
Submitted:
21 August 2024
Posted:
22 August 2024
You are already at the latest version
Abstract
Cumulative evidence attests to the essential roles of commensal microbes in the physiology of the hosts. Although microbiome has been a major research subject since the time of Luis Pasteur and William Russell over 140 years ago, recent findings that certain intracellular bacteria contribute to the pathophysiology of healthy or diseased tissues have brought the field of microbiome to a new era of investigation. In particular, in the field of breast cancer research, breast-tumor resident bacteria are deemed to be the essential players of tumor initiation and progression. This is a resurrection of Russel’s bacterial cause of cancer theory abandoned over 100 years ago. This review will introduce some of the recent findings that exemplify the roles of breast tumor-resident microbes in breast carcinogenesis and metastasis and provide mechanistic explanations of these phenomena. Such information would be able to justify the utility of breast-tumor resident microbes as biomarkers for disease progression and therapeutic targets.
Keywords:
breast cancer
; cancer metastasis
; tumor-resident bacteria
; microbiome
; metabolites
1. Introduction
External and internal surfaces of animal bodies are entirely covered by microorganisms. For humans, each person contains about 40 trillion microbes, which is as much as the total 30 trillion host cells [1]. These commensals are mostly found in the gut, where their density reaches almost 10 trillion microbes per ml and they weigh about one kilogram per person [2]. It is increasingly evident that these symbionts are not merely passive passengers, but are essential players for fundamental functions of the body, including immunity, metabolism, and energy balance [3]. This notation is well supported by the fact that germ-free (GF) mice manifest serious defects in lymphoid tissue structure and functions [4]. In particular, their gut mucosal immunity is severely compromised due to low numbers of lymphocytes and antibody production [5,6,7].
It is increasingly evident that different physiological conditions between healthy cohorts and cancer patients could be largely attributed to discrete characterization of microbial floras. It is established that not only tumor microbiotas, but also gut microbiotas, of cancer patients are far less diverse than the normal counter-parts—a condition of ‘dysbiosis’. Thus, correcting the microbiome of cancer patients has gained traction as an adjuvant approach. [8,9,10]. For example, normalizing the microbiome of cancer patients through FMTs from healthy cohorts or treatment-responders has been proven for its therapeutic benefits. In fact, treating patients of Trastuzumab-resistant HER2-positive breast cancer with FMT from drug responders improves Trastuzumab-response and tumor immunity [11]. However, the detailed mechanisms of FMT-induced anti-cancer effects have not been fully elucidated.
Along with gut microbiota, breast microbiota is proposed to play critical roles in the breast health and carcinogenesis [8,9,10]. Breast-resident microbes are originated from either the skin/nipple microbiota or those translocated from the guts along with immune cells, such as dendritic cells and macrophages [8,12,13]. Breast microbiota, however, could also be modified by environmental agents, such as aseptic solutions affecting skin microbiota [14]. Gut-breast microbial translocation, termed ‘gut-breast axis’, greatly contributes to the composition of microbiotas of breast tissues and milk [15,16,17,18]. This phenomenon, however, has mostly been conceived in relation to pregnancy and its influence by female sexual hormones [15,16,19,20,21,22]. Thus, it remains unclear whether gut-breast axis exists outside pregnancy on a regular basis and contributes to breast pathophysiology. If this holds true, gut-breast axis may contribute to FMT-mediated anti-breast cancer effects.
2. Breast Tissue Microbiota
The human breast contains a unique microbiota different from those in other parts of the body, playing critical roles in breast health as well as the health of offsprings [23]. Breast tissue microbiota is more diverse (higher α-diversity) than the skin tissue, while each species’ relative abundance (Shannon Index) is similar between them [24]. Sample-to-sample differences of microbiota compositions (β-diversities) are also higher in the breast compared to the skin, owing to major differences found in less abundant microbial species [24]. These features of the breast microbiota are independent of its location within the breast, parity, age, and nationality of the individual [23]. Based on the microbiotas of healthy livers or breast tumors, healthy breast-resident bacteria are expected to mostly reside inside parenchymal cells [25,26]. However, there are clear differences between healthy breast microbiota and breast tumor microbiota [24]. For example, the most abundant bacterial phyla in the healthy breast tissues of women older than 18 years old are Proteobacteria and Firmicutes, whereas these bacteria are under-represented in tumors (Table 1) [27]. Many of these bacteria abundant in healthy breast tissues produce beneficial biomaterials that confer anti-tumor and pro-immunogenic activities to protect the healthy tissue microenvironment.
3. Breast Milk Microbiota
Breast milk microbiota is proposed to be linked to breast tissue microbiota, although there has been no study to confirm their direct relationships. Breast milk microbiota is detectable from the third trimester of pregnancy through lactation. Breast milk, in particular, colostrum (first milk after giving birth), is the primary source of commensals to the newborn [57], whereas maternal-neonatal microbial transfers during pregnancy are conducted through the placenta and amniotic fluid [58,59]. This bacterial transfer through breast milk greatly contributes to the bacterial composition of infants’ guts which is similar to that of breast milk [60]. Breast milk microbiota plays critical roles in the infant’s immune development and his/her health of early and later life. Thus, dysbiosis of the breast milk microbiota would greatly influence infant development [61]. Typically, a baby ingests 1 × 105 to 1 × 107 bacteria a day while consuming approximately 800 mL/day of breast milk [62]. These breast milk bacteria include the genera Lactobacillus, Staphylococcus, Enterococcus, and Bifidobacterium (Table 1) [57]. Breast milk contains olisaccharides which are indigestible by the host, but are digested by enzymes produced by specific gut bacteria, such as bifidobacterial and lactobacilli, which utilize the metabolites for their expansion [63]. In addition, breast milk contains bacterial species that produce short-chain fatty acids (SCFAs), such as butylate, acetate, and formic acid (e.g., Coprococcus, Faecalibacterium, and Roseburia spp). These SCFA-producing bacteria would repopulate the neonatal guts and play a beneficial role in weight gain and adiposity [59,64,65,66].
4. Breast Tumor Microbiota
Microbes within tumors are mostly localized within tumor parenchyma as well as immune cells [26,53]. Since normal tissue microbes presumably reside within parenchymal cells as well, they are proposed to be a major source of intratumoral microbes [67,68]. Nevertheless, breast tumor microbiota is greatly different from healthy breast microbiota, indicating substantial influences of bacterial transfer from other parts of the body during tumorigenesis (Table 1). Breast tumor microbiotas are in general abundant in Fusobacterium, Atopobium, Gluconacterobacter, Hydrogenophaga, and Lactobacillus [24], unlike normal breast microbiotas abundant in Proteobacteria and Firmicutes [27]. Breast tumor microbiota is associated with dysregulation of cell proliferation, metabolic pathways, and immunological responses, contributing to tumor growth and progression (Table 1) [40,45,46,69,70]. Conversely, normal breast microbiota is associated with increased cysteine and methionine metabolism, glycosyltransferases, and fatty acid biosynthesis, promoting immunological responses [24,71,72]. Furthermore, breast tumor microbiota is enriched for Enterobacteriaceae and Staphylococcus, compared to healthy breast microbiota [27]. Both bacteria are known to produce genotoxins that induce DNA damage to help induce malignant progression of host cells [38,39]. In addition, lactic acid producing Lactobacilli, also abundant in breast tumor microbiota, could lower pH and induce metabolic rewiring of the tumor microenvironment (TME), leading to chemotherapy and radiation resistance of tumors [73]. These three taxa of breast tumor-associated bacteria are also found to promote tumor metastasis and colonization, while being transported along with tumor cells to the metastatic site [74]. For colorectal cancer, on the contrary, a different bacterial taxon Fusobacterium is transported along with colon cancer cells to the metastatic site [75], suggesting the roles of different bacteria in metastasis of different types of cancers. The involvement of breast microbiota in tumor metastasis will be further discussed below.
4.1. Breast Cancer Subtype-Specific Microbiota
Different tumor types have distinct microbial compositions, indicating the impacts of different tissue/TME (Table 2) [26]. Furthermore, even among breast tumors, different tumor subtypes (Luminal A, Luminal B, Her2+, and triple negative (TN) types) have distinct microbial compositions (Table 3). This indicates that the heterogeneity of molecular and metabolic profiles and cells of origin among different breast tumor subtypes impacts the fitness of different microbial communities [56,76,77]. For example, luminal subtypes (Luminal A and B) are abundant for the phyla Tenericutes, Proteobacteria, and Planctomycetes. Above all, the most abundant genus in Luminal A tumors is Xanthomonadales (phylum Proteobacteria), while that for Luminal B tumors is Clostridium (phylum Firmicutes) [78]. Conversely, HER2+ breast tumors are abundant in Akkermansia (phylum Verrucomicrobia), Thermi, Firmicutes (Filibacter, Anaerostipes, and Granulicatella_US31), Bacteroidetes (Cloacibacterium, Alloprevotella, and Dyadobacter), and Proteobacteria (Burkholderiales, Helicobacter pylori, PRD01a011B, Stakelama, and Blastomonas) [26,78,79,80,81]. In contrast, TN breast tumors are enriched for Euryarchaeota, Cyanobacteria, Firmicutes, Prevotella, Arcanobacterium, and Brevundimonas [79,82]. In particular, the presence of Listeria fleischmannii (Firmicutes) in TN tumors is shown to be strongly associated with activation of epithelial-to-mesenchymal transition pathway, while the presence of Haemophilus influenza (Pseudomonadota) is correlated with tumor growth and cell cycle progression [83].
4.2. Race/Ethnicity-Specific Breast Cancer Microbiota
In addition to differences in microbiotas among different breast tumor subtypes, races and ethnicities of individuals greatly impact the composition of breast tumor microbiota. This is at least in part attributed to distinct cellular and immunological patterns of breast tumors of Asian, Black and white women (e.g., Asian: high Th1 cells (IFNγ), and megakaryocytes; Black: high dendritic cells, B cells, mesenchymal stem cells, and CXCL9 expression; White: high adipocytes, hematopoietic stem cells, and endothelial cells) [108]. Smith et al. showed that Xanthomonadaceae was the most abundant member in breast tumors from Non-Hispanic White women, whereas genus Ralstonia was most abundant in breast tumors from Non-Hispanic Black women. They also showed that tumors from Non-Hispanic white women were richer in Phylum Bacteroidetes compared to Non-Hispanic Black women [78]. Similarly, Thyagarajan et al. reported that Phylum Bacteroidetes was significantly over-represented in TN breast tumors from white women. Conversely, in TN breast tumors from Black women, phyla Actinobacteria and Thermi and genus Bradyrhizobiaceae were underrepresented. TN tumors from Black women showed reduction of Shannon diversity than adjacent normal tissue, while the trend was reversed for white women [109].
Furthermore, Parida et al. reported racially distinct bacterial biomarkers for breast tumors. For Asian, Pseudomonas, Terrabacter, Clostridiodes, Aestuariibacter, Succinimonas, Catellicoccus, Leucobacter, Rhizobium, Rhodococcus, Methylobacter and Planctopirus are elevated; for Black, Xanthomonas, Amycolatopsis, Aphanizomenon, Anaerovorax, Aminiphilus, Trichormus, Chlorobium, and Sulfurovum are elevated; and for White: Halonatronum, Salinarchaeum and Amorphus are elevated. Such racial differences in microbial components of breast tumors are proposed to be linked to differential metastasis predictors among races [108].
5. Origin of Breast Tissue Microbiota
The origins of breast tissues and milk microbiotas are currently unclear; however, they are proposed to be derived from the breast skin, the oral cavity of the suckling infant, and the maternal gut through gut-breast axis (Figure 1) [58].
5.1. Microbial Transfer from Breast Skin
Mechanisms of microbial transfer from the skin to breast have not yet been clearly determined, although there are several possible scenarios based on the transfer of pathogenic bacteria from the skin to mammary glands. Abnormal microbiota of the breast skin could contribute to the pathogenicity of breast tissue, attesting to microbial transfer from the skin to the breast tissue [14]. Skin microbes such as Pseudomonas aeruginosa possess fatty-acid metabolising capabilities and could become pathogenic in the breast tissue [110]. Additionally, Staphylococcus aureus enriched in the skin of atopic dermatitis could lead to the formation of breast abscess also colonized by S. aureus [111]. In fact, Staphylococcus is among the most abundant genera in breast tumors and strongly linked to breast cancer metastasis, attesting to the role of skin bacteria in breast tumors [40,47,109,112]. In particular, bacteraemia and colonization of S. aureus in certain tissues could promote the incidence of primary tumors [113,114]. Other bacterial taxa linked to increased breast cancer risk include Bacillus, Bacteroidetes, Brevundimonas, Comamonadaceae, Enterobacteriaceae, and Methylobacterium which are also found in the skin microbiota, supporting the possibility of their transfer from the skin [47,106,109,110]. Furthermore, increased numbers of Corynebacterium and Pseudomonas, usually only found in normal skin flora, could break the skin barrier and penetrate deep into the breast tissue to induce granular lobular mastitis [111,112].
Iatrogenic breakdown of the skin barrier during medical procedures could also result in contamination of the underlying breast tissue by skin commensals [113]. For example, breast skin microbes, such as Staphylococcus epidermidis, play roles in the pathogenesis of breast implant complications including anaplastic large cell lymphoma [113,114,115,116,117]. On the contrary, mechanisms of microbial transfer from the skin to the breast tissue without damaging the skin barrier are more uncertain. Proposed scenarios include retrograde transfer through the nipple and ducts (see the details below) [57] and contamination during nipple aspirate fluid procedure [106].
5.2. Microbial Transfer from Nipple
The nipple of mammary gland contains about ten orifices of milk ducts [123]. It was initially proposed that these ductal openings facilitate bacterial transfer from mother’s skin into the breast milk [124]. However, this possibility was ruled out by the finding that microbial compositions of the nipple skin and nipple aspirate fluid are significantly different [125]. Especially, there are strictly anaerobic species, such as Lactobacilli and bifidobacteria, enriched in the breast milk which are unlikely to have originated from skin microbiota [29]. As a likely mechanism of bacterial transfer through nipples, there are some degrees of retrograde flow of milk back into the mammary glands from the infant’s mouth during suckling [57,126]. Such oscillating milk flows allow mothers to respond to pathogens afflicting infants, build antibodies for them, and transfer these antibodies back to the infants so that they can fight against illnesses. Nevertheless, such retrograde bacterial transfer from infants only serves to influence, but not act as, the original source of the maternal breast microbiota. In fact. certain anerobic bacteria, such as Lactobacillus vini and paracasei, are more abundant in the breast of nulliparous or never-breastfed women than breast-fed women than [35], suggesting that these bacteria are potentially derived from the maternal guts.
5.3. Microbial Transfer via Gut-Breast Axis or Oro-Breast Axis
Translocation of gut bacteria to external tissues is commonly associated with disease conditions that impair the intestinal epithelial tight junctions allowing luminal bacteria to move across the epithelial barrier and get into the bloodstream [127]. However, such bacterial translocation also takes place, although to a lesser extent, in healthy individuals, involving beneficial gut bacteria such as Lactobacilli and Bifidobacteria [128,129,130]. Such non-disease-related bacterial translocation appears to involve select species and be associated with immunotaining and immunomodulation of the host [131,132,133].
During pregnancy and lactation, maternal gut bacteria translocate to mammary glands so that they could be transferred to offsprings for colonizing their guts. Especially, during late pregnancy, there are synchronous changes in maternal mammary glands and guts. Mammary glands undergo structural and functional remodeling to become specialized organs that produce and transmit nutrients and other components necessary for neonatal growth [134]. Lactating mammary glands are also an effector site of the mucosal-associated lymphoid tissue system, playing essential roles in infants’ immunity [135]. Along with soluble immune factors, breast milk, especially, colostrum, contains select types of leukocytes, such as neutrophils, macrophages, and lymphocytes [136], facilitated by looser tight junctions of breast epithelia after giving a birth [137]. In particular, leukocytes exposed to antigens in the guts may migrate to mammary glands and be transferred to infants through breast milk for their defense and immune training [138,139]. Synchronously, in maternal guts epithelial permeability increases to facilitate bacterial transmigration [140]. Furthermore, gut microbiota undergoes metabolic adaptation to elevated glucose levels, which would further modify their ability to translocate across the intestinal epithelium and reach mammary glands [141].
Over decades, it has been known that breast milk, maternal feces and infant feces share same bacterial species, attesting to their physical connections [16]. Then, Perez et al. reported that a group of gut bacteria appeared to be physically translocated to mesenteric lymph nodes and then to mammary glands during late pregnancy and lactation. Bacteria translocated to the mammary glands would then enter breast milk and be transferred to the infants to establish the microflora of the neonatal guts. Furthermore, the same study showed that viable gut bacteria found in milk-producing breast cells were also detected in peripheral blood mononuclear cells (PBMC), indicating that PBMC helped translocation of these gut bacteria to the breast [131].
The theory of gut-breast bacterial translocation was confirmed by studies demonstrating that orally administered Lactobacilli strains reached the breast milk of mothers [142,143]. Such studies also support another theory of an oro-mammary bacterial translocation brought up by a finding that maternal oral bacteria and milk microbiota partially overlap [144]. Here, the majority of oral bacteria are expected to travel through the gastrointestinal (GI) tract to reach the guts and then be transported to the breast via gut-breast axis, whereas a small fraction of them. In contrast, a small fraction of oral bacteria could directly enter oral and maxillofacial blood circulation to spread to distant tissues/organs [145]. Oro-mammary translocation is in particular important as a cause for the abundant oral bacteria (e.g., Fusobacteria and Streptococci) in breast tumors [145].
6. Mechanisms of Bacterial Translocation
Although the pathway and mechanisms that some bacteria could enter the breast tissue has not been elucidated yet, some works have offered a plausible scientific basis. So far, there are two major mechanisms proposed: internalization/transcytosis by gut epithelia; and direct sampling by phagocytic cells (Figure 2).
6.1. Internalization into Epithelial Cells
Luminal bacteria could be internalized into gut epithelial cells and subsequently taken by DCs or macrophages in the mucosal environment. There are several potential mechanisms for internalization of non-invasive bacteria into gut epithelial cells. First, intestinal epithelia harbor specialized microfold (M) cells that transcytose luminal bacteria to make them available to mucosal immune cells [146]. Alternatively, upon activation of TLR4, non-specialized enterocytes or kidney epithelial cells were found to transcytose gram-negative gut bacteria [147]. Second, metabolic and oxidative stress, including hypoxia, low dose of nitric oxide, and uncoupling of mitochondrial oxidative phosphorylation, could damage tight junction of gut epithelia and induce transcytotic bacterial transfer to epithelia [148,149,150,151]. Third, low concentrations of IFNγ could cause the influx of noninvasive E. coli bacteria into human colon epithelia without affecting cell viability and tight junctions [152]. Such IFNγ-mediated transcytotic bacterial transfer was shown to depend on extracellular signal-regulated kinase (ERK) 1/2 and ADP-ribosylation factor (ARF)-6 [153]. Fourth, infection of intestines with the parasites Giardia lamblia or Campylobacter jejuni could damage gut epithelial barriers and tight junctions and induce penetration of luminal bacteria to the epithelia [154,155]. Fifth, viable non-pathogenic bacteria could enter host cells through endocytic pathways associated with lipid rafts and caveolin-1. Caveolin-1 or cholesterol was in fact found colocalized with bacteria-containing endosomes in epithelial cells [155,156]. These methods exploited by non-invasive bacteria are different from those of invasive pathogens using specialized needle-like systems to inject effector proteins into epithelial cells and manipulate the host cytoskeletons for anchorage and entry [157].
6.2. Sampling and Transportation by Immune Cells
It has been known that certain type of immune cells, especially, those denoted as CD18+ cells such as dendritic cells (DCs) and macrophages, could penetrate the gut epithelial barrier and directly take up non-pathogenic commensal bacteria from the lumen [158,159]. Especially, DCs are capable of opening tight junctions of intestinal epithelia and sampling lumina bacteria without destroying the epithelial integrity because of their ability to repair damage. This allows non-invasive gut bacteria to spread to extra-intestinal organs [158]. Similarly, macrophages could promote the extra-intestinal dissemination of non-invasive gut bacteria [159]. It was shown that induction of DCs with viable commensal bacteria, but not with dead bacteria, stimulates DC maturation, indicated by the increase in the class II major histocompatibility complex and the B7.2 protein on the cell surface, and their translocation across colon epithelium [160,161,162]. DCs, and possibly macrophages, that have taken up luminal bacteria then migrate to the nearby mesenteric lymph nodes and could stay there up to several days [163]. Such lymphoid tissue resident gut commensals are found to elevate anti-inflammatory signaling to help establish mutualism with host immunity [164]. Alternatively, these lymphoid-resident gut commensal could be taken up by lymphocytes, and transported to distant tissues, such as lactating mammary glands [165,166]. In lactating mammary glands, colonization of immune cells and their bacteria cargos is selective due to regulation by lactogenic hormones and retrograde signaling from suckling infants requesting specific immune cells to fight against their ailments [136,167,168].
7. Functions of Intracellular Microbiota
Increasing evidence demonstrates the existence of different intracellular bacteria in humans and mice [169–172]. The prevalence of intracellular bacteria over extracellular bacteria in tissues is largely attributed to more efficient immunological clearance of the latter than the former [173]. These intracellular microbes are proposed to play direct roles of in the pathophysiology of normal tissues and tumors [174]. This view has been increasingly solidified since the groundbreaking discoveries of the roles of Helicobacter pylori in stomach ulcers and, then, gastric cancer 30 years ago [175]. The recent surge of the next-generation sequencing technologies has allowed investigators to profile microbial compositions of tumor tissues, identify tumor-associated microbes, and study their specific functions. Certain commensal bacteria invade cancer cells and remain inside cells during tumor progression and even metastases [75,174]. This is largely attributed to the fact that intracellular bacteria are better protected than extracellular bacteria in a highly immunologic TME [74].
8. Bacterially Produced Metabolites
As discussed above, microbiotas in human breast milk and breast tissue play essential roles in infants’ development and healthy intestinal microbiota and immunity. In particular, different bacterially-produced metabolites contribute to differences in breast tissue microenvironment and health of offsprings [19]. A group of bacterially produced metabolites are found to exert beneficial effects. For example, short-chain fatty acids (SCFAs), such as butyrate, acetate and formic acid, in the breast milk play promote weight gain and adiposity of infants [176]. Cadaverine, a metabolite produced by bacterial lysine decarboxylase, is found to suppress breast cancer progression and metastasis, although the synthesis is downmodulated in breast cancer patients [177]. Also, indolepropionic acid (IPA) is a bacterial tryptophan metabolite and has cytostatic properties through activation of aryl hydrocarbon and pregnane X receptors. Ectopic application of IPA to breast cancer cells is found to suppress their growth and metastasis [178]. On the other hand, another group of bacteria-derived metabolites exacerbate breast cancer growth. For example, queuine is a nucleobase mostly synthesized by certain pathogenic bacteria, such as Clostridioides difficile and Chlamydia trachomatis, to promote their virulence. Queuine is incorporated into specific transfer RNAs (tRNAs) which drive the expression of genes involved in cell proliferation and migration of breast cancer cells [179,180]. Furthermore, recent studies report anti-tumor effects of bacterial metabolite trimethylamine N-oxide (TMAO) produced by a group of commensal bacteria, such as Clostridia, Bifidobacteria, and Coriobacteria. TMAO could promote tumor cell killing activities of CD8+ T cells and M1-type macrophages. Analysis of clinical tumor samples found that TNBC tumors abundant in Clostridiales are enriched in TMAO and exhibit activated immune microenvironment [181].
9. Breast Tumor-Associated Bacteria
Different levels of bacterial metabolites in normal vs. cancerous breast tissues discussed above are largely attributed to differences in microbial compositions. Decreased ratios of Sphingomonas yanoikuyae to Methylobacterium radiotolerans in the breast tissues are linked to elevated breast cancer risks [106]. Lactobacillus, Staphylococcus, and Enterobacteriaceae are more abundant in tumor-adjacent normal breast tissues compared to healthy breast tissues, indicating their contributions to neoplastic processes [47]. These different tumor-associated bacteria play differential roles in the development of breast cancer. Pro- and ant-tumor roles of select commensal bacteria are discussed below.
9.1. Origin of Breast-Tumor Resident Bacteria
Breast tumor-resident bacteria are proposed to be derived from the similar origins as those in normal breast tissues, namely, breast skin, the oral cavity of the suckling infant, maternal gut through gut-breast axis, and maternal oral cavity through oro-breast axis [182]. However, how these bacteria have traveled to distant tumors remains largely unknown. Bacterial strains found in tumors are mostly present in the gut microbiome, supporting the possibility of gut-breast axis [183]. Oral microbiota is also one of the potential sources of breast tumor resident bacteria [184]. In particular, Fusobacterium nucleatum, a major human oral bacterium, is commonly found in breast tumor cells [30], while it is rarely found in the intestine and thus is expected to reach tumors through the circulatory system [185]. Furthermore, a study by Nejman, et al. showed that bacteria in tumor-adjacent normal breast tissues had intermediate compositions between those of breast tumors and normal tissues [26]. This indicates that there are bacterial transfers between neighboring tissues that result in heterogeneity within tumors.
9.2. Major Breast-Tumor Resident Bacterial Species
There are several major bacterial species frequently found in breast tumor samples.
9.2.1. Fusobacterium nucleatum
Fusobacterium nucleatum is a common opportunistic bacterium in the oral cavity and is a potential causative agent of periodontitis and oral carcinomas [186]. This bacterium is also elevated in various types of solid tumors, especially in colorectal tumors, compared to the matched healthy tissues [187]. It is also associated with liver metastasis, indicating the broad spread of this oral pathogen [75]. F. nucleatum localizes at tumor sites by attaching to the cell surface galactose-N-acetylgalactosamine (Gal-GalNAc) through its lectin Fap2 [29,174]. In particular, intravascularly injected Fap2-expressing F. nucleatum strain ATCC 23726 specifically colonizes mammary tumors in mice, whereas Fap2-deficient bacteria fail to do so. Furthermore, F. nucleatum secretes an amyloid-like filament FadA which not only helps the attachment and invasion of the bacterium to host cells [188], but also serves as the scaffold of biofilm formation and promotes cancer progression [189]. Within tumor cells F. nucleatum induces proinflammatory signaling through TNFα, NF-kB and IL-6/IL-8 pathways [190,191,192] and promotes tumor growth, epithelial-to-mesenchymal transition (EMT), metastasis, and therapy resistance, while also suppressing NK cell-mediated tumor cell killing and T cell infiltration into tumors [29,94].
9.2.2. Streptococcus
Streptococcus is an oral bacterium found to promote metastasis and colonization of metastasized breast cancer cells [74]. Streptococcus mutans is a gram positive bacterium associated with dental caries (cavities). This bacterium could invade endothelial cells through Toll-like receptor 2 that triggers the production of proinflammatory IL-6/IL-8 and monocyte chemoattractant protein-1 (MCP1). Inflamed endothelial cells elevate permeability of the blood vessel, leading to various systemic conditions. For example, intravenously injected S. mutans is shown to induce lung vascular inflammation (e.g., thrombosis) and promote breast cancer metastasis to the lungs [193,194]. In addition, S. cuniculiIn, originally isolated from the respiratory tract of wild animals [195], is shown to promote metastatic potential of tumor cells by reorganizing actin cytoskeletons to resist sheer stress during invasion [74]. In contrast, another strains of Streptococcus confer beneficial effects on breast cancer treatment. S. salivarius, an abundant probiotic bacterium found in breast milk, is shown to suppress breast cancer growth when applied ectopically. Similarly, S. pneumoniae, the bacterium responsible for pneumonia and lung cancer [196], produces endopeptidase O (PepO) virulence protein. Ectopic administration of PepO to mouse model of triple-negative breast cancer (TNBC) is shown to activate TLR2/4 in tumor-associated macrophages and suppresses breast tumor growth [197].
9.2.3. Staphylococcus, and Enterobacteriaceae
Staphylococcus and Enterobacteriaceae are intestinal bacteria that could induce DNA damage within host cells. Staphylococci produce a toxin alpha phenol-soluble modulins (PSMα) and specific lipoproteins (Lpls). PSMα could incur DNA damage, whereas Lpls dampen DNA damage repair signaling, compromising genomic integrity of the host cell [39]. Furthermore, S. aureus, the bacterium usually found in the upper respiratory tract and skin, lowers the immunogenicity of the TME by suppressing effector T cells and promoting regulatory T cells [198]. In addition, S. xylosus, a skin commensal, promotes metastatic potential of tumor cells by reorganizing actin cytoskeletons to resist fluid sheer stress (FSS) during invasion [74]. Enterobacteriaceae (e.g., E. coli and Salmonella) are mostly intestinal commensals, and systemic infection with these bacteria is a common complication in cancer patients [199]. Similar to Staphylococcus, Enterobacteriaceae, especially those harboring the polyketide synthase (pks) island, produce a genotoxin Colibactin that causes DNA double strand breaks [200]. Enterobacteriaceae also impairs the expression of p53 tumor suppressor upon DNA damage, contributing to genomic instability of the host cell [201]. Furthermore, Enterobacteriaceae-infected host tumor cells produce the bactericidal lysophosphatidylcholines, which is found to be elevated in breast tumors and promote tumor growth and metastasis [202,203].
9.3. Roles of Intracellular Microbes in Breast Tumor Initiation/Development
According to the International Agency for Research on Cancer (IARC), 18-20% of cancers are caused by biological carcinogens such as oncogenic viruses and bacteria [204]. The roles of these microbes in cancer initiation and development involve six major mechanisms: genome instability/mutation, epigenetic modification, chronic inflammation, immune evasion, metabolic regulation, and metastasis [205].
9.3.1. Genome Instability/Mutation
The induction of genomic instability and mutation is one of the major carcinogenic mechanisms of the microbes. Oncoviruses are one of the major breast tumor-causing agents, including human papilloma virus (HPV), mouse mammary tumor virus (MMTV), Epstein-Barr virus (EBV), and bovine leukemia virus (BLV) [206]. They integrate the viral genome into the host chromosome to induce genetic mutations, while oncoproteins are produced by the integrated viral genome. For example, the HPV E7 oncoprotein directly inhibits cGas-STING pathway involved in the expression of type I interferon and pro-inflammatory factors, leading to immune escape [207,208]. EBV LMP1 oncoprotein upregulates oncogenic signaling pathways, such as the NF-κB pathway, involved in cell proliferation [209], while MMTV oncovirus-infected cells escape apoptosis by activation of Src tyrosine kinase pathway [210].
Likewise, certain carcinogenic bacteria, such as pks+ Escherichia coli and Bacteroides fragilis, secrete carcinogenic toxins that induce DNA damage, which results in elevated tumor onset and mortality [211]. The toxic produced by Bacteroides fragilis also promotes the expression of the enzyme spermine oxidase producing reactive oxygen species (ROS) that causes DNA damage [212]. The oncobacterium Fusobacterium nucleatum secretes FadA, a key adhesin, that activates the E-cadherin/β-catenin pathway to upregulate checkpoint kinase 2 (CHK2), inducing DNA damage [213]. Fusobacterium nucleatum infection also downmodulates Ku70/p53 DNA damage repair pathway, exacerbating DNA double-strand breaks (DSBs) [214]. E. coli and Staphylococcus epidermis isolated from breast tumors could cause DSBs even in cervical cancer cells, demonstrating non-tissue-specific tumorigenicity [47]. Furthermore, H. pylori and E. coli expressing EspF effector protein could suppress DNA mismatch repair mechanisms, augmenting genome instability and tumorigenesis [211,215].
Bacterial metabolites could also induce DNA damage to promote tumor development. Breast tumor tissues contain the elevated levels β-glucuronidase, a carcinogenic enzyme [125,216], that generates reactive intermediates from 2-amino-3-methylimidazo [4,5-f]quinoline to induce DNA damage [217]. Furthermore, Streptococcus anginosus and Porphyromonas gingivalis can convert ethanol to acetaldehyde that could form DNA adducts or inhibit DNA repair enzymes, causing DNA damage [218,219].
9.3.2. Tumor Metastasis
Over the past decade, it has been unveiled that intratumoral bacteria play critical roles in tumor metastasis. The initial study by Bullman et al. reported that primary colorectal tumors and their metastases shared the same viable bacterial components and that these bacteria were able to promote tumor cell growth and survival [75]. Furthermore, recent studies demonstrated that these tumor-resident bacteria are in fact localized in the cytosol of tumor cells and transported to the metastatic sites by tumor cells. During tumor cell metastasis, the intracellular bacteria promote tumor cell survival by allowing them to overcome physical and biochemical hurdles of unfavorable environment through adaptations termed pro-metastatic processes [67,74,220]. During pro-metastatic adaptations, tumor cells acquire capabilities of breaking tissue boundaries, controlling the local environment, conferring immune suppression and resistance to mechanical stress, and remodulating tumor cell intrinsic properties, such as epithelial-to-mesenchymal transition (EMT), stemness, and adhesion (Figure 3) [220]. Heer are some examples of intratumoral bacteria playing role in pro-metastasis and metastasis of tumor cells.
Food bacteria Listeria, in particular, Listeria monocytogenes, is an intracellular pathogen found in decaying food [221]. L. monocytogenes has been long utilized to develop cancer vaccines because it induces potent innate and adaptive immunological responses [222]. However, this bacterium could also reside within breast tumor cells and promote the growth and metastasis of tumor cells, worsening prognosis of patients [223,224]. Intratumoral L. monocytogenes induces cytoskeletal reorganization of tumor cells through its actin nucleation protein ActA and promotes tumor cell survival under FSS in the circulation [224]. Furthermore, such actin nucleation also recruits the ubiquitin conjugating enzyme Ube2N that activates TAK1-p38 MAP kinase signaling that controls tumor cell metastasis [225,226]. Not only the pathogenic strain, but also a non-pathogenic strain L. fleischmannii resides within breast tumors and is strongly associated with elevated expression of EMT-associated genes [83].
Another tumor metastasis-associated bacterium is Fusobacterium nucleatum, an opportunistic bacterium usually found in the oral cavity, but also abundant in breast tumors [29]. Intratumoral F. nucleatum promotes tumor cell invasion and suppresses immunological response through several different mechanisms [29]. First, F. nucleatum produces a virulence factor FadA, an amyloid protein that helps binding of the pathogen to host cells [227]. FadA upregulates Mir4435-2HG, which then induces the expression of SNAIL1 triggering EMT of host cells [94]. Second, F. nucleatum elevates the expression of MMP-9 that degrades extracellular matrix to assist tumor cell invasion. Third, F. nucleatum upregulates the expression of an adhesion molecule ICAM1 through the ALPK1/NF-κB axis that promotes tumor cell adhesion to endothelial cells during intravasation [228]. Fourth, F. nucleatum induces the production of extracellular vesicles that promote the expression of TLR4 in neighboring tumor cells to help their growth and metastasis [229]. Fifth, F. nucleatum elevates the expression of immune checkpoint receptors, TIGIT and CEACAM1, that suppress immunological responses. Lastly, F. nucleatum directly invades and kills tumor-infiltrating lymphocytes, including NK cells and T cells [29].
The third tumor metastasis-associated bacterium is Bacteroides fragilis, an abundant commensal bacterium in the colon and breast tumors. B. fragilis produces an enterotoxin, B. fragilis toxin (BFT), a zinc-dependent metalloprotease commonly associated with inflammatory colon diseases. Intratumorally produced BFT could elevate the growth and metastatic potentials of breast tumor cells by inducing the expression of stem cell/EMT-associated genes such as Slug and Twist [108,230,231]. As the mechanisms of this phenomenon, it is shown that BFT induces the cleavage of E-cadherin on the surface of tumor cells, which then triggers nuclear localization of β-catenin and Notch effector NICD. Activation of both the Wnt and Notch signaling pathways greatly promotes stemness and metastasis of tumors [232].
Furthermore, Staphylococcus and Lactibacillus, commensal bacteria abundant in breast tumor cells, have been shown to translocate to the lungs along with metastasizing tumor cells. These intracellular bacteria inhibit RhoA/ROCK-induced contractility of tumor cells while being exposed to FSS, conferring protection against mechanical force-induced apoptosis of tumor cells during metastasis [74]. As a potential mechanism, the same group proposed the possible involvement of ADP-ribosyltransferase C3 exoenzyme produced by these bacteria. This enzyme inhibits Rho GTPases to counteract immune cell activities and is well studied as their virulence factor of select bacterial species, such as Staphylococcus and Bacilus [233,234].
10. Discussion
Cumulative evidence unveils that intratumoral microbes are not only the mere biomarkers for breast cancer phenotype and prognosis, but also the causes of breast cancer initiation and metastasis. Such roles of intratumoral microbes suggest that they could serve as potential targets for breast cancer treatment and prevention. Thus, the efficacy of the use antibiotic(s) in combination with chemotherapy has been tested for breast cancer treatment. However, these antibiotics are reported to show both positive and negative effects, depending on whether they target tumor-resident bacteria or intestinal microbes [235]. For example, a Phase II study of combining Moxifloxacin, a fourth-generation quinolone with broad-spectrum coverage of breast tumor-resident bacteria, with treatment of physician’s choice (TPC: capecitabine, eribulin, gemcitabine, paclitaxel, or nab-paclitaxel) reported a promising efficacy and well-tolerated toxicities in patients with metastatic breast cancer [236]. On the other hand, a retrospective study on 772 women with triple-negative breast cancer treated with antimicrobials along with standard cytotoxic chemotherapy found that these patients had overall poorer survivals than those without antimicrobials [237]. Such deleterious effects of antimicrobial use for breast cancer patients are largely due to intestinal microbiota disorder that impairs immune function and triggers a systemic inflammatory response [235]. One possible solution to such corundum is to repopulate beneficial bacterial flora by supplementing probiotics and prebiotics to cancer patients treated with or without antibiotics. Furthermore, more recent strategies include fecal matter transplant of health cohorts to patients with breast cancer resistant to standard chemotherapy. Therapeutic manipulation of tumor microbiome is an emerging research field which would revolutionize cancer therapy in the near future.
11. Conclusion
In summary, recent studies have unveiled that intratumoral microbes play critical roles in breast cancer development and metastasis and serve as potential targets for breast cancer treatment and prevention.
Author Contributions
Conceptualization: S.F.; Writing—Original Draft Preparation: S.F.; Writing—Review & Editing: S.F., and Funding Acquisition: S.F.
Funding
This work was supported by the startup fund from the Department of Medicine, MetroHealth Medical Center/Case Western Reserve University to SF; American Cancer Society Research Scholar Grant (RSG-18-238-01-CSM) to S.F; and National Cancer Institute Research Grants (R01CA248304 and R21CA288449) to SF.
Data availability statement
All data generated or analyzed during this study are included in this published article [and its supplementary information file].
Acknowledgments
The author would like to thank all the laboratory members in Furuta laboratory at MetroHealth Medical Center/Case Western Reserve University for constructive suggestions for the subject matter of the manuscript.
Conflicts of Interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Davenport, E.R.; Clark, A.G.; Ley, R.E. The Relationship Between the Human Genome and Microbiome Comes into View. Annu Rev Genet 2017, 51, 413–433. [Google Scholar] [CrossRef]
- Smith, K.; McCoy, K.D.; Macpherson, A.J. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin Immunol 2007, 19, 59–69. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Research 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Umesaki, Y.; Setoyama, H.; Matsumoto, S.; Okada, Y. Expansion of alpha beta T-cell receptor-bearing intestinal intraepithelial lymphocytes after microbial colonization in germ-free mice and its independence from thymus. Immunology 1993, 79, 32–37. [Google Scholar]
- Hapfelmeier, S.; Lawson, M.A.; Slack, E.; Kirundi, J.K.; Stoel, M.; Heikenwalder, M.; Cahenzli, J.; Velykoredko, Y.; Balmer, M.L.; Endt, K.; et al. Reversible microbial colonization of germ-free mice reveals the dynamics of IgA immune responses. Science 2010, 328, 1705–1709. [Google Scholar] [CrossRef]
- Ivanov, I.I; Frutos Rde, L.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008, 4, 337–349. [Google Scholar] [CrossRef]
- Di Modica, M.; Arlotta, V.; Sfondrini, L.; Tagliabue, E.; Triulzi, T. The Link Between the Microbiota and HER2+ Breast Cancer: The New Challenge of Precision Medicine. Front Oncol 2022, 12, 947188. [Google Scholar] [CrossRef]
- Tzeng, A.; Sangwan, N.; Jia, M.; Liu, C.-C.; Keslar, K.S.; Downs-Kelly, E.; Fairchild, R.L.; Al-Hilli, Z.; Grobmyer, S.R.; Eng, C. Human breast microbiome correlates with prognostic features and immunological signatures in breast cancer. Genome Medicine 2021, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Bodai, B.I.; Nakata, T.E. Breast Cancer: Lifestyle, the Human Gut Microbiota/Microbiome, and Survivorship. Perm J 2020, 24. [Google Scholar] [CrossRef] [PubMed]
- Di Modica, M.; Gargari, G.; Regondi, V.; Bonizzi, A.; Arioli, S.; Belmonte, B.; De Cecco, L.; Fasano, E.; Bianchi, F.; Bertolotti, A.; et al. Gut Microbiota Condition the Therapeutic Efficacy of Trastuzumab in HER2-Positive Breast Cancer. Cancer Res 2021, 81, 2195–2206. [Google Scholar] [CrossRef]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.A.; Knight, R. The microbiome and human cancer. Science 2021, 371. [Google Scholar] [CrossRef]
- Nagpal, R.; Yadav, H. Bacterial Translocation from the Gut to the Distant Organs: An Overview. Ann Nutr Metab 2017, 71 Suppl 1, 11–16. [Google Scholar] [CrossRef]
- Wang, K.; Nakano, K.; Naderi, N.; Bajaj-Elliott, M.; Mosahebi, A. Is the skin microbiota a modifiable risk factor for breast disease?: A systematic review. Breast 2021, 59, 279–285. [Google Scholar] [CrossRef]
- Selvamani, S.; Dailin, D.J.; Gupta, V.K.; Wahid, M.; Keat, H.C.; Natasya, K.H.; Malek, R.A.; Haque, S.; Sayyed, R.Z.; Abomoelak, B.; et al. An Insight into Probiotics Bio-Route: Translocation from the Mother’s Gut to the Mammary Gland. Applied Sciences 2021, 11, 7247. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Fernández, L.; Verhasselt, V. The Gut‒Breast Axis: Programming Health for Life. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Farache, J.; Koren, I.; Milo, I.; Gurevich, I.; Kim, K.W.; Zigmond, E.; Furtado, G.C.; Lira, S.A.; Shakhar, G. Luminal bacteria recruit CD103+ dendritic cells into the intestinal epithelium to sample bacterial antigens for presentation. Immunity 2013, 38, 581–595. [Google Scholar] [CrossRef]
- Soto-Pantoja, D.R.; Gaber, M.; Arnone, A.A.; Bronson, S.M.; Cruz-Diaz, N.; Wilson, A.S.; Clear, K.Y.J.; Ramirez, M.U.; Kucera, G.L.; Levine, E.A.; et al. Diet Alters Entero-Mammary Signaling to Regulate the Breast Microbiome and Tumorigenesis. Cancer Res 2021, 81, 3890–3904. [Google Scholar] [CrossRef]
- Zhang, J.; Xia, Y.; Sun, J. Breast and gut microbiome in health and cancer. Genes Dis 2021, 8, 581–589. [Google Scholar] [CrossRef]
- Flood, T.R.; Kuennen, M.R.; Blacker, S.D.; Myers, S.D.; Walker, E.F.; Lee, B.J. The effect of sex, menstrual cycle phase and oral contraceptive use on intestinal permeability and ex-vivo monocyte TNFα release following treatment with lipopolysaccharide and hyperthermia. Cytokine 2022, 158, 155991. [Google Scholar] [CrossRef] [PubMed]
- Atashgaran, V.; Wrin, J.; Barry, S.C.; Dasari, P.; Ingman, W.V. Dissecting the Biology of Menstrual Cycle-Associated Breast Cancer Risk. Front Oncol 2016, 6, 267. [Google Scholar] [CrossRef] [PubMed]
- Schaadt, N.S.; Alfonso, J.C.L.; Schönmeyer, R.; Grote, A.; Forestier, G.; Wemmert, C.; Krönke, N.; Stoeckelhuber, M.; Kreipe, H.H.; Hatzikirou, H.; et al. Image analysis of immune cell patterns in the human mammary gland during the menstrual cycle refines lymphocytic lobulitis. Breast Cancer Res Treat 2017, 164, 305–315. [Google Scholar] [CrossRef]
- Younes, J.A.; Lievens, E.; Hummelen, R.; van der Westen, R.; Reid, G.; Petrova, M.I. Women and Their Microbes: The Unexpected Friendship. Trends in Microbiology 2018, 26, 16–32. [Google Scholar] [CrossRef]
- Hieken, T.J.; Chen, J.; Hoskin, T.L.; Walther-Antonio, M.; Johnson, S.; Ramaker, S.; Xiao, J.; Radisky, D.C.; Knutson, K.L.; Kalari, K.R.; et al. The Microbiome of Aseptically Collected Human Breast Tissue in Benign and Malignant Disease. Scientific Reports 2016, 6, 30751. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.W.; Zhang, H.; Zhang, X.; Xin, P.F.; Gao, X.; Li, H.R.; Zhou, C.Y.; Gao, W.M.; Kou, X.X.; Zhang, J.G. Liver Microbiome in Healthy Rats: The Hidden Inhabitants of Hepatocytes. Cellular Microbiology 2023, 2023, 7369034. [Google Scholar] [CrossRef]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef]
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O’Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of Human Breast Tissue. Applied and Environmental Microbiology 2014, 80, 3007–3014. [Google Scholar] [CrossRef]
- Lawani-Luwaji, E.U.; Alade, T. Sphingomonadaceae: Protective against breast cancer? Bulletin of the National Research Centre 2020, 44, 191. [Google Scholar] [CrossRef]
- Parhi, L.; Alon-Maimon, T.; Sol, A.; Nejman, D.; Shhadeh, A.; Fainsod-Levi, T.; Yajuk, O.; Isaacson, B.; Abed, J.; Maalouf, N.; et al. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat Commun 2020, 11, 3259. [Google Scholar] [CrossRef]
- Little, A.; Tangney, M.; Tunney, M.M.; Buckley, N.E. Fusobacterium nucleatum: a novel immune modulator in breast cancer? Expert Rev Mol Med 2023, 25, e15. [Google Scholar] [CrossRef] [PubMed]
- Brook, I. Chapter 173—Anaerobic bacteria. In Infectious Diseases (Third Edition), Cohen, J., Opal, S.M., Powderly, W.G., Eds.; Mosby: London, 2010; pp. 1757–1776. [Google Scholar]
- Allali, I.; Delgado, S.; Marron, P.I.; Astudillo, A.; Yeh, J.J.; Ghazal, H.; Amzazi, S.; Keku, T.; Azcarate-Peril, M.A. Gut microbiome compositional and functional differences between tumor and non-tumor adjacent tissues from cohorts from the US and Spain. Gut Microbes 2015, 6, 161–172. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Kwon, H.; Kim, Y.J. The Firmicutes/Bacteroidetes Ratio as a Risk Factor of Breast Cancer. J Clin Med 2023, 12. [Google Scholar] [CrossRef]
- Hieken, T.J.; Chen, J.; Chen, B.; Johnson, S.; Hoskin, T.L.; Degnim, A.C.; Walther-Antonio, M.R.; Chia, N. The breast tissue microbiome, stroma, immune cells and breast cancer. Neoplasia 2022, 27, 100786. [Google Scholar] [CrossRef] [PubMed]
- German, R.; Marino, N.; Hemmerich, C.; Podicheti, R.; Rusch, D.B.; Stiemsma, L.T.; Gao, H.; Xuei, X.; Rockey, P.; Storniolo, A.M. Exploring breast tissue microbial composition and the association with breast cancer risk factors. Breast Cancer Res 2023, 25, 82. [Google Scholar] [CrossRef]
- Yao, Z.; Ma, Q.; Cai, Z.; Raza, M.F.; Bai, S.; Wang, Y.; Zhang, P.; Ma, H.; Zhang, H. Similar Shift Patterns in Gut Bacterial and Fungal Communities Across the Life Stages of Bactrocera minax Larvae From Two Field Populations. Front Microbiol 2019, 10, 2262. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; McMillan, A.; Angelini, M.; Gloor, G.B.; Sumarah, M.; Burton, J.P.; Reid, G. Effect of chemotherapy on the microbiota and metabolome of human milk, a case report. Microbiome 2014, 2, 24. [Google Scholar] [CrossRef]
- Boopathi, S.; Priya, P.S.; Haridevamuthu, B.; Nayak, S.P.R.R.; Chandrasekar, M.; Arockiaraj, J.; Jia, A.-Q. Expanding germ-organ theory: Understanding non-communicable diseases through enterobacterial translocation. Pharmacological Research 2023, 194, 106856. [Google Scholar] [CrossRef]
- Deplanche, M.; Mouhali, N.; Nguyen, M.-T.; Cauty, C.; Ezan, F.; Diot, A.; Raulin, L.; Dutertre, S.; Langouet, S.; Legembre, P.; et al. Staphylococcus aureus induces DNA damage in host cell. Scientific Reports 2019, 9, 7694. [Google Scholar] [CrossRef] [PubMed]
- Klann, E.; Williamson, J.M.; Tagliamonte, M.S.; Ukhanova, M.; Asirvatham, J.R.; Chim, H.; Yaghjyan, L.; Mai, V. Microbiota composition in bilateral healthy breast tissue and breast tumors. Cancer Causes Control 2020, 31, 1027–1038. [Google Scholar] [CrossRef]
- Wahid, M.; Dar, S.A.; Jawed, A.; Mandal, R.K.; Akhter, N.; Khan, S.; Khan, F.; Jogaiah, S.; Rai, A.K.; Rattan, R. Microbes in gynecologic cancers: Causes or consequences and therapeutic potential. Seminars in Cancer Biology 2022, 86, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Nakajima-Adachi, H.; Tamai, M.; Nakanishi, H.; Hachimura, S. Extracts of Gluconacetobacter hansenii GK-1 induce Foxp3(+)T cells in food-allergic mice by an IL-4-dependent or IL-4-independent mechanism. Biosci Microbiota Food Health 2022, 41, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Lilian, M.; Rawlynce, B.; Charles, G.; Felix, K. Potential role of rumen bacteria in modulating milk production and composition of admixed dairy cows. Letters in Applied Microbiology 2023, 76. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, M.; Menakha, M.; Vijayakumar, S. Cryptophycin F—A potential cyanobacterial drug for breast cancer. Biomedicine & Aging Pathology 2014, 4, 229–234. [Google Scholar] [CrossRef]
- Yang, Y.; Fukui, R.; Jia, H.; Kato, H. Amaranth Supplementation Improves Hepatic Lipid Dysmetabolism and Modulates Gut Microbiota in Mice Fed a High-Fat Diet. Foods 2021, 10. [Google Scholar] [CrossRef]
- Milton-Laskibar, I.; Cuevas-Sierra, A.; Portillo, M.P.; Martínez, J.A. Effects of Resveratrol Administration in Liver Injury Prevention as Induced by an Obesogenic Diet: Role of Ruminococcaceae. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The Microbiota of Breast Tissue and Its Association with Breast Cancer. Appl Environ Microbiol 2016, 82, 5039–5048. [Google Scholar] [CrossRef]
- Bindels, L.B.; Porporato, P.; Dewulf, E.M.; Verrax, J.; Neyrinck, A.M.; Martin, J.C.; Scott, K.P.; Buc Calderon, P.; Feron, O.; Muccioli, G.G.; et al. Gut microbiota-derived propionate reduces cancer cell proliferation in the liver. British Journal of Cancer 2012, 107, 1337–1344. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, K.; Wei, J.; Ding, Y.; Wang, X.; Hou, H.; Wu, J.; Liu, T.; Wang, B.; Cao, H. Gut microbiota-derived short-chain fatty acids regulate gastrointestinal tumor immunity: a novel therapeutic strategy? Front Immunol 2023, 14, 1158200. [Google Scholar] [CrossRef]
- Derqaoui, S.; Oukessou, M.; Attrassi, K.; Elftouhy, F.Z.; Nassik, S. Detection of Sutterella spp. in Broiler Liver and Breast. Front Vet Sci 2022, 9, 859902. [Google Scholar] [CrossRef]
- Gutierrez-Orozco, F.; Thomas-Ahner, J.M.; Galley, J.D.; Bailey, M.T.; Clinton, S.K.; Lesinski, G.B.; Failla, M.L. Intestinal microbial dysbiosis and colonic epithelial cell hyperproliferation by dietary α-mangostin is independent of mouse strain. Nutrients 2015, 7, 764–784. [Google Scholar] [CrossRef] [PubMed]
- Thu, M.S.; Chotirosniramit, K.; Nopsopon, T.; Hirankarn, N.; Pongpirul, K. Human gut, breast, and oral microbiome in breast cancer: A systematic review and meta-analysis. Front Oncol 2023, 13, 1144021. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, F.; Li, P. Intratumor microbiota in cancer pathogenesis and immunity: from mechanisms of action to therapeutic opportunities. Front Immunol 2023, 14, 1269054. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yu, B.; Rao, B.; Sun, Y.; Yu, J.; Wang, D.; Cui, G.; Ren, Z. The effect of the intratumoral microbiome on tumor occurrence, progression, prognosis and treatment. Front Immunol 2022, 13, 1051987. [Google Scholar] [CrossRef]
- Li, Y.; Ren, L.; Wang, Y.; Li, J.; Zhou, Q.; Peng, C.; Li, Y.; Cheng, R.; He, F.; Shen, X. The Effect of Breast Milk Microbiota on the Composition of Infant Gut Microbiota: A Cohort Study. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Dieleman, S.; Aarnoutse, R.; Ziemons, J.; Kooreman, L.; Boleij, A.; Smidt, M. Exploring the Potential of Breast Microbiota as Biomarker for Breast Cancer and Therapeutic Response. The American Journal of Pathology 2021, 191, 968–982. [Google Scholar] [CrossRef]
- Fernández, L.; Langa, S.; Martín, V.; Maldonado, A.; Jiménez, E.; Martín, R.; Rodríguez, J.M. The human milk microbiota: Origin and potential roles in health and disease. Pharmacological Research 2013, 69, 1–10. [Google Scholar] [CrossRef]
- Gomez-Gallego, C.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Seminars in Fetal and Neonatal Medicine 2016, 21, 400–405. [Google Scholar] [CrossRef]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Scientific Reports 2016, 6, 23129. [Google Scholar] [CrossRef] [PubMed]
- Favier, C.F.; Vaughan, E.E.; Vos, W.M.D.; Akkermans, A.D.L. Molecular Monitoring of Succession of Bacterial Communities in Human Neonates. Applied and Environmental Microbiology 2002, 68, 219–226. [Google Scholar] [CrossRef]
- Macia, L.; Mackay, C.R. Dysfunctional microbiota with reduced capacity to produce butyrate as a basis for allergic diseases. Journal of Allergy and Clinical Immunology 2019, 144, 1513–1515. [Google Scholar] [CrossRef]
- Heikkilä, M.P.; Saris, P.E.J. Inhibition of Staphylococcus aureus by the commensal bacteria of human milk. Journal of Applied Microbiology 2003, 95, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Sela, D.A.; Mills, D.A. Nursing our microbiota: molecular linkages between bifidobacteria and milk oligosaccharides. Trends in Microbiology 2010, 18, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Benito, D.; Lozano, C.; Jiménez, E.; Albújar, M.; Gómez, A.; Rodríguez, J.M.; Torres, C. Characterization of Staphylococcus aureus strains isolated from faeces of healthy neonates and potential mother-to-infant microbial transmission through breastfeeding. FEMS Microbiology Ecology 2015, 91. [Google Scholar] [CrossRef]
- Martín, V.; Maldonado-Barragán, A.; Moles, L.; Rodriguez-Baños, M.; Campo, R.d.; Fernández, L.; Rodríguez, J.M.; Jiménez, E. Sharing of Bacterial Strains Between Breast Milk and Infant Feces. Journal of Human Lactation 2012, 28, 36–44. [Google Scholar] [CrossRef]
- Prentice, P.M.; Schoemaker, M.H.; Vervoort, J.; Hettinga, K.; Lambers, T.T.; van Tol, E.A.F.; Acerini, C.L.; Olga, L.; Petry, C.J.; Hughes, I.A.; et al. Human Milk Short-Chain Fatty Acid Composition is Associated with Adiposity Outcomes in Infants. The Journal of Nutrition 2019, 149, 716–722. [Google Scholar] [CrossRef]
- Liu, J.; Luo, F.; Wen, L.; Zhao, Z.; Sun, H. Current Understanding of Microbiomes in Cancer Metastasis. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Jiang, Z.; Wei, C.; Ma, Y.; Zhao, J.; Wang, F.; Zhao, B.; Wang, D.; Tang, D. Intratumoural microbiota: from theory to clinical application. Cell Communication and Signaling 2023, 21, 164. [Google Scholar] [CrossRef]
- Kim, W.; Kim, E.; Min, H.; Kim, M.G.; Eisenbeis, V.B.; Dutta, A.K.; Pavlovic, I.; Jessen, H.J.; Kim, S.; Seong, R.H. Inositol polyphosphates promote T cell-independent humoral immunity via the regulation of Bruton’s tyrosine kinase. Proc Natl Acad Sci U S A 2019, 116, 12952–12957. [Google Scholar] [CrossRef]
- Weinberg, S.E.; Sun, L.Y.; Yang, A.L.; Liao, J.; Yang, G.Y. Overview of Inositol and Inositol Phosphates on Chemoprevention of Colitis-Induced Carcinogenesis. Molecules 2021, 26, 31. [Google Scholar] [CrossRef]
- Marth, J.D.; Grewal, P.K. Mammalian glycosylation in immunity. Nat Rev Immunol 2008, 8, 874–887. [Google Scholar] [CrossRef]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.A.; Ren, W.; Yin, Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Colbert, L.E.; El Alam, M.B.; Wang, R.; Karpinets, T.; Lo, D.; Lynn, E.J.; Harris, T.A.; Elnaggar, J.H.; Yoshida-Court, K.; Tomasic, K.; et al. Tumor-resident Lactobacillus iners confer chemoradiation resistance through lactate-induced metabolic rewiring. Cancer Cell 2023, 41, 1945–1962.e1911. [Google Scholar] [CrossRef]
- Fu, A.; Yao, B.; Dong, T.; Chen, Y.; Yao, J.; Liu, Y.; Li, H.; Bai, H.; Liu, X.; Zhang, Y.; et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell 2022, 185, 1356–1372.e1326. [Google Scholar] [CrossRef] [PubMed]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Skibinski, A.; Kuperwasser, C. The origin of breast tumor heterogeneity. Oncogene 2015, 34, 5309–5316. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, V.; Iorio, E.; Miodini, P.; Silvestri, M.; Dugo, M.; Daidone, M.G. Metabolic Footprints and Molecular Subtypes in Breast Cancer. Dis Markers 2017, 2017, 7687851. [Google Scholar] [CrossRef]
- Smith, A.; Pierre, J.F.; Makowski, L.; Tolley, E.; Lyn-Cook, B.; Lu, L.; Vidal, G.; Starlard-Davenport, A. Distinct microbial communities that differ by race, stage, or breast-tumor subtype in breast tissues of non-Hispanic Black and non-Hispanic White women. Sci Rep 2019, 9, 11940. [Google Scholar] [CrossRef]
- Wang, H.; Altemus, J.; Niazi, F.; Green, H.; Calhoun, B.C.; Sturgis, C.; Grobmyer, S.R.; Eng, C. Breast tissue, oral and urinary microbiomes in breast cancer. Oncotarget 2017, 8, 88122–88138. [Google Scholar] [CrossRef]
- Hadzega, D.; Minarik, G.; Karaba, M.; Kalavska, K.; Benca, J.; Ciernikova, S.; Sedlackova, T.; Nemcova, P.; Bohac, M.; Pindak, D.; et al. Uncovering Microbial Composition in Human Breast Cancer Primary Tumour Tissue Using Transcriptomic RNA-seq. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Tzeng, A.; Sangwan, N.; Jia, M.; Liu, C.C.; Keslar, K.S.; Downs-Kelly, E.; Fairchild, R.L.; Al-Hilli, Z.; Grobmyer, S.R.; Eng, C. Human breast microbiome correlates with prognostic features and immunological signatures in breast cancer. Genome Med 2021, 13, 60. [Google Scholar] [CrossRef]
- Banerjee, S.; Wei, Z.; Tan, F.; Peck, K.N.; Shih, N.; Feldman, M.; Rebbeck, T.R.; Alwine, J.C.; Robertson, E.S. Distinct microbiological signatures associated with triple negative breast cancer. Sci Rep 2015, 5, 15162. [Google Scholar] [CrossRef]
- Thompson, K.J.; Ingle, J.N.; Tang, X.; Chia, N.; Jeraldo, P.R.; Walther-Antonio, M.R.; Kandimalla, K.K.; Johnson, S.; Yao, J.Z.; Harrington, S.C.; et al. A comprehensive analysis of breast cancer microbiota and host gene expression. PLoS One 2017, 12, e0188873. [Google Scholar] [CrossRef] [PubMed]
- Chng, K.R.; Chan, S.H.; Ng, A.H.Q.; Li, C.; Jusakul, A.; Bertrand, D.; Wilm, A.; Choo, S.P.; Tan, D.M.Y.; Lim, K.H.; et al. Tissue Microbiome Profiling Identifies an Enrichment of Specific Enteric Bacteria in Opisthorchis viverrini Associated Cholangiocarcinoma. EBioMedicine 2016, 8, 195–202. [Google Scholar] [CrossRef]
- Audirac-Chalifour, A.; Torres-Poveda, K.; Bahena-Román, M.; Téllez-Sosa, J.; Martínez-Barnetche, J.; Cortina-Ceballos, B.; López-Estrada, G.; Delgado-Romero, K.; Burguete-García, A.I.; Cantú, D.; et al. Cervical Microbiome and Cytokine Profile at Various Stages of Cervical Cancer: A Pilot Study. PLoS One 2016, 11, e0153274. [Google Scholar] [CrossRef]
- Łaniewski, P.; Barnes, D.; Goulder, A.; Cui, H.; Roe, D.J.; Chase, D.M.; Herbst-Kralovetz, M.M. Linking cervicovaginal immune signatures, HPV and microbiota composition in cervical carcinogenesis in non-Hispanic and Hispanic women. Sci Rep 2018, 8, 7593. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; Fathi, P.; Craig, J.M.; Boleij, A.; Taddese, R.; Geis, A.L.; Wu, X.; DeStefano Shields, C.E.; Hechenbleikner, E.M.; Huso, D.L.; et al. Patients with familial adenomatous polyposis harbor colonic biofilms containing tumorigenic bacteria. Science 2018, 359, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Guo, L.; Liu, J.J.; Zhao, H.P.; Zhang, J.; Wang, J.H. Alteration of the esophageal microbiota in Barrett’s esophagus and esophageal adenocarcinoma. World J Gastroenterol 2019, 25, 2149–2161. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Man, S.M.; Mitchell, H.M. Is Campylobacter to esophageal adenocarcinoma as Helicobacter is to gastric adenocarcinoma? Trends Microbiol 2015, 23, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Li, S.; Ma, Z.; Liang, S.; Shan, T.; Zhang, M.; Zhu, X.; Zhang, P.; Liu, G.; Zhou, F.; et al. Presence of Porphyromonas gingivalis in esophagus and its association with the clinicopathological characteristics and survival in patients with esophageal cancer. Infect Agent Cancer 2016, 11, 3. [Google Scholar] [CrossRef]
- Avilés-Jiménez, F.; Guitron, A.; Segura-López, F.; Méndez-Tenorio, A.; Iwai, S.; Hernández-Guerrero, A.; Torres, J. Microbiota studies in the bile duct strongly suggest a role for <em>Helicobacter pylori</em> in extrahepatic cholangiocarcinoma. Clinical Microbiology and Infection 2016, 22, 178.e111–178.e122. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Loza, E.; Villa-Gomez, G.; Trujillo, C.C.; Baez, S.; Asai, T.; Ikoma, T.; Endoh, K.; Nakamura, K. Metagenomics of Microbial Communities in Gallbladder Bile from Patients with Gallbladder Cancer or Cholelithiasis. Asian Pac J Cancer Prev 2018, 19, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Buti, L.; Spooner, E.; Van der Veen, A.G.; Rappuoli, R.; Covacci, A.; Ploegh, H.L. Helicobacter pylori cytotoxin-associated gene A (CagA) subverts the apoptosis-stimulating protein of p53 (ASPP2) tumor suppressor pathway of the host. Proc Natl Acad Sci U S A 2011, 108, 9238–9243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, C.; Liu, J.; Geng, F.; Shi, X.; Li, Q.; Lu, Z.; Pan, Y. Fusobacterium nucleatum promotes epithelial-mesenchymal transiton through regulation of the lncRNA MIR4435-2HG/miR-296-5p/Akt2/SNAI1 signaling pathway. Febs j 2020, 287, 4032–4047. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.; Avenaud, P.; Ménard, A.; Le Bail, B.; Balabaud, C.; Bioulac-Sage, P.; de Magalhães Queiroz, D.M.; Mégraud, F. Association of Helicobacter species with hepatitis C cirrhosis with or without hepatocellular carcinoma. Gut 2005, 54, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; von Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the microbiome and TP53 in human lung cancer. Genome Biology 2018, 19, 123. [Google Scholar] [CrossRef]
- Yu, G.; Gail, M.H.; Consonni, D.; Carugno, M.; Humphrys, M.; Pesatori, A.C.; Caporaso, N.E.; Goedert, J.J.; Ravel, J.; Landi, M.T. Characterizing human lung tissue microbiota and its relationship to epidemiological and clinical features. Genome Biology 2016, 17, 163. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.L.; Kuczynski, J.; Bhattacharya, A.; Huey, B.; Corby, P.M.; Queiroz, E.L.; Nightingale, K.; Kerr, A.R.; DeLacure, M.D.; Veeramachaneni, R.; et al. Changes in abundance of oral microbiota associated with oral cancer. PLoS One 2014, 9, e98741. [Google Scholar] [CrossRef]
- Chan, P.J.; Seraj, I.M.; Kalugdan, T.H.; King, A. Prevalence of mycoplasma conserved DNA in malignant ovarian cancer detected using sensitive PCR-ELISA. Gynecol Oncol 1996, 63, 258–260. [Google Scholar] [CrossRef]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov 2018, 8, 403–416. [Google Scholar] [CrossRef]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Cavarretta, I.; Ferrarese, R.; Cazzaniga, W.; Saita, D.; Lucianò, R.; Ceresola, E.R.; Locatelli, I.; Visconti, L.; Lavorgna, G.; Briganti, A.; et al. The Microbiome of the Prostate Tumor Microenvironment. Eur Urol 2017, 72, 625–631. [Google Scholar] [CrossRef]
- Cohen, R.J.; Shannon, B.A.; McNeal, J.E.; Shannon, T.; Garrett, K.L. Propionibacterium acnes associated with inflammation in radical prostatectomy specimens: a possible link to cancer evolution? J Urol 2005, 173, 1969–1974. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Baba, Y.; Ishimoto, T.; Tsutsuki, H.; Zhang, T.; Nomoto, D.; Okadome, K.; Yamamura, K.; Harada, K.; Eto, K.; et al. Fusobacterium nucleatum confers chemoresistance by modulating autophagy in oesophageal squamous cell carcinoma. Br J Cancer 2021, 124, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Xuan, C.; Shamonki, J.M.; Chung, A.; Dinome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS One 2014, 9, e83744. [Google Scholar] [CrossRef]
- Bernardo, G.; Le Noci, V.; Di Modica, M.; Montanari, E.; Triulzi, T.; Pupa, S.M.; Tagliabue, E.; Sommariva, M.; Sfondrini, L. The Emerging Role of the Microbiota in Breast Cancer Progression. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Siddharth, S.; Xia, Y.; Sharma, D. Concomitant analyses of intratumoral microbiota and genomic features reveal distinct racial differences in breast cancer. npj Breast Cancer 2023, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, S.; Zhang, Y.; Thapa, S.; Allen, M.S.; Phillips, N.; Chaudhary, P.; Kashyap, M.V.; Vishwanatha, J.K. Comparative analysis of racial differences in breast tumor microbiome. Sci Rep 2020, 10, 14116. [Google Scholar] [CrossRef]
- Yuan, Y.; Leeds, J.A.; Meredith, T.C. Pseudomonas aeruginosa directly shunts β-oxidation degradation intermediates into de novo fatty acid biosynthesis. J Bacteriol 2012, 194, 5185–5196. [Google Scholar] [CrossRef]
- Park, S.M.; Choi, W.S.; Yoon, Y.; Jung, G.H.; Lee, C.K.; Ahn, S.H.; Wonsuck, Y.; Yoo, Y. Breast abscess caused by Staphylococcus aureus in 2 adolescent girls with atopic dermatitis. Korean J Pediatr 2018, 61, 200–204. [Google Scholar] [CrossRef]
- Chiba, A.; Bawaneh, A.; Velazquez, C.; Clear, K.Y.J.; Wilson, A.S.; Howard-McNatt, M.; Levine, E.A.; Levi-Polyachenko, N.; Yates-Alston, S.A.; Diggle, S.P.; et al. Neoadjuvant Chemotherapy Shifts Breast Tumor Microbiota Populations to Regulate Drug Responsiveness and the Development of Metastasis. Molecular Cancer Research 2020, 18, 130–139. [Google Scholar] [CrossRef]
- Gotland, N.; Uhre, M.L.; Sandholdt, H.; Mejer, N.; Lundbo, L.F.; Petersen, A.; Larsen, A.R.; Benfield, T. Increased risk of incident primary cancer after Staphylococcus aureus bacteremia: A matched cohort study. Medicine 2020, 99, e19984. [Google Scholar] [CrossRef] [PubMed]
- Kullander, J.; Forslund, O.; Dillner, J. Staphylococcus aureus and squamous cell carcinoma of the skin. Cancer Epidemiol Biomarkers Prev 2009, 18, 472–478. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nature Reviews Microbiology 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Yu, H.-j.; Deng, H.; Ma, J.; Huang, S.-j.; Yang, J.-m.; Huang, Y.-f.; Mu, X.-p.; Zhang, L.; Wang, Q. Clinical metagenomic analysis of bacterial communities in breast abscesses of granulomatous mastitis. International Journal of Infectious Diseases 2016, 53, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Li, Z.; Lin, X.; Li, F.; Xu, H.; Yu, X.; Liu, L.; Liang, Y.; Xu, Z.; Wang, J.; et al. Etiology of granulomatous lobular mastitis based on metagenomic next-generation sequencing. International Journal of Infectious Diseases 2021, 113, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, J.; Carvajal, M.; Hernández, G. Back to Basics: Could the Preoperative Skin Antiseptic Agent Help Prevent Biofilm-Related Capsular Contracture? Aesthet Surg J 2019, 39, 848–859. [Google Scholar] [CrossRef] [PubMed]
- Bachour, Y.; Poort, L.; Verweij, S.P.; van Selms, G.; Winters, H.A.H.; Ritt, M.J.P.F.; Niessen, F.B.; Budding, A.E. PCR Characterization of Microbiota on Contracted and Non-Contracted Breast Capsules. Aesthetic Plastic Surgery 2019, 43, 918–926. [Google Scholar] [CrossRef]
- Rieger, U.M.; Mesina, J.; Kalbermatten, D.F.; Haug, M.; Frey, H.P.; Pico, R.; Frei, R.; Pierer, G.; Lüscher, N.J.; Trampuz, A. Bacterial biofilms and capsular contracture in patients with breast implants. British Journal of Surgery 2013, 100, 768–774. [Google Scholar] [CrossRef]
- Hu, H.; Johani, K.; Almatroudi, A.; Vickery, K.; Van Natta, B.; Kadin, M.E.; Brody, G.; Clemens, M.; Cheah, C.Y.; Lade, S.; et al. Bacterial Biofilm Infection Detected in Breast Implant–Associated Anaplastic Large-Cell Lymphoma. Plastic and Reconstructive Surgery 2016, 137. [Google Scholar] [CrossRef]
- Walker, J.N.; Hanson, B.M.; Pinkner, C.L.; Simar, S.R.; Pinkner, J.S.; Parikh, R.; Clemens, M.W.; Hultgren, S.J.; Myckatyn, T.M. Insights into the Microbiome of Breast Implants and Periprosthetic Tissue in Breast Implant-Associated Anaplastic Large Cell Lymphoma. Scientific Reports 2019, 9, 10393. [Google Scholar] [CrossRef] [PubMed]
- Love, S.M.; Barsky, S.H. Anatomy of the nipple and breast ducts revisited. Cancer 2004, 101, 1947–1957. [Google Scholar] [CrossRef]
- West, P.A.; Hewitt, J.H.; Murphy, O.M. Influence of methods of collection and storage on the bacteriology of human milk. The Journal of applied bacteriology 1979, 46 2, 269–277. [Google Scholar] [CrossRef]
- Chan, A.A.; Bashir, M.; Rivas, M.N.; Duvall, K.; Sieling, P.A.; Pieber, T.R.; Vaishampayan, P.A.; Love, S.M.; Lee, D.J. Characterization of the microbiome of nipple aspirate fluid of breast cancer survivors. Scientific Reports 2016, 6, 28061. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, D.T.; Kent, J.C.; Owens, R.A.; Hartmann, P.E. Ultrasound Imaging of Milk Ejection in the Breast of Lactating Women. Pediatrics 2004, 113, 361–367. [Google Scholar] [CrossRef]
- Berg, R.D. Bacterial translocation from the gastrointestinal tract. Adv Exp Med Biol 1999, 473, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Sedman, P.C.; Macfie, J.; Sagar, P.; Mitchell, C.J.; May, J.; Mancey-Jones, B.; Johnstone, D. The prevalence of gut translocation in humans. Gastroenterology 1994, 107, 643–649. [Google Scholar] [CrossRef]
- Rodriguez, A.V.; Baigorí, M.D.; Alvarez, S.; Castro, G.R.; Oliver, G. Phosphatidylinositol-specific phospholipase C activity in Lactobacillus rhamnosus with capacity to translocate. FEMS Microbiol Lett 2001, 204, 33–38. [Google Scholar] [CrossRef]
- Berg, R.D. Bacterial translocation from the gastrointestinal tract. Trends Microbiol 1995, 3, 149–154. [Google Scholar] [CrossRef]
- Perez, P.F.; Doré, J.l.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J.; Donnet-Hughes, A. Bacterial Imprinting of the Neonatal Immune System: Lessons From Maternal Cells? Pediatrics 2007, 119, e724–e732. [Google Scholar] [CrossRef]
- Cronin, M.; Morrissey, D.; Rajendran, S.; El Mashad, S.M.; van Sinderen, D.; O’Sullivan, G.C.; Tangney, M. Orally administered bifidobacteria as vehicles for delivery of agents to systemic tumors. Mol Ther 2010, 18, 1397–1407. [Google Scholar] [CrossRef]
- Danino, T.; Prindle, A.; Kwong, G.A.; Skalak, M.; Li, H.; Allen, K.; Hasty, J.; Bhatia, S.N. Programmable probiotics for detection of cancer in urine. Sci Transl Med 2015, 7, 289ra284. [Google Scholar] [CrossRef]
- Mira, A.; Rodríguez, J. Prebiotics and Probiotics in Human Milk. 2017. [Google Scholar]
- Brandtzaeg, P. The mucosal immune system and its integration with the mammary glands. J Pediatr 2010, 156, S8–15. [Google Scholar] [CrossRef]
- Goldman, A.S.; Goldblum, R.M. Transfer of maternal leukocytes to the infant by human milk. Curr Top Microbiol Immunol 1997, 222, 205–213. [Google Scholar] [CrossRef]
- Trend, S.; de Jong, E.; Lloyd, M.L.; Kok, C.H.; Richmond, P.; Doherty, D.A.; Simmer, K.; Kakulas, F.; Strunk, T.; Currie, A. Leukocyte Populations in Human Preterm and Term Breast Milk Identified by Multicolour Flow Cytometry. PLoS One 2015, 10, e0135580. [Google Scholar] [CrossRef]
- Tuaillon, E.; Valea, D.; Becquart, P.; Al Tabaa, Y.; Meda, N.; Bollore, K.; Van de Perre, P.; Vendrell, J.P. Human milk-derived B cells: a highly activated switched memory cell population primed to secrete antibodies. J Immunol 2009, 182, 7155–7162. [Google Scholar] [CrossRef]
- Cabinian, A.; Sinsimer, D.; Tang, M.; Zumba, O.; Mehta, H.; Toma, A.; Sant’Angelo, D.; Laouar, Y.; Laouar, A. Transfer of Maternal Immune Cells by Breastfeeding: Maternal Cytotoxic T Lymphocytes Present in Breast Milk Localize in the Peyer’s Patches of the Nursed Infant. PLoS One 2016, 11, e0156762. [Google Scholar] [CrossRef]
- Hammond, K.A. Adaptation of the maternal intestine during lactation. J Mammary Gland Biol Neoplasia 1997, 2, 243–252. [Google Scholar] [CrossRef]
- Gosalbes, M.J.; Compte, J.; Moriano-Gutierrez, S.; Vallès, Y.; Jiménez-Hernández, N.; Pons, X.; Artacho, A.; Francino, M.P. Metabolic adaptation in the human gut microbiota during pregnancy and the first year of life. EBioMedicine 2019, 39, 497–509. [Google Scholar] [CrossRef]
- Jiménez, E.; Fernández, L.; Maldonado, A.; Martín, R.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Oral Administration of <i>Lactobacillus</i> Strains Isolated from Breast Milk as an Alternative for the Treatment of Infectious Mastitis during Lactation. Applied and Environmental Microbiology 2008, 74, 4650–4655. [Google Scholar] [CrossRef]
- Arroyo, R.; Martín, V.; Maldonado, A.; Jiménez, E.; Fernández, L.; Rodríguez, J.M. Treatment of Infectious Mastitis during Lactation: Antibiotics versus Oral Administration of Lactobacilli Isolated from Breast Milk. Clinical Infectious Diseases 2010, 50, 1551–1558. [Google Scholar] [CrossRef]
- Moossavi, S.; Azad, M.B. Origins of human milk microbiota: new evidence and arising questions. Gut Microbes 2020, 12, 1667722. [Google Scholar] [CrossRef]
- Li, S.; He, M.; Lei, Y.; Liu, Y.; Li, X.; Xiang, X.; Wu, Q.; Wang, Q. Oral Microbiota and Tumor-A New Perspective of Tumor Pathogenesis. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Petris, C.K.; Golomb, M.; Phillips, T.E. Bacterial Transcytosis across Conjunctival M Cells. Investigative Ophthalmology & Visual Science 2007, 48, 2172–2177. [Google Scholar] [CrossRef]
- Neal, M.D.; Leaphart, C.; Levy, R.; Prince, J.; Billiar, T.R.; Watkins, S.; Li, J.; Cetin, S.; Ford, H.; Schreiber, A.; et al. Enterocyte TLR4 mediates phagocytosis and translocation of bacteria across the intestinal barrier. J Immunol 2006, 176, 3070–3079. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Moniri, N.H.; Ozawa, K.; Stamler, J.S.; Daaka, Y. Nitric oxide regulates endocytosis by S-nitrosylation of dynamin. Proc Natl Acad Sci U S A 2006, 103, 1295–1300. [Google Scholar] [CrossRef] [PubMed]
- Wells, C.L.; VandeWesterlo, E.M.; Jechorek, R.P.; Erlandsen, S.L. Effect of hypoxia on enterocyte endocytosis of enteric bacteria. Crit Care Med 1996, 24, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Keita Å, V.; Phan, V.; McKay, C.M.; Schoultz, I.; Lee, J.; Murphy, M.P.; Fernando, M.; Ronaghan, N.; Balce, D.; et al. Targeting mitochondria-derived reactive oxygen species to reduce epithelial barrier dysfunction and colitis. Am J Pathol 2014, 184, 2516–2527. [Google Scholar] [CrossRef]
- Lewis, K.; Lutgendorff, F.; Phan, V.; Söderholm, J.D.; Sherman, P.M.; McKay, D.M. Enhanced translocation of bacteria across metabolically stressed epithelia is reduced by butyrate. Inflamm Bowel Dis 2010, 16, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.; Hoare, C.; Tanianis-Hughes, J.; Carlson, G.L.; Warhurst, G. Interferon gamma induces translocation of commensal Escherichia coli across gut epithelial cells via a lipid raft-mediated process. Gastroenterology 2005, 128, 1258–1267. [Google Scholar] [CrossRef]
- Smyth, D.; McKay, C.M.; Gulbransen, B.D.; Phan, V.C.; Wang, A.; McKay, D.M. Interferon-gamma signals via an ERK1/2-ARF6 pathway to promote bacterial internalization by gut epithelia. Cell Microbiol 2012, 14, 1257–1270. [Google Scholar] [CrossRef]
- Chen, T.L.; Chen, S.; Wu, H.W.; Lee, T.C.; Lu, Y.Z.; Wu, L.L.; Ni, Y.H.; Sun, C.H.; Yu, W.H.; Buret, A.G.; et al. Persistent gut barrier damage and commensal bacterial influx following eradication of Giardia infection in mice. Gut Pathog 2013, 5, 26. [Google Scholar] [CrossRef]
- Kalischuk, L.D.; Inglis, G.D.; Buret, A.G. Campylobacter jejuni induces transcellular translocation of commensal bacteria via lipid rafts. Gut Pathog 2009, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.L.; Peng, W.H.; Kuo, W.T.; Huang, C.Y.; Ni, Y.H.; Lu, K.S.; Turner, J.R.; Yu, L.C. Commensal bacterial endocytosis in epithelial cells is dependent on myosin light chain kinase-activated brush border fanning by interferon-γ. Am J Pathol 2014, 184, 2260–2274. [Google Scholar] [CrossRef] [PubMed]
- Tosi, T.; Pflug, A.; Discola, K.F.; Neves, D.; Dessen, A. Structural basis of eukaryotic cell targeting by type III secretion system (T3SS) effectors. Res Microbiol 2013, 164, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.-P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nature Immunology 2001, 2, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Jones-Carson, J.; Bäumler, A.J.; Falkow, S.; Valdivia, R.; Brown, W.; Le, M.; Berggren, R.; Parks, W.T.; Fang, F.C. Extraintestinal dissemination of Salmonella by CD18-expressing phagocytes. Nature 1999, 401, 804–808. [Google Scholar] [CrossRef]
- Langa, S. Interactions between lactic acid bacteria, intestinal epithelial cells and immune cells. Development of in vitro models; Complutense University of Madrid: Madrid, 2006. [Google Scholar]
- Langa, S.; Maldonado-Barragán, A.; Delgado, S.; Martín, R.; Martín, V.; Jiménez, E.; Ruíz-Barba, J.L.; Mayo, B.; Connor, R.I.; Suárez, J.E.; et al. Characterization of Lactobacillus salivarius CECT 5713, a strain isolated from human milk: from genotype to phenotype. Applied Microbiology and Biotechnology 2012, 94, 1279–1287. [Google Scholar] [CrossRef]
- Rescigno, M.; Citterio, S.; Thèry, C.; Rittig, M.; Medaglini, D.; Pozzi, G.; Amigorena, S.; Ricciardi-Castagnoli, P. Bacteria-induced neo-biosynthesis, stabilization, and surface expression of functional class I molecules in mouse dendritic cells. Proceedings of the National Academy of Sciences 1998, 95, 5229–5234. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Uhr, T. Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 2004, 303, 1662–1665. [Google Scholar] [CrossRef]
- Fung, T.C.; Bessman, N.J.; Hepworth, M.R.; Kumar, N.; Shibata, N.; Kobuley, D.; Wang, K.; Ziegler, C.G.K.; Goc, J.; Shima, T.; et al. Lymphoid-Tissue-Resident Commensal Bacteria Promote Members of the IL-10 Cytokine Family to Establish Mutualism. Immunity 2016, 44, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Roitt, I.M. Essential immunology. 1974. [Google Scholar]
- Martı́n, R.o.; Langa, S.; Reviriego, C.; Jiménez, E.; Marı́n, M.a.L.; Olivares, M.; Boza, J.; Jiménez, J.; Fernández, L.; Xaus, J. The commensal microflora of human milk: new perspectives for food bacteriotherapy and probiotics. Trends in Food Science & Technology 2004, 15, 121–127. [Google Scholar]
- Bertotto, A.; Gerli, R.; Castellucci, G.; Scalise, F.; Vaccaro, R. Human milk lymphocytes bearing the gamma/delta T-cell receptor are mostly delta TCS1-positive cells. Immunology 1991, 74, 360. [Google Scholar]
- Zheng, Y.; Corrêa-Silva, S.; de Souza, E.C.; Maria Rodrigues, R.; da Fonseca, F.A.M.; Gilio, A.E.; Carneiro-Sampaio, M.; Palmeira, P. Macrophage profile and homing into breast milk in response to ongoing respiratory infections in the nursing infant. Cytokine 2020, 129, 155045. [Google Scholar] [CrossRef]
- Lluch, J.; Servant, F.; Païssé, S.; Valle, C.; Valiere, S.; Kuchly, C.; Vilchez, G.; Donnadieu, C.; Courtney, M.; Burcelin, R. The characterization of novel tissue microbiota using an optimized 16S metagenomic sequencing pipeline. PloS one 2015, 10, e0142334. [Google Scholar] [CrossRef] [PubMed]
- Dunn, C.M.; Velasco, C.; Rivas, A.; Andrews, M.; Garman, C.; Jacob, P.B.; Jeffries, M.A. Identification of cartilage microbial DNA signatures and associations with knee and hip osteoarthritis. Arthritis & Rheumatology 2020, 72, 1111–1122. [Google Scholar]
- Berthelot, J.-M.; Lioté, F.; Sibilia, J. Tissue microbiota: A’secondary-self’, first target of autoimmunity? 2022, 89, 105337. [Google Scholar] [CrossRef]
- Hosang, L.; Canals, R.C.; van der Flier, F.J.; Hollensteiner, J.; Daniel, R.; Flügel, A.; Odoardi, F. The lung microbiome regulates brain autoimmunity. Nature 2022, 603, 138–144. [Google Scholar] [CrossRef]
- Immunity to Infection. Primer to the Immune Response 2014, 295–332. [CrossRef]
- Cullin, N.; Antunes, C.A.; Straussman, R.; Stein-Thoeringer, C.K.; Elinav, E. Microbiome and cancer. Cancer Cell 2021, 39, 1317–1341. [Google Scholar] [CrossRef]
- WHO, I. IARC monographs program on the evaluation of carcinogenic risks to humans-some industrial-chemicals, Lyon, 15-22 February 1994-preamble. IARC monographs on the evaluation of carcinogenic risks to humans 1994, 60, 13–33. [Google Scholar]
- Prentice, P.M.; Schoemaker, M.H.; Vervoort, J.; Hettinga, K.; Lambers, T.T.; Van Tol, E.A.; Acerini, C.L.; Olga, L.; Petry, C.J.; Hughes, I.A. Human milk short-chain fatty acid composition is associated with adiposity outcomes in infants. The Journal of nutrition 2019, 149, 716–722. [Google Scholar] [CrossRef]
- Kovács, T.; Mikó, E.; Vida, A.; Sebő, É.; Toth, J.; Csonka, T.; Boratkó, A.; Ujlaki, G.; Lente, G.; Kovács, P.; et al. Cadaverine, a metabolite of the microbiome, reduces breast cancer aggressiveness through trace amino acid receptors. Sci Rep 2019, 9, 1300. [Google Scholar] [CrossRef]
- Sári, Z.; Mikó, E.; Kovács, T.; Jankó, L.; Csonka, T.; Lente, G.; Sebő, É.; Tóth, J.; Tóth, D.; Árkosy, P.; et al. Indolepropionic Acid, a Metabolite of the Microbiome, Has Cytostatic Properties in Breast Cancer by Activating AHR and PXR Receptors and Inducing Oxidative Stress. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, R.; Zhang, Y.; Matuszek, Ż.; Zhang, W.; Xia, Y.; Pan, T.; Sun, J. tRNA Queuosine Modification Enzyme Modulates the Growth and Microbiome Recruitment to Breast Tumors. Cancers 2020, 12, 628. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zallot, R.; Grove, T.L.; Payan, D.J.; Martin-Verstraete, I.; Šepić, S.; Balamkundu, S.; Neelakandan, R.; Gadi, V.K.; Liu, C.F.; et al. Discovery of novel bacterial queuine salvage enzymes and pathways in human pathogens. Proc Natl Acad Sci U S A 2019, 116, 19126–19135. [Google Scholar] [CrossRef]
- Hoyles, L.; Jiménez-Pranteda, M.L.; Chilloux, J.; Brial, F.; Myridakis, A.; Aranias, T.; Magnan, C.; Gibson, G.R.; Sanderson, J.D.; Nicholson, J.K.; et al. Metabolic retroconversion of trimethylamine N-oxide and the gut microbiota. Microbiome 2018, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, Q.; Liu, Y.; Guo, Y.; Li, Q. Awareness of intratumoral bacteria and their potential application in cancer treatment. Discover Oncology 2023, 14, 57. [Google Scholar] [CrossRef]
- Viswanathan, S.; Parida, S.; Lingipilli, B.T.; Krishnan, R.; Podipireddy, D.R.; Muniraj, N. Role of Gut Microbiota in Breast Cancer and Drug Resistance. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Feng, K.; Ren, F.; Shang, Q.; Wang, X.; Wang, X. Association between oral microbiome and breast cancer in the east Asian population: A Mendelian randomization and case–control study. Thoracic Cancer 2024, 15, 974–986. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef]
- Groeger, S.; Zhou, Y.; Ruf, S.; Meyle, J. Pathogenic Mechanisms of Fusobacterium nucleatum on Oral Epithelial Cells. Front Oral Health 2022, 3, 831607. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Yamada, M.; Li, M.; Liu, H.; Chen, S.G.; Han, Y.W. FadA from Fusobacterium nucleatum Utilizes both Secreted and Nonsecreted Forms for Functional Oligomerization for Attachment and Invasion of Host Cells*. Journal of Biological Chemistry 2007, 282, 25000–25009. [Google Scholar] [CrossRef]
- Meng, Q.; Gao, Q.; Mehrazarin, S.; Tangwanichgapong, K.; Wang, Y.; Huang, Y.; Pan, Y.; Robinson, S.; Liu, Z.; Zangiabadi, A.; et al. Fusobacterium nucleatum secretes amyloid-like FadA to enhance pathogenicity. EMBO Rep 2021, 22, e52891. [Google Scholar] [CrossRef]
- Han, Y.W.; Redline, R.W.; Li, M.; Yin, L.; Hill, G.B.; McCormick, T.S. Fusobacterium nucleatum Induces Premature and Term Stillbirths in Pregnant Mice: Implication of Oral Bacteria in Preterm Birth. Infection and Immunity 2004, 72, 2272–2279. [Google Scholar] [CrossRef]
- Han, Y.W.; Shi, W.; Huang, G.T.J.; Kinder Haake, S.; Park, N.H.; Kuramitsu, H.; Genco, R.J. Interactions between periodontal bacteria and human oral epithelial cells: Fusobacterium nucleatum adheres to and invades epithelial cells. Infection and Immunity 2000, 68, 3140–3146. [Google Scholar] [CrossRef]
- Engevik, M.A.; Danhof, H.A.; Ruan, W.; Engevik, A.C.; Chang-Graham, A.L.; Engevik, K.A.; Shi, Z.; Zhao, Y.; Brand, C.K.; Krystofiak, E.S. Fusobacterium nucleatum secretes outer membrane vesicles and promotes intestinal inflammation. MBio 2021, 12, 10–1128. [Google Scholar]
- Nagata, E.; Oho, T. Invasive Streptococcus mutans induces inflammatory cytokine production in human aortic endothelial cells via regulation of intracellular toll-like receptor 2 and nucleotide-binding oligomerization domain 2. Mol Oral Microbiol 2017, 32, 131–141. [Google Scholar] [CrossRef]
- Yu, L.; Maishi, N.; Akahori, E.; Hasebe, A.; Takeda, R.; Matsuda, A.Y.; Hida, Y.; Nam, J.-M.; Onodera, Y.; Kitagawa, Y.; et al. The oral bacterium Streptococcus mutans promotes tumor metastasis by inducing vascular inflammation. Cancer Science 2022, 113, 3980–3994. [Google Scholar] [CrossRef]
- Vela, A.I.; Rey, V.S.d.; Zamora, L.; Casamayor, A.; Domínguez, L.; Fernández-Garayzábal, J.F. Streptococcus cuniculi sp. nov., isolated from the respiratory tract of wild rabbits. International journal of systematic and evolutionary microbiology 2014, 64 Pt 7, 2486–2490. [Google Scholar]
- Li, N.; Zhou, H.; Holden, V.K.; Deepak, J.; Dhilipkannah, P.; Todd, N.W.; Stass, S.A.; Jiang, F. Streptococcus pneumoniae promotes lung cancer development and progression. iScience 2023, 26, 105923. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Huang, J.; Xiao, J.; Xu, W.; Zhang, H.; Yuan, Y.; Yin, Y.; Zhang, X. The Streptococcus virulence protein PepO triggers anti-tumor immune responses by reprograming tumor-associated macrophages in a mouse triple negative breast cancer model. Cell & Bioscience 2023, 13, 198. [Google Scholar] [CrossRef]
- Ma, J.; Gnanasekar, A.; Lee, A.; Li, W.T.; Haas, M.; Wang-Rodriguez, J.; Chang, E.Y.; Rajasekaran, M.; Ongkeko, W.M. Influence of Intratumor Microbiome on Clinical Outcome and Immune Processes in Prostate Cancer. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Henao-Martínez, A.F.; González-Fontal, G.R.; Castillo-Mancilla, J.R.; Yang, I.V. Enterobacteriaceae bacteremias among cancer patients: an observational cohort study. Int J Infect Dis 2013, 17, e374–378. [Google Scholar] [CrossRef]
- Oliero, M.; Calvé, A.; Fragoso, G.; Cuisiniere, T.; Hajjar, R.; Dobrindt, U.; Santos, M.M. Oligosaccharides increase the genotoxic effect of colibactin produced by pks+ Escherichia coli strains. BMC cancer 2021, 21, 1–10. [Google Scholar]
- Aschtgen, M.-S.; Fragkoulis, K.; Sanz, G.; Normark, S.; Selivanova, G.; Henriques-Normark, B.; Peuget, S. Enterobacteria impair host p53 tumor suppressor activity through mRNA destabilization. Oncogene 2022, 41, 2173–2186. [Google Scholar] [CrossRef]
- Giallourou, N.; Urbaniak, C.; Puebla-Barragan, S.; Vorkas, P.A.; Swann, J.R.; Reid, G. Characterizing the breast cancer lipidome and its interaction with the tissue microbiota. Communications biology 2021, 4, 1229. [Google Scholar]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sciences 2020, 247, 117443. [Google Scholar] [CrossRef]
- Parkin, D.M. The global health burden of infection-associated cancers in the year 2002. Int J Cancer 2006, 118, 3030–3044. [Google Scholar] [CrossRef]
- Cao, Y.; Xia, H.; Tan, X.; Shi, C.; Ma, Y.; Meng, D.; Zhou, M.; Lv, Z.; Wang, S.; Jin, Y. Intratumoural microbiota: a new frontier in cancer development and therapy. Signal Transduction and Targeted Therapy 2024, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Lawson, J.S.; Heng, B. Viruses and breast cancer. Cancers (Basel) 2010, 2, 752–772. [Google Scholar] [CrossRef] [PubMed]
- Lau, L.; Gray, E.E.; Brunette, R.L.; Stetson, D.B. DNA tumor virus oncogenes antagonize the cGAS-STING DNA-sensing pathway. Science 2015, 350, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Donnelly, C.R.; Gong, W.; Heath, B.R.; Hao, Y.; Donnelly, L.A.; Moghbeli, T.; Tan, Y.S.; Lin, X.; Bellile, E.; et al. HPV16 drives cancer immune escape via NLRX1-mediated degradation of STING. J Clin Invest 2020, 130, 1635–1652. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; Jiang, S.; Gewurz, B.E. Epstein-Barr Virus LMP1-Mediated Oncogenicity. J Virol 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Floor, S.L.; Dumont, J.E.; Maenhaut, C.; Raspe, E. Hallmarks of cancer: of all cancer cells, all the time? Trends Mol Med 2012, 18, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Brianti, M.T.; Almeida, V.R.; Ortega, M.M.; Fischer, W.; Haas, R.; Matheu, A.; Ribeiro, M.L. Helicobacter pylori infection modulates the expression of miRNAs associated with DNA mismatch repair pathway. Mol Carcinog 2017, 56, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, A.C.; Destefano Shields, C.E.; Wu, S.; Huso, D.L.; Wu, X.; Murray-Stewart, T.R.; Hacker-Prietz, A.; Rabizadeh, S.; Woster, P.M.; Sears, C.L.; et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc Natl Acad Sci U S A 2011, 108, 15354–15359. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Tian, Z.; Kong, X.; Yang, L.; Shan, X.; Dong, B.; Ding, X.; Jing, X.; Jiang, C.; Jiang, N.; et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J Exp Clin Cancer Res 2020, 39, 202. [Google Scholar] [CrossRef] [PubMed]
- Geng, F.; Zhang, Y.; Lu, Z.; Zhang, S.; Pan, Y. Fusobacterium nucleatum Caused DNA Damage and Promoted Cell Proliferation by the Ku70/p53 Pathway in Oral Cancer Cells. DNA Cell Biol 2020, 39, 144–151. [Google Scholar] [CrossRef]
- Maddocks, O.D.; Scanlon, K.M.; Donnenberg, M.S. An Escherichia coli effector protein promotes host mutation via depletion of DNA mismatch repair proteins. mBio 2013, 4, e00152–00113. [Google Scholar] [CrossRef]
- Kim, D.H.; Jin, Y.H. Intestinal bacterial beta-glucuronidase activity of patients with colon cancer. Arch Pharm Res 2001, 24, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Humblot, C.; Murkovic, M.; Rigottier-Gois, L.; Bensaada, M.; Bouclet, A.; Andrieux, C.; Anba, J.; Rabot, S. β-Glucuronidase in human intestinal microbiota is necessary for the colonic genotoxicity of the food-borne carcinogen 2-amino-3-methylimidazo [4, 5-f] quinoline in rats. Carcinogenesis 2007, 28, 2419–2425. [Google Scholar] [CrossRef]
- Mizumoto, A.; Ohashi, S.; Hirohashi, K.; Amanuma, Y.; Matsuda, T.; Muto, M. Molecular Mechanisms of Acetaldehyde-Mediated Carcinogenesis in Squamous Epithelium. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.; Yilmaz, Ö. Possible role of Porphyromonas gingivalis in orodigestive cancers. Journal of oral microbiology 2019, 11, 1563410. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Banerjee, P.; Guo, H.F.; Ireland, S.; Pankova, D.; Ahn, Y.H.; Nikolaidis, I.M.; Liu, X.; Zhao, Y.; Xue, Y.; et al. Epithelial-to-mesenchymal transition drives a pro-metastatic Golgi compaction process through scaffolding protein PAQR11. J Clin Invest 2017, 127, 117–131. [Google Scholar] [CrossRef]
- Vivant, A.L.; Garmyn, D.; Piveteau, P. Listeria monocytogenes, a down-to-earth pathogen. Front Cell Infect Microbiol 2013, 3, 87. [Google Scholar] [CrossRef]
- Lauvau, G.; Vijh, S.; Kong, P.; Horng, T.; Kerksiek, K.; Serbina, N.; Tuma, R.A.; Pamer, E.G. Priming of memory but not effector CD8 T cells by a killed bacterial vaccine. Science 2001, 294, 1735–1739. [Google Scholar] [CrossRef]
- Huang, B.; Zhao, J.; Shen, S.; Li, H.; He, K.L.; Shen, G.X.; Mayer, L.; Unkeless, J.; Li, D.; Yuan, Y.; et al. Listeria monocytogenes promotes tumor growth via tumor cell toll-like receptor 2 signaling. Cancer Res 2007, 67, 4346–4352. [Google Scholar] [CrossRef]
- Duan, Y.; Ma, F.; Guo, B.; Chen, Z.; Liu, Y.; Jiang, X.; Rong, Z.; Zou, Y.; Zhang, H.; Wang, T.; et al. Listeria monocytogenes promotes breast cancer proliferation and enhances the survival rate of circulating breast cancer cells. 2023. [Google Scholar] [CrossRef]
- Chua, M.D.; Mineva, G.M.; Guttman, J.A. Ube2N is present and functions within listeria Actin-rich structures and lamellipodia: A localization and pharmacological inhibition study. Anat Rec (Hoboken) 2023, 306, 1140–1148. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, W.; Font-Burgada, J.; Palmer, T.; Hamil, A.S.; Biswas, S.K.; Poidinger, M.; Borcherding, N.; Xie, Q.; Ellies, L.G.; et al. Ubiquitin-conjugating enzyme Ubc13 controls breast cancer metastasis through a TAK1-p38 MAP kinase cascade. Proc Natl Acad Sci U S A 2014, 111, 13870–13875. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, Z.; Tang, Z.; Huang, Y.; Huang, M.; Liu, H.; Ziebolz, D.; Schmalz, G.; Jia, B.; Zhao, J. More Than Just a Periodontal Pathogen -the Research Progress on Fusobacterium nucleatum. Front Cell Infect Microbiol 2022, 12, 815318. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Zheng, S.; Li, M.; Xu, C.; Jia, D.; Qi, Y.; Hou, T.; Wang, L.; Wang, B. Fusobacterium nucleatum promotes colorectal cancer cells adhesion to endothelial cells and facilitates extravasation and metastasis by inducing ALPK1/NF-κB/ICAM1 axis. Gut Microbes 2022, 14, 2038852. [Google Scholar] [CrossRef]
- Li, G.; Sun, Y.; Huang, Y.; Lian, J.; Wu, S.; Luo, D.; Gong, H. Fusobacterium nucleatum-derived small extracellular vesicles facilitate tumor growth and metastasis via TLR4 in breast cancer. BMC Cancer 2023, 23, 473. [Google Scholar] [CrossRef]
- Franco, A.A.; Buckwold, S.L.; Shin, J.W.; Ascon, M.; Sears, C.L. Mutation of the zinc-binding metalloprotease motif affects Bacteroides fragilis toxin activity but does not affect propeptide processing. Infect Immun 2005, 73, 5273–5277. [Google Scholar] [CrossRef] [PubMed]
- Pradella, D.; Naro, C.; Sette, C.; Ghigna, C. EMT and stemness: flexible processes tuned by alternative splicing in development and cancer progression. Molecular Cancer 2017, 16, 8. [Google Scholar] [CrossRef]
- Parida, S.; Wu, S.; Siddharth, S.; Wang, G.; Muniraj, N.; Nagalingam, A.; Hum, C.; Mistriotis, P.; Hao, H.; Talbot, C.C., Jr.; et al. A Procarcinogenic Colon Microbe Promotes Breast Tumorigenesis and Metastatic Progression and Concomitantly Activates Notch and β-Catenin Axes. Cancer Discov 2021, 11, 1138–1157. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.; Yao, B.; Dong, T.; Cai, S. Emerging roles of intratumor microbiota in cancer metastasis. Trends in Cell Biology 2023, 33, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Rohrbeck, A.; Just, I. Cell Entry of C3 Exoenzyme from Clostridium botulinum. In Uptake and Trafficking of Protein Toxins, Barth, H., Ed.; Springer International Publishing: Cham, 2017; pp. 97–118. [Google Scholar]
- Gao, Y.; Shang, Q.; Li, W.; Guo, W.; Stojadinovic, A.; Mannion, C.; Man, Y.G.; Chen, T. Antibiotics for cancer treatment: A double-edged sword. J Cancer 2020, 11, 5135–5149. [Google Scholar] [CrossRef]
- Wang, X.; Li, J.; Shi, W.; Huang, Z.; Xia, W.; Huang, J.; Su, Y.; Wang, S.; Shi, Y.; Bi, X.; et al. Efficacy of Moxifloxacin plus Treatment of Physician’s Choice in Patients with Metastatic Breast Cancer. Oncologist 2020, 25, e1439–e1445. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, J.D.; Ritter, V.; Purington, N.; Andrade, K.; Han, S.; Liu, M.; Liang, S.-Y.; John, E.M.; Gomez, S.L.; Telli, M.L.; et al. Antimicrobial exposure is associated with decreased survival in triple-negative breast cancer. Nature Communications 2023, 14, 2053. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Potential origins of breast tissue/milk microbiota. A. The structure of human breast. The inset: the structure of the skin. B. three potential routs of microbial transfer to the breast milk/tissue. 1) Breast skin microbiota migration. Microbes of the adjacent skin could enter the mammary gland through the areola. 2) Infant mouth microbiota transfer. During suckling, the oral microbes of the infant could enter the mammary gland through retrograde transfer. 3) Gut-breast axis. Gut mucosal dendritic cells (DCs) occasionally sample commensal bacteria in the lumen and transfer them to lymphoid tissues and eventually reach the mammary gland.
Figure 1.
Potential origins of breast tissue/milk microbiota. A. The structure of human breast. The inset: the structure of the skin. B. three potential routs of microbial transfer to the breast milk/tissue. 1) Breast skin microbiota migration. Microbes of the adjacent skin could enter the mammary gland through the areola. 2) Infant mouth microbiota transfer. During suckling, the oral microbes of the infant could enter the mammary gland through retrograde transfer. 3) Gut-breast axis. Gut mucosal dendritic cells (DCs) occasionally sample commensal bacteria in the lumen and transfer them to lymphoid tissues and eventually reach the mammary gland.
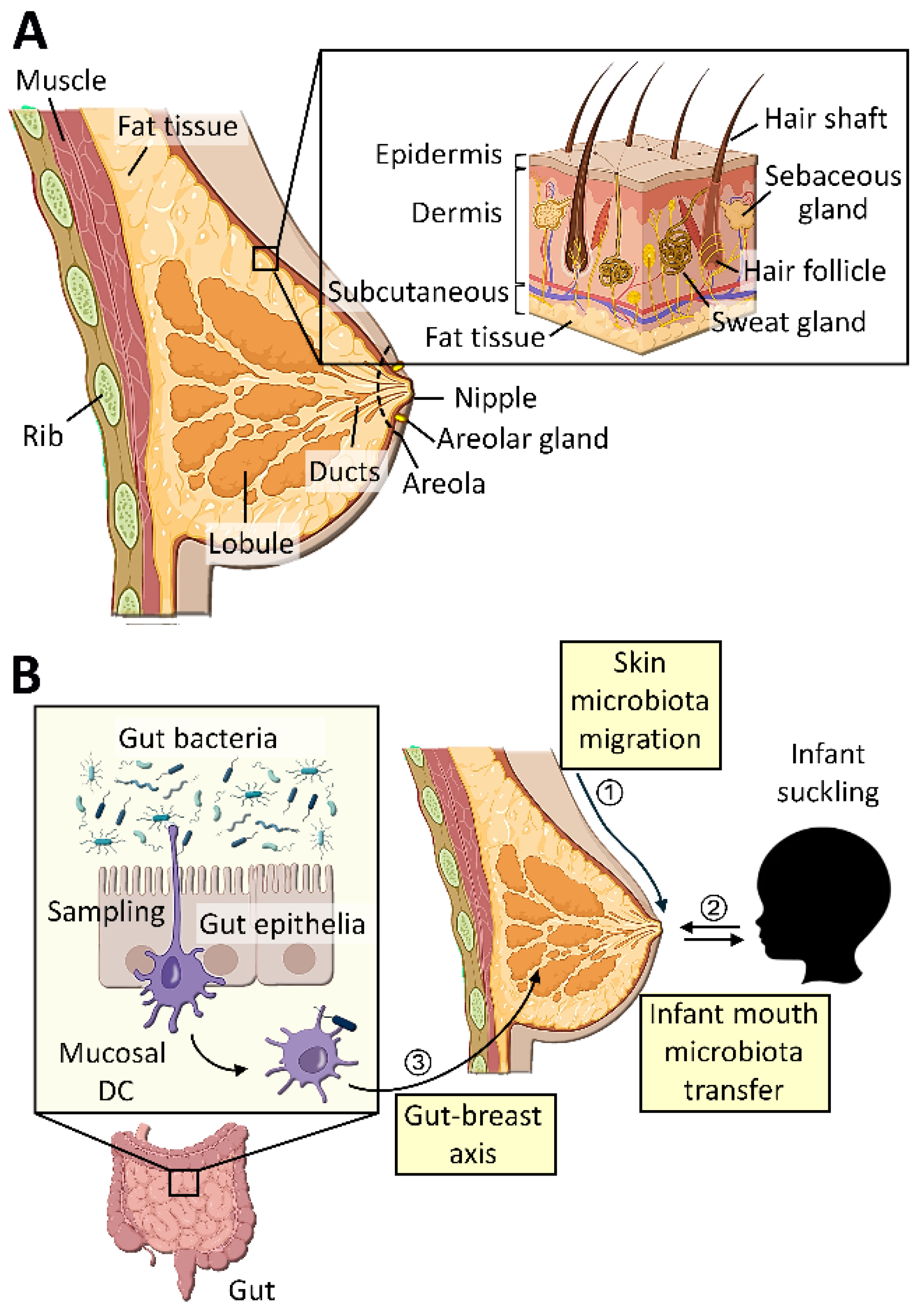
Figure 2.
Two major modes of bacterial translocation. 1) Luminal bacteria are internalized by specialized microfold (M) cells present in the gut epithelial layer. These bacteria are transcytosed and made available to mucosal immune cells. 2) Luminal bacteria are directly taken up by CD18+ phagocytic cells in the mucosal layer that penetrate gut epithelial layer. These immune cells which have taken up bacteria migrate to gut associated lymphoid tissues (GALTs) and then, mesenteric lymph nodes, where they say until they will be disseminated to thymus, spleen, and other distant tissues.
Figure 2.
Two major modes of bacterial translocation. 1) Luminal bacteria are internalized by specialized microfold (M) cells present in the gut epithelial layer. These bacteria are transcytosed and made available to mucosal immune cells. 2) Luminal bacteria are directly taken up by CD18+ phagocytic cells in the mucosal layer that penetrate gut epithelial layer. These immune cells which have taken up bacteria migrate to gut associated lymphoid tissues (GALTs) and then, mesenteric lymph nodes, where they say until they will be disseminated to thymus, spleen, and other distant tissues.
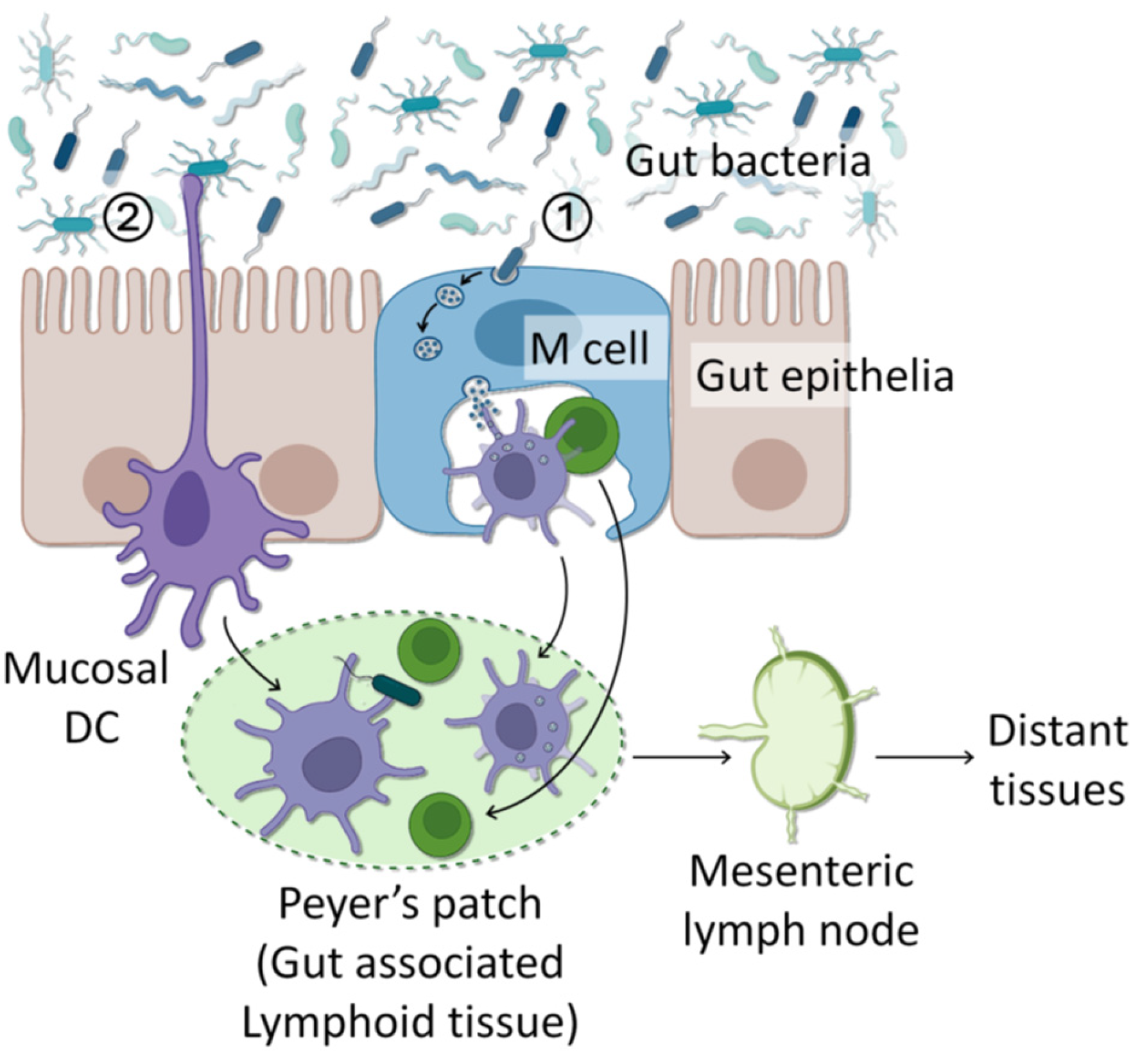
Figure 3.
Roles of intratumoral bacteria in tumor metastasis. Bacteria inside the primary tumor could be transported to the metastasized tumor along with tumor cells. During metastasis, intratumoral bacteria assist the process in several different aspects. These include promotion of EMT, production of MMP, resistance to fluid sheer stress (FSS), and immune suppression. .
Figure 3.
Roles of intratumoral bacteria in tumor metastasis. Bacteria inside the primary tumor could be transported to the metastasized tumor along with tumor cells. During metastasis, intratumoral bacteria assist the process in several different aspects. These include promotion of EMT, production of MMP, resistance to fluid sheer stress (FSS), and immune suppression. .
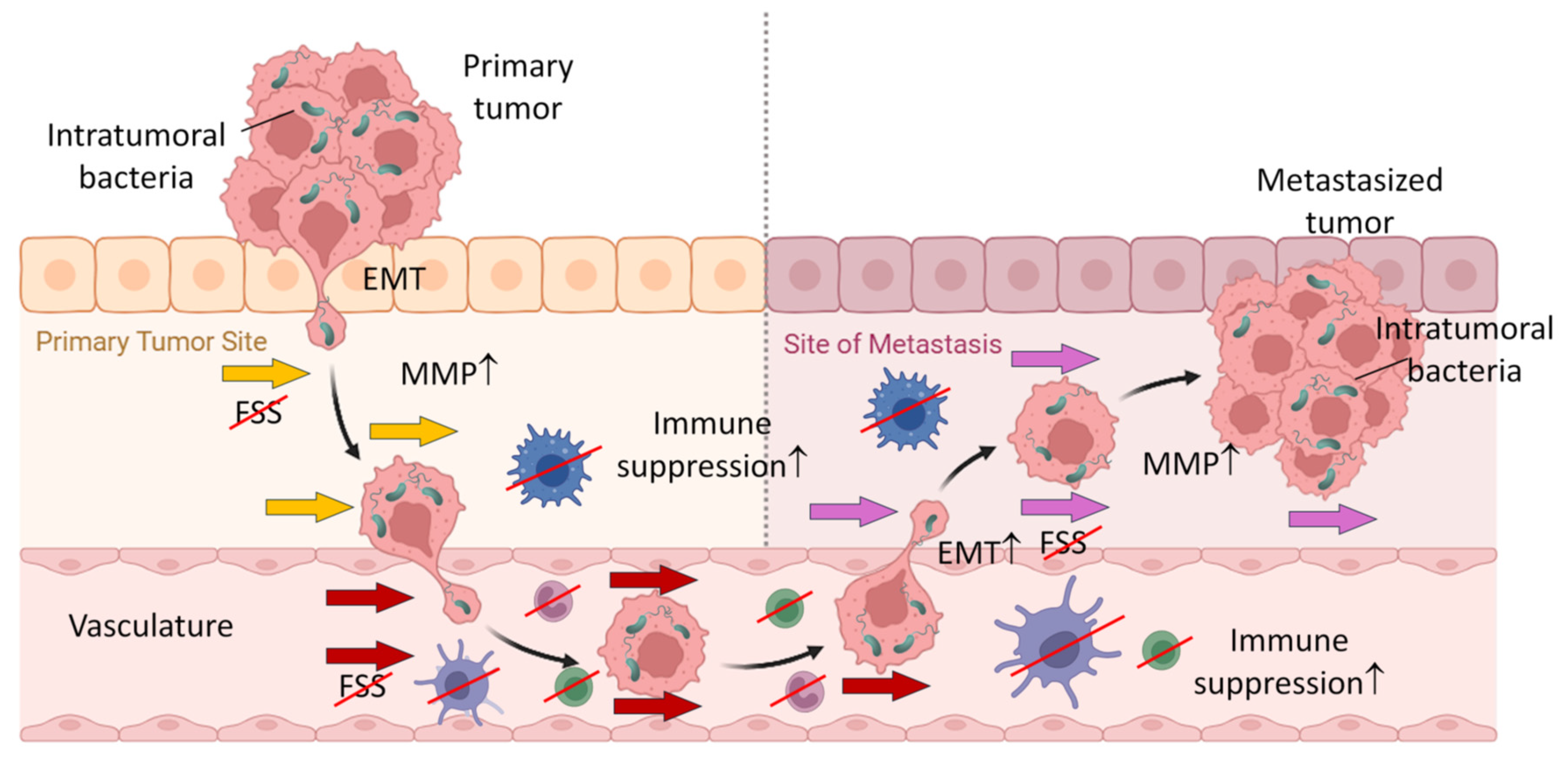

Table 1.
Bacteria differentially represented in normal vs. tumorous breast tissues.
| Normal Breast | Breast Cancer | ||||||
|---|---|---|---|---|---|---|---|
| Microbes | Levels | Functions | Ref. | Microbes | Levels | Functions | Ref. |
| Sphingomonas | Higher | Degrades environmental carcinogens, aromatic hydrocarbons and polycyclic aromatic hydrocarbons; protective against ER+ breast cancer | [24,28] | Fusobacterium nucleatum | Higher | Promotes breast cancer cell attachment, invasion and colonization during metastasis; impairs immunity and therapy response; activates β-catenin-mediated oncogene transcription and cell proliferation; produces β-lactamase for resistance to β-lactam antibiotics (e.g., penicillin) | [24,29,30,31] |
|
Firmicutes, /Actinobacteria |
Higher | Negatively correlate with stromal fibrosis and breast cancer risk; enriched in breast milk | [32,33,34] | ||||
|
Lactobacillaceae, Acetobacterraceae, Leuconostocaceae Xanthomonadaceae |
Higher | Induce fructose and mannose metabolism and immune-related genes; enriched in breast milk of healthy women | [35,36,37] | Enterobacteriaceae, Staphylococcus | Higher | Induce DNA double-strand break of host cells | [38,39] |
| Ralstonia | Higher | Dysregulates genes involved in the carbohydrate metabolism | [35] | ||||
| Cyanobacteria, | Higher | Produces anti-cancer molecule (e.g, Cryptophycin F) | [40] | Atopobium, Gluconacetobacter, | Higher | Modulate immunological responses | [24,41,42] |
| Proteobacteria, Synergistetes, Tenericutes | Higher | Regulate milk composition and production; | [43,44] | Porphyromonadaceae, Ruminococcaceae, | Higher | Participates in aberrant host metabolism | [40,45,46] |
|
Prevotellaceae Butyricimonas, |
Higher | Produce short-chain fatty acids (SCFAs) propionate and butyrate that exert anti-tumor activities | [40,47,48,49] |
Sutterella, Verrucomicrobiaceae |
Higher | Also found in cecal microbiota | [40,50,51] |
| Acinetobacter, | Higher | Abundant in HR+ and HER2+ breast cancer | [40,52] | ||||
|
Flavobacterium Hydrogenophaga |
Higher | Abundant in metastatic breast cancer | [40,53,54] | ||||
|
Alcaligenaceae, Moraxellaceae, Parabacteroides |
Higher | Enriched in breast milk | [40,55] |
Akkermansia (phylum Verrucomicrobia), Thermia, |
Higher | Abundant in TNBC | [40,56] |
Table 2.
Representative bacteria in different types of tumors.
| Cancer types | Microbes | Levels | Protumor mechanisms | Ref. |
|---|---|---|---|---|
| Breast | Fusobacterium nucleatum | Increased | Suppresses T cell infiltration into tumors; promotes tumor growth and metastatic progression | [29] |
| Anaerococcus, Caulobacter Propionibacterium, Streptococcus, Staphylococcus | Decreased | Positively correlated with oncogenic immune features and T-cell activation-related genes | [81] | |
| Bile duct |
Bifidobacteriaceae, Enterobacteriaceae, Enterococcaceae |
Increased | Increased production of bile acids and ammonia, leading to DNA damage in host cells and carcinogenesis | [84] |
| Cervical | Fusobacterium spp. | Increased | Associated with increased IL-4 and TGF-β1 mRNA in cervical cells | [85] |
|
Anaerotruncus, Anaerostipes, Atopobium, Arthrospira, Bacteroides, Dialister, Peptoniphilus, Porphyromonas, Ruminococcus, Treponema |
Increased | Elevates vaginal pH to weaken host defense against infection and promotes tumor formation | [86] | |
| Colorectal | Bacteroides fragilis | Increased | Increased interleukin-17 in the colon and DNA damage in colonic epithelium that accelerate tumor onset and elevate host mortality | [87] |
| Fusobacterium | Increased | Cancer cell proliferation and distant metastasis | [75] | |
| Esophageal | Lactobacillus fermentum | Increased | Establishes acidic environment for growth advantage | [88] |
| Helicobacter pylori | Increased | Spread from gastric colonization | [88] | |
| Campylobacter spp. | Increased | Causes inflammation that could contribute to carcinogenesis | [89] | |
| Porphyromonas gingivalis | Increased | Accelerate cell cycle, promotes cellular migration, and metabolism of potentially carcinogenic substances such as ethanol to carcinogenic derivative, acetaldehyde | [90] | |
| Extrahepatic Bile duct |
Helicobacter pylori | Increased | Increases in virulence genes cagA and vacA abundance and promotes tumor formation | [85] |
| Helicobacter bilis | Increased | Induces inflammation to contribute to tumor formation | [91] | |
| Gallbladder | Fusobacterium nucleatum, Escherichia coli, Enterobacter spp. | Increased | Promotes gallstones development and chronic cholecystitis to contribute to tumor formation | [92] |
| Gastric | Helicobacter pylori | Increased | CagA protein suppresses p53-mediated apoptosis of host cells while increasing cell motility, and metastatic phenotype | [93] |
| Fusobacterium nucleatum | Increased | Induces epithelial-to-mesenchymal transition | [94] | |
| Liver cancer | Helicobacter bifidus | Increased | Contributes to formation of chronic hepatitis that promotes tumor progression | [95] |
| Lung | Acidovorax spp. | Increased | Associated with carcinomas with p53 mutations | [96] |
| Thermus, Legionella | Increased | Associated with the advanced stage and metastatic cancer | [97] | |
| Oral cancer | Fusobacterium nucleatum | Increased | Induces epithelial-to-mesenchymal transition | [94] |
| Firmicutes (esp. Streptococcus), Actinobacteria (esp. Rothia) | Increased | Elevated in normal oral tissues | [98] | |
| Ovarian | Mycoplasma | Increased | Prevalent in 60% of tumors | [99] |
| Pancreatic | Enterobacteriaceae, Pseudomonas spp., Mycobacterium avium, Pseudoxanthomonas, Streptomyces, Bacillus cereus | Increased | Contributes to chemotherapy resistance and immune suppression | [100,101] |
| Malassezia globosa | Increased | Induces the complement cascade through the activation of mannose-binding lectin C3 to promote tumorigenesis | [102] | |
| Prostate | Pseudomonas, Escherichia, Immunobacterium, Propionibacterium spp. | Increased | Induces prostatitis and differentiation of prostate basal cells into ductal cells to promote tumor formation | [103] |
| Propionibacterium acnes spp. | Increased | Induces prostatitis and promotes tumor formation | [104] | |
| Staphylococcus | Increased | Induce inflammation of the prostate tissue and promotes tumor formation | [103] | |
| Fusobacterium nucleatum, Streptococcus oligosporus | Increased | Induces chemoresistance by regulating autophagy | [105] |
Table 3.
Representative bacteria in different breast tumor subtypes.
| Breast cancer Subtypes |
Microbes | Levels | Sample Type | Ref. |
|---|---|---|---|---|
| Luminal A | Proteobacteria (Xanthomonadale,) | Increased | Breast tumor | [78] |
| Tenericutes, Proteobacteria, Planctomycetes | Increased | Breast tumor | [106] | |
| Luminal B | Firmicutes (Clostridium) | Increased | Breast tumor | [78] |
| Tenericutes, Proteobacteria, and Planctomycetes | Increased | Breast tumor | [106] | |
| HER2+ | Thermi, Verrucomicrobia (Akkermasia) | Increased | Breast tumor | [78] |
| Firmicutes (Granulicatella:US31),Bacteroidetes (Dyadobacter) | Increased | Breast tumor | [26] | |
| Firmicutes (Filibacter, Anaerostipes), Bacteroides (Cloacibacterium, Alloprevotella), Proteobacteria (PRD01a011B, Stakelama Blastomonas) | Increased | Breast Tumor | [9] | |
| Proteobacteria (Burkholderiales, Helicobacter pylori) | Increased | Breast Tumor | [80] | |
| TNBC | Streptococcaceae, Ruminococcus | Increased | Breast tumor | [78] |
| Actinomycetaceae, Caulobacteriaceae, Sphingobacteriaceae, Enterobacteriaceae, Prevotellaceae, Brucellaceae, Bacillaceae, Peptostreptococcaceae, Flavobacteriaceae | Increased | Breast tumor | [82] | |
| Prevotella, Brevundimonas, Actinomyces, Aerococcus, Arcobacter, Geobacillus, Orientia, Rothia, Streptococcaceae, Ruminococcus, phyla Euryarchaeota | Increased | Breast tumor | [78,82] | |
| Bartonella, Coxiella, Mobiluncus, Mycobacterium, Rickettsia, Sphingomonas,Azomonas, Alkanindiges, Proteus, Brevibacillus, Kocuria, Parasediminibacterium | Increased | Breast tumor | [107] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



