
Submitted:
22 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
The floating freshwater fern Azolla is the only plant that retains an endocyanobiont, Nostoc azollae (aka Anabaena azollae) during its sexual and asexual reproduction. The increased interest in Azolla as a potential source of food, and its unique evolutionary history have raised questions about its cyanotoxin content and genome. Cyanotoxins are potent toxins synthesized by cyanobacteria which have an anti-herbivore effect, but also have been linked to neurodegenerative disorders including Alzheimer’s and Parkinson's diseases, liver and kidney failure, muscle paralysis and other severe health issues. In this study, we investigated 48 accessions of the Azolla-Nostoc symbiosis for the presence of genes coding microcystin, nodularin, cylindrospermopsin and saxitoxin, and BLAST analysis for anatoxin-a. We also investigated the presence of the neurotoxin β-N-methylamino-L-alanine (BMAA) in Azolla and N. azollae through LC-MS/MS. The PCR amplification of saxitoxin, cylindrospermospin, microcystin, and nodularin genes showed that Azolla and its cyanobiont N. azollae do not have the genes to synthesize these cyanotoxins. Additionally, the matching of the anatoxin-a gene to the sequenced N. azollae genome does not indicate the presence of the anatoxin-a gene. The LC-MS/MS analysis showed that BMAA and its isomers AEG and DAB are absent from Azolla and Nostoc azollae. Azolla therefore has the potential to safely feed millions of people due to its rapid growth while free-floating on shallow fresh water without the need for nitrogen fertilizers.
Keywords:
Azolla
; Food
; Nostoc
; Anabaena
; symbiosis
; BMAA
; microcystins
; nodularin
; anatoxin-a
; cylindrospermopsin
; saxitoxin
1. Introduction
Azolla Lam. is the only plant with a permanent nitrogen-fixing cyanobacterial symbiont (cyanobiont) that has chains of cells (filaments) comprising photosynthetic vegetative cells and thicker-walled heterocysts that contain the nitrogen-fixing enzyme nitrogenase [1]. The cyanobiont has been assigned to both Anabaena azollae and Nostoc azollae because its morphology resembles free-living species of both genera, including their change into motile hormogonia and akinetes (resting cells) that ensure survival during stressed conditions [2].
Genetic and paleontological data indicate that the Azolla – N. azollae symbiosis originated 80 million years ago in North America following Whole Genome Duplication (WGD) that increased the genome of Azolla’s immediate ancestor [3]. Nostoc azollae’s subsequent coevolution with Azolla caused extensive changes in the cyanobiont’s genome compared to free-living species of Anabaena and Nostoc [3,4,5,6,7,8]. Some changes involved the upregulation of genes that enhanced N. azollae’s sequestration of atmospheric nitrogen and provision of nitrogen-based compounds to Azolla, increasing the plant’s speed of growth free-floating on fresh water. The downregulation, loss, or conversion to pseudogenes of other genes changed N. azollae’s ancestors from independent free-living organisms into obligate endosymbionts, reflecting N. azollae‘s permanent location inside the leaves and female megasporocarps of Azolla. These included genes that previously expressed proteins involved in the synthesis of carotenoid and chlorophyll pigments, so that A. azollae is reliant on Azolla’s cellular pigments for protection against photooxidative damage [6].
Colonies of N. azollae live in specialized cavities in Azolla’s dorsal floating leaves, providing nitrogen-based nutrients to the plant that enable it to double its biomass in less than two days while free-floating on fresh water [9,10]. As a result, Azolla has been used for hundreds of years in India and the Far East as a nitrogen biofertilizer for paddy rice, reducing mosquito breeding populations by 95% [11,12,13] and emissions of the potent greenhouse gas methane from paddies by 25-50% [14,15,16]. Azolla also provides livestock feed, biofuel and biofertilizer for other plants, alleviating shortages of the ‘three Fs’ that increasingly threaten food supplies globally: feed, fuel, and fertilizer. It absorbs and removes phosphates and nitrates from water contaminated by chemical fertilizers, industrial pollutants, and animal and human waste that trigger toxic cyanobacterial (aka blue-green algal) blooms in rivers and lakes. The symbionts’ combined CO2 sequestration also increases Azolla’s carbon capture, so that it can sequester large amounts of atmospheric CO2, with the plants being compressed and stored to reduce anthropogenic climate change through Carbon Capture and Storage (CCS). Azolla can, therefore, mitigate many of the threats arising from a Perfect Storm as our population increases by more than a million every three days. Its remarkable properties are increasingly recognised, and it has designated as a unique superorganism by [1].
Azolla has the potential to help feed millions of people because of its rapid growth, ease of outdoor cultivation in tropical and temperate regions, plus global production using the indoor Azolla Biosystem described by Bujak & Bujak (2020) in ‘The Azolla Story’ [2]. The use of Azolla for human consumption was thought to be limited by its high total polyphenolic content (TPC), but Winstead et al. (2024) [17] showed that the TPC of raw Azolla caroliniana, which is native in the eastern United States, have only 4.26 g gallic acid equivalent (GAE) kg−1 DW, and that simple cooking methods can decrease TPC in all Azolla species. They also demonstrated that its protein content is 19% DW and its apparent protein digestibility is 78.45%, with a yield 173 g FW m−2 day−1 and 5.53 g DW m−2 day−1, confirming Azolla‘s potential for cultivation and domestication as a nutritious food. This raises the question of whether Azolla is safe to eat because of the presence of harmful cyanotoxins in many cyanobacteria.
Cyanotoxins are produced by cyanobacteria of the genera Anabaena and Nostoc among others and include some of the most powerful natural poisons that target the nervous system (neurotoxins: BMAA, saxitoxin and anatoxin-a), the liver (hepatotoxins: microcystins and nodularins), protein synthesis and DNA modification (cylindrospermopsin) and the skin (dermatoxins; nodularin). Cyanotoxins are alkaloids (anatoxin-a, saxitoxin, cylindrospermopsin) or peptides (pentapeptide nodularin or the heptapeptide microcystin) [18,19,20,21]. Upon their release in water, they are ingested by zooplankton and animals, or absorbed by phytoplankton and plants that can have acute or chronic effects when eaten by humans. This is a global health issue owing to bioconcentration and bioaccumulation in the food chain and poisoning through ingestion of contaminated food, so that cyanotoxins are now widely analyzed and studied to determine their effects on plants and animals [22]. For example, the World Health Organization (WHO) recommends a value of 1 μg/L for microcystin-LR in drinking water [23].
BMAA (β-N-methylamino-L-alanine) is a non-proteinogenic amino acid is produced by free-living cyanobacteria in marine, freshwater, and terrestrial environments [24,25]. It has been detected in plants with endosymbiotic cyanobacteria including lichens, hornworts, the leaf petioles of the tropical flowering plant Gunnera, and the cycad Cycas circinalis [24,25,26] and linked to the amyotrophic lateral sclerosis/parkinson-dementia complex (ALS/PDC) detected among the Chamorro people living on the Pacific island of Guam [24,25]. BMAA, like other cyanotoxins, can be biomagnified in seafood eaten by people, including fish [27,28], shrimps [29], mussels, oysters, and crabs [30]. BMAA can also be synthetized by eukaryotes such as diatoms [31] and dinoflagellates [32] which are food sources for crustaceans, fish and shellfish [33].
These observations raise the question of whether eating Azolla may be harmful to humans due to the possible production of BMAA and other cyanotoxins by N. azollae. Unlike free-living Anabaena and Nostoc, the loss or conversion to pseudogenes of genes involved in cyanotoxin and/or BMAA production may have occurred in N. azollae because they were no longer needed by the permanently enclosed cyanobiont. The following analyses were therefore undertaken on all seven extant Azolla species and their cyanobionts to determine if they can be safely eaten by people:
- The presence of genes coding for microcystin, nodularin, cylindrospermopsin and saxitoxin.
- The presence of the anatoxin-a/homoanatoxin-a gene cluster by bioinformatic tools.
- The presence of BMAA.
The seven examined species of Azolla are A. caroliniana, A. filiculoides, A. mexicana, A. microphylla, A. nilotica, A. rubra and A. pinnata including its two subspecies A. pinnata subsp. pinnata and A. pinnata subsp. imbricata. Table 1 lists the 48 accessions that provided the Azolla species and subspecies used in this study.
2. Results
2.1. The Cyanotoxins Microcystin, Nodularin, Saxitoxin, Cylindrospermopsin and Anatoxin-a
The PCR amplification of 12 genes that encode for four cyanotoxins (cylindrospermopsin, nodularin, saxitoxin and microcystin) was determined for all seven Azolla species and two A. pinnata subspecies from 48 Azolla accessions listed in Table 1. Global distribution of the accession countries is shown in Supplementary Material Figure S25. The results indicate that cyanotoxin gene amplification on Azolla accessions were negative when matched with positive and negative controls for 12 genes: cyl, mcy A, mcy B, mcy B domain A, mcy C, mcy C domain A, mcy D domain ACP, mcy D domain KS, mcyE/ndaF, mcy E domain GSA-AMT, mcy G domain CM, sxt.
The results also showed that same 12 genes in N. azollae isolated from the 47 Azolla accessions were not amplified compared with positive and negative controls. This indicates that both Azolla and N. azollae do not have genes that biosynthesize those cyanotoxins. Photographs of all gels from the PCR amplifications are shown in Supplementary Material Figures S1 to S24.
The BLAST search for the anatoxin-a and homoanatoxin-a gene cluster showed a query cover of only 3% and percent identity of 73.04%. Most of these alignments were partial segments comprising less than 250 bp of the anaH transposase gene with identities around 70%. The aligned genes in N. azollae are only associated with pseudogenes and not with protein coding genes. All other alignments were partial, and none included any of the whole genes associated with the anatoxin-a biosynthesis gene cluster.
2.2. Detection of BMAA (β-N-methylamino-L-alanine)
The detection of the non-proteinogenic amino acid BMAA by LC-MS/MS with method 1 on all seven Azolla species (A. caroliniana, A. filiculoides, A. microphylla, A. mexicana, A. nilotica, A. rubra and the two subspecies of A. pinnata) showed that the BMAA was absent from both Azolla and its cyanobiont N. azollae since there is not found the retention time for BMAA and the isomer 2,4-DAB (2,4-diaminobutyric acid) when compared with their standards which are not shown).
Re-analysis of six of the Azolla species (A. rubra that was not re-analyzed) using method 2, in which the samples were derivatized, corroborated the results obtained with method 1. This indicates that the Azolla species shown in Figure 1 did not show the retention times for BMAA (RT=13.92 min) and the two isomers 2,4-DAB (RT=14.93 min), and AEG (N-(2-aminoethyl)-glycine) (RT=13.22 min) when compared with their standards (Figure 2).
3. Discussion
Most genera of free-living cyanobacteria synthesize cyanotoxins and include cyanobacteria that have temporary symbiosis with some plants, so that the host plant has the potential to assimilate and bioaccumulate the cyanotoxins, discussed above. There are few published studies documenting genetic and chromatographic detection of cyanotoxins in cyanobionts. Cyanobacteria from lichens have been analysed and contain genes that encode nodularin and microcystin, and can translate the peptides nodularin and microcystins [34,35,36]. The synthesis of those two cyanotoxins may be linked to the temperature and humidity in which the lichens grow and may be important for the maintenance of lichens in diverse ecological habitats [37].
Unlike lichens, the fern Azolla has a permanent symbiosis with the cyanobacteria N. azollae giving this symbiosis a unique evolution pattern and, ultimately, the loss of genes by the cyanobiont [4,5,6,7]. Our genetic analyses show, for or the first time, that all seven Azolla species and their cyanobiont, N. azollae, do not possess genes associated with the synthesis of microcystin, nodularin, saxitoxin, cylindrospermopsin, anatoxin-a, and homoanatoxin-a. The biosynthetic pathways of microcystins [38] and anatoxin-a/homoanatoxin-a [39] are a multi-step process that requires several genes to synthesize both cyanotoxins. All genes associated with microcystin synthesis were, therefore, amplified by specific primers with Azolla and N. azollae DNA, and the complete anatoxin-a gene cluster was BLAST searched against the N. azollae genome. There were no matches, supporting the model that N. azollae lost the ability to synthesise microcystin, nodularin, saxitoxin, cylindrospermopsin, anatoxin-a, and homoanatoxin-a due to downregulation of cyanotoxin biosynthesis genes or loss of the genes during the co-evolution of Azolla and N. azollae.
BMAA was isolated in 1967 from seeds of Cycas circinalis (cycad) [24,25,26] and identified as the primary cause of amyotrophic lateral sclerosis/parkinson-dementia complex (ALS/PDC) in the Chamorro people on the Pacific island of Guam [40,41], with the high levels of the neurotoxin resulting from biomagnification through the food chain [27,28,33,42,43]. BMAA is a cyanotoxin that can cross the blood-brain barrier where it forms a reservoir [44]) and can be inserted into proteins instead of the amino acid L-serine, causing protein misfolding and aggregation [45,46,47]. BMAA can induce changes in the expression of genes in brain cells and thus resulting in a wide range of other neurodegenerative disorders [19]. Alzheimer’s, Parkinson’s and other neurological diseases including amyotrophic lateral sclerosis (ALS), progressive supranuclear palsy (PSP) and dementia with Lewy bodies (DLB) [48] may therefore be partially caused or facilitated by BMAA. However, the gene/genes for the codification of BMAA are not known in any cyanobacteria and plant, so that their presence can only be detected by analytical methods, including those used in this study.
BMAA was detected in plant-cyanobacteria symbiosis such as hornworts, liverwort, lichens, cycads and Gunnera [24] and also in A. filiculoides with 2 μg/g [42]. Some analytical methods to detect this non-proteinogenic molecule can result in erroneous interpretations due to structural isomers DAB (2,4-diaminobutyric) and AEG (N-(2-aminoethyl)-glycine which can co-elute and be mis-identified as BMAA [49,50]. For the present study, two methods were therefore used to detect BMAA, DAB and AEG, and did not detect BMAA, AEG, or DAB in any of the analyzed Azolla and N. azollae. These data indicate that the previous reported detection of 2 μg/g BMAA in Azolla [42] is incorrect.
Harmful algal blooms (HABs) of other cyanobacteria species also release cyanotoxins upon cell necrosis. The uptake and bioaccumulation of cyanotoxins from irrigated water for crop and non-crop therefore also needs to be evaluated to determine if Azolla species may bioaccumulate cyanotoxins. Previous studies show that A. filiculoides does now uptake or bioaccumulate microcystin [51] or cylindrospermospin [52], confirming that Azolla can be safely eaten.
4. Materials and Methods
4.1. Azolla Accessions and Culturing
The seven Azolla species including two A. pinnata subspecies from the germplasm collection at IRRI (International Rice Research Institute) and two A. filiculoides accessions from Portugal (FI-BGLU and FI-BGM) were used to detect the cyanotoxin genes of microcystin, nodularin, cylindrospermospin and saxitoxin by PCR, and BMAA by LC-MS/MS (Table 1.) The 48 Azolla accessions have a global distribution from 33 countries (Supplementary Material, Figure S25). The Azolla species were cultured in Hoagland medium (H-40), pH 6.1-6.2, at controlled temperature, photoperiod, and light intensity [53]. The biomass was collected, washed in distilled water, frozen at -80°C, lyophilized, and then weighed.
4.2. Isolation of Nostoc azollae from Azolla accessions
Nostoc azollae cyanobionts were isolated from the dorsal foliar cavities of 48 Azolla accessions (Table 1) using the gentle roller method [54,55] with the following modifications. Roots were cut-off and sporophytes were disinfected in aqueous sodium hypochlorite (1 ml NaClO:10 ml distilled water, v:v) for 20 minutes, followed by three washes in ultrapure water (Millipore, Madrid, Spain). Sporophytes were sectioned and squeezed with a roller to separate the cyanobiont from Azolla cavities. The extract (Azolla+water+N. azollae) was centrifuged twice at 3000 g for 3 minutes to settle fern debris. The recovered supernatant with N. azollae filaments was centrifuged twice at 1000 g for 1 minute to free cyanobionts from the cellular debris. The recovered dark-green pellet was centrifuged at 11000 g for 10 minutes, stored at -20ºC, frozen at -80°C, lyophilized and weighted.
4.3. Detection and Analysis of BMAA (β-N-Methylamino-L-Alanine)
4.3.1. Method 1
The methodology, including reagents and materials, described by Baptista et al. (2015) [56] was used., with extraction of BMAA and quantification by LC-MS/MS using validated analytical methods [50,56]. Lyophilized Azolla and N. azollae (10 mg each sample) were acid-digested in 6 M HCl at 90°C for 20 minutes, using a high-pressure microwave system (Milestone-Ethos 1). After evaporation with nitrogen, 20 mM of HCl was added to samples and filtered (0.22 μm Millipore).
Analyses of BMAA by LC-MS/MS were performed in a Thermo LCQ Fleet Ion Trap LC/MSn system (Thermo Scientific) using a 2.1×100 mm, 5-μm diameter ZIC-HILIC column (SeQuant) and a 14×1 mm, 5-μm guard column (SeQuant). The mobile phase was acetonitrile (0.1% formic acid) and deionized water (0.1% formic acid). A linear gradient of 90% acetonitrile for 20 minutes was followed by 60% acetonitrile for 15 minutes and 90% acetonitrile for 5 minutes. The flow rate was 0.5 ml min-1, injection volume was 10 μl, column temperature at 40°C and the positive mode on the electrospray ionization (ESI). Nitrogen was the sheath gas at a rate of 45 (unitless), and auxiliary gas at a rate of 20 (unitless). The capillary temperature was held at 250°C. Mass-to-charge ratio (m/z) scan was performed from 50 to 150 and the ion m/z 119 to assess 2,4-DAB (2,4-diaminobutyric acid) was monitored. The occurrence of the product ions m/z 102, 88 and 76 was verified at collision energy of 14 V for the presence of BMAA.
4.3.2. Method 2
Method 2 followed that described by Pravadali-Cekic S. et al. (2023) [49] with some modifications for the amount of starting material, chromatographic column, eluents and mode of mass detection. Lyophilized Azolla biomass, (100 mg) was dissolved in 3 ml of trichloacetic acid (TCA) 10% (v/v) and sonicated on ice (5 min, 70% amplitude, 20Hz), followed by overnight precipitation at 4°C. The mixture was then centrifuged (5000 g, 15 min, 4°C), the supernatant reserved and the pellet submitted to a second extraction cycle. The third extraction step used 10% TCA/acetone. The pellet as the bound fraction was transferred to a glass vial with acetone (100 %), centrifuged, and the supernatant added to the_free fraction. The pooled free fraction was then evaporated to dry in a SpeedVac and kept at -80°C. Pellets were also dried using the SpeedVac and acid hydrolysis by adding 3 ml of 6 M HCl overnight at 110°C. The hydrolyzed pellet was re-suspended in 1 ml ultrapure water and added to the free fraction. Samples were then filtered through a This was carried out with a 20 µl standard mix solution or sample extract, 20 µl of derivatizing reagent, and 60 µl of borate buffer. Following AccQ-Tag Ultra Derivatisation Kit in accordance with the manufacturer’s guidelines, the mixture was vortexed for several seconds and placed in a thermocycler at 55°C for 10 minutes. The final extract was then transferred to a 1.5mL vial for LC/MS/MS analysis.
Samples were injected in a Liquid Chromatograph Thermo Finnigan Surveyor HPLC System (Thermo Scientific, MA, USA), coupled with a Mass Spectrometry LCQ Fleet™ Ion Trap Mass Spectrometer (Thermo Scientific, MA, USA). XcaliburTM version 2. Mass Spectrometer Tune Method parameters optimization was used for data acquisition and processing using direct injection of BMMA and co-occurring isomers in a solution of 1 ppm in LCMS grade water (Table 2). The Mass Spectrometer operated in electrospray positive polarity mode using Collision Ionisation Dissociation (CID) corresponding to the [M+H]+ BMAA, AEG (N-(2-aminoethyl)-glycine), and 2,4-DAB molecules ion precursors and respective diagnostic fragments. The spray voltage was maintained at 3.5 kV, capillary temperature at 350°C, and capillary voltage at 20 kV and tube lens at 120 kV. Nitrogen was used as the sheath and auxiliary gas, with collision energy at 20 eV in Colission Induced Dissociation Mode. Separation was achieved on an ACE Excel C18 (50 × 2.1 mm I.D., 1.7 μm, Batch: V17-1253, Avantor® ACE ®, VWR, PT) at 18°C, with a flow rate of 0.3 ml/min injected at a volume of 10 μL in a no-waste mode. The eluents used were methanol (A) and water (B), both acidified with formic acid at 0.1% (v/v). The gradient program started at 13% A, increasing to 90% A in 20 minutes, turning back to initial conditions in 5 minutes, equilibrating for an additional 10 minutes with 20% A. See Table 2 for the chromatographic and mass parameters.
4.4. Cyanotoxin Genes at Azolla Accessions and Nostoc Azollae
4.4.1. DNA Extraction
DNA from N. azollae was extracted with PureLink® Genomic DNA MiniKit (Invitrogen, Carlsbad, California, USA) and DNA from Azolla accessions was extracted with Genomic DNA from Plant NucleoSpin® Plant II (Macherey-Nagel, Düren, Germany) according to the manufacturer instructions. DNA was stored at -20°C. The DNA was quantified in a Qubit fluorometer (Invitrogen) using the Quant-iT® dsDNA HS assay following the manufacturer instructions. A working DNA concentration of 0.1 μg/μl was made with sterile ultrapure water, and saxitoxin, nodularin, and microcystin were assessed by specific primers (Table 3). A Biometra TProfessional (Goettingen) thermocycler was used for PCR amplification using the conditions listed in Table 4 for each gene, with a hold at 4°C for all the programs. Each 20 μl reaction contained 1 μl of 0.5 µM of each primer (Invitrogen), 2 μl of 0.1 μg/μl DNA, 9 μl Supreme NZYTaq 2x Green Master Mix (NZYTech) and 7 μl of ultrapure sterile water. Negative (with sterile ultrapure water) and positive (Microcystis aeruginosa LEGE91094 for microcystin and microcystin/nodularin genes, Aphanizomenon ovalisporum for cylindrospermopsin gene and Aphanizomenon gracillaris LMECYA 40 from INSA for saxitoxin gene) controls were included. The amplification products were separated in 1.5% agarose gel electrophoresis running in TAE 1x at 150 V, 25-30 min and stained with 0.2 μg/ml ethidium bromide (BioRad). The 1 Kb Plus DNA ladder (Invitrogen) was used as molecular size marker.
4.4.3. BLAST of Anatoxin-a Genes against Nostoc azollae
To determine if N. azollae produces anatoxin-a, a nucleotide BLAST (BLASTN) search for anatoxin-a coding genes was performed. Since the anatoxin-a gene cluster was discovered after the PCR and gel analysis of the other cyanotoxins performed in this study, analysis of its presence was made separately through BLAST rather than as the query sequence [57]. This was a 34682 bp sequence encoding for proteins associated with the biosynthesis of these toxins. The nucleotide query was applied to the full genome of Nostoc azollae 0708 (taxid: 551115). Matches with E-values from the BLASTN less than 0.01 were investigated and analyzed.
5. Conclusions
Our LC-MS/MS results show the Azolla-Nostoc azollae superorganism does not contain BMAA or their isomers DAB and AEG, and that Azolla and N. azollae do not synthesize other common cyanotoxins, indicating that Azolla is a nutritious food that can be safely eaten.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Materials 1, Figures S1-S24: Agarose gel from the PCR amplification of genes in Azolla and N. azollae. Figure S25: Map showing the countries of Azolla accessions origin used in the present study. Figure S1: Agarose gel from the PCR amplification of the gene microcystin/nodularin synthetase for nodularin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder; Figure S2: Agarose gel from the PCR amplification of the gene saxitoxin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (A. gracillaris LMECYA 40), C-: negative control, 1st line is Ladder. Figure S3: Agarose gel from the multiplex PCR amplification of the genes poliketide synthase and peptide synthase for cylindrospermopsin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (A. ovalisporum), C-: negative control, 1st line is Ladder. Figure S4: Agarose gel from the PCR amplification of the gene microcystin synthetase (mcy A) for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S5: Agarose gel from the PCR amplification of the gene microcystin synthetase (mcy B) for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S6: Agarose gel from the PCR amplification of the gene microcystin C for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S7: Agarose gel from the PCR amplification of the gene microcystin B A-domain for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S8: Agarose gel from the PCR amplification of the gene microcystin C A-domain for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S9: Agarose gel from the PCR amplification of the gene microcystin D ACP-domain for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S10: Agarose gel from the PCR amplification of the gene microcystin D KS-domain for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S11: Agarose gel from the PCR amplification of the gene microcystin E GSA-AMT-domain for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S12: Agarose gel from the PCR amplification of the gene microcystin G CM-domain for microcystin in 48 Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S13: Agarose gel from the PCR amplification of the gene microcystin/nodularin synthetase for nodularin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S14: Agarose gel from the PCR amplification of the gene saxitoxin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (A. gracillaris LMECYA 40), C-: negative control, 1st line is Ladder. Figure S15: Agarose gel from the multiplex PCR amplification of the genes poliketide synthase and peptide synthase for cylindrospermopsin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (A. ovalisporum), C-: negative control, 1st line is Ladder. Figure S16: Agarose gel from the PCR amplification of the gene microcystin synthetase (mcy A) for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S17: Agarose gel from the PCR amplification of the gene microcystin synthetase (mcy B) for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S18: Agarose gel from the PCR amplification of the gene microcystin C for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S19: Agarose gel from the PCR amplification of the gene microcystin B A-domain for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S20: Agarose gel from the PCR amplification of the gene microcystin C A-domain for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S21: Agarose gel from the PCR amplification of the gene microcystin D ACP-domain for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S22: Agarose gel from the PCR amplification of the gene microcystin D KS-domain for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S23: Agarose gel from the PCR amplification of the gene microcystin E GSA-AMT-domain for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S24: Agarose gel from the PCR amplification of the gene microcystin G CM-domain for microcystin in 47 Nostoc azollae isolated from Azolla accessions (see Table 1 from the manuscript). C+ -: positive control (M. aeruginosa LEGE 91094), C-: negative control, 1st line is Ladder. Figure S25: Map showing the countries of Azolla accessions origin used in the present study. Legend: a – A. pinnata subsp. imbricata; b - A. filiculoides; c – A. mexicana; d – A. caroliniana; e – A. microphylla; f - A. nilotica; g – A. rubra; h – A. pinnata subsp. Pinnata. Supplementary Materials 2: Sequence alignment from the BLASTN query between the Anatoxin-a gene cluster and the Nostoc azollae genome.
Author Contributions
Conceptualization, Jonathan Bujak, Ana L. Pereira, Alexandra Bujak, Victor Leshyk, Timo Stadtlander and Daniel Winstead; Formal analysis, Ana L. Pereira, Joana Azevedo, Vitor Vasconcelos and Daniel Winstead; Investigation, Ana L. Pereira, Vitor Vasconcelos and Daniel Winstead; Methodology, Jonathan Bujak, Ana L. Pereira, Joana Azevedo, Timo Stadtlander and Daniel Winstead; Project administration, Jonathan Bujak; Resources, Joana Azevedo; Supervision, Jonathan Bujak; Validation, Ana L. Pereira; Visualization, Jonathan Bujak, Ana L. Pereira, Alexandra Bujak, Victor Leshyk and Daniel Winstead; Writing – original draft, Jonathan Bujak, Ana L. Pereira, Alexandra Bujak and Daniel Winstead; Writing – review & editing, Ana L. Pereira, Alexandra Bujak and Daniel Winstead.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carrapiço, F. Azolla as a Superorganism. Its Implication in Symbiotic Studies. In Symbioses and Stress: Joint Ventures in Biology; Seckbach, J., Grube, M., Eds.; Springer Netherlands: Dordrecht, 2010; pp. 225–241 ISBN 978-90-481-9449-0.
- Singh, P.; Khan, A.; Srivastava, A. Chapter 16 - Heterocyst and Akinete Differentiation in Cyanobacteria: A View toward Cyanobacterial Symbiosis. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press, 2020; pp. 235–248 ISBN 978-0-12-819311-2.
- Bujak, J.P.; Bujak, A.A. Origin and Evolution of the Azolla Superorganism. Plants 2024, 13, Article number 2106 (online publication with no page numbers). [CrossRef]
- Ekman, M.; Tollbäck, P.; Klint, J.; Bergman, B. Protein Expression Profiles in an Endosymbiotic Cyanobacterium Revealed by a Proteomic Approach. Mol. Plant-Microbe Interactions® 2006, 19, 1251–1261. [CrossRef]
- Ekman, M.; Tollbäck, P.; Bergman, B. Proteomic Analysis of the Cyanobacterium of the Azolla Symbiosis: Identity, Adaptation, and NifH Modification. J. Exp. Bot. 2008, 59, 1023–1034. [CrossRef]
- Larsson, J.; Nylander, J.A.; Bergman, B. Genome Fluctuations in Cyanobacteria Reflect Evolutionary, Developmental and Adaptive Traits. BMC Evol. Biol. 2011, 11, 187. [CrossRef]
- Ran, L.; Larsson, J.; Vigil-Stenman, T.; Nylander, J.A.A.; Ininbergs, K.; Zheng, W.-W.; Lapidus, A.; Lowry, S.; Haselkorn, R.; Bergman, B. Genome Erosion in a Nitrogen-Fixing Vertically Transmitted Endosymbiotic Multicellular Cyanobacterium. PLoS ONE 2010, 5, e11486. [CrossRef]
- Bujak, J.; Bujak, A. The Azolla Story: A Message from the Future.; The Azolla Foundation, 2020; ISBN 1-5272-8335-6.
- Bujak, A.; Bujak, J. Azolla’s Use as a Biofertilizer and Livestock Feed. In Ferns; Marimuthu, J., Fernández, H., Kumar, A., Thangaiah, S., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 671–695 ISBN 9789811661693.
- Watanabe, I.; Berja, N.S. The Growth of Four Species of Azolla as Affected by Temperature. Aquat. Bot. 1983, 15, 175–185. [CrossRef]
- Ansari, M.A.; Sharma, V.P. Role of Azolla in Controlling Mosquito Breeding in Ghaziabad District Villages (U.P.). Indian J. Malariol. 1991, 28, 51–54.
- Mwingira, V.; Mayala, B.; Senkoro, K.; Rumisha, S.; Shayo, H., Elizabeth; Mlozi, P.; Mboera, L. Mosquito Larval Productivity in Rice-Fields Infested with Azolla in Mvomero District, Tanzania. Tanzan. J. Health Res. 2009, 11, 17–22. [CrossRef]
- Rajendran, R.; Reuben, R. Evaluation of the Water Fern Azolla Microphylla for Mosquito Population Management in the Rice-Land Agro-Ecosystem of South India. Med. Vet. Entomol. 1991, 5, 299–310. [CrossRef]
- Bharati, K. Influence of Incorporation or Dual Cropping of Azolla on Methane Emission from a Flooded Alluvial Soil Planted to Rice in Eastern India. Agric. Ecosyst. Amp Environ. 2000.
- Mujiyo; Sunarminto, B.; Hanudin, E.; Widada, J.; Syamsiyah, J. Methane Emission on Organic Rice Experiment Using Azolla. Int. J. Appl. Environ. Sci. 2016, 11, 295–308.
- Xu, H.; Zhu, B.; Liu, J.; Li, D.; Yang, Y.; Zhang, K.; Jiang, Y.; Hu, Y.; Zeng, Z. Azolla Planting Reduces Methane Emission and Nitrogen Fertilizer Application in Double Rice Cropping System in Southern China. Agron. Sustain. Dev. 2017, 37, 29. [CrossRef]
- Winstead, D.; Di Gioia, F.; Jauregui, M.; Jacobson, M. Nutritional Properties of Raw and Cooked Azolla Caroliniana Willd., an Aquatic Wild Edible Plant. Food Sci. Nutr. 2024, 12, 2050–2060. [CrossRef]
- Bláha, L.; Babica, P.; Maršálek, B. Toxins Produced in Cyanobacterial Water Blooms - Toxicity and Risks. Interdiscip. Toxicol. 2009, 2, 36–41. [CrossRef]
- Burton, B.; Collins, K.; Brooks, J.; Marx, K.; Renner, A.; Wilcox, K.; Moore, E.; Osowski, K.; Riley, J.; Rowe, J.; et al. The Biotoxin BMAA Promotes Dysfunction via Distinct Mechanisms in Neuroblastoma and Glioblastoma Cells. PLOS ONE 2023, 18, e0278793. [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management.; CRC Press, 1999;
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water - Second Edition; CRC Press, 2021; ISBN 978-1-00-308144-9.
- Merel, S.; D, W.; R, C.; S, S.; E, B.; O, T. State of Knowledge and Concerns on Cyanobacterial Blooms and Cyanotoxins. Environ. Int. 2013, 59. [CrossRef]
- Funari, E.; Testai, E. Human Health Risk Assessment Related to Cyanotoxins Exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse Taxa of Cyanobacteria Produce β-N-Methylamino-l-Alanine, a Neurotoxic Amino Acid. Proc. Natl. Acad. Sci. 2005, 102, 5074–5078. [CrossRef]
- Esterhuizen, M.; Downing, T.G. Beta-N-Methylamino-L-Alanine (BMAA) in Novel South African Cyanobacterial Isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [CrossRef]
- Vega, A.; Bell, E.A. α-Amino-β-Methylaminopropionic Acid, a New Amino Acid from Seeds of Cycas Circinalis. Phytochemistry 1967, 6, 759–762. [CrossRef]
- Al-Sammak, M.A.; Hoagland, K.D.; Cassada, D.; Snow, D.D. Co-Occurrence of the Cyanotoxins BMAA, DABA and Anatoxin-a in Nebraska Reservoirs, Fish, and Aquatic Plants. Toxins 2014, 6, 488–508. [CrossRef]
- Hammerschlag, N.; Davis, D.A.; Mondo, K.; Seely, M.S.; Murch, S.J.; Glover, W.B.; Divoll, T.; Evers, D.C.; Mash, D.C. Cyanobacterial Neurotoxin BMAA and Mercury in Sharks. Toxins 2016, 8, 238. [CrossRef]
- Holtcamp, W. The Emerging Science of BMAA: Do Cyanobacteria Contribute to Neurodegenerative Disease? Environ. Health Perspect. 2012, 120, a110–a116. [CrossRef]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA Exposure in an Amyotrophic Lateral Sclerosis Cluster from Southern France. PloS One 2013, 8, e83406. [CrossRef]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A Novel Source for the Neurotoxin BMAA in Aquatic Environments. PloS One 2014, 9, e84578. [CrossRef]
- Jiang, L.; Ilag, L. Detection of Endogenous BMAA in Dinoflagellate (Heterocapsa Triquetra) Hints at Evolutionary Conservation and Environmental Concern. PubRaw Sci. 2014, 1, 1–8.
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in Shellfish from Two Portuguese Transitional Water Bodies Suggests the Marine Dinoflagellate Gymnodinium Catenatum as a Potential BMAA Source. Aquat. Toxicol. Amst. Neth. 2014, 152, 131–138. [CrossRef]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria Produce a High Variety of Hepatotoxic Peptides in Lichen Symbiosis. Proc. Natl. Acad. Sci. 2012, 109, 5886–5891. [CrossRef]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Lichen Species Identity and Diversity of Cyanobacterial Toxins in Symbiosis. New Phytol. 2013, 198, 647–651.
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.; Fieker, C.; Neilan, B.A. Nodularin, a Cyanobacterial Toxin, Is Synthesized in Planta by Symbiotic Nostoc Sp. ISME J. 2012, 6, 1834–1847.
- Koksharova, O.A.; Safronova, N.A. Non-Proteinogenic Amino Acid β-N-Methylamino-L-Alanine (BMAA): Bioactivity and Ecological Significance. Toxins 2022, 14, 539.
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes Coding for Hepatotoxic Heptapeptides (Microcystins) in the Cyanobacterium Anabaena Strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [CrossRef]
- Méjean, A.; Paci, G.; Gautier, V.; Ploux, O. Biosynthesis of Anatoxin-a and Analogues (Anatoxins) in Cyanobacteria. Toxicon 2014, 91, 15–22.
- Kurland, L.T.; Mulder, D.W. Epidemiologic Investigations of Amyotrophic Lateral Sclerosis. 2. Familial Aggregations Indicative of Dominant Inheritance. I. Neurology 1955, 5, 182–196. [CrossRef]
- Kurland, L.T.; Mulder, D.W. Epidemiologic Investigations of Amyotrophic Lateral Sclerosis. 2. Familial Aggregations Indicative of Dominant Inheritance. II. Neurology 1955, 5, 249–268. [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of Cyanobacterial Neurotoxins and Neurodegenerative Disease among the Chamorro People of Guam. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 13380–13383. [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A Mechanism for Slow Release of Biomagnified Cyanobacterial Neurotoxins and Neurodegenerative Disease in Guam. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 12228–12231. [CrossRef]
- Xie, X.; Basile, M.; Mash, D.C. Cerebral Uptake and Protein Incorporation of Cyanobacterial Toxin β-N-Methylamino-L-Alanine. NeuroReport 2013, 24, 779. [CrossRef]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. Beta-N-Methylamino-L-Alanine Enhances Neurotoxicity through Multiple Mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [CrossRef]
- Rush, T.; Liu, X.; Lobner, D. Synergistic Toxicity of the Environmental Neurotoxins Methylmercury and β-N-Methylamino-L-Alanine. Neuroreport 2012, 23, 216–219. [CrossRef]
- Weiss, J.H.; Koh, J.Y.; Choi, D.W. Neurotoxicity of Beta-N-Methylamino-L-Alanine (BMAA) and Beta-N-Oxalylamino-L-Alanine (BOAA) on Cultured Cortical Neurons. Brain Res. 1989, 497, 64–71. [CrossRef]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The Non-Protein Amino Acid BMAA Is Misincorporated into Human Proteins in Place of L-Serine Causing Protein Misfolding and Aggregation. PloS One 2013, 8, e75376. [CrossRef]
- Pravadali-Cekic, S.; Vojvodic, A.; Violi, J.P.; Mitrovic, S.M.; Rodgers, K.J.; Bishop, D.P. Simultaneous Analysis of Cyanotoxins β-N-Methylamino-L-Alanine (BMAA) and Microcystins-RR, -LR, and -YR Using Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS). Molecules 2023, 28, 6733. [CrossRef]
- Faassen, E.J.; Gillissen, F.; Lürling, M. A Comparative Study on Three Analytical Methods for the Determination of the Neurotoxin BMAA in Cyanobacteria. PLoS One 2012, 7, e36667.
- Pereira, A.L.; Monteiro, B.; Azevedo, J.; Campos, A.; Osório, H.; Vasconcelos, V. Effects of the Naturally-Occurring Contaminant Microcystins on the Azolla Filiculoides–Anabaena Azollae Symbiosis. Ecotoxicol. Environ. Saf. 2015, 118, 11–20.
- Santos, C.; Azevedo, J.; Campos, A.; Vasconcelos, V.; Pereira, A.L. Biochemical and Growth Performance of the Aquatic Macrophyte Azolla Filiculoides to Sub-Chronic Exposure to Cylindrospermopsin. Ecotoxicology 2015, 24, 1848–1857. [CrossRef]
- Pereira, A.; Carrapico, F. Culture of Azolla Filiculoides in Artificial Conditions. Plant Biosyst. 2009, 2009, 431–434. [CrossRef]
- Peters, G.A.; Mayne, B.C. The Azolla, Anabaena Azollae Relationship: I. Initial Characterization of the Association. Plant Physiol. 1974, 53, 813–819.
- Rai, A.K.; Rai, V. Effect of NaCl on Growth, Nitrate Uptake and Reduction and Nitrogenase Activity of Azolla Pinnata–Anabaena Azollae. Plant Sci. 2003, 164, 61–69.
- Baptista, M.S.; Vasconcelos, R.G.W.; Ferreira, P.C.; Almeida, C.M.R.; Vasconcelos, V.M. Assessment of the Non-Protein Amino Acid BMAA in Mediterranean Mussel Mytilus Galloprovincialis after Feeding with Estuarine Cyanobacteria. Environ. Sci. Pollut. Res. 2015, 22, 12501–12510. [CrossRef]
- Méjean, A.; Mann, S.; Maldiney, T.; Vassiliadis, G.; Lequin, O.; Ploux, O. Evidence That Biosynthesis of the Neurotoxic Alkaloids Anatoxin-a and Homoanatoxin-a in the Cyanobacterium Oscillatoria PCC 6506 Occurs on a Modular Polyketide Synthase Initiated by l -Proline. J. Am. Chem. Soc. 2009, 131, 7512–7513. [CrossRef]
Figure 1.
Total ion chromatogram of six Azolla species for the detection of BMAA with method 2. From top to bottom: A. caroliniana (CA 3001), A. filiculoides (FI 1507), A. pinnata subsp. pinnata (PP 7001), A. nilotica (NI 5001), A. mexicana (ME 2026), A. microphylla (MI 4021), A. pinnata subsp. imbricata (PI 1).
Figure 1.
Total ion chromatogram of six Azolla species for the detection of BMAA with method 2. From top to bottom: A. caroliniana (CA 3001), A. filiculoides (FI 1507), A. pinnata subsp. pinnata (PP 7001), A. nilotica (NI 5001), A. mexicana (ME 2026), A. microphylla (MI 4021), A. pinnata subsp. imbricata (PI 1).
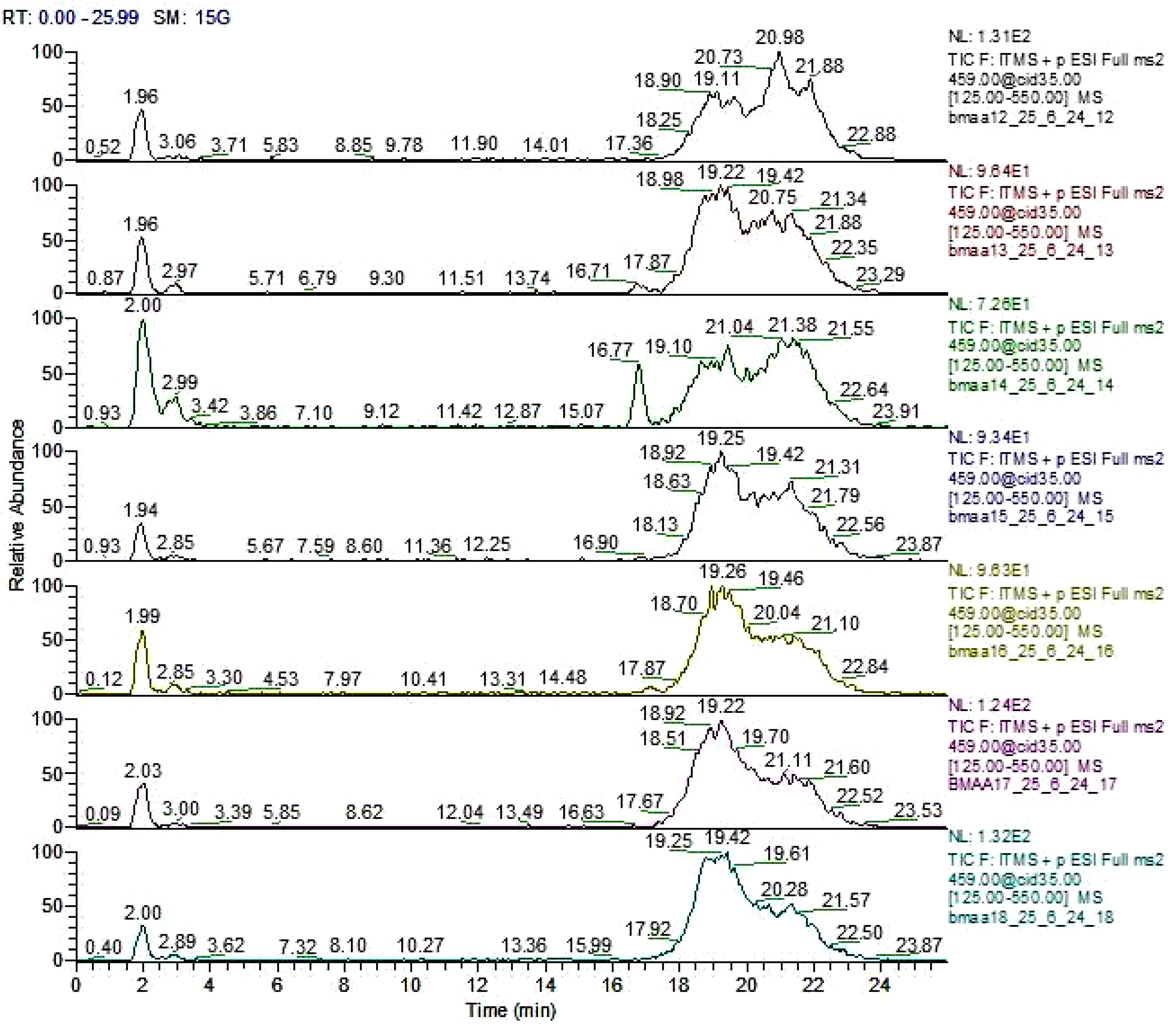
Figure 2.
Total ion chromatogram (left) and CID spectra (right) of derivatized standards at 1 ppm. BMAA (top, RT=13.92 min), AEG (middle, RT=13.22 min), and 2,4-DAB (bottom, RT=14.93 min).
Figure 2.
Total ion chromatogram (left) and CID spectra (right) of derivatized standards at 1 ppm. BMAA (top, RT=13.92 min), AEG (middle, RT=13.22 min), and 2,4-DAB (bottom, RT=14.93 min).
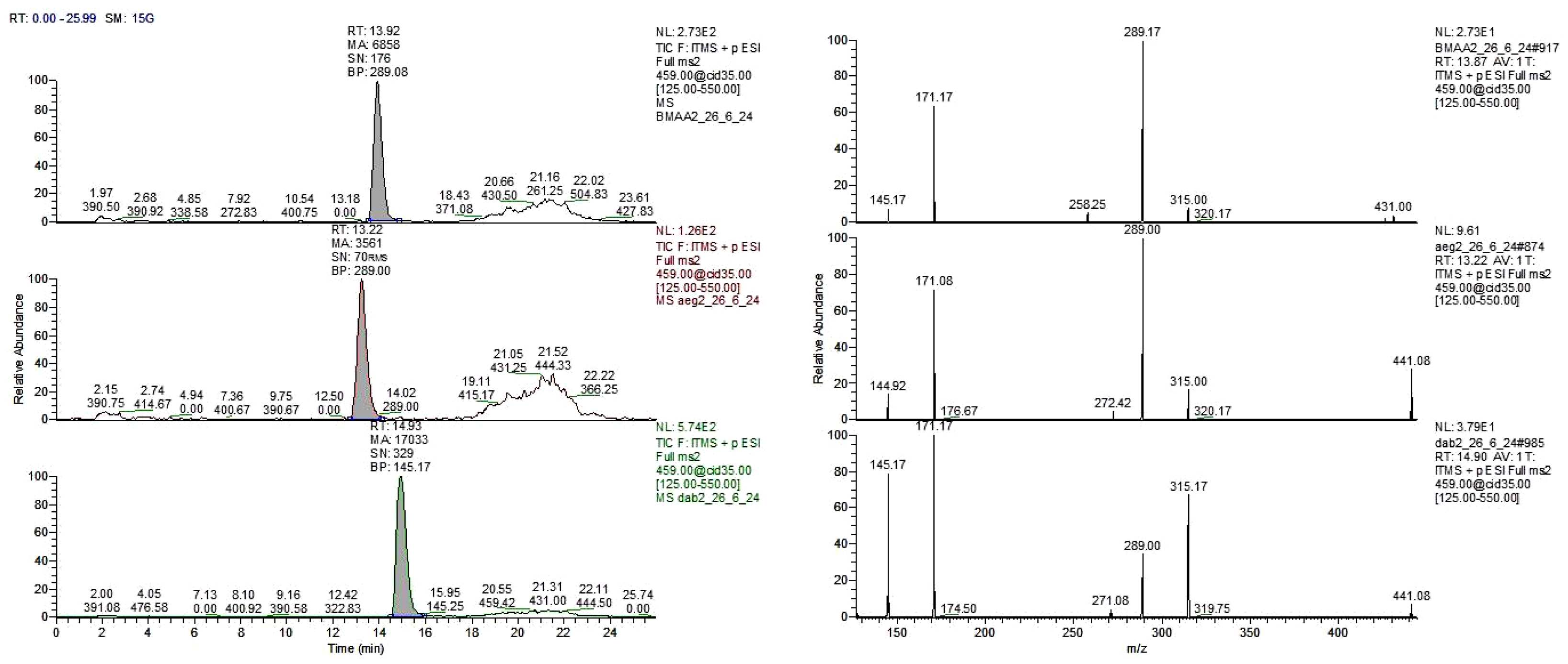

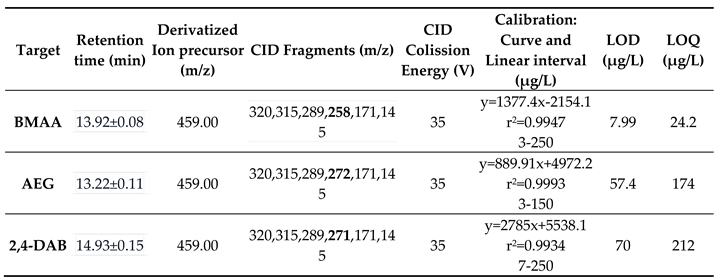
Table 1.
List of Azolla accessions from worldwide countries.
| Accessiona | Species name | Origin and harvest year | Sourceb/collector |
|---|---|---|---|
| PI1*,$ | A. pinnata subsp. imbricata | Philippines, Sto Domingo, Albay, 1975 | IRRI |
| PI2 | Malaysia, Bumbong Lima, Butterworth, 1977 | IRRI | |
| PI23 | India, Cuttack, Orissa, 1978 | CRRI | |
| PI68 | Sri Lanka, Tissa, 1984 | S. Kulasooriya | |
| PI102 | Japan, Okinawa, 1987 | O. Mochida | |
| PI503 | Australia, Murdoch, 1978 | M. Dilworth | |
| PI531 | Indonesia, Bali, 1983 | - | |
| PI540 | China, Putian, 1989 | C. van Hove | |
| FI1001* | A. filiculoides | East Germany (ex- GDR), 1979 | IB China |
| FI1008 | USA, Cranmore Road, Sutter Co., California, 1981 | D. Rains | |
| FI1010 | Peru, PUFFI, Lima, 1982 | CIAT | |
| FI1042 | Brazil, Parana, 1987 | I. Watanabe | |
| FI1052 | South of France, North of Lyon, 1989 | P. Roger | |
| FI1090 | Japan, Tanabe-cho, 1992 | S. Kitoh | |
| FI1501 | Belgian, Harchies, 1987 | A. Lawalree | |
| FI1505 | South Africa, Verwoerd dam, 1987 | D. Toerien | |
| FI1507$ | Colombia, Zipaquira, 1987 | Y. Lopez | |
| FI1522 | Switzerland, Zurich Botanical Garden, 1987 | - | |
| FI-BGLU | Botanical Garden of Lisbon University, 2009 | A.L. Pereira | |
| FI-BGM | Botanical Garden of Madeira, Funchal, 2010 | C. Lobo | |
| ME2001* | A. mexicana | USA, Graylodge, California, 1978 | D. Rains |
| ME2008 | Colombia, CIAT, Cali, 1982 | CIAT | |
| ME2011 | Japan, Osaka, 1984 | T. Lumpkin | |
| ME2026$ | Brazil, Solimoes river, Pacencia Island, Iranduba, Amazonas (BLCC 18), 1984 | T. Lumpkin | |
| CA3001*,$ | A. caroliniana | USA, Ohio, 1978 | D. Rains |
| CA3017 | Brazil, Rio Grande do Sul, 1987 | I. Watanabe | |
| CA3502 | Egypt, Moshtohr University, 1987 | C. Myttenaere | |
| CA3507 | Suriname, Boxel, 1987 | H. Lardinois | |
| CA3513 | Zimbabwe, Causeway Botanical Garden, 1987 | T. Muller | |
| CA3524 | Holland, 1987 | E. Ohoto | |
| CA3525 | Ruanda, Cyili Rice Research Center, 1987 | C. van Hove | |
| MI4018* | A. microphylla | Paraguay, 1981 | D. Rains |
| MI4021$ | Equator, Santa Cruz Island, Galapagos, 1982 | T. Lumpkin | |
| MI4028 | Philippines, hybrid (MI4018xFI1001), 1985 | Do Van Cat | |
| MI4054 | Brazil, Baía, 1987 | I. Watanabe | |
| MI4510 | Philippines, Los Baños, IRRI, 1987 | C. van Hove | |
| NI5001*,$ | A. nilotica | Sudan, Kosti, 1982 | T. Lumpkin |
| NI5002# | Sudan, Kosti, 1989 | T. Lumpkin | |
| NI5501 | Burundi, Bujumbura, 1987 | J. Bouharmont | |
| RU6010* | A. rubra | New Zealand, Nouville, 1986 | C. van Hove |
| RU6502 | Australia, Victoria (37.40S-144.40E), 1985 | - | |
| RU6503 | New Zealand, between Lumdsen and Kingston, 1986 | C. van Hove | |
| PP7001*,$ | A. pinnata subsp. pinnata | Australia, Kakadu Northern Park Northern Territory, 1982 | Yatazawa |
| PP7506 | Sierra Leone, 1982 | C Dixon | |
| PP7509 | Nigeria, Moor plantation, 987 | C. van Hove | |
| PP7511 | Guinea-Bissau, Contuboel, 1987 | H. Diara | |
| PP7512 | Zaire, Kisantu, 1987 | B. Bruyneel | |
| PP7546 | Madagascar, Antsahavory, East zone, 1991 | C. van Hove |
aAccession numbers were listed according to IRRI code number except for Portuguese accessions (FI-BGLU and FI-BGM); - Unknown collector or germplasm source. bCIAT-International Centre for Tropical Agriculture, Colombia, CRRI-Cyili Rice Research Center, IB China-Institute of Botany, Academia Sinica, Beijing, China, IRRI-International Rice Research Institute. # N. azollae was not isolated from this Azolla accession. *BMAA extracted from Azolla and Nostoc azollae (isolated from Azolla, see 4.2.) with method 1 (see 4.3.1.). $BMAA extracted from Azolla accessions with method 2 (see 4.3.2.).
Table 2.
Chromatographic and Mass parameters for the BMAA and the isomers AEG and DAB detection.
LOD - limit of detection; LOQ - limit of quantification; CID - collision induced dissociation.
Table 3.
Primers used to amplify cyanotoxic genes in Azolla and N. azollae DNA.
| Gene | Primer | Sequence primer (5’→3´) | Size (bp) | Reference |
|---|---|---|---|---|
| Saxitoxin (sxt) | SXT683F | GGATCTCAAACATGATCCCA | 195 | Lopes et al. 2012 |
| SXT877R | GCCAAACGCAGTACCACTT | |||
| Cylindrospermopsin (cyl) (poliketide synthase) | K18F | CCTCGCACATAGCCATTTGC | 422 | Schembri et al. 2001 |
| M4R | GAAGCTCTGGAATCCGGTAA | |||
| Cylindrospermopsin (cyl) (peptide synthase) | M13 | GGCAAATTGTGATAGCCACGAGC | 597 | Fergusson 2003 |
| M14 | GATGGAACATCGCTCACTGGTG | Schembri et al. 2001 | ||
| Microcystin/Nodularin synthetase (mcyE/ndaF) | HepF | TTTGGGGTTAACTTTTTTGGCCATAGTC | 472 | Jungblut 2006 |
| HepR | AATTCTTGAGGCTGTAAATCGGGTTT | |||
| Microcystin synthetase (mcy A) | mcyA-Cd1F | AAAATTAAAAGCCGTATCAAA | 297 | Hisbergues et al. 2003 |
| mcyA-Cd1R | AAAAGTGTTTTATTAGCGGCTCAT | |||
| Microcystin synthetase (mcy B) | 2959F | TGGGAAGATGTTCTTCAGGTATCCAA | 350 | Nonneman & Zimba 2002 |
| 3278R | AGAGTGGAAACAATATGATAAGCTAC | |||
| Microcystin (mcy C) | FAA | CTATGTTATTTATACATCAGG | 758 | Neilan 1999 |
| RAA | CTCAGCTTAACTTGATTATC | |||
| Microcystin (mcy B, domain A) | 2156F | ATCACTTCAATCTAACGACT | 955 | Mikalsen 2003 |
| 3111R | GTTGCTGCTGTAAGAAA | |||
| Microcystin (mcy C, domain A) | PSCF1 | GCAACATCCCAAGAGCAAAG | 674 | Ouahid 2005 |
| PSCR1 | CCGACAACATCACAAAGGC | |||
| Microcystin (mcy D, domain ACP) | PKDF1 | GACGCTCAAATGATGAAACT | 647 | Ouahid 2005 |
| PKDR1 | GCAACCGATAAAAACTCCC | |||
| Microcystin (mcy D, domain KS) | PKDF2 | AGTTATTCTCCTCAAGCC | 859 | Ouahid 2005 |
| PKDR2 | CATTCGTTCCACTAAATCC | |||
| Microcystin (mcy E, domain GSA-AMT) | PKEF1 | CGCAAACCCGATTTACAG | 755 | Ouahid 2005 |
| PKER1 | CCCCTACCATCTTCATCTTC | |||
| Microcystin (mcy G, domain CM) | PKGF1 | ACTCTCAAGTTATCCTCCCTC | 425 | Ouahid 2005 |
| PKGR1 | AATCGCTAAAACGCCACC |
Table 4.
Amplification conditions for the cyanotoxic genes in Azolla and N. azollae DNA.
| Gene | Initial denaturation | Denaturation | Annealing | Extension | Final extension | Reference |
|---|---|---|---|---|---|---|
| sxt | 94°C; 3 min | 35 cycles | 72°C; 7 min | Lopes et al. 2012 | ||
| 94°C; 10 s | 52°C; 20 s | 72°C; 1 min | ||||
| cyl | 94°C; 10 min | 30 cycles | 72°C; 7 min | Fergusson 2003 | ||
| 94°C; 30 s | 55°C; 30 s | 72°C; 7 min | ||||
| mcyE/ndaF | 92°C; 2 min | 35 cycles | 72°C; 5 min | Jungblut 2006 | ||
| 92°C; 20 s | 56°C; 30 s | 72°C; 1 min | ||||
| mcy A | 95°C; 2 min | 35 cycles | 72°C; 7 min | Hisbergues et al. 2003 | ||
| 95°C; 90 s | 56°C; 30 s | 72°C; 50 s | ||||
| mcy B | 94°C; 2 min | 35 cycles | 72°C; 5 min | Nonneman 2002 | ||
| 94°C; 30 s | 59°C; 45 s | 72°C; 1 min | ||||
| mcy C | 94°C; 2 min | 35 cycles | 72°C; 7 min | Neilan 1999 | ||
| 94°C; 10 s | 50°C; 20 s | 72°C; 1 min | ||||
| mcy B, domain A | 94°C; 4 min | 30 cycles | 72°C; 7 min | Mikalsen 2003 | ||
| 95°C; 30 s | 52°C; 30 s | 72°C; 1 min | ||||
| mcy C, domain A | 94°C; 5 min | 35 cycles | 72°C; 7 min | Ouahid 2005 | ||
| 95°C; 1 min | 52°C; 30 s | 72°C; 1 min | ||||
| mcy D, domain ACP | 94°C; 5 min | 35 cycles | 72°C; 7 min | Ouahid 2005 | ||
| 95°C; 1 min | 52°C; 30 s | 72°C; 1 min | ||||
| mcy D, domain KS | 94°C; 5 min | 35 cycles | 72°C; 7 min | Ouahid 2005 | ||
| 95°C; 1 min | 52°C; 30 s | 72°C; 1 min | ||||
| mcy E, domain GST-AMT | 94°C; 5 min | 35 cycles | 72°C; 7 min | Ouahid 2005 | ||
| 95°C; 1 min | 52°C; 30 s | 72°C; 1 min | ||||
| mcy G, domain CM | 94°C; 5 min | 35 cycles | 72°C; 7 min | Ouahid 2005 | ||
| 95°C; 1 min | 52°C; 30 s | 72°C; 1 min | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



