A: Molecular & Integrative Physiology. 2013, 165(1), 89–96. [CrossRef]
- Morris, S., Van Aardt, W.J., & Ahern, M.D. The effect of lead on the metabolic and energetic status of the Yabby, Cherax destructor, during environmental hypoxia. Aquatic Toxicology. 2005, 75(1), 16–31. [CrossRef]
- Yao, C.L., Wang, Z.Y., & Xiang, J.H. Structure and function of arginine kinase in crustacean. Chinese Journal of Biochemistry and Molecular Biology. 2008, 24(3), 203–208.
- Huang, L.N., Liu, N., & Zhao, J. The research prospect of Arginine Kinase. Life Science Research. 2015, 19(05), 452–456.
- Voncken, F., Gao, F., & Wadforth, C. The phosphoarginine energy-buffering system of Trypanosoma brucei involves multiple arginine kinase isoforms with different subcellular locations. PLoS One. 2013, 8(6), e65908. [CrossRef]
- Gäde, G., Weeda, E., & Gabbott, P.A. Changes in the level of octopine during the escape responses of the scallop, Pecten maximus (L.). Journal of comparative physiology. 1978, 124, 121–127. [CrossRef]
- Qin, Y.L., Peng, H.L.Y., & Fu, S.J. Effects of food deprivation on fast-start swimming and predator-prey interaction between a predator and prey fish species. Chinese Journal of Ecology. 2016, 35(09), 2429–2434.
- Hirai, H., Pang, Z., & Bao, D. Cbln1 is essential for synaptic integrity and plasticity in the cerebellum. Nature Neuroscience. 2005, 8(11), 1534–1541. [CrossRef]
- Matsuda, K., Yuzaki, M. Cbln family proteins promote synapse formation by regulating distinct neurexin signaling pathways in various brain regions. European Journal of Neuroscience. 2011, 33(8), 1447–1461. [CrossRef]
- Stevens, B., Allen, N.J., & Vazquez, L.E. The classical complement cascade mediates CNS synapse elimination. Cell. 2007, 131(6), 1164–1178. [CrossRef]
- Uemura T., Lee S J., & Yasumura M. Trans-synaptic interaction of GluRδ2 and Neurexin through Cbln1 mediates synapse formation in the cerebellum. Cell. 2010, 141(6), 1068–1079. [CrossRef]
- Reid, K.B.M., Gagnon, J., & Frampton, J. Completion of the amino acid sequences of the A and B chains of subcomponent C1q of the first component of human complement. Biochemical Journal. 1982, 203(3), 559–569. [CrossRef]
- Yamauchi T., Kamon J., Waki H., Imai, Y., Shimozawa, N., Hioki, K., Uchida, S., Ito, Y., Takakuwa, K., Matsui, J., Takata, M., Eto, K., Terauchi, Y., Komeda, K., Tsunoda., M., Murakami, K., Ohnishi, Y., Naitoh, T., Yamamura, K., Ueyama, Y., Froguel, P., Kimura, S., Nagai, R., Kadowaki, T. Globular adiponectin protected ob/ob mice from diabetes and ApoE-deficient mice from atherosclerosis. Journal of Biological Chemistry. 2003, 278(4), 2461–2468. [CrossRef]
- Bolliger, M.F., Martinelli, D.C., & Südhof, T.C. The cell-adhesion G protein-coupled receptor BAI3 is a high-affinity receptor for C1q-like proteins. Proceedings of the National Academy of Sciences. 2011, 108(6), 2534–2539. [CrossRef]
- Ressl, S., Vu, B.K., & Vivona, S. Structures of C1q-like proteins reveal unique features among the C1q/TNF superfamily. Structure. 2015, 23(4), 688–699. [CrossRef]
- Gao, Z. Identification, expression and functional characterization of complement components C1q-like and C3a molecules in amphioxus. M.A. Thesis. Ocean University Of China. Qingdao. 2015.
- Nayak, A., Ferluga, J., & Tsolaki, A.G. The non-classical functions of the classical complement pathway recognition subcomponent C1q. Immunology Letters. 2010, 131(2), 139–150. [CrossRef]
- Nauta, A.J., Trouw, L.A., & Daha, M.R. Direct binding of C1q to apoptotic cells and cell blebs induces complement activation. European Journal of Immunology. 2002, 32(6), 1726–1736. [CrossRef]
- Korb, L.C., & Ahearn, J.M. C1q binds directly and specifically to surface blebs of apoptotic human keratinocytes: complement deficiency and systemic lupus erythematosus revisited. Journal of Immunology (Baltimore, Md.: 1950). 1997, 158(10), 4525–4528. [CrossRef] [PubMed]
- Young Jr, K.R., Ambrus Jr, J.L., & Malbran, A. Complement subcomponent C1q stimulates Ig production by human B lymphocytes. Journal of Immunology (Baltimore, Md.: 1950). 1991, 146(10), 3356–3364.
- Ferry, H., Potter, P.K., & Crockford, T.L. Increased positive selection of B1 cells and reduced B cell tolerance to intracellular antigens in c1q-deficient mice. The Journal of Immunology. 2007, 178(5), 2916–2922. [CrossRef]
- Kobayashi, H., Hirashima, Y., & Terao, T. Human myometrial cells in culture express specific binding sites for urinary trypsin inhibitor. Molecular Human Reproduction. 2000, 6(8), 735–742. [CrossRef]
- Bordin, S., Ghebrehiwet, B., & Page, R.C. Participation of C1q and its receptor in adherence of human diploid fibroblast. Journal of immunology. 1990, 145(8), 2520–2526. [CrossRef]
- Naito, A.T., Sumida, T., & Nomura, S. Complement C1q activates canonical Wnt signaling and promotes aging-related phenotypes. Cell. 2012, 149(6), 1298–1313. [CrossRef]
- Bossi, F., Tripodo, C., & Rizzi, L. C1q as a unique player in angiogenesis with therapeutic implication in wound healing. Proceedings of the National Academy of Sciences. 2014, 111(11), 4209–4214. [CrossRef]
- Tan, A., Ke, S.Y., & Chen, Y. Expression patterns of C1ql4 and its cell-adhesion GPCR Bai3 in the murine testis and functional roles in steroidogenesis. The FASEB Journal. 2019, 33(4), 4893–4906. [CrossRef]
- Siddiqui, N., & Straube, A. (2020). The Kinesin–3 Family: Long-Distance Transporters. The Kinesin Superfamily Handbook.
- Venkateswarlu, K., Hanada, T., & Chishti, A. H. Centaurin-α1 interacts directly with kinesin motor protein KIF13B. Journal of Cell Science. 2005, 118(11), 2471–2484. [CrossRef]
- Willemsen, M.H., Ba, W., & Wissink-Lindhout, W.M. Involvement of the kinesin family members KIF4A and KIF5C in intellectual disability and synaptic function. Journal of Medical Genetics. 2014, 51(7).
- Kaori, H., Yamada, Y., & Geyer. KIF13B regulates angiogenesis through Golgi to plasma membrane trafficking of VEGFR2. Journal of cell science. 2014, 127(20), 4518-4530.
- Amiri, M., Conserva, F., & Panayiotou, C. The human adenylate kinase 9 is a nucleoside mono-and diphosphate kinase. The international journal of biochemistry & cell biology. 2013, 45(5), 925–931. [CrossRef]
- Dzeja, P., & Terzic, A. Adenylate Kinase and AMP signaling networks: metabolic monitoring, signal communication and body energy sensing. International Journal of Molecular Sciences. 2009, 10(4), 1729–1772. [CrossRef]
- Yang, A.L. Effects of adenylate kinase ak4 knockout on germ cell apoptosis of male zebrafish. M.A. Thesis. ShanDong University. Shandong. 2017.
- Vogel, B.E., Muriel, J.M., & Dong, C. Hemicentins: what have we learned from worms. Cell Research. 2006, 16(11), 872–878. [CrossRef]
- Vogel, B.E, Hedgecock, E.M. Hemicentin, a conserved extracellular member of the immunoglobulin superfamily, organizes epithelial and other cell attachments into oriented line-shaped junctions. Development. 2001, 128(6), 883-894.
- Chowdhury, A., Herzog, C., & Hasselbach, L. Expression of fibulin-6 in failing hearts and its role for cardiac fibroblast migration. Cardiovasc Research. 2014, 103(4), 509–520. [CrossRef]
- Carney, T.J., Natália M.F., & Sonntag, C. (2010) Genetic Analysis of Fin Development in Zebrafish Identifies Furin and Hemicentin1 as Potential Novel Fraser Syndrome Disease Genes. PLoS Genetics. [CrossRef]
- Lin, M.H., Pope III, B.D., & Sasaki, T. Mammalian hemicentin 1 is assembled into tracks in the extracellular matrix of multiple tissues. Developmental Dynamics. 2020, 249(6), 775–788.
- Yang, L.D., Jiang, H.F., & He, S.P. Comparative genomics reveals accelerated evolution of fright reaction genes in ostariophysan fishes. Frontiers in Genetics. 2019, 10, 1283. [CrossRef]
Figure 1.
Schematic diagram of the experimental device (left) and operation diagram (right) used to measure the shell closure force and movement of M. yessoensis.
Figure 1.
Schematic diagram of the experimental device (left) and operation diagram (right) used to measure the shell closure force and movement of M. yessoensis.
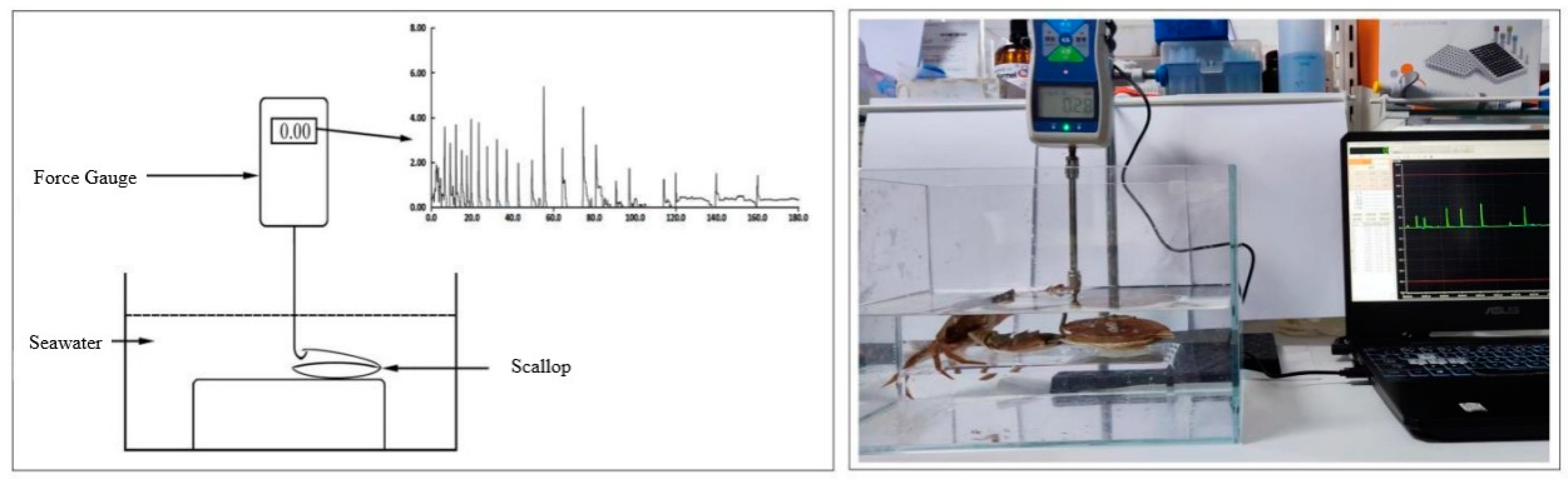
Figure 2.
M. yessoensis behavior in experiment II. X indicates C. japonica; lowercase s, m, and l represent small, medium, and large scallops, respectively, and the subscript 0 indicates the control group (no C. japonica).
Figure 2.
M. yessoensis behavior in experiment II. X indicates C. japonica; lowercase s, m, and l represent small, medium, and large scallops, respectively, and the subscript 0 indicates the control group (no C. japonica).

Figure 3.
Typical force recording during a movement response for different sizes of M. yessoensis. sB, mB, and lB indicate small, medium, and large scallops in the control group, respectively; X-sB, X-mB, and X-lB indicate small, medium, and large scallops in the crab-stimulated group, respectively.
Figure 3.
Typical force recording during a movement response for different sizes of M. yessoensis. sB, mB, and lB indicate small, medium, and large scallops in the control group, respectively; X-sB, X-mB, and X-lB indicate small, medium, and large scallops in the crab-stimulated group, respectively.

Figure 4.
Maximum contraction force and clap rate of adductor muscles of different sizes of M. yessoensis with and without exposure to the crab predator. Different uppercase letters indicate significant differences (P < 0.05) among different size groups without the presence of the predator. Different lowercase letters indicate significant differences (P < 0.05) among different size groups during crab stimulation. * indicates a significant difference (P < 0.05) between the same size scallops with and without crab stimulation.
Figure 4.
Maximum contraction force and clap rate of adductor muscles of different sizes of M. yessoensis with and without exposure to the crab predator. Different uppercase letters indicate significant differences (P < 0.05) among different size groups without the presence of the predator. Different lowercase letters indicate significant differences (P < 0.05) among different size groups during crab stimulation. * indicates a significant difference (P < 0.05) between the same size scallops with and without crab stimulation.
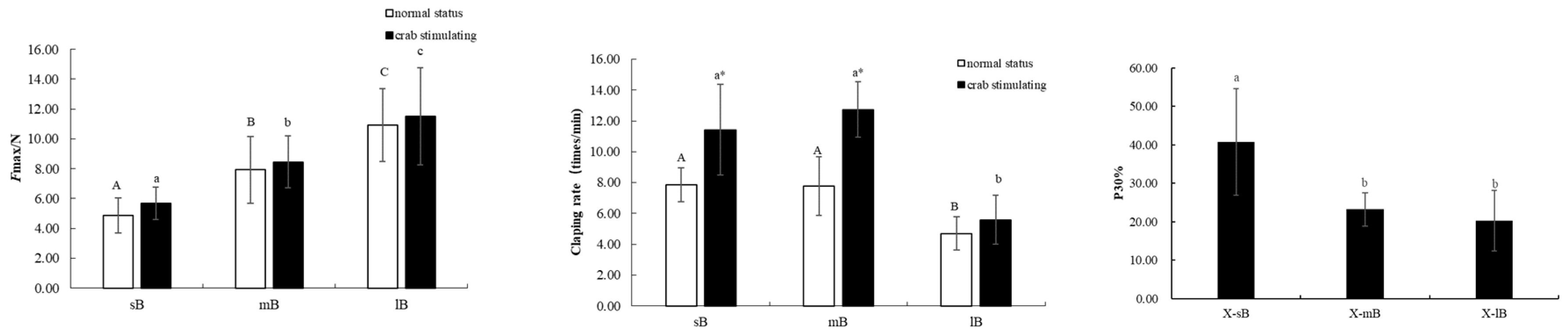
Figure 5.
Tonic, phasic, and total contraction forces of adductor muscles from different sized M. yessoensis with and without exposure to the crab predator.
Figure 5.
Tonic, phasic, and total contraction forces of adductor muscles from different sized M. yessoensis with and without exposure to the crab predator.
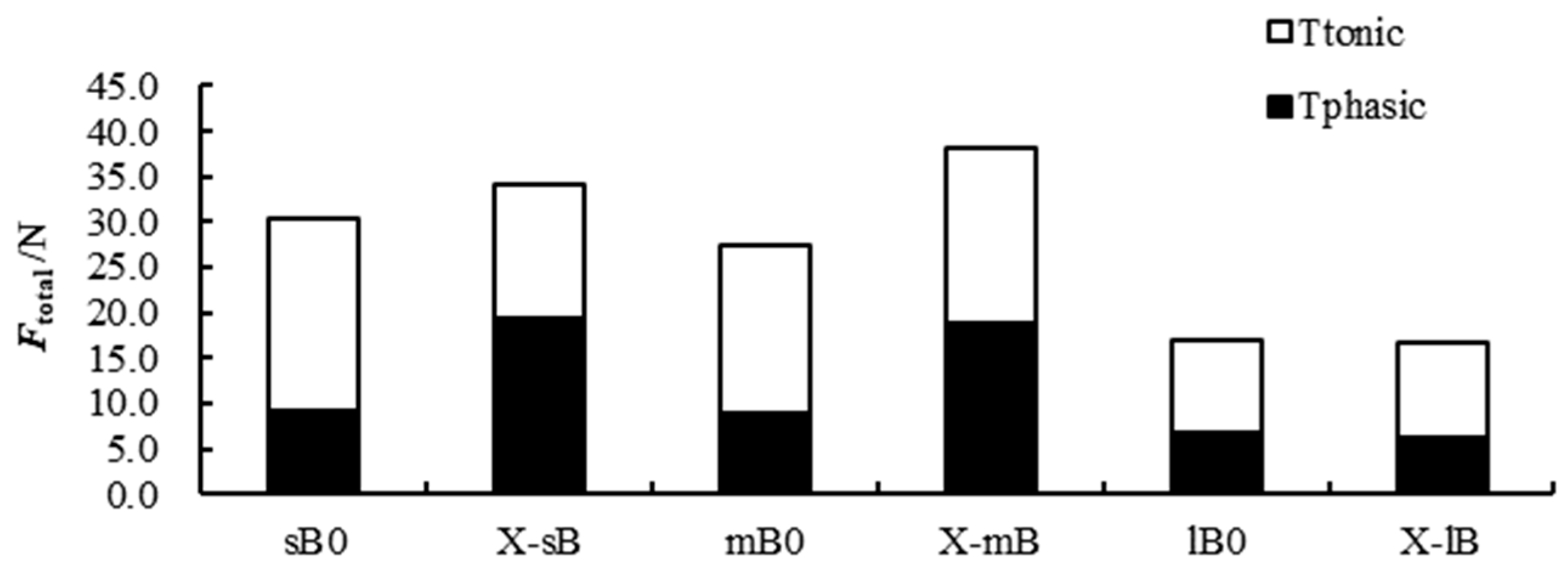
Figure 6.
Comparison of tissue enzyme viability in M. yessoensis tissues: (A) gill, (B) mantle, and (C) adductor muscle. Different uppercase letters indicate significant differences (P < 0.05) among different size groups without the presence of the predator. Different lowercase letters indicate significant differences (P < 0.05) among different size groups in the presence of crabs. “*”represents a significant difference (P < 0.05) between the same size in different stimulus states. “**” represents a highly significant difference (P < 0.01) between scallops of the same size with and without crab stimulation.
Figure 6.
Comparison of tissue enzyme viability in M. yessoensis tissues: (A) gill, (B) mantle, and (C) adductor muscle. Different uppercase letters indicate significant differences (P < 0.05) among different size groups without the presence of the predator. Different lowercase letters indicate significant differences (P < 0.05) among different size groups in the presence of crabs. “*”represents a significant difference (P < 0.05) between the same size in different stimulus states. “**” represents a highly significant difference (P < 0.01) between scallops of the same size with and without crab stimulation.
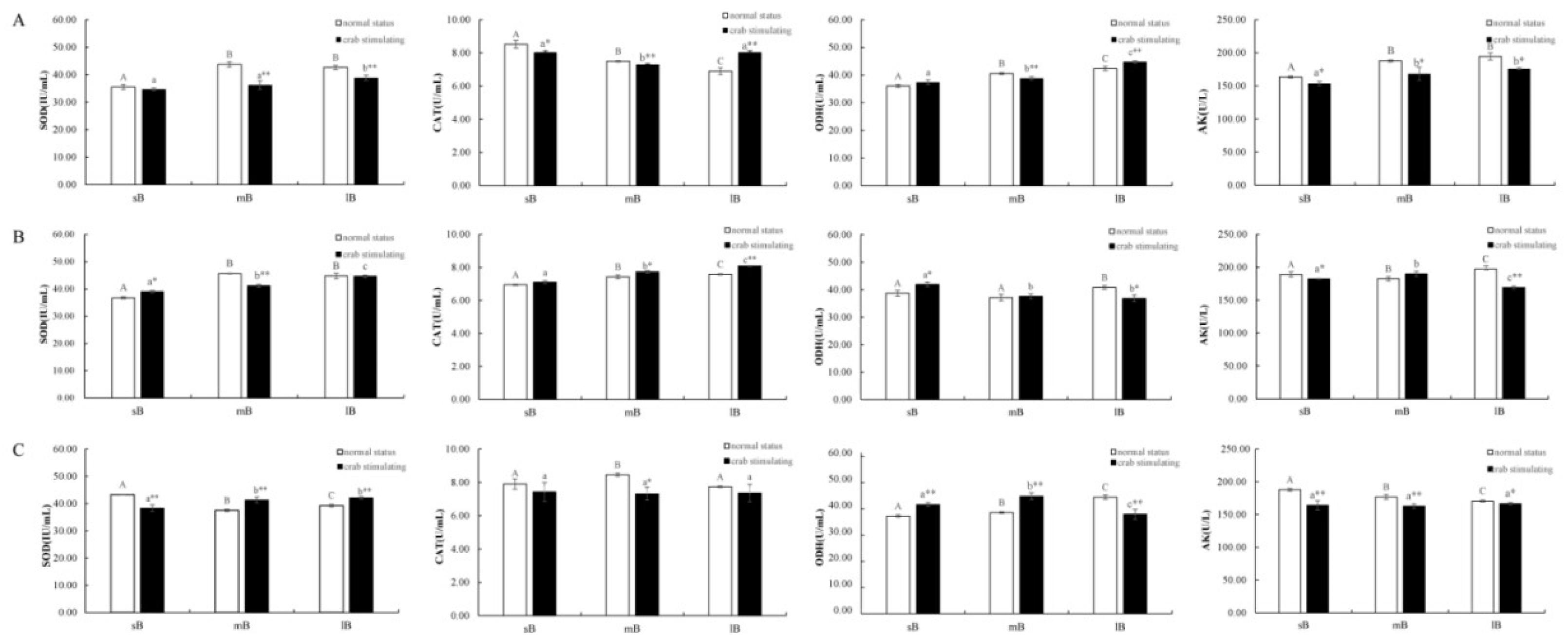
Figure 7.
Volcano map of DEGs.
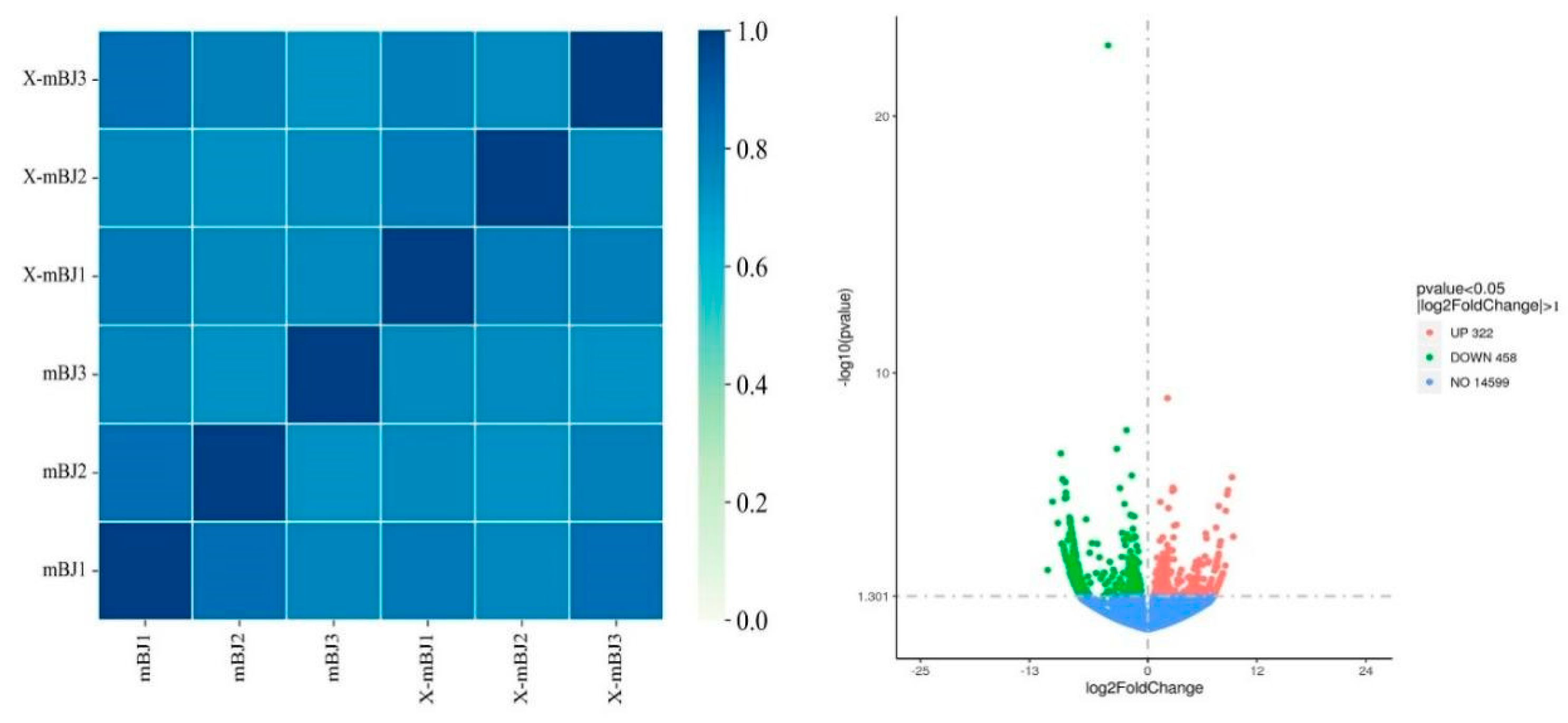
Figure 8.
GO analysis of DEGs.
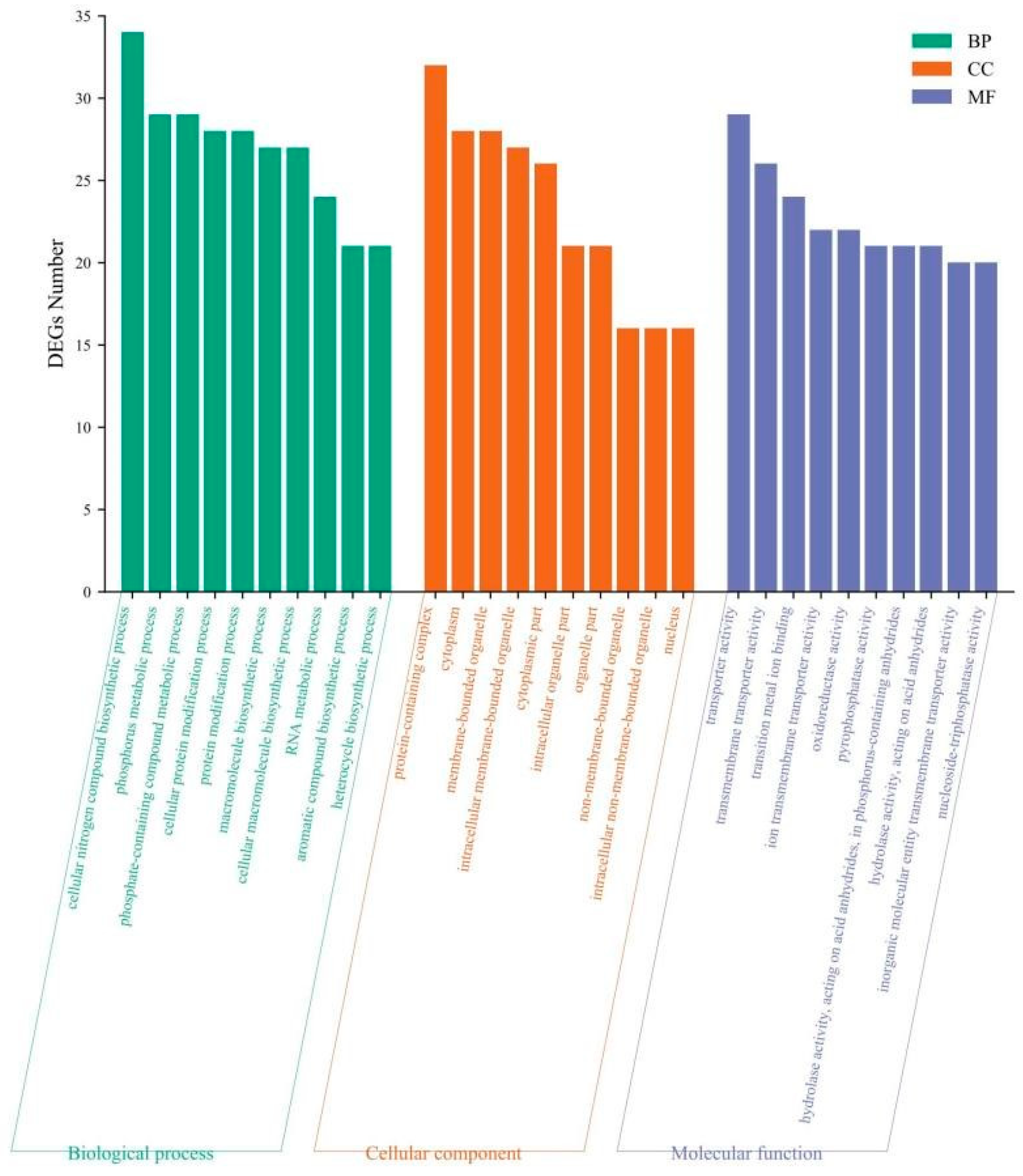
Figure 9.
KEGG bubble diagram
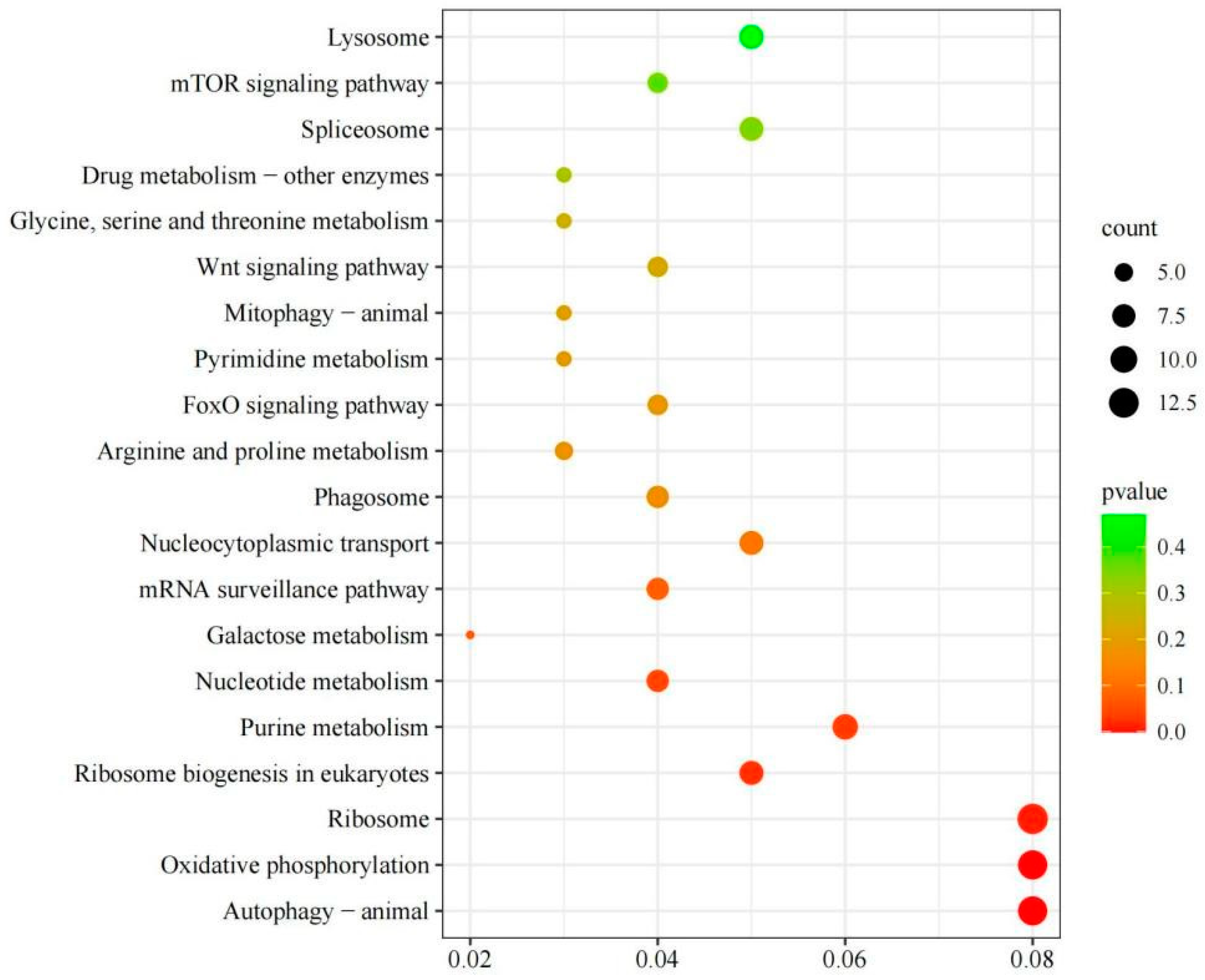
Figure 10.
Comparison of the RNA-Seq and qRT-PCR results
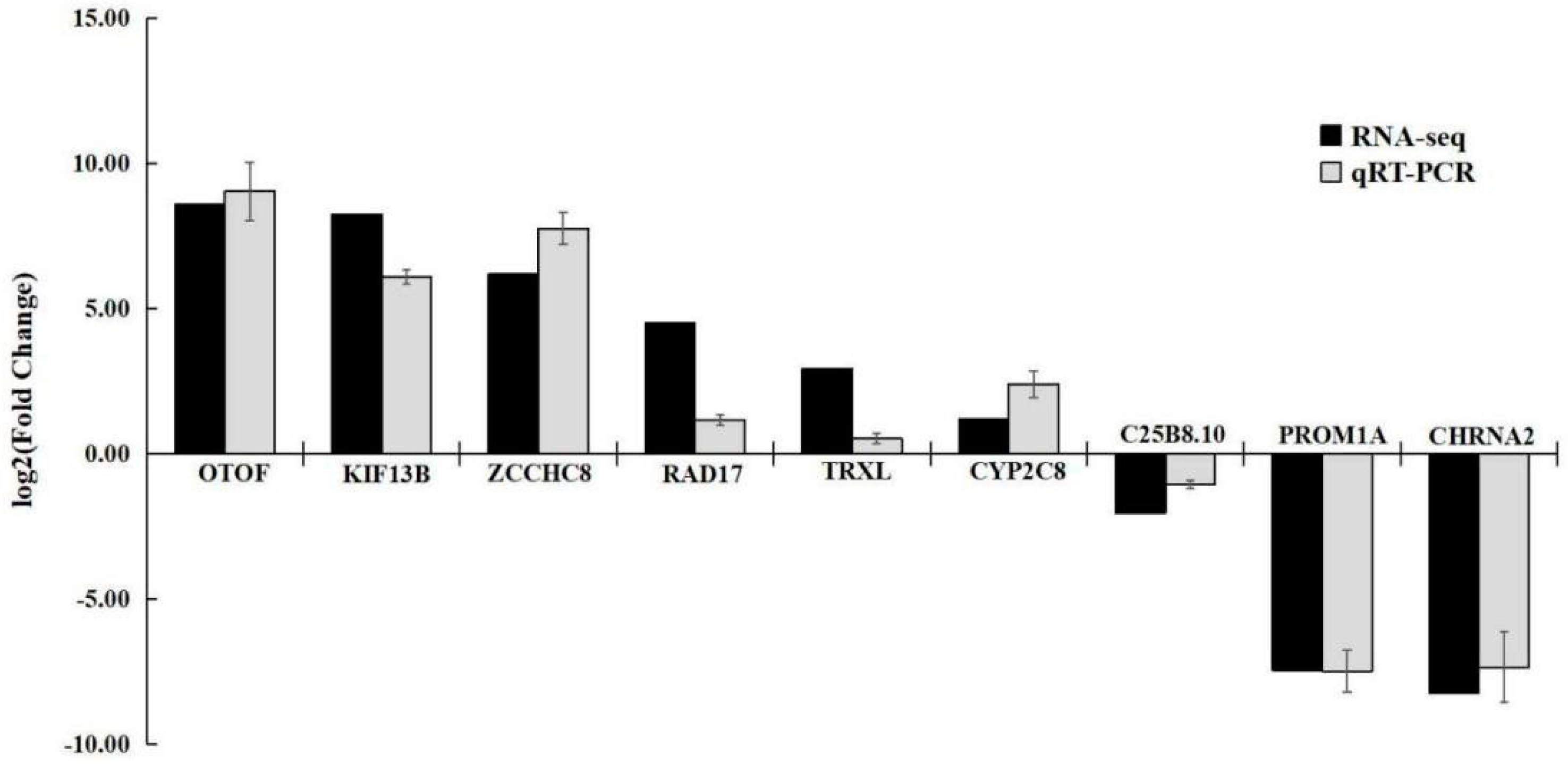

Table 1.
Specifications of the three size classes of M. yessoensis
| Large size (l) | Middle size (m) | Small size (s) | |
|---|---|---|---|
| Shell length/mm | 119.85±3.23 | 89.24±3.77 | 60.10±3.23 |
| Shell height/mm | 116.92±6.02 | 87.46±3.55 | 61.39±6.02 |
| Shell width/mm | 26.98±2.98 | 26.98±2.98 | 16.21±2.63 |
| Total wet weight/g | 176.50±28.57 | 83.70±13.97 | 30.16±5.29 |
Table 2.
Primers used for the qRT-PCR validation
| Gene name | Forword Primer (5’-3’) | Reverse Primer (5’-3’) |
|---|---|---|
| Gapdh | TGGTATGGCTTTCCGTGTGC | TCCTCTGTGTAACCAAGGAACC |
| KIF13B | GCAGCCAACCTCAGTCCTAACAG | TCGTGCTCGTCCTCTACCATCAT |
| CYP2C8 | GTTGCTCCTCTTGGCGTTCCT | GGCGACCGACAGAGAATGCT |
| ZCCHC8 | ACCACCACTGCCAATCAACACTC | CCATCACCTGTAGCTCCACCTCT |
| TRXL | TGTCTACAACACCCGCCAGAAT | ACACCACGAAGCATGGAAGTC |
| RAD17 | ACGAGTCGGAGTTGTGGTCTG | TGCCTGTGCCTTGAGATGTGT |
| OTOF | GTTGACGGACTCGGACGACATC | GCCTTCAGCACTCGCACAGT |
| C25B8.10 | GTTGAGCTTGGAGCTGGAACAG | GCCACCACAGTCCTAACAGAGT |
| CHRNA2 | GCCGTGCTCAGAATCCACAACT | TCCCGACGACACGCCACAATA |
| PROM1A | GGTTTGGCTTGGGATGGTGTCT | GCGTGGCTGACCTTGTTGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



