
Submitted:
20 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Croton grewioides Baill., a species native to the Caatinga, popularly known as canela de cunhã, is used in traditional medicine to treat gastrointestinal diseases such as diarrhea, gastritis and stomach ulcers. This study aimed to identify the chemical composition of the essential oil of C. grewioides (OECG) and evaluate its antibacterial and antibiotic-modulating activities against both standard and multidrug-resistant bacteria. To analyze the compounds, present in the oil, gas chromatography coupled with mass spectrometry (GC/MS) and gas chromatography with flame ionization detector (GC/FID) techniques were used. In the bacteriological tests, the Minimum Inhibitory Concentration (MIC) was obtained using the broth microdilution technique. The modulating effect of the essential oil was determined by calculating the subinhibitory concentration followed by serial microdilution of the antibiotics. The MIC reduction factor (FRC) was calculated and its data expressed as a percentage. Analysis of the chemical composition identified the presence of the major compound estragole with a relative abundance of 50.34%. The MIC values obtained demonstrated efficacy in K. pneumoniae isolated from urine with MIC values of 512 µg/mL. OECG potentiated the effects of all antibiotics tested on S. aureus ATCC 29213, K. pneumoniae Carbapnemase, E. coli ATCC 25922 and S. aureus ATCC 29213 strains with their FRC of 97.65 %, 99.80 %, 99.85 % and 99.88 %, respectively. Thus, the essential oil of C. growioides presents synergistic effects when combined with the tested antibiotics, in addition to acting in the fight against bacterial resistance to antibiotics.
Keywords:
antibacterial activity
; Croton grewioides Baill
; essential oil
; synergistic effect
1. Introduction
The discovery of antibiotics led to a reduction in mortality rates caused by bacterial infections in hospital settings. However, the widespread and uncontrolled use of these drugs has promoted the emergence of resistant microorganisms [1], making research into natural products a promising alternative for combating microbial resistance [2].
In Brazil, the native fauna and flora have been used since before colonization by native peoples to treat illnesses, accumulating enormous knowledge of medicinal flora [3]. Despite having one of the greatest biodiversities in the world, research on native products of Brazilian flora is relatively underdeveloped [4]. Among the Brazilian biomes, the Caatinga encompasses a large number of plant species with therapeutic potential, but with few studies focused on the area of chemistry. Therefore, research on species from the Caatinga is still in its infancy and their use is reserved, in most cases, for use by traditional and local communities [5].
The species Croton grewioides Baill. (Euphorbiaceae), commonly known as canela de cunhã, is widely distributed throughout the Nordeste region of Brazil and is characteristic of the Caatinga biome [6]. Its use in traditional medicine is through teas and infusions of fresh leaves, being commonly used in the treatment of gastrointestinal disorders such as diarrhea, gastritis and in the treatment of stomach ulcers, in addition to being used as a sedative agent, relieving pain symptoms and having a calming effect [7, 8, 9, 10, 11]. Its anti-inflammatory, hepatoprotective and insecticidal effect is also reported, being used by small and medium-sized farmers for pest control [12]. These reported activities are likely associated with the chemical constituents anethole and estragole, which are present in the essential oil of the leaves of C. grewioides.
Given the high prevalence of antibiotic-resistant microorganisms and the limited microbiological studies on C. grewioides, this study aimed to identify the chemical composition of the essential oil of C. grewioides and evaluate its antibacterial and antibiotic-modulating activities against both standard and multidrug-resistant bacteria.
2. Materials and Methods
2.1. Collection and obtaining of essential oil
Aerial parts of C. grewioides were collected in the municipality of Crato, Ceará - Brazil, between August and September 2022 in the morning. A specimen was deposited in the Herbarium Caririense Dárdano de Andrade Lima of the Regional University of Cariri, where it was assigned registration number 1619. The essential oil was obtained by hydrodistillation (Clevenger type apparatus), where the sample of the dry plant material (2 kg) was crushed and subjected to distillation for 4h. The obtained oil was dried over Na₂SO₄, yielding 0.20% (w/w).
2.2. GC-MS analysis
The essential oil was analyzed using a Shimadzu GC–MS QP2010 series, provided by Shimadzu Scientific Instruments Inc. (Columbia, MD, USA), with fused silica capillary column SH-Rtx-5 (30 m × 0.25 mm I.D.; 0.25 m film thickness) and the following temperature program: 80–180 ºC at 4 ºC/min, then to 246 ºC at 6.6 ºC/min, closing with 10 min at 280 ºC, at 3.4 ºC/min, totaling analysis time of 30 min. He was used as the carrier gas, flow rate of 1.5 mL/min, split mode (1:100), and injection port was set at 220 ºC. The quadrupole MS operating parameters: interface temperature (280 ºC) and ion source (200 ºC); electron impact ionization at 70 eV; scan mass range of 40–350 m/z with sampling rate of 1.0 scan/s. Injection volume: 1 µL of 500 ppm solution prepared with dichloromethane. The constituents were identified by computational search using digital libraries of mass spectral data (NIST 08) and by comparing their authentic mass spectra and reported in the literature [13].
2.3. GC-FID analysis
The essential oil was analyzed using a Shimadzu GC-2010 Plus equipped with flame ionization detector (FID), provided by Shimadzu Scientific Instruments Inc. (Columbia, MD, USA), with fused silica capillary column SH-Rtx-5 (30 m × 0.25 mm I.D.; 0.25 m film thickness) and the following temperature program: 80–180 ºC at 4 ºC/min, then to 246 ºC at 6.6 ºC/min, closing with 10 min at 280 ºC, at 3.4 ºC/min, totaling analysis time of 30 min. He was used as the carrier gas, flow rate of 1.5 mL/min, split mode (1:15), and injection port was set at 220 ºC. Injection volume: 1 µL of 500 ppm solution prepared with dichloromethane. The detector temperature was set to 300 °C. The linear retention index was obtained by injecting a mixture of C8-C40 linear hydrocarbons under the same conditions as the samples, as described by Van Den Dool and Kratz (1963). The identity of the compounds was confirmed by comparing their retention indices and mass spectra with those taken from the literature [13].
2.4. Antibacterial activity
2.4.1. Determination of minimum inhibitory concentration (MIC)
To determine the antibacterial activity, the broth microdilution method containing the Brain Heart Infusion Broth (BHI) culture medium was used based on the CLSI document [14]. The following strains were used: Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 29213, Klebsiella pneumoniae Clinical Urine Isolate (ICU), Escherichia coli 06, Staphylococcus aureus 10, Klebsiella pneumoniae Carbapnemase (KPC) and Klebsiella pneumoniae Betalactamase (BLSE).
The essential oil of C. grewioides (OECG) was prepared at a concentration of 1024 µg/mL diluted in sterile water and DMSO. In the assay, serial microdilutions of 100 µL of OECG were performed where the control group contained only the inoculum. The tests were conducted in triplicate and incubated in the microbiological incubator at 37º C for 24 h. The readings were performed by the colorimetric method using 25 µL of 0.01% sodium resazurin [14].
2.4.2. Antibiotic modulation tests
The antibiotics used to evaluate the modulating activity of OECG were amoxicillin, penicillin, gentamicin and amikacin. The methodology for performing the test was adapted from Coutinho et al. [15]. Based on the calculation of the subinhibitory concentrations (MIC/8), serial microdilution of the respective antibiotics was performed. The control group contained only OECG and the inoculum. The plates were placed in an oven at 37°C for 24 h and then bacterial growth was evaluated by the colorimetric method with sodium resazurin (0.01%).
2.4.3. Determination of the MIC reduction factor
To determine the MIC Reduction Factor (FRC) expressed as a percentage, the value of X was calculated using the formula below. To do this, a simple rule of three was used, which obtained the following conclusions: X is equal to the values of the test MIC (which contains the essential oil together with the antibiotic) multiplied by the value of 100 divided by the values of the control MIC (antibiotics and bacteria). Thus, the values obtained in the CRF calculation represent the percentage of reduction in the MIC values in the presence of the essential oil.
FRC = 100 - X
X = CIM test x 100 / CIM control
FCR = 100 – (CIM test x 100 / CIM control)
2.4.4. Statistical analysis
The tests were analyzed by two-way ANOVA followed by the Bonferroni test using GranphPad Prism 10.0 software. Results expressed at p<0.05 were considered statistically significant.
3. Results and discussion
3.1. Composition of C. grewioides essential oil
The evaluation of the chemical composition of the essential oil extracted from the aerial parts of C. grewioides revealed the presence of 2.92 % of monoterpenoid compounds, 5.45 % of oxygenated monoterpenes, 4.40 % of sesquiterpenes, 0.42 % of oxygenated sesquiterpenes, 69.73 % of phenylpropanoids and 10.99 % of ethers. The major compound identified in the chemical analysis was estragole with 50.34 %, followed by the compounds methyl eugenol with 19.39 % and anisole with 10.99 %. In total, 18 compounds were identified by chromatography analysis (Figure 1 and Figure 2), whose percentages are described in Table 1.
In the study by Santos et al. [16] that evaluated the chemical composition of essential oils from three chemotypes of C. grewioides extracted from branches, in the first chemotype the major component was (E)-anethole with 70.5 %. In the second chemotype, eugenol with 84.2 %. Finally, in the third chemotype, (Z)-methyl with 53.4 % was presented. In another study carried out by Silva et al. [17], the composition of the essential oil of C. grewioides extracted from branches revealed (E)-anethole as the major component with 47.8 %. Comparing these studies with the components identified in this research, the absence of the compound (E)-anethole in the chromatographic analyses is demonstrated, which can be explained by environmental factors such as collection site, collection period, part of the plant used, vegetative state of the plant, genetic factors, among others [18, 19].
Studies evaluating the chemical composition of the essential oil extracted from the leaves of C. grewioides, reported by Costa et al. [20] and Siqueira et al. [21], reveal the presence of estragole with 76.8 % and 45.95 % respectively as the major components present in its oil. As in the studies by Leite et al. [22] and Andrade et al. [8], its percentage present in the essential oil is 78 % and 84.7 %, whose estragole compound and its percentages are close to those found in this study.
The variation in the chemical composition of essential oils is likely a result of the plant's adaptive mechanisms, which utilize secondary metabolites to defend against predators and to adapt to variations in both biotic and abiotic environmental conditions [23].
3.2. Antibacterial activity
The antibacterial activity of C. grewioides essential oil tested on standard strains demonstrated significant results against the K. pneumoniae (ICU) strain with an MIC value of 512 µg/mL. In multidrug-resistant strains, the oil presented better results on the K. pneumoniae (KPC) and K. pneumoniae (BLSE) strains, with MIC values of 512 and 256 µg/mL, respectively. The other MIC values of the strains used in this study are described in Table 2.
In the strains tested, there was no inhibition of bacterial growth even at a concentration of 1024 µg/mL of OECG, this was due to the fact that the strains tested present biochemical mechanisms related to bacterial resistance, which implies a high degree of pathogenicity, since the strains tested in this study are involved in various infections in hospitalized patients, mainly due to urinary tract infections, as is the case with strains of E. coli and K. pneumoniae [24].
Studies such as those by Rodrigues and collaborators [25], which demonstrate the antibacterial potential of C. grewioides essential oil against standard strains of Pseudomonas aeruginosa ATCC 15442 and S. aureus ATCC 12692, obtaining minimum inhibitory dose (MID) results of 1 and 0.5 mg/L of air in S. aureus. These findings corroborate the results obtained in this research, suggesting that compounds present in the essential oil, such as estragole and anethole, which are commonly reported in chemical studies, are directly related to the antibacterial effects observed in the tests.
3.3. Modulating activity
The beta-lactam antibiotics used in the study were amoxicillin and penicillin, which have a mechanism of action associated with the inhibition of bacterial cell wall synthesis, leading to destabilization of the cell wall's macrostructure and leakage of intracellular fluid due to membrane lysis. The aminoglycoside antibiotics were amikacin and gentamicin, which act by inhibiting the protein synthesis mechanism specifically in the 30S subunit of bacterial ribosomes [26].
From the results obtained in the modulation tests, OECG enhanced the effects of amoxicillin on strains of S. aureus 10, K. pneumoniae (ICU) and S. aureus ATCC 29213, with their respective MIC Reduction Factors (FRC) of 85 %, 84.16 % and 97.65 %. As for penicillin, it presented results on strains of S. aureus 10 and S. aureus ATCC 29213, with its most potent effects on K. pneumoniae (KPC) with a FRC of 99.95 %, E. coli 06 with 99.80 % and K. pneumoniae (ICU) with 98.02 %. The remaining results are detailed in Table 3.
When associated with gentamicin, OECG demonstrated promising results in both standard and multidrug-resistant strains, except for K. pneumoniae (KPC), which presented negative FRC, indicating an antagonistic effect. Of these results, the standard strains obtained FRC of 99.85 % in E. coli ATCC 25922, 99.80 % in S. aureus ATCC 29213 and 99.75 % in K. pneumoniae (ICU). As for amikacin, OECG was more effective against S. aureus ATCC 29213 with FRC of 99.88 %.
In the graph shown in Figure 3, the modulating effect of OECG associated with the antibiotic amoxicillin against the strains used in this study, a synergistic effect was observed in the multiresistant strain S. aureus 10, and in all standard strains tested. In the other strains tested, an antagonistic effect was observed in S. aureus 10, E. coli 06, K. pneumoniae (BLSE), K. pneumoniae (KPC).
OECG also showed synergistic effects in association with penicillin. Figure 4 demonstrates the synergistic effects in all strains tested, with the exception of K. pneumoniae (BLSE) and E. coli ATCC 29213, which showed equivalence of means. One of the main mechanisms of bacterial resistance to beta-lactam antibiotics is the production of the beta-lactamase enzyme that hydrolyzes the beta-lactam ring of these antibiotics, inhibiting their action [27]. The lack of effects of OECG associated with penicillin against K. pneumoniae (BLSE) may be related to its resistance mechanism of the type of producer of the beta-lactamase enzyme that acts to inhibit the action of these drugs [28].
Carbapinemase-producing bacteria exhibit a broad resistance mechanism against several beta-lactam antibiotics, acting in the hydrolysis of drugs such as penicillins, cephalosporins, monobactams and carbapenems [29, 30]. In this study, OECG potentiated the effect of penicillin, giving evidence that it may be involved in increasing membrane permeability, in which the entry of the drug into the cell is facilitated, in addition to acting at the genetic level, altering its resistance mechanism, since the drug is able to enter the cell, overcoming enzymatic barriers and inhibiting peptidoglycan synthesis, leading to rupture and extravasation of intracellular content [31].
Amoxicillin, which also acts to inhibit cell wall synthesis, showed synergism in standard strains and multiresistant S. aureus 10. The mechanism is similar to that of penicillin. Furthermore, more effective inhibition by the drug was expected in standard strains, since these do not have as many resistance mechanisms as multiresistant strains.
In corroboration with these data, Medeiros et al. [32] evaluated the effects of C. grewioides essential oil on the modulation of the antibiotics norfloxacin and tetracycline against Staphylococcus aureus (SA-1199B) with efflux pump resistance mechanism, and demonstrated that in the absence of the oil, the antibiotic presents an inhibitory concentration of 64 µg/mL for norfloxacin and 32 µg/mL for tetracycline. In the presence of the oil, these values dropped to 16 µg/mL for norfloxacin and 0.5 µg/mL for tetracycline, enhancing the effect of the antibiotic even in the presence of the overexpressed gene for efflux pump.
The effect of OECG against aminoglycoside antibiotics showed that amikacin (Figure 5) presented synergistic effects in all strains tested except K. pneumoniae (BLSE), which presented an antagonistic effect, and K. pneumoniae (KPC), which demonstrated equivalence to the control. As for gentamicin (Figure 6), synergistic effects were observed in all strains tested, except for K. pneumoniae (KPC), which exhibited an antagonistic effect.
Protein synthesis inhibitors that act on bacterial ribosomes in the 30S and 50S subunits are mainly classified as aminoglycosides, tetracyclines, and aphenicols, among others. These antibacterial drugs have greater selectivity precisely because they target components of the bacterial ribosome that are distinct from those of eukaryotic ribosomes [26]. For amikacin, promising results were observed in both standard and multidrug-resistant strains, with synergistic effects reported in all standard strains as well as in the multidrug-resistant E. coli 06 and S. aureus 10.
Essential oils, due to their hydrophobic nature, interfere in essential biochemical processes for the cell when they come into contact with bacterial membranes, such as the respiratory chain and energy production, which leads to the deregulation of membrane permeability, facilitating the entry of antibiotic drugs, in addition to interfering in the efflux pump resistance mechanism, since this requires the binding of the adenosine triphosphate (ATP) molecule for its functioning in the “expulsion” of toxic substances [25, 33].
Linked to the hydrophobic character of essential oils, their effects on the antibacterial and modulating activity reported in this research may be related to the various compounds of the terpene class present in their composition that have an intrinsic relationship to this activity [34].
4. Conclusions
The data obtained in this research allow us to conclude that OECG has synergistic effects when associated with the antibiotics amoxicillin, penicillin, amikacin and gentamicin against all strains tested, with the most potent effects observed in E. coli 06, K. pneumoniae (KPC), S. aureus ATCC 29213 and E. coli ATCC 25922. Thus, OECG has the potential to be used in future antimicrobial therapies, acting as a modulating agent in the fight against bacterial resistance to antibiotics. However, further research on its action on bacterial membrane permeability, genetic alterations of its resistance mechanism, in addition to analysis of isolated compounds are necessary to verify whether the efficacy of the essential oil is linked to the major compound and thus be able to use it in future antibacterial treatments.
Author Contributions
Author Contribution Statement: Research, writing, formatting, methodology, investigation, J. J.F,V.; methodology, investigation, C.J.C.; formal analysis, data curation, N.K.G.C.; formal analysis, data curation, writing, J.B.N.; methodology, investigation, M.I.S.; methodology, investigation, M.P.S.; formal analysis, data curation, J. W. G. C.; methodology, investigation, G.G.O.S.; methodology, investigation, validation, writing, F.F.G.R.; conceitualization, methodology, supervision, writing, J.G.M.C.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
This work was carried out at the Laboratório de Pesquisa de Produtos Naturais (LPPN) of the Departamento de Química Biológica at the Universidade Regional do Cariri (URCA), with support from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Financiadora de Estudos e Projetos (FINEP), Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP), Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA) and Instituto Nacional de Ciência e Tecnologia – Alimentos (INCT-ALIM).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Costa, A. R., Lima Silva, J., Lima, K. R. R., Rocha, M. I., Barros, L. M., da Costa, J. G. M., ... & Coutinho, H. D. M. Rhaphiodon echinus (Nees & Mart.) Schauer: Chemical, toxicological activity and increased antibiotic activity of antifungal drug activity and antibacterial. Microb. Pathog., v. 107, p. 280-286, 2017. [CrossRef]
- Silva, Líllian Oliveira Pereira; Nogueira, Joseli Maria da Rocha. Resistência bacteriana: potencial de plantas medicinais como alternativa para antimicrobianos. Rev. bras. anal. clin, p. 21-27, 2021.
- <named-content content-type="background:white">Marisco, G. Marisco, G., & Rocha, R. Estudos etnobotânicos em comunidades indígenas no Brasil. 2016. https://www.arca.fiocruz.br/handle/icict/19250.
- Souza, R. K. D. , da Silva, M. A. P., de Menezes, I. R. A., Ribeiro, D. A., Bezerra, L. R., & de Almeida Souza, M. M. Ethnopharmacology of medicinal plants of carrasco, Nordeste Brazil. Journal of Ethnopharmacology, v. 157, p. 99-104, 2014.
- Oliveira, S. D. D. S. , De Oliveira E Silva, A. M., Blank, A. F., Nogueira, P. C. D. L., Nizio, D. A. D. C., Almeida-Pereira, C. S., Pereira, R.O, Menezes-Sá, T. S. A., Santana, M. H. S. & Arrigoni-Blank, M. D. F. Radical scavenging activity of the essential oils from Croton grewioides Baill accessions and the major compounds eugenol, methyl eugenol and methyl chavicol. J. Essent. Oil Res. v. 33, n. 1, p. 94-103, 2021. [CrossRef]
- Lima, F. C. , Sousa, D. F., Ferreira, J. M., Lima Jr, R. C., Tomé, A. R., Cardoso, J. H. L., Queiroz, M. G. R. & Campos, A. R. Croton zehntneri Essential Oil Prevents Acetaminophen-Induced Acute Hepatotoxicity in Mice. v. 2, n. 4, 2008.
- Sousa, A. J. , Oliveira, G. L., Fonseca, L., Rocha, M. S., Rai, M., Santos, F. E., & Lima, S. G. D. Antioxidant properties of Croton zehntneri Pax et Hoffm. Essential oil and its inclusion complex with β-cyclodextrin prepared by spray drying. Journal of the Brazilian Chemical Society, v. 33, p. 1244-1253, 2022.
- Andrade, T. C. , Lima, S. G., Freitas, R. M., Rocha, M. S., Islam, T., SILVA, T. G., & Militao, G. C.Isolation, characterization and evaluation of antimicrobial and cytotoxic activity of estragole, obtained from the essential oil of Croton zehntneri (euphorbiaceae). An. Acad. Bras. Ciênc. v. 87, p. 173-182, 2015. [CrossRef]
- Coelho-de-Souza, A. N. , Lahlou, S., Barreto, J. E., Yum, M. E., Oliveira, A. C., Oliveira, H. D., Celedônioa, N.R., Feitosaa, R.G.F., Duarte, G.P., Santosa, C.F., De Albuquerque A.A.C. & Leal-Cardoso, J. H. Essential oil of Croton zehntneri and its major constituent anethole display gastroprotective effect by increasing the surface mucous layer. Fundamental & clinical pharmacology, v. 27, n. 3, p. 288-298, 2013. [CrossRef]
- Cabral, P. H. B. , de Morais Campos, R., Fonteles, M. C., Santos, C. F., Cardoso, J. H. L., & do Nascimento, N. R. F. Effects of the essential oil of Croton zehntneri and its major components, anethole and estragole, on the rat corpora cavernosa. Life sci, v. 112, n. 1-2, p. 74-81. [CrossRef]
- Sá, N. A. R. , Bruno, J. B., Guerreiro, D. D., Cadenas, J., Alves, B. G., Cibin, F. W. S., Leal-Cardoso, J.H., Gastal, E.L. & Figueiredo, J. R. Anethole reduces oxidative stress and improves in vitro survival and activation of primordial follicles. Braz J Med Biol Res. v. 51, 2018. [CrossRef]
- Cavalcanti, J. M. , Leal-Cardoso, J. H., Diniz, L. R. L., Portella, V. G., Costa, C. O., Linard, Alves, C.K., Rocha, M.V.A.P., Lima, C.C., Cecatto, V.M.& Coelho-de-Souza, A. N. The essential oil of Croton zehntneri and trans-anethole improves cutaneous wound healing. J. Ethnopharmacol, v. 144, n. 2, p. 240-247, 2012. [CrossRef]
- Adams, R.P. , 2001. Identification of Essential Oil Components by Gas Chromatography/ Quadrupole Mass Spectroscopy. Allured Pub, Corp, Illinois.
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-. Ninth Edition. CLSI document Clinical and Laboratory Standards Institute, 2012.
- Coutinho, H. D. , Costa, J. G., Lima, E. O., Falcão-Silva, V. S., & Siqueira-Júnior, J. P. Enhancement of the antibiotic activity against a multiresistant Escherichia coli by Mentha arvensis L. and chlorpromazine. Chemotherapy, v. 54, n. 4, p. 328-330, 2008.
- Santos, H. S. , Furtado, E. F., Bertini, L. M., Bandeira, P. N., Albuquerque, M. R., Menezes, J. E., Trevisan, M. T. S. & Lemos, T. L. Chemical composition and cholinesterase inhibition of essential oils of three chemotypes from Croton zehntneri. Rev. Latinoam. Quím, v. 38, n. 1, p. 45-51, 2010. [CrossRef]
- Silva, C. G. , Zago, H. B., Júnior, H. J., da Camara, C. A., de Oliveira, J. V., Barros, R., Schwartz, M. O. E. & Lucena, M. F. Composition and insecticidal activity of the essential oil of Croton grewioides Baill. against Mexican bean weevil (Zabrotes subfasciatus Boheman). J. Essent. Oil Res, v. 20, n. 2, p. 179-182. 2008. [CrossRef]
- Ribeiro, S. M. , Bonilla, O. H., & Lucena, E. M. P. Influência da sazonalidade e do ciclo circadiano no rendimento e composição química dos óleos essenciais de Croton spp. da Caatinga. Iheringia, Sér. Bot. v. 73, n. 1, p. 31-38, 2018. [CrossRef]
- Morais, Lilia Aparecida Salgado. Influência dos fatores abióticos na composição química dos óleos essenciais. Hortic Bras, v. 27, n. 2, p. S4050-4063, 2009. http://www.alice.cnptia.embrapa.br/alice/handle/doc/577686.
- Costa, J. G. , Rodrigues, F. F., Angélico, E. C., Pereira, C. K., Souza, E. O. D., Caldas, G. F.R., Silva, M. R., Santos, N. K. A., Mota, M.L. & Santos, P. F. D. Composição química e avaliação da atividade antibacteriana e toxicidade do óleo essencial de Croton zehntneri (variedade estragol). Rev. Bras. Farmacogn, v. 18, p. 583-586, 2008. [CrossRef]
- Siqueira, R. J. B. , Leal-Cardoso, J. H., Couture, R., & Lahlou, S. Role of capsaicin-sensitive sensory nerves in mediation of the cardiovascular effects of the essential oil of Croton zehntneri leaves in anaesthetized rats. Clin. Exp. Pharmacol. Physiol. v. 33, n. 3, p. 238-247, 2006. [CrossRef]
- Leite, G. D. O. , Penha, A. R. S., Costa, J. G. M. D., & Campos, A. R. Anxiogenic like effect of Croton zehntneri Pax et Hoffm leaves essential oil in mice. J. Essent. Oil-Bear. Plants, v. 12, n. 5, p. 546-550, 2009. [CrossRef]
- Taveira, F. S. N. , De Lima, W. N., Andrade, E. H. A., & Maia, J. G. S. Seasonal essential oil variation of Aniba canelilla. Biochem. Syst. Ecol. v. 31, n. 1, p. 69-75, 2003. [CrossRef]
- Oliveira, A. L. D. , Soares, M. M., Santos, T. C. D., & Santos, A. Mecanismos de resistência bacteriana a antibióticos na infecção urinária. UNINGÁ Rev. v. 20, n. 3, 2014.
- Rodrigues, Fabíola FG; Costa, JGM; Coutinho, HDM. Synergy effects of the antibiotics gentamicin and the essential oil of Croton zehntneri. Phytomedicine, v. 16, n. 11, p. 1052-1055, 2009. [CrossRef]
- Scherer, C. B. , Botoni, L. S., & Costa-Val, A. P. Mecanismos de ação de antimicrobianos e resistência bacteriana. Veterinária, v. 4, n. 13, p. 12-20, 2016.
- Arruda, C. J. M. , de Almeida Siqueira, V. F., de Souza, F. J. M., das Neves Silva, J. L., Ferreira, K., & Faro, A. Revisão bibliográfica de antibióticos beta-lactâmicos. Revista Saúde em Foco, 2019. [Google Scholar]
- Cornaglia, G. , Giamarellou, H., & Rossolini, G. M. Metallo-β-lactamases: a last frontier for β-lactams? Lancet Infect. Dis, v. 11, n. 5, p. 381-393, 2011. [CrossRef]
- Queenan, A. M. , & Bush, K. Carbapenemases: the versatile β-lactamases. Clin Microbiol Rev, v. 20, n. 3, p. 440-458, 2007. [CrossRef]
- Nogueira, H. S. , de Oliveira Xavier, A. R. E., de Sousa Xavier, M. A., Carvalho, A. A., Monção, G. A., & Barreto, N. A. P. Antibacterianos: principais classes, mecanismos de ação e resistência. Unimontes Cient. v. 18, n. 2, p. 96-108, 2016.
- Monge, K. M. M. (2013). Carbapenémicos: tipos y mecanismos de resistência bacterianos. Rev. méd. Costa Rica Centroam. 70(608), 599-605.
- Medeiros, V. M. , do Nascimento, Y. M., Souto, A. L., Madeiro, S. A. L., de Oliveira Costa, V. C., Silva, S. M. P., Silva, V. S. F., Agra, M. F., Siqueira-Júnior J. P. & Tavares, J. F. Chemical composition and modulation of bacterial drug resistance of the essential oil from leaves of Croton grewioides. Microb. Pathog, v. 111, p. 468-471, 2017. [CrossRef]
- Coutinho, H. D. M. , Matias, E. F. F., Santos, K. K. A., Tintino, S. R., Souza, C. E. S., Guedes, G. M. M., Santos F.A.D, Costa J.G.M., Falcão-Silva V.S. & Siqueira-Júnior, J. P. Enhancement of the norfloxacin antibiotic activity by gaseous contact with the essential oil of Croton zehntneri. JYP, v. 2, n. 4, p. 362-364, 2010. [CrossRef]
- Nicolson, K. , Evans, G., & O'Toole, P. W. Potentiation of methicillin activity against methicillin-resistant Staphylococcus aureus by diterpenes. FEMS Microbiol. Lett. v. 179, n. 2, p. 233-239, 1999. [CrossRef]
Figure 1.
Chromatogram GC/MS.
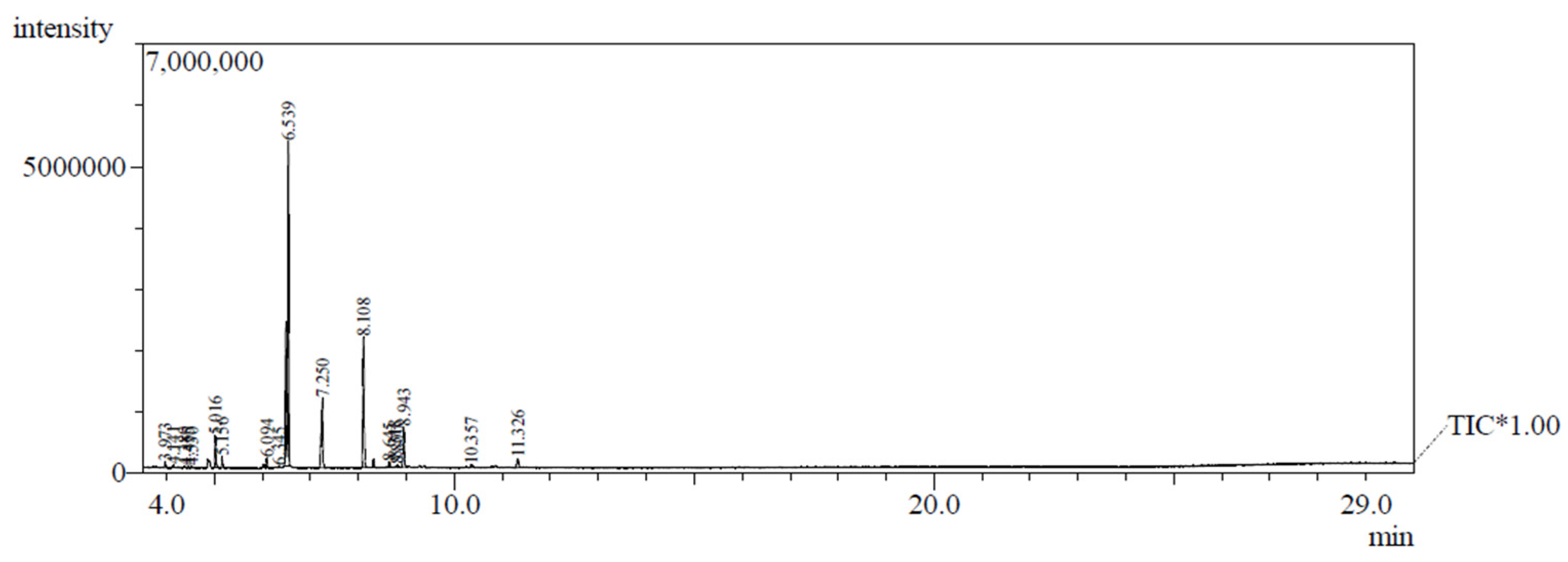
Figure 2.
Chromatogram GC/MS.
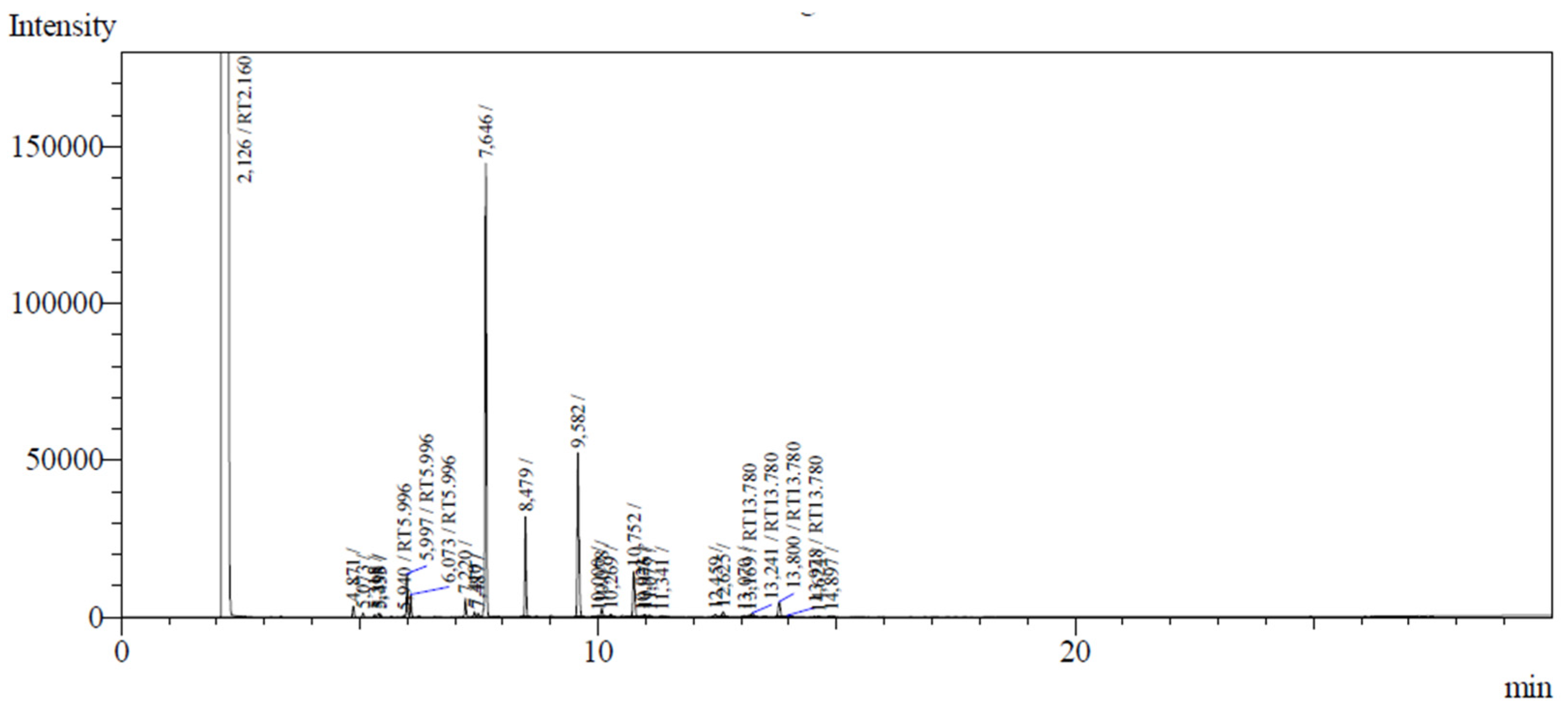
Figure 3.
Modulating potential of OECG on the antibiotic activity of amoxicillin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
Figure 3.
Modulating potential of OECG on the antibiotic activity of amoxicillin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
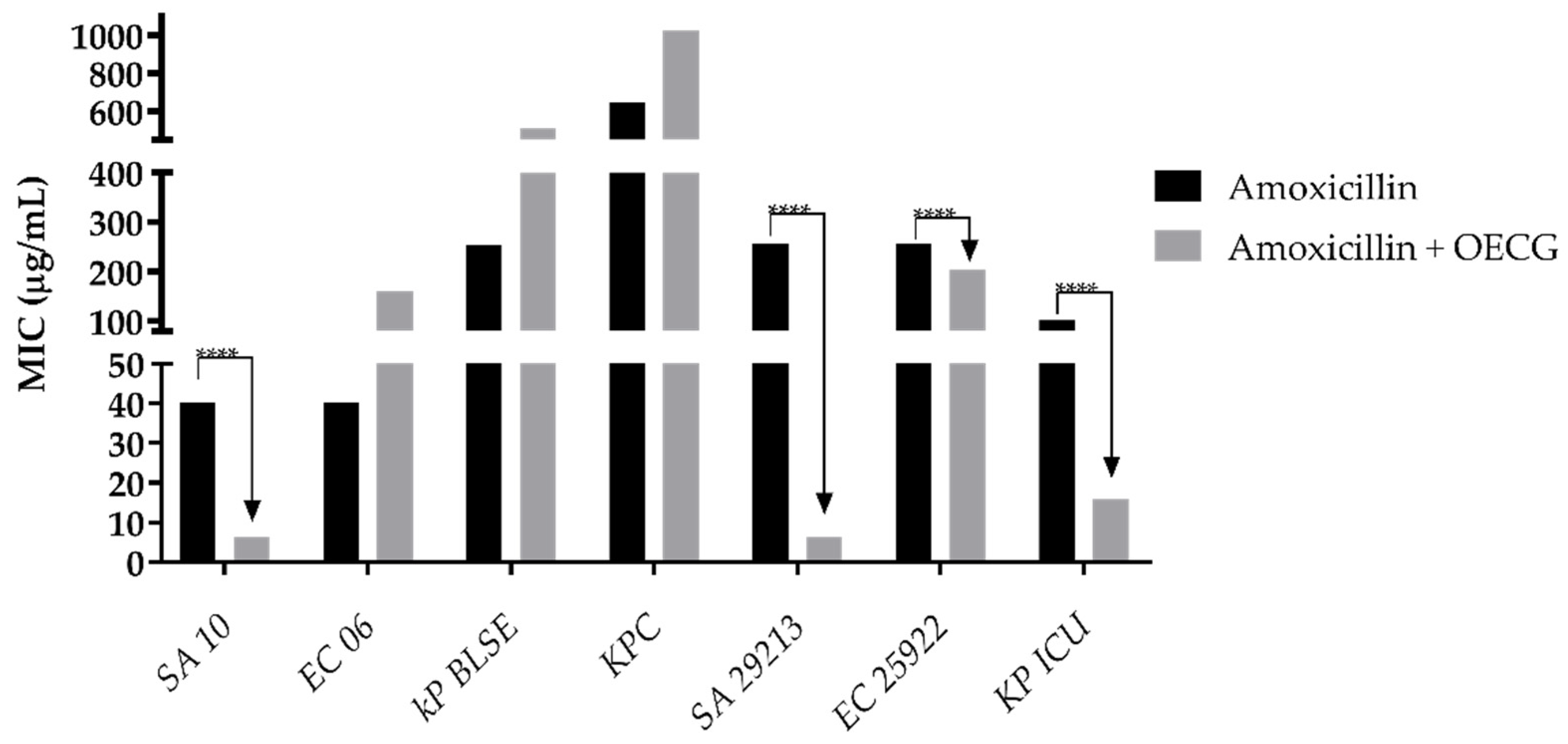
Figure 4.
Modulating potential of OECG on the antibiotic activity of penicillin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
Figure 4.
Modulating potential of OECG on the antibiotic activity of penicillin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
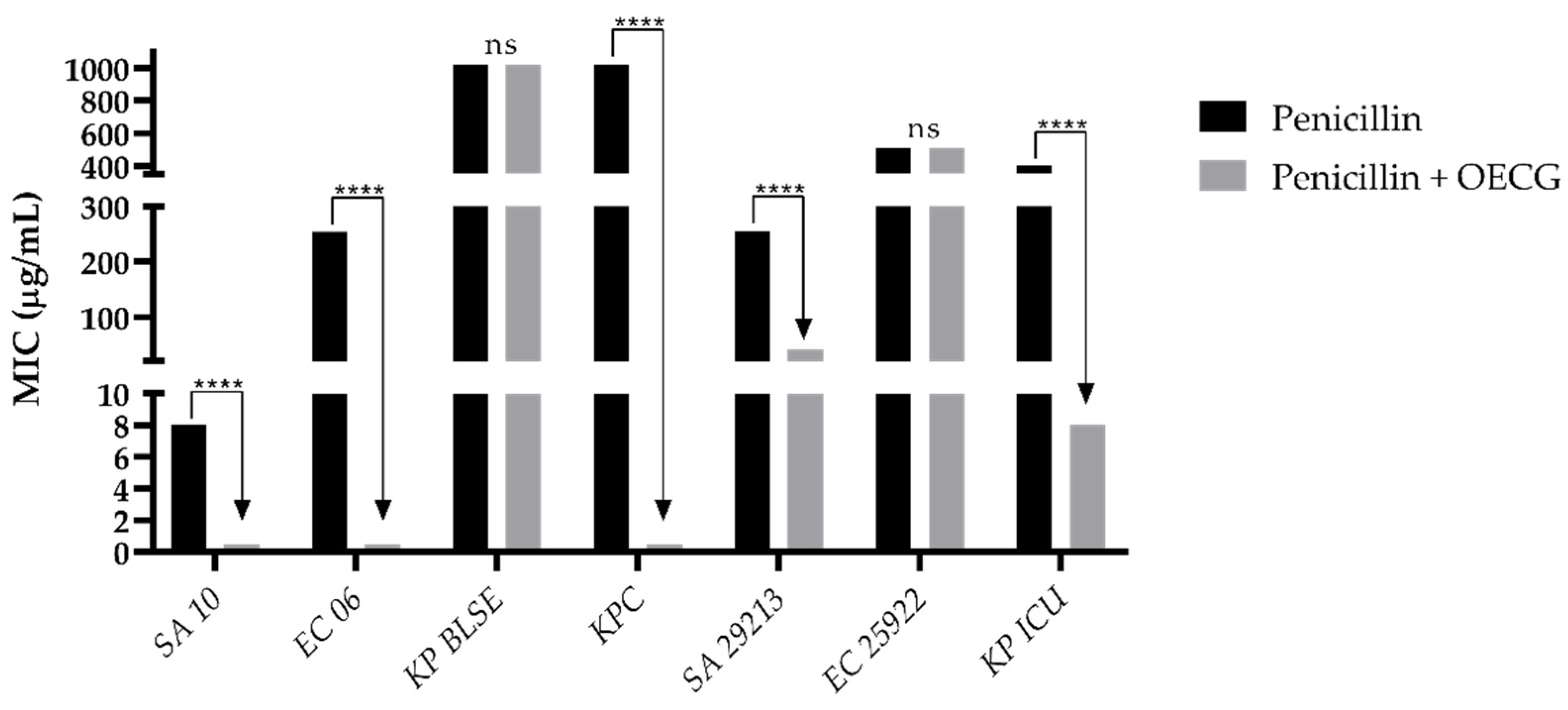
Figure 5.
Modulating potential of OECG on the antibiotic activity of amikacin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
Figure 5.
Modulating potential of OECG on the antibiotic activity of amikacin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
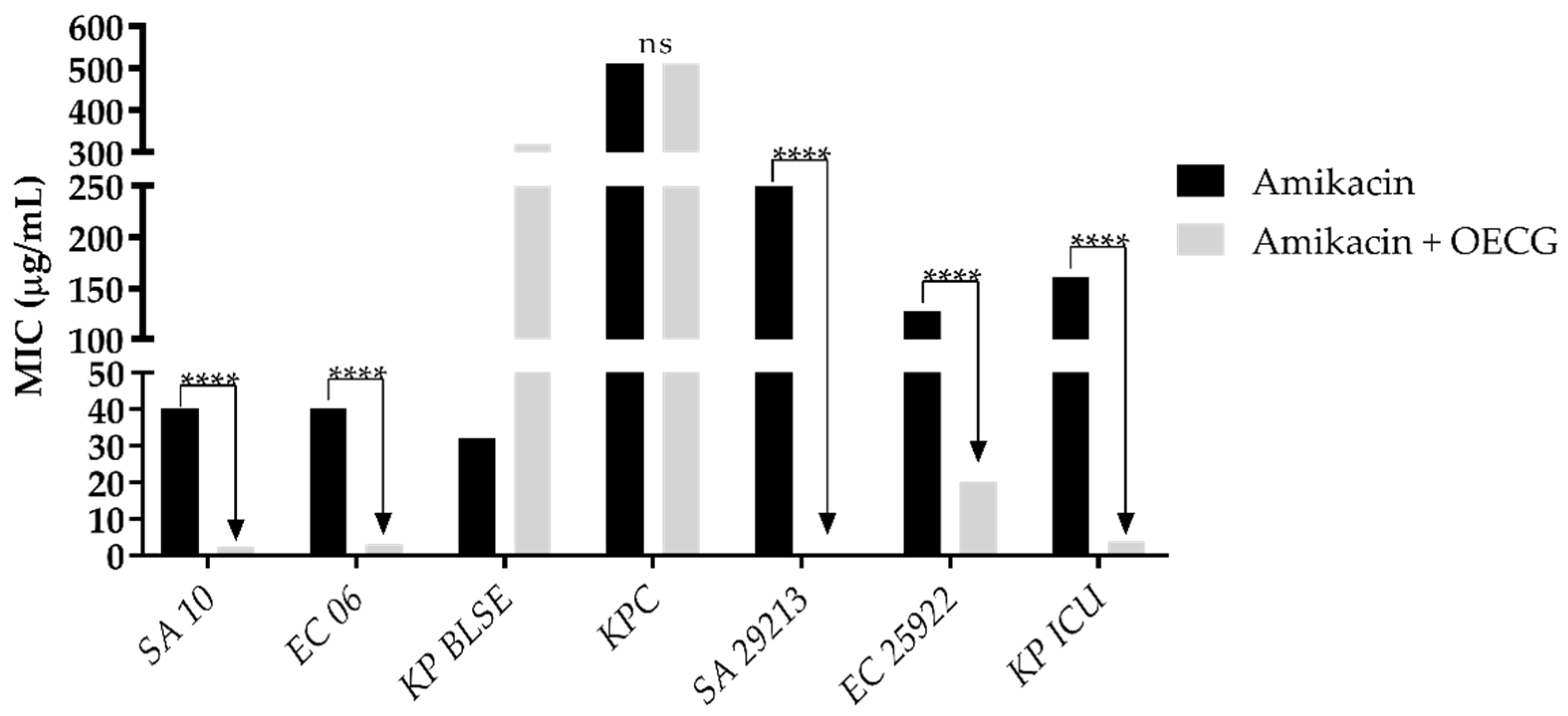
Figure 6.
Modulating potential of OECG on the antibiotic activity of gentamicin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
Figure 6.
Modulating potential of OECG on the antibiotic activity of gentamicin against strains SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922. Source: Author. Two-way ANOVA followed by Bonferroni post-test, using GraphPad Prism 10.0 software. * p <0.0001; **p <0.0005.
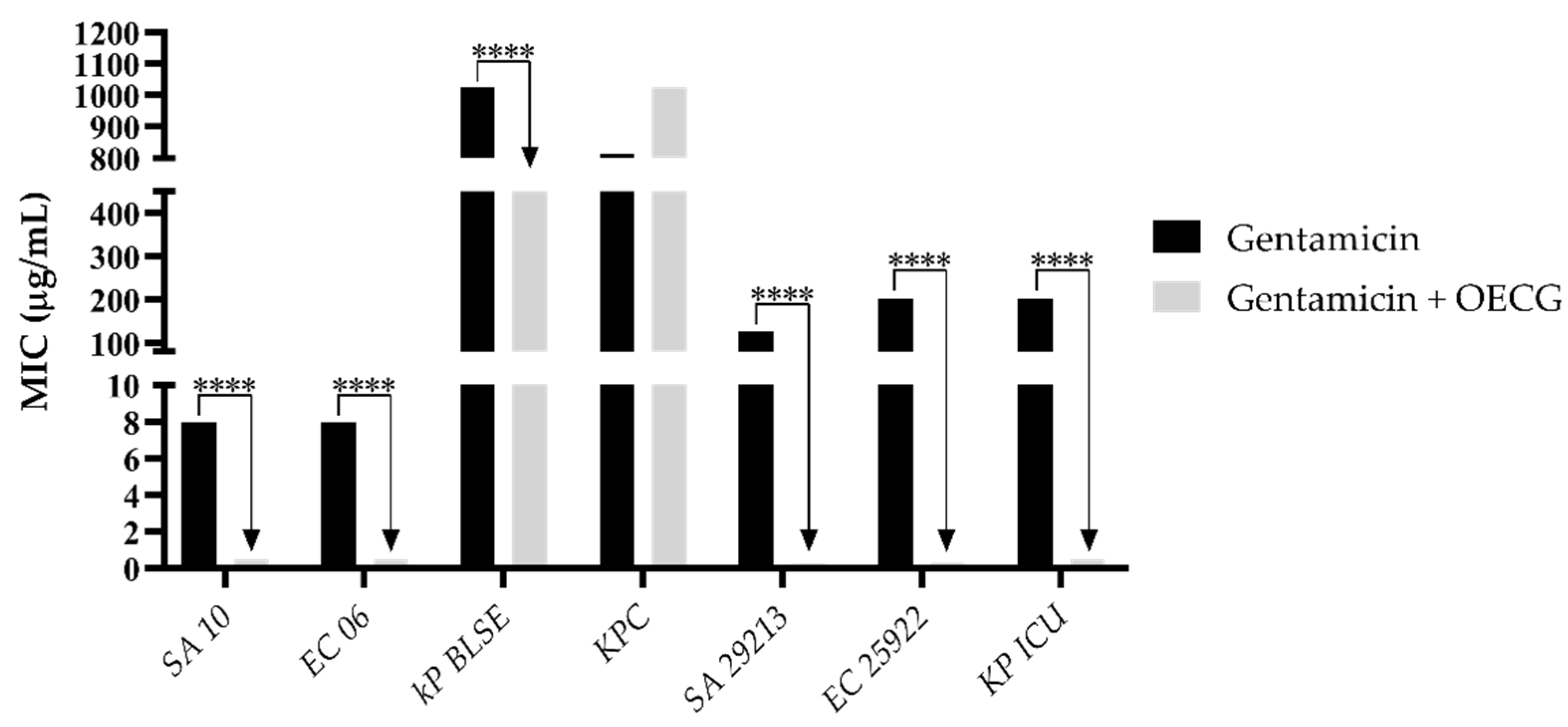

Table 1.
Compounds identified in the essential oil of the aerial parts of C. grewioides.
| Compounds | (%) | RT (min) | IR1 | IR2 |
| estragole | 50.34 | 6.53 | 1.246 | 1.226 |
| methyl eugenol | 19.39 | 8.10 | 1.339 | 1.359 |
| anisole | 10.99 | 7.25 | 1.283 | 1.264 |
| methyl-trans-isoeugenol | 6.09 | 8.94 | 1.403 | 1.422 |
| 1,8-cineole | 4.2 | 5.01 | 1.156 | 1.164 |
| α-selinene | 1.97 | 11.32 | 1.628 | 1.648 |
| β-elemene | 1.37 | 8.31 | 1.355 | 1.375 |
| β-ocimene | 1.09 | 5.15 | 1.165 | 1.145 |
| camphor | 1.08 | 6.09 | 1.223 | 1.243 |
| α-pinene | 0.82 | 3.97 | 1.070 | 1.050 |
| trans-caryphyllene | 0.81 | 8.64 | 1.380 | 1.400 |
| caryphyllene oxide | 0.42 | 10.35 | 1.553 | 1.572 |
| camphene | 0.36 | 4.14 | 1.090 | 1.097 |
| β-myrcene | 0.29 | 4.53 | 1.122 | 1.142 |
| 2-β-pinene | 0.25 | 4.45 | 1.116 | 1.119 |
| isoledene | 0.25 | 8.80 | 1.391 | 1.373 |
| isoborneol | 0.17 | 6.34 | 1.236 | 1.255 |
| sabinense | 0.11 | 4.38 | 1.111 | 1.123 |
| Total | 100 % |
RT: Retention time (min); IR1: Linear retention index obtained; IR2: Linear retention index obtained from the literature.
Table 2.
Minimum Inhibitory Concentration (MIC) valuesof OECG against standard and multiresistant bacteria.
Table 2.
Minimum Inhibitory Concentration (MIC) valuesof OECG against standard and multiresistant bacteria.
| Organisms | MIC (µg/mL) | |
| Multidrug-resistant strains | SA 10 | ≥1024 |
| EC 06 | ≥1024 | |
| KPC | 512 | |
| KP BLSE | 256 | |
| Standard strains | EC 25922 | ≥1024 |
| SA 29213 | ≥1024 | |
| KP ICU | 512 |
SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922;.
Table 3.
Minimum inhibitory concentrations (µg/mL) of antibiotics in the absence and presence of C. growioides essential oil and their MIC reduction factors (FRC).
Table 3.
Minimum inhibitory concentrations (µg/mL) of antibiotics in the absence and presence of C. growioides essential oil and their MIC reduction factors (FRC).
| Substance | Organisms | MIC of antibiotics (µg/mL) | |||
| Amoxicillin | Penicillin | Gentamicin | Amikacin | ||
| Control | SA 10 | 40 | 8 | 8 | 40 |
| EC 06 | 40 | 256 | 8 | 40 | |
| KP BLSE | 256 | 1024 | 1024 | 32 | |
| KPC | 645 | 1024 | 812 | 512 | |
| KP ICU | 101 | 406 | 203 | 161 | |
| SA 29213 | 256 | 256 | 128 | 256 | |
| EC 25922 | 256 | 512 | 203 | 128 | |
| OECG test | SA 10 | 6 | 0.5 | 0.5 | 2 |
| EC 06 | 161 | 0.5 | 0.5 | 3 | |
| KP BLSE | 256 | 1024 | 256 | 161 | |
| KPC | 1024 | 0.5 | 1024 | 512 | |
| KP ICU | 16 | 8 | 0.5 | 4 | |
| SA 29213 | 6 | 40 | 0.25 | 0.30 | |
| EC 25922 | 203 | 512 | 0.30 | 20 | |
| FRC | SA 10 | 85 % | 93.75 % | 93.75 % | 95 % |
| EC 06 | - 302.5 % | 99.80 % | 93.75 % | 92.5 % | |
| KP BLSE | 0 % | 0 % | 75 % | - 403.12 % | |
| KPC | - 58.76 % | 99.95 % | - 26.10 % | 0 % | |
| KP ICU | 84.16 % | 98.02 % | 99.75 % | 97.51 % | |
| SA 29213 | 97.65 % | 84.37 % | 99.80 % | 99.88 % | |
| EC 25922 | 20.7 % | 0 % | 99.85 % | 84.37 % | |
SA 10 - S. aureus 10; EC 06 - E. coli 06; KP BLSE - K. pneumoniae BLSE; KPC - K. pneumoniae Carbapenemáse; KP ICU – K. pneumoniae Clinical Urine Isolate; SA 29213 - S. aureus ATCC 29213; EC 25922 - E. coli ATCC 25922; OECG - C. grewioides essential oil; FRC - MIC reduction factor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



