
Submitted:
15 August 2024
Posted:
16 August 2024
You are already at the latest version
Abstract
The role of astro- and microglial cells in the pathogenesis of epilepsy is currently under active in-vestigation. It is proposed that the activity of these cells may be regulated by agonists of peroxi-some proliferator-activated nuclear receptors (PPARs). The objective of this study was to investi-gate the effects of the PPAR β/δ agonist GW0742 (Fitorine) on the behavior and gene expression of astro- and microglial proteins involved in the regulation of epileptogenesis in rat brain in a lithi-um-pilocarpine model of temporal lobe epilepsy (TLE). A seven-day administration of GW0742 at a dose of 5 mg/kg/day has been shown to suppress TLE-related behavioral hyperactivity in the Open Field test and to attenuate the TLE-induced increase in gene expression and protein levels of astro- and microglial activation markers Gfap and Aif1 in the dorsal hippocampus and temporal cortex. Furthermore, GW0742 reversed the TLE-related increase in Lcn2 gene expression (a marker of the A1 phenotype of astrocytes) in the temporal cortex, the increase in the ratio of Nos2/Arg1 gene expression (markers of the M1/M2 phenotypes of microglia), and the decrease in Arg1 gene ex-pression in the dorsal hippocampus. The results suggest that GW0742 may mitigate behavioral and biochemical abnormalities in a lithium-pilocarpine model of TLE.
Keywords:
gene expression
; microglia
; astroglia
; neuroinflammation
; temporal lobe epilepsy
; PPAR β/δ ago-nists
; GW0742
; fitorine
1. Introduction
Despite the availability of a wide range of anticonvulsant drugs, up to 30% of patients with epilepsy remain refractory to treatment [1]. Accordingly, the identification of novel therapeutic agents is an urgent goal. In recent years, a considerable amount of research has been devoted to investigating the potential involvement of astro- and microglial cells in the pathogenesis of epilepsy. Neuroinflammation associated with increased astroglial and microglial cell activity plays an important role in the pathogenesis of epilepsy [2,3]. Pro-inflammatory proteins produced by glial cells, in particular interleukin-1β and tumor necrosis factor, have been shown to lower seizure thresholds in models of acute seizures [4,5], and to contribute to the development of chronic epileptic processes in the brain [6,7] and to the formation of epilepsy-related behavioral disorders [8].
Reactive astro- and microgliosis represents one of the most distinctive histopathological alterations associated with epilepsy [9]. However, the mere activation of glial cells does not yet indicate an increase in the production of pro-inflammatory proteins. Astrocytes and microglia can exist in different functional states, or phenotypes, which are associated with the production of neurotoxic (proepileptogenic) or neuroprotective (antiepileptogenic) proteins. In previous years, the classification of astrocytes and microglia was a widely used method of distinguishing between A1 and M1 phenotypes (pro-inflammatory) and A2 and M2 phenotypes (protective) [10,11]. At present, this classification is considered too simplistic, and its use is currently being debated [11,12]. However, the view that when examining the role of glial cells in various neuropathologies, it is essential to study the gene expression of both damaging and protective proteins remains unchanged [12].
Pharmacological agents that can inhibit pro-inflammatory processes while simultaneously activating the protective properties of glial cells represent a promising avenue of treatment for epilepsy [13]. In particular, peroxisome proliferator-activated receptor (PPAR) agonists exhibit these characteristics [11,14]. PPARs are nuclear receptor proteins that regulate the expression of genes essential for various metabolic processes, cell differentiation, and neuroinflammation [15]. There are three types of PPARs: α, β/δ, and γ. All of these receptor types are targets for free fatty acids and selected products of lipid metabolism, particularly eicosanoids [16]. Nevertheless, specific ligands have been identified for distinct subtypes of PPARs. For instance, PPARγ is activated by the prostaglandin PGJ2 [17] while PPARα is activated by leukotriene B4 [18]. Additionally, numerous fatty acids, particularly arachidonic acid derivatives, can bind to PPARβ/δ. However, the specific fatty acids that act as endogenous ligands for PPARβ/δ remain unresolved [19].
All types of PPARs are expressed in various cells throughout the body, including those in the brain. They are found in neurons, oligodendrocytes, astrocytes, as well as in relatively smaller amounts in microglia [20,21,22]. Additionally, distinct patterns of distribution are observed for the three types of PPARs within different regions of the central nervous system (CNS) and among various cell types within the CNS [21].
The anti-inflammatory properties characteristic of agonists of all types of PPARs are realized through negative regulation of NF-κB, AP-1, and C/EBP-mediated signaling pathways [23]. Consequently, the effects of PPAR agonists are manifested by the suppression of oxidative stress, a reduction in the production of inducible NO synthase and pro-inflammatory cytokines, and an increase in the expression of anti-inflammatory genes [23]. The neuroprotective properties of synthetic PPAR agonists have been demonstrated in models of diverse neuropathologies, including epilepsy [24,25]. Meanwhile, in models of epilepsy and acute seizures, studies have primarily concentrated on examining the protective properties of PPARγ agonists [26,27,28,29]. There have been few studies conducted using PPARα agonists [30].The effects of PPAR β/δ agonists have yet to be sufficiently studied.
The objective of this study was to examine the impact of the selective PPAR β/δ agonist GW0742 on the behavior and gene expression of astro- and microglial proteins associated with epileptogenesis in the rat brain, using the lithium-pilocarpine model of temporal lobe epilepsy. GW0742, also known as GW610742 and Fitorine, is a selective PPAR β/δ agonist [31]. It is used in sports nutrition as a performance-enhancing, fat-burning drug. A number of studies have demonstrated the drug's broad spectrum of action, including its ability to inhibit the interaction between the vitamin D receptor and steroid receptor coactivator 2 [32]. Additionally, GW0742 has been identified as a potential anti-diabetic drug [33]. GW0742 has been demonstrated to possess a marked anti-inflammatory effect, as evidenced by a number of research findings [34,35], including studies conducted in models of neuropathology [36]. The neuroprotective effects of GW0742 have been demonstrated in models of neurodegenerative disease [37,38]. Nevertheless, the effects of GW0742 in models of epilepsy and seizures remain poorly understood.
2. Results
2.1. Survival and Weight Trend Analysis
The studies were conducted using a lithium-pilocarpine model of TLE on male Wistar rats aged 7–8 weeks. GW0742 (5 mg/kg) was administered once daily for a week following the induction of status epilepticus with pilocarpine. The mortality rates observed in the initial period following the induction of status epilepticus with pilocarpine were 8% in the control group of rats and 33% in the GW0742-treated group. Nevertheless, a comparison of the survival curves (log-rank test, Figure 1a) reveals no statistically significant differences between the groups (χ2 = 2.47; p = 0.16). These results are consistent with those previously reported for this model. Mortality in this model is typically 20–30% of rats [6].
The initial weight of the animals did not differ significantly between groups. Following the administration of pilocarpine, a decrease was observed, with a reduction of 12–24% (Figure 1b, two-way ANOVA, TLE factor: F(1,35) = 118; p < 0.001;). GW0742 had no effect on the weight dynamics of control and TLE rats (treatment factor: F(1,35) = 0.97; p = 0.33).
2.2. Behavior
The results of the social interaction test (Figure 1c) revealed a significant reduction in communicative activity in TLE rats, with a 6-fold decrease observed (two-way ANOVA, TLE factor – F(1, 32) = 84; p < 0.001). Furthermore, the TLE rats exhibited increased anxiety in the open field test, evidenced by an elevated duration of grooming behavior (Figure 1d; F(1, 34) = 7.7; p < 0.01). GW0742 had no effect on these changes.
Hyperactivity represents one of the most salient behavioral disturbances that is consistently observed in the lithium-pilocarpine model [6,39]. Indeed, TLE rats exhibited increased locomotor activity (Figure 1d-g), as evidenced by elevated locomotion time and length of distance traveled in the Open Field test (TLE factor: F(1, 35) = 11.8; p < 0.01 (locomotion time) and F(1, 34) = 13.3; p < 0.001 (distance traveled)). The administration of GW0742 was shown to prevent the development of these impairments. A post hoc analysis employing the Sidak test demonstrated that the only statistically significant differences were between the untreated control and TLE groups (p < 0.01).
The results demonstrated that GW0742 is effective in mitigating specific pathological processes that occur during the development of TLE. Accordingly, we proceeded to the second stage of the study, which entailed an examination of the impact of GW0742 on the gene expression of astro- and microglial proteins that are implicated in epileptogenesis regulation.
2.3. Gene Expression Analysis of Glial Protein Genes at the mRNA and Protein Level
The analysis of gene expression of markers of astrocyte (Gfap) and microglia (Aif1) activation in treated and untreated TLE rats at the mRNA level (Figure 2) revealed a notable elevation in both markers in the dorsal hippocampus (TLE factor: Gfap – F(1, 20) = 62, p < 0.001; Aif1 – F(1, 24) = 47, p < 0.001) and temporal cortex (TLE factor: Gfap – F(1, 20) = 280, p < 0.001; Aif1 – F(1, 20) = 75, p < 0.001) of TLE rats. GW0742 exhibited a modest inhibitory effect on Gfap gene expression in the dorsal hippocampus (treatment factor: F(1, 20) = 5.4, p = 0.03) and Aif1 gene expression in the temporal cortex (treatment factor: F(1, 20) = 7.3, p = 0.01).
Analysis of the expression of the same genes at the protein level also reveals an increase in GFAP production (Figure 3, TLE factor: dorsal hippocampus – F(1, 19) = 26; p < 0.0001; temporal cortex – F(1, 20) = 5.3; p = 0.03) and IBA1 (Figure 4, TLE factor: dorsal hippocampus – F(1, 19) = 57; p < 0.0001; temporal cortex – F(1, 20) = 16.1, p < 0.001).
Treatment with GW0742 resulted in a significant normalization of expression levels of GFAP. Posterior comparisons of GFAP protein levels reveal significant differences between the untreated control group and the TLE group, as evidenced by Sidak's test (Sidak's test, p < 0.001). However, no such differences were identified in the treated groups (p = 0.21). The analysis revealed a significant effect of GW0742 treatment on IBA1 protein levels in the dorsal hippocampus (F(1, 19) = 5.3; p = 0.03). A posteriori comparisons of the temporal cortex revealed that the untreated TLE group exhibited enhanced IBA1 production compared to the control group (Sidak's test, p = 0.01). However, no such differences were observed in the treated animal groups (p = 0.13). These findings suggest that GW0742 attenuates, though does not fully block, the micro- and astroglial cell activation associated with epileptogenesis.
Given that the activation of astro- and microglial cells does not provide sufficient evidence to determine whether pro- or anti-inflammatory protein gene expression is predominant, we conducted further analysis of the gene expression of pro- and anti-inflammatory cytokines, which are characteristic of both astrocytes and microglia. Additionally, we examined the gene expression of pro-inflammatory and protective proteins that are specific to microglia or astrocytes.
The induction of TLE resulted in an increased expression of pro-inflammatory genes. Tnfa was upregulated in the temporal cortex (TLE factor: F(1, 19) = 12.4; p = 0.002) but not in the dorsal hippocampus (Figure 5a and 5b). There was a significant increase in Il1b levels in both the dorsal hippocampus and temporal cortex (F(1, 22) = 10.7; p < 0.01 and F(1, 21) = 6.73; p = 0.02; Figure 5c and 5d). The direction and severity of these changes were similar in GW0742-treated and untreated rats. Two-way ANOVA revealed no significant effect of GW0742 administration on Tnfa and Il1b expression.
Additionally, there was a significant increase in the expression of the anti-inflammatory cytokine Il1rn (TLE factor: dorsal hippocampus - F(1, 22) = 151; p < 0.001; temporal cortex - F(1, 21) = 68; p < 0.001). Furthermore, the ratio of Il1b and Il1rn gene expression decreased (dorsal hippocampus – F(1, 23) = 34; p < 0.001; temporal cortex – F(1, 23) = 26; p < 0.001). This indicates that during the latent period of the lithium-pilocarpine model, compensatory mechanisms against pathological processes remained highly active. Treatment with GW0742 resulted in a decrease in these changes in the temporal cortex (treatment factor for Il1rn: F(1, 21) = 6.0; p = 0.02; for Il1b/Il1rn – F(1, 23) = 5.8; p = 0.02). This may be attributed to the reduced severity of pathological changes observed in treated rats, which suggests that the activation of compensatory mechanisms was less pronounced.
The NOD-like receptor protein 3 (NLRP3), a basic inflammasome protein, has been identified as another protein associated with the triggering of pro-inflammatory signaling pathways [40]. The primary sources of NLRP3 in the brain are microglial cells [41]. The expression of the Nlrp3 gene was observed to increase by more than twofold in both the dorsal hippocampus (TLE factor: F(1, 24) = 44, p < 0.001) and the temporal cortex (F(1, 21) = 88, p < 0.001) (Figure 6a and 6b). Furthermore, in the temporal cortex, GW0742 treatment resulted in a reduction in Nlrp3 gene expression (treatment factor: F(1, 21) = 5.1, p = 0.03), although post hoc comparisons did not yield statistically significant differences.
Subsequently, the gene expression of two microglial proteins was examined, namely the inducible NO synthase (Nos2 gene, Figures 6c and 6d) and the arginase enzyme (Arg1 gene, Figures 6e and 6f). It is established that in macrophages and microglia, these proteins compete for the substrate L-arginine, which can activate intracellular cascades associated with the production of damaging (Nos2) or protective (Arg1) factors, respectively [42]. Consequently, an additional analysis was conducted on the Nos2/Arg1 ratio as an indicator of the equilibrium between damaging and neuroprotective mechanisms (Figure 6g and 6h).
No significant alterations in Nos2 gene expression were observed in the studied brain structures. In the dorsal hippocampus, Arg1 gene expression was significantly decreased (TLE factor: F(1, 21) = 12.0, p = 0.002) and the Nos2/Arg1 ratio increased (TLE factor: F(1, 24) = 5.7; p = 0.02). In both cases, post hoc comparisons reveal significant changes only in untreated rats.
Next, we analyzed the expression of two astrocytic protein genes: lipocalin (Lcn2 gene, Figure 7a and 7b), which plays an important role in the induction of neuronal death in inflammatory and pathological conditions of the CNS [43,44] and a marker of the protective phenotype of astroglia calcium-binding protein S100A10 (S100a10 gene, Figure 7c and 7d). Lcn2 gene expression was increased in the dorsal hippocampus (TLE factor: F(1, 23) = 8.417; p < 0.01) and in the temporal cortex (TLE factor: F(1, 20)= 11.07; p < 0.01). Nevertheless, post hoc comparisons indicate a statistically significant increase exclusively in the temporal cortex of untreated TLE rats. The expression of the S100a10 gene was increased in the temporal cortex of both treated and untreated TLE animals (F(1, 21) = 34; p < 0.001).
In addition, the expression of three astroglial protein genes that may be associated with epileptogenesis were examined: guanylate binding protein 2 (Gbp2 gene) [45,46] pentraxin-3 (Ptx3) [46,47], and the glutamate transporter EAAT2 (Slc1a2) [48,49]. No significant differences were observed between the groups regarding these genes (Figure 8).
3. Discussion
In this study, we investigated the effects of the PPARβ/δ agonist GW0742 on the progression of epileptogenesis in a rat lithium-pilocarpine model of TLE. Treatment attenuated hyperactivity in rats that is a characteristic of the lithium-pilocarpine model of TLE, and diminished activation of astrocytes and microglial cells, which was revealed by analyzing the expression level of a number of glial genes. At the same time, administration of GW0742 to control animals did not affect behavior of rats or gene expression.
PPARβ/δ agonists, and GW0742 as one such agonist, have been demonstrated to possess pronounced neuroprotective and anti-inflammatory properties [22,36]. Given the pivotal role of neuroinflammation in the mechanisms of epileptogenesis [50], PPAR β/δ agonists may emerge as a valuable therapeutic tool for the treatment and prevention of epilepsy. Moreover, PPAR β/δ agonists have been demonstrated to exert a significant influence on oxidative stress [51,52], lipid and carbohydrate metabolism[53]. These effects may potentially contribute to the inhibition of epileptogenesis [54,55,56]. For example, our recent findings demonstrated that cardarin, a PPARβ/δ agonist, effectively mitigates the behavioral disturbances that emerge during the chronic phase of the TLE model [57].
The effects of the PPAR β/δ agonist GW0742 were examined in the latent phase of the TLE model. One week following pilocarpine administration, rats exhibited hyperactivity in the open field test and a significant reduction in communicative behavior in the social test. These behavioral changes are characteristic of the model used and have been consistently reproduced in our studies [6,58,59] and in other laboratories [60,61,62]. The behavioral abnormalities observed in the lithium-pilocarpine model may be specifically related to neuroinflammation, as evidenced by the attenuation of these abnormalities by anti-inflammatory therapy [6]. The present study demonstrates that administration of GW0742 attenuates hyperactivity.
The neuroprotective effects of GW0742 and other PPAR β/δ agonists are thought to be related to their effects on astroglial and microglial cells [63,64,65]. The present study demonstrates that the utilized model of TLE activates glial cells. During the latent phase of the lithium-pilocarpine model, we observed that Gfap and Aif1 are upregulated at both the mRNA and protein levels in the dorsal hippocampus and temporal cortex. The upregulation of these genes has been identified as an indicator of astrogliosis and microgliosis and has been demonstrated in numerous models of seizures and epilepsy [66,67,68,69,70].
The colony-stimulating factor 1 receptor (CSF1R)-related pathway has been identified as a key regulator of microglial activation [67], while astroglial activation has been linked to an increase in JAK2 tyrosine kinase and STAT3 protein production [66]. The latter pathway is subject to regulation by PPARs [71]. This study is the first to demonstrate that GW0742 treatment can attenuate the increased expression of Gfap and Aif1 genes at the mRNA and protein levels in a TLE model. Notably, similar results have been reported in the treatment of epilepsy with a ketogenic diet [72], the effects of which are presumably mediated by PPARs [73].
The impact of GW0742 on Gfap and Aif1 gene expression is of functional significance, as microglia and astrocyte activation contribute to epileptic processes [74]. One of the primary reasons for this is the close relationship between glial activation and the development of neuroinflammation [67], which plays a pivotal role in the pathogenic mechanisms of epileptogenesis [50]. The elevated gene expression of pro-inflammatory proteins, including Il1b, Tnfa, and Nlrp3, in the brains of rats with TLE, as observed in this study, is in accordance with the findings of previous studies conducted by our research group and other researchers [39,75,76]. It is noteworthy that in the latent phase of the lithium-pilocarpine model, the expression of not only pro-inflammatory protein genes but also Il1rn, the gene encoding the anti-inflammatory cytokine was increased. Furthermore, the production of Il1rn mRNA was more prominent than that of Il1b, as indicated by a reduction in the Il1b/Il1rn ratio. GW0742 diminished the elevated Il1rn gene expression and restored equilibrium in the Il1b/Il1rn ratio. It is plausible that the augmentation of Il1rn expression is contingent upon elevated glial cell activation, and this was mitigated following GW0742 administration.
Furthermore, we investigated the gene expression of several proteins that are traditionally regarded as markers of polar (pro-inflammatory or protective) functional states of microglia and astrocytes. The classification of microglia and astrocytes into M1/M2 and A1/A2 phenotypes has been the subject of criticism, as these states are rarely observed in pure form [12]. Nevertheless, even outside this classification framework, the genes under consideration are of interest for research purposes, as they play a role in regulating damaging or neuroprotective processes during epileptogenesis.
Our findings demonstrate that a TLE-induced increase in the expression of the lipocalin-2 gene (Lcn2, a marker of A1 astrocytes) occurs in the rat brain. Lipocalin-2 is an iron transport protein that contributes to oxidative stress and inflammation [77]. Elevated levels of lipocalin-2 are considered a biomarker of brain injury [78,79]. Previous studies have demonstrated a significant elevation in lipocalin-2 levels in the hippocampus of wild-type mice subjected to a kainate-induced seizure model [77]. Conversely, lipocalin-2 deficiency in knockout mice has been shown to mitigate kainate-induced iron overload and oxidative stress in hippocampal cells [77]. Our findings indicate that GW0742 exerts a protective effect, as evidenced by the high levels of Lcn2 gene expression observed exclusively in untreated TLE rats.
We also analyzed the gene expression of the A2 phenotype astrocyte marker gene S100a10; it was increased in both treated and untreated rats in the hippocampus and temporal cortex during the latent phase of epileptogenesis. S100A10 is a polyfunctional protein; in complex with annexin A2, it is involved in the organization of lipid microdomains on the cell membrane, binding of actin filaments and cytoskeleton scaffolds, in membrane transport and fibrinolysis [80,81,82], and in the regulation of the activity of some ion channels [83]. The neuroprotective role of S100A10 in epilepsy may be related, in particular, to its effect on the serotoninergic system [83], which plays a protective role in epileptogenesis [84]. Previously, enhanced expression of the S100a10 gene in the hippocampus was shown in rats in a kainate model of TLE [85]. It is likely that increased expression of the S100a10 gene is associated with the triggering of neuroprotective mechanisms at the initial stages of epileptogenesis.
We also found decreased Arg1 gene expression (a marker of M2 microglia) and increased ratio of the M1 marker Nos2 to Arg1 in the hippocampus of untreated TLE rats. It is known that macrophages and related microglial cells are characterized by alternative pathways of arginine metabolism via inducible NO synthase or arginase with different functional consequences [42]. The first pathway leads to NO synthesis, cell death due to oxidative stress, impaired energy metabolism, DNA damage and other negative consequences [86]. The second pathway is associated with increased synthesis of ornithine decarboxylase and consequent attenuation of inflammation [87]. GW0742 offset the TLE-induced increase in the Nos2/Arg1 ratio, which may indicate its neuroprotective effect.
The expression of glutamate transporter genes EAAT2 (gene is Slc1a2), astroglia marker A1 guanylate-binding protein 2 (Gbp2), and astroglia marker A2 pentraxin-3 (Ptx3) was not altered in rat brain during the latent phase of the TLE model in this experiment. For Slc1a2 and Ptx3 gene, similar results were obtained in our previous studies [6,88]. Increased Gbp2 gene expression in rat temporal cortex was previously detected in the chronic phase of the lithium-pilocarpine model [88]. GW0742 did not affect the expression of the listed genes.
The present study investigated the effects of GW0742 in a lithium-pilocarpine rat model, demonstrating a spectrum of protective properties for GW0742. This evidence suggests that PPARβ/δ agonists may represent a novel and promising approach for the prevention of epileptogenesis and the treatment of temporal lobe epilepsy.
4. Materials and Methods
4.1. Experimental Design
The experimental design is illustrated in Figure 9. The studies were conducted on male Wistar rats aged 7–8 weeks. The lithium-pilocarpine model was utilized for the purpose of modeling TLE. This experimental model accurately reproduces the pathophysiological, histopathological, and biochemical processes associated with the development of human TLE [83]. The study was conducted during the initial period of epileptogenesis, which is the latent period of the model and coincides with the absence of spontaneous recurrent seizures.
To induce convulsions, rats were intraperitoneally (i.p.) administered 127 mg/kg of LiCl (Sigma-Aldrich, St. Louis, MO, USA). On the subsequent day, pilocarpine (Sigma-Aldrich) was administered. One hour prior to administration, an injection of (-) scopolamine methyl bromide (1 mg/kg, i.p.; Sigma-Aldrich) was administered to prevent the peripheral effects of pilocarpine. The dosage of pilocarpine was selected on an individual basis, based on the observed reaction of the rats to its administration. Pilocarpine was initially administered at a dose of 10 mg/kg (i.p.), with subsequent doses administered at 30-minute intervals, until the development of stage 4 seizures, as defined by the Racine scale [89]. The requisite dose of pilocarpine for the induction of such convulsions was observed to be within the range of 20 to 40 mg/kg. It has been demonstrated that fractional administration of pilocarpine enhances animal survival, with the formation of temporal lobe epilepsy occurring in all rats that received the indicated dose range of pilocarpine [6]. Rats that did not develop seizures after the fourth injection (40 mg/kg) were excluded from the study, as this constituted no more than 5% of the total number of animals. Seizures were terminated 90 min after the onset of stage 4 by the administration of diazepam (10 mg/kg, i.p., Sigma-Aldrich). A total of four groups were established: 1) untreated control (Ctrl+Veh; n = 9); 2) GW0742 treated control (Ctrl+GW0742; n = 8); 3) untreated TLE rats (TLE+Veh; n = 13; 12 survived); 4) GW0742 treated TLE rats (TLE+GW0742; n = 15; 10 survived). In order to ensure an equal distribution of animals into the different groups, the dose of pilocarpine administered was taken into consideration. This ensured that the number of animals in the treated and untreated groups with TLE was balanced. However, the distribution of animals into groups was otherwise random.
GW0742 (5 mg/kg, i.p., Clearsynth, Mumbai, India) was administered in a single daily dose for a period of one week following the induction of status epilepticus by pilocarpine. The initial injection was administered 24 hours following pilocarpine administration. Dimethyl sulfoxide (DMSO) was utilized as the solvent, given that GW0742 is essentially insoluble in water. The control animals were administered DMSO. The dosage of the drug was selected based on its demonstrated anti-inflammatory effects in an Alzheimer's disease model [90].
The weight and survival of the animals were assessed during the whole period of drug administration. To increase the survival rate of TLE rats, in the first days after pilocarpine-induced epileptic status they were fed with water from a pipette and wet food (porridge, fruit).
4.2. Behavioral Tests
Behavioral testing was conducted on the 7th and 8th days following pilocarpine administration, utilizing the open field and social tests (Figure 9).
The open field test was conducted in a circular arena with a diameter of 1 m and walls 30 cm in height. The illumination of the experimental setup was 8 lux. The rat was positioned in the center of the field and its behavior was recorded for a period of five minutes via video. Subsequently, the behavior was analyzed using the software Tracking and Field V4.0 (Institute of Experimental Medicine, St. Petersburg, Russia). Locomotor activity (quantified in terms of distance traveled and time spent in motion) and an indicator of anxiety (measured in terms of the duration of grooming behavior) were evaluated.
The social interaction test was conducted in transparent Plexiglas cages measuring 60×30×40 cm. The male rat was placed in the cage setup one day prior to the test to allow for adaptation and space exploration. Another adult male rat was then placed into the experimental cage for a five-minute observation period. During this time, the cage host's communicative (sniffing and grooming of the partner) and aggressive behaviors were evaluated.
4.3. Real-Time RT-PCR
Following the completion of behavioral testing, the animals were euthanized, their brains were removed, and stored at -80°C until the subsequent biochemical studies could be conducted. The temporal cortex and dorsal hippocampus (Figure 9) were isolated from the brain using a microtome-cryostat (OTF5000, Bright Instrument, Luton, UK). The required brain regions were identified using a rat brain atlas [91]. The temporal cortex and dorsal hippocampus were selected for study due to their critical involvement in the pathogenesis of epilepsy [92].
Total RNA was isolated using ExtractRNA reagent (Evrogen, Moscow, Russia) according to the manufacturer's recommendations. RNA concentration and purity were measured on a NanoDrop™ Lite spectrophotometer (Thermo Fisher Scientific, Waltham, Massachusetts, USA), by absorbance at 260 nm and 260/280 nm ratio, respectively.
cDNA synthesis was performed using 1 μg of total RNA, oligo-dT (0.5 μg per 1 μg RNA) and 9-mer random primers (0.25 μg per 1 μg RNA, DNA Synthesis LLC, Moscow, Russia) and 100 units of M-MLV reverse transcriptase (Evrogen). The reaction was performed in a total volume of 20 μl, and the procedure was according to the recommendations of the reverse transcriptase manufacturers. Before further PCR, the obtained cDNA was diluted 10-fold.
Next, we analyzed gene expression of proteins that may influence the processes of epileptogenesis: markers of astroglial (Gfap) and microglial (Aif1) cell activation; pro-inflammatory and anti-inflammatory proteins (Nlrp3, Il1b, Tnfa, Il1rn); markers of pro-inflammatory and anti-inflammatory phenotypes of microglia (M1 – Nos2; M2 – Tgfb1, Arg1); markers of pro-inflammatory and neuroprotective phenotype of astroglia (A1 – Ptx3, Lcn2; A2 – Gbp2, S100a10) neurotrophic and growth factors Fgf2, Bdnf, Gdnf, glutamate transporter Slc1a2. The expression of all the above genes was analyzed by real-time RT-PCR.
Real-time PCR (TaqMan technology) was performed on a C1000 Touch thermal cycler with a CFX384 Touch™ detector (Bio-Rad, Hercules, CA, USA). The reaction was performed in a total volume of 6 μl with 0.8 μl of cDNA, 0.5 units of TaqM-polymerase (Alkor Bio, St. Petersburg, Russia), 3.5 mM MgCl2 and specific forward and reverse primers and probes (see Appendix A Table A1) synthesized by DNA-Synthesis LLC. Samples were analyzed in 4 repeats.
The relative expression of the genes of interest was calculated using the method of 2-∆∆Ct [93]. Normalization was performed using the geometric mean of the expression of the three most stable reference genes selected from 9 reference genes (Gapdh, Actb, Rpl13a, B2m, Pgk1, Ppia, Hprt1, Ywhaz, Sdha). Reference genes were selected based on the ranking obtained using the online tool RefFinder® (http://blooge.cn/RefFinder/ accessed on 10 May 2024) according to the procedure described previously [94]. In the conducted study, the following reference genes were used for data normalization: Actb, Rpl13a, and Pgk1 for dorsal hippocampus; Sdha, Gapdh, and Ppia for temporal cortex.
4.4. Western Blot Analysis
The isolated brain structures were homogenized on ice in an optimized lysis buffer proposed [95] containing 100 mM Tris-HCl pH 8.0, 140 mM NaCl, 20 mM EDTA, 5% dodecylс sodium sulfate, 1X protease inhibitor cocktail (Pierce Protease Inhibitor Tablets, ThermoFisher Scientific), 1 mM sodium orthovanadate, 20 mM sodium fluoride. The homogenate was incubated for one hour at room temperature with constant stirring. The samples were then centrifuged (15 min, 14000 g, 20°C), and the supernatant was used for protein concentration quantification and Western blotting. Protein concentration was determined by the Lowry method as modified by Hartree [96]. The supernatant was diluted 1:1 with 2X application buffer (125 mM Tris-HCl pH 6.8, 40% glycerol, 4% sodium dodecyl sulfate, 5% β-mercaptoethanol, 0.02% bromphenol blue) and incubated for 15 min at 70°C, then stored at -20°C until electrophoresis.
Prior to electrophoresis, protein concentration was equalized by diluting the samples with 1X plating buffer.
Electrophoretic separation was performed under reducing and denaturing conditions [97] in a 13.5% polyacrylamide gel together with a molecular weight standard (Thermo Scientific PageRuler Prestained Protein Ladder 10-170 kDA; ThermoFisher Scientific) at an amperage of 125 V. 12 μg of protein was taken for plating because this concentration allowed us to fall within the linear region of the densitometric analysis for all antibodies used under our conditions. Together with the analyzed samples, a calibrator sample obtained by mixing several samples from animals from different groups was applied to each gel.
Proteins were transferred onto nitrocellulose membrane (0.2 μm pore diameter) by semi-dry transfer with Invitrogen Power Blotter 1-Step Transfer Buffer (ThermoFisher Scientific) according to the manufacturer's instructions, for 13 minutes at a current of 2.5A. After transfer, the membrane was stained with a 0.1% solution of Ponseau S dye in 5% acetic acid and documented using a ChemiDoc MP gel imaging system (Bio-Rad). The membrane was then washed with phosphate buffered saline (0.01 M phosphate buffer pH 7.4, 137 mM NaCl, 2.7 mM KCl) containing 0.1% Tween-20 (PBST). Blocking was performed using the SNAP I.D. 2.0 vacuum blot hybridization system (Merck Millipore, Burlington, MA, USA) in 0.5% skim milk powder solution (Sigma-Aldrich) diluted in PBST, according to the device manufacturer's instructions. The membrane was then incubated overnight at +4°C in a solution of primary antibodies in PBST containing 0.05% sodium azide against GFAP (ab7260, 1:10000, rabbit polyclonal, Abcam, Cambridge, UK), Iba1 (1:1000, rabbit polyclonal, Novus Biologicals, Littleton, Colorado, USA), EAAT2 (ab205248, 1:1000, rabbit monoclonal, Abcam). The membrane was then washed with PBST from primary antibodies, treated with a solution of secondary antibodies (antibodies against rabbit immunoglobulin G, cat. nom. 31460, 1:20000, Pierce Goat anti-rabbit IgG-HRP, ThermoFisher Scientific) and washed from secondary antibodies using the SNAP I.D. 2.0 vacuum blot hybridization system (Merck Millipore, USA) according to the manufacturer's recommendations. Chemiluminescent signal was obtained using SuperSignal™ West Pico PLUS chemiluminescent substrate (ThermoFisher Scientific) and documented using the ChemiDoc MP system (Bio-Rad). Densitometric analysis was performed using Image Lab 6.0.1 software (Bio-Rad): the optical specific signal was normalized to the calibrator sample by the Ponseau S (total protein) signal of the corresponding lane.
4.5. Statistical Analysis
The statistical analysis was conducted using the SPSS Statistics 23 software (IBM, Armonk, NY, USA) and the GraphPad Prism software (GraphPad Software, San Diego, CA, USA). Outliers were identified through the application of the quartile method. The log-rank test was employed for the analysis of survival curves in rats. The Kolmogorov-Smirnov test was employed to ascertain the normality of the distributions. The assumption of homogeneity of variance was evaluated using the Levene test. A three-way mixed ANOVA (test days × TLE × treatment) was employed to analyze weight dynamics during drug administration. For the remaining data, which exhibited a normal distribution, a two-way ANOVA (TLE × treatment) with Sidak's posterior multiple comparisons test was employed. Statistical significance was set at p ≤ 0.05. Graphs display mean and standard errors. The results of statistical processing are described in detail in the text for the variables for which statistically significant differences were identified.
Author Contributions
Conceptualization, O.E.Z. and A.V.Z.; methodology, O.E.Z., A.P.S., A.V.Z.; formal analysis, O.E.Z., A.A.K., M.V.Z., D.S.S.; investigation, O.E.Z., A.A.K., A.I.R., A.R.K., M.V.Z., D.S.S., writing—original draft preparation, O.E.Z., A.R.K., A.I.R., A.A.K., M.V.Z., A.P.S., D.S.S.; writing—review and editing, O.E.Z., A.V.Z., A.A.K.; supervision, A.V.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Science Foundation, grant number 23-25-00480.
Institutional Review Board Statement
In The study was conducted according to the EU Directive 2010/63/EU for animal experiments and approved by the Ethics Committee of the Sechenov Institute of Evolutionary Physiology and Biochemistry of the Russian Academy of Sciences (ethical permit number 1-16, 26 January 2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
The authors thank Marat R. Subkhankulov for excellent assistance with behavioral experiments.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A

Table A1.
Primers and probes used in RT-qPCR.
| Gene Symbol RefSeq Accession Number |
Nucleotide Sequences (Forward, Reverse, TaqMan Probe) |
Final Primers and Probe Concentration (nM) | Reference |
|---|---|---|---|
|
Actb (actin beta) NM_031144 |
TGTCACCAACTGGGACGATA GGGGTGTTGAAGGTCTCAAA FAM-CGTGTGGCCCCTGAGGAGCAC-BHQ1 |
200 200 |
[98] (primers) [99] (probe) |
|
Gapdh (glyceraldehyde-3-phosphate dehydrogenase) NM_017008 |
TGCACCACCAACTGCTTAG GGATGCAGGGATGATGTTC R6G-ATCACGCCACAGCTTTCCAGAGGG-BHQ2 |
200 100 |
[100] |
|
B2m (beta-2-microglobulin) NM_012512 |
TGCCATTCAGAAAACTCCCC GAGGAAGTTGGGCTTCCCATT ROX-ATTCAAGTGTACTCTCGCCATCCACCG-BHQ1 |
200 100 |
[101] |
|
Rpl13a (ribosomal protein L13a) NM_173340 |
GGATCCCTCCACCCTATGACA CTGGTACTTCCACCCGACCTC FAM-CTGCCCTCAAGGTTGTGCGGCT-BHQ1 |
200 100 |
[102] (primers) [99] (probe) |
|
Sdha (succinate dehydrogenase complex flavoprotein subunit A) NM_130428 |
AGACGTTTGACAGGGGAATG TCATCAATCCGCACCTTGTA R6G-ACCTGGTGGAGACGCTGGAGCT-BHQ2 |
200 100 |
[103] (primers) [99] (probe) |
|
Ppia (peptidylprolyl isomerase A) NM_017101 |
AGGATTCATGTGCCAGGGTG CTCAGTCTTGGCAGTGCAGA ROX-CACGCCATAATGGCACTGGTGGCA-BHQ1 |
200 100 |
[104] |
|
Hprt1 (hypoxanthine phosphoribosyltransferase 1) NM_012583 |
TCCTCAGACCGCTTTTCCCGC TCATCATCACTAATCACGACGCTGG FAM-CCGACCGGTTCTGTCATGTCGACCCT-BHQ1 |
200 100 |
[105] (primers) [99] (probe) |
|
Pgk1 (phosphoglycerate kinase 1) NM_053291 |
ATGCAAAGACTGGCCAAGCTAC AGCCACAGCCTCAGCATATTTC R6G-TGCTGGCTGGATGGGCTTGGA-BHQ2 |
200 100 |
[106] (primers) [99] (probe) |
|
Ywhaz (tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zet) NM_013011 |
GATGAAGCCATTGCTGAACTTG GTCTCCTTGGGTATCCGATGTC ROX-TGAAGAGTCGTACAAAGACAGCACGC-BHQ1 |
200 100 |
[106] (primers) [99] (probe) |
|
Il1b (interleukin 1 beta) NM_031512 |
CACCTCTCAAGCAGAGCACAG GGGTTCCATGGTGAAGTCAAC FAM-TGTCCCGACCATTGCTGTTTCCTAG-BHQ1 |
400 200 |
[107] |
|
Tnfa (tumor necrosis factor) NM_012675 |
CCAGGTTCTCTTCAAGGGACAA CTCCTGGTATGAAATGGCAAATC ROX-CCCGACTATGTGCTCCTCACCCACA-BHQ2 |
200 200 |
[107] |
|
Aif1 (allograft inflammatory factor 1) NM_017196.3 |
CAACACACTGCAGCCTCATC AAGCTTTTCCTCCCTGCAAA Cy5-CCCCACCTAAGGCCACCAGCGTCTGA-BHQ3 |
200 100 |
[39] |
|
Gfap (glial fibrillary acidic protein) NM_017009.2 |
TGGCCACCAGTAACATGCAA CAGTTGGCGGCGATAGTCAT HEX-CGGTCCAAGTTTGCAGACCTCACAG-BHQ2 |
200 200 |
[108] (primers) [6] (probe) |
|
Slc1a2 (solute carrier family 1 member 2) NM_017215.2 |
CCAGTGCTGGAACTTTGCCT TAAAGGGCTGTACCATCCAT FAM-AGCGTGTGACCAGATTCGTCCTCCCA-BHQ1 |
200 150 |
[109] (primers) [6] (probe) |
|
Il1rn (interleukin 1 receptor antagonist) NM_022194.2 |
GGGGACCTTACAGTCACCTAAT GGTTAGTATCCCAGATTCTGAAGG ROX-AGTCAGCTGGCCACCCTGCTGGGA-BHQ2 |
400 100 |
[39] |
|
Nlrp3 (NLR family pyrin domain containing 3) NM_001191642 |
CAGACCCTCATGTTGCCTGT AGACCTCGGCAGAAGCTAGA FAM-CCAGACTGGTGAACTGCTGCCTCA-BHQ1 |
200 100 |
[46] |
|
Lcn2 (lipocalin 2) NM_130741.1 |
AGCTACGATGTGCAAGTGGC CCCCTTGGTTCTTCCGTACA FAM-CGACACTGACTACGACCAGTTTGCCA-BHQ1 |
200 150 |
[46] |
|
Arg1 (arginase 1) NM_017134.3 |
AGCTGGGAATTGGCAAAGTG AACTCAGGTGAATGGGCCTT HEX-TGGAAGAGACCTTCAGCTACCTGC-BHQ2 |
300 100 |
[110] (primers) [46] (probe) |
|
S100a10 (S100 calcium binding protein A10) NM_031114.1 |
CATTTCACAGGTTTGCAGGGG GCACTGGTCCAGGTCTTTCA Cy5-AGGACCCTCTGGCTGTGGACA-BHQ3 |
200 250 |
[46] |
|
Ptx3 (pentraxin 3) NM_001109536.2 |
AAACTTCGCCTCTCCAGCAA CATGGTGTGGGGTCCTCG HEX-TGCTCTCTGGTCTGCAGTGTTGGC-BHQ2 |
400 200 |
[46] |
|
Gbp2 (guanylate binding protein 2) NM_133624.2 |
AGTCAATGGGCCACGTCTAA AGTGGGTGATGGCCTTTTGT HEX-AGCAGTGGGTCTCTCCCCTGCA-BHQ2 |
200 100 |
[46] |
|
Nos2 (nitric oxide synthase 2) NM_012611.3 |
CAGAAGCAGAATGTGACCATCAT CGGAGGGACCAGCCAAATC ROX-CCACCACACAGCCTCAGAGTCCTT-BHQ2 |
200 200 |
[111] |
References
- Löscher, W.; Klitgaard, H.; Twyman, R.E.; Schmidt, D. New avenues for anti-epileptic drug discovery and development. Nat. Rev. Drug Discov. 2013, 12, 757–776. [Google Scholar] [CrossRef]
- Sanz, P.; Rubio, T.; Garcia-Gimeno, M.A. Neuroinflammation and Epilepsy: From Pathophysiology to Therapies Based on Repurposing Drugs. Int. J. Mol. Sci. 2024, Vol. 25, Page 4161 2024, 25, 4161. [Google Scholar] [CrossRef]
- Devinsky, O.; Vezzani, A.; Najjar, S.; De Lanerolle, N.C.; Rogawski, M.A. Glia and epilepsy: excitability and inflammation. Trends Neurosci. 2013, 36, 174–184. [Google Scholar] [CrossRef]
- Riazi, K.; Galic, M.A.; Kuzmiski, J.B.; Ho, W.; Sharkey, K.A.; Pittman, Q.J. Microglial activation and TNFα production mediate altered CNS excitability following peripheral inflammation. Proc. Natl. Acad. Sci. 2008, 105, 17151–17156. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, K.M.; Hutchinson, M.R.; Northcutt, A.; Maier, S.F.; Watkins, L.R.; Barth, D.S. The cortical innate immune response increases local neuronal excitability leading to seizures. Brain 2009, 132, 2478–2486. [Google Scholar] [CrossRef]
- Dyomina, A. V.; Zubareva, O.E.; Smolensky, I. V.; Vasilev, D.S.; Zakharova, M. V.; Kovalenko, A.A.; Schwarz, A.P.; Ischenko, A.M.; Zaitsev, A. V. Anakinra Reduces Epileptogenesis, Provides Neuroprotection, and Attenuates Behavioral Impairments in Rats in the Lithium–Pilocarpine Model of Epilepsy. Pharmaceuticals 2020, 13, 340. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Han, S.-H.; Lee, K.-W. Kainic acid-induced seizures cause neuronal death in infant rats pretreated with lipopolysaccharide. Neuroreport 2000, 11, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Suleymanova, E.M. Behavioral comorbidities of epilepsy and neuroinflammation: Evidence from experimental and clinical studies. Epilepsy Behav. 2021, 117, 107869. [Google Scholar] [CrossRef]
- Li, D.; Wang, Y.; Guo, Y.; Wang, W. Bioinformatics analysis reveals multiple functional changes in astrocytes in temporal lobe epilepsy. Brain Res. 2024, 1831. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Fan, Y.-Y.; Huo, J. A1/A2 astrocytes in central nervous system injuries and diseases: Angels or devils? Neurochem. Int. 2021, 148, 105080. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Sierra, A.; Stevens, B.; Tremblay, M.-E.; Aguzzi, A.; Ajami, B.; Amit, I.; Audinat, E.; Bechmann, I.; Bennett, M.; et al. Microglia states and nomenclature: A field at its crossroads. Neuron 2022, 110, 3458–3483. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ji, X.; Shan, M.; Wang, Y.; Dai, X.; Yin, M.; Liu, Y.; Guan, L.; Ye, L.; Cheng, H. Melatonin regulates microglial polarization to M2 cell via RhoA/ROCK signaling pathway in epilepsy. Immunity, Inflamm. Dis. 2023, 11. [Google Scholar] [CrossRef]
- Zhao, Q.; Wu, X.; Yan, S.; Xie, X.; Fan, Y.; Zhang, J.; Peng, C.; You, Z. The antidepressant-like effects of pioglitazone in a chronic mild stress mouse model are associated with PPARγ-mediated alteration of microglial activation phenotypes. J. Neuroinflammation 2016, 13, 259. [Google Scholar] [CrossRef]
- Yonutas, H.M.; Sullivan, P.G. Targeting PPAR isoforms following CNS injury. Curr. Drug Targets 2013, 14, 733–742. [Google Scholar] [CrossRef]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: nutritional and clinical implications--a review. Nutr. J. 2014, 13. [Google Scholar] [CrossRef]
- Li, J.; Guo, C.; Wu, J. 15-Deoxy-∆-12,14-Prostaglandin J2 (15d-PGJ2), an Endogenous Ligand of PPAR- γ: Function and Mechanism. PPAR Res. 2019, 2019. [Google Scholar] [CrossRef]
- Narala, V.R.; Adapala, R.K.; Suresh, M. V.; Brock, T.G.; Peters-Golden, M.; Reddy, R.C. Leukotriene B4 is a physiologically relevant endogenous peroxisome proliferator-activated receptor-alpha agonist. J. Biol. Chem. 2010, 285, 22067–22074. [Google Scholar] [CrossRef] [PubMed]
- Giordano Attianese, G.M.P.; Desvergne, B. Integrative and systemic approaches for evaluating PPARβ/δ (PPARD) function. Nucl. Recept. Signal. 2015, 13, e001. [Google Scholar] [CrossRef]
- Tufano, M.; Pinna, G. Is There a Future for PPARs in the Treatment of Neuropsychiatric Disorders? Molecules 2020, 25. [Google Scholar] [CrossRef]
- Warden, A.; Truitt, J.; Merriman, M.; Ponomareva, O.; Jameson, K.; Ferguson, L.B.; Mayfield, R.D.; Harris, R.A. Localization of PPAR isotypes in the adult mouse and human brain. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Strosznajder, A.K.; Wójtowicz, S.; Jeżyna, M.J.; Sun, G.Y.; Strosznajder, J.B. Recent Insights on the Role of PPAR-β/δ in Neuroinflammation and Neurodegeneration, and Its Potential Target for Therapy. Neuromolecular Med. 2021, 23, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Genolet, R.; Wahli, W.; Michalik, L. PPARs as drug targets to modulate inflammatory responses? Curr. Drug Targets. Inflamm. Allergy 2004, 3, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Senn, L.; Costa, A.-M.; Avallone, R.; Socała, K.; Wlaź, P.; Biagini, G. Is the peroxisome proliferator-activated receptor gamma a putative target for epilepsy treatment? Current evidence and future perspectives. Pharmacol. Ther. 2023, 241, 108316. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Segura, I.; Santiago-Balmaseda, A.; Rodríguez-Hernández, L.D.; Morales-Martínez, A.; Martínez-Becerril, H.A.; Martínez-Gómez, P.A.; Delgado-Minjares, K.M.; Salinas-Lara, C.; Martínez-Dávila, I.A.; Guerra-Crespo, M.; et al. PPARs and Their Neuroprotective Effects in Parkinson’s Disease: A Novel Therapeutic Approach in α-Synucleinopathy? Int. J. Mol. Sci. 2023, 24, 3264. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Shao, X.G.; Sun, H.; Li, Y.N.; Yang, J.; Deng, Y.C.; Huang, Y.G. Activation of cerebral peroxisome proliferator-activated receptors gamma exerts neuroprotection by inhibiting oxidative stress following pilocarpine-induced status epilepticus. Brain Res. 2008, 1200, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Adabi Mohazab, R.; Javadi-Paydar, M.; Delfan, B.; Dehpour, A.R. Possible involvement of PPAR-gamma receptor and nitric oxide pathway in the anticonvulsant effect of acute pioglitazone on pentylenetetrazole-induced seizures in mice. Epilepsy Res. 2012, 101, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wang, K.; Xiang, W.; Li, Y.; Hao, Y.; Guan, Y. Rosiglitazone polarizes microglia and protects against pilocarpine-induced status epilepticus. CNS Neurosci. Ther. 2019, 25, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chen, L.; Qu, Y.; Wang, D.; Zhu, Y.; Zhu, Y. Rosiglitazone Prevents Autophagy by Regulating Nrf2-Antioxidant Response Element in a Rat Model of Lithium-pilocarpine-induced Status Epilepticus. Neuroscience 2021, 455, 212–222. [Google Scholar] [CrossRef]
- Puligheddu, M.; Pillolla, G.; Melis, M.; Lecca, S.; Marrosu, F.; De Montis, M.G.; Scheggi, S.; Carta, G.; Murru, E.; Aroni, S.; et al. PPAR-Alpha Agonists as Novel Antiepileptic Drugs: Preclinical Findings. PLoS One 2013, 8, e64541. [Google Scholar] [CrossRef]
- Sznaidman, M.L.; Haffner, C.D.; Maloney, P.R.; Fivush, A.; Chao, E.; Goreham, D.; Sierra, M.L.; LeGrumelec, C.; Xu, H.E.; Montana, V.G.; et al. Novel selective small molecule agonists for peroxisome proliferator-activated receptor delta (PPARdelta)--synthesis and biological activity. Bioorg. Med. Chem. Lett. 2003, 13, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Nandhikonda, P.; Yasgar, A.; Baranowski, A.M.; Sidhu, P.S.; McCallum, M.M.; Pawlak, A.J.; Teske, K.; Feleke, B.; Yuan, N.Y.; Kevin, C.; et al. Peroxisome proliferation-activated receptor δ agonist GW0742 interacts weakly with multiple nuclear receptors, including the vitamin D receptor. Biochemistry 2013, 52, 4193–4203. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.-S.; Ku, P.-M.; Niu, C.-S.; Cheng, J.-T.; Lee, K.-S. Development of PPAR-agonist GW0742 as antidiabetic drug: study in animals. Drug Des. Devel. Ther. 2015, 9, 5625–5632. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Esposito, E.; Mazzon, E.; Paterniti, I.; Galuppo, M.; Cuzzocrea, S. GW0742, a selective PPAR-beta/delta agonist, contributes to the resolution of inflammation after gut ischemia/reperfusion injury. J. Leukoc. Biol. 2010, 88, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Haskova, Z.; Hoang, B.; Luo, G.; Morgan, L.A.; Billin, A.N.; Barone, F.C.; Shearer, B.G.; Barton, M.E.; Kilgore, K.S. Modulation of LPS-induced pulmonary neutrophil infiltration and cytokine production by the selective PPARbeta/delta ligand GW0742. Inflamm. Res. Off. J. Eur. Histamine Res. Soc.... [et al.] 2008, 57, 314–321. [Google Scholar] [CrossRef]
- Tang, X.; Yan, K.; Wang, Y.; Wang, Y.; Chen, H.; Xu, J.; Lu, Y.; Wang, X.; Liang, J.; Zhang, X. Activation of PPAR-β/δ Attenuates Brain Injury by Suppressing Inflammation and Apoptosis in a Collagenase-Induced Intracerebral Hemorrhage Mouse Model. Neurochem. Res. 2020, 45, 837–850. [Google Scholar] [CrossRef]
- Das, N.R.; Gangwal, R.P.; Damre, M. V; Sangamwar, A.T.; Sharma, S.S. A PPAR-β/δ agonist is neuroprotective and decreases cognitive impairment in a rodent model of Parkinson’s disease. Curr. Neurovasc. Res. 2014, 11, 114–124. [Google Scholar] [CrossRef] [PubMed]
- An, Y.-Q.; Zhang, C.T.; Du, Y.; Zhang, M.; Tang, S.S.; Hu, M.; Long, Y.; Sun, H.B.; Hong, H. PPARδ agonist GW0742 ameliorates Aβ1-42-induced hippocampal neurotoxicity in mice. Metab. Brain Dis. 2016, 31, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Zubareva, O.E.; Dyomina, A. V; Kovalenko, A.A.; Roginskaya, A.I.; Melik-Kasumov, T.B.; Korneeva, M.A.; Chuprina, A. V; Zhabinskaya, A.A.; Kolyhan, S.A.; Zakharova, M. V; et al. Beneficial Effects of Probiotic Bifidobacterium longum in a Lithium-Pilocarpine Model of Temporal Lobe Epilepsy in Rats. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Xiao, L.; Zheng, H.; Li, J.; Wang, Q.; Sun, H. Neuroinflammation Mediated by NLRP3 Inflammasome After Intracerebral Hemorrhage and Potential Therapeutic Targets. Mol. Neurobiol. 2020, 57, 5130–5149. [Google Scholar] [CrossRef]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front. Immunol. 2014, 5, 120091. [Google Scholar] [CrossRef]
- Kim, J.H.; Ko, P.W.; Lee, H.W.; Jeong, J.Y.; Lee, M.G.; Kim, J.H.; Lee, W.H.; Yu, R.; Oh, W.J.; Suk, K. Astrocyte-derived lipocalin-2 mediates hippocampal damage and cognitive deficits in experimental models of vascular dementia. Glia 2017, 65, 1471–1490. [Google Scholar] [CrossRef]
- Bi, F.; Huang, C.; Tong, J.; Qiu, G.; Huang, B.; Wu, Q.; Li, F.; Xu, Z.; Bowser, R.; Xia, X.-G.; et al. Reactive astrocytes secrete lcn2 to promote neuron death. Proc. Natl. Acad. Sci. 2013, 110, 4069–4074. [Google Scholar] [CrossRef] [PubMed]
- El Baassiri, M.G.; Rahal, S.S.; Fulton, W.B.; Sodhi, C.P.; Hackam, D.J.; Nasr, I.W. Pharmacologic Toll-like receptor 4 inhibition skews toward a favorable A1/A2 astrocytic ratio improving neurocognitive outcomes following traumatic brain injury. J. Trauma Acute Care Surg. 2023, 95, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, M. V; Dyomina, A. V; Kovalenko, A.A.; Zubareva, O.E.; Ischenko, A.M.; Zaitsev, A. V Anakinra Promotes M2 Microglia Activation during the Latent Phase of the Lithium-Pilocarpine Model of Temporal Lobe Epilepsy. J. Evol. Biochem. Physiol. 2024, 60, 672–689. [Google Scholar] [CrossRef]
- Ding, Z.-B.; Song, L.-J.; Wang, Q.; Kumar, G.; Yan, Y.-Q.; Ma, C.-G. Astrocytes: a double-edged sword in neurodegenerative diseases. Neural Regen. Res. 2021, 16, 1702. [Google Scholar] [CrossRef]
- Zaitsev, A. V.; Smolensky, I. V.; Jorratt, P.; Ovsepian, S. V. Neurobiology, Functions, and Relevance of Excitatory Amino Acid Transporters (EAATs) to Treatment of Refractory Epilepsy. CNS Drugs 2020, 34, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Zubareva, O.E.; Kovalenko, A.A.; Kalemenev, S. V.; Schwarz, A.P.; Karyakin, V.B.; Zaitsev, A. V. Alterations in mRNA expression of glutamate receptor subunits and excitatory amino acid transporters following pilocarpine-induced seizures in rats. Neurosci. Lett. 2018, 686, 94–100. [Google Scholar] [CrossRef]
- Soltani Khaboushan, A.; Yazdanpanah, N.; Rezaei, N. Neuroinflammation and Proinflammatory Cytokines in Epileptogenesis. Mol. Neurobiol. 2022, 59, 1724–1743. [Google Scholar] [CrossRef]
- Aleshin, S.; Reiser, G. Role of the peroxisome proliferator-activated receptors (PPAR)-α, β/δ and γ triad in regulation of reactive oxygen species signaling in brain. Biol. Chem. 2013, 394, 1553–1570. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, R.; Toral, M.; Gómez-Guzmán, M.; Romero, M.; Sanchez, M.; Mahmoud, A.M.; Duarte, J. The Role of Nrf2 Signaling in PPARβ/δ-Mediated Vascular Protection against Hyperglycemia-Induced Oxidative Stress. Oxid. Med. Cell. Longev. 2018, 2018, 5852706. [Google Scholar] [CrossRef] [PubMed]
- Magadum, A.; Engel, F.B. PPARβ/δ: Linking Metabolism to Regeneration. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Chen, Z.-P.; Wang, S.; Zhao, X.; Fang, W.; Wang, Z.; Ye, H.; Wang, M.-J.; Ke, L.; Huang, T.; Lv, P.; et al. Lipid-accumulated reactive astrocytes promote disease progression in epilepsy. Nat. Neurosci. 2023, 26, 542–554. [Google Scholar] [CrossRef]
- Boison, D.; Steinhäuser, C. Epilepsy and astrocyte energy metabolism. Glia 2018, 66, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Geronzi, U.; Lotti, F.; Grosso, S. Oxidative stress in epilepsy. Expert Rev. Neurother. 2018, 18, 427–434. [Google Scholar] [CrossRef]
- Subkhankulov, M.R.; Sinyak, D.S.; Guk, V.A.; Postnikova, T.Y.; Roginskaya, A.I.; Zubareva, O.E. Cardarin Effect on the Formation of Histopathological and Behavioral Abnormalities in the Lithium-Pilocarpine Model of Temporal Lobe Epilepsy in Rats. J. Evol. Biochem. Physiol. 2024 601 2024, 60, 316–331. [Google Scholar] [CrossRef]
- Dyomina, A. V; Smolensky, I. V; Zaitsev, A. V Refinement of the Barnes and Morris water maze protocols improves characterization of spatial cognitive deficits in the lithium-pilocarpine rat model of epilepsy. Epilepsy Behav. 2023, 147, 109391. [Google Scholar] [CrossRef]
- Smolensky, I. V; Zubareva, O.E.; Kalemenev, S. V; Lavrentyeva, V. V; Dyomina, A. V; Karepanov, A.A.; Zaitsev, A. V Impairments in cognitive functions and emotional and social behaviors in a rat lithium-pilocarpine model of temporal lobe epilepsy. Behav. Brain Res. 2019, 372, 112044. [Google Scholar] [CrossRef]
- Suleymanova, E.M.; Gulyaev, M. V.; Abbasova, K.R. Structural alterations in the rat brain and behavioral impairment after status epilepticus: An MRI study. Neuroscience 2016, 315, 79–90. [Google Scholar] [CrossRef]
- Wulsin, A.C.; Franco-Villanueva, A.; Romancheck, C.; Morano, R.L.; Smith, B.L.; Packard, B.A.; Danzer, S.C.; Herman, J.P. Functional disruption of stress modulatory circuits in a model of temporal lobe epilepsy. PLoS One 2018, 13. [Google Scholar] [CrossRef]
- Rojas, J.J.; Deniz, B.F.; Miguel, P.M.; Diaz, R.; Hermel, É. do E.S.; Achaval, M.; Netto, C.A.; Pereira, L.O. Effects of daily environmental enrichment on behavior and dendritic spine density in hippocampus following neonatal hypoxia-ischemia in the rat. Exp. Neurol. 2013, 241, 25–33. [Google Scholar] [CrossRef]
- Pan, H.-C.; Yang, C.-N.; Lee, W.-J.; Sheehan, J.; Wu, S.-M.; Chen, H.-S.; Lin, M.-H.; Shen, L.-W.; Lee, S.-H.; Shen, C.-C.; et al. Melatonin Enhanced Microglia M2 Polarization in Rat Model of Neuro-inflammation Via Regulating ER Stress/PPARδ/SIRT1 Signaling Axis. J. Neuroimmune Pharmacol. 2024, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xue, L.; Zheng, J.; Tian, X.; Zhang, Y.; Tong, Q. PPARß/δ agonist alleviates NLRP3 inflammasome-mediated neuroinflammation in the MPTP mouse model of Parkinson’s disease. Behav. Brain Res. 2019, 356, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Konttinen, H.; Gureviciene, I.; Oksanen, M.; Grubman, A.; Loppi, S.; Huuskonen, M.T.; Korhonen, P.; Lampinen, R.; Keuters, M.; Belaya, I.; et al. PPARβ/δ-agonist GW0742 ameliorates dysfunction in fatty acid oxidation in PSEN1ΔE9 astrocytes. Glia 2019, 67, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xue, T.; Zhang, Z.; Wang, X.; Xu, P.; Zhang, J.; Lei, X.; Li, Y.; Xie, Y.; Wang, L.; et al. Role of signal transducer and activator of transcription-3 in up-regulation of GFAP after epilepsy. Neurochem. Res. 2011, 36, 2208–15. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Murugan, M.; Bosco, D.B.; Liu, Y.; Peng, J.; Worrell, G.A.; Wang, H.L.; Ta, L.E.; Richardson, J.R.; Shen, Y.; et al. Microglial proliferation and monocyte infiltration contribute to microgliosis following status epilepticus. Glia 2019, 67, 1434–1448. [Google Scholar] [CrossRef]
- Kyriatzis, G.; Bernard, A.; Bôle, A.; Khrestchatisky, M.; Ferhat, L. In the Rat Hippocampus, Pilocarpine-Induced Status Epilepticus Is Associated with Reactive Glia and Concomitant Increased Expression of CD31, PDGFRβ, and Collagen IV in Endothelial Cells and Pericytes of the Blood–Brain Barrier. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef]
- Dyomina, A. V.; Kovalenko, A.A.; Zakharova, M. V.; Postnikova, T.Y.; Griflyuk, A. V.; Smolensky, I. V.; Antonova, I. V.; Zaitsev, A. V. MTEP, a Selective mGluR5 Antagonist, Had a Neuroprotective Effect but Did Not Prevent the Development of Spontaneous Recurrent Seizures and Behavioral Comorbidities in the Rat Lithium-Pilocarpine Model of Epilepsy. Int. J. Mol. Sci. 2022, 23, 497. [Google Scholar] [CrossRef]
- Postnikova, T.Y.; Diespirov, G.P.; Amakhin, D. V.; Vylekzhanina, E.N.; Soboleva, E.B.; Zaitsev, A. V. Impairments of Long-Term Synaptic Plasticity in the Hippocampus of Young Rats during the Latent Phase of the Lithium-Pilocarpine Model of Temporal Lobe Epilepsy. Int. J. Mol. Sci. 2021, 22, 13355. [Google Scholar] [CrossRef]
- Li, J.; Xiang, X.; Gong, X.; Shi, Y.; Yang, J.; Xu, Z. Cilostazol protects mice against myocardium ischemic/reperfusion injury by activating a PPARγ/JAK2/STAT3 pathway. Biomed. Pharmacother. 2017, 94, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.T.; Kao, N.J.; Cross, T.W.L.; Lee, W.J.; Lin, S.H. Effects of ketogenic diet on cognitive functions of mice fed high-fat-high-cholesterol diet. J. Nutr. Biochem. 2022, 104. [Google Scholar] [CrossRef]
- Simeone, T.A.; Matthews, S.A.; Samson, K.K.; Simeone, K.A. Regulation of brain PPARgamma2 contributes to ketogenic diet anti-seizure efficacy. Exp. Neurol. 2017, 287, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Robel, S.; Sontheimer, H. Glia as drivers of abnormal neuronal activity. Nat. Neurosci. 2016 191 2015, 19, 28–33. [Google Scholar] [CrossRef]
- Qi, R.; Wang, M.; Zhong, Q.; Wang, L.; Yang, X.; Huang, B.; Yang, Z.; Zhang, C.; Geng, X.; Luo, C.; et al. Chronic vagus nerve stimulation (VNS) altered IL-6, IL-1β, CXCL-1 and IL-13 levels in the hippocampus of rats with LiCl-pilocarpine-induced epilepsy. Brain Res. 2022, 1780. [Google Scholar] [CrossRef] [PubMed]
- Pohlentz, M.S.; Müller, P.; Cases-Cunillera, S.; Opitz, T.; Surges, R.; Hamed, M.; Vatter, H.; Schoch, S.; Becker, A.J.; Pitsch, J. Characterisation of NLRP3 pathway-related neuroinflammation in temporal lobe epilepsy. PLoS One 2022, 17, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Jeong, E.A.; Lee, J.Y.; An, H.S.; Jang, H.M.; Ahn, Y.J.; Lee, J.; Kim, K.E.; Roh, G.S. Lipocalin-2 Deficiency Reduces Oxidative Stress and Neuroinflammation and Results in Attenuation of Kainic Acid-Induced Hippocampal Cell Death. Antioxidants (Basel, Switzerland) 2021, 10, 1–13. [Google Scholar] [CrossRef]
- Liu, R.; Wang, J.; Chen, Y.; Collier, J.M.; Capuk, O.; Jin, S.; Sun, M.; Mondal, S.K.; Whiteside, T.L.; Stolz, D.B.; et al. NOX activation in reactive astrocytes regulates astrocytic LCN2 expression and neurodegeneration. Cell Death Dis. 2022, 13. [Google Scholar] [CrossRef]
- Jha, M.K.; Suk, K. Glia-based biomarkers and their functional role in the CNS. Expert Rev. Proteomics 2013, 10, 43–63. [Google Scholar] [CrossRef]
- Hoellenriegel, J.; Meadows, S.A.; Sivina, M.; Wierda, W.G.; Kantarjian, H.; Keating, M.J.; Giese, N.; O’Brien, S.; Yu, A.; Miller, L.L.; et al. The phosphoinositide 3’-kinase delta inhibitor, CAL-101, inhibits B-cell receptor signaling and chemokine networks in chronic lymphocytic leukemia. Blood 2011, 118, 3603–3612. [Google Scholar] [CrossRef]
- Babiychuk, E.B.; Draeger, A. Annexins in cell membrane dynamics. Ca(2+)-regulated association of lipid microdomains. J. Cell Biol. 2000, 150, 1113–1123. [Google Scholar] [CrossRef]
- Milosevic, A.; Liebmann, T.; Knudsen, M.; Schintu, N.; Svenningsson, P.; Greengard, P. Cell- and region-specific expression of depression-related protein p11 (S100a10) in the brain. J. Comp. Neurol. 2017, 525, 955–975. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Greengard, P. p11 (S100A10)--an inducible adaptor protein that modulates neuronal functions. Curr. Opin. Pharmacol. 2007, 7, 27–32. [Google Scholar] [CrossRef]
- Bagdy, G.; Kecskemeti, V.; Riba, P.; Jakus, R. Serotonin and epilepsy. J. Neurochem. 2007, 100, 857–873. [Google Scholar] [CrossRef] [PubMed]
- Dahal, A.; Govindarajan, K.; Kar, S. Administration of Kainic Acid Differentially Alters Astrocyte Markers and Transiently Enhanced Phospho-tau Level in Adult Rat Hippocampus. Neuroscience 2023, 516, 27–41. [Google Scholar] [CrossRef]
- Murphy, M.P. Nitric oxide and cell death. Biochim. Biophys. Acta 1999, 1411, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Hardbower, D.M.; Asim, M.; Luis, P.B.; Singh, K.; Barry, D.P.; Yang, C.; Steeves, M.A.; Cleveland, J.L.; Schneider, C.; Piazuelo, M.B.; et al. Ornithine decarboxylase regulates M1 macrophage activation and mucosal inflammation via histone modifications. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E751–E760. [Google Scholar] [CrossRef]
- Kharisova, A.R.; Roginskaya, A.I.; Zubareva, O.E. Effect of Cardarine on Gene Expression of Proteins Involved in Epileptogenesis in Rat Hippocampus in the Lithium-Pilocarpine Model of Temporal Lobe Epilepsy. J. Evol. Biochem. Physiol. 2024 603 2024, 60, 1064–1081. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation: cortical areas. Electroencephalogr. Clin. Neurophysiol. 1975, 38, 1–12. [Google Scholar] [CrossRef]
- Malm, T.; Mariani, M.; Donovan, L.J.; Neilson, L.; Landreth, G.E. Activation of the nuclear receptor PPARδ is neuroprotective in a transgenic mouse model of Alzheimer’s disease through inhibition of inflammation. J. Neuroinflammation 2015, 12, 7. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The rat brain in stereotaxic coordinates: hard cover edition; Elsevier: Amsterdam, The Netherlands, 2006; ISBN 9780080475134. [Google Scholar]
- Chauhan, P.; Philip, S.E.; Chauhan, G.; Mehra, S. The Anatomical Basis of Seizures. In Epilepsy; Exon Publications, 2022; pp. 15–24.
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schwarz, A.P.; Kovalenko, A.A.; Malygina, D.A.; Postnikova, T.Y.; Zubareva, O.E.; Zaitsev, A. V. Reference gene validation in the brain regions of young rats after pentylenetetrazole-induced seizures. Biomedicines 2020, 8, 1–12. [Google Scholar] [CrossRef]
- Kopec, A.M.; Rivera, P.D.; Lacagnina, M.J.; Hanamsagar, R.; Bilbo, S.D. Optimized solubilization of TRIzol-precipitated protein permits Western blotting analysis to maximize data available from brain tissue. J. Neurosci. Methods 2017, 280, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Harrington, C.R. Lowry protein assay containing sodium dodecyl sulfate in microtiter plates for protein determinations on fractions from brain tissue. Anal. Biochem. 1990, 186, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970. [Google Scholar] [CrossRef]
- Bonefeld, B.E.; Elfving, B.; Wegener, G. Reference genes for normalization: A study of rat brain tissue. Synapse 2008, 62, 302–309. [Google Scholar] [CrossRef]
- Schwarz, A.P.; Malygina, D.A.; Kovalenko, A.A.; Trofimov, A.N.; Zaitsev, A. V. Multiplex qPCR assay for assessment of reference gene expression stability in rat tissues/samples. Mol. Cell. Probes 2020, 53. [Google Scholar] [CrossRef]
- Lin, W.; Burks, C.A.; Hansen, D.R.; Kinnamon, S.C.; Gilbertson, T.A. Taste Receptor Cells Express pH-Sensitive Leak K + Channels. J. Neurophysiol. 2004, 92, 2909–2919. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Yamauchi, A.; Nishimura, M.; Ueda, N.; Naito, S. Soybean Oil Fat Emulsion Prevents Cytochrome P450 mRNA Down-Regulation Induced by Fat-Free Overdose Total Parenteral Nutrition in Infant Rats. Biol. Pharm. Bull. 2005, 28, 143–147. [Google Scholar] [CrossRef]
- Swijsen, A.; Nelissen, K.; Janssen, D.; Rigo, J.-M.; Hoogland, G. Validation of reference genes for quantitative real-time PCR studies in the dentate gyrus after experimental febrile seizures. BMC Res. Notes 2012, 5, 685. [Google Scholar] [CrossRef] [PubMed]
- Pohjanvirta, R.; Niittynen, M.; Lindén, J.; Boutros, P.C.; Moffat, I.D.; Okey, A.B. Evaluation of various housekeeping genes for their applicability for normalization of mRNA expression in dioxin-treated rats. Chem. Biol. Interact. 2006, 160, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Malkin, S.L.; Amakhin, D. V; Veniaminova, E.A.; Kim, K.K.; Zubareva, O.E.; Magazanik, L.G.; Zaitsev, A. V Changes of AMPA receptor properties in the neocortex and hippocampus following pilocarpine-induced status epilepticus in rats. Neuroscience 2016, 327, 146–55. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.L.; Vink, R.; Donkin, J.J.; van den Heuvel, C. Validation of reference genes for normalization of real-time quantitative RT-PCR data in traumatic brain injury. J. Neurosci. Res. 2009, 87, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Langnaese, K.; John, R.; Schweizer, H.; Ebmeyer, U.; Keilhoff, G. Selection of reference genes for quantitative real-time PCR in a rat asphyxial cardiac arrest model. BMC Mol. Biol. 2008, 9, 53. [Google Scholar] [CrossRef]
- Rioja, I.; Bush, K.A.; Buckton, J.B.; Dickson, M.C.; Life, P.F. Joint cytokine quantification in two rodent arthritis models: kinetics of expression, correlation of mRNA and protein levels and response to prednisolone treatment. Clin. Exp. Immunol. 2004, 137, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, V.; Tanga, F.Y.; DeLeo, J.A. Attenuation of Morphine Tolerance, Withdrawal-Induced Hyperalgesia, and Associated Spinal Inflammatory Immune Responses by Propentofylline in Rats. Neuropsychopharmacology 2004, 29, 327–334. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, S.M.; Hasselfeld, K.; Bauer, D.; Simmons, M.; Roussos, P.; Haroutunian, V.; Meador-Woodruff, J.H.; McCullumsmith, R.E. Glutamate transporter splice variant expression in an enriched pyramidal cell population in schizophrenia. Transl. Psychiatry 2015, 5, e579. [Google Scholar] [CrossRef]
- Su, J.; Zhang, Y.; Cheng, C.; Zhu, Y.; Ye, Y.; Sun, Y.; Xiang, S.; Wang, Y.; Liu, Z.; Zhang, X. Hydrogen regulates the M1/M2 polarization of alveolar macrophages in a rat model of chronic obstructive pulmonary disease. Exp. Lung Res. 2021, 47, 301–310. [Google Scholar] [CrossRef]
- Sang, N.; Yun, Y.; Li, H.; Hou, L.; Han, M.; Li, G. SO2 inhalation contributes to the development and progression of ischemic stroke in the brain. Toxicol. Sci. 2010, 114, 226–236. [Google Scholar] [CrossRef]
Figure 1.
Effects of GW0742 treatment on survival and behavior of rats in a lithium-pilocarpine model of TLE. (a) Kaplan–Meier survival curves. (b) Body weight dynamics. Behavior of control and TLE rats in the social test (c) and open field test (d-g): (c) Communication time in the social test. (d) Grooming time. (e) Length of distance traveled. (f) Locomotion time. (g) Examples of tracks in the open field. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. * p < 0.05; ** p < 0.01; *** p < 0.001, multiple comparisons Sidak's test.
Figure 1.
Effects of GW0742 treatment on survival and behavior of rats in a lithium-pilocarpine model of TLE. (a) Kaplan–Meier survival curves. (b) Body weight dynamics. Behavior of control and TLE rats in the social test (c) and open field test (d-g): (c) Communication time in the social test. (d) Grooming time. (e) Length of distance traveled. (f) Locomotion time. (g) Examples of tracks in the open field. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. * p < 0.05; ** p < 0.01; *** p < 0.001, multiple comparisons Sidak's test.
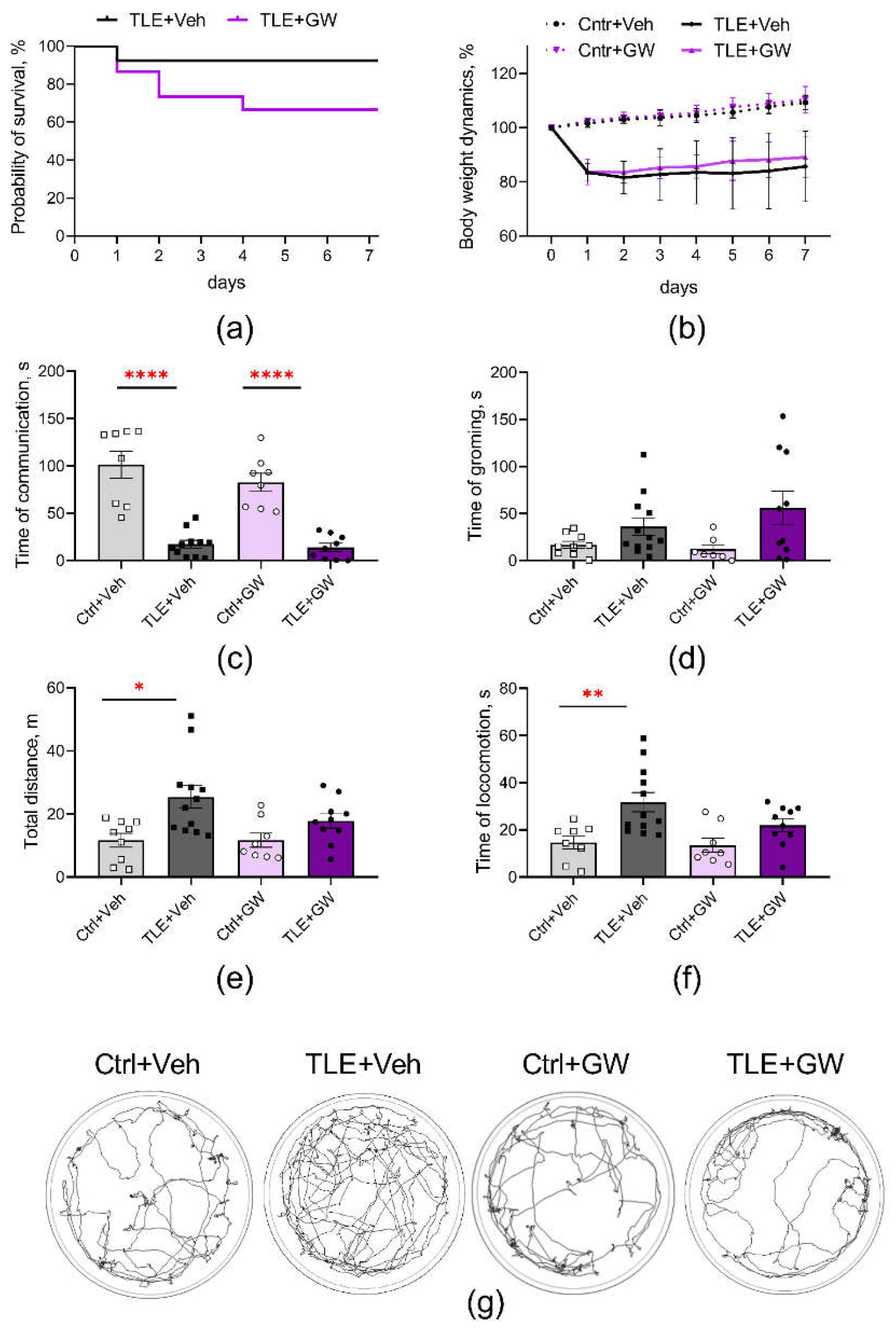
Figure 2.
Relative gene expression of astrocyte activation marker Gfap (a, b) and microglial cell activation marker Aif1 (c, d) in the dorsal hippocampus and temporal cortex of TLE and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05, *** p < 0.001.
Figure 2.
Relative gene expression of astrocyte activation marker Gfap (a, b) and microglial cell activation marker Aif1 (c, d) in the dorsal hippocampus and temporal cortex of TLE and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05, *** p < 0.001.
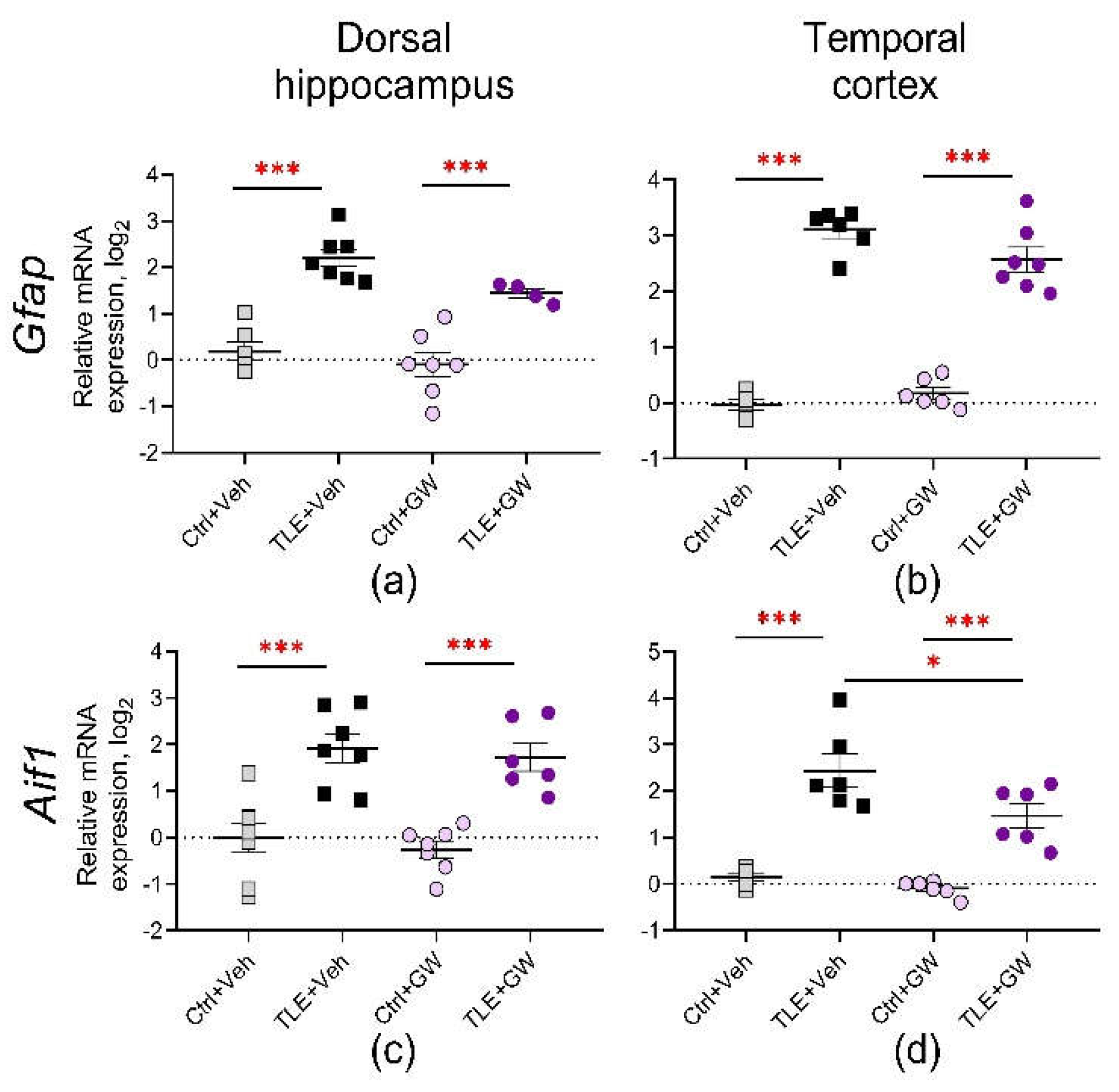
Figure 3.
Changes in relative protein levels of the astrocyte marker GFAP in the dorsal hippocampus (a, c) and temporal cortex (b, d) of rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: *** – p < 0.001.
Figure 3.
Changes in relative protein levels of the astrocyte marker GFAP in the dorsal hippocampus (a, c) and temporal cortex (b, d) of rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: *** – p < 0.001.
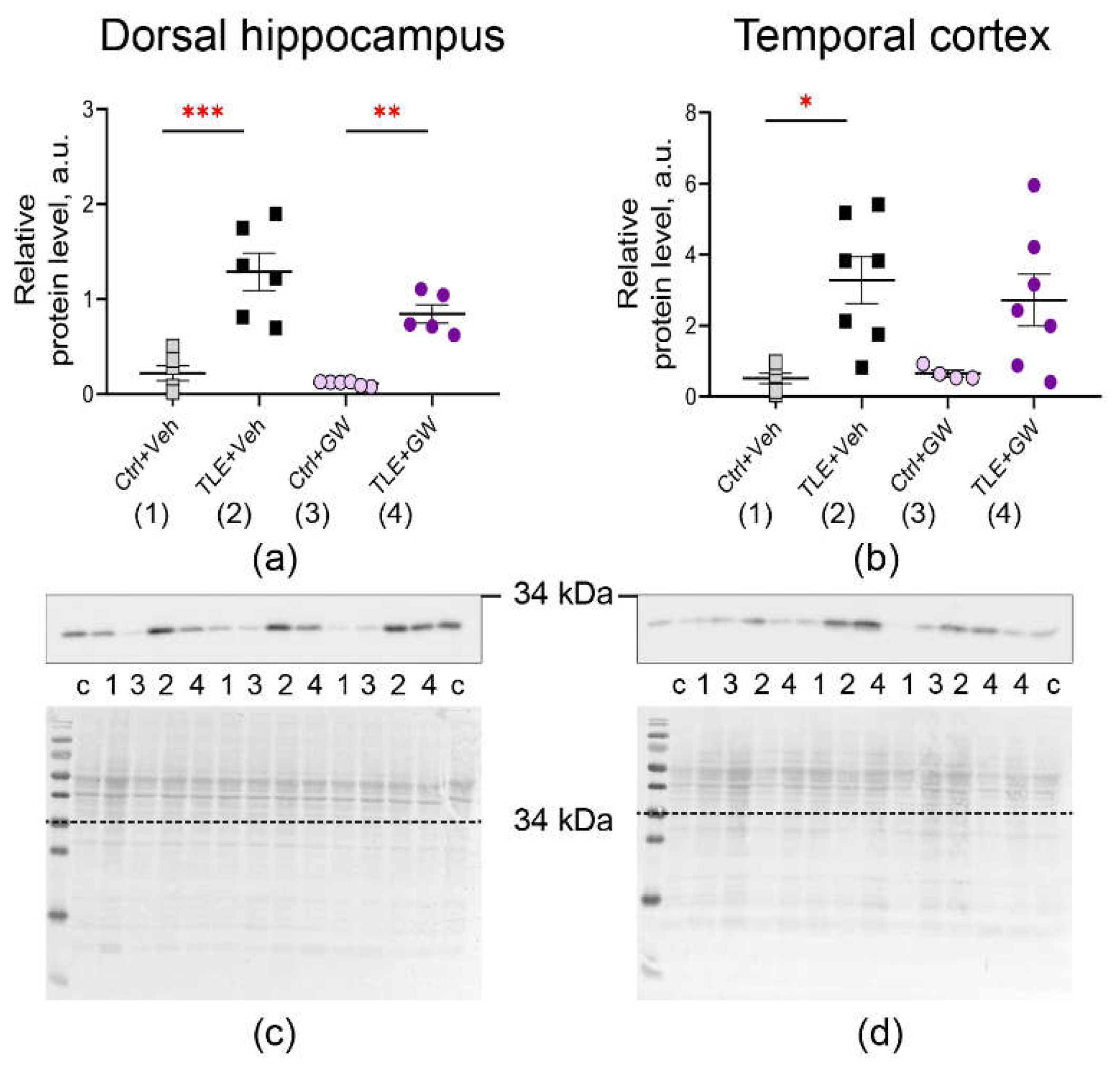
Figure 4.
Changes in relative protein levels of the microglial cell marker IBA1 in the dorsal hippocampus (a, c) and temporal cortex (b, d) of rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 4.
Changes in relative protein levels of the microglial cell marker IBA1 in the dorsal hippocampus (a, c) and temporal cortex (b, d) of rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05, ** p < 0.01, *** p < 0.001.
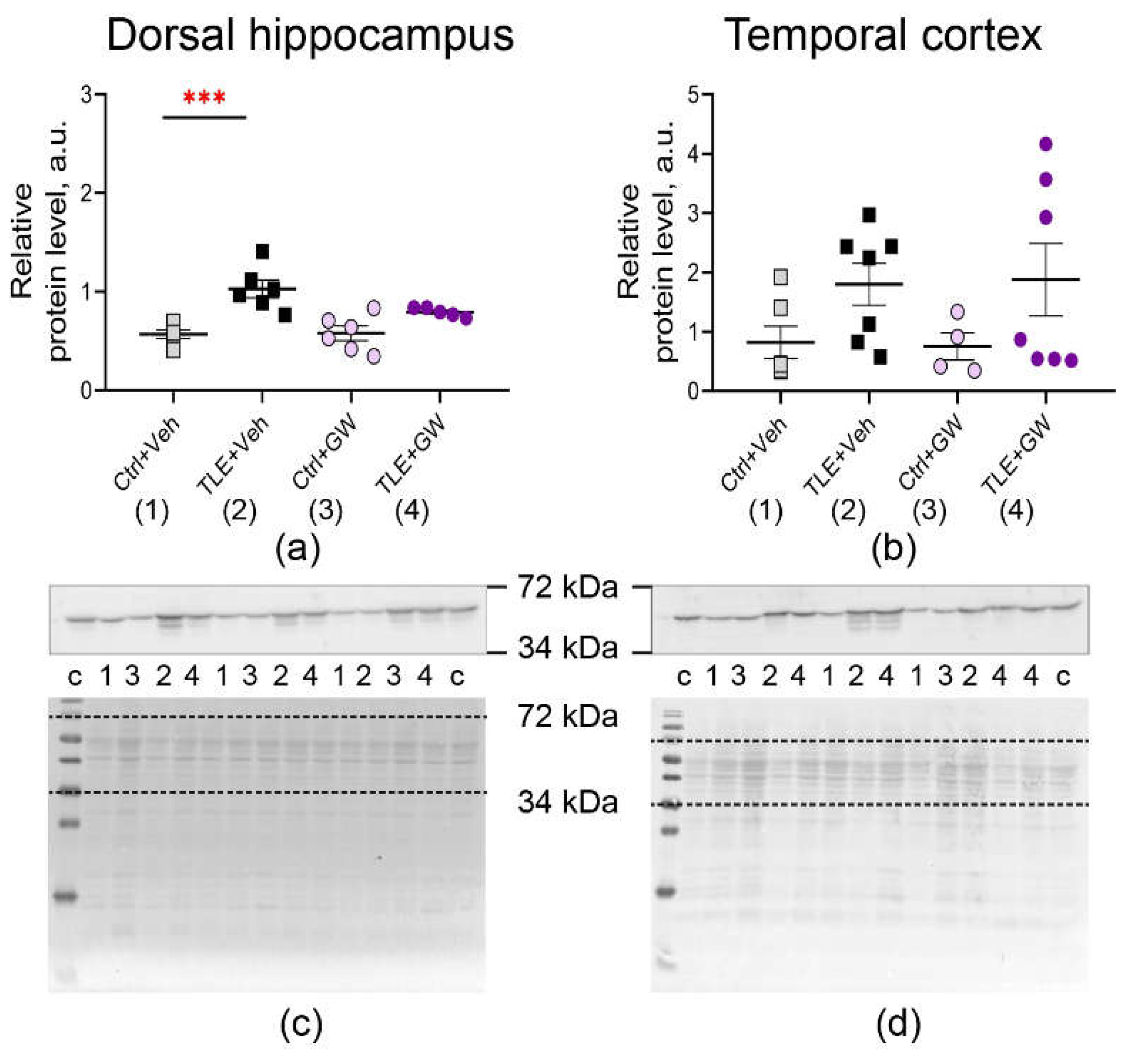
Figure 5.
Relative gene expression of pro-inflammatory cytokines Tnfa and Il1b (a-d) and anti-inflammatory cytokine Il1rn (e, f) and their ratio (g, h) in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05, *** p < 0.001.
Figure 5.
Relative gene expression of pro-inflammatory cytokines Tnfa and Il1b (a-d) and anti-inflammatory cytokine Il1rn (e, f) and their ratio (g, h) in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05, *** p < 0.001.
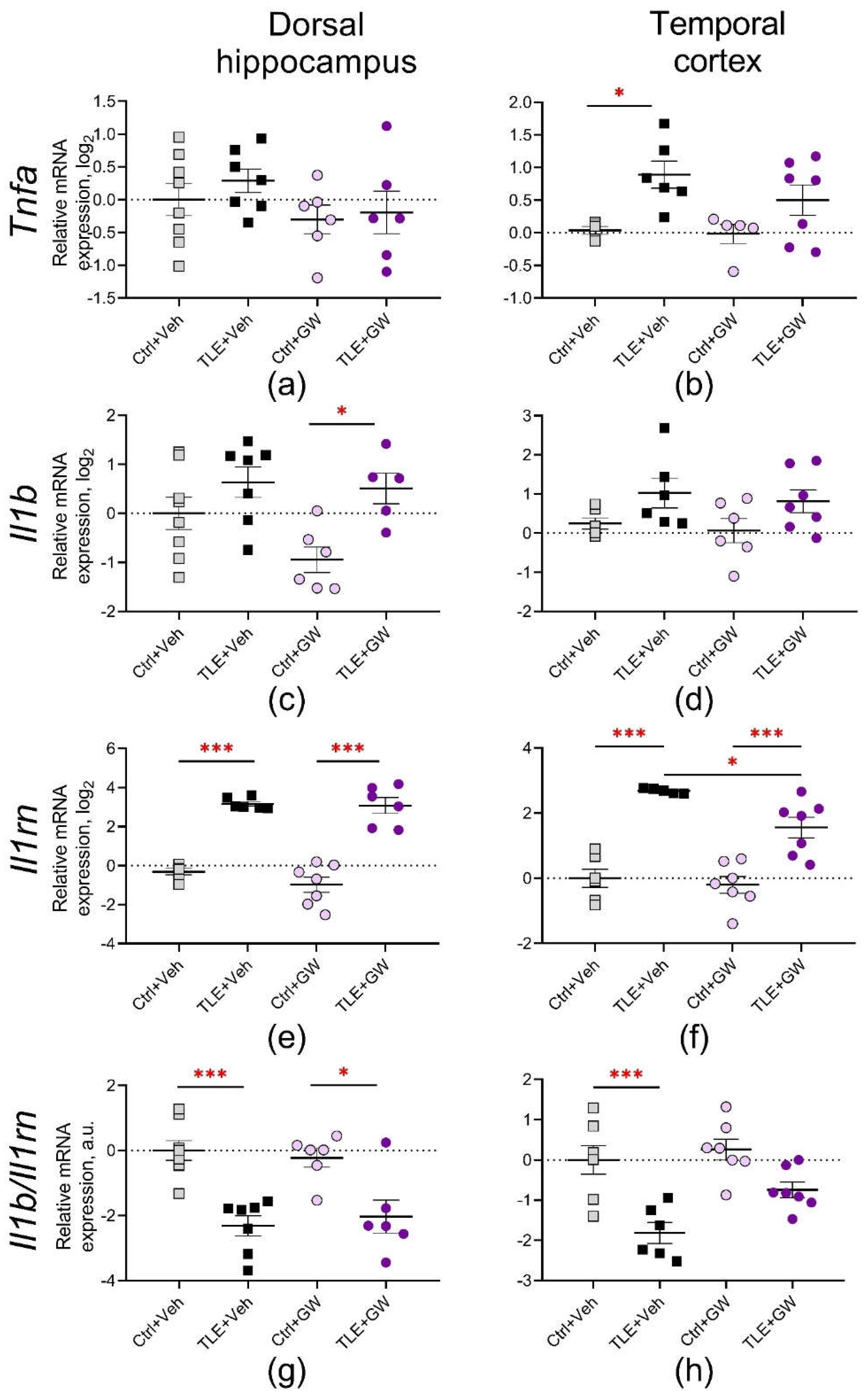
Figure 6.
Relative gene expression of microglial pro-inflammatory (a-d) and protective (e, f) proteins and their ratio (g, h) in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's posteriori test: * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 6.
Relative gene expression of microglial pro-inflammatory (a-d) and protective (e, f) proteins and their ratio (g, h) in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's posteriori test: * p < 0.05; ** p < 0.01; *** p < 0.001.
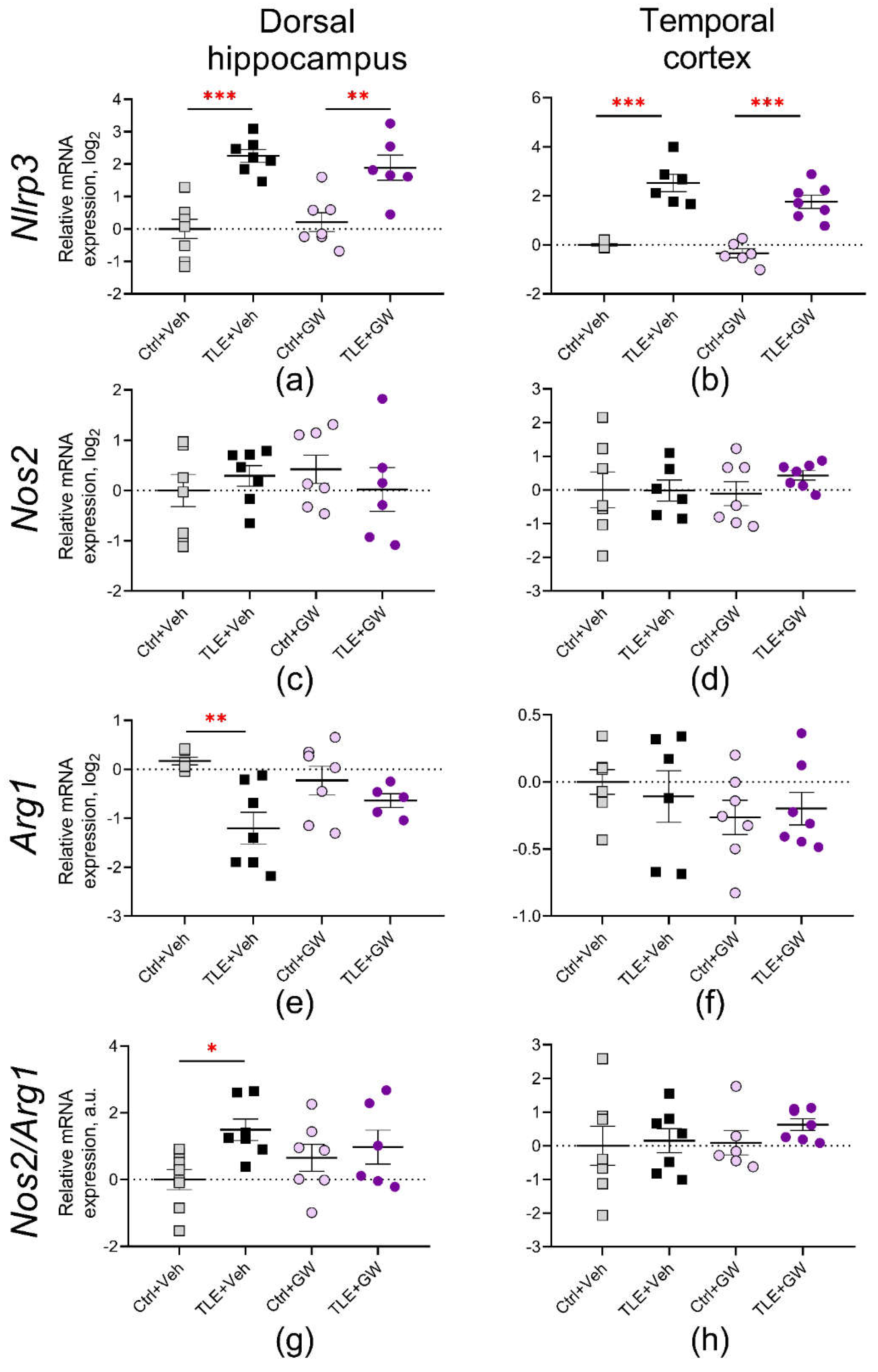
Figure 7.
Relative gene expression of astroglial proteins Lcn2 (a, b) and S100a10 (c, d) in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05; *** p < 0.001.
Figure 7.
Relative gene expression of astroglial proteins Lcn2 (a, b) and S100a10 (c, d) in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal. Asterisks indicate significant differences between groups according to Sidak's test: * p < 0.05; *** p < 0.001.
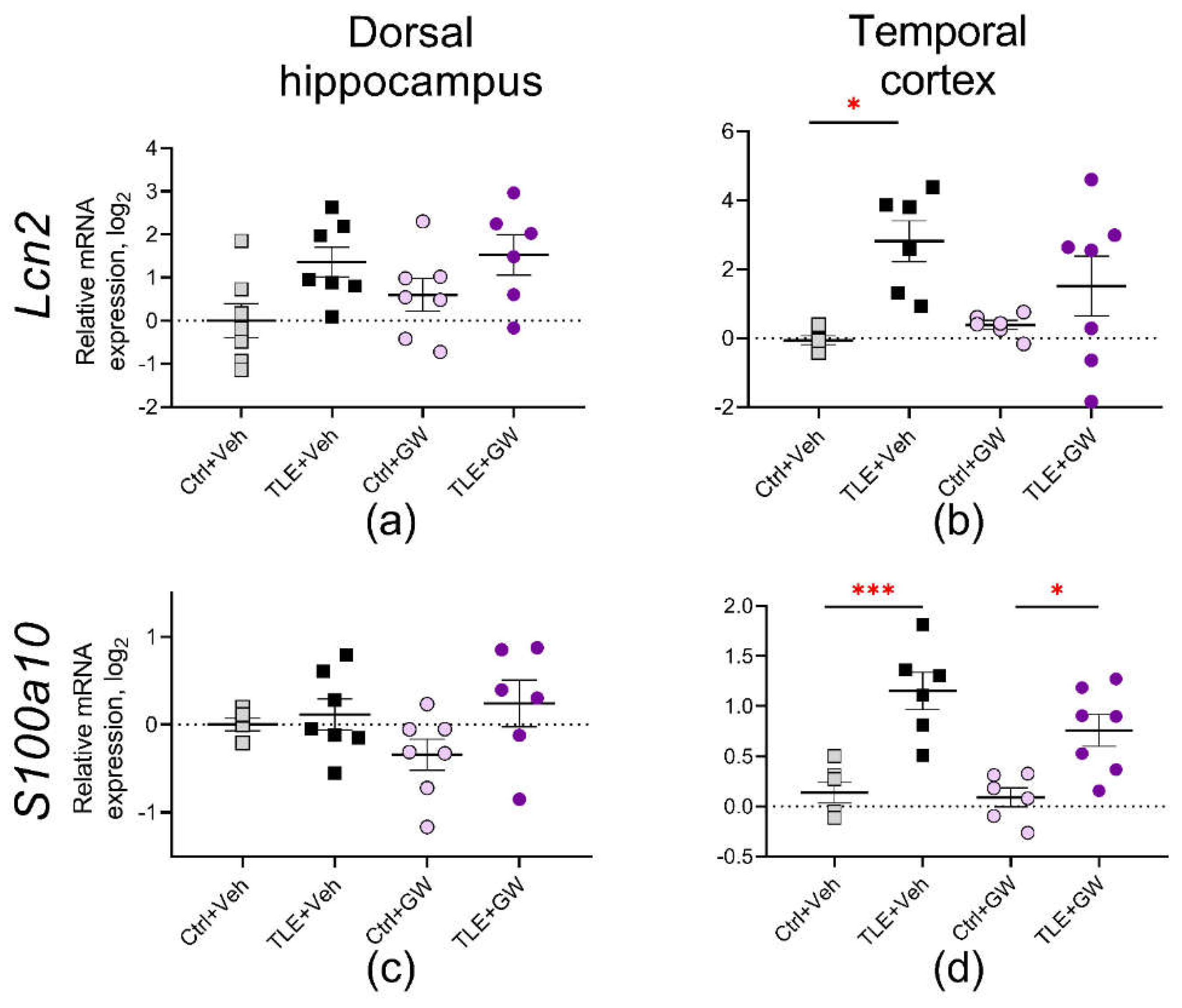
Figure 8.
Relative gene expression of astroglial proteins in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal.
Figure 8.
Relative gene expression of astroglial proteins in the dorsal hippocampus and temporal cortex of experimental and control rats. Ctrl+Veh, control rats without treatment; Ctrl+GW, GW0742-treated control rats; TLE+Veh, TLE rats; TLE+GW, GW0742-treated TLE rats. Data are presented as mean and standard error of the mean. Each point represents the value of a single animal.
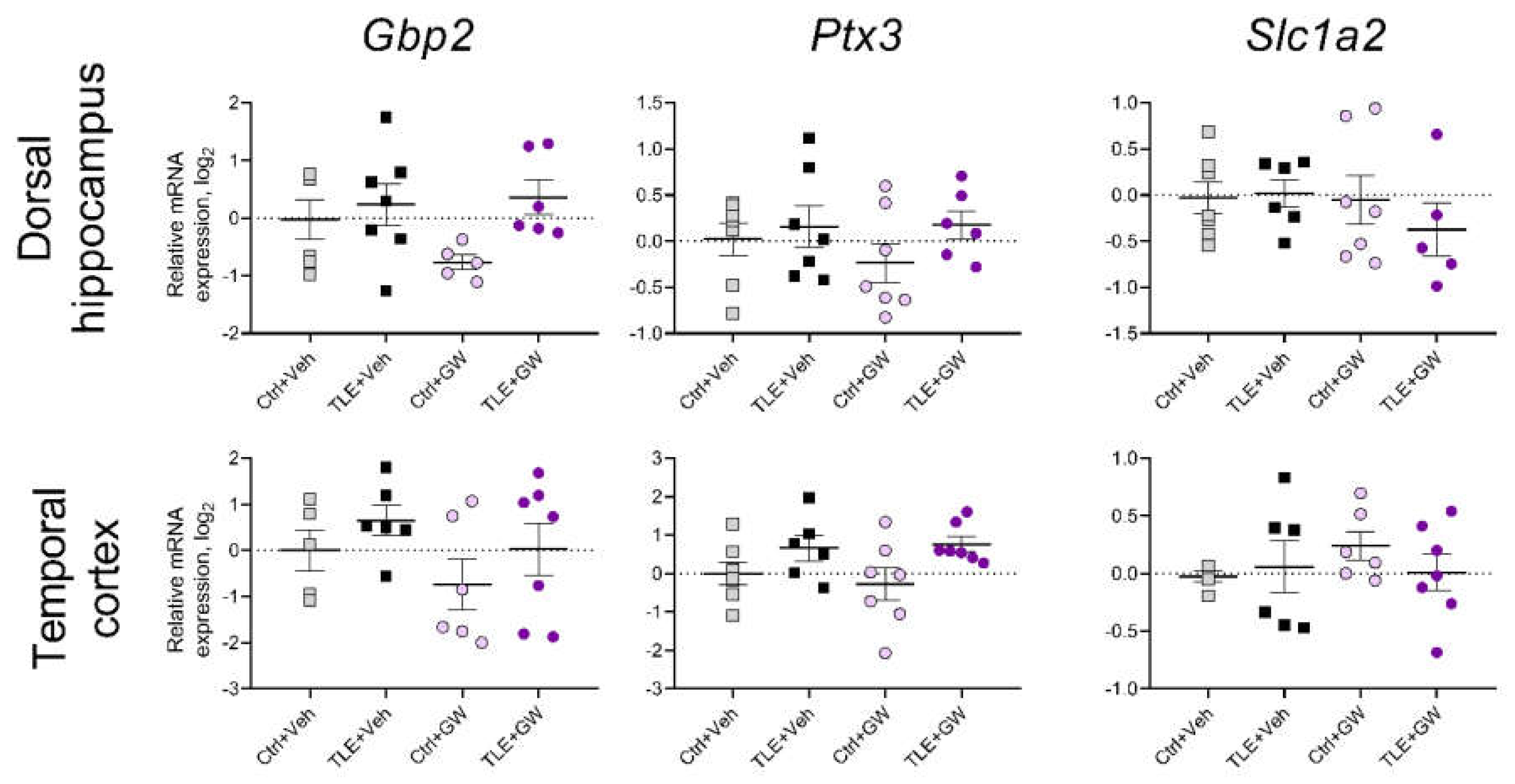
Figure 9.
Design of the experiment and scheme of brain regions dissection for analysis. TC - temporal cortex, DH - dorsal hippocampus, GW – GW0742.
Figure 9.
Design of the experiment and scheme of brain regions dissection for analysis. TC - temporal cortex, DH - dorsal hippocampus, GW – GW0742.
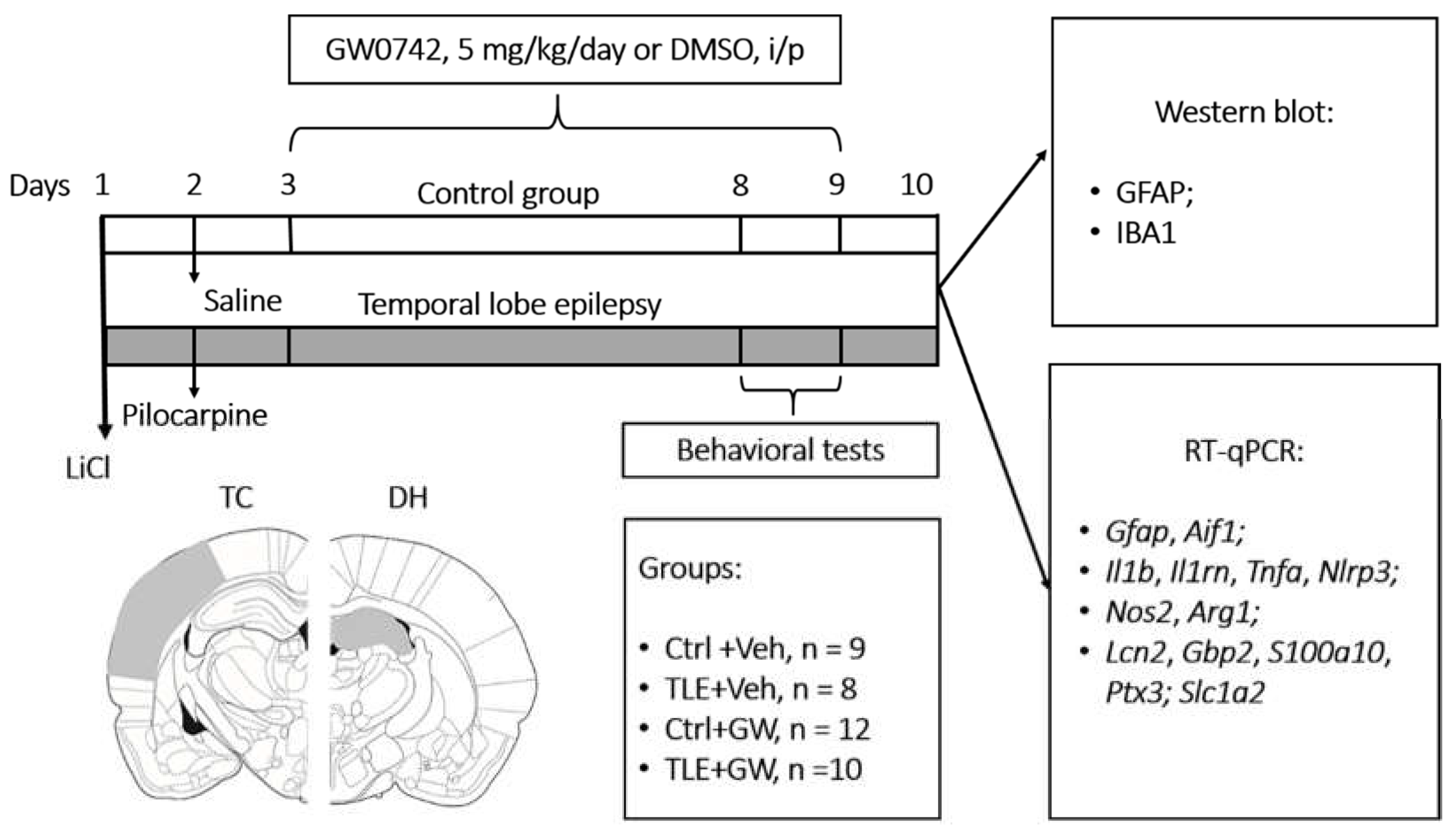
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



