
Submitted:
21 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
This study aimed to analyze the population dynamics of the mite Varroa destructor in honey bee (Apis mellifera) colonies in a temperate semi-arid climate in Mexico. Ten colonies homogeneous in population, food stores, and levels of mite infestation, were used. The mite infestation rate in brood and adult bees, total number of mites, daily mite fall, brood and adult bee population, and food stores, were determined periodically for 10 months. There was a significant effect (P <0.05) of sampling period on the population of V. destructor in adult bees, brood, total mite population, and daily fallen mites. The total mite population increased 26% on average per colony. The increase in brood amount reduced the mite infestation rate in adult bees, and the opposite occurred when the brood decreased. Monitoring V. destructor populations by recording fallen mites is more reliable than determining mite infestation rates in bees, as mite fall has a dynamic pattern similar to that of the total mite population. The best period to apply an acaricide treatment in the region of study is between November and December because most mites were in the phoretic phase, since there was less brood in the colonies compared to other times.
Keywords:
Apis mellifera
; Varroa destructor
; population dynamics
; temperate semi-arid climate
1. Introduction
The mite Varroa destructor (Acari: Varroidae) [1], is the main health problem for the beekeeping industry worldwide [2]. This is because the mite has had a relatively recent association with Apis mellifera, the western honey bee, and has rapidly spread to almost all regions where western honey bee colonies are managed [3,4]. The mite feeds on the hemolymph and fat tissue of the brood and adult bees, inhibiting their immune system, making them more susceptible to bacteria, viruses, fungi, and pesticides [5,6,7,8,9]. Additionally, V. destructor transmits several viruses to its hosts [10,11,12]. It also shortens the lifespan of parasitized bees [13], reducing the populations and honey yields of their colonies [14,15,16]. For all these reasons, V. destructor is one of the main factors associated with the high loss of colonies worldwide [17,18,19].
Besides the damage caused by V. destructor to honey bee colonies, the application of acaricides (synthetic and organic) used for its control represents another problem because, in general, all have shown adverse effects to the bees [20,21,22,23,24].
To control mite populations in honey bee colonies and at the same time reduce the use of acaricides, it is necessary to identify the appropriate times to apply treatments. This knowledge would help reduce the negative effects of acaricides on bee health, the selection pressure for mites to develop resistance to the active ingredients of the chemicals, the risk of contamination of hive products, and the production cost for beekeepers due to unnecessary or excessive treatment applications [25,26,27,28,29].
Several reports on the population dynamics of V. destructor in honey bee colonies from different countries have been published [30,31,32,33,34], but none from Mexico. These reports are valuable for specific countries, but it is critical to take into consideration that the population dynamics of the mite varies regionally due to the seasonality of the brood period in the colonies and its effects on mite reproduction [27,28]. Additionally, very few studies have jointly evaluated the population levels of V. destructor in its different life phases (phoretic and reproductive) along with recording the daily mite drop and the population and food stores of honey bee colonies.
Studies on the population dynamics of V. destructor in honey bee colonies provide information on its evolution and changes in a given region and allow for the establishment of timely control strategies. This work reports for the first time in Mexico the population dynamics of V. destructor, considering its phoretic and reproductive phases, as well as natural mite drop, in relation to the population dynamics and food stores of honey bee colonies established in a temperate semi-arid climate.
2. Materials and Methods
2.1. Study Region
The study was conducted at an experimental apiary belonging to the Veterinary and Animal Science Academic Unit of the Autonomous University of Zacatecas in El Cordovel, General Enrique Estrada, municipality of the state of Zacatecas, Mexico (22°59'42’’N, 102°44'24’’W). This region is characterized by a temperate semi-arid climate, with an average annual precipitation of 400 to 700 mm and an average annual temperature of 18ºC [35].
2.2. Experimental Colonies
Ten nucleus colonies homogeneous in bee population, food stores, and V. destructor infestation levels were used. Six commercial colonies, which had not been exposed to acaricide treatments for more than two years, were used to create the nucleus colonies. The bees from these six colonies were shaken into a large wire cage to blend them. Each nucleus colony was composed of three brood frames and one frame containing honey and pollen. The frames were randomly taken from the six source colonies, while 2 kg of bees were taken from the cage for each nucleus colony [36]. These new colonies were housed in Langstroth hives identified with a number and distributed in a circular arrangement in the apiary, separated by 2 m from one another. Three days after establishing the colonies, sister queens of the same age and origin were introduced to the colonies. The colonies were fed weekly with 1.5 L of a 1:1 water/sucrose syrup and 250 g of a commercial food supplement containing 20% protein (Nutra®, X-Nox, Aguascalientes, Mexico). Additionally, frames with wax foundation were added as needed by the colonies.
2.3. V. destructor Infestation in Adult bees, Brood, and Daily Mite Fall
The infestation level of V. destructor in adult bees and worker brood, as well as the number of mites fallen in the experimental colonies, was determined on five occasions over a 10 month period (July 3, September 25, December 6, 2018, February 28, and May 6, 2019).
The mite infestation level in adult bees was determined as per De Jong et al. [37]. This technique involves collecting a sample of approximately 300 adult bees from the brood nest of each colony into a container with 75% ethanol. Subsequently, mechanical agitation was used to remove the mites adhered to the bees. The mite infestation rate was determined by dividing the total number of mites counted by the number of bees analyzed, and the result was multiplied by 100. The infestation level in worker brood was determined by dividing the number of mite infested cells in a section of capped brood comb (10 x 10 cm) by the number of cells analyzed, multiplied by 100 [38].
With the data on V. destructor infestation levels in adult bees and brood, and the population estimates of adult bees and capped brood (described later), the total population of mites in adult bees (phoretic phase) and in the brood (reproductive phase) of the experimental colonies was estimated. This was done by multiplying the average number of mites per bee or brood by the estimated population of adult bees or brood, respectively.
To record fallen mites, a galvanized metallic sheet (28 x 43.5 cm) impregnated with petrolatum was placed on the bottom board of each hive, and a metal mesh (3 mm) of the dimensions of the hive was installed between the sticky sheet and the brood chamber, so that the fallen mites would pass through the mesh and adhere to the sticky sheet. The average daily fallen mites was obtained by dividing the number of recorded parasites by four, which was the number of days the adhesive sheets remained in the hives [38].
2.4. Bee Population, Brood, and Food Stores of Experimental Colonies
The adult bee population, as well as the comb area containing honey and pollen in the colonies, was calculated by visually estimating the proportion of the surface of each side of combs occupied by these variables. This estimation was performed twice by two different observers, and the values were averaged. To calculate the population of adult bees, the surface proportion occupied by adult bees on each comb was multiplied by 2,430, which is the estimated number of adult individuals for a Langstroth brood chamber frame [36]. The proportions for honey and pollen were converted to area (cm²) using the surface area of a Langstroth frame on both sides (1,760 cm²) [36]. The brood population was estimated by determining the area (cm²) of capped brood in the colonies. For this purpose, all the combs of each colony were photographed on both sides, and using the ImageJ® software (National Institute of Health, Maryland, USA), the area in cm² of capped brood in each comb and colony was determined. The amount of brood was estimated by multiplying the areas of capped brood by the number of brood cells per cm² (3.9) [36]. Measurements were taken between 6:00 PM and 7:00 PM, when most of the bees were inside the hives. The estimates of the population size of bees, brood, and food stores were also done every 72 days for 10 months, starting in July 2018.
2.5. Statistical Analyses
The data on V. destructor infestation rates in brood and adult bees were arcsine-square root transformed to normalize their distribution. For non-percentage data, normality was verified using the Shapiro-Wilk test. Analysis of variance was used to compare the initial values of V. destructor infestation rates, daily mite fall, bee population, and food reserves of the experimental colonies. Measures of central tendency and dispersion were obtained for V. destructor infestation levels, bee population, and food stores of the colonies. Repeated measures analysis of variance and Tukey tests were also used to compare the effect of sampling time on the measured variables. Additionally, Pearson correlation tests were performed to establish relationships between the evaluated variables. All analyses were conducted using SAS software, version 9.0. [39].
3. Results
The colonies did not differ at the beginning of the study for infestation levels in adult bees (F1,9 = 2.51, P = 0.35), brood (F1,9 = 2.42, P = 0.36), and daily fall of V. destructor (F1,9 = 3.0, P = 0.33). There were also no differences between colonies for initial bee population (F1,9 = 1.72, P = 0.41), amount of brood (F1,9 = 0.81, P = 0.53), and areas of pollen (F1,9 = 0.48, P = 0.61) and honey (F1,9 = 1.69, P = 0.42).
During the course of the study, significant differences were found between sampling periods for V. destructor infestation rates in adult bees (F4,46 = 4.08, P = 0.008) and brood (F4,46 = 11.82, P < 0.0001), as well as for the number of fallen mites (F4,46 = 7.81, P = 0.0001), amount of brood (F4,46 = 39.27, P < 0.0001), adult bee population (F4,46 = 13.33, P < 0.0001), mite population in brood (F4,46 = 10.33, P < 0.0001), mite population in adult bees (F4,46 = 5.36, P = 0.001), total mite population (F4,46 = 13.51, P < 0.0001), areas of pollen (F4,46 = 15.19, P < 0.0001), and areas of honey (F4,46 = 25.59, P < 0.0001).
In September, the colonies had the lowest infestation rates in brood and adult bees and the lowest number of estimated total mites, while in December, the colonies had the highest mite infestation rate in the brood. In May, the colonies had the highest number of mites fallen onto the adhesive sheets, and the highest estimated number of total mites in the colonies (Figure 1).
Figure 2 shows that there was a greater population of adult bees and brood during the months of September and May, while the estimated number of V. destructor mites on adult bees showed significant differences only between September and May. Additionally, the estimated number of mites in the brood was significantly higher in May compared to the period between the previous June and December. Moreover, a reduction in the population of V. destructor in adult bees was observed in response to an increase in the amount of brood in the colonies, and an increase in the population of V. destructor in adult bees was observed when the amount of brood in the colonies decreased. It was also observed that in December, there was the lowest estimated number of V. destructor mites in the brood compared to the number of mites on adult bees.
A decrease (although not significant) in the estimated total number of V. destructor mites (including mites in both the reproductive and phoretic phases) relative to the populations of brood and adult bees, which increased significantly, was observed from July to September. The estimated total population of V. destructor in the colonies was lowest in September, with an average of 782±168 mites, a number that progressively and significantly increased to reach 2,715±350 mites in May. Following a similar pattern, the daily average of fallen mites showed no significant differences from July to February, with an average of 27.6±1.6 parasites, while the mean of that variable was significantly higher in May (75.3±5.6; Figure 3).
From September and until February, there was a parallel and significant increase in the total population of V. destructor and the honey and pollen stores in the brood chambers of the experimental colonies (Figure 4).
Significant correlations were found between the rates of V. destructor infestation (in adult bees and brood) and daily mite drop, total V. destructor population, and amount of brood. The daily mite drop was related to the infestation rate in adult bees. Additionally, the amount of brood was also related to the adult bee population, while the honey areas were significantly correlated with the pollen areas of the colonies (Table 1).
4. Discussion
The differences found in the estimated populations of V. destructor in brood, adult bees, and total mites, as well as for the number of parasites fallen between the analyzed time periods, are likely due to changes in environmental conditions and availability of food resources for the colonies over the 10 months monitored. These factors are known to influence variations in the queen's egg-laying rate [27,28,40], and consequently, the availability of brood for the reproduction of V. destructor.
There is a close relationship between the population dynamics of the parasite and its host, which is apparently confirmed by the reduction in mite infestation rate in brood and adult bees between July and September. This appears to be due to the increase in the brood and worker bee populations observed during that period, causing a dilution effect on the mite infestation rates. The increase in the amount of brood encouraged that many mites in the phoretic phase left the adult bees to enter cells containing larvae to reproduce. Conversely, the increase in the infestation rate of V. destructor in adult bees in December is likely a consequence of the mite reproduction between July and September and a reduction in the bee and brood populations (see Figure 1 and Figure 2). From September to January, the amount of brood decreased, thereby reducing the possibilities for mite reproduction and population growth. From February to May (spring blooming season), the amount of brood increased, leading to a significant multiplication of V. destructor, probably due to greater availability of food resources for the colonies during that time of year, resulting in greater production and availability of larvae for the mite to reproduce. Although there was no significant correlation between V. destructor population and the stores of honey and pollen, Figure 4 shows an increase in the mite population when food stores increased. In September and October, as well as between February and May, there are many plants blooming in the study area, which favored colony development and the storage of honey and pollen in the combs. It has been reported that the amount of pollen in the colonies is related to the fertility of the mite [41].
The results of this study show that when the amount of brood increases (between July and September), the mite infestation rate in adult bees decreases because many parasites migrate from the adult bees to the brood. Inversely, when the amount of brood decreases (September to December), the number of mites and the infestation rate in adult bees increases because there are not enough larvae for the parasites to reproduce [34].
The estimated total mite population tended to decrease when the bee population in the colonies increased (see Figure 3). This is possibly because before July there was little brood available for the parasite to reproduce, and this is simply a delayed effect of the population dynamics resulting in a decrease in mite population due to low reproduction during the period of little brood before July. However, from September onward, there was a progressive increase in the mite population.
The results presented in Figure 1 and Figure 3 show that monitoring V. destructor populations by recording fallen mites is more reliable than determining mite infestation rates in bees, as mite fall has a dynamic pattern similar to that of the estimated total mite population. Additionally, the techniques for determining infestation levels in brood and adult bees are destructive, negatively impacting the colony population, particularly when infestation levels are determined frequently [42]. This conclusion is reinforced by the correlation found between daily mite drop and the estimated total mite population in the colonies. Overall, the results confirm that mite fall is a relatively effective technique for determining V. destructor infestation levels in colonies.
Contrary to what was reported by Branco et al. [42], our results suggest that the rate of V. destructor in adult bees and brood is an imprecise and uninformative measure for estimating the total mite population in the colonies. This is due to the migration of V. destructor during the phoretic and reproductive phases of the mite and the fact that the population dynamics of the parasite are related to the cycle and amount of brood and adult bees in the colonies, which vary across different seasons of the year [27,28]. The increase of brood and adult bee population in the colonies reduces the proportion of mites relative to the bee population, while a decrease in the colony population leads to a higher mite infestation rate [43]. Additionally, when the amount of brood decreases and the number of adult bees increases, the percentage of mite infestation in adult bees rises, as occurred during December, the time of year with the least brood area in the colonies, which caused most mites to remain in the phoretic phase, concentrating the V. destructor population on adult bees (see Figure 2).
Clearly, the period from November to December is optimal for applying an acaricide, as most of these chemical products do not affect mites inside capped brood cells but do affect them in the phoretic phase. Thus, during these months (at the end of the fall honey harvest), there is a window of opportunity to apply an acaricide treatment in regions with a temperate semi-arid climate of the Mexican highlands. Specifically, this is the time of year when products such as oxalic acid are most effective [44]. Additionally, it is important to consider that acaricide treatment during this period will limit V. destructor reproduction before the expected increase in bee population in the early part of the year (starting in February). It is also important to consider that the treatment threshold suggested for Mexico by SAGARPA [45] is 5% infestation in adult bees and/or 10 mites fallen in 24 h, which aligns with the results of a study conducted in the central highlands region of Mexico, where it was found that infestation levels of 4.5% in adult bees do not significantly affect population parameters, food stores, and colony weight [46]. Proper and timely application of a treatment helps reduce the negative effects of acaricides on bees and brood, as well as the selection pressure for resistant mites, the risks of contamination of hive products, the production cost for beekeepers due to unnecessary treatment applications, and prevents relaxing the selection pressure to develop mite-resistant honey bee colonies [25,26,27,28,29].
In a study conducted with Africanized honey bee colonies in the Mexican tropics, it was found that the population of V. destructor decreased by more than 1,000 mites over twelve months. This was attributed to the low production of fertile mites in each reproductive cycle of V. destructor (0.7) during a time of year with high mortality of worker larvae [47]. Conversely, in the present study conducted in a temperate semi-arid region of Mexico with colonies of bees with greater European ancestry, an increase of 1,136 mites per colony on average over ten months was observed, representing a 26% increase in the total mite population. Considering the record of mites fallen onto the adhesive boards of the hives, there was a 39% increase in final infestation levels compared to the initial levels (33±1.5 mites in July vs. 75.3±5.6 in May). The increase in mite population observed in this study is similar to that reported in honey bee colonies in Costa Rica [32]. The population size of drone brood in the colonies and the low expression of resistance mechanisms to the mite may have been factors that partly explain the population increase of V. destructor in this study, although many more factors not studied here could have contributed to the mite's population growth and its temporal variations.
The population dynamics of V. destructor are variable and depend on multiple factors and their interactions [44,48]. The main characteristics that determine the population growth rate of V. destructor and its pathogenicity in honey bee colonies are its reproductive ability and longevity [44,49], as well as the climatic conditions and nectar flow that influence the availability of brood, drones, and the generation of swarms [27,28,40], management practices [48], and the overcrowding of colonies, which can favor bee drift and robbing [50,51,52]. Other factors include the genotype of the colonies [53,54,55] and the expression of bee defense mechanisms that limit the reproduction and survival of the mite. The mechanisms of resistance that most restrain the population growth of V. destructor in honey bee populations of Latin America include hygienic behavior, grooming behavior, low brood attractiveness, suppression of mite reproduction and other mite non-reproduction related mechanisms that result in low fertility and fecundity of the mite [56,57].
Studies have reported variable results regarding the correlations between colony population parameters and V. destructor infestation levels, as well as correlations between infestation levels measured by different methods [26,30,32,41,42,47,53]. Therefore, there is no consensus on the effect of these parameters on the population dynamics of the mite. Moreover, complex multifactorial interactions exist that make it difficult to accurately predict mite populations in colonies [45]. The results of this study provide new insights into the population dynamics of V. destructor in a temperate semi-arid climate region and suggest that more studies are needed to identify the limiting factors of mite reproduction, as well as the seasonality of mite population dynamics in different regions and climates. This knowledge will help establish sustainable control strategies for this harmful parasite of honey bees.
5. Conclusions
The results of this study demonstrate a close relationship between the population dynamics of V. destructor and that of honey bee colonies. In December, there was an increase in the number of mites infesting adult bees, as well as a decrease in the number of mites infesting brood. Therefore, it is advisable to apply an acaricide treatment during this period. Another significant finding of this study is that estimating V. destructor infestation levels of honey bee colonies relying on mite drop data, provides a more reliable indication of the mite population in the colonies than data on brood and adult bee infestation rates. Similar studies are needed for other regions and climates, as well as studies that relate climatic variables to the population dynamics of V. destructor to generate knowledge that will enable the development of effective and sustainable mite control strategies.
Author Contributions
Conceptualization, all authors; data curation, A.S.R and C.A.M.-F.; methodology and formal analysis, C.A.M.-F., E.G.-N and L.A.G.; Investigation, A.S.R.; visualization, E.G.-N., writing—original draft preparation, C.A.M.-F. and E.G.-N.; writing—review and editing, C.A.M.-F., E.G.-N and L.A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data are available in this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari: Varroidae) Is More than One Species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the major ectoparasite of the western honey bee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Traynor, K.S.; Mondet, F.; de Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.Y.; Chantawannakul, P.; McAfee, A. Varroa destructor: A complex parasite, crippling honey bees worldwide. Trends Parasitol. 2020, 36, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.C.; Colin, T.; Cook, J.; da Silva, C.R.B.; Gloag, R.; Hogendoorn, K.; Howard, S.R.; Remnant, E.J.; Roberts, J.M.K.; Tierney, S.M.; et al. The final frontier: Ecological and evolutionary dynamics of a global parasite invasion. Biol. Lett. 2023, 19, 20220589. [Google Scholar] [CrossRef] [PubMed]
- Bowen-Walker, P.L.; Gunn, A. The effect of the ectoparasitic mite, Varroa destructor on adult worker honeybee (Apis mellifera) emergence weights, water, protein, carbohydrate, and lipid levels. Entomol. Exp. Appl. 2001, 101, 207–217. [Google Scholar] [CrossRef]
- vanEngelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef]
- Koleoglu, G.; Goodwin, P.H.; Reyes-Quintana, M.; Hamiduzzaman, M.M; Guzman-Novoa, E. Effect of Varroa destructor, wounding and Varroa homogenate on gene expression in brood and adult honey bees. PLoS ONE 2017, 12, e0169669. [Google Scholar] [CrossRef]
- Koleoglu, G.; Goodwin, P.H.; Reyes-Quintana, M.; Hamiduzzaman, M.M.; Guzman-Novoa, E. Varroa destructor parasitism reduces hemocyte concentrations and prophenol oxidase gene expression in bees from two populations. Parasitol. Res. 2018, 117, 1175–1183. [Google Scholar] [CrossRef]
- Morfin, N.; Anguiano-Baez, R.; Guzman-Novoa, E. Honey Bee (Apis mellifera) Immunity. Vet. Clin. N. Am. Food Anim. 2021, 37, 521–533. [Google Scholar] [CrossRef]
- Chen, Y.; Pettis, J.S.; Evans, J.D.; Kramer, M.; Feldlaufer, M.F. Transmission of kashmir bee virus by the ectoparasitic mite Varroa destructor. Apidologie 2004, 35, 441–448. [Google Scholar] [CrossRef]
- Martin, S.J.; Ball, B.V.; Carreck, N.L. Prevalence and persistence of Deformed wing virus (DWV) in untreated or acaricide-treated Varroa destructor infested honey bee (Apis mellifera) colonies. J. Apic. Res. 2010, 49, 72–79. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Childers, A.K.; Chen, Y.; Madella, S.; Nessa, A.; vanEngelsdorp, D.; Evans, J.D. Recent spread of Varroa destructor virus-1, a honey bee pathogen, in the United States. Sci. Rep. 2017, 7, 17447. [Google Scholar] [CrossRef]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumanna, P. Dead or alive: Deformed wing virus and Varroa destructor reduce the life span of winter honeybees. Appl. Environ. Microbiol. 2012, 78, 981–987. [Google Scholar] [CrossRef]
- Arechavaleta-Velasco, M.E.; Guzman-Novoa, E. Producción de miel de colonias de abejas (Apis mellifera L.) tratadas y no tratadas con fluvalinato contra Varroa jacobsoni Oudemans en Valle de Bravo, Estado de México. Vet. Méx. 2000, 31, 379–382. [Google Scholar]
- Medina-Flores, C.A.; Guzmán-Novoa, E.; Aréchiga-Flores, C.; Aguilera-Soto, J.; Gutiérrez-Piña, F. Efecto del nivel de infestación de Varroa destructor sobre la producción de miel de colonias de Apis mellifera en el altiplano semiárido de México. Rev. Mex. Cienc. Pecu. 2011, 2, 313–317. [Google Scholar]
- Emsen, B.; Guzman-Novoa, E.; Kelly, P.G. Honey production of honey bee (Hymenoptera: Apidae) colonies with high and low Varroa destructor (Acari: Varroidae) infestation rates in eastern Canada. Can. Entomol. 2014, 146, 236–240. [Google Scholar] [CrossRef]
- Chauzat, M.-P.; Martel, A.-C.; Zeggane, S.; Drajnudel, P.; Schurr, F.; Clément, M.-C.; Ribière-Chabert, M.; Aubert, M.; Faucon, J.-P. A case control study and a survey on mortalities of honey bee colonies (Apis mellifera) in France during the winter of 2005-6. J. Apic. Res. 2010, 49, 40–51. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Eccles, L.; Calvete, Y.; Mcgowan, J.; Kelly, P.G.; Correa-Benítez, A. Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 2010, 41, 443–450. [Google Scholar] [CrossRef]
- Medina-Flores, C.A.; López-Carlos, M.; Carrillo-Muro, O.; Gray, A. Honey Bee Colony Losses in Mexico’s Semi-Arid High Plateau for the Winters 2016–2017 to 2021–2022. Insects 2023, 14, 453. [Google Scholar] [CrossRef]
- Rodríguez-Dehaibes, S.R.; Otero-Colina, G.; Villanueva-Jiménez, J.A.; Corcuera, P. ; Susceptibility of Varroa destructor (Gamasida: Varroidae) to four pesticides used in three Mexican apicultural regions under two different management systems. Int. J. Acarol. 2011, 37, 441–447. [Google Scholar] [CrossRef]
- Berry, J.A.; Hood, W.M.; Pietravalle, S.; Delaplane, K.S. Field-level sublethal effects of approved bee hive chemicals on Honey Bees (Apis mellifera L). PLoS ONE 2013, 8, e76536. [Google Scholar] [CrossRef] [PubMed]
- Martínez, P.J.F.; Medina, M.L.A. Evaluación de la resistencia del ácaro Varroa destructor al fluvalinato en colonias de abejas (Apis mellifera) en Yucatán, México. Rev. Mex. Cienc. Pecu. 2011, 2, 93–99. [Google Scholar]
- Kamler, M.; Nesvorna, M.; Stara, J.; Erban, T.; Hubert, J. Comparison of tau-fluvalinate, acrinathrin, and amitraz effects on susceptible and resistant populations of Varroa destructor in a vial test. Exp. Appl. Acarol. 2016, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gashout, H.A.; Goodwin, P.H.; Guzman-Novoa, E. Lethality of synthetic and natural acaricides to worker honey bees (Apis mellifera) and their impact in the expression of health and detoxification-related genes. Environ. Sci. Pollut. Res. 2018, 25, 34730–34739. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Hood, W.M. Effects of delayed acaricide treatment in honey bee colonies parasitized by Varroa jacobsoni and a late-season treatment threshold for the South-Eastern USA. J. Apic. Res. 1997, 36, 125–132. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Hood, W.M. Economic Threshold for Varroa jacobsoni Oud. in the Southeastern USA. Apidologie 1999, 30, 383–395. [Google Scholar] [CrossRef]
- Delaplane, K.S. Varroa control: timing is everything. Am. Bee J 1998, 138, 575–576. [Google Scholar]
- Caron, D. Delaware bee mites survey. Am. Bee J. 1999, 139, 631–633. [Google Scholar]
- González-Cabrera, J.; Rodríguez-Vargas, S.; Davies, T. G E.; Field, L.M.; Schmehl, D.; Ellis, J.D.; Krieger, K.; Williamson, M.S. Novel Mutations in the Voltage-Gated Sodium Channel of Pyrethroid-Resistant Varroa destructor Populations from the Southeastern USA. PloS ONE 2016, 11, e0155332. [Google Scholar] [CrossRef]
- Gatien, P.; Currie, R.W. Timing of acaracide treatments for control of low-level populations of Varroa destructor (Acari: Varroidae) and implications for colony performance of honey bees. Can. Entomol. 2003, 135, 749–763. [Google Scholar] [CrossRef]
- Kokkinis, M.; Liakos, V. Population dynamics of Varroa destructor in colonies of Apis mellifera macedonica in Greece. J. Apic. Res. 2004, 43, 150–154. [Google Scholar] [CrossRef]
- Calderón, R.A.; van Veen, J.W. Varroa destructor (Mesostigmata: Varroidae) in Costa Rica: population dynamics and its influence on the colony condition of Africanized honey bees (Hymenoptera: Apidae). Rev. Biol. Trop. 2008, 56, 1741–1747. [Google Scholar] [CrossRef] [PubMed]
- Strauss, U.; Pirk, C.W.W.; Crewe, R.M.; Human, H.; Dietemann, V. Impact of Varroa destructor on honeybee (Apis mellifera scutellata) colony development in South Africa. Exp. Appl. Acarol. 2015, 65, 89–106. [Google Scholar] [CrossRef]
- da Silva, L.A.; da Silva, A.D.; Domingos, H.G.T.; Bergamo, G.C.; Message, D.; Gramacho, K.P. Varroa destructor mite population dynamics in africanized honeybee (Apis mellifera) colonies in a semi-arid region. Exp. Appl. Acarol. 2024, 1–11. [Google Scholar] [CrossRef]
- INEGI. Instituto Nacional de Estadística Geografía e Informática. Aspectos geográficos de Zacatecas. Available online: https://www.inegi.org.mx/contenidos/app/areasgeograficas/resumen/resumen_32.pdf (accessed on 8 June 2024).
- Guzman-Novoa, E.; Morfin, N.; Dainat, B.; Williams, G.R.; van der Steen, J.; Correa-Benítez, A.; Delaplane, K.S. Standard methods to estimate strength parameters, flight activity, comb construction, and fitness of Apis mellifera colonies 2.0. J. Apic. Res. 2024. [Google Scholar] [CrossRef]
- De Jong, D.; De Andrea Roma, D.; Gonçalves, L.S. A comparative analysis of shaking solutions for the detection of Varroa jacobsoni on adult honeybees. Apidologie 1982, 13, 297–306. [Google Scholar] [CrossRef]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard methods for varroa research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef]
- SAS. Statistical Analysis Software, version 9.0; SAS Institute: Raleigh, NC, USA, 2002. [Google Scholar]
- Fries, I.; Hansen, H.; Imdorf, A.; Rosenkranz, P. Swarming in honey bees (Apis mellifera) and Varroa destructor population development in Sweden. Apidologie 2003, 34, 389–397. [Google Scholar] [CrossRef]
- Moretto, G.; Goncalves, L.S.; De Jong, D. Relationship between food availability and the reproductive ability of the mite Varroa jacobsoni in africanized bee colonies. Am Bee J. 1997, 137, 67–9. [Google Scholar]
- Branco, M.R.; Kidd, N.A.C.; Pickard, R.S. A comparative evaluation of sampling methods for Varroa destructor (Acari: Varroidae) population estimation. Apidologie 2006, 37, 452–461. [Google Scholar] [CrossRef]
- Moretto, G.; Goncalves, L.S.; De Jong, D. Africanized bees are more efficient at removing Varroa jacobsoni: preliminary report. Am. Bee J. 1991, 131, 434. [Google Scholar]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- SAGARPA (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación). Modificación a la Norma Oficial Mexicana NOM-001-ZOO-1994, Campaña Contra la Varroasis de las Abejas. 2005. Available online: https://www.agricultura.gob.mx/sites/default/files/sagarpa/document/2018/08/07/1274/180919-nom-001-zoo-1994-y-nom-002-zoo-1994.pdf (accessed on 24 July 2024).
- Maya-Martínez, O.; Medina-Flores, C.; Aquino-Pérez, G.; Olmos-Oropeza, G.; López-Carlos, M. Seasonal treatment with amitraz against Varroa destructor and its effects in honey bee colonies of Apis mellifera. Abanico Vet. 2020, 10, e–129. [Google Scholar] [CrossRef]
- Medina, L.M.; Martin, S.J.; Espinosa-Montaño, L.; Ratnieks, F.L. Reproduction of Varroa destructor in worker brood of Africanized honey bees (Apis mellifera). Exp. Appl. Acarol. 2002, 27, 79–88. [Google Scholar] [CrossRef]
- Dynes, T.L.; Berry, J.A.; Delaplane, K.S.; De Roode, J.C.; Brosi, B.J. Assessing virulence of Varroa destructor mites from different honey bee management regimes. Apidologie 2020, 51, 276–289. [Google Scholar] [CrossRef]
- Calis, J.N.M.; Fries, I.; Ryrie, S.C. Population modelling of Varroa jacobsoni Oud. Apidologie 1999, 30, 111–124. [Google Scholar] [CrossRef]
- Frey, E.; Rosenkranz, P. Autumn Invasion Rates of Varroa destructor (Mesostigmata: Varroidae) Into Honey Bee (Hymenoptera: Apidae) Colonies and the Resulting Increase in Mite Populations. J. Econ. Entomol. 2014, 107, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Seeley, T.D.; Smith, M.L. Crowding Honey bee Colonies in Apiaries Can Increase Their Vulnerability to the Deadly Ectoparasite Varroa destructor. Apidologie 2015, 46, 716–727. [Google Scholar] [CrossRef]
- Messan, K.; DeGrandi-Hoffman, G.; Castillo-Chavez, C.; Kang, Y. Migration effects on population dynamics of the honybee-mite interactions. Math. Model. Nat. Phenom. 2017, 12, 84–115. [Google Scholar] [CrossRef]
- de Guzman, L.I.; Rinderer, T.E.; Frake, A.M. Growth of Varroa destructor (acari: Varroidae) populations in Russian honey bee (hymenoptera: Apidae) colonies. Ann. Entomol. Soc. Am. 2007, 100, 187–195. [Google Scholar] [CrossRef]
- Medina-Flores, C.A.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Aréchiga-Flores, C.F.; López-Carlos, M.A. Africanized honey bees (Apis mellifera) have low infestation levels of the mite Varroa destructor in different ecological regions in Mexico. Genet. Mol. Res. 2014, 13, 7282–7293. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Cuellar, A.K.; De la Mora, A.; Contreras-Escareño, F.; Morfin, N.; Tapia-González, J.M.; Macías-Macías, J.O.; Petukhova, T.; Correa-Benítez, A.; Guzman-Novoa, E. Genotype, but not climate, affects the resistance of honey bees (Apis mellifera) to viral infections and to the mite Varroa destructor. Vet. Sci. 2022, 9, 358. [Google Scholar] [CrossRef] [PubMed]
- Morfin, N.; Goodwin, P.H.; Guzman-Novoa, E. Varroa destructor and its impacts on honey bee biology. Front. Bee Sci. 2023, 1, 1272937. [Google Scholar] [CrossRef]
- Guzman-Novoa, E.; Corona, M.; Alburaki, M.; Reynaldi, F.J.; Invernizzi, C.; Fernández de Landa, G.; Maggi, M. Honey bee populations surviving Varroa destructor parasitism in Latin America and their mechanisms of resistance. Front. Ecol. Evol. 2024, 12, 1434490. [Google Scholar] [CrossRef]
Figure 1.
Estimated total population of V. destructor, brood and adult bee infestation (%), and average daily mite fall (± SE) at different times of the year in honey bee colonies (n = 10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
Figure 1.
Estimated total population of V. destructor, brood and adult bee infestation (%), and average daily mite fall (± SE) at different times of the year in honey bee colonies (n = 10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
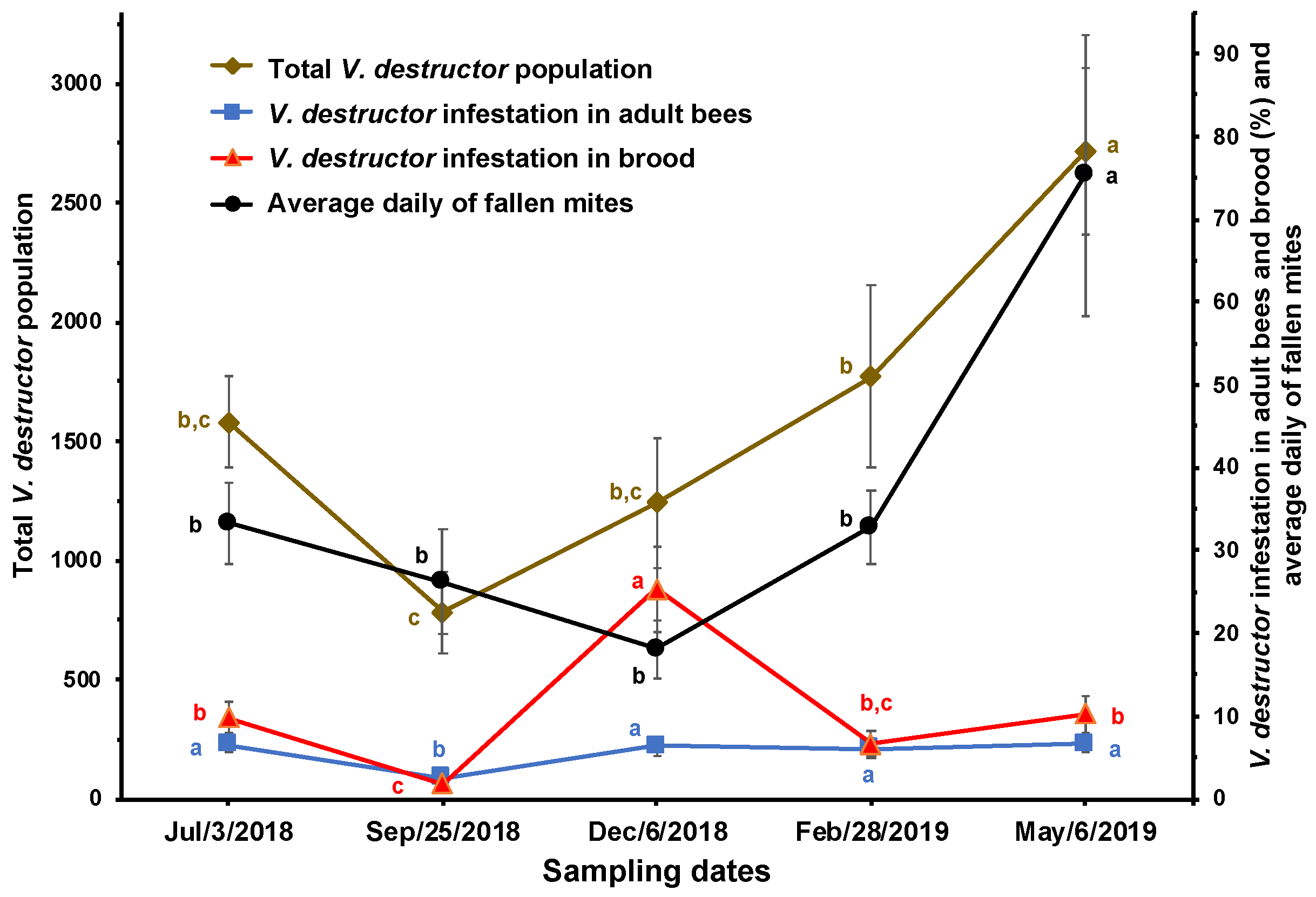
Figure 2.
Estimated population of brood, adult bees, and V. destructor in brood and adult bees (± SE) at different times of the year in honey bee colonies (n = 10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
Figure 2.
Estimated population of brood, adult bees, and V. destructor in brood and adult bees (± SE) at different times of the year in honey bee colonies (n = 10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
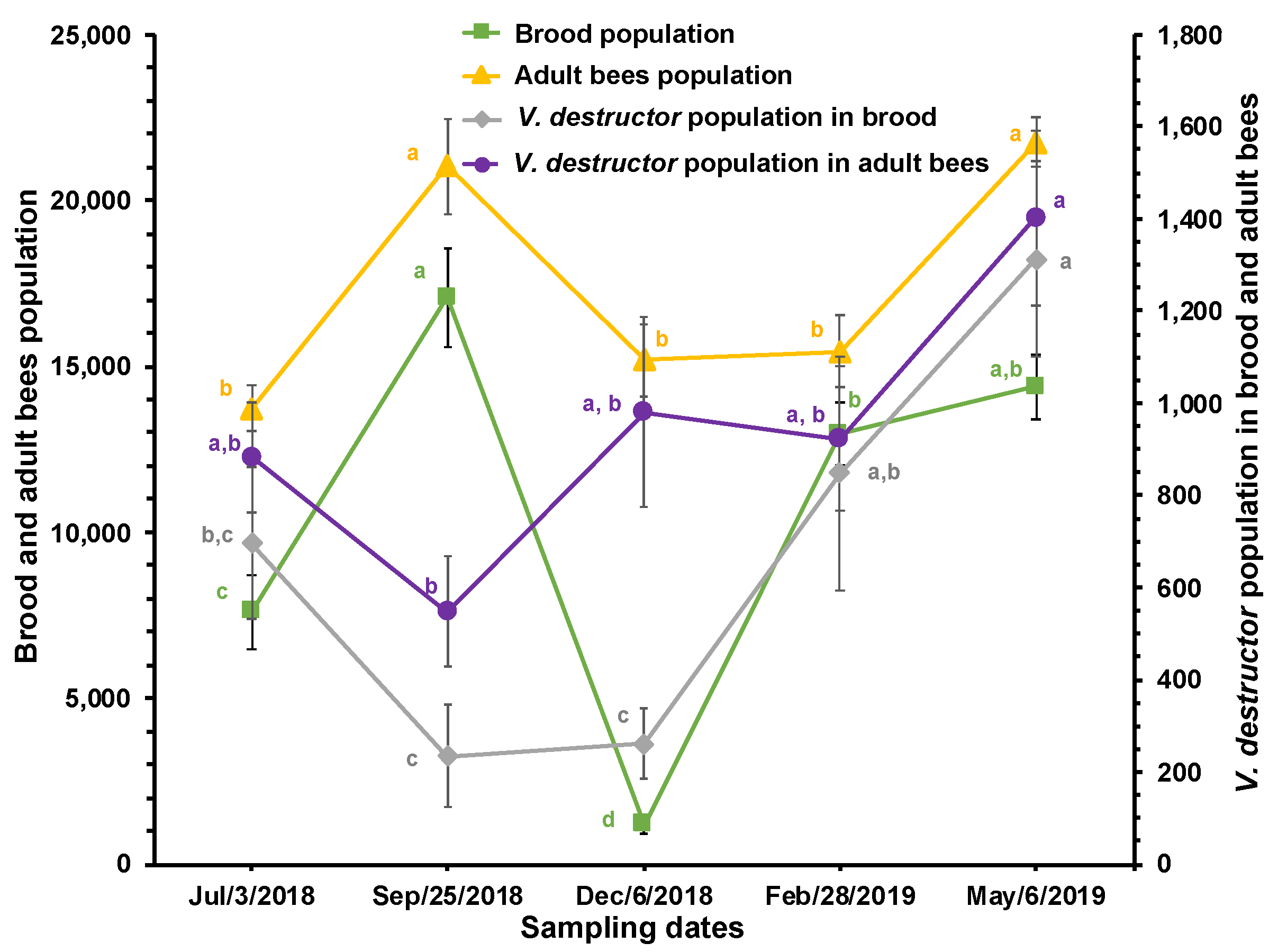
Figure 3.
Estimated brood, adult bees, and total V. destructor populations and average daily mite drop (± SE) at different times of the year in honey bee colonies (n = 10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
Figure 3.
Estimated brood, adult bees, and total V. destructor populations and average daily mite drop (± SE) at different times of the year in honey bee colonies (n = 10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
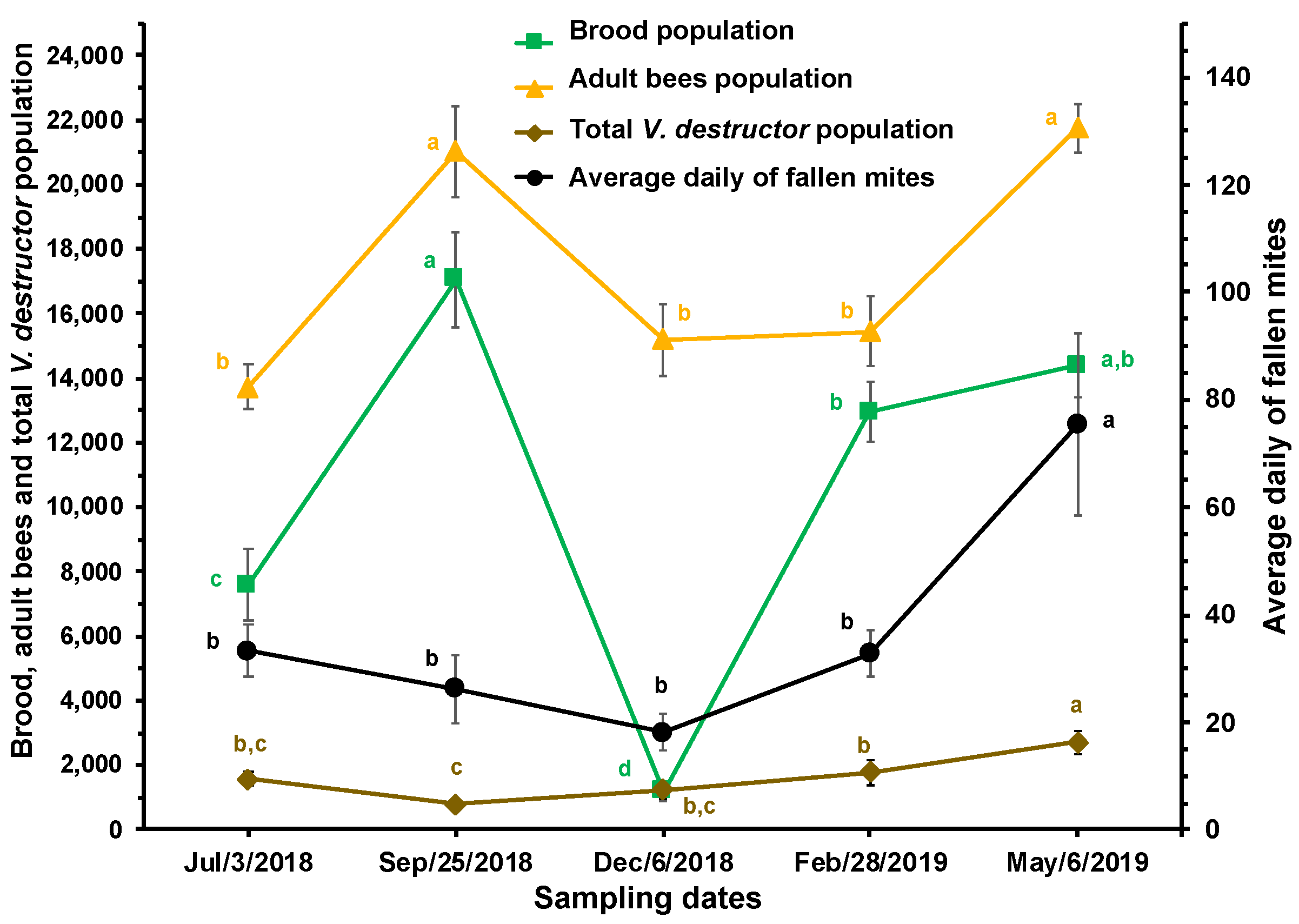
Figure 4.
Honey and pollen areas (cm2) and estimated total population of V. destructor mites (± S.E.) at different times of the year in honey bee colonies (n=10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
Figure 4.
Honey and pollen areas (cm2) and estimated total population of V. destructor mites (± S.E.) at different times of the year in honey bee colonies (n=10) established in a temperate semi-arid climate. Different letters between means for each variable, indicate significant differences based on analyses of variance and Tukey tests.
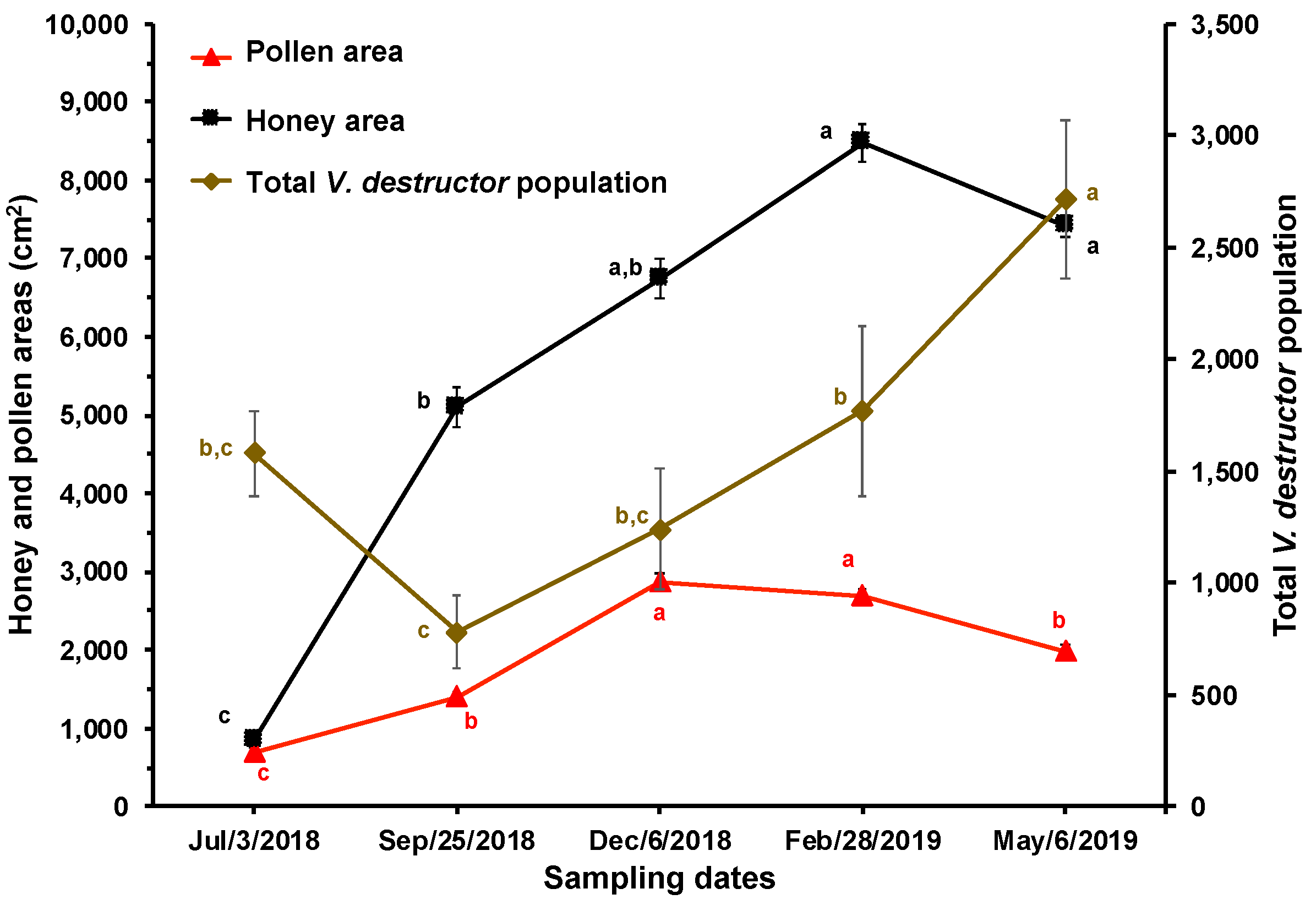

Table 1.
Correlation coefficients between V. destructor infestation rates in brood and adult bees, daily mite drop, total V. destructor population, adult bee population, amount of brood, and areas of honey and pollen in honey bee colonies (n=10) established in a temperate semi-arid climate.
Table 1.
Correlation coefficients between V. destructor infestation rates in brood and adult bees, daily mite drop, total V. destructor population, adult bee population, amount of brood, and areas of honey and pollen in honey bee colonies (n=10) established in a temperate semi-arid climate.
| Variable | Infestation adult | Infestation brood | Fallen of mites | Total mites | Brood amount | Adult population | Pollen area |
|---|---|---|---|---|---|---|---|
| Infestation brood | 0.48*** | ||||||
| Fallen mites | 0.43** | 0.12ns | |||||
| Total mites | 0.73*** | 0.40** | 0.62*** | ||||
| Brood amount | -0.30* | -0.65*** | 0.13ns | 0.04ns | |||
| Adult population | -0.23ns | -0.18ns | 0.17ns | 0.15ns | 0.59*** | ||
| Pollen area | 0.20ns | 0.19ns | 0.18ns | 0.10ns | -0.12ns | -0.02ns | |
| Honey area | 0.17ns | 0.18ns | 0.10ns | 0.30ns | 0.15ns | 0.21ns | 0.45*** |
***, P < 0.001; **, P < 0.01; *, P < 0.05; ns = Not significant. The correlation coefficients were obtained with Pearson's test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



