
Submitted:
21 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Six solvent fractions isolated from flower bud and leaf ethanolic extracts of Cleistocalyx operculatus were analyzed for their phytochemical contents including phenolics, flavonoids, saponins, tannins, and alkaloids. Antioxidant activities were measured using the ABTS, DPPH, and FRAP assays. The results showed that flower bud aqueous fraction (BAF) and leaf aqueous fraction (LAF) rich in phenolic content (768.18 and 490.74 mg GAE/g dry extract, respectively) exhibited significantly higher antioxidant activities than other fractions. Flower bud hexane fraction (BHF) had remarkably high flavonoid and saponin contents (134.77 mg QE/g and 153.33 mg OA/g dry extract, respectively), followed by that of leaf hexane fraction (LHF) (76.54 mg QE/g and 88.25 mg OA/g dry extract, respectively). BHF and LHF were found to have extremely high antibacterial activity against two H. pylori strains ATCC 51932 and 43504 (MICs of 125 µg/ml). Interestingly, DMC (2’,4’-Dihydroxy-6’-methoxy-3’,5’-dimethylchalcone) isolated from BHF displayed greater antibacterial activity against the bacterial strains (MICs of 25–50 µg/ml) than those of the fractions. In addition, DMC presented potent inhibitory effects on H. pylori-urease (IC50 of 3.2 µg/ml) and α-amylase (IC50 of 83.80 µg/ml), but no inhibition against α-glucosidase. It was also demonstrated that DMC showed its pronounced inhibitory effects on urease activity and biofilm formation of H. pylori, and could increase the membrane permeability of the bacterial cells. Scanning electron micrographs depict that BHF and DMC had strong effects on the cell shape and significantly induced distortion and damage of the cell membrane. The fractions and DMC showed no significant toxicity to four tested human cell lines. Efforts to reduce antibiotic use indicate the need for further researches of the flower buds and DMC as potential products to prevent or treat gastric H. pylori infections.

Keywords:
Cleistocalyx operculatus
; antioxidant effect
; anti-H. pylori activity
; antibiofilm formation
; membrane permeability
; morphological transformation
1. Introduction
Flower buds and leaves of Cleistocalyx operculatus (Roxb.) Merr. and L.M. Perry (or Syzygium nervosum DC.) are rich sources of bioactive compounds, predominately containing flavonoids, chalcones, and triterpenoids [1,2]. Several solvent extracts of the flower buds have been reported to exert various pharmacological activities in vitro and in vivo, including anti-hyperglycemic and cardio tonic effects [3,4,5,6]. DMC (2’,4’-Dihydroxy-6’-methoxy-3’,5’-dimethylchalcone), a major constituent of the flower buds, was found to inhibit significantly the growth of human liver cancer cells and human umbilical vein endothelial cells [2,7]. Methanolic extract of the leaves was shown to have inhibitory activity against Gram-positive bacteria (e.g., Staphylococcus aureus, Bacillus subtilis and Streptococcus mutans) and the yeast Candida maltosa, but no effect on Gram-negative bacteria (e.g., Escherichia coli and Pseudomonas aeruginosa) [4]. However, a previous study indicated that crude ethanolic extract of the leaves showed high antibacterial activity against Helicobacter pylori ATCC 51932 and three clinical isolates of H. pylori [8]. Recent research has reported that crude hexane extract from flower buds of C. operculatus displayed the most growth-inhibitory activity against H. pylori and Salmonella Typhimurium, while crude ethanol and methanol extracts of the flower buds exhibited the strongest antioxidant activities [9]. Furthermore, the hexane extract was found to have the strongest inhibitory effect on H. pylori urease activity [9].
H. pylori is a Gram-negative bacterium with curved or spiral shape that infects and colonizes the human gastric mucosa [10]. The bacterial infections are prevalent in over half of the world’s population, particularly in developing regions where the infections can affect as much as 90% of the population and tend to endure for a person’s lifetime [11]. Despite being asymptomatic in most infected individuals, H. pylori has been identified as the highly causative agent of chronic active gastritis, peptic ulcer disease, gastric adenocarcinoma, and mucosa-associated lymphoid tissue (MALT) lymphoma [12,13]. Several important factors, including vacuolating cytotoxin A (VacA), cytotoxin-associated gene A (CagA), chemotactic motility, adhesins, biofilm formation and urease production of H. pylori, are known to contribute to the virulence of this organism [14,15,16]. Its spiral shape is the predominant form involved in the ability to thrive and colonize the gastric epithelial cells. Under unfavorable conditions, the spiral forms can convert into coccoid forms as a survival mechanism, but the coccoid are much less infective and virulent, and less likely to colonize and cause inflammation [16]. In addition, biofilm formation of H. pylori provides the bacterium with protection and resistance to antimicrobial agents [17]. Nowadays, global antibiotic resistance in H. pylori is on the rise in many parts of the world leading treatment failures and reinfections [10].
This study aims to investigate phytochemical contents in six solvent fractions of the C. operculatus flower buds and leaves, and evaluate their antioxidant, enzyme inhibitory, and anti-H. pylori activities. Effects of the fractions and DMC on biofilm formation, membrane permeability and cell morphology of H. pylori were also examined.
2. Materials and Methods
Chemicals, reagents, and enzymes used in bioactivity studies were purchased from Sigma-Aldrich (Schnelldorf, Germany) and Merck (Damstadt, Germany). Two strains of Helicobacter pylori (ATCC 51932) and (ATCC 43504), and MCF-7 cells (HTB-22, human breast cancer cell line), HeLa cells (CCL-2, cervical carcinoma cell line), and Jurkat cells (TIB-152, blood cancer cell line) were provided by ATCC (The American Type Culture Collection, Manassas, Rockville), and fibroblast cells derived from human foreskins were provided by the Laboratory of Molecular Biology, Department of Genetics, VNUHCM-University of Science [18]. Media and serum were provided by HIMEDIA (India), Becton, Dickinson and Company, Sparks, Gibco (New Zealand), and Sigma-Aldrich. All of the solvents and other chemicals were reagent grade and commercially available.
2.1. General Experimental Procedures
An Agilent 1260 infinity series HPLC system (San Jose, CA, USA) was performed using a ZORBAX Eclipse Plus C18 column (4.6 x 150 mm, 3.5 μm) maintained at 25°C with UV detection at 320 nm. The mobile phase compositions were acetonitrile and formic acid 0.1% (70:30, v/v) with a flow rate of 1 ml/min. The injection volume was 10 μl running time for 75 minutes. 1H and 13C NMR (Nuclear Magnetic Resonance) spectra were obtained in CDCl3 at 25°C using a 500MHz Bruker AVANCE III HD spectrometer (Germany). The fourier transform infrared spectroscopy (FT-IR) spectrum was recorded by Bruker Tensor 27 FT-IR Spectrometer (Bruker Optik GmbH, Ettlingen, Germany). Silica Gel 60 (0.06-0.2 mm) (Scharlau, Spain) were used in a silica gel column (∅2 × 30 cm).
2.2. Extraction and Isolation of Plant Material
Flower bud and leaf ethanolic crude extracts of Cleistocalyx operculatus were previously obtained and described by Thanh et al. (2024). The ethanolic extracts of flower buds (90 g) and leaves (81 g) were partitioned by liquid-liquid fractionation to yield 1.8 g of flower bud hexane fraction (BHF), 78 g of flower bud ethyl acetate fraction (BEF), and 10.2 g of flower bud aqueous fraction (BAF) and 26.7 g of leaf hexane fraction (LHF), 24.3 g of leaf ethyl acetate fraction (LEF) and 30 g of leaf aqueous fraction (LAF), respectively, presented as supplementary material (Figure S1).
Guided by anti-H. pylori tests, BHF (1.8 g), one of the most active fractions, was subjected to silica gel column chromatography eluted with a gradient of n-hexane (H) and ethyl acetate (EA) (100:0–0:100) to obtain six column fractions, fraction 1 (34.7 mg), fraction 2–3 (17.6 mg), and fraction 4–6 (56.4 mg) described previously by (7) (Figure S1). Repetitive column chromatography of the fraction 2–3 (17.6 mg) eluted with H:AE = 90:10 (Figure S1) resulted in an active principle 1 (14.2 mg) as a yellow needle-shaped crystals.
The purity of principle 1 was found to be greater than 95 % by HPLC analysis (Figure S2) and its spectroscopic data matched the data reported in previous research of Choommongkol et al. (2022) [19]. The principle 1, DMC (2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone) was identified on the following evidence: FT-IR ν (KBr): the stretching vibrations of the O-H (3421 cm-1), νAr-H and ν=C–H (3001–3028 cm-1), the stretching vibrations of the C–H (2855–2925 cm-1), the C=O (1626 cm-1), the CAr=CAr (1451–1540 cm-1), the deformation vibrations of the C-H (1359–1419 cm-1), the stretching vibrations of the C-O (1111–1220 cm-1) and the out of plane deformation vibrations of the CAr-H and =C–H (612–987 cm-1) (Figure S3). The data of 1H NMR, 13C NMR, and HMBC of DMC were shown in Table S1, Figure S4 and S5. The spectral data were identical to the published data of Choommongkol et al. (2022) [19]. Additionally, 1H - 13C HMBC-correlations of DMC were presented in Figure S6.
2.3. Total Phytochemical Contents
Total phenolic content (TPC) was quantified using the Folin-Ciocalteu assay indicated by Temesgen et al. (2022) [20]. An aliquot of each fraction (0.9 ml) was shaken vigorously with 4.5 ml of Folin-Ciocalteu reagent (10%) and incubated for 5 min in the dark. Then 1.8 ml of Na2CO3 (7%) was added into the mixture and continued to incubate for 30 min. Absolute ethanol was used as a control (blank), and acid gallic (0–100 µg/ml) was also prepared to build a standard curve (y = 0.0095x + 0.0029, R² = 0.9978). Absorbance at 765 nm was subsequently measured using a spectrophotometer (UV-5100, Metash, Shanghai, China). The TPC was expressed as the mg gallic acid equivalent (GAE)/g of dry fraction.
Total flavonoid content (TFC) was determined following the aluminum chloride colorimetric method [21]. An aliquot of each fraction (300 µL) was mixed with 150 µL of NaNO2 solution (5%) and incubated for 6 min at room temperature. The mixture was then added with 300 µL of AlCl3 (5%) and 1 ml of NaOH 1 M and stand for 6 min. Absolute ethanol was prepared as a control (blank) to replace the extract. Quercetin (0–100 µg/ml) was used to estimate the standard curve (y = 0.0075x - 0.0017, R² = 0.9998). Absorbance of the mixture was measured at 510 nm. TFC was expressed as mg of quercetin equivalent (QE)/g of dry fraction.
Total alkaloid content (TAC) was quantified using the modified method of Ncube et al. (2015) [22]. Briefly, 1 ml of each fraction diluted in HCl (2N) was transferred to a separating funnel containing 5 ml of bromocresol green (BCG) solution (0.01%) and 5 ml phosphate buffer solution at pH 4.7. The mixture was shaken vigorous and extracted twice with 5 ml of chloroform. The alkaloid extracts were collected and diluted in a 10 ml volumetric flask with chloroform. The absorbance of the mixture was measured at 470 nm. Atropine (0–100 µg/ml) was used as standard solution (y = 0.0128x - 0.0493, R² = 0.9927) and absolute ethanol was control. The total content of alkaloids was shown as mg atropine equivalent (AE)/g of dry fraction.
Total tannin content (TTC) was quantified using a previously reported method [21]. An aliquot of each fraction (0.5 ml) was added into 3.0 ml of vanillin solution (4% in methanol, w/v). Then, the mixture was mixed with 1.5 ml of HCl and incubated for 15 min in the dark. Absorbance of the mixture was read at 500 nm, and absolute ethanol was used as control (blank) to replace the extract. The TTC was calculated using catechin standard curve (y = 0.0012x + 0.0261, R² = 0.9984) and expressed as mg of catechin equivalent (CE)/g of dry fraction.
Total saponins content (TSC) was determined using the vanillin method [23]. Briefly, each fraction (0.5 ml) was mixed with 0.1 ml of vanillin solution (5% in acetic acid, w/v) and 0.4 ml of perchloric acid 70%. The mixture was heated at 60 °C for 15 min in water bath and then cooling it down to room temperature. After that, 5 ml of absolute acetic acid was mixed well into the mixture. Oleanolic acid (0–300 µg/ml) was used to measure the standard curve (y = 0.0041x - 0.0256, R² = 0.9934). The absorbance values were measured at 548 nm. The TSC was expressed as mg oleanolic acid equivalent (OAE)/g of dry fraction.
2.4. Antioxidant Assay
DPPH (2,2-diphenyl-1-(2,4,6-trinitrophenyl)hydrazyl) radical scavenging assay was followed the previous study as described by Elouafy et al. (2023) [24] to assess antioxidant activity. Briefly, different concentrations of each fraction (0–150 μg/ml) were prepared in methanol. Each dilution (1 ml) was mixed well with 1 ml of DPPH methanolic solution (25 µg/ml) and incubated at 37 °C in the dark condition for 30 min. Absorbance of the mixture was subsequently measured at 517 nm. Absolute methanol and ascorbic acid were used as blank and positive control, respectively. The DPPH scavenging activity was calculated using Equation (1).
where OD1 and OD2 are the absorbance values of the blank and each fraction or ascorbic acid, respectively. The concentration required to inhibit 50% of the free DPPH radial (IC50) was calculated by plotting the DPPH scavenging activity versus the sample concentration.
The measurement of antioxidant activity was also performed using the ABTS•+ (2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt) assay presented by Olszowy-Tomczyk and Typek (2024) [25] with some modifications. Briefly, the initial solutions consisted of 7 mM ABTS•+ solution (5 ml) and 140 mM K2S2O8 solution (88 μl) to 2.45 mM final concentration. The working solution was reacted in the dark at room temperature for 16 hours and then diluted in ethanol until the absorbance value of 0.7 ± 0.002 at 734 nm using a spectrophotometer. Fresh ABTS•+ solution was created for each assay. For the analysis, each fraction (0.01 ml) was mixed with 0.99 ml of the ABTS•+ solution. After 6 min storage in the dark, the absorbance was measured at 734 nm. Absolute methanol and trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) were used as negative control and positive control, respectively. The ABTS•+ scavenging capacity was evaluated as the percentage inhibition of ABTS radical scavenging activity, using Equation (2).
where OD1 and OD2 was absorbance of the ABTS•+ radical cation in methanol and in each fraction or Trolox, respectively. The concentration required to inhibit 50% ABTS•+ solution (IC50) was subsequently estimated by plotting the ABTS radical scavenging activity versus the sample concentration.
The FRAP assay was determined using a previously reported method [21] with some modifications. Briefly, the FRAP reagent was prepared by mixing 300 mM sodium acetate buffer pH 3.6 (25 ml), 10 mM TPTZ (2, 4, 6- tripyridyl-s-triazine) solution in 40 mM HCl (2.5 ml), and 20 mM FeCl3.6H2O solution (2.5 ml) and then warmed at 37°C before using. To initiate the reaction, 150 μl of each fraction was mixed with 2850 μl of the FRAP reagent and then incubated at 37°C for 30 min in the dark condition. Standard curve of the assay was also prepared by using a serial concentration of Trolox (0–500 µg/ml). The absorbance of the colored product was read at 593 nm. The antioxidant capacity was expressed as mg Trolox equivalent (TE)/g of dry fraction.
2.5. Anti-Helicobacter pylori Assay
Two H. pylori strains were stored in BHI (brain heart infusion) broth medium supplemented with 10% NBS (newborn bovine serum), which contained 10 mg/l vancomycin, 5 mg/l polymyxin B, 5 mg/l trimethoprim, and 2 mg/l amphotericin B, and 25% glycerol in a liquid nitrogen container until use. The minimal inhibitory concentrations (MICs) of the fractions and DMC were determined using a broth dilution assay in sterile 96-well plates for the bacterial strains [26]. Different concentrations (1 to 1000 μg/ml) of tested samples were diluted in DMSO and then performed in 75 μl Brucella broth supplemented with 10% NBS to get the final concentration of DMSO in all assay less than 2.5%. Generally, 30 μl of bacterial suspension (5 × 106 CFU/ml) of each strain was added to 10 μl tested samples. Then, the plates were incubated at 37oC and shaken at 50 rpm for 48 h in a microaerophilic jar using Oxoid CampyGen gas packs (Thermo Scientific, UK). The lowest concentration that visibly inhibited bacterial growth using resazurin as an indicator was defined as MIC value.
2.6. Enzyme Inhibitory Assay
Crude urease of H. pylori ATCC 43504 was prepared according to the method of Ngan et al. (2012) [27]. Briefly, 10 μl each fraction or DMC at various concentrations of 0–500 μg/ml were added in 30 μl EDTA-sodium phosphate buffer 20 mM (pH 7.3). Then 10 μl urease solution (75 μl urease/1 ml of the buffer) was added and incubated at room temperature for 1 h before adding 50 μl urea solution (0.24 mg urea/1 ml of the buffer) and continue to incubate at room temperature for 30 min. After that, 40 μl of solution A (40% sodium salicylate and 0.3% sodium nitroprusside) and 60 μl solution B (0.5% sodium hydroxide and 0.042% sodium hypochlorite) were added into the mixture. The ammonia released by the urease was quantified by measuring absorbance on a Microlisa Plus microplate reader (Micro Lab Instruments, India) at 625 nm with ammonium chloride as a standard and buffer solution as control.
The α-glucosidase activity was determined by measuring the release of p-nitrophenol from pNPG (p-nitrophenyl-α-D-glucopyranoside) according to the method of Shai et al. (2011) [28] with slightly modification. Each fraction and DMC were dissolved in DMSO (dimethyl sulfoxide) to a serial concentration (0–500 μg/ml). A mixture containing 100 µl of potassium phosphate buffer (100 mM, pH = 6.8), 20 µl of α-glucosidase (2.0 U/ml) and 40 µl of tested samples was prepared and pre-incubated at 37 °C for 15 min. The reaction was initiated by adding 40 µl of pNPG 5 mM and incubated at 37 °C for 20 min. To stop the reaction, 100 µl Na2CO3 0.1 M was added to the solution. The amount of p-nitrophenol released was determined by measuring absorbance at 405 nm. Acarbose was used as standard inhibitor. The control solution contained buffer solution (40 μl) instead of the fractions, DMC or standard inhibitor.
The α-amylase inhibitory activity of the fractions and DMC were caried out according to the method of Ogunyemi et al. (2022) [29] with slightly modifications. A mixture of 100 μl of various concentrations (0–500 μg/ml in DMSO) of each fraction or DMC and 100 μl of sodium phosphate buffer (0.02 M, pH 6.9) containing 200 μl of α-amylase solution (2.5 U/ml) were incubated at room temperature for 10 min. After preincubation, 200 μl soluble starch solution 1% in sodium phosphate buffer were added to the mixture. The reaction mixtures were then incubated at 30°C for 10 min followed by addition of 300 μl of HCl 1 N to stop the reaction. The mixture was added 90 μl of iodine solution 10% to observe the color change and diluted with 10 ml of distilled water. The sodium phosphate buffer and acarbose were used as control and standard inhibitor, respectively. The absorbance was read at 540 nm.
These enzyme inhibitory activities were determined using Equation (3).
where Ac was the absorbance of the control and As was the absorbance of the fractions, DMC or standard inhibitors.
2.7. Biofilm Formation Inhibitory Assay
The biofilm inhibition of H. pylori ATCC 43504 by the fractions and DMC was carried out in 96-well plates as reported previously [30]. In brief, 30 μl the bacterial suspension (108 CFU/ml) and 10 μl the tested samples at sub-MICs (MIC/2, MIC/4 and MIC/8) were added into 60 μl of Brucella broth in each well of sterile 96-well plates. Blank wells containing DMSO and background wells containing samples was also similarly arranged as control wells without the bacterial suspension. After 48h incubation at 37°C in microaerobic environment with shaking at 150 rpm, the medium was removed and the wells washed with PBS (phosphate buffer saline 0.01 M, pH 7.2). The plates were then air dried before fixing with absolute methanol for 15 min and dried against. Each well was dyed with a 0.1% crystal violet solution for 10 min and washed with distilled water. The dried plates were dissolved crystal violet using 95% ethanol for 15 min and measured the absorbance at 595 nm using Microlisa Plus microplate reader.
2.8. Scanning Electron Microscopy
To determine the efficacy of BHF and DMC against H. pylori through the morphological changes, scanning electron microscopy (SEM) analysis was performed [31]. Overnight broth cultures of H. pylori ATCC43504 were prepared in Brucella broth. The cell suspension of H. pylori was cultured after 48 h with or without tested samples at MIC concentrations. Then the bacteria were harvested by centrifugation at 14000 rpm for 5 min. In brief, the specimens were fixed in modified Karnovsky’s fixative (2% glutaraldehyde and 2% paraformaldehyde in 0.05 M sodium cacodylate buffer, pH 7.2) for 4 h and post-fixed in 1% osmium tetroxide in 0.05 M sodium cacodylate buffer for 4h at 4 oC. The specimens were dehydrated through a series of increasing concentrations of ethanol (20, 40, 60, 80, 95% and absolute), then treated twice with hexamethyldisilazane, each for 15 min, and dried at room temperature overnight. SEM was performed by Institute of Chemical Technology, Ho Chi Minh city, Vietnam. The specimens were then mounted on SEM stubs by double-sided carbon conductive tape and coated with gold (JEC-3000FC ion sputter, JEOL Co., Japan). The SEM images were obtained using a scanning electron microscope JSM-IT200 (JEOL Co., Japan) operating at acceleration voltage of 5 kV.
2.9. Membrane Permeability Assay
The alteration in membrane permeability of H. pylori was detected by crystal violet assay [32]. Shortly, 30 μl suspensions of H. pylori ATCC 43504 (108 CFU/ml) were prepared in 60 μl of BB medium and 10 μl of each fraction and DMC at different concentrations (MIC, MIC/2, MIC/4 and MIC/8). The mixture was incubated in 2 h and the cells were harvested at 4500 rpm for 5 min at 4°C. The cells were washed twice in PBS (0.01 M, pH 7.2). After that the cells were resuspended in 1 ml PBS containing 100 μl crystal violet 0.1 % and incubated for 5 min at 37 °C. The suspension was then centrifuged at 14000 rpm for 15 min and the OD590 of the supernatant was measured in Microlisa Plus microplate reader. The percentage of crystal violet uptake of the H. pylori cells in all the tested samples was calculated using Equation (4).
2.10. Cell line Cultures and Cytotoxicity Assay
MCF-7, HeLa, Jurkat, and fibroblast cell lines were grown in EMEM (Eagle’s minimal essential medium) for MCF-7 and HeLa cells, RPMI (Roswell Park Memorial Institute) medium for Jurkat cells, and DMEM/F12 (Dulbecco’s modified eagle medium: nutrient mixture F12) medium for fibroblast cells. These media were supplemented with 10% FBS (fetal bovin serum), 2 mM L-glutamine, 20 mM HEPES, 0.025 μg/ml amphotericin B, 100 IU/ml penicillin G, and 100 μg/ml streptomycin, at 37°C and 5% CO2. Fibroblast cells used were between passages 2 and 5, and HeLa, MCF7, and Jurkat cells used were between passages 4 and 20.
The SRB (Sulforhodamine B) assay was performed as previously done by Nguyen and Huynh (2016) [18]. In brief, cells were seeded in 96-well plates at a density of 10,000 cells/well for MCF-7, HeLa, and fibroblast, and 50,000 cells/well for Jurkat cells. These cells were then cultured for 24 h before exposure to varying concentrations of each fraction or DMC for a duration of 48 h. Treated cells were fixed with a cold 50 % (w/v) trichloroacetic acid solution for 1–3 h, then washed and stained with 0.2 % (w/v) SRB for 20 min. After five subsequent washes with 1 % acetic acid, protein-bound dye was solubilized in a 10 mM Tris base solution. Optical density values were determined using Microlisa Plus microplate reader at wavelengths of 492 nm and 620 nm. Camptothecin served as a positive control and DMSO 0.25% as negative control. The percentage of growth inhibition was determined using Equation (5).
In which ODt and ODc are the optical density value of the tested sample and the control sample, respectively.
2.11. Statistical Analysis
MIC values of each tested extract and amoxicillin were obtained from at least three independent experiments performed in triplicate (n ≥ 9). Tested materials with MIC values of ≤ 130, > 130–< 630, 630–1250, > 1250–< 2500, and ≥ 2500 µg/ml was classified as the extremely high, high, moderate, low, and no inhibitory activity against the growth of tested bacteria, respectively [33]. All other experiments were performed in triplicate and data were shown as mean ± standard derivations (SD) (n ≥ 3). Analysis of variance (ANOVA) by Tukey’s multiple comparison tested as P < 0.05 and half maximal inhibitory concentration value (IC50) and half cytotoxicity concentration (CC50) values were conducted using GraphPad Prism 8 software program (San Diego, CA).
3. Results
3.1. Total Phytochemical Contents
The total phytochemical contents of different fractions from flower buds and leaves of C. operculatus were shown in Table 1. BAF and LAF revealed the highest values of TPC with 768.18 and 490.74 mg GAE/g dry extract, respectively. The lowest TPC was found in LHF (201.63 mg GAE/g dry extract). In contrast, the BHF showed the highest TFC and TSC (134.77 mg QE/g dry extract and 153.33 mg OA/g dry extract, respectively) and following with BEF (85.88 mg QE/g dry extract and 158.10 mg OA/g dry extract, respectively), and LHF and LEF (71.72–76.54 mg QE/g dry extract and 75.24–88.25 mg OA/g dry extract, respectively). LAF contained the lowest levels of both TFC and TSC (25.56 mg QE/g dry extract and 25.56 mg OA/g dry extract,) while BAF had the lowest level of TFC (11.04 mg QE/g dry extract). In the experiment, the flower bud and leaf fractions of C. operculatus presented relatively low values of TTC (4.36–42,97 mg CE/g dry extract) and significantly low values of TAC (1.50–5.41 mg AE/g dry extract).
3.2. Antioxidant Activity
The antioxidant activities of different fractions from flower buds and leaves of C. operculatus were exhibited in Table 2. Depending on the solvent used for extracting fractions, the antioxidant activity of the fractions was significantly varied. In the FRAP assay, LAF exhibited the highest antioxidant activity (301.82 mg TE/g dry extract), followed with BAF (201.80 mg TE/g extract). The lowest FRAP activity was obtained with LHF (55.57 mg TE/g extract) and BHF (69.01 mg TE/g extract). The result of DPPH and ABTS radical scavenging abilities indicated that LAF exhibited the strongest antioxidant activity (IC50 values of 11.24 and 0.55 μg/ml, respectively) followed by LEF (16.05 and 0.98 μg/ml) and BAF (24.69 and 1.08 μg/ml). However, result from ABTS method showed that LEF, LHF and BAF gave similar ABTS radical scavenging activity (0.98–1,08 μg/ml). Weak DPPH radical scavenging activity was found in the BHF, LHF and BEF (IC50 range of 33.99–58.46 μg/ml), while weak ABTS radical scavenging activity was found in BEF and BHF (1.48–1.70 μg/ml). All the tested fractions produced the lower antioxidant effects than that of ascorbic acid (IC50 of 3.34 μg/ml) but higher than that of the trolox (IC50 of 2.63 μg/ml).
3.3. Anti-H. pylori Activity
The bacterial growth inhibitory activities of the C. operculatus fractions and DMC against two H. pylori strains ATCC 51932 and ATCC 43504 were shown in Table 3. Both the BHF and LHF showed extremely high growth inhibitory activity against the strains ATCC 51932 and ATCC 43504 with MIC value of 125 µg/ml. Other fractions displayed high growth inhibitory effect with MIC values ranged from 250–500 µg/ml on the both bacterial strains, except the LAF with moderate effect (MIC value of 1000 µg/ml) on the strain 43504. Remarkably, DMC revealed stronger antibacterial activity (MIC values of 25 and 50 µg/ml) than those of BHF and LHF. The MIC values of amoxicillin against the both strains of H. pylori ATCC 51932 and ATCC 43504 (0.01 µg/ml) indicated that these strains were susceptible to amoxicillin because MIC resistance breakpoints for amoxicillin against H. pylori was reported to be >0.5 µg/ml [34].
3.4. Enzyme Inhibitory Activity
The inhibitory activities of the C. operculatus fractions and DMC against H. pylori-urease, α-glucosidase and α-amylase were presented in Table 4. All tested fractions and DMC exhibited strong inhibition against H. pylori-urease (IC50 values ranging from 2.3–3.6 μg/ml) except for BEF (IC50 of 4.9 μg/ml) and LAF (IC50 of 6.8 μg/ml). These fractions and DMC showed better inhibitory activity against urease than that of thiourea (IC50 of 44.3 μg/ml) as positive control. These fractions also had strong inhibitory effect on α-glucosidase (IC50 values ranging from 0.6–2.6 μg/ml) and significantly stronger than the positive control, acarbose (IC50 of 25.6 μg/ml) whereas DMC had no inhibition against α-glucosidase (IC50 of 94.6 μg/ml). Nevertheless, these fractions exhibited weak (IC50 values ranging from 191.3–497.2 μg/ml) and no (IC50 of 1281.7 μg/ml) inhibitory activities against α-amylase, compared with acarbose and DMC which had similar inhibitory effect (IC50 values of 75.0 and 83.8 μg/ml, respectively).
3.5. Effect on Biofilm Formation of H. pylori
The results of biofilm inhibition revealed that all tested fractions and DMC at sub-MICs inhibited biofilm formation of H. pylori after 48 of treatments (Figure 1). At MIC/2, all fractions (62.5–500 µg/ml) and DMC (25 µg/ml) demonstrated potential antibiofilm activity and reduced biofilm formation by 82.2±0.74–93.6±1.53% with nonsignificant difference (P>0.05). At MIC/4, LEF (125 µg/ml) and DMC (12.5 µg/ml) induced the pronounced antibiofilm effect with the inhibitory percents of 78.4±0.98 and 77.8±1.37%, respectively, followed by the slightly lower activity of BHF (31.25 µg/ml) and BEF (125 µg/ml), reducing biofilm formation by 73.5±0.98% and 70.2±2.55%, respectively. However, LHF (31.25 µg/ml), BAF (125 µg/ml) and LAF (250 µg/ml) reduced biofilm formation by 50.9±6.86%, 54.3±2.43%, and 57.8±0.88%, respectively. Notably, at MIC/8, BEF and LEF (62.5 µg/ml) remained able to inhibit biofilm formation by 58.2±0.59% and 57.1±0.98%, respectively, followed by DMC (6.25 µg/ml) that inhibited by 46.6%±0.49%. Other fractions had weak inhibitory effect (21.1±0.49–36.4±2.84%).
3.6. Effect on the Morphology of H. pylori
The scanning electron micrographs for the untreated H. pylori showed the cells appeared mainly in a spiral-shaped form successfully incorporated within biofilms (Figure 2A,B). The cell surface was smooth and regular with an intact cell membrane. However, after 48h-treatment with BHF at MIC = 125 µg/ml (Figure 2C,D) and DMC at MIC = 50 µg/ml (Figure 2E,F), 86.2±3.56 and 93.7±3.68 % of the spiral cells were converted into coccoid-shaped cells, respectively. These figures indicated that BHF and DMC could inhibit bacterial biofilm formation and induce clusters of coccoid cells. The clusters were aggregated and stuck to each other with rough or deformed membrane surface and cell debris. This suggested that most of the coccoids induced by BHF and DMC were the morphological appearance of bacterial cell death or destruction.
3.7. Effect on Membrane Permeability
The alteration in membrane permeability of H. pylori caused by C. operculatus fractions and DMC was presented in Figure 3. The crystal violet uptake by untreated cells was only by 13.8±1.04–26.0±0.98% but the crystal violet uptake by treated cells increased from 63.0±0.87 to 71.3±1.46% caused by all tested fractions and 79.8±1.06% caused by DMC at MIC/2 after 2h-treatments. At MIC/4, DMC (12.5 µg/ml) induced a significant increase in the uptake of crystal violet by 64.4±2.76%, followed by those of LHF and BHF (31.25 µg/ml), LEF and BAF (125 µg/ml), and LAF (250 µg/ml) increasing by 49.5±2.43% and 50.3±1.54%, 46.3±4.17% and 54.8±2.49%, and 50.1±1.36%, respectively, and BEF (125 µg/ml) increasing only by 33.82±1.73%. At MIC/8, the membrane permeability of the bacterial cells did not alter much (27.4±1.90–40.4±2.06%) compared with those of the untreated groups.
3.8. Cytotoxicity Effects
In order to investigate the selectivity of the anti-H. pylori activity of C. operculatus fractions and DMC, the cytotoxic effect of these tested samples on four human cell lines was examined in Table 5. All the fractions and DMC showed no significant cytotoxicity toward fibroblast cells with CC50 values of >100 µg/ml. Among these factions, BAF exhibited no cytotoxic activity against the four tested cell lines (CC50 values >100 µg/ml) whereas BHF displayed the most cytotoxic effect on three cancer cell lines, Jurkat, MCF-7, and HeLa cells with CC50 values of 18.51, 30.79, and 31.70 µg/ml, respectively. Other fractions and DMC had no cytotoxicity to the three cancer cell lines (CC50 values ranging from 85.43 to >100 µg/ml) or weak cytotoxicity (CC50 values ranging from 51.06 to 77.07 µg/ml). However, the four tested cell lines were less sensitive to these fractions and DMC when compared with the positive control camptothecin, CC50 of 0.005–1.57 µg/ml (P<0.001).
4. Discussion
Phytochemicals (e.g., polyphenols, flavonoids, alkaloids, tannins and saponins) are known to play an important role in overall health and disease prevention. In the C. operculatus flower bud and leaf fractions, the contents of phytochemicals significantly varied depending on the solvents used. Similarly with other previous studies, high polyphenols content was mostly found in the aqueous extracts of both C. operculatus flower buds and leaves [20,21,35,36,37]. Minh et al. (2023) [38] recently showed that flavonoids content was only present at low level in the aqueous extracts of C. operculatus fresh leaves (2.73 mg QE/g) when compared with that in the hexane and ethyl acetate extracts. The flavonoid contents were present at low level in aqueous extracts of C. operculatus fresh leaves (2.73 mg QE/g) [38] and flower buds (81.1 mg QE/g) [9] when compared with that in the hexane fraction.
In our study, flavonoids have been found to be remarkably rich in the hexane and ethyl acetate fractions. Chalcones belonging to flavonoids family, especially DMC, have been presented as bioactive agents isolated from the non- or low-polar solvent extracts of the flower buds [1,2,7,39,40]. The contents of saponin and tannin were also high in these hexane and ethyl acetate fractions, particularly in the flower buds of C. operculatus. The tannin content in hexane fraction was reported to be higher than that in aqueous extract of C. nervosum pulp [41].
It is well known that the higher polyphenol and flavonoid contents could significantly increase the antioxidant capacity of plant extracts [42]. In the present study, hexane and ethyl acetate fractions exhibited weaker antioxidant capacities than the aqueous counterparts. This may be because the content of flavonoids was present at lower levels than those of other phenolic compounds in the flower buds and leaves of C. operculatus. The aqueous fractions in our study exhibited similar antioxidant capacity to those of aqueous extracts from the flower buds and leaves of C. operculatus reported by Mai et al. (2009) [35]. Comparing to the other studies, most of the C. operculatus fractions performed higher antioxidant capacity than those of Psidium guajava L. [43], Halimium halimifolium [21] and Syzygium aromaticum flower buds [20].
The researches of Mai et al. (2009) [35] and Minh et al. (2023) [38] have shown that there were very little to no alkaloids in C. operculatus. Our study identified alkaloids in all of the tested fractions at significantly lower concentrations than other phytochemicals. Although the antioxidant activity of alkaloids was also demonstrated [4], the presence of alkaloids at low levels in the fractions of the C. operculatus flower buds and leaves could make them contribution secondary when compared with polyphenols and flavonoids.
Among the fractions of C. operculatus flower buds and leaves, the hexane fractions and the isolated compound DMC presented the strongest growth inhibitory activity against H. pylori. This indicated that flavonoids and other phenolic compounds could have a high synergistic activity against the growth of H. pylori. Especially, the single compound DMC, isolated from the flower buds of C. operculatus, was found to have 2.5 to 5 times more potent inhibitory activity than these hexane fractions. Similarly, terpenes (e.g., diterpenes and sesquiterpenes) and phenolic compounds from leaf extract of Caseria sylvestris have been reported to provide a high synergism against H. pylori [44]. Several studies have shown that extracts from the flower buds and leaves possessed antibacterial activity against many types of Gram-negative and -positive bacteria such as Xanthomonas spp. [45], Staphylococcus aureus, S. epidermidis, S. haemolyticus, Bacillus subtilis, and Streptococcus mutans [4], H. pylori [8], and S. pyogenes [46]. In addition, DMC has been also known as a bioactive agent against cellular oxidative stress [2] and cytotoxic effect [19,47]. Crude ethanolic extract from leaves [8] and crude hexane extract from flower buds of C. operculatus [9] had previously reported to have high antibacterial activity against H. pylori. However, to date, there is no data reported on antibacterial activity of DMC isolated from C. operculatus flower buds against the pathogenic bacterium.
The antibacterial activity of C. operculatus fractions against H. pylori was found to be higher than those of other reported medicinal plants such as Cichorium intybus (MICs of 1.25–10 mg/ml), Cinnamomum zeylanicum (MICs of 1.25 to 5 mg/ml), and Foeniculum vulgare (MICs of >10 mg/ml) [48], and H. rosa sinensis (MICs of 0.2–0.25 mg/ml) [26]. It was previously reported that 2’,4’- dihydroxychalcone isolated from leaves of Muntingia calabura provided antibacterial activity against methicillin-susceptible and -resistant Staphylococcus aureus (MICs of 50 and 100 mg/ml, respectively) [49]. Recently, the flavonoids naringenin, myricetin, and luteolin isolated form red flowers of H. rosa sinensis were reported to display effective anti-H. pylori properties (MICs of 100–150 μg/ml) [30]. In our study, DMC (2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone) was found to possess the potent growth inhibitory activity against H. pylori (MICs of 25–50 µg/ml).
H. pylori can produce urease for successful survival in acidic condition and colonization in gastric mucosa of the human stomach. Therefore, inhibition of the bacterial enzyme could prevent the bacterial growth and colonization. The crude hexane extract from flower buds C. operculatus was found to have a strong inhibitory effect on H. pylori-urease activity [9] The C. operculatus fractions and DMC in our present study have also been found to possess potent inhibitory effects on the urease of H. pylori and much stronger than those of Fagonia arabica L. and Casuarina equisetifolia L. reported by Amin et al. (2013) [50]. The flavonoid quercetin, present in acetone extract of Heterotheca inuloides Cass. (Asteraceae), has been known to display a high in vitro inhibition against the enzyme with IC50 = 132.4 μg/ml [51]. Several other flavonoids (such as naringenin, myricetin, and luteolin) and phenolic acid (protocatechuic acid) from red flowers of Hibiscus rosa sinensis were also reported to produce the potent inhibition of H. pylori-urease activity [30].
In addition, inhibition of α-amylase and α-glucosidase activities has been known to be one of the treatments for diabetes since it helps to control glucose levels in the blood. Previously reported by Zhang and Lu (2012) [6], the aqueous extract of C. operculatus flower buds and DMC inhibited α-amylase with IC50 values of 73.10 and 20.67 µg/ml, respectively. Recently, Chukiatsiri et al. (2023) [52] indicated that hexane extract of C. nervosum had no inhibitory activity against both α-amylase and α-glucosidase, but the aqueous extract of this plant revealed depressing effect on both of the enzymes with IC50 values of 0.61 and 0.44 mg/ml, respectively. In the recent study, we found that the C. operculatus fractions had strong inhibitory effect on α-glucosidase, while DMC displayed inhibitory activity against α-amylase.
Moreover, adverse effects on the bacterial biofilm formation, cell morphology and membrane permeability have been also well described as mode of action of plant secondary metabolites on the bacterial survival [53,54]. In the current study, we found that all C. operculatus fractions and DMC inhibited urease with the IC50 values significantly smaller than their MIC values. At sub-MICs, they also exhibited the pronounced antibiofilm activity as the biofilm growth can be reservoirs for the spread of the pathogenic bacterium, persistent infection and resistance to adverse factors [55]. EtOAc fraction of H. rosa-sinensis red flowers at MIC/2 (0.125 mg/ml) has been reported to inhibit biofilm formation of H. pylori by 79.3% and cause considerable transformation of the spiral forms to the coccoid forms (91 vs 18% at 1.5 and 0.75 mg/ml after 48 h of treatment) [26]. The flavanone naringenin was found to display the most antibiofilm activity (85.9 versus 52.7% of inhibition at MIC/2 and MIC/4 = 25 μg/ml) [30]. The flavanone also induced morphological conversion of H. pylori to the coccoid forms (95 versus 16.5% at 1000 and 500 μg/ml) followed by the flavone luteolin (87.5 versus 14.7%) which caused higher conversion to the coccoid forms than the flavonol myricetin (79 versus 15%) [30]. Previously, the methanolic extract of C. operculatus leaves have been known to have the anticaries activity against Streptococcus mutans in terms of inhibition of acid production and biofilm formation [4]. This current study is first report of anti-H. pylori and antibiofilm activities and morphological conversion of the bacterial cells caused by C. operculatus flower bud and leaf fractions and DMC.
Furthermore, naturally occurring chalcones found in many medicinal and edible plants have been known to be precursors of plant flavonoids [56]. Flavonoids possessing antibacterial properties could penetrate the lipid bilayer membrane causing increase in membrane permeability and alleviating the bacterial pathogenicity [57]. In our study, the C. operculatus flower bud and leaf fractions, especially the isolated DMC were proven to effectively increase the membrane permeability of H. pylori. Results from SEM images indicated that hexane fraction of C. operculatus flower buds and DMC produced extensive morphological damages causing increase in the membrane permeability and exerted bactericidal effects. The H. pylori cells treated with hesperetin, naringenin, 7-O-butylnaringenin were shown to be damaged and resulted in morphological alterations or irregular shapes and rough surfaces [58,59]. Hesperidin was reported to interact with bacterial cells and induce membrane disruption leading to leakage of cytoplasmic components prior to cell death [60]. Ergüden and Ünver (2021) [61] proposed that phenolic chalcones induced ion leakage from Gram-positive bacterial cytoplasm prior to the membrane deformation and cell death.
Cytotoxicity experiments showed that the C. operculatus fractions and DMC are not toxic to the tested cell lines, in which the flower bud hexane fraction (DMC-rich fraction) was more toxic to the three cancer cell lines (MCF-7, Jurkat, and HeLa) than to the fibroblast cells. The DMC-rich extract obtained from fruits of S. nervosum was also reported to have stronger anticancer activities against A549 (human lung cancer cells) and HepG2 (human liver cancer cells) than the individual DMC [19] These showed that the fractions and DMC have selective inhibitory effects on H. pylori-urease and induce morphological conversions and membrane disruption leading to the cell death.
5. Conclusions
The fractions from flower buds and leaves of C. operculatus exerted various antioxidant activities. The results demonstrated that the aqueous fractions of flower buds and leaves, which contain high phenolic contents, exhibited significantly antioxidant effect. Flower bud and leaf hexane fractions with rich contents of flavonoids and saponins possessed considerable antibacterial activity towards H. pylori. The pronounced anti-H. pylori activity of the hexane fractions and DMC (2’,4’-dihydroxy-6’-methoxy-3’,5’-dimethylchalcone) may result from its potential role in urease inhibitory and antibiofilm properties. Additionally, the growth-inhibiting and bactericidal effects of the flower bud hexane fraction and DMC have been attributed to causing morphological changes, increasing permeability and damaging cell membrane of H. pylori. Moreover, inhibitory activities of C. operculatus fractions against α-glucosidase and DMC against α-amylase, and their safety for human cells indicate the need to evaluate their biological effectiveness in vivo. Flower bud and leaf-derived materials rich in chalcones and flavonoids could be a potential source of natural antioxidant and antibacterial agents for food and pharmaceutical products.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org, Figure S1: Scheme of flower bud and leaf solvent fractions partitioned by liquid-liquid fractionation from flower bud and leaf ethanolic extracts of Cleistocalyx operculatus and isolation of principle 1 derived from flower bud hexane fraction (BHF); Figure S2: HPLC of DMC isolated from the flower bud hexane fraction (BHF); Figure S3: FT-IR data of DMC isolated from flower bud hexane fraction (BHF); Figure S4: 1H-NMR data of DMC isolated from the flower bud hexane fraction (BHF); Figure S5: 13C-NMR data of DMC isolated from flower bud hexane fraction (BHF); Figure S6: 1H - 13C HMBC-correlations of DMC isolated from flower bud hexane fraction (BHF); Table S1: 1H NMR, 13C-NMR, and HMBC data of DMC isolated from flower bud hexane fraction (BHF).
Author Contributions
Conceptualization, D.T.T., L.T.M.N. and T.T.H.; methodology, D.T.T., M.T.T., N.T.M.T., P.N.P.T., P.T.H.T, P.T.G.T.,L.T.M.N. and T.T.H.; software, D.T.T.; validation, D.T.T., L.T.M.N. and T.T.H.; formal analysis, D.T.T., M.T.T., P.T.G.T., investigation, D.T.T., M.T.T., N.T.M.T., P.N.P.T., P.T.H.T, P.T.G.T., L.T.M.N. and T.T.H.; resources, D.T.T., L.T.M.N. and T.T.H.; data curation, D.T.T., M.T.T., N.T.M.T.; writing—original draft preparation, D.T.T. and T.T.H.; writing—review and editing, D.T.T. and T.T.H.; supervision, L.T.M.N. and T.T.H.; project administration, L.T.M.N.; funding acquisition, L.T.M.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Vietnam National Foundation for Science and Technology Development (NAFOSTED) under grant number 106-YS.06-2015.17
Institutional Review Board Statement
The study was approved by the Ethical Committee of the Vietnam National University, Ho Chi Minh City (No. 702/DHQG-KHCN).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Ye, C.L.; Lu, Y.H.; Li, X.D.; Wei, D.Z. HPLC analysis of a bioactive chalcone and triterpene in the buds of Cleistocalyx operculatus. S. Afr. J. Bot. 2005; 71, 312–315. [Google Scholar] [CrossRef]
- Ye, C.L.; Liu, X.G.; Huang, Q. Antioxidant activity and protection of human umbilical vein endothelial cells from hydrogen peroxide-induced injury by DMC, a chalcone from buds of Cleistocalyx operculatus. S. Afr. J. Bot. 2013, 86, 36–40. [Google Scholar] [CrossRef]
- Dung, N.T.; Kim, J.M.; Kang, S.C. Chemical composition, antimicrobial and antioxidant activities of the essential oil and the ethanol extract of Cleistocalyx operculatus (Roxb.) Merr and Perry buds. Food Chem. Toxicol. 2018, 46, 3632–3639. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.T.M.; Schultze, N.; Boger, C.; Alresley, Z.; Bolhuis, A.; Lindequist, U. Anticaries and antimicrobial activities of methanolic extract from leaves of Cleistocalyx operculatus L. Asian Pac. J. Trop. Biomed. 2017, 7(1), 43–48. [Google Scholar] [CrossRef]
- Woo, A.Y.H.; Waye, M.M.Y.; Kwan, H.S.; Chan, M.C.Y.; Chau, C.F.; Cheng, C.H.K. Inhibition of ATPases by Cleistocalyx operculatus. A possible mechanism for the cardiotonic actions of the herb. Vasc. Pharmacol. 2002, 38(3), 163–8. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Y. Inhibitory activities of extracts from Cleistocalyx operculatus flower buds on pancreatic lipase and α-amylase. Eur. Food Res. Technol. 2012, 235(6), 1133–1139. [Google Scholar] [CrossRef]
- Ye, C.L.; Lu, Y.H.; Wei, D.Z. Flavonoids from Cleistocalyx operculatus. Phytochem. 2004, 65(4), 445–447. [Google Scholar] [CrossRef]
- Ngan, L.T.M.; Dung, P.P.; Nhi, N.V.T.Y.; Hoang, N.V.M.; Hieu, T.T. Antibacterial activity of ethanolic extracts of some Vietnamese medicinal plants against Helicobacter pylori. AIP Conf. Proc. 2017, 1878, 020030. [Google Scholar] [CrossRef]
- Thanh, D.T.; Oanh, V.K.; Nguyen, H.C.; Ngan, L.T.M.; Hieu, T.T. Phytochemical composition, antioxidant, antibacterial, and enzyme inhibitory activities of organic extracts from flower buds of Cleistocalyx operculatus (Roxb.) Merr. et Perry. BioTechnologia. 2024; 105, 137–147. [Google Scholar] [CrossRef]
- Elbehiry, A.; Marzouk, E.; Aldubaib, M.; Abalkhail, A.; Anagreyyah, S.; Anajirih, N.; Almuzaini, A.M.; Rawway, M.; Alfadhel, A.; Draz, A.; Abu-Okail, A. Helicobacter pylori Infection: Current Status and Future Prospects on Diagnostic, Therapeutic and Control Challenges. Antibiotics. 2023, 12(2), 191. [Google Scholar] [CrossRef]
- Wroblewski, L.E.; Peek, R.M.; Wilson, K.T. Helicobacter pylori and gastric cancer: Factors that modulate disease risk. Clin. Microbiol. Rev. 2010, 23(4), 713–739. [Google Scholar] [CrossRef]
- Tsugane, S.; Sasazuki, S. Diet and the risk of gastric cancer: Review of epidemiological evidence. Gastric Cancer. 2007, 10(2), 75–83. [Google Scholar] [CrossRef]
- Eusebi, L.H.; Zagari, R.M.; Bazzoli, F. Epidemiology of Helicobacter pylori Infection. In Helicobacter. Blackwell Publishing Ltd. 2014, 19, 1–5. [Google Scholar] [CrossRef]
- Kusters, J.G.; Van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin Microbiol Rev. 2006, 19(3), 449–490. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Camargo, M.C.; El-Omar, E.; Liou, J.M.; Peek, R.; Schulz, C.; Smith, S.I.; Suerbaum, S. Helicobacter pylori infection. Nat. Rev. Dis. Primers. 2023, 9(1), 19. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, H.; Osaki, T.; Kamiya, S. Biofilm formation by Helicobacter pylori and its involvement for antibiotic resistance. BioMed Res. Int. 2015, 2015, 914791. [Google Scholar] [CrossRef]
- Gaddy, J.A.; Radin, J.N.; Cullen, T.W.; Chazin, W.J.; Skaar, E.P.; Trent, M.S.; Algood, H.M.S. Helicobacter pylori resists the antimicrobial activity of calprotectin via lipid a modification and associated biofilm formation. mBio. 2015, 6(6), e01349-15. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.N.T.; Huynh, T.D.H. Selective cytotoxicity of a Vietnamese traditional formula, Nam Dia long, against MCF-7 cells by synergistic effects. BMC Complement. Altern. Med. 2016, 16(1), 220. [Google Scholar] [CrossRef]
- Choommongkol, V.; Punturee, K.; Klumphu, P.; Rattanaburi, P.; Meepowpan, P.; Suttiarporn, P. Microwave-Assisted Extraction of Anticancer Flavonoid, 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethyl Chalcone (DMC), Rich Extract from Syzygium nervosum Fruits. Molecules 2022, 27(4), 1397. [Google Scholar] [CrossRef]
- Temesgen, S.; Sasikumar, J.M.; Egigu, M.C. Effect of Extraction Solvents on Total Polyphenolic Content and Antioxidant Capacity of Syzygium Aromaticum L. Flower Bud from Ethiopia. Biomed Res. Int. 2022, 2022, 4568944. [Google Scholar] [CrossRef]
- Rebaya, A.; Belghith, S.I.; Baghdikian, B.; Leddet, V.M.; Mabrouki, F.; Olivier, E.; Cherif, J.K.; Ayadi, M.T. Total Phenolic, Total Flavonoid, Tannin Content, and Antioxidant Capacity of Halimium halimifolium (Cistaceae). J. Appl. Pharm. Sci. 2015, 5(1), 052–057. [Google Scholar] [CrossRef]
- Ncube, B.; Nair, J.J.; Rárová, L.; Strnad, M.; Finnie, J.F.; Van Staden, J. Seasonal pharmacological properties and alkaloid content in Cyrtanthus contractus N.E. Br. S. Afr. J. Bot. 2015, 97, 69–76. [Google Scholar] [CrossRef]
- Benyong, H.; Ying, C.; Ying, R.; Chaoyin, C. Content determination of total saponins from Opuntia. BTAIJ. 2014, 10(18), 2014. [https://www.tsijournals.com/articles/content-determination-of-total-saponins-from-opuntia.pdf].
- Elouafy, Y.; El Yadini, A.; Mortada, S.; Hnini, M.; Harhar, H.; Khalid, A.; Abdalla, A.; Bouyahya, A.; Goh, K.; Ming, L.; Faouzi, M.; Tabyaoui, M. Antioxidant, antimicrobial, and α-glucosidase inhibitory activities of saponin extracts from walnut (Juglans regia L.) leaves. Asian Pac. J. Trop. Biomed. 2023, 13, 60–69. [Google Scholar] [CrossRef]
- Olszowy-Tomczyk, M.; Typek, R. Transformation of phenolic acids during radical neutralization. J. Food Sci. Technol. 2024, 61(4), 790–797. [Google Scholar] [CrossRef]
- Ngan, L.T.M.; Tan, M.T.; Hoang, N.V.M.; Thanh, D.T.; Linh, N.T.T.; Hoa, T.T.H.; Nuong, N.T.M.; Hieu, T.T. Antibacterial activity of Hibiscus rosa-sinensis l. red flower against antibiotic-resistant strains of Helicobacter pylori and identification of the flower constituents. Braz. J. Med. Biol. Res. 2021, 54, e10889. [Google Scholar] [CrossRef] [PubMed]
- Ngan, L.T.M.; Moon, J.K.; Kim, J.H.; Shibamoto, T.; Ahn, Y.J. Growth-inhibiting effects of Paeonia lactiflora root steam distillate constituents and structurally related compounds on human intestinal bacteria. World J. Microbiol. Biotechnol. 2012, 28(4), 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Shai, L.J.; Magano, S.R.; Lebelo, S.L.; Mogale, A.M. Inhibitory effects of five medicinal plants on rat alpha-glucosidase: Comparison with their effects on yeast alpha-glucosidase. J. Med. Plant. Res. 2011, 18(1), 2863–2867. [http://www.academicjournals.org/JMPR].
- Ogunyemi, O.M.; Gyebi, G.A.; Saheed, A.; Paul, J.; Nwaneri-Chidozie, V.; Olorundare, O.; Adebayo, J.; Koketsu, M.; Aljarba, N.; Alkahtani, S.; Batiha, G.E.S.; Olaiya, C.O. Inhibition mechanism of alpha-amylase, a diabetes target, by a steroidal pregnane and pregnane glycosides derived from Gongronema latifolium Benth. Front. Mol. Biosci. 2022, 9, 866719. [Google Scholar] [CrossRef] [PubMed]
- Hieu, T.T.; Truong, T.H.H.; Nguyen, T.T.L.; Nguyen, V.M.H.; Nguyen, T.M.N.; Luong, T.M.N. Growth-inhibiting, bactericidal, antibiofilm, and urease inhibitory activities of Hibiscus rosa sinensis L. flower constituents toward antibiotic sensitive- and resistant-strains of Helicobacter pylori. ACS Omega. 2020, 5, 20080–20089. [Google Scholar] [CrossRef]
- Guzman, J.; Téné, N.; Touchard, A.; Castillo, D.; Belkhelfa, H.; Haddioui-Hbabi, L.; Treilhou, M.; Sauvain, M. Anti-Helicobacter pylori properties of the ant-venom peptide bicarinalin. Toxins (Basel). 2018, 10(1), 21. [Google Scholar] [CrossRef]
- Devi, K.P.; Nisha, S.A.; Sakthivel, R.; Pandian, S.K. Eugenol (an essential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane. J. Ethnopharmacol. 2010, 130(1), 107–115. [Google Scholar] [CrossRef]
- Ngan, L.T.M.; Vi, N.T.; Tham, D.T.M.; Loan, L.T.T.; Ho, P.T.; Hieu, T.T. Antioxidant and anti-Helicobacter pylori activities of Hericium erinaceus mycelium and culture filtrate. Biomed. Res. Ther. 2021, 8(3), 4266–4275. [Google Scholar] [CrossRef]
- Schwarz, S.; Böttner, A.; Goosens, L.; Hafez, H.M.; Hartmann, K.; Kaske, M.; Kehrenberg, C.; Kietzmann, M.; Klarmann, D.; Klein, G.; Krabisch, P.; Luhofer, G.; Richter, A.; Schulz, B.; Sigge, C.; Waldmann, K.H.; Wallmann, J.; Werckenthin, C. A proposal of clinical breakpoints for amoxicillin applicable to porcine respiratory tract pathogens. Vet. Microbiol. 2008, 126(1), 178–188. [Google Scholar] [CrossRef]
- Mai, T.T.; Fumie, N.; Chuyen, N.V. Antioxidant activities and hypolipidemic effects of an aqueous extract from flower buds of Cleistocalyx operculatus (Roxb.) Merr. and Perry. J. Food Biochem. 2009, 33, 790–807. [Google Scholar] [CrossRef]
- Seo, J.; Lee, S.; Elam, M.L.; Johnson, S.A; Kang, J.; Arjmandi, B.H. Study to find the best extraction solvent for use with guava leaves (Psidium guajava L.) for high antioxidant efficacy. Food Sci. Nutr. 2014, 2, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Wijekoon, M.M.J.O.; Bhat, R.; Karim, A.A. Effect of extraction solvents on the phenolic compounds and antioxidant activities of bunga kantan (Etlingera elatior Jack.) inflorescence. J. Food Compost. Anal. 2011, 24, 615–619. [Google Scholar] [CrossRef]
- Minh, T.T. L.; Kieu, L.T.B.; Mai, S.T.T.; Ngoc, D.L.B.; Thuy, L.T.B.; Quyen, N.T.; Anh, T.T.; Huy, L. V.; Phong, N.V.; Duyen, C.T.M.; Minh, N.H.; Eric, G. Addition of Mentha arvensis in infusions of Cleistocalyx operculatus improves the Hedonic Score and retains the high antioxidant and anti-lipid-peroxidation effects. Appl. Sci. 2023, 13(5), 2873. [Google Scholar] [CrossRef]
- Dung, N.X.; Van, L.H.; Khoi, T.T.; Leclercq, P.A. GC and GC/MS analysis of the leaf oil of Cleistocalyx operculatus Roxb. Merr. et Perry (Syn. Eugenia operculata Roxb.; Syzygicum mervosum DC.). J. Essent. Oil Res. 1994, 6, 661–662. [Google Scholar] [CrossRef]
- Min, B.S.; Thu, C.V.; Dat, N.T.; Nguyen, C.; Dang, H.; Jang, H.S.; Hung, T.M. Antioxidative Flavonoids from Cleistocalyx operculatus Buds. Chem. Pharm. Bull. 2008, 56(12), 1725–1728. [Google Scholar] [CrossRef]
- Charoensin, S.; Taya, S.; Wongpornchai, S.; Wongpoomchai, R. Assessment of genotoxicity and antigenotoxicity of an aqueous extract of Cleistocalyx nervosum var. paniala in in vitro and in vivo models. Interdiscip. Toxicol. 2012, 5(4), 201–206. [Google Scholar] [CrossRef] [PubMed]
- Saeed, N.; Khan, M.R.; Shabbir, M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complement. Altern. Med. 2012, 12, 221. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compost. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Spósito, L.; Oda, F.B.; Vieira, J.H.; Carvalho, F.A.; dos Santos Ramos, M.A.; de Castro, R.C.; Crevelin, E.J.; Crotti, A.E.M.; Santos, A.G.; da Silva, P.B.; Chorilli, M.; Bauab, T.M. In vitro and in vivo anti-Helicobacter pylori activity of Casearia sylvestris leaf derivatives. J. Ethnopharmacol 2019, 233, 1–12. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Dung, N.T.; Suh, H.J.; Kang, S.C. Antibacterial activity of essential oil and extracts of Cleistocalyx operculatus buds against the bacteria of Xanthomonas spp. J. Am. Oil Chem. Soc. 2010, 87, 1341–1349. [Google Scholar] [CrossRef]
- Thuy, B.T.P.; Hieu, L.T.; My, T.T.A.; Hai, N.T.T.; Loan, H.T.P.; Thuy, N.T.T.; Triet, N.T.; Van Anh, T.T.; Dieu, N.T.X.; Quy, P.T.; Van Trung, N.; Quang, D.T.; Huynh, L.K.; Nhung, N.T.A. Screening for Streptococcus pyogenes antibacterial and Candida albicans antifungal bioactivities of organic compounds in natural essential oils of Piper betle L., Cleistocalyx operculatus L. and Ageratum conyzoides L. Chem. Pap. 2021, 75, 1507–1519. [Google Scholar] [CrossRef]
- Utama, K.; Khamto, N.; Meepowpan, P.; Aobchey, P.; Kantapan, J.; Sringarm, K.; Roytrakul, S.; Sangthong, P. Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Syzygium nervosum Seeds on Antiproliferative, DNA Damage, Cell Cycle Arrest, and Apoptosis in Human Cervical Cancer Cell Lines. Molecules. 2022, 27(4), 1154. [Google Scholar] [CrossRef]
- Nostro, A.; Cellini, L.; Di Bartolomeo, S.; Di Campli, E.; Grande, R.; Cannatelli, M.A.; Marzio, L.; Alonzo, V. Antibacterial effect of plant extracts against Helicobacter pylori. Phytother. Res. 2005, 19(3), 198–202. [Google Scholar] [CrossRef] [PubMed]
- Sufian, A.S.; Ramasamy, K.; Ahmat, N.; Zakaria, Z.A.; Yusof, M.I.M. Isolation and identification of antibacterial and cytotoxic compounds from the leaves of Muntingia calabura L. J. Ethnopharmacol. 2013, 146(1), 198–204. [Google Scholar] [CrossRef]
- Amin, M.; Anwar, F.; Naz, F.; Mehmood, T.; Saari, N. Anti-Helicobacter pylori and urease inhibition activities of some traditional medicinal plants. Molecules. 2013, 18(2), 2135–2149. [Google Scholar] [CrossRef]
- Egas, V.; Salazar-Cervantes, G.; Romero, I.; Méndez-Cuesta, C.A.; Rodríguez-Chávez, J.L.; Delgado, G. Anti-Helicobacter pylori metabolites from Heterotheca inuloides (Mexican arnica). Fitoterapia. 2018, 127, 314–321. [Google Scholar] [CrossRef]
- Chukiatsiri, S.; Wongsrangsap, N.; Ratanabunyong, S.; Choowongkomon, K. In vitro Evaluation of Antidiabetic Potential of Cleistocalyx nervosum var. paniala Fruit Extract. Plants 2023, 12(1), 112. [Google Scholar] [CrossRef]
- Wink, M. Importance of plant secondary metabolites for protection against insects and microbial infections. In Advances in Phytomedicine; Rai, M., M. C. Carpinella, M.C., Eds.; Publisher: Elsevier, Amsterdam, Netherlands, 2006; Volume 3. [Google Scholar] [CrossRef]
- Tran, T.H.; Luong, T.M.N.; Bui, V.L.; Tran, T.H. Effects of plant essential oils and their constituents on Helicobacter pylori: A Review. Plant Sci. Today. 2023, 10(2), 334–344. [Google Scholar] [CrossRef]
- Singh, A.; Amod, A.; Pandey, P.; Bose, P.; Pingali, S.; Shivalkar, S.; Varadwaj, P.; Sahoo, A.; Samanta, S. Bacterial biofilm infections, their resistance to antibiotics therapy and current treatment strategies. Biomed. Mater. 2022, 17, 022003. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khan, J.; Dukhyil, A.A.B.; Alarousy, R.M.I.I.; Attah, E.I.; Sharma, T.; Khairnar, S.J.; Bendale, A.R. Chalcone Scaffolds, Bioprecursors of Flavonoids: Chemistry, Bioactivities, and Pharmacokinetics. Molecules. 2021, 26(23), 7177. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial Activities of Flavonoids: Structure-Activity Relationship and Mechanism. Curr. Med. Chem. 2014, 22(1), 132–149. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Wang, C.M.; Su, C.H.; Ho, H.C.; Chang, C.H.; Chou, C.H.; Hsu, Y.M. Eudesmin attenuates Helicobacter pylori-induced epithelial autophagy and apoptosis and leads to eradication of H. pylori infection. Exp. Ther. Med. 2018, 15(3), 2388–2396. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Lee, J.H.; Kim, K.T.; Park, Y.S.; Nah, S.Y.; Ahn, D.U.; Paik, H.D. Antimicrobial effect of 7-O-butylnaringenin, a novel flavonoid, and various natural flavonoids against Helicobacter pylori strains. Int. J. Environ. Res. Public Health. 2013, 10(11), 5459–5469. [Google Scholar] [CrossRef]
- Sharaf, M.; Arif, M.; Hamouda, H.I.; Khan, S.; Abdalla, M.; Shabana, S.; Rozan, H.E.; Khan, T.U.; Chi, Z.; Liu, C. Preparation, urease inhibition mechanisms, and anti-Helicobacter pylori activities of hesperetin-7-rhamnoglucoside. Curr. Res. Microb. Sci. 2022, 3, 100103. [Google Scholar] [CrossRef]
- Ergüden, B.; Ünver, Y. Phenolic chalcones lead to ion leakage from Gram-positive bacteria prior to cell death. Arch. Microbiol. 2021, 204(1), 3. [Google Scholar] [CrossRef]
Figure 1.
Effect of C. operculatus fractions and DMC at sub-MICs on H. pylori biofilm formation 48 h post-treatment. Data are reported as means ± SD.
Figure 1.
Effect of C. operculatus fractions and DMC at sub-MICs on H. pylori biofilm formation 48 h post-treatment. Data are reported as means ± SD.
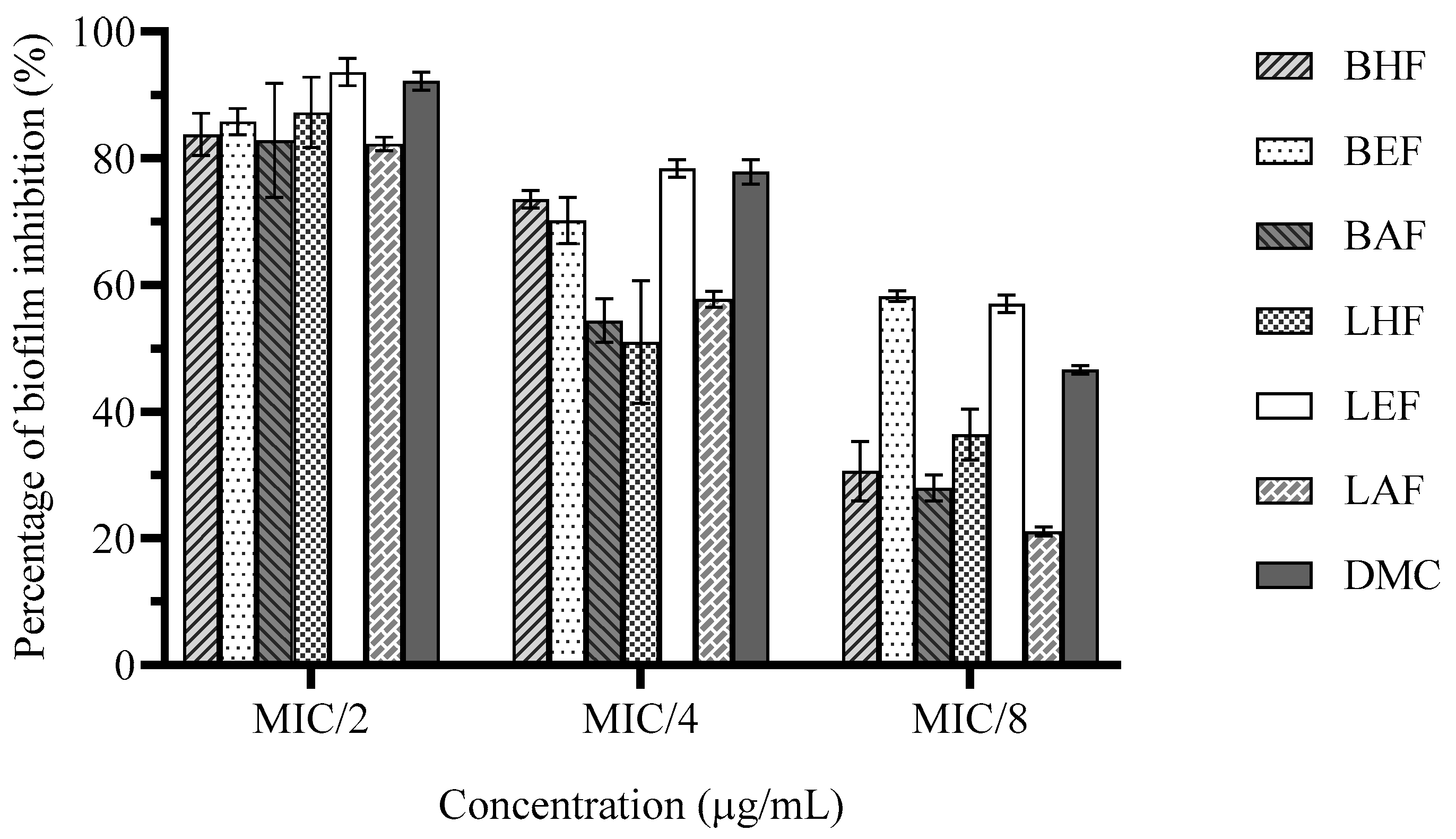
Figure 2.
SEM micrographs of H. pylori ATCC 43504 depicted untreated cells (A and B) and cells treated with 125 µg/ml BHF (C and D) and 50 µg/ml DMC (E and F), bar 5 and 1 µm, respectively.
Figure 2.
SEM micrographs of H. pylori ATCC 43504 depicted untreated cells (A and B) and cells treated with 125 µg/ml BHF (C and D) and 50 µg/ml DMC (E and F), bar 5 and 1 µm, respectively.

Figure 3.
Effect of C. operculatus fractions and DMC at sub-MICs on the uptake of crystal violet by H. pylori ATCC 43504 after 2h treatment. Data are reported as means ± SD (n = 3).
Figure 3.
Effect of C. operculatus fractions and DMC at sub-MICs on the uptake of crystal violet by H. pylori ATCC 43504 after 2h treatment. Data are reported as means ± SD (n = 3).
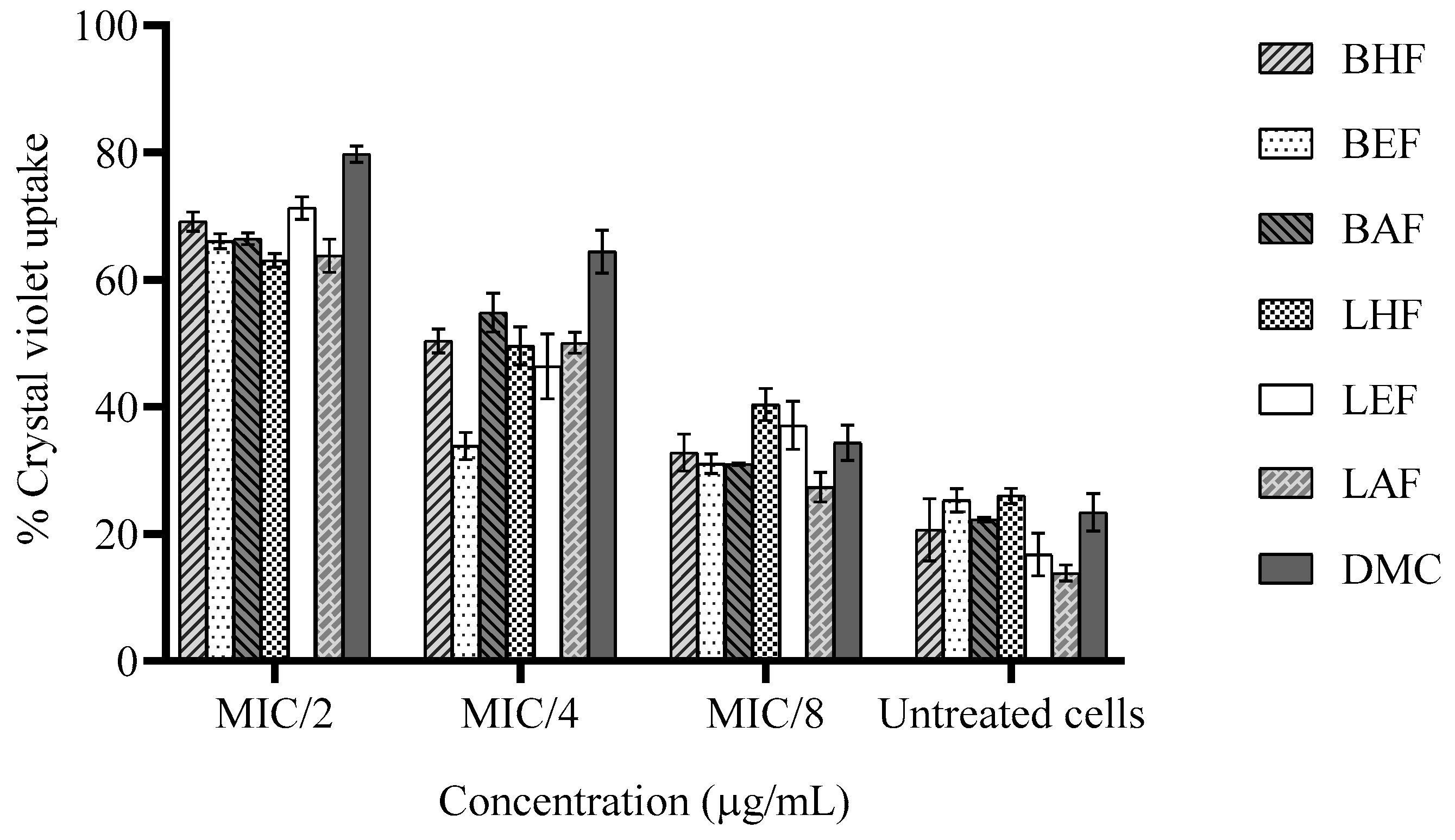

Table 1.
Total phytochemical contents in different fractions from flower buds and leaves of C. operculatus.
Table 1.
Total phytochemical contents in different fractions from flower buds and leaves of C. operculatus.
| Fractions | TPC | TFC | TSC | TTC | TAC |
|---|---|---|---|---|---|
| BHF | 426.77c ± 1.22 | 134.77a ± 7.75 | 153.33a ± 4.69 | 42.97a ± 2.93 | 1.66d ± 0.09 |
| BEF | 280.46d ± 11.06 | 85.88b ± 2.52 | 158.10a ± 5.97 | 22.97c ± 1.73 | 1.50d ± 0.17 |
| BAF | 768.18a ± 12.20 | 11.04e ± 0.53 | 81.59b ± 1.20 | 13.53d ± 2.93 | 3.04c ± 0.20 |
| LHF | 201.63f ± 3.45 | 76.54bc ± 1.72 | 88.25b ± 9.46 | 33.17b ± 2.10 | 4.81b ± 0.09 |
| LEF | 238.47e ± 4.18 | 71.72c ± 1.74 | 75.24b ± 5.95 | 10.39d ± 1.29 | 4.73b ± 0.05 |
| LAF | 490.74b ± 7.29 | 25.56d ± 0.21 | 25.56c ± 2.44 | 4.36e ± 0.96 | 5.41a ± 0.08 |
1 BHF: flower bud hexane fraction, BEF: flower bud ethyl acetate fraction, and BAF: flower bud aqueous fraction. LHF: leaf hexane fraction, LEF: leaf ethyl acetate fraction, and LAF: leaf aqueous fraction. TPC: total phenolic content (mg GAE/g dry extract), TFC: total flavonoid content (mg QE/g dry extract), TSC: total saponins content (mg OA/g dry extract), TTC: total tannin content (mg CE/g dry extract), TAC: total alkaloid content (mg AE/g dry extract). Data are shown as mean ± SD (n = 3). Means within a column with different letters indicate significant difference at p < 0.05.
Table 2.
Antioxidant activity of different fractions from flower buds and leaves of C. operculatus.
| Samples | FRAP (mg TE/g extract) |
DPPH IC50 (µg/ml) |
ABTS IC50 (µg/ml) |
|---|---|---|---|
| BHF | 69.01e ± 1.833 | 33.99c ± 0.76 | 1.70b ± 0.09 |
| BEF | 91.95c ± 1.302 | 58.46a ± 1.370 | 1.48b ± 0.162 |
| BAF | 201.80b ± 4.502 | 24.69d ± 0.194 | 1.08c ± 0.013 |
| LHF | 55.57f ± 1.265 | 39.96b ± 0.237 | 1.06c ± 0.084 |
| LEF | 78.56d ± 1.13 | 16.05e ± 0.031 | 0.98c ± 0.007 |
| LAF | 301.82a ± 2.306 | 11.24f ±1.524 | 0.55d ± 0.004 |
| Ascorbic acid | ND | 3.34g ± 0.017 | ND |
| Trolox | ND | ND | 2.63a ± 0.05 |
1 BHF: flower bud hexane fraction, BEF: flower bud ethyl acetate fraction, and BAF: flower bud aqueous fraction. LHF: leaf hexane fraction, LEF: leaf ethyl acetate fraction, and LAF: leaf aqueous fraction. Data are shown as mean ± SD (n = 3). Means within a column with different letters indicate significant difference at p < 0.05. ND: not determined.
Table 3.
In vitro minimal inhibitory concentration (MIC) values of C. operculatus fractions and DMC against H. pylori.
Table 3.
In vitro minimal inhibitory concentration (MIC) values of C. operculatus fractions and DMC against H. pylori.
| Samples | MIC (µg/ml) | |
|---|---|---|
| H. pylori ATCC 51932 | H. pylori ATCC 43504 | |
| BHF | 125 | 125 |
| BEF | 250 | 500 |
| BAF | 500 | 500 |
| LHF | 125 | 125 |
| LEF | 250 | 500 |
| LAF | 500 | 1000 |
| DMC | 25 | 50 |
| Amoxicillin | 0.01 | 0.01 |
1 BHF: flower bud hexane fraction, BEF: flower bud ethyl acetate fraction, and BAF: flower bud aqueous fraction. LHF: leaf hexane fraction, LEF: leaf ethyl acetate fraction, and LAF: leaf aqueous fraction. Test materials with MIC values of ≤ 130, >130–<630, 630–1250, >1250–< 2500, and ≥2500 µg/ml was classified as the extremely high, high, moderate, low, and no inhibitory activity against the growth of test bacteria, respectively [26].
Table 4.
Inhibitory effect of C. operculatus fractions and DMC on H. pylori-urease, α-glucosidase and α-amylase.
Table 4.
Inhibitory effect of C. operculatus fractions and DMC on H. pylori-urease, α-glucosidase and α-amylase.
| Samples | IC50 (µg/ml) | ||
|---|---|---|---|
| H. pylori-Urease | α-Glucosidase | α-Amylase | |
| BHF | 2.3e ± 0.13 | 1.5c ± 0.09 | 398.5d ± 5.3 |
| BEF | 4.9c ± 0.40 | 0.9c ± 0.01 | 497.2b ± 17.1 |
| BAF | 2.5de ± 0.21 | 0.8c ± 0.01 | 444.3c ±14.3 |
| LHF | 3.2de ± 0.26 | 2.6c ± 0.16 | 191.3f ± 9.5 |
| LEF | 3.6d ± 0.01 | 1.2c ± 0.06 | 292.6e ± 16.6 |
| LAF | 6.8b ± 0.07 | 0.6c ± 0.02 | 1281.7a ±23.7 |
| DMC | 3.2de ± 0.03 | 94.6a ± 2.57 | 83.80g ± 0.08 |
| Thiourea | 44.3a ± 1.12 | ND | ND |
| Acarbose | ND | 25.6b ± 0.70 | 75.0g ± 3.86 |
1 BHF: flower bud hexane fraction, BEF: flower bud ethyl acetate fraction, and BAF: flower bud aqueous fraction. LHF: leaf hexane fraction, LEF: leaf ethyl acetate fraction, and LAF: leaf aqueous fraction. Data are shown as mean ± SD (n = 3). Means within a column with different letters indicate significant difference at p < 0.05. ND: not determined.
Table 5.
Cytotoxicity of C. operculatus fractions and DMC against four human cell lines.
| Samples | CC50 (µg/ml) | |||
|---|---|---|---|---|
| MCF-7 | Jurkat | HeLa | Fibroblast | |
| BHF | 30.79d ± 0.83 | 18.51d ± 1.37 | 31.70c ± 5.74 | >100 |
| BEF | 57.74c ± 1.13 | 51.06c ± 1.42 | 92.53a ± 4.90 | >100 |
| BAF | > 100 | >100 | >100 | >100 |
| LHF | 89.00a ± 1.33 | 56.78c ± 2.62 | 86.45ab ± 4.06 | >100 |
| LEF | 85.43a ± 2.76 | 66.73b ± 1.02 | 91.72a ± 1.43 | >100 |
| LAF | >100 | 97.25a ± 1.28 | 77.07b ± 5.06 | >100 |
| DMC | 71.41b ± 2.14 | 73.82b ± 7.23 | >100 | >100 |
| Camptothecin | 0.007e ± 0.002 | 0.005e ± 0.001 | 0.89d ± 0.088 | 1.57 ± 0.84 |
1 BHF: flower bud hexane fraction, BEF: flower bud ethyl acetate fraction, and BAF: flower bud aqueous fraction. LHF: leaf hexane fraction, LEF: leaf ethyl acetate fraction, and LAF: leaf aqueous fraction. Data are shown as mean ± SD (n = 3). Means within a column with different letters indicate significant difference at p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



