Submitted:
20 August 2024
Posted:
22 August 2024
You are already at the latest version
Abstract
Manure-compost-soil is the main pathway for the use of agricultural livestock waste. Biochar, regarded as the conditioner, was beneficial to nitrogen fixation during aerobic composting, and was to the enhancement of soil organic matter (OM) and nitrogen content, when applied with compost to the soil. But, the manure-compost-soil process was also related to microbial succession, whether biochar could affect this succession was not studied. In this paper, we investigated the influence of biochar on the manure-compost-soil process and its microbial community succession. Compared with the control group, biochar increased the OM and total nitrogen content of the compost. Biochar, compost and biochar-based compost improved the total carbon (TC) and OM of the soil. The genera of the initial composting sample were Saccharomonospora, Atopostipes and Lactobacillus, and were eliminated after high-temperature fermentation. Lysobacter, Glutamicibacter and Streptomyces were the dominant genera in the soil samples, promoting plant growth. Nocardiopsis, Saccharomonospora, Bacillus and Oceanobacillus dominated the genera in the whole manure-compost-soil process. Thus, aerobic composting could eliminate the toxic or negative bacteria that were directly input into the soil by manure. Those genera from compost or biochar-based compost flux into the soil could make an important contribution to OM’s cycle.
Keywords:
biochar
; pig manure
; aerobic composting
; black soil
; microbial migration
; 16S rRNA
1. Introduction
In recent years, livestock and poultry manure production in China has surged to approximately 4.2 billion tons annually [1]. If not handled properly, manure can cause environmental pollution [2]. Aerobic composting offers a green technology for treating agricultural waste [3]. With microbial activity and ample oxygen, aerobic composting produces high-quality compost suitable for soil enhancement [4]. These composts, regarded as organic fertilizers, can enrich soil fertility, enhancing agronomy and crop productivity [5,6]. Compost application boosts soil carbon and nitrogen content, stabilizes soil organic matter, improves soil structure for root growth, and introduces microorganisms into the soil [7]. In summary, the manure–compost–soil pathway is crucial for managing agricultural livestock and poultry waste.
The manure–compost–soil process plays a pivotal role in microbial transfer and succession. Microbial diversity undergoes significant changes during aerobic composting, and the composition of different composts varies due to factors like raw materials [8,9,10]. The soil microenvironment was rich in microbiological composition, making it a key focused in soil ecosystem management [11]. Fierer et al. [12] analyzed data from 198 publications and found that soil carbon and nitrogen utilization were critical for plant growth, and microorganisms significantly influence soil nitrogen mineralization. Soil health is also related to the bacterial structure within the soil. Fierer et al. [12] also highlighted the ability to infer specific soil properties by quantifying the relative abundance of microbial taxa or genes. Therefore, investigating microbial diversity and structure in the manure–compost–soil process is crucial for understanding the effects of manure utilization.
Material transformation occurs throughout the manure–compost–soil process. Applying compost and manure to soil can improve fertility, but organic matter rapidly mineralizes, with only a small fraction remaining stable in the soil long-term [13]. Recent studies have shown that biochar and compost application can effectively increase organic matter retention, enhancing soil quality and farm productivity [14,15,16]. Biochar is a highly aromatic, insoluble solid produced by biomass thermal cracking and carbonization anoxia conditions. It has physicochemical properties such as high stability and developed pore structure, promoting composted organic matter degradation and enhancing microbial richness [17,18]. Applying biochar to soil can improve physical structure aspects, such as the pore structure and soil water holding capacity, facilitating the conversion of organic nutrients [16]. Agegnehu et al. [19] also noted that biochar and compost can improve soil quality and crop yields. We hypothesized that biochar-based compost made by aerobic composting combines the functions of compost and biochar, promoting the formation of carboxyl and phenolic groups, and improving reactivity. Mawof et al. [15] found that biochar, compost, and biochar–compost mixtures significantly improved soil’s physicochemical properties and crop yield under wastewater irrigation (p < 0.05). Appling biochar and biochar-based compost to soil can improve soil fertility, water-holding capacity, crop yield, and carbon sequestration, which is vital agriculture’s green and sustainable development [20,21,22,23,24]. Recent studies have focused only on the effect of compost or biochar on the structure of soil microbes. For instance, Yin et al. [25] found that biochar accelerated the humification composting process, improving compost quality. Xu et al. [26] explored the effects of corn stover biochar in soil on nitrogen leaching and bacterial community structure, and found that biochar addition helped reduce soil nitrogen leaching and increased bacterial diversity in the soil. However, the mechanism of microbial transfer or succession during the production and application of biochar-based compost remains unclear. We assume that manure carries some nutrients and microbes. When manure becomes fertilizer or compost via aerobic composting through high-temperature fermentation and microbial activity, the microbial structure changes. When compost is applied, compost microbes flow into the soil, which also has its own microbial structure. Thus, a key question is whether compost and biochar application can alter microbial succession.
This paper innovatively delves into the multifaceted effects of incorporating biochar as an additive in the manure–compost–soil process, with a particular emphasis on its unprecedented impact on bacterial succession. By examining the complex interactions between biochar, compost, and soil microbial communities, we conducted an aerobic composting experiment using pig manure with biochar as an additive. Subsequently, we utilized the compost products in a potting experiment. Throughout this process, we collected representative samples from the raw composting materials, final compost, and soil samples near the cucumber root system in the experimental group. We measured and analyzed the basic physicochemical properties of the soil samples and utilized 16S rRNA high-throughput data. Our research pioneers a deeper comprehension of how biochar not only modifies the physical and chemical properties of compost and soil but also fundamentally transforms the microbial landscape, fostering a more diverse and beneficial bacterial succession.
2. Materials and Methods
2.1. Aerobic Composting and cucumber Potting Experiment
This study encompassed two primary sets of experiments: aerobic composting and cucumber potting. The composting experiment was divided into control (A and C) and experimental (B and D) groups, while the potting experiment evaluated different combinations of compost and biochar.
The primary raw materials for composting were fresh pig manure and wheat straw cut to 1~3 cm. These were sourced from the pig farm of the Chinese Academy of Agricultural Sciences (Beijing, China) and the Shangzhuang experimental base of China Agricultural University in Beijing, China. The control groups (A and C) consisted of a mixture of swine manure and wheat straw at an initial moisture content of 60% and a C/N ratio of 15, with a weight ratio of 5:1 (Table 1) [4]. The experimental groups (B and D) incorporated wheat straw biochar, prepared via high-temperature pyrolysis at 500 °C, into the mix at a ratio of 10:2:1 swine manure: wheat straw: biochar according to Liu et al. [27]. The mixtures were placed into four 90 L aerobic composting reactors and composted for 28 days under intermittent oxygen supply (30 min on/30 min off) with a ventilation rate of 0.2 L·min⁻¹·VS⁻¹·kg⁻¹ [28]. Samples were collected from the top, middle, and bottom of the reactors on days 0, 3, 7, 14, 21, and 28 for physicochemical and microbiological analysis.
The cultivation substrate for the potting trials consisted of a 5:1 mix of black soil and vermiculite. Six groups were set up, including a blank control (no additives), a biochar-only treatment, and four treatments with composts from the composting experiment (Table 1). The additive ratio of compost and biochar was based on a fertilizer application rate of 250 kg N/hm2 [26]. The details of group name and ingredients are shown in Table 1. Figure S1 shows the cucumber potting experiment set up with six groups (one group for each six parallel pots). Each pot contained ~0.25 kg of substrate and was seeded with five cucumber seeds (Zhongnong 6 variety). The plants were grown in an artificial climate chamber (MGC-350HP-2, China) at 25-30 °C with a 16:8 h light-dark cycle for two months. The moisture content of the material in the pots was adjusted to 60-70%. Soil samples around the root were collected on days 0, 7, 21, 42, and 64 for physicochemical and microbiological analysis. The basic physicochemical properties of the raw materials for aerobic composting and pot experiments are shown in Table 2.
2.2. Basic physical and chemical indicators.
The samples were dried at 105 °C for 24 h until a constant weight to measure the moisture content (MC). Total carbon (TC) and total nitrogen (TN) content were determined using an elemental analyzer (Vario EL CHNOS; Elemental Analysensystems GmbH, Langenselbold, Germany). These values were used to calculate the C/N ratio [29]. Organic matter (OM) and germination index (GI) were determined and calculated according to the methods provided by He et al. [28]. NH4+-N were extracted from the compost samples by leaching with 2 mol/L KCL and the content was determined by a continuous flow analyzer (SEAL Analytical AutoAnalyzer 3, Germany) [30]. The supernatant was extracted and filtered at a solid–liquid ratio of 1:10 (w/v), and the pH values were determined using a pH meter (SC8231, Japan) [29]. The chemical functional groups of the samples were determined using a Fourier transform infrared spectrometer (FT-IR, PerkinElmer Spotlight 400, Waltham, MA, USA), according to Fang et al. [31].
2.3. Microbial Diversity Determination and Analysis
2.3.1. DNA Extraction and Purification
Total DNA from compost samples was extracted using a soil genomic DNA extraction kit (DP336, Tianhe Biotech (Beijing, China). Extracted DNA was purified using a Puc-T TA cloning kit (CWBIO, Beijing, China), and then purified DNA was analyzed by 1% (w/v) agarose gel electrophoresis.
2.3.2. High-Throughput Sequencing
A fluorescence spectrophotometer (NANODROP 2000, Thermo Scientific, USA) was used to detect the concentration of DNA, and the quality of DNA was checked via 1% agarose gel electrophoresis. The concentration of the DNA solution was adjusted, and the DNA working solution was stored at -4 °C. The V3–V4 region of the 16S rRNA gene was selected for bacterial-specific fragmentation with primers 338F (5‘-ACTCCTACGGGGAGGCAGCAG-3’) and 806R (3‘-GGACTACHVGGGTWTCTAAT-5’) [32]. The PCR amplification program consisted of pre-denaturation at 95 °C for 5 min, denaturation at 95 °C for 45 s, annealing at 55 °C for 50 s, and extension at 72 °C for 45 s for a total of 28 cycles; and a final extension at 72 °C for 10 min. The PCR amplification products were sequenced using the Illumina MiSeq PE300 sequencing platform.
Raw data generated from high-throughput sequencing runs were processed and analyzed following the flow of the QIIME platform (Quantitative insights into microbial ecology; v1.2.1). Sequence reads were trimmed so that the average quality of each read was higher than 20 and then assembled using Flash software. Chimeric sequences were identified and removed using UCHIME. Taxonomic analysis of OTU representative sequences with a 97% similarity level was performed using uclust classification or a RDP classifier Bayesian algorithm. Sequence clustering was performed by uclust (QIIME) with a similarity cut-off value of 97%, after which the samples were clustered into operational taxonomic units. QIIME software was used to calculate the Chao1 index and Shannon index of the samples, and the R language toolkit plotted the remaining graphs. A typical correlation analysis (CCA) was performed using Canoco for Windows (v.5.0) to determine the correlation between biochar and microorganisms in compost and microorganisms in soil. Firstly, DCA analysis was performed on the sample data, and then the maximum Pearson correlation coefficient of the environmental factors with the differences in the distribution of the sample communities and the subset of the environmental factors were determined by the bioenv function. The distribution tables of the sample species with respect to the environmental factors or subsets of environmental factors were then analyzed separately, and finally their significance was obtained by using ANOVA software.
2.4. Statistics and Data Analysis
Statistical analyses of the basic physical and chemical indicators were performed using Excel 2019 (Microsoft Corporation, USA). The data were expressed as the mean ± standard deviation values of repeated measurements. Dynamic changes in the physicochemical properties were plotted using OriginPro 2022b software (OriginLab, USA). Significant difference analysis was calculated by ANOVA analysis using SPSS statistics v19.0 (IBM, USA) (p < 0.05).
3. Results and Discussion
3.1. Physicochemical Changes in the Manure–Compost–Soil Process
3.1.1. Manure–Compost during Aerobic Composting
The key findings from Figure 1 highlight the beneficial effects of incorporating biochar into composting. Notably, while there were no significant differences in temperature or oxygen concentration among the four groups (control A/C and experimental B/D) (p > 0.05). Figure 1(a) shows the four groups treated at temperatures >50°C for more than three days; the compost met the non-hazardous criteria [4]. Besides, throughout the composting process, the experimental groups (B and D) exhibited a longer duration of high temperatures (>50°C) and higher oxygen concentrations during the initial days when temperatures exceeded 50°C. This statistically significant improvement (p < 0.05) in oxygen penetration indicates that biochar enhances the internal aeration of the compost heap [33,34].
Regarding the physicochemical properties, although pH, germination index (GI), organic matter (OM), ammonia (NH4+), total nitrogen (TN), and total carbon (TC) did not differ significantly between the control and experimental groups within their respective categories (A vs. C and B vs. D) (p>0.05), some interesting trends emerged. As shown in Figure 1(b), the pH initially rose and then declined in all groups, likely due to the production and subsequent dissipation of ammonium ions [33]. Notably, the experimental groups (B and D) maintained lower pH values, suggesting ongoing microbial activity and organic acid production [35]. In Figure 1(b), the germination index (GI) values of the four groups increased gradually due to the degradation of toxic substances such as low-molecular-weight fatty acids, ammonia, and toxic nitrogen compounds [36]. The GI of the compost products of all the four groups was greater than 80%, which indicated that the final compost maturity was up to the required level [37,38]. The GI of group D was significantly higher than that of the other three groups (p < 0.05), indicating that the addition of biochar may reduce harmful substances in compost to an extent. In Figure 1(c), organic matter was continuously consumed during the composting process and the degradation rate of the experimental groups (B and D) was higher than that of the control groups (A and C). The organic matter’s degradation rates in the four groups were 3.92%, 10.45%, 4.73%, and 7.41%, respectively. This may be because biochar promoted organic matter degradation and water evaporation during aerobic composting [39]. The NH4+ content of the four groups showed a decreasing trend (Figure 1(c)). Eventually, the NH4+ content of the control groups (A and C) was slightly higher than that of the experimental groups (B and D), which was due to the retention of NH4+ by the biochar slowing down the activity of denitrifying bacteria [40]. In Figure 1(d), the total nitrogen content in the experimental groups (B and D) was slightly higher than that in the blank groups (A and C). This is due to the ability of biochar to absorb ammonia and other nitrogenous substances, which explains the higher total nitrogen levels in treatments using biochar as an additive [41]. The total carbon content of the experimental groups (B and D) with biochar addition was significantly higher than in the blank groups (A and C) (p < 0.05). Biochar had a better absorption ability to conserve carbon [41]. Therefore, the addition of biochar increased the carbon/nitrogen ratio in the composting experiment.
3.1.2. Compost–Soil in the Potting Experiment
The cucumbers in the pot trial germinated and grew well, sprouting around day 7 (Figure S2). Based on the statistical analysis, there was no significant difference in pH, OM, TC and TN contents between the AA and CC groups (P>0.05), as well as between the BB and DD groups. As shown in Figure 2(a), the pH changes of soil samples in the BLANK group fluctuated less. The pH of soil samples in other groups showed a tendency of decreasing and then increasing, with the range mostly between 6.7 and 7.8. On days 0, 10, and 60, the pH of the BIOCHAR group was higher than that of the BLANK group. The pH of the AA, BB, CC, and DD groups was significantly higher than that of the BLANK group (p < 0.05). Adding biochar and compost increased the soil pH, as also noted by Naeem et al. [42]. This is mainly because biochar and compost are mostly weakly alkaline [43]. As shown in Figure 2(b), the organic matter and total carbon contents of BIOCHAR group were significantly higher than in the control group (BLANK) (p < 0.05). This indicated that compost can be used as a source of organic matter, providing much-needed nutrients to the soil [44]. As shown in Figure 2(c), (d), the total nitrogen contents of the AA and CC groups were significantly higher than that of the control group (BLANK) (p < 0.05). This is because biochar can directly improve the nitrogen cycle in the soil by dissolving organic nitrogen and fixing nitrogen [44]. Conclusively, the application of compost and biochar increased the organic matter, total carbon, and total nitrogen contents in the soil samples.
In Figure S3, the wave peaks at 1077 cm-1 for different substances were Si-O-Si vibrational peaks [45]. Vermiculite and black soil had higher absorption values compared to the other substances, probably due to the greater abundance of elemental Si. The peak distributions of the added biochar (BIOCHAR) were similar to those of the control (BLANK) groups. The compost samples from A, B, C, and D showed C-H vibrational peaks at 2928 cm-1 and N-H vibrational peaks at 3313 cm-1, which represent the alkyl group and amide, respectively. This indicated a higher content of carbon-based material and protein content flow into soil samples of the AA, BB, CC, and DD groups. This is because compost is rich in nitrogen and carbon-containing nutrients. Agegnehu et al. [19] proposed that the application of biochar to acidic soils not only improves the pH of the soil, but also provides the soil with influential nitrogen. Compost is also rich in NH4+, which can be utilized to improve the physicochemical properties of soil and thus provide nutrients for cucumber seedlings.
3.2. Analysis of the Bacterial Diversity in the Manure–Compost–Soil Process
In Figure 3(a), the microbial composition of the initial compost sample was significantly different (p < 0.05) from that of the final compost sample and soil sample. However, there was a high similarity in microbial composition between the final compost samples (A28, B28, C28, and D28) and the potting soil samples (AA0, BB0, CC0, and DD0) to which the compost was applied. This indicates that as the compost was applied to the soil, the microorganisms that survived in the compost samples flowed into the soil. As shown in Table 3, the microbial abundance of compost decreased significantly during the heating period of the aerobic composting process because bacterial activity was inhibited by the high temperature [9,46,47]. The microbial diversity index of the potted final soil samples (AA60, BB60, CC60, DD60, and BLANK60) increased compared to their respective initial soil samples. The microbial diversity index was lower in soil samples with added compost and biochar-based compost (AA0, BB0, CC0, and DD0) than in the control soil group (BLANK0), because of more bacteria from compost (Figure 3(b)). The microbial diversity index was higher in the soil sample with biochar application (BIOCAHR0) compared to the control (BLANK0) due to the increased nutrient retention capacity of the soil [48]. Xu et al. [26] found that the microbial diversity in the soil increased as the amount of biochar applied increased. Overall, the microbial composition of the soil was richer than that of the raw compost material (agricultural solid waste), making the study of microbial function in the soil microstructure complex.
Notes: Initial aerobic composting samples on Day 0 of A, B, C, and D were named A0, B0, C0, and D0. Final aerobic composting samples on Day 28 of A, B, C, and D were named A28, B28, C28, and D28. Initial soil samples of potting experiment on Day 0 of BlANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_0, BIOCHAR_0, AA_0, BB_0, CC_0, and DD_0. Final soil samples of potting experiment on Day 60 of BALNK, BIOCHAR, AA, BB, CC, and DD were named BLANK_60, BIOCHAR_60, AA_60, BB_60, CC_60, and DD_60.
3.3. Bacterial Community Composition in the Manure–Compost–Soil Process
As seen in Figure 4(a), the dominant phyla in the composting stage of the composting samples were Firmicutes, Actinobacteriota, and Proteobacteria, whose relative abundance (RA) accounted for more than 90% of the RA of the total bacterial flora. Mao et al. [47] found that the dominant bacterial phylum in the high-temperature period samples of pig manure compost with added bamboo biochar and bacterial agents were Firmicutes and Proteobacteria, and that the activity of the bacteria in the high-temperature period was affected by the soluble organic carbon content and temperature. The RA of Firmicutes exceeded 90% in the initial compost samples. Actinobacteriota gradually developed into the dominant bacterial phylum during the composting reaction, and biochar increased its RA in the compost samples. The dominant phyla that together accounted for more than 90% of RA in the soil samples were Chloroflexi, Bacterioidota, and Acidobacteriota. Previous studies found that Proteobacteria, Acidobacteria, Chloroflexi, and Bacteroidetes were also more abundant in the nitrogen-laced soil samples [26,47]. Biochar-containing soil samples (BIOCHAR group) had lower RAs of Acidobacteria, Chloroflexi, and Gemmatimonadetes and higher RAs of Proteobacteria, Bacteroidetes, and Actinobacteria. Thus, Firmicutes was the most abundant bacterial phylum in the compost samples. Among the soil samples, the largest number of bacterial species belonged to the Actinobacteriota. The composition of the bacterial phylum did not change much from the compost samples to the soil samples, but there were some differences in the subordinate bacterial species.
Notes: Initial aerobic composting samples on Day 0 of A, B, C, and D were named A0, B0, C0, and D0. Final aerobic composting samples on Day 28 of A, B, C, and D were named A28, B28, C28, and D28. Initial soil samples of potting experiment on Day 0 of BLANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_0, BIOCHAR_0, AA_0, BB_0, CC_0, and DD_0. Final soil samples of potting experiment on Day 60 of BLANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_60, BIOCHAR_60, AA_60, BB_60, CC_60, and DD_60.
As known in Figure 4(b), Nocardiopsis was more abundant in soil samples of compost (AA, CC) or biochar-based compost (BB, DD) application groups than of control (BLANK) and biochar additive (BIOCHAR) groups. Nocardiopsis belongs to the Actinobacteriota and has been detected in compost and soil, producing a number of antimicrobial compounds, including thiopeptides [49,50]. It contributes to the recycling of organic compounds. Streptococcus was mostly present in initial samples of compost, originating from manure, and was almost absent after aerobic high-temperature fermentation. It serves as an indicator of harmful microorganisms for the environmentally sound treatment of agricultural wastes [51]. Atopostipes and Lactobacillus were only present in the initial samples of the compost and had low tolerance to the high-temperature fermentation of the compost [52]. Clostridium_sensu_stricto_1 and Terrisporobacter were present in the later stages of composting and belong to the Firmicutes, the main genera carrying resistance genes [53]. Lysimachia belong to the genus Ganoderma in the family Xanthomonadaceae and was a member of an ecologically important microbial community associated with soils and plants. It has been shown to induce systemic resistance in certain plant species, thereby protecting plants from pathogen infection [54,55]. Glutamicibacter is derived from soil samples that show a variety of potential plant-growth-promoting properties and can tolerate high NaCl concentrations and a wide pH range [56]. Streptomyces is derived from soil samples and produces volatile organic compounds that have the potential to inhibit soil diseases [57]. Thus, aerobic composting can eliminate toxic or negative bacteria that are directly deposited in the soil by manure. Compost or biochar-based compost applied to soil as a fertilizer can introduce some genera into the soil, such as Nocardiopsis and Clostridium_sensu_stricto_1, which have a positive influence on the microbial diversity of the soil.
3.4. Difference Analysis of Bacteria from Pig Manure–Compost to Compost–Soil
Figure 5 presents a detailed analysis of bacterial diversity at the genus level in both compost and soil samples. While certain genera like Nocardiopsis, Saccharomonospora, Bacillus, and Oceanobacillus exhibited differences in abundance between compost and soil, these differences were not statistically significant. However, at a 95% confidence interval, notable variations emerged for other genera, including Clostridium_sensu_stricto_1, Streptococcus, Terrisporobacter, Lysobacter, Atopostipes, Lactobacillus, Glutamicibacter, Romboutsia, and Streptomyces. Notably, Lysobacter, Glutamicibacter, and Streptomyces were considerably more abundant in soil samples compared to compost samples, highlighting their dominance in the soil environment.
Intriguingly, the manure initially contained potentially contaminating and toxic genera such as Saccharomonospora (from 2.68% to 0.80%), Atopostipes (from 5.71% to 0.13%), and Lactobacillus (from 5.27% to 0.04%), which were effectively eliminated through the high-temperature aerobic composting process. This finding underscores the sanitizing effect of composting on manure-derived microbial contaminants.
In contrast, soil samples from the potting experiment revealed Lysobacter, Glutamicibacter, and Streptomyces as the dominant genera, contributing positively to the microbial diversity of the soil. Furthermore, the application of compost or biochar-amended compost as fertilizer introduced additional beneficial genera like Nocardiopsis and Clostridium_sensu_stricto_1 into the soil, further enriching its microbial diversity. These results demonstrate the potential of biochar-based compost to promote a healthy and diverse microbial ecosystem in agricultural soils.
Notes: Compost containing composting samples: A0, B0, C0, D0, A28, B28, C28, D28; Soil containing soil samples: AA0, BB0, CC0, DD0, AA60, BB60, CC60, DD60. Initial aerobic composting samples on Day 0 of A, B, C, and D were named A0, B0, C0, and D0. Final aerobic composting samples on Day 28 of A, B, C, and D were named A28, B28, C28, and D28. Initial soil samples of potting experiment on Day 0 of AA, BB, CC, and DD were named AA_0, BB_0, CC_0, and DD_0. Final soil samples of potting experiment on Day 60 of AA, BB, CC, and DD were named AA_60, BB_60, CC_60, and DD_60.
3.5. Correlation Analysis of Bacteria and Physicochemical Properties in the Manure-Compost-Soil Process
In Figure 6(a), the structure of bacterial communities in the four soil experimental groups (AA, BB, CC, and DD) underwent a notable transformation over the 60-day period. Initially dispersed across different regions, the groups converged into a single region by the end of the experiment, indicating that both compost and biochar-amended compost exert comparable influences on the bacterial genetics in soil. In contrast, during the composting phase, the four compost groups remained clustered together, whereas the biochar-supplemented group in the soil experiment exhibited a distinct spatial pattern. This suggests that while biochar did not directly alter the genetic makeup of bacteria during composting, it began to exert its effects once introduced into the soil system. Regarding the nutrient dynamics, the synchronous variations in total carbon and nitrogen concentrations reflect their mutual dependence on organic matter degradation [33].
In Figure 6b, notable associations were observed between specific genera and physicochemical properties. For instance, Pseudohyphobacterium and Marsupialia displayed strong correlations with pH, while Aerococcus and Facklamia were positively linked to moisture content (MC). Moreover, bacterial genera such as Romboutsia, Turicibacter, and Lactobacillus exhibited positive correlations with organic matter (OM), total carbon (TC), total nitrogen (TN), and NH4+ levels. In contrast, Lysobacter, Streptomyces, and Ornithinimicrobium showed negative correlations with these parameters[57]. Notably, Lactobacillus, known for its strict fermentation and anaerobic nature, was among the genera positively associated with soil fertility indicators.
Notes: MC: moisture content, %; OM: organic matter, %; TC: total carbon, %; TN: total nitrogen, %. Initial aerobic composting samples on Day 0 of A, B, C, and D were named A0, B0, C0, and D0. Final aerobic composting samples on Day 28 of A, B, C, and D were named A28, B28, C28, and D28. Initial soil samples of potting experiment on Day 0 of BLANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_0, BIOCHAR_0, AA_0, BB_0, CC_0, and DD_0. Final soil samples of potting experiment on Day 60 of BLANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_60, BIOCHAR_60, AA_60, BB_60, CC_60, and DD_60.
4. Conclusion
This paper lies in its innovative exploration of the multifaceted effects of incorporating biochar as an additive. Specifically, the paper contributes new insights into how biochar not only modifies the physical and chemical properties of compost and soil but also fundamentally transforms the microbial landscape, fostering a more diverse and beneficial bacterial succession in the manure–compost–soil process.
- Biochar addition during aerobic composting increased the C/N ratio and total carbon content of the compost, as well as improved the germination index.
- When applied to soil, composts containing biochar increased the soil's pH, organic matter, total carbon, and total nitrogen contents compared to composts without biochar.
- Microbial diversity decreased during the high-temperature phase of aerobic composting but increased significantly in the final compost products. Soil samples amended with composts showed higher microbial diversity than unamended soil.
- The microbial communities in the composts and soil samples were distinct, but some bacterial genera (e.g., Nocardiopsis, Clostridium_sensu_stricto_1) persisted and contributed positively to the microbial diversity of the soil.
- Correlation analysis revealed significant relationships between bacterial genera and soil physicochemical properties, providing insights into the microbial mechanisms underlying the observed effects of biochar on soil quality.
In summary, the paper's novelty stems from its comprehensive analysis of the complex interactions between biochar, compost, and soil microbial communities, which led to new insights into how biochar-based compost can enhance soil health and microbial succession.
In a future study, the microbial structure during the manure–compost–soil process could be further analyzed by taking plant samples in conjunction with assessing plant growth. The main soil used in this study was black soil, which had nutrients suitable for plant growth to some extent. We could consider using acidic or saline–alkaline soils to conduct in-depth and systematic research related to improving the physicochemical properties or microbial succession induced by compost, biochar, or biochar-based compost.
Supplementary Materials
Figure S1. Soil raw materials and mixed actual substrate of pot experiments in different groups. Figure S2. Progress in cucumber potting trials. Figure S3. Wave number diagrams for different substances.
Author Contributions
Conceptualization, X.H.(Xiang He) and X.H.(Xueqin He); methodology, X.H.(Xueqin He) and X.H.(Xiang He); software, X.H.(Xiang He) and L.H.(Li He); validation, X.H.(Xiang He), F.J.(Feiyang Jiao) and K.Z.(Kan Zhang).; formal analysis, X.H.(Xiang He) and X.H.(Xueqin He); resources, X.H.(Xueqin He); data curation, X.H.(Xueqin He) and X.H.(Xiang He); writing—original draft preparation, X.H.(Xiang He); writing—review and editing, X.H.(Xiang He) and X.H.(Xueqin He); visualization, X.H.(Xiang He) and L.H.(Li He); supervision, X.H.(Xueqin He); project administration X.H.(Xueqin He), funding acquisition, X.H.(Xueqin He).
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
This work was supported by the National Key Research and Development Program of China (2022YFD200210304).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Awasthi, S.K.; Duan, Y.; Liu, T.; Zhang, Z.; Pandey, A.; Varjani, S.; Awasthi, M.K.; Taherzadeh, M.J. Can biochar regulate the fate of heavy metals (Cu and Zn) resistant bacteria community during the poultry manure composting? J Hazard Mater 2021, 406, 124593. [CrossRef]
- Ma, S.; Sun, X.; Fang, C.; He, X.; Han, L.; Huang, G. Exploring the mechanisms of decreased methane during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane. Waste Manage 2018, 78, 393-400. [CrossRef]
- Ravindran, B.; Nguyen, D.D.; Chaudhary, D.K.; Chang, S.W.; Kim, J.; Lee, S.R.; Shin, J.; Jeon, B.; Chung, S.; Lee, J. Influence of biochar on physico-chemical and microbial community during swine manure composting process. J Environ Manage 2019, 232, 592-599. [CrossRef]
- Haug, R. The practical handbook of compost engineering. CRC Press: 1993; p. [CrossRef]
- Zhou, S.; Chang, T.; Zhang, Y.; Shaghaleh, H.; Zhang, J.; Yang, X.; Qin, H.; Talpur, M.M.A.; Alhaj Hamoud, Y. Organic fertilizer compost alters the microbial composition and network structure in strongly acidic soil. Appl Soil Ecol 2024, 195, 105263. [CrossRef]
- Morra, L.; Bilotto, M.; Baldantoni, D.; Alfani, A.; Baiano, S. A seven-year experiment in a vegetable crops sequence: Effects of replacing mineral fertilizers with Biowaste compost on crop productivity, soil organic carbon and nitrates concentrations. Sci Hortic-Amsterdam 2021, 290, 110534. [CrossRef]
- Scotti, R.; Pane, C.; Spaccini, R.; Palese, A.M.; Piccolo, A.; Celano, G.; Zaccardelli, M. On-farm compost: a useful tool to improve soil quality under intensive farming systems. Appl Soil Ecol 2016, 107, 13-23. [CrossRef]
- Awasthi, M.K.; Zhang, Z.; Wang, Q.; Shen, F.; Li, R.; Li, D.; Ren, X.; Wang, M.; Chen, H.; Zhao, J. New insight with the effects of biochar amendment on bacterial diversity as indicators of biomarkers support the thermophilic phase during sewage sludge composting. Bioresource Technol 2017, 238, 589-601. [CrossRef]
- He, X.; Yin, H.; Han, L.; Cui, R.; Fang, C.; Huang, G. Effects of biochar size and type on gaseous emissions during pig manure/wheat straw aerobic composting: Insights into multivariate-microscale characterization and microbial mechanism. Bioresource Technol 2019, 271, 375-382. [CrossRef]
- Ma, S.; Fang, C.; Sun, X.; Han, L.; He, X.; Huang, G. Bacterial community succession during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane under slight positive pressure. Bioresource Technol 2018, 259, 221-227. [CrossRef]
- Babin, D.; Leoni, C.; Neal, A.L.; Sessitsch, A.; Smalla, K. Editorial to the Thematic Topic "Towards a more sustainable agriculture through managing soil microbiomes". Fems Microbiol Ecol 2021, 97. [CrossRef]
- Fierer, N.; Wood, S.A.; Bueno De Mesquita, C.P. How microbes can, and cannot, be used to assess soil health. Soil Biology and Biochemistry 2021, 153, 108111. [CrossRef]
- Bol, R.; Amelung, W.; Friedrich, C.; Ostle, N. Tracing dung-derived carbon in temperate grassland using 13C natural abundance measurements. Soil Biol Biochem 2000, 32, 1337-1343. [CrossRef]
- Duan, M.; Li, H.; Gu, J.; Tuo, X.; Sun, W.; Qian, X.; Wang, X. Effects of biochar on reducing the abundance of oxytetracycline, antibiotic resistance genes, and human pathogenic bacteria in soil and lettuce. Environ Pollut 2017, 224, 787-795. [CrossRef]
- Mawof, A.; Prasher, S.; Bayen, S.; Nzediegwu, C. Effects of Biochar and Biochar-Compost Mix as Soil Amendments on Soil Quality and Yield of Potatoes Irrigated with Wastewater. J Soil Sci Plant Nut 2021, 21, 2600-2612. [CrossRef]
- Purakayastha, T.J.; Bera, T.; Bhaduri, D.; Sarkar, B.; Mandal, S.; Wade, P.; Kumari, S.; Biswas, S.; Menon, M.; Pathak, H.; et al. A review on biochar modulated soil condition improvements and nutrient dynamics concerning crop yields: Pathways to climate change mitigation and global food security. Chemosphere 2019, 227, 345-365. [CrossRef]
- Nguyen, M.K.; Lin, C.; Hoang, H.G.; Sanderson, P.; Dang, B.T.; Bui, X.T.; Nguyen, N.S.H.; Vo, D.N.; Tran, H.T. Evaluate the role of biochar during the organic waste composting process: A critical review. Chemosphere (Oxford) 2022, 299, 134488. [CrossRef]
- Sanchez-Monedero, M.A.; Cayuela, M.L.; Roig, A.; Jindo, K.; Mondini, C.; Bolan, N. Role of biochar as an additive in organic waste composting. Bioresource Technol 2018, 247, 1155-1164. [CrossRef]
- Agegnehu, G.; Srivastava, A.K.; Bird, M.I. The role of biochar and biochar-compost in improving soil quality and crop performance: A review. Appl Soil Ecol 2017, 119, 156-170. [CrossRef]
- Fu, T.; Shangguan, H.; Wu, J.; Tang, J.; Yuan, H.; Zhou, S. Insight into the synergistic effects of conductive biochar for accelerating maturation during electric field-assisted aerobic composting. Bioresource Technol 2021, 337, 125359. [CrossRef]
- Guo, X.; Liu, H.; Zhang, J. The role of biochar in organic waste composting and soil improvement: A review. Waste Manage 2020, 102, 884-899. [CrossRef]
- Kung, C.; Mu, J.E. Prospect of China's renewable energy development from pyrolysis and biochar applications under climate change. Renewable and Sustainable Energy Reviews 2019, 114, 109343. [CrossRef]
- Sun, X.; Ma, S.; Han, L.; Li, R.; Schlick, U.; Chen, P.; Huang, G. The effect of a semi-permeable membrane-covered composting system on greenhouse gas and ammonia emissions in the Tibetan Plateau. J Clean Prod 2018, 204, 778-787. [CrossRef]
- Xiao, L.; Feng, L.R.; Yuan, G.D.; Wei, J. Low-cost field production of biochars and their properties. Environ Geochem Hlth 2020, 42, 1569-1578. [CrossRef]
- Yin, Y.; Tao, X.; Du, Y.; Li, M.; Yang, S.; Zhang, W.; Yang, C.; Li, H.; Wang, X.; Chen, R. Biochar improves the humification process during pig manure composting: Insights into roles of the bacterial community and metabolic functions. J Environ Manage 2024, 355, 120463. [CrossRef]
- Xu, N.; Tan, G.C.; Wang, H.Y.; Gai, X.P. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur J Soil Biol 2016, 74, 1-8. [CrossRef]
- Liu, N.; Zhou, J.; Han, L.; Ma, S.; Sun, X.; Huang, G. Role and multi-scale characterization of bamboo biochar during poultry manure aerobic composting. Bioresource Technol 2017, 241, 190-199. [CrossRef]
- He, X.; Yin, H.; Fang, C.; Xiong, J.; Han, L.; Yang, Z.; Huang, G. Metagenomic and q-PCR analysis reveals the effect of powder bamboo biochar on nitrous oxide and ammonia emissions during aerobic composting. Bioresource Technol 2021, 323, 124567. [CrossRef]
- He, X.; Chen, L.; Han, L.; Liu, N.; Cui, R.; Yin, H.; Huang, G. Evaluation of biochar powder on oxygen supply efficiency and global warming potential during mainstream large-scale aerobic composting. Bioresource Technol 2017, 245, 309-317. [CrossRef]
- Xiong, J.; Ma, S.; He, X.; Han, L.; Huang, G. Nitrogen transformation and dynamic changes in related functional genes during functional-membrane covered aerobic composting. Bioresource Technol 2021, 332, 125087. [CrossRef]
- Fang, C.; Yuan, X.; Liao, K.; Qu, H.; Han, L.; He, X.; Huang, G. Micro-aerobic conditions based on membrane-covered improves the quality of compost products: Insights into fungal community evolution and dissolved organic matter characteristics. Bioresource Technol 2022, 362, 127849. [CrossRef]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and Experimental Application of a Novel Non-Degenerate Universal Primer Set that Amplifies Prokaryotic 16S rRNA Genes with a Low Possibility to Amplify Eukaryotic rRNA Genes. Dna Res 2014, 21, 217-227. [CrossRef]
- López-Cano, I.; Roig, A.; Cayuela, M.L.; Alburquerque, J.A.; Sánchez-Monedero, M.A. Biochar improves N cycling during composting of olive mill wastes and sheep manure. Waste Management (Elmsford) 2016, 49, 553-559. [CrossRef]
- Sánchez-García, M.; Alburquerque, J.A.; Sánchez-Monedero, M.A.; Roig, A.; Cayuela, M.L. Biochar accelerates organic matter degradation and enhances N mineralisation during composting of poultry manure without a relevant impact on gas emissions. Bioresource Technol 2015, 192, 272-279. [CrossRef]
- Wei, Y.; Zhao, Y.; Shi, M.; Cao, Z.; Lu, Q.; Yang, T.; Fan, Y.; Wei, Z. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresource Technol 2018, 247, 190-199. [CrossRef]
- Xie, T.; Zhang, Z.; Zhang, D.; Wei, C.; Lin, Y.; Feng, R.; Nan, J.; Feng, Y. Effect of hydrothermal pretreatment and compound microbial agents on compost maturity and gaseous emissions during aerobic composting of kitchen waste. Sci Total Environ 2023, 854, 158712. [CrossRef]
- Tiquia, S.M.; Tam, N.F.Y. Elimination of phytotoxicity during co-composting of spent pig-manure sawdust litter and pig sludge. Bioresource Technol 1998, 65, 43-49. [CrossRef]
- Kong, Y.; Zhang, J.; Yang, Y.; Liu, Y.; Zhang, L.; Wang, G.; Liu, G.; Dang, R.; Li, G.; Yuan, J. Determining the extraction conditions and phytotoxicity threshold for compost maturity evaluation using the seed germination index method. Waste Management (Elmsford) 2023, 171, 502-511. [CrossRef]
- Zhang, L.; Sun, X. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar. Bioresource Technol 2014, 171, 274-284. [CrossRef]
- Ottani, F.; Parenti, M.; Santunione, G.; Moscatelli, G.; Kahn, R.; Pedrazzi, S.; Allesina, G. Effects of different gasification biochar grain size on greenhouse gases and ammonia emissions in municipal aerated composting processes. J Environ Manage 2023, 331, 117257. [CrossRef]
- Yu, H.; Xie, B.; Khan, R.; Shen, G. The changes in carbon, nitrogen components and humic substances during organic-inorganic aerobic co-composting. Bioresource Technol 2019, 271, 228-235. [CrossRef]
- Naeem, M.A.; Khalid, M.; Aon, M.; Abbas, G.; Amjad, M.; Murtaza, B.; Khan, W.; Ahmad, N. Combined application of biochar with compost and fertilizer improves soil properties and grain yield of maize. J Plant Nutr 2018, 41, 112-122. [CrossRef]
- Liu, H.; Awasthi, M.K.; Zhang, Z.; Syed, A.; Bahkali, A.H. Evaluation of gases emission and enzyme dynamics in sheep manure compost occupying with peach shell biochar. Environmental Pollution (1987) 2024, 351, 124065. [CrossRef]
- Aina Najwa Mohd Nor Azman, N.; Inderakusumowati Md Khalid, P.; Aishah Saidina Amin, N.; Yamani Zakaria, Z.; Mohammad Zainol, M.; Ilham, Z.; Phaiboonsilpa, N.; Asmadi, M. Effects of biochar, compost, and composted biochar soil amendments on okra plant growth. Materials Today : Proceedings 2023. [CrossRef]
- Neville, F.; Seyfaee, A. Real-Time Monitoring of in Situ Polyethyleneimine-Silica Particle Formation. Langmuir 2013, 29, 14681-14690. [CrossRef]
- He, X.; Han, L.; Fu, B.; Du, S.; Liu, Y.; Huang, G. Effect and microbial reaction mechanism of rice straw biochar on pore methane production during mainstream large-scale aerobic composting in China. J Clean Prod 2019, 215, 1223-1232. [CrossRef]
- Mao, H.; Lv, Z.; Sun, H.; Li, R.; Zhai, B.; Wang, Z.; Awasthi, M.K.; Wang, Q.; Zhou, L. Improvement of biochar and bacterial powder addition on gaseous emission and bacterial community in pig manure compost. Bioresource Technol 2018, 258, 195-202. [CrossRef]
- Wang, P.; Wang, S.; Chen, F.; Zhang, T.; Kong, W. Preparation of two types plant biochars and application in soil quality improvement. Sci Total Environ 2024, 906, 167334. [CrossRef]
- Bennur, T.; Kumar, A.R.; Zinjarde, S.; Javdekar, V. Nocardiopsis species: Incidence, ecological roles and adaptations. Microbiol Res 2015, 174, 33-47. [CrossRef]
- Ibrahim, A.H.; Desoukey, S.Y.; Fouad, M.A.; Kamel, M.S.; Gulder, T.A.M.; Abdelmohsen, U.R. Natural Product Potential of the Genus Nocardiopsis. Mar Drugs 2018, 16. [CrossRef]
- Jepsen, S.E.; Krause, M.; Gruttner, H. Reduction of Fecal Streptococcus and Salmonella by selected treatment methods for sludge and organic waste. Water Sci Technol 1997, 36, 203-210. [CrossRef]
- Li, X.; Shi, X.; Lu, M.; Zhao, Y.; Li, X.; Peng, H.; Guo, R. Succession of the bacterial community and functional characteristics during continuous thermophilic composting of dairy manure amended with recycled ceramsite. Bioresource Technol 2019, 294, 122044. [CrossRef]
- Wang, J.; Gu, J.; Wang, X.; Song, Z.; Dai, X.; Guo, H.; Yu, J.; Zhao, W.; Lei, L. Enhanced removal of antibiotic resistance genes and mobile genetic elements during swine manure composting inoculated with mature compost. J Hazard Mater 2021, 411, 125135. [CrossRef]
- Exposito, R.G.; Postma, J.; Raaijmakers, J.M.; De Bruijn, I. Diversity and Activity of Lysobacter Species from Disease Suppressive Soils. Front Microbiol 2015, 6. [CrossRef]
- Puopolo, G.; Tomada, S.; Pertot, I. The impact of the omics era on the knowledge and use of Lysobacter species to control phytopathogenic micro-organisms. J Appl Microbiol 2018, 124, 15-27. [CrossRef]
- Qin, S.; Feng, W.; Zhang, Y.; Wang, T.; Xiong, Y.; Xing, K. Diversity of Bacterial Microbiota of Coastal Halophyte Limonium sinense and Amelioration of Salinity Stress Damage by Symbiotic Plant Growth-Promoting Actinobacterium Glutamicibacter halophytocola KLBMP 5180. Appl Environ Microb 2018, 84. [CrossRef]
- Cordovez, V.; Carrion, V.J.; Etalo, D.W.; Mumm, R.; Zhu, H.; van Wezel, G.P.; Raaijmakers, J.M. Diversity and functions of volatile organic compounds produced by Streptomyces from a disease-suppressive soil. Front Microbiol 2015, 6. [CrossRef]
Figure 1.
Dynamical changes in (a) temperature and oxygen concentration, (b) pH and germination index (GI), (c) organic matter and ammonia (NH4+), (d) total nitrogen (TN) and total carbon (TC) in the physicochemical properties of compost samples.
Figure 1.
Dynamical changes in (a) temperature and oxygen concentration, (b) pH and germination index (GI), (c) organic matter and ammonia (NH4+), (d) total nitrogen (TN) and total carbon (TC) in the physicochemical properties of compost samples.
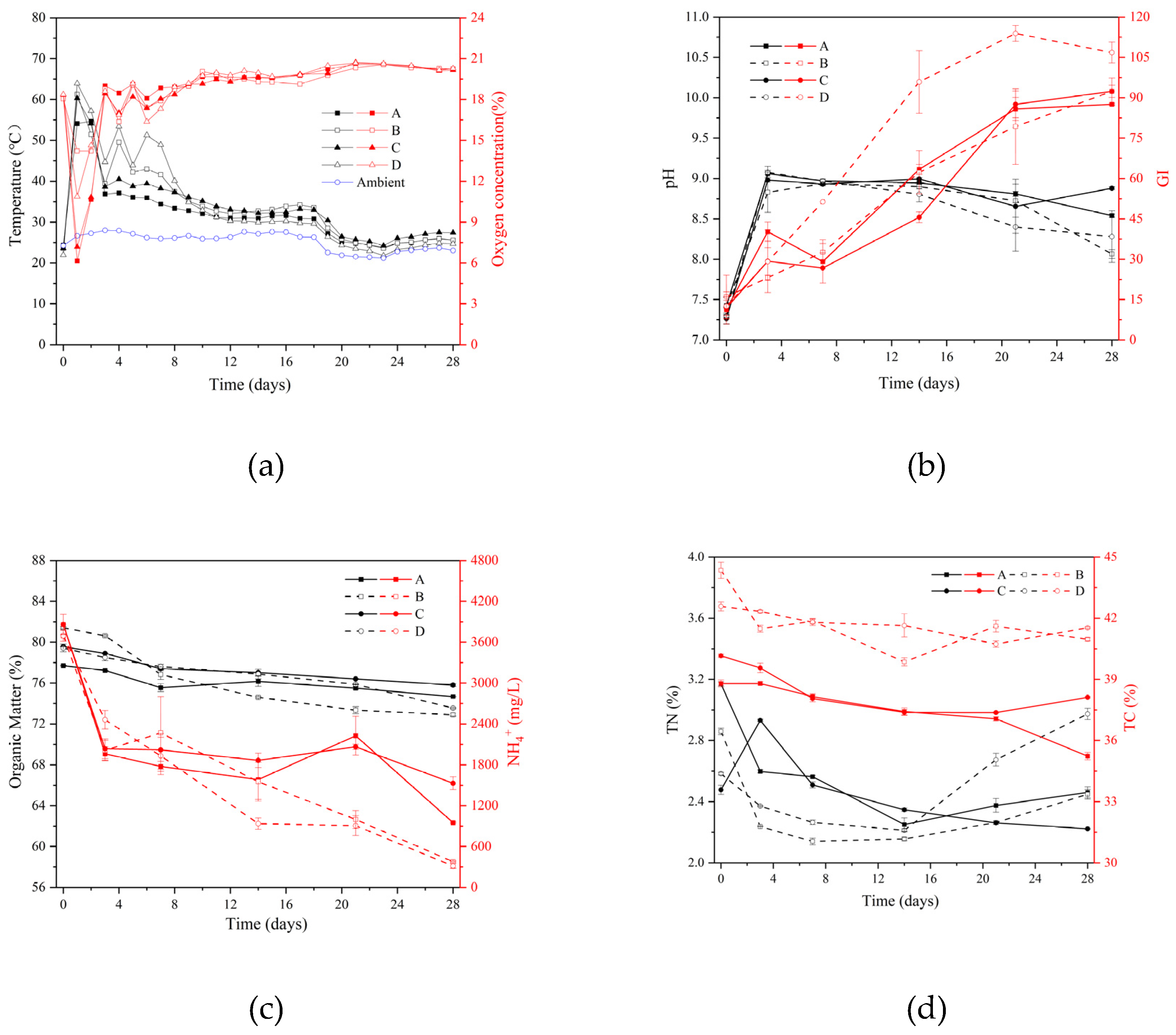
Figure 2.
Dynamic changes in soil samples’ physicochemical properties: (a) pH, (b) organic matter, (c) total carbon (TC) and (d) total nitrogen (TN).
Figure 2.
Dynamic changes in soil samples’ physicochemical properties: (a) pH, (b) organic matter, (c) total carbon (TC) and (d) total nitrogen (TN).
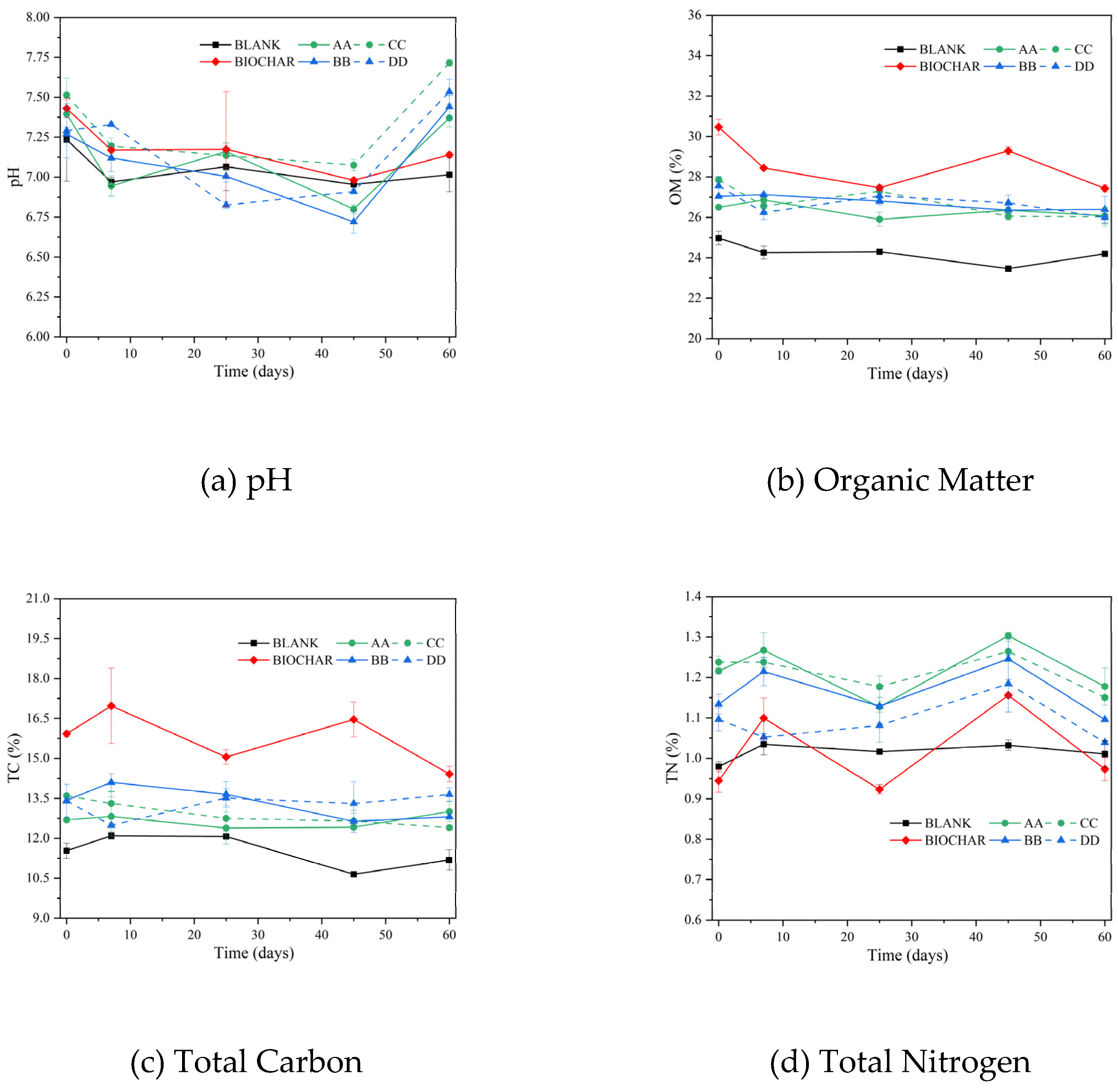
Figure 3.
Hierarchical clustering of microbes (a) and typing analysis (b) on the genus level.
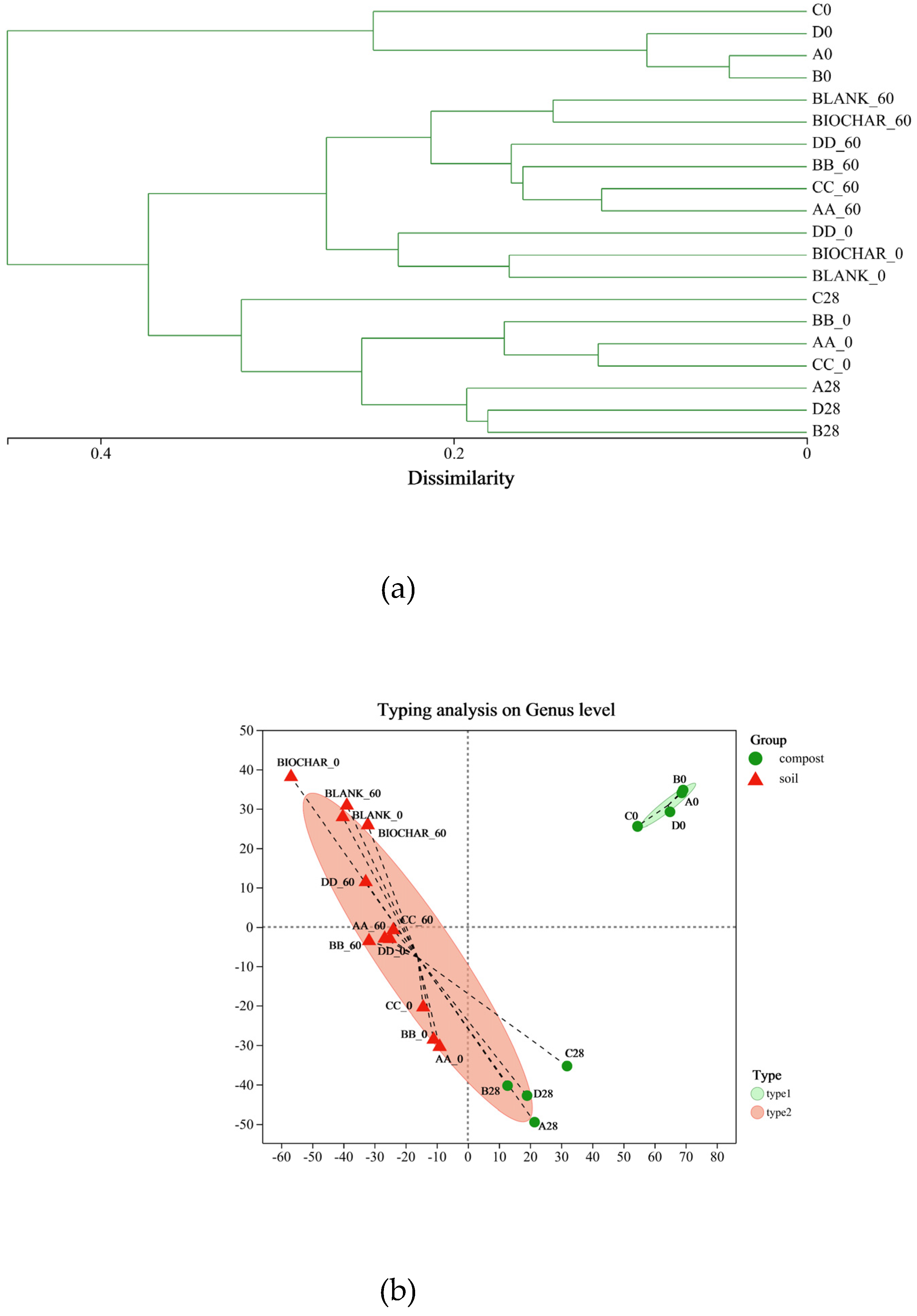
Figure 4.
Microbial composition histogram at phylum level (a) and genus level (b).
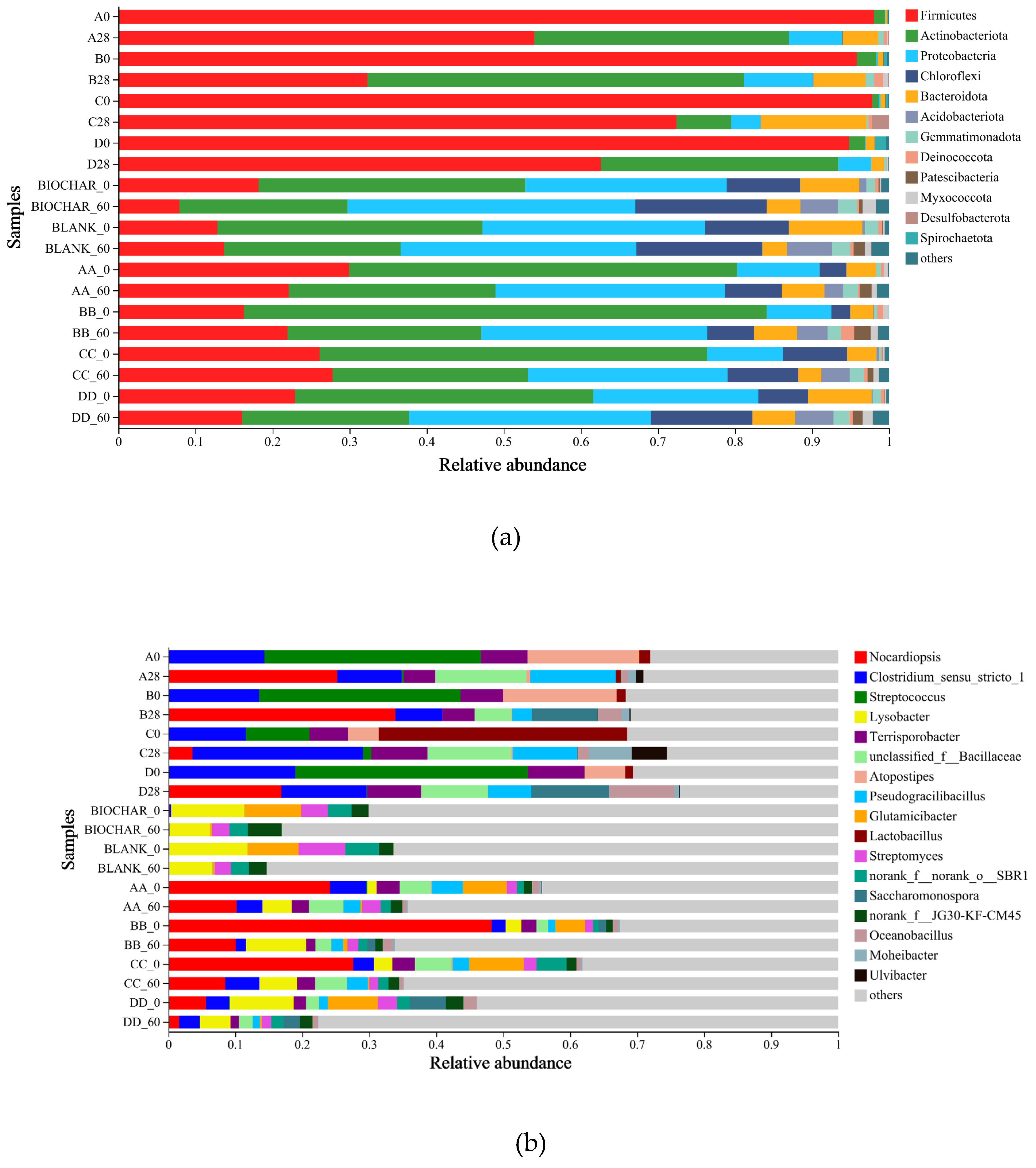
Figure 5.
Difference analysis of microbes at the genus level in compost and soil samples.
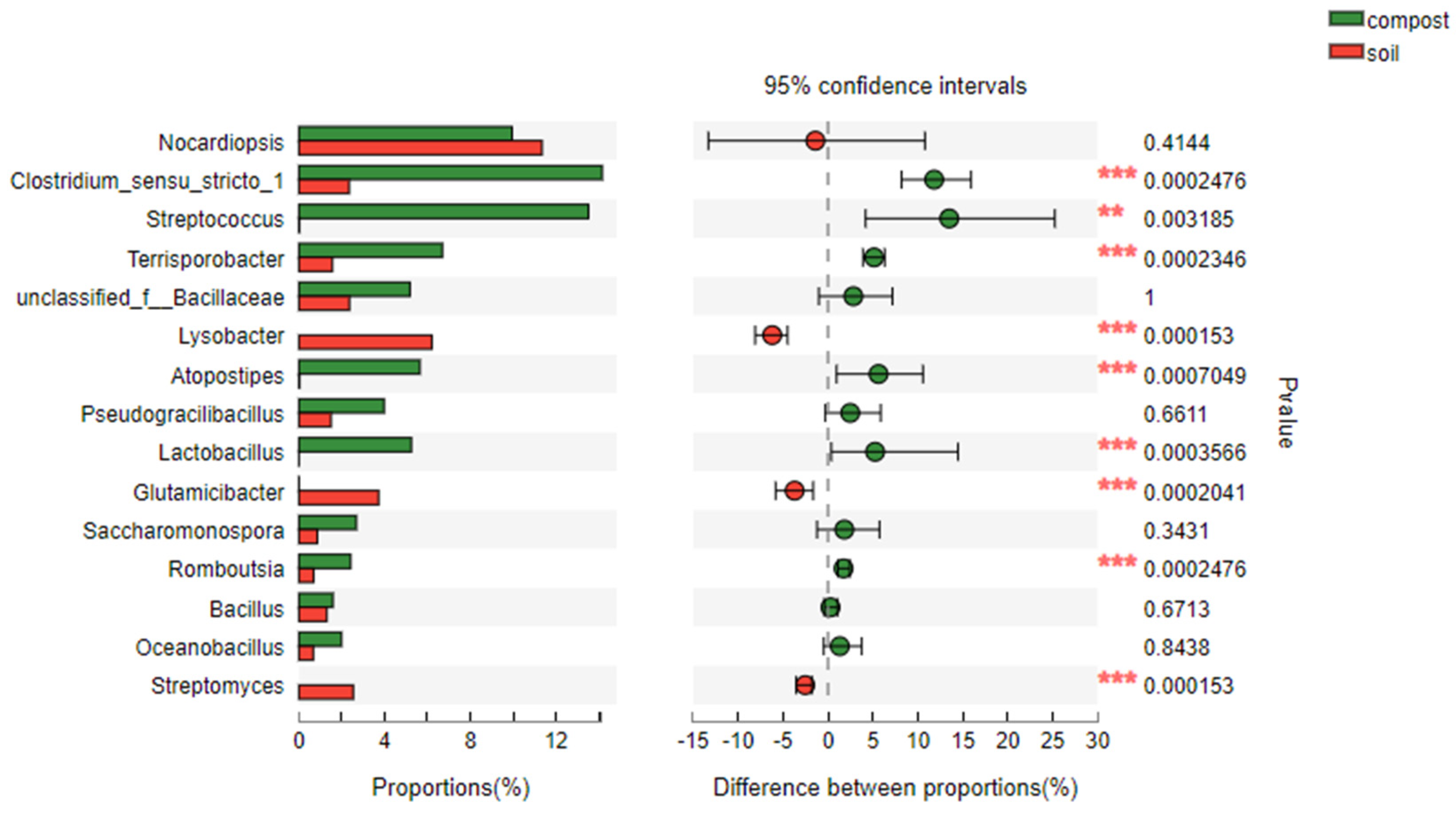
Figure 6.
Canonical correlation analysis (CCA) (a) and Spearman correlation heatmap (b) between physicochemical properties and microbes at the genus level.
Figure 6.
Canonical correlation analysis (CCA) (a) and Spearman correlation heatmap (b) between physicochemical properties and microbes at the genus level.
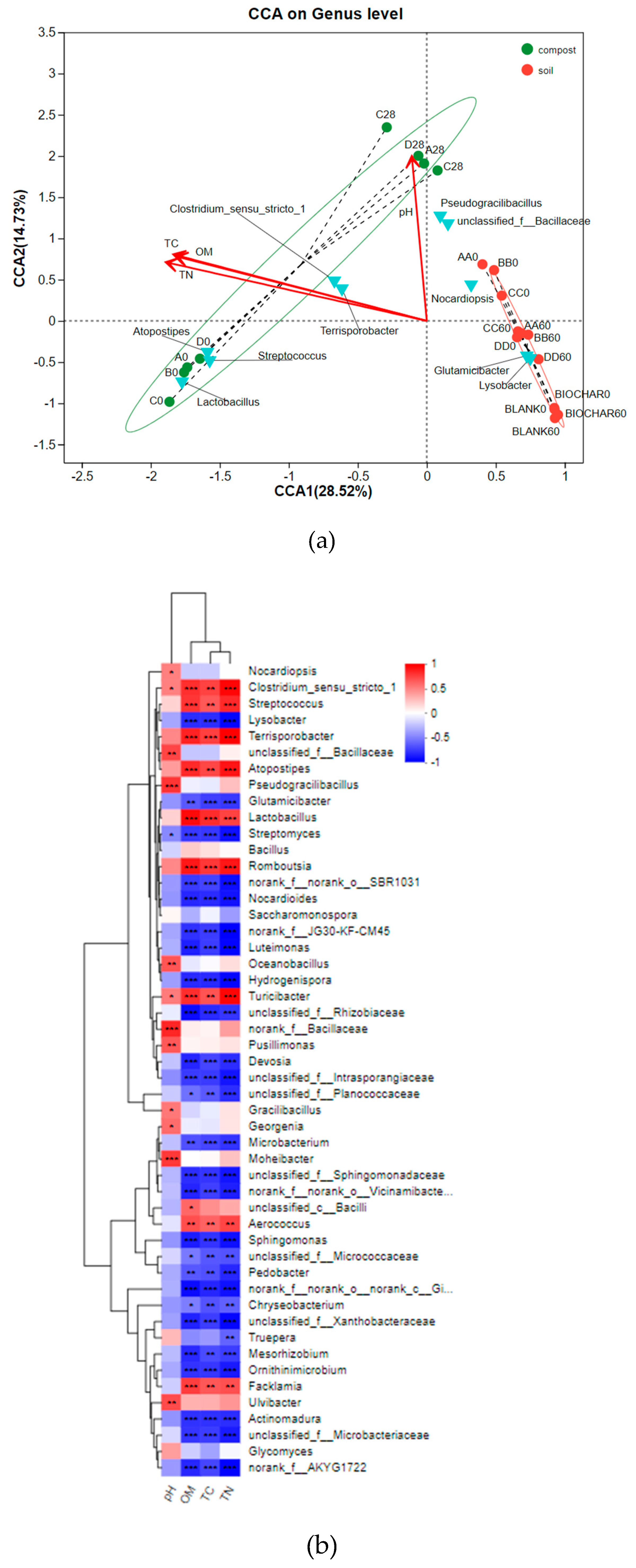

Table 1.
Ingredients and group name of aerobic composting and potting experiments.
| Experiment | Group name | Ingredients a |
|---|---|---|
| Aerobic Composting | Control groups (A and C) | Swine manure and wheat straw (Ratio of 5:1) |
| Experimental groups (B and D) | Swine manure, wheat straw, and wheat straw biochar (Ratio of 10:2:1) |
|
| Potting b | BLANK | 0.25 kg nutrient soil |
| BIOCHAR | 0.225 kg nutrient soil + 0.025 kg biochar | |
| AA | 0.225 kg nutrient soil + 0.025 kg compost from the Group A composting | |
| BB | 0.225 kg nutrient soil + 0.025 kg biochar-based compost from Group B | |
| CC | 0.225 kg nutrient soil + 0.025 kg compost from Group C | |
| DD | 0.225 kg nutrient soil + 0.025 kg biochar-based compost from Group D |
Notes: Nutrient soil: Black soil and vermiculite in the ratio of 5:1. a: Based on wet basis. b: The weight of ingredients in the potting experiment was for one pot (six pots in each group).
Table 2.
Basic physicochemical properties of raw materials for aerobic composting and pot experiments.
Table 2.
Basic physicochemical properties of raw materials for aerobic composting and pot experiments.
| Name | MC a | pH a | OM b | TC b | TN b |
|---|---|---|---|---|---|
| Pig manure | 66.00 ± 0.72 | / | 74.70 ± 0.01 | 37.41 ± 0.14 | 3.19 ± 0.01 |
| Wheat straw | 6.07 ± 0.14 | / | 89.81 ± 0.03 | 43.43 ± 0.09 | 0.99 ± 0.06 |
| Biochar | 3.21 ± 0.15 | 10.21 ± 0.07 | 80.99 ± 0.28 | 70.60 ± 0.16 | 1.08 ± 0.01 |
| Black soil | 2.71 ± 0.15 | 6.87 ± 0.01 | 4.51 ± 0.06 | 12.60 ± 0.24 | 1.12 ± 0.02 |
| Vermiculite | 5.62 ± 0.03 | 7.25 ± 0.01 | 25.05 ±0.68 | 0.16 ± 0.01 | 0.20 ± 0.01 |
| Compost A | 13.32 ± 0.06 | 7.96 ± 0.01 | 65.57 ± 0.07 | 33.36 ± 0.59 | 3.97 ± 0.06 |
| Compost C | 13.38 ±0.15 | 8.07 ± 0.03 | 65.30 ± 0.22 | 32.51 ± 0.02 | 3.86 ± 0.01 |
| Compost B | 10.80 ± 0.36 | 8.54 ± 0.04 | 67.98 ± 0.09 | 41.41 ± 0.15 | 3.34 ± 0.01 |
| Compost D | 10.31 ± 0.10 | 8.15 ± 0.03 | 71.47 ± 0.28 | 43.20 ± 0.06 | 2.67 ± 0.02 |
Notes: MC: moisture content, %; OM: organic matter, %; TC: total carbon, %; TN: total nitrogen, %, a: Based on wet basis, b: Based on dry basis. Composts A and C: without biochar additive during aerobic composting. Composts B and D: with biochar additive during aerobic composting.
Table 3.
Microbial diversity by Alpha index of compost and soil samples.
| Sample name | Chao | Shannon | Simpson | Shannoneven | Simpsoneven |
|---|---|---|---|---|---|
| A0 | 252 | 3.314 | 0.111 | 0.599 | 0.036 |
| C0 | 243 | 3.131 | 0.155 | 0.570 | 0.026 |
| B0 | 294 | 3.542 | 0.098 | 0.623 | 0.035 |
| D0 | 260 | 3.273 | 0.128 | 0.589 | 0.030 |
| A28 | 199 | 3.840 | 0.060 | 0.725 | 0.084 |
| B28 | 136 | 3.231 | 0.123 | 0.658 | 0.060 |
| C28 | 268 | 4.092 | 0.058 | 0.732 | 0.064 |
| D28 | 145 | 3.430 | 0.068 | 0.689 | 0.101 |
| BLANK_0 | 635 | 5.449 | 0.014 | 0.844 | 0.114 |
| AA_0 | 351 | 4.466 | 0.045 | 0.762 | 0.063 |
| BB_0 | 278 | 3.372 | 0.199 | 0.599 | 0.018 |
| CC_0 | 330 | 4.515 | 0.031 | 0.778 | 0.096 |
| DD_0 | 436 | 4.967 | 0.019 | 0.817 | 0.121 |
| BIOCHAR_0 | 1035 | 5.739 | 0.014 | 0.827 | 0.070 |
| BLANK_60 | 896 | 6.259 | 0.004 | 0.921 | 0.298 |
| AA_60 | 761 | 5.830 | 0.008 | 0.879 | 0.171 |
| BB_60 | 856 | 5.771 | 0.014 | 0.855 | 0.083 |
| CC_60 | 661 | 5.739 | 0.008 | 0.884 | 0.179 |
| DD_60 | 987 | 6.288 | 0.004 | 0.912 | 0.243 |
| BIOCHAR_60 | 758 | 6.145 | 0.004 | 0.927 | 0.332 |
Notes: Alpha index is mainly used to study diversity of communities in certain habitats (or sample), reflecting information such as species, richness, and diversity of samples. Chao reflects community richness; Shannon and Simpson reflect community diversity; and Simpsoneven and Shannoneven reflect community evenness. Initial aerobic composting samples on Day 0 of A, B, C, and D were named A0, B0, C0, and D0. Final aerobic composting samples on Day 28 of A, B, C, and D were named A28, B28, C28, and D28. Initial soil samples of potting experiment on Day 0 of BLANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_0, BIOCHAR_0, AA_0, BB_0, CC_0, and DD_0. Final soil samples of potting experiment on Day 60 of BLANK, BIOCHAR, AA, BB, CC, and DD were named BLANK_60, BIOCHAR_60, AA_60, BB_60, CC_60, and DD_60.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



