
Submitted:
22 August 2024
Posted:
27 August 2024
You are already at the latest version
Abstract
Finding, safe and reliable alternatives to fungicides is one of the most actual challenges in agriculture. In this regard, our experiment investigated the effectiveness of different elicitors, botanical extracts and essential oils to control grey mould (Botrytis cinerea) and powdery mildew (Podosphaera aphanis) on strawberry plants. Furthermore, the defence mechanisms activated in plants after treatments were investigated. The trial was conducted in field conditions under plastic tunnel with strawberry plants ‘Elsanta’. In the first experiment, a group of strawberry plants was treated before flowering with elicitors [acibenzolar-S-Methyl – (BTH), chitosan], botanical extracts (seaweed extract, alfalfa hydrolysate) and essential oils (thyme and juniper) to evaluate their ability to reduce the incidence of grey mould infection on flowers. In the second experiment, another group of plants was treated before and after experimental inoculation with P. aphanis. Results indicated that the incidence of flower infected by B. cinerea was significantly reduced by thyme and juniper essential oils, showing values that were not significantly different from the commercial fungicide penconazole (positive control). As a consequence, final yield of essential oil-treated plants was +27% higher as compared to control. No significant differences emerged for other tested products against grey mould. However, gene expression analysis showed an up-regulation of FaEDS1, FaLOX and PR gene expression (FaPR1, FaPR5, FaPR10) in leaves treated with BTH. The other tested natural substances also elicited defence-related genes, however at a lower level than BTH. In the experiment, all treatments applied prior inoculation significantly reduced the incidence and severity of powdery mildew as compared to control. At 28 days after inoculation, chitosan and thyme essential oil applications performed similarly to their positive controls (BTH and penconazole, respectively), showing a significant reduction of disease incidence (by -84 and -92%) as compared to control. Post-inoculum application of essential oils showed similar effective action to fungicide (penconazole) in reducing incidence and severity of powdery mildew. These results indicate that the tested substances could be used as alternatives to fungicides for the control of grey mould and powdery mildew in strawberry, therefore representing a valuable tool for the control of these fungal diseases under the framework of the sustainable agriculture.
Keywords:
Fragaria x ananassa
; Botrytis cinerea
; Podosphaera aphanis
; resistance inducers
; defence-related genes
; acibenzolar-S-Methyl
; chitosan
; thyme essential oil
; juniper essential oils
; seaweed extracts
1. Introduction
Strawberry is the most important berry fruit crop produced globally with an increase of 39.4% between 2008-2021 [1]. In EU, Spain (360 kt), Poland (162 kt), Germany (130 kt) and Italy (117 kt) are leader producing countries. Europe produces nearly 1.7 Kt of strawberries (21% of the world’s quantity). From 2007 to 2021, the EU strawberry market increased yearly by +2.1%, reaching in 2020 ~€3,282 billion [2]. This growth responds to the increasing demand for red fruits, and consumers’ awareness about their nutritional value and antioxidant benefits [3]. However, strawberry cultivation relies on an average of 36 sprays per year with a pesticide application of 18 kg ha-1. Indeed, strawberry recurrently ranks at the top of the ‘dirty dozen’ (a list of 12 top pesticide-contaminated fruit/vegetables), supporting the need to develop zero-residues production methods [2,4]. Powdery mildew, caused by the obligate fungus Podosphaera aphanis (Wallr.), and grey mould, which etiological agent is Botrytis cinerea Pers., are among the diseases of strawberry requiring the highest pesticide applications. Their control requires periodic application of fungicides such as penconazole, quinoxyfen, and trifloxystrobin [5,6,7].
Strawberry production is increasingly shifting from open fields into protected environments [8]. Unfortunately, phytosanitary problems related mainly to grey mould and powdery mildew are increasing inside the new growing environment since these diseases find the optimal conditions such as the lack of precipitation (for powdery mildew), reduced light intensities, high humidity and longer periods of temperature above 20°C [9,10].
Xenobiotic pesticides are still massively used for crop protection, being fungicides the most widely applied chemical [11]. Indeed, despite IPM has been key European cornerstone for crop production, between 2011 and 2020, pesticide sales in the EU were almost stable, around 350,000 t year-1 [12]. Pesticides use poses serious environmental, health and social risks [13]. For example, EFSA reports that in 2019 approx. 4% of fruits and vegetables exceeded residue limits [14]. Moreover, pesticide may include a rise in pathogen resistance and contribute to the environmental resistome [6,7,15]. In Europe, the awareness of the risks of pesticides led to the development of policies for their sustainable use (Directive 2009/128/EC) and actions, such as the Farm2ForK Strategy, aiming at a reduction of 50% of pesticide use by 2030 [16].
The use of botanical extract, essential oil and natural elicitors represents a promising and safe alternative to the use of fungicides [17,18,19]. Resistance inducers are considered a reliable a sustainable method for crop protection since they promote natural plant defences against pathogens [20]. These compounds may act on systemic acquired resistance (SAR) or induced systemic resistance (ISR). Among the signalling molecules regulating plant defences, salicylic acid (SA), jasmonic acid (JA) and ethylene play a crucial role [21]. SAR acts mainly against biotrophic pathogens such as powdery mildew and this resistance is dependent on the salicylic acid (SA) pathway and associated with both systemic expression of pathogenesis-related (PR) genes and systemic increase of SA levels [22,23,24]. SA biosynthesis is activated by PAD4 (Phytoalexin Deficient 4) and EDS1 proteins (Enhanced Disease Susceptibility 1) [25,26]. Several PR proteins were identified such as PR-1 (antifungal); PR-5 (thaumatin-like protein); PR-10 (ribonuclease-like protein) [27,28]. JA is an important regulator involved in ISR signalling pathway, being lipoxygenase (LOX) a key enzyme for its biosynthesis [29,30,31]. JA activates the expression of genes encoding antifungal compounds such as the LOX-derived oxylipins which are synthetized in response to biotic stress [32,33]. Furthermore, plants can activate defences with the biosynthesis of particular defensive substances (secondary metabolites) such as phenylpropanoids, isoprenoids and alkaloids which come from the phenylpropanoid, isoprenoid and alkaloid pathways, respectively [34,35,36,37,38].
In the absence of any biotic stress, the plant defence mechanisms might be trigged through exogenous chemical elicitors (or resistance inducers) which does not directly kill the harmful organism, but promotes the plant defence [37,39,40].
In the present study, we investigated different resistance inducers (acibenzolar-S-methyl, chitosan), botanicals extracts (seaweed extract, alfalfa protein hydrolysate) and essential oils (thyme and juniper). Chitosan is the deacetylated form of the biopolymer chitin which is a component of crustacean and insect exoskeletons and fungal cell walls. Chitosan was found to be able to activate L-phenylalanine ammonia-lyase (PAL) biosynthesis in different crop species, leading to a higher final accumulation of phenolic compounds with antipathogenic activity in tomato [41], papaya [42] and grape [43]. For this reason, chitosan is often used as a plant elicitor, rather than a product with a direct antimicrobial activity [44]. Seaweed extracts are a complex mixture of components such as mineral elements, amino acids, vitamins, phytohormones, betaines, sterols and polysaccharides (e.g. ulvans, agarans, carrageenans, alginates, fucans and laminarins) [45,46,47,48]. Researches have attributed to seaweed polysaccharides the role of effective elicitors of plant defence against biotic stresses [48,49,50,51]. Protein hydrolysates are defined as a “mixtures of polypeptides, oligopeptides and amino acids that are manufactured from protein sources, using partial hydrolysis” [52]. They exert a biostimulating activity tanks to their content in plant hormones, peptides and amino acids [53,54,55]. Although biostimulant properties of legume-derived protein hydrolysates are well studied, few indications are present in scientific literature regarding their eliciting action against pathogens. Essential oils have received a great interest in recent years due to their antimicrobial, antiviral, biodegradable and eco-friendly properties [56,57,58]. Essential oils have a complex composition containing monoterpenes, sesquiterpines and phenylpropanoids. As a consequence, resistance against them is rarely developed by pathogens [59]. In particular, Thymus essential oil has been demonstrated to have a pronounced direct antimicrobial properties and in vitro experiments showed that this activity is mediated by two phenolic components such as thymol and carvacrol [56,60,61]. The antimicrobial activity of Juniper essential oil, linked to some of its components such as limonene and terpinen-4-ol, has been demonstrated on fungal pathogens (e.g. B. cinerea) [62]. In addition to its direct antifungal activity, essential oils can have an elicitor activity against pathogens (e.g. downy mildew) on grapevine plants [63].
The aim of this work was to evaluate the efficacy of selected commercial products (elicitors and essential oils) to control grey mold and powdery mildew in strawberry plants cultivated under plastic tunnel conditions. Moreover, the impact of the tested products on the plant defence mechanism was investigated.
2. Materials and Methods
2.1. Plant Uaterial, Urowth Uonditions and Uested Uroducts
The experiments were conducted under soilless conditions inside a plastic tunnel at the Laimburg Research Centre, in the municipality of Ora/Auer (46° 22’ North; 11° 17’ East; 237 m a.s.l.) in Alto-Adige/South Tyrol, Italy. The temperatures inside greenhouse ranged between 20-25°C (day) and 10-15°C (night) and relative humidity (RH) was kept at high values (Figure 1), thanks to a fog system placed under the soilless structure.
Cold stored strawberry tray plants (TP) ‘Elsanta’ were transplanted in individual pots (16 cm diameter) containing a mixture of natural clay, white peat, perlite and mineral fertilizer (pH 5.8, EC 0.40 dSm-1, total porosity 90%, bulk density 130kg m-3; Profi Substrat, SP VM, Einheits Erde, Germany). All plants were watered daily by drip irrigation and fertilized weekly alternating a granulated long-term NPK fertilizer (10 g per pot; Berry Fertilizer 15-8-12, Hack, Germany) and a water-soluble fertilizer containing macro- and microelements (200 mL per pot at a dosage of 200 g h L-1; Floral 20-20-20, Cifo, Italy). No fungicidal applications were carried out during the experiment, whereas the acaricide (Vertimec EC, Syngenta, Italy) was sprayed twice before flowering at a concentration of 0.5 mL L-1.
Details on the names, abbreviations and application doses of the tested products are reported in Table 1. FUN and BTH were used as positive controls and water as the negative control. In all cases, Adesil A surfactant (Serpan, Italy) was added as wetting agent (0.03 %) to improve the uniform leaf coverage and solubility of different products. Products were applied as a fine mist on the upper surface of the leaves until the runoff point (50 mL per plant) using a hand sprayer before (Experiment 1) or after flowering (Experiment 2 and 3).
2.2. Experimental Uesign
The research activity was subdivided into three experiments:
Experiment 1: Pre-flowering treatment
To assess the efficacy of commercial products in preventing floral infection by B. cinerea, a group of 66 strawberry plants was sprayed before flowering (BBCH 56 – Inflorescence elongating) with a single application of the products listed in Table 1. The same plants were also sampled for gene expression analysis to investigate the molecular bases of products efficacy. The experimental setup was organized as a completely randomized block design with four replicates of two plant each per treatment. In addition to gene expression analysis (see section 2.5.), some parameters such as yield and fruit qualitative traits were evaluated (see section 2.4.), as well as the incidence of flowers infected by B. cinerea (see section 2.3.).
Experiment 2: Post-flowering treatment at pre-inoculation
A second group of 84 strawberry plants was sprayed in post-flowering (BBCH 71 – Receptacle protruding from sepal whorl). Plants were treated with a single application of FUN, BTH, APH and THY (Table 1). SEA was applied 2 times (second application after 3 days) while all other products only one time. 7 days after treatment, plants were artificially inoculated with Podosphaera aphanis (see section 2.3.). Powdery mildew assessment (section 2.3.) was conducted considering each strawberry plant as a single replication (8 plants per treatment).
Experiment 3: Post-flowering treatment at post-inoculation
The experiment aimed at investigating the potential curative effect of essential oils (THY and JUN) against P. aphanis. For this aim, 34 strawberry plants were sprayed with THY and JUN 24 hours after inoculation. Penconazole and water were used as positive and negative control, respectively.
2.3. Disease Ussessment and Unoculation Urocedure of Podosphaera Uphanis
Grey mould incidence was assessed in condition of natural infection. Incidence was calculated taking in account the percentage of symptomatic flowers in each plant [64].
Powdery mildew incidence and severity were assessed after experimental inoculation of the plants. Conidia of P. aphanis were collected from naturally infected, symptomatic strawberry plants ‘Elsanta’ obtained from a local farm. One hour before inoculation, a powdery mildew conidia suspension was produced by harvesting conidia by washing symptomatic strawberry leaves into a flask of 0.03% Adesil A (Serpan, Italy) in distilled water. Conidia concentration was quantified with a haemocytometer and adjusted to 105 conidia mL-1, according to the methodology of [65] with some modifications. Powdery mildew conidia suspension was sprayed onto the upper surface of the leaves (100 mL per plant). Approximately every 2 weeks after inoculation, the assessment of powdery mildew incidence and severity was performed. The percentage of infected leaves per plant was calculated by counting the number of healthy and infected leaves (presenting at least one powdery mildew patch per leaf) [8]. Disease severity was determined considering 12 leaves per plant (8 plants per treatment) and was evaluated with a plant image analyser (BioLeaf – Foliar Analysis™) and expressed as percentage of infected area compared to total leaf area.
2.4. Yield and Uruit Uuality
Ripe strawberry fruits (fully red color) were harvested every three days, starting at 50 days after transplanting. Data coming from each fruit picking were used to calculate the final cumulative yield per plant (as grams plant-1). Fruit quality was determined on four mature and homogeneous fruits per plant. The firmness (FF as kg cm-2), the total soluble solids (TSS as °Brix) and the titratable acidity (TA as g L-1 of citric acid) were evaluated, respectively, with a penetrometer (6 mm in diameter), a portable refractometer (PAL-1, ATAGO, Tokyo, Japan) and with a titrator (TitroLine easy, SCHOTT, Mainz, Germany) by titrating strawberry pulp to pH 8.2 using 0.1 M NaOH. Extraction and quantification of phenolic compounds was determined using a method described by Meyers et al. [66] and Wolfe et al. [67]. Briefly, phenolics were extracted from lyophilized leaves/fruits using 80% methanol acidified with H3PO4 and NaF. The total phenolic content was determined with the Folin-Ciocalteu colorimetric method and expressed as milligrams of gallic acid equivalents (GAE) per 100 grams of dry weight (mg GAE 100g-1 DW).
2.5. Gene Uxpression Unalysis
For gene expression analysis, leaf samples were collected at 0, 48, 72, 96 and 168 hours after treatment taking two young single leaves per plant. The leaves were immersed in liquid nitrogen and stored at -80°C. At every sampling time, two plants per treatment were considered. Each plant was sampled only once to avoid a wounding-effect. Changes in expression of genes related to downstream components of the SAR pathway (Table 2) were determined by a reverse transcription-quantitative real-time polymerase chain reaction (RT-qPCR) as described in Cellini et al. [68]. Briefly, total RNA was extracted from strawberry leaves following the manufacturer’s instructions in Spectrum™ Plant Total RNA Kit (Sigma, USA). RNA was used for cDNA synthesis with reverse-transcription PCR. Specific primers were designed using Primer3 software and previously tested for specificity (Table 2). The expression of genes was measured using a StepOne Plus Real-Time PCR instrument with a SYBR Green-based assay. Strawberry leaves pre-treated with different resistance inducers were examined for FaPR1, FaPR5, FaPR10, FaEDS1 and FaLOX genes.
All quantifications were normalized by the threshold cycle value (CT) compared to the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (FaGPDH2). Gene expression analysis in Experiment 1 was conducted on plants treated with BTH, SEA, CHI and THY.
2.6. Statistical Unalysis
One-way analysis of variance (ANOVA) was performed to determine significance among treatments, after validating data for normal distribution (Shapiro-Wilk normality test, p > 0.05) and equality of variances (Bartlett’s test, p > 0.05). For non-normal data, Kruskal-Wallis test was applied. Data expressed in percentage were arcsin-transformed prior to analysis of variance. In case of significance, mean separation was done with the least significant difference (LSD) test. All analyses were carried out in R v. 3.3.1. (R Development Core Team 2016). Results are expressed as mean ± standard error (SE).
3. Results
3.1. Efficacy against Uatural Unfection by Botrytis Uinerea (Experiment 1)
Natural disease incidence in untreated plants was around 50%, whereas treatment with fungicide was able to halve the percentage of infected flowers per plant compared to control (Figure 2). An effect similar to that of the fungicide was observed in plants treated with THY and JUN (40-50% less infections in comparison to control). No significant differences were detected among other treatments (33-44% of infected flowers per plant) in comparison to untreated plants.
3.2. Preventive Uffects against Uowdery Uildew Uncidence and Ueverity on Ueaves (Experiment 2)
Symptoms of powdery mildew on strawberry leaves started to be visible about 14 days after inoculation. Considering preventive application, results indicated that all tested products controlled powdery mildew development on strawberry leaves (Figure 3A and B). More in detail, at 28 days after inoculation, CHI and THY applications performed a similar action to their positive controls (BTH and FUN, respectively), showing a significant incidence reduction (by -84 and -92%) as compared to control. SEA and APH treatments showed to significantly reduce powdery mildew incidence (-42 and -58%, respectively, as compared to control). The statistical differences among treatments remained similar at 42 days (Figure 3A). Powdery mildew was well established in untreated plants at almost 2 months after inoculation, when more 65% of the leaves presented at least one infection site (Figure 3A). At that time, all treatments maintained an inhibitory action towards powdery mildew. More in detail, fungal incidence in CHI-, SEA- and APH-treated plants was similar to that observed on BTH-treated plants, whereas THY effect was analogue to FUN. Assessment of disease severity showed that all treatments supressed development of powdery mildew. Despite disease severity remained low, with symptoms developing on less than 5% of leaf area, differences among treatments were detected (Figure 3B). In particular, as observed for disease incidence, the application of FUN, CHI and THY demonstrated to significantly reduce the extension of red blotches (up to 94% of reduction as compared to untreated).
3.3. Curative Uffects against Uowdery Uildew Uncidence and Ueverity on Ueaves (Experiment 3)
Post-inoculum application of products showed the effective action of essential oils in reducing disease incidence and severity (Figure 4A and B). Although both essential oils (THY and JUN) showed to inhibit powdery mildew development with an action similar to positive control (FUN). THY seemed to have a higher inhibitory action against the pathogen, being the disease severity at 56 days the lowest among the treatments.
3.4. Fruit Uroduction and Uuality
Plants treated with essential oils (THY and JUN) showed to increase the final yield per plant. In the case of JUN-treated plants, yield result was comparable to that obtained with the fungicide treatment (around 160 g plant-1) (Table 2). The enhanced yield in THY- and JUN-treated plants (+23 and +31% as compared to control, respectively) was a direct consequence of the reduction in fruit losses due to the infection (about 15% more than untreated fruits) (Table 2).
Treatments such as CHI and SEA demonstrated also to significantly increase the yield (135 g compared to 120 g plant-1 in control plants). BTH- and APH-treated plants yielded the same amount as control plants. Regarding fruit weight, no significant differences were observed among treatments (around 10-12 g fruit-1) (Table 2).
A higher sugar and acidity content was observed in untreated fruits, though differences with treated fruits were not significant (Table 2). As to phenolic content in strawberry fruits, no significant differences were detected. FUN-treated fruits showed a tendency to increase this parameter (+24% as compared to control fruits).

Table 2.
Yield parameters (total yield per plant, number of fruits per plant and mean fruit weight) and fruits quality attributes as affected by tested products.
Table 2.
Yield parameters (total yield per plant, number of fruits per plant and mean fruit weight) and fruits quality attributes as affected by tested products.
| Treatment | Total yield (g plant-1) |
Number fruits plant-1 (N°) |
Mean fruit weight (g) |
Total soluble solids (°Brix) |
Titratable acidity (g citric acid L-1) |
Total phenol content (mg GAE 100 g-1 DW) |
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | ||||||||||||||||||||
| CON | 120.80 | ± 2.471 | e | 11.63 | ± 0.71 | de | 10.62 | ± 0.57 | 13.13 | ± 0.27 | 4.29 | ± 0.49 | 2,139.09 | ± 63.54 | ||||||
| FUN | 161.05 | ± 3.63 | a | 14.00 | ± 0.46 | ab | 11.57 | ± 0.38 | 11.43 | ± 0.52 | 3.75 | ± 0.61 | 2,602.84 | ± 303.15 | ||||||
| BTH | 121.46 | ± 3.28 | de | 12.00 | ± 0.60 | cde | 10.21 | ± 0.29 | 12.73 | ± 1.06 | 3.48 | ± 0.17 | 2,250.58 | ± 145.74 | ||||||
| SEA | 133.04 | ± 3.29 | cd | 13.13 | ± 0.90 | abcd | 10.44 | ± 0.72 | 10.68 | ± 0.72 | 3.11 | ± 0.17 | 2,204.47 | ± 92.48 | ||||||
| CHI | 135.64 | ± 6.60 | c | 12.38 | ± 0.80 | bcde | 11.10 | ± 0.48 | 11.23 | ± 1.62 | 2.95 | ± 0.33 | 2,286.84 | ± 125.48 | ||||||
| THY | 148.55 | ± 2.71 | b | 13.50 | ± 0.63 | abc | 11.17 | ± 0.55 | 11.60 | ± 0.69 | 3.12 | ± 0.24 | 2,499.77 | ± 256.05 | ||||||
| APH | 127.61 | ± 4.70 | cde | 11.25 | ± 0.67 | e | 11.54 | ± 0.54 | 12.53 | ± 0.17 | 3.24 | ± 0.14 | 2,431.74 | ± 57.76 | ||||||
| JUN | 157.69 | ± 4.82 | ab | 14.38 | ± 0.32 | a | 10.98 | ± 0.26 | 11.38 | ± 0.36 | 2.91 | ± 0.14 | 2,235.11 | ± 114.87 | ||||||
| Significance | *** | ** | ns | ns | ns | ns | ||||||||||||||
1 Means ± SE within the same column followed by the same letter, do not significantly differ according to LSD test at p < 0.05 (n=4). Anova significant differences: ***p < 0.001; **p < 0.01; *p < 0.05; ns: not significant.
3.5. Gene Uxpression Unalysis
Results showed an up-regulation of PR genes expression (FaPR1, FaPR5, FaPR10) in leaves treated with BTH (Figure 5A, 5B and 5C). More in detail, gene expression was maximal 4 days after BTH treatment and then decreased to standard level. BTH application induced a long lasting (up to 7 days) increase of FaEDS1 and FaLOX genes expression. The expression levels of target genes appeared weaker in plants treated with the natural elicitors (SEA and CHI) and THY than the positive control (BTH). As shown in Figure 5, FaPR1, FaPR5, FaPR10 and FaLOX exhibited a peak of expression at 2 and 3 days after SEA application (fold change <2). These effects completely disappeared at 4 (for FaPR5, FaPR10 and FaLOX) and 7 days (for FaPR1). FaEDS1 was not expressed after treatment. Contrary to what observed in SEA, starting at 2 days after CHI treatment, FaPR1, FaPR5, FaEDS1 and FaLOX were induced and their expression levels continued to be stable even at 7 days (except for FaLOX). Thyme essential oil application demonstrated to have a low effect on activation of defence-related genes (fold-change <1). SEA, CHI and THY proved to enhance defence-related gene expression, even though at a lower level as compared to BTH (fold change expression <2).
4. Discussion
Pre-flowering applications of essential oils reduced the incidence of B. cinerea in flowers by directly inhibiting pathogen growth. This effect is likely due to the well-known antifungal activity of essential [58,69,70]. More in details, essential oils can directly inhibit biosynthesis of chitin, β-glucans and ergosterol, alter fungal mitochondrial electron transport chain, cell division, and modify RNA/DNA and protein synthesis [71,72,73,74,75,76]. Essential oils may also induce defence mechanisms in plant tissues, including the activation of antioxidant and defence-related enzymes and the accumulation of phenolic compounds [57,77]. Positive effect of thyme essential oil against grey mould and Fusarium wilt in tomato plants was observed by Ben-Jabeur et al. [60]. The authors attributed the protective action of thyme extract to a combination of a direct antifungal activity and the induction of defences in plants. However, in our experiment, application of THY did not induce any significant defence-related gene expression in plant tissues, nor altered total phenolic content (Figure 5; Table 3). The results may be related to the time required for strawberries to enhance their plant defences or to the activation of defence mechanisms that are not associated with SAR. Concerning the induction of phenolic compounds, we measure it only on fruit that were not directly exposed to essential oil. Furthermore, differences in phenolic compounds may be related to the induction of specific compounds rather than a generalized induction of this chemical class [78]. The reduction of flower losses by B. cinerea in plants treated with essential oils and fungicide could explain the increased final yield measured in those treated plants.
Apart from the THY and JUN essential oils, none of the other tested products, including the commercial resistance inducer BTH, were able to control grey mould flower infections (Figure 2). BTH-induced resistance against diseases was observed on several plants such as tomato, Arabidopsis, peach, apple, strawberry and papaya [79,80,81,82,83,84]. Although our results confirmed the induction of defence responses in BTH-treated strawberry leaves (the highest level of gene expression in comparison to all tested products), incidence of infected flowers, as well as the final yield was not significantly different to control treatment.
The lack of efficacy of BTH could be related to the timing of resistance induction. Several studies showed that the rise of plant defences needs a minimum of 3-4 days, reaches a peak in two-three weeks and decreases to normal level in approximately 4 weeks [85]. Indeed, in our study a rise of defence related genes was observed at 4 days after the treatment. This indicates that, also in our experimental conditions, it can be assumed a similar timing of plant resistance as found in other studies [86]. However, B. cinerea incidence was assessed only more than 60 days after treatment, when the BTH effect was probably already fading. Furthermore, under natural disease pressure, B. cinerea inoculum can built up rapidly under favourable conditions making results less predictable. In this view, a product like essential oils, which have a dual mode of action—direct antifungal effects and induction of resistance—may result in a more consistent protective effect throughout the season. When considering the experiments in controlled inoculation with P. aphanis, the positive action of post-flowering application of elicitors and essential oils against powdery mildew infections was shown. Considering both preventive (Experiment 2) and curative actions (Experiment 3), the thyme essential oil had a protective effect against powdery mildew similar to that of the fungicide. This finding is in agreement with studies conducted by Sturchio et al. [87] and Mostafa et al. [88], where clove, lemongrass, peppermint, rosemary, tea tree and thyme essential oils demonstrated to be valid fungicidal alternatives to control powdery mildew infections on Cucurbitaceae crops. In our experiment, chitosan, seaweed extract, alfalfa hydrolysate showed an action similar to BTH treatment against powdery mildew infection (Figure 3). Induction of resistance by chitosan application is also a well-studied topic. Being a polysaccharide, chitosan could act in the same way as described for algal polysaccharides. Accumulation of PR proteins, phytoalexins, lignin biosynthesis and callose formation were among some of the defence mechanisms induced by chitosan in treated plants [44]. As an example, grapes treated with chitosan showed to be less susceptible to powdery mildew and to enhance phenolic concentration in berry peel [89]. Similar results were also obtained by Faoro et al. [90] with powdery mildew infection on barley.
Carrot leaves treated with seaweed extract (Ascophyllum nodosum) showed to be less infected by Alternaria and Botrytis, due to activation of defence genes (e.g. PR1, PR5 and PAL), as well as to an increased activity of defence enzymes and accumulation of phenols and phytoalexins [91]. Polysaccharides contained in seaweed extracts could be considered signalling molecules because they activate salicylic acid (SA) and jasmonic acid (JA) signalling pathway, promoting the systemic acquired (SAR) and the induced systemic resistance (ISR) in treated plants [48]. Marine polysaccharides such as alginates, fucans and laminarin, which are components of the utilized brown seaweed extract, could have contributed to enhance plant defences. The mechanism of action of alfalfa protein hydrolysate against diseases probably involves the complex biochemical composition of the hydrolysate. More in detail, two main categories of APH components such as peptides and amino acids are to be considered as signalling molecules [92,93,94]. Cappelletti et al. [95] observed the reduced infection by powdery mildew in zucchini plants treated with different protein hydrolysates. The authors explained this result with the involvement of amino acids and peptides in the regulation of innate immune response in plants.
5. Conclusions
The effectiveness of natural products for the control of grey mould and powdery mildew were evaluated in this experiment. Essential oils showed an effect similar to synthetic fungicide both against B. cinerea and P. aphanis, significantly reducing incidence and severity of infections. The highest defence-related gene expression level was induced by BTH application, even though only incidence of powdery mildew was reduced in those treated plants. As for the other foliar applications (seaweed extract, alfalfa protein hydrolysate and chitosan), they also demonstrated to be valid alternatives to fungicides against powdery mildew.
Appropriate application way (dosages, type of formulation, time and number of application) seems to be of pivotal relevance to improve the efficiency of this group of products. Therefore, further investigations should lead to this direction.
Author Contributions
S.S. investigation, data analysis, writing—original draft preparation; C.A. supervision, conceptualization, writing—review and editing; A.C., I.D., G.B. molecular, pathological and biochemical analysis and experimental design, formal analysis; F.S. supervision, conceptualization, formal analysis, manuscript writing and revisions.
Funding
The research was co-founded by the Free University of Bozen-Bolzano (Project BIO_TOOL TN 1B07). S.S. was funded by a Ph.D. grant from the Free University of Bozen-Bolzano.
Acknowledgments
The authors would like to thank the Laimburg Research Centre for providing the experimental site where the study has been conducted.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 20 October 2022).
- IndexBox, Inc. EU - Strawberries - Market Analysis, Forecast, Size, Trends and Insights; 2021.
- van der Wolf, J.M.; Evenhuis, A.; Kastelein, P.; Krijger, M.C.; Funke, V.Z.; van den Berg, W.; Moene, A.F. Risks for Infection of Strawberry Plants with an Aerosolized Inoculum of Xanthomonas Fragariae. Eur. J. Plant Pathol. 2018, 152, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Rosello, C.; Bélanger, R.; Ratti, C. Fate of Residual Pesticides in Fruit and Vegetable Waste (FVW) Processing. Foods 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-O.; Shin, J.-H.; Gumilang, A.; Chung, K.; Choi, K.Y.; Kim, K.S. Effectiveness of Different Classes of Fungicides on Botrytis Cinerea Causing Gray Mold on Fruit and Vegetables. Plant Pathol. J. 2016, 32, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of Pesticides Use in Agriculture: Their Benefits and Hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Wightwick, A.; Walters, R.; Allinson, G.; Reichman, S.; Menzies, N. Environmental Risks of Fungicides Used in Horticultural Production Systems. In Fungicides; InTech, 2010.
- Hukkanen, A.T.; Kokko, H.I.; Buchala, A.J.; McDougall, G.J.; Stewart, D.; Kärenlampi, S.O.; Karjalainen, R.O. Benzothiadiazole Induces the Accumulation of Phenolics and Improves Resistance to Powdery Mildew in Strawberries. J. Agric. Food Chem. 2007, 55, 1862–1870. [Google Scholar] [CrossRef]
- Elad, Y.; Malathrakis, N.E.; Dik, A.J. Biological Control of Botrytis-Incited Diseases and Powdery Mildews in Greenhouse Crops. Crop Prot. 1996, 15, 229–240. [Google Scholar] [CrossRef]
- Sylla, J.; Alsanius, B.W.; Krüger, E.; Becker, D.; Wohanka, W. In Vitro Compatibility of Microbial Agents for Simultaneous Application to Control Strawberry Powdery Mildew (Podosphaera Aphanis). Crop Prot. 2013, 51, 40–47. [Google Scholar] [CrossRef]
- EUROSTAT European Statistics. Agri-Environmental Indicator - Consumption of Pesticides; 2021.
- EUROSTAT Statistics on Agricultural Use of Pesticides in the EU; 2019.
- EEA How Pesticides Impact Human Health and Ecosystems in Europe (Briefing No. 06/2023); The European Environment Agency (EEA), 2023.
- EFSA EFSA- The 2019 European Union Report on Pesticide Residues in Food. 2019. [CrossRef]
- Cernava, T.; Erlacher, A.; Soh, J.; Sensen, C.W.; Grube, M.; Berg, G. Enterobacteriaceae Dominate the Core Microbiome and Contribute to the Resistome of Arugula (Eruca Sativa Mill.). Microbiome 2019, 7, 13. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Donati, I.; Ferrari, E.; Tanunchai, B.; Fareed Mohamed Wahdan, S.; Sadubsarn, D.; Farneti, B.; Checcucci, A.; Buscot, F.; et al. Taxonomical and Functional Composition of Strawberry Microbiome Is Genotype-Dependent. Adv. Plant Genet. Genomics 2022, 42, 189–204. [Google Scholar] [CrossRef]
- Borges, A.A.; Sandalio, L.M. Induced Resistance for Plant Defense. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Gutiérrez Martínez, P.; Alkan, N. Induced Resistance to Control Postharvest Decay of Fruit and Vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Terry, L. Elicitors of Induced Disease Resistance in Postharvest Horticultural Crops: A Brief Review. Postharvest Biol. Technol. 2004, 32, 1–13. [Google Scholar] [CrossRef]
- Reglinski, T.; Vanneste, J.; Wurms, K.; Gould, E.; Spinelli, F.; Rikkerink, E. Using Fundamental Knowledge of Induced Resistance to Develop Control Strategies for Bacterial Canker of Kiwifruit Caused by Pseudomonas Syringae Pv. Actinidiae. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Phuntumart, V.; Marro, P.; Métraux, J.-P.; Sticher, L. A Novel Cucumber Gene Associated with Systemic Acquired Resistance. Plant Sci. 2006, 171, 555–564. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. Intervention of Phytohormone Pathways by Pathogen Effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef] [PubMed]
- Mishina, T.E.; Zeier, J. The Arabidopsis Flavin-Dependent Monooxygenase FMO1 Is an Essential Component of Biologically Induced Systemic Acquired Resistance. PLANT Physiol. 2006, 141, 1666–1675. [Google Scholar] [CrossRef]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from Red Seaweeds As Promoters of Growth and Elicitors of Defense Response in Plants. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Garrido-Gala, J.; Gadea, J.; Blanco-Portales, R.; Muñoz-Mérida, A.; Trelles, O.; de los Santos, B.; Arroyo, F.T.; Aguado-Puig, A.; Romero, F.; et al. Partial Activation of SA- and JA-Defensive Pathways in Strawberry upon Colletotrichum Acutatum Interaction. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Cui, H.; Gobbato, E.; Kracher, B.; Qiu, J.; Bautor, J.; Parker, J.E. A Core Function of EDS1 with PAD4 Is to Protect the Salicylic Acid Defense Sector in Arabidopsis Immunity. New Phytol. 2017, 213, 1802–1817. [Google Scholar] [CrossRef]
- Carr, J.P.; Lewsey, M.G.; Palukaitis, P. Chapter 3 - Signaling in Induced Resistance. In Advances in Virus Research; Carr, J.P., Loebenstein, G., Eds.; Academic Press, 2010; Vol. 76, pp. 57–121 ISBN 0065-3527.
- Ebrahim, S.; Usha, K.; Singh, B. Pathogenesis Related (PR) Proteins in Plant Defense Mechanism. Sci Microb Pathog 2011, 2, 1043–1054. [Google Scholar]
- Chernin, L.; Glick, B.R. The Use of ACC Deaminase to Increase the Tolerance of Plants to Various Phytopathogens. In Bacteria in Agrobiology: Stress Management; Maheshwari, D.K., Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 2012; pp. 279–299. ISBN 978-3-642-23465-1. [Google Scholar]
- Pieterse, C.M.J.; Van Pelt, J.A.; Ton, J.; Parchmann, S.; Mueller, M.J.; Buchala, A.J.; Métraux, J.-P.; Van Loon, L.C. Rhizobacteria-Mediated Induced Systemic Resistance (ISR) in Arabidopsis Requires Sensitivity to Jasmonate and Ethylene but Is Not Accompanied by an Increase in Their Production. Physiol. Mol. Plant Pathol. 2000, 57, 123–134. [Google Scholar] [CrossRef]
- Turner, J.G.; Ellis, C.; Devoto, A. The Jasmonate Signal Pathway. Plant Cell 2002, 14, S153–S164. [Google Scholar] [CrossRef] [PubMed]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and Action of Jasmonates in Plants. Annu. Rev. Plant Biol. 1997, 48, 355–381. [Google Scholar] [CrossRef] [PubMed]
- Vellosillo, T.; Aguilera, V.; Marcos, R.; Bartsch, M.; Vicente, J.; Cascon, T.; Hamberg, M.; Castresana, C. Defense Activated by 9-Lipoxygenase-Derived Oxylipins Requires Specific Mitochondrial Proteins. PLANT Physiol. 2013, 161, 617–627. [Google Scholar] [CrossRef]
- Iriti, M.; Rossoni, M.; Borgo, M.; Ferrara, L.; Faoro, F. Induction of Resistance to Gray Mold with Benzothiadiazole Modifies Amino Acid Profile and Increases Proanthocyanidins in Grape: Primary versus Secondary Metabolism. J. Agric. Food Chem. 2005, 53, 9133–9139. [Google Scholar] [CrossRef]
- Korkina, L.G. Phenylpropanoids as Naturally Occurring Antioxidants: From Plant Defense to Human Health. Cell Mol Biol 2007, 53, 15–25. [Google Scholar]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Jakopic, J.; Cunja, V.; Veberic, R.; Munda, A.; Stampar, F. Phenolic Compounds as Defence Response of Pepper Fruits to Colletotrichum Coccodes. Physiol. Mol. Plant Pathol. 2013, 84, 138–145. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Gómez-Plaza, E. Elicitors: A Tool for Improving Fruit Phenolic Content. Agriculture 2013, 3, 33–52. [Google Scholar] [CrossRef]
- Sivankalyani, V.; Feygenberg, O.; Diskin, S.; Wright, B.; Alkan, N. Increased Anthocyanin and Flavonoids in Mango Fruit Peel Are Associated with Cold and Pathogen Resistance. Postharvest Biol. Technol. 2016, 111, 132–139. [Google Scholar] [CrossRef]
- Boubakri, H.; Poutaraud, A.; Wahab, M.A.; Clayeux, C.; Baltenweck-Guyot, R.; Steyer, D.; Marcic, C.; Mliki, A.; Soustre-Gacougnolle, I. Thiamine Modulates Metabolism of the Phenylpropanoid Pathway Leading to Enhanced Resistance to Plasmopara Viticolain Grapevine. BMC Plant Biol. 2013, 13, 1–15. [Google Scholar] [CrossRef]
- Cluzet, S.; Torregrosa, C.; Jacquet, C.; Lafitte, C.; Fournier, J.; Mercier, L.; Salamagne, S.; Briand, X.; Esquerre’-Tugaye’, M.-T.; Dumas, B. Gene Expression Profiling and Protection of Medicago Truncatula against a Fungal Infection in Response to an Elicitor from Green Algae Ulva Spp. Plant Cell Environ. 2004, 27, 917–928. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Rabea, E.I. Potential of the Biopolymer Chitosan with Different Molecular Weights to Control Postharvest Gray Mold of Tomato Fruit. Postharvest Biol. Technol. 2009, 51, 110–117. [Google Scholar] [CrossRef]
- Ali, A.; Mohamed, M.T.M.; Siddiqui, Y. Control of Anthracnose by Chitosan through Stimulation of Defence-Related Enzymes in Eksotika II Papaya (Carica Papaya L.) Fruit. J. Biol. Life Sci. Vol 3 No 1 2012 2012.
- Meng, X.; Tian, S. Effects of Preharvest Application of Antagonistic Yeast Combined with Chitosan on Decay and Quality of Harvested Table Grape Fruit. J. Sci. Food Agric. 2009, 89, 1838–1842. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Sivasankari, S.; Venkatesalu, V.; Anantharaj, M.; Chandrasekaran, M. Effect of Seaweed Extracts on the Growth and Biochemical Constituents of Vigna Sinensis. Bioresour. Technol. 2006, 97, 1745–1751. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection against Pathogens in Plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis Cinerea in Grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Klarzynski, O.; Descamps, V.; Plesse, B.; Yvin, J.-C.; Kloareg, B.; Fritig, B. Sulfated Fucan Oligosaccharides Elicit Defense Responses in Tobacco and Local and Systemic Resistance against Tobacco Mosaic Virus. Mol. Plant. Microbe Interact. 2003, 16, 115–122. [Google Scholar] [CrossRef]
- Stadnik, M.J.; Freitas, M.B. de Algal Polysaccharides as Source of Plant Resistance Inducers. Trop. Plant Pathol. 2014, 39, 111–118. [Google Scholar] [CrossRef]
- Schaafsma, G. Safety of Protein Hydrolysates, Fractions Thereof and Bioactive Peptides in Human Nutrition. Eur. J. Clin. Nutr. 2009, 63, 1161. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Sambo, P.; Nicoletto, C.; Santagata, S.; Schiavon, M.; Nardi, S. The Use of Organic Biostimulants in Hot Pepper Plants to Help Low Input Sustainable Agriculture. Chem. Biol. Technol. Agric. 2015, 2. [Google Scholar] [CrossRef]
- Matsubayashi, Y.; Sakagami, Y. Peptide Hormones in Plants. Annu. Rev. Plant Biol. 2006, 57, 649–674. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar Applications of a Legume-Derived Protein Hydrolysate Elicit Dose-Dependent Increases of Growth, Leaf Mineral Composition, Yield and Fruit Quality in Two Greenhouse Tomato Cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Boubaker, H.; Karim, H.; El Hamdaoui, A.; Msanda, F.; Leach, D.; Bombarda, I.; Vanloot, P.; Abbad, A.; Boudyach, E.H.; Ait Ben Aoumar, A. Chemical Characterization and Antifungal Activities of Four Thymus Species Essential Oils against Postharvest Fungal Pathogens of Citrus. Ind. Crops Prod. 2016, 86, 95–101. [Google Scholar] [CrossRef]
- Cindi, M.D.; Soundy, P.; Romanazzi, G.; Sivakumar, D. Different Defense Responses and Brown Rot Control in Two Prunus Persica Cultivars to Essential Oil Vapours after Storage. Postharvest Biol. Technol. 2016, 119, 9–17. [Google Scholar] [CrossRef]
- Soppelsa, S.; Van Hemelrijck, W.; Bylemans, D.; Andreotti, C. Essential Oils and Chitosan Applications to Protect Apples against Postharvest Diseases and to Extend Shelf Life. Agronomy 2023, 13. [Google Scholar] [CrossRef]
- Farzaneh, M.; Kiani, H.; Sharifi, R.; Reisi, M.; Hadian, J. Chemical Composition and Antifungal Effects of Three Species of Satureja (S. Hortensis, S. Spicigera, and S. Khuzistanica) Essential Oils on the Main Pathogens of Strawberry Fruit. Postharvest Biol. Technol. 2015, 109, 145–151. [Google Scholar] [CrossRef]
- Ben-Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme Essential Oil as a Defense Inducer of Tomato against Gray Mold and Fusarium Wilt. Plant Physiol. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef]
- de Lira Mota, K.; de Oliveira Pereira, F.; de Oliveira, W.; Lima, I.; de Oliveira Lima, E. Antifungal Activity of Thymus Vulgaris L. Essential Oil and Its Constituent Phytochemicals against Rhizopus Oryzae: Interaction with Ergosterol. Molecules 2012, 17, 14418–14433. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-X.; Zhang, Y.-D.; Li, N.; Wu, D.-D.; Li, Q.-M.; Chen, Y.-Z.; Zhang, G.-C.; Yang, J. Inhibitory Effect and Mechanism of Action of Juniper Essential Oil on Gray Mold in Cherry Tomatoes. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Maia, A.J.; Oliveira, J.S.B.; Schwan-Estrada, K.R.F.; Faria, C.M.R.; Batista, A.F.; Costa, W.F.; Batista, B.N. The Control of Isariopsis Leaf Spot and Downy Mildew in Grapevine Cv. Isabel with the Essential Oil of Lemon Grass and the Activity of Defensive Enzymes in Response to the Essential Oil. Crop Prot. 2014, 63, 57–67. [Google Scholar] [CrossRef]
- Bulger, M.A.; Ellis, M.A.; L, V. ; Madden Influence of Temperature and Wetness Duration on Infection of Strawberry Flowers by Botrytis Cinerea and Disease Incidence of Fruit Originating from Infected. Am. Phytopathol. Soc. 1987, 77, 1225–1230. [Google Scholar] [CrossRef]
- Wang, S.Y.; Tzeng, D.D.-S. Methionine-Riboflavin Mixtures with Surfactants and Metal Ions Reduce Powdery Mildew Infection in Strawberry Plants. J. Am. Soc. Hortic. Sci. 1998, 123, 987–991. [Google Scholar] [CrossRef]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and Antiproliferative Activities of Strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant Activity of Apple Peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Cellini, A.; Buriani, G.; Rocchi, L.; Rondelli, E.; Savioli, S.; Rodriguez Estrada, M.T.; Cristescu, S.M.; Costa, G.; Spinelli, F. Biological Relevance of Volatile Organic Compounds Emitted during the Pathogenic Interactions between Apple Plants and Erwinia Amylovora. Mol. Plant Pathol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F. Antibacterial and Antifungal Activities of Essential Oils. In Lipids and Essential Oils as Antimicrobial Agents; H. Thormar, 2010.
- Sivakumar, D.; Bautista-Baños, S. A Review on the Use of Essential Oils for Postharvest Decay Control and Maintenance of Fruit Quality during Storage. Crop Prot. 2014, 64, 27–37. [Google Scholar] [CrossRef]
- Bhatia, R.; Shreaz, S.; Khan, N.; Muralidhar, S.; Basir, S.F.; Manzoor, N.; Khan, L.A. Proton Pumping ATPase Mediated Fungicidal Activity of Two Essential Oil Components. J. Basic Microbiol. 2011, 52, 504–512. [Google Scholar] [CrossRef]
- Freiesleben, S.H.; Jager, A. Correlation between Plant Secondary Metabolites and Their Antifungal Mechanisms–a Review. Med. Aromat. Plants 2014, 03. [Google Scholar] [CrossRef]
- Ghannoum, M.A.; Rice, L.B. Antifungal Agents: Mode of Action, Mechanisms of Resistance, and Correlation of These Mechanisms with Bacterial Resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Calderone, R. Exploiting Mitochondria as Targets for the Development of New Antifungals. Virulence 2017, 8, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Ban, X.; Zeng, H.; He, J.; Chen, Y.; Wang, Y. The Mechanism of Antifungal Action of Essential Oil from Dill (Anethum Graveolens L.) on Aspergillus Flavus. PLoS ONE 2012, 7, e30147. [Google Scholar] [CrossRef]
- Wei, Y.; Wei, Y.; Xu, F.; Shao, X. The Combined Effects of Tea Tree Oil and Hot Air Treatment on the Quality and Sensory Characteristics and Decay of Strawberry. Postharvest Biol. Technol. 2018, 136, 139–144. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Testolin, R.; Zanotelli, D.; Andreotti, C. Effect of Biostimulants on Apple Quality at Harvest and After Storage. Agronomy 2020, 10, 1214. [Google Scholar] [CrossRef]
- Benhamou, N.; Bélanger, R.R. Benzothiadiazole-Mediated Induced Resistance to Fusarium Oxysporum f. Sp. Radicis-Lycopersici in Tomato. Plant Physiol. 1998, 118, 1203–1212. [Google Scholar] [CrossRef]
- Hien Dao, T.T.; Puig, R.C.; Kim, H.K.; Erkelens, C.; Lefeber, A.W.M.; Linthorst, H.J.M.; Choi, Y.H.; Verpoorte, R. Effect of Benzothiadiazole on the Metabolome of Arabidopsis Thaliana. Plant Physiol. Biochem. 2009, 47, 146–152. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, W.; Bi, Y.; Luo, Y. Postharvest BTH Treatment Induces Resistance of Peach (Prunus Persica L. Cv. Jiubao) Fruit to Infection by Penicillium Expansum and Enhances Activity of Fruit Defense Mechanisms. Postharvest Biol. Technol. 2005, 35, 263–269. [Google Scholar] [CrossRef]
- Skłodowska, M.; Gajewska, E.; Kuźniak, E.; Mikiciński, A.; Sobiczewski, P. BTH-Mediated Antioxidant System Responses in Apple Leaf Tissues. Sci. Hortic. 2010, 125, 34–40. [Google Scholar] [CrossRef]
- Terry, L.A.; Joyce, D.C. Suppression of Grey Mould on Strawberry Fruit with the Chemical Plant Activator Acibenzolar. Pest Manag. Sci. Former. Pestic. Sci. 2000, 56, 989–992. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Qiu, X.; Moore, P.H.; Borth, W.; Hu, J.; Ferreira, S.; Albert, H.H. Systemic Acquired Resistance Induced by BTH in Papaya. Physiol. Mol. Plant Pathol. 2003, 63, 237–248. [Google Scholar] [CrossRef]
- Cellini, A.; Fiorentini, L.; Buriani, G.; Yu, J.; Donati, I.; Cornish, D.A.; Novak, B.; Costa, G.; Vanneste, J.L.; Spinelli, F. Elicitors of the Salicylic Acid Pathway Reduce Incidence of Bacterial Canker of Kiwifruit Caused by Pseudomonas Syringae Pv. Actinidae. Ann. Appl. Biol. 2014, 165, 441–453. [Google Scholar] [CrossRef]
- Bokshi, A.I.; Jobling, J.; McConchie, R. A Single Application of Milsana® Followed by Bion® Assists in the Control of Powdery Mildew in Cucumber and Helps Overcome Yield Losses. J. Hortic. Sci. Biotechnol. 2008, 83, 701–706. [Google Scholar] [CrossRef]
- Sturchio, E.; Donnarumma, L.; Annesi, T.; Milano, F.; Casorri, L.; Masciarelli, E.; Zanellato, M.; Meconi, C.; Boccia, P. Essential Oils: An Alternative Approach to Management of Powdery Mildew Diseases. Phytopathol. Mediterr. 2014, 53, 385–395. [Google Scholar]
- Mostafa, Y.S.; Hashem, M.; Alshehri, A.M.; Alamri, S.; Eid, E.M.; Ziedan, E.-S.H.E.; Alrumman, S.A. Effective Management of Cucumber Powdery Mildew with Essential Oils. Agriculture 2021, 11. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; Di Tommaso, G.; D’Amico, S.; Borgo, M.; Faoro, F. New Chitosan Formulation Prevents Grapevine Powdery Mildew Infection and Improves Polyphenol Content and Free Radical Scavenging Activity of Grape and Wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Faoro, F.; Maffi, D.; Cantu, D.; Iriti, M. Chemical-Induced Resistance against Powdery Mildew in Barley: The Effects of Chitosan and Benzothiadiazole. BioControl 2008, 53, 387–401. [Google Scholar] [CrossRef]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K. Seaweed Extract Reduces Foliar Fungal Diseases on Carrot. Crop Prot. 2008, 27, 1360–1366. [Google Scholar] [CrossRef]
- Franssen, H.J.; Bisseling, T. Peptide Signaling in Plants. Proc. Natl. Acad. Sci. 2001, 98, 12855–12856. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, K.; Casson, S.; Chilley, P. Peptides: New Signalling Molecules in Plants. Trends Plant Sci. 2002, 7, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Sonawala, U.; Dinkeloo, K.; Danna, C.H.; McDowell, J.M.; Pilot, G. Review: Functional Linkages between Amino Acid Transporters and Plant Responses to Pathogens. Plant Sci. 2018, 277, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, M.; Perazzolli, M.; Nesler, A.; Giovannini, O.; Pertot, I. The Effect of Hydrolysis and Protein Source on the Efficacy of Protein Hydrolysates as Plant Resistance Inducers against Powdery Mildew. J. Bioprocess. Biotech. 2017, 07. [Google Scholar] [CrossRef]
Figure 1.
Climatic conditions: average daily temperature (T°C) and relative humidity (RH %) inside the greenhouse during the experimental period.
Figure 1.
Climatic conditions: average daily temperature (T°C) and relative humidity (RH %) inside the greenhouse during the experimental period.
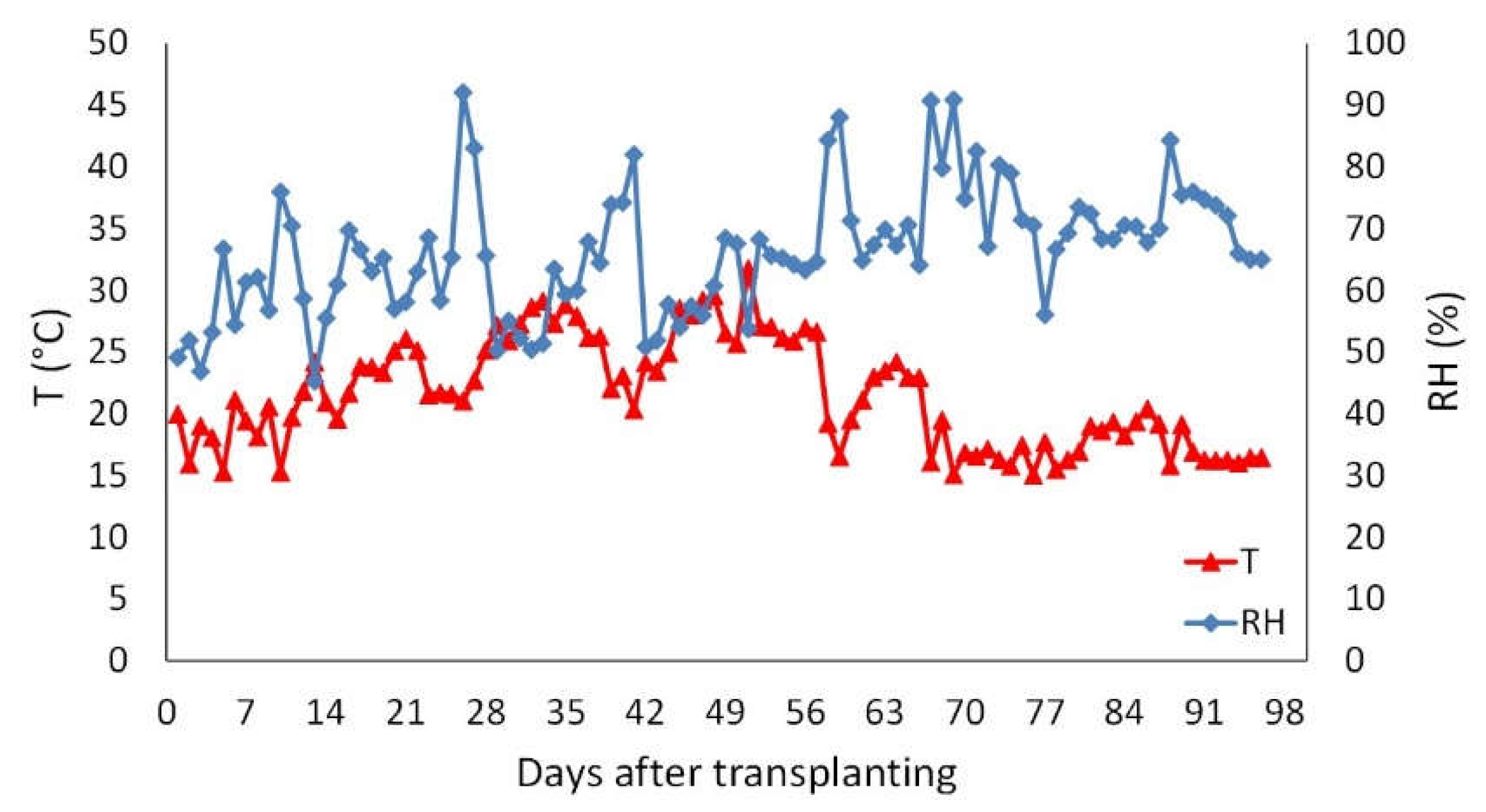
Figure 2.
Incidence of flowers affected by natural infections by Botrytis cinerea at the end of harvest (56 days after flowering). Different letters at the top of each bar indicate significant differences among treatments according to LSD test at p < 0.05 (n=4). Treatments’ legend: CON, control; FUN, fungicide; BTH, benzothiadiazole; CHI, chitosan; SEA, seaweed extract; APH, alfalfa protein hydrolysate; THY, thyme essential oil; JUN, juniper essential oil.
Figure 2.
Incidence of flowers affected by natural infections by Botrytis cinerea at the end of harvest (56 days after flowering). Different letters at the top of each bar indicate significant differences among treatments according to LSD test at p < 0.05 (n=4). Treatments’ legend: CON, control; FUN, fungicide; BTH, benzothiadiazole; CHI, chitosan; SEA, seaweed extract; APH, alfalfa protein hydrolysate; THY, thyme essential oil; JUN, juniper essential oil.
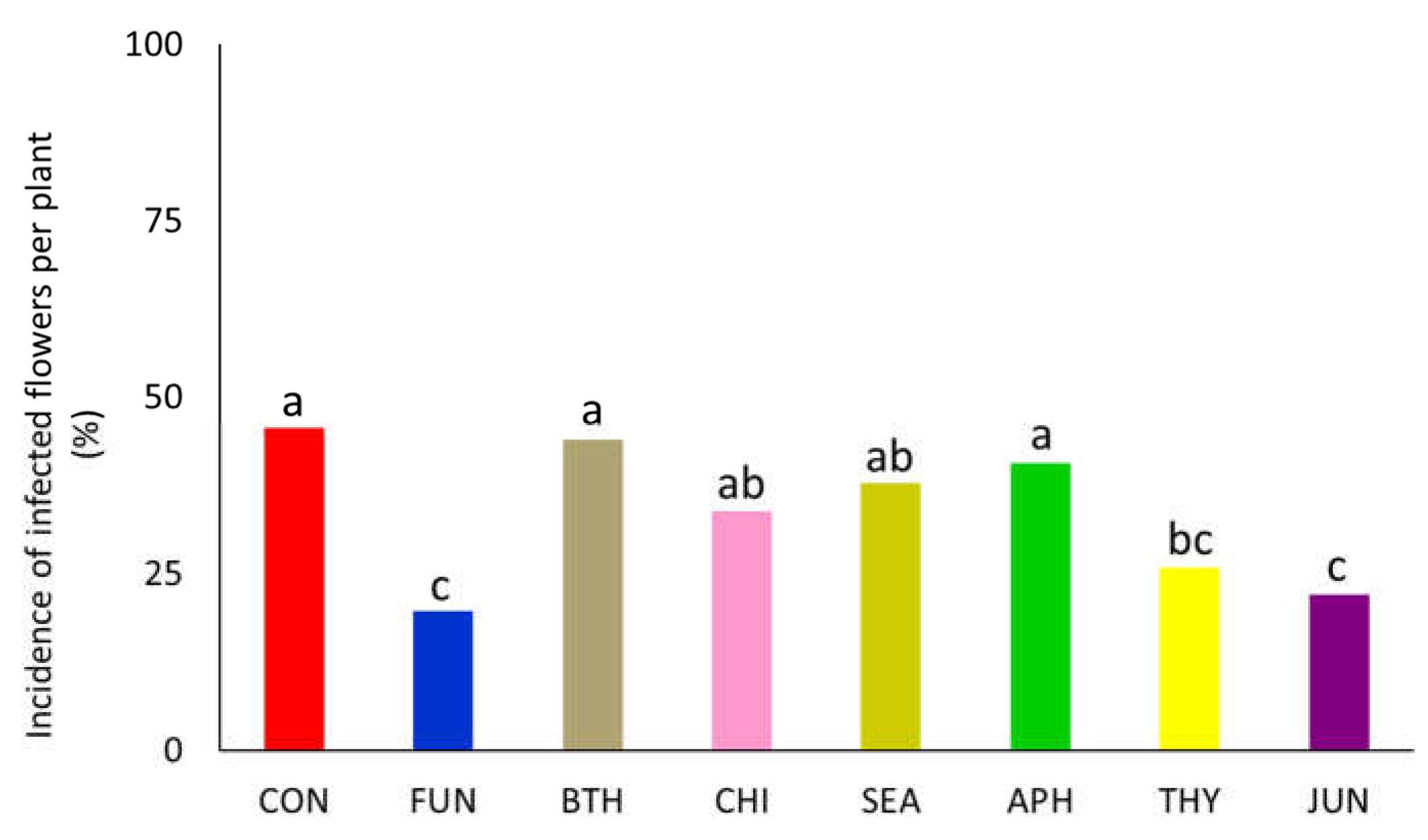
Figure 3.
Powdery mildew incidence on strawberry leaves after inoculation (A) and severity at 56 days after inoculation (B). Tested products were applied 7 days before inoculation at BBCH 71 (Experiment 2). Different letters at the top of each bar indicate significant differences among treatments according to LSD test at p < 0.05 (n=8). ns: not significant.
Figure 3.
Powdery mildew incidence on strawberry leaves after inoculation (A) and severity at 56 days after inoculation (B). Tested products were applied 7 days before inoculation at BBCH 71 (Experiment 2). Different letters at the top of each bar indicate significant differences among treatments according to LSD test at p < 0.05 (n=8). ns: not significant.
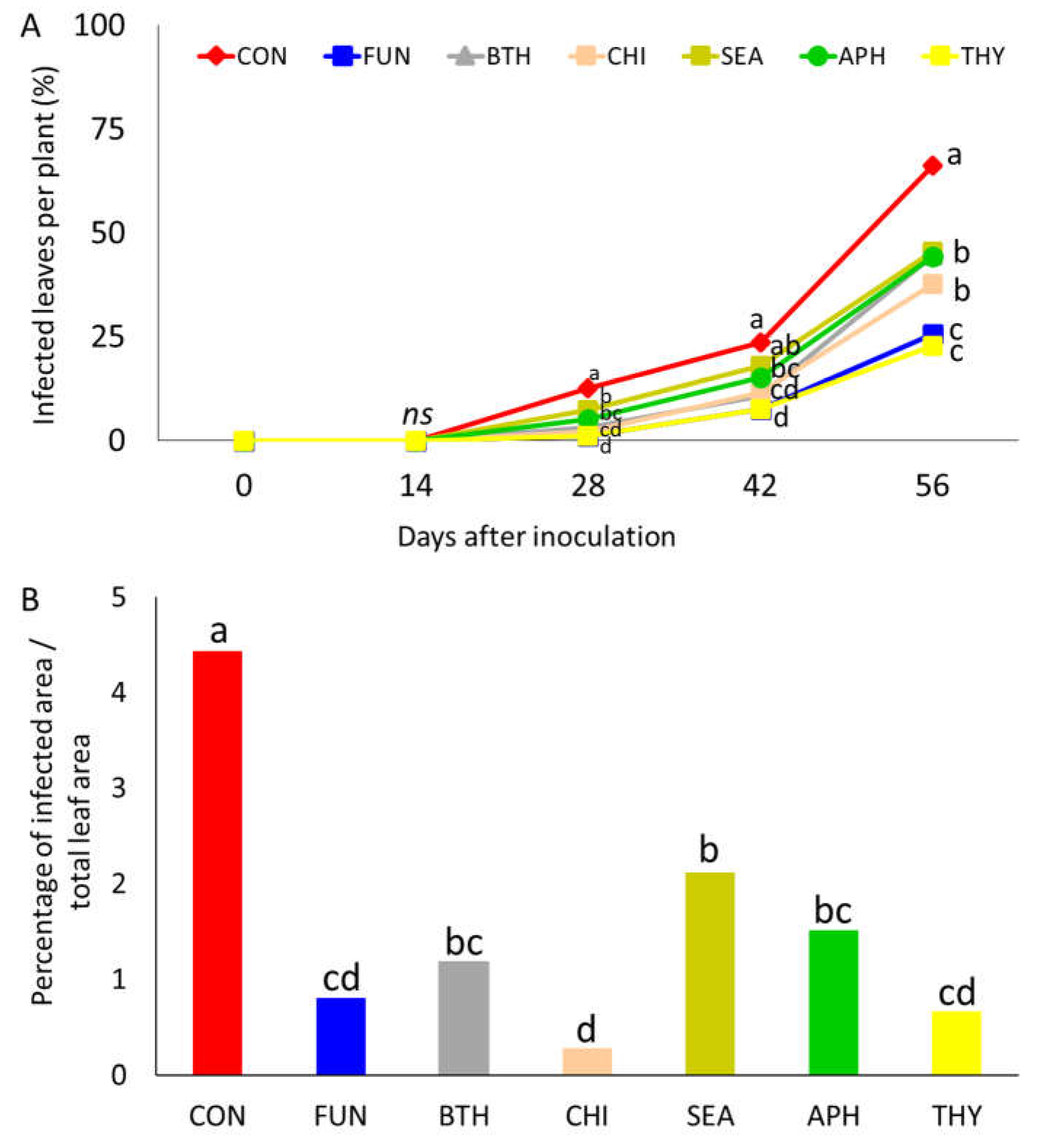
Figure 4.
Powdery mildew incidence on strawberry leaves after inoculation (A) and severity at 56 days after inoculation (B). Tested products were applied 24 hours after inoculation at BBCH 73 (Experiment 3). Different letters at the top of each bar indicate significant differences among treatments according to LSD test at p < 0.05 (n=8). ns: not significant.
Figure 4.
Powdery mildew incidence on strawberry leaves after inoculation (A) and severity at 56 days after inoculation (B). Tested products were applied 24 hours after inoculation at BBCH 73 (Experiment 3). Different letters at the top of each bar indicate significant differences among treatments according to LSD test at p < 0.05 (n=8). ns: not significant.
Figure 5.
Expression of defence-related genes (FaPR1 (A), FaPR5 (B), FaPR10 (C), FaEDS1 (D) and FaLOX (E)) in strawberry leaves treated with benzothiadiazole, seaweed extracts, chitosan and thyme essential oil. RT-qPCR was performed using FaGPDH2 as housekeeping gene. Values were normalized to the control at each time point. Data are expressed as mean ± SE.
Figure 5.
Expression of defence-related genes (FaPR1 (A), FaPR5 (B), FaPR10 (C), FaEDS1 (D) and FaLOX (E)) in strawberry leaves treated with benzothiadiazole, seaweed extracts, chitosan and thyme essential oil. RT-qPCR was performed using FaGPDH2 as housekeeping gene. Values were normalized to the control at each time point. Data are expressed as mean ± SE.
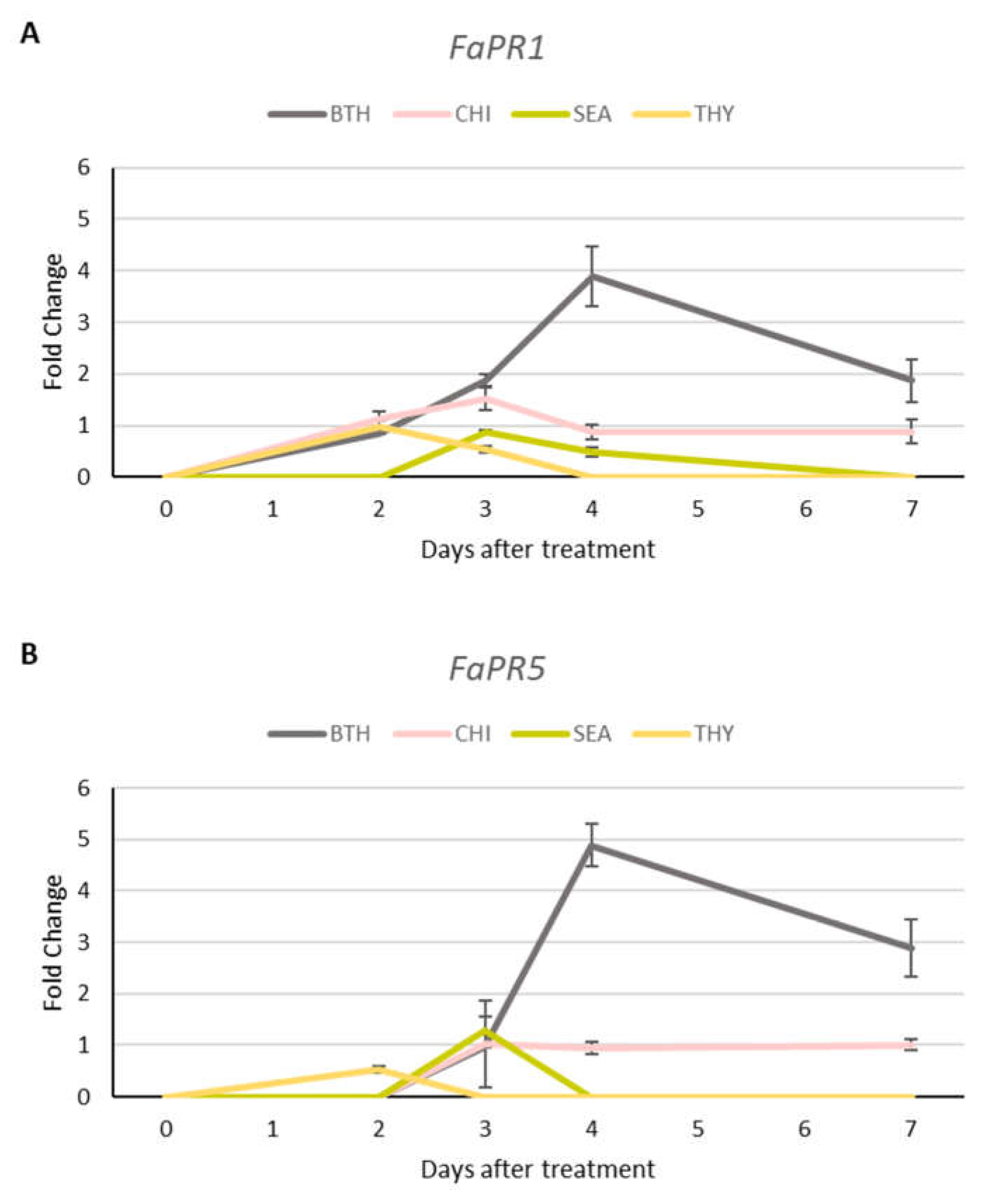
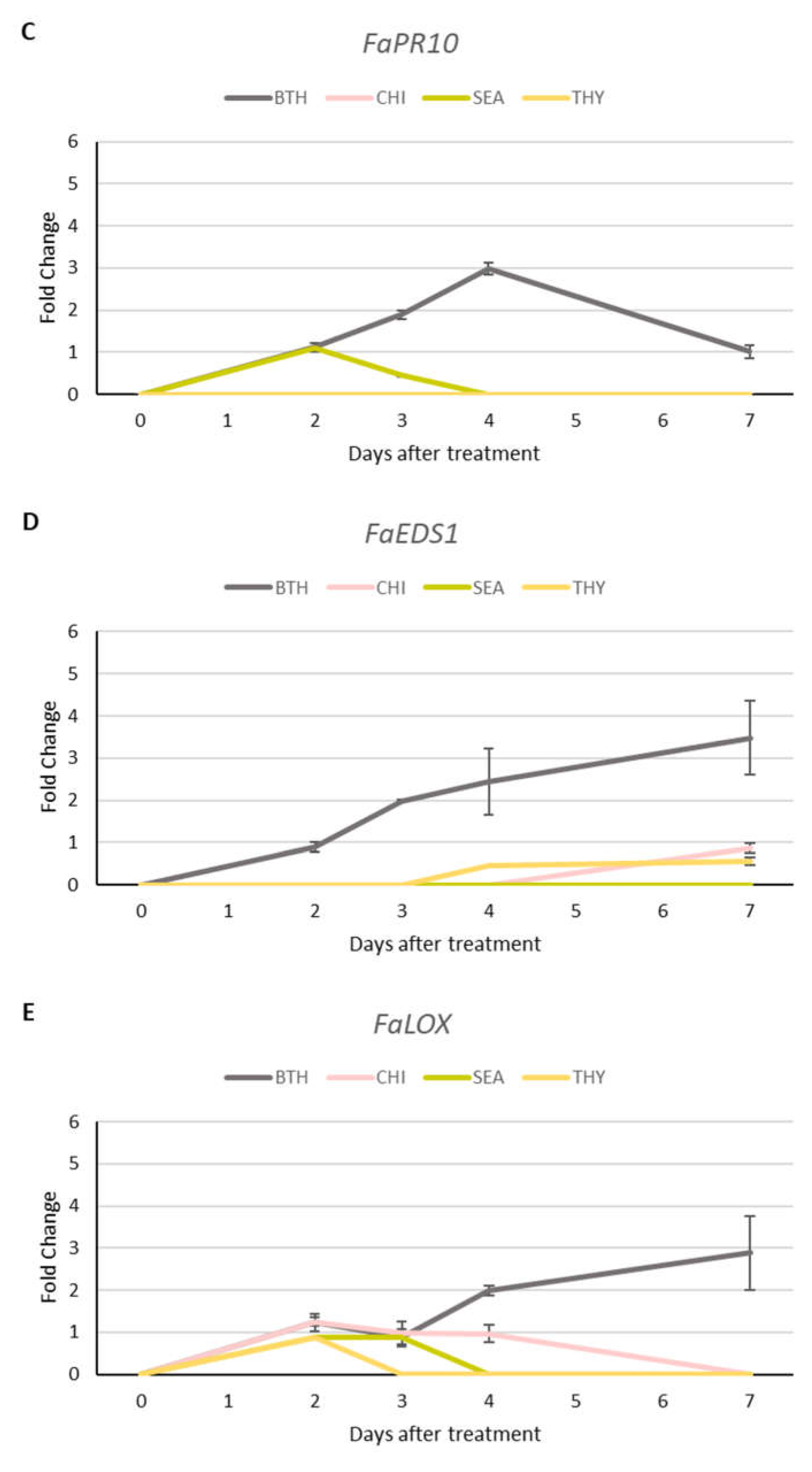
Table 1.
Product characteristics and mode of application.
| Treatment | Name | Active ingredient | Concentration | Commercial name |
|---|---|---|---|---|
| CON | Untreated | water | - | - |
| FUN | Fungicide | penconazole | 0.25 mL L-1 | Topas® 200 EW, Syngenta Crop protection, Italy |
| BTH | Benzothiadiazole | acibenzolar-S-methyl | 0.4 g L-1 | Bion® 50 WG, Syngenta Crop protection, Italy |
| CHI | Chitosan solution | chitosan | 10 mL L-1 | ChitoPlant Solution, Agritalia, Italy |
| SEA | Seaweed extract (Ascophyllum nodosum) |
mix of components | 4 g L-1 | Experimental product, ILSA S.p.a., Italy |
| APH | Alfalfa protein hydrolysate | mix of components | 4 g L-1 | Experimental product, ILSA S.p.a., Italy |
| THY | Thyme essential oil (Thymus vulgaris) |
thymol | 1 mL L-1 | Essential oil, Vitalis Dr. Joseph, Italy |
| JUN | Juniper essential oil (Juniperus communis) |
α-pinene | 1 mL L-1 | Essential oil, Vitalis Dr. Joseph, Italy |
Table 2.
List of primers used for RT-qPCR analysis.
| Gene name | Gene description | Forward primer sequence [5’ – 3’] |
Reverse primer sequence [5’ – 3’] |
|---|---|---|---|
| FaPR1 | Pathogenesis related protein 1 | TGCTAATTCACATTATGGCG | GTTAGAGTTGTAATTATAGTAGG |
| FaPR5 | Pathogenesis related protein 5 | CGATGCCCCTGCTTACAGTTACCCTAAGGATG | CCTCGTAATTGCTTCAAGGGCAGAACACAACC |
| FaPR10 | Pathogenesis related protein 10 | CGAGGAATACAACTAAACCTTGCCGTCT | TACAATTTGCCACACATACACCGAAGTG |
| FaEDS1 | Enhanced disease susceptibility 1 | AAAAGAGAGACTTCAATGCCAATGTG | CTTCGTTCTTTGCGTGTCTGTAGTAGTT |
| FaLOX | Lipoxygenase | CGACGACGACTGGATACACCGCAGGG | GAGGTTGGCCGCTGTTTCTTGCACCGTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



