
Submitted:
23 August 2024
Posted:
24 August 2024
You are already at the latest version
Abstract
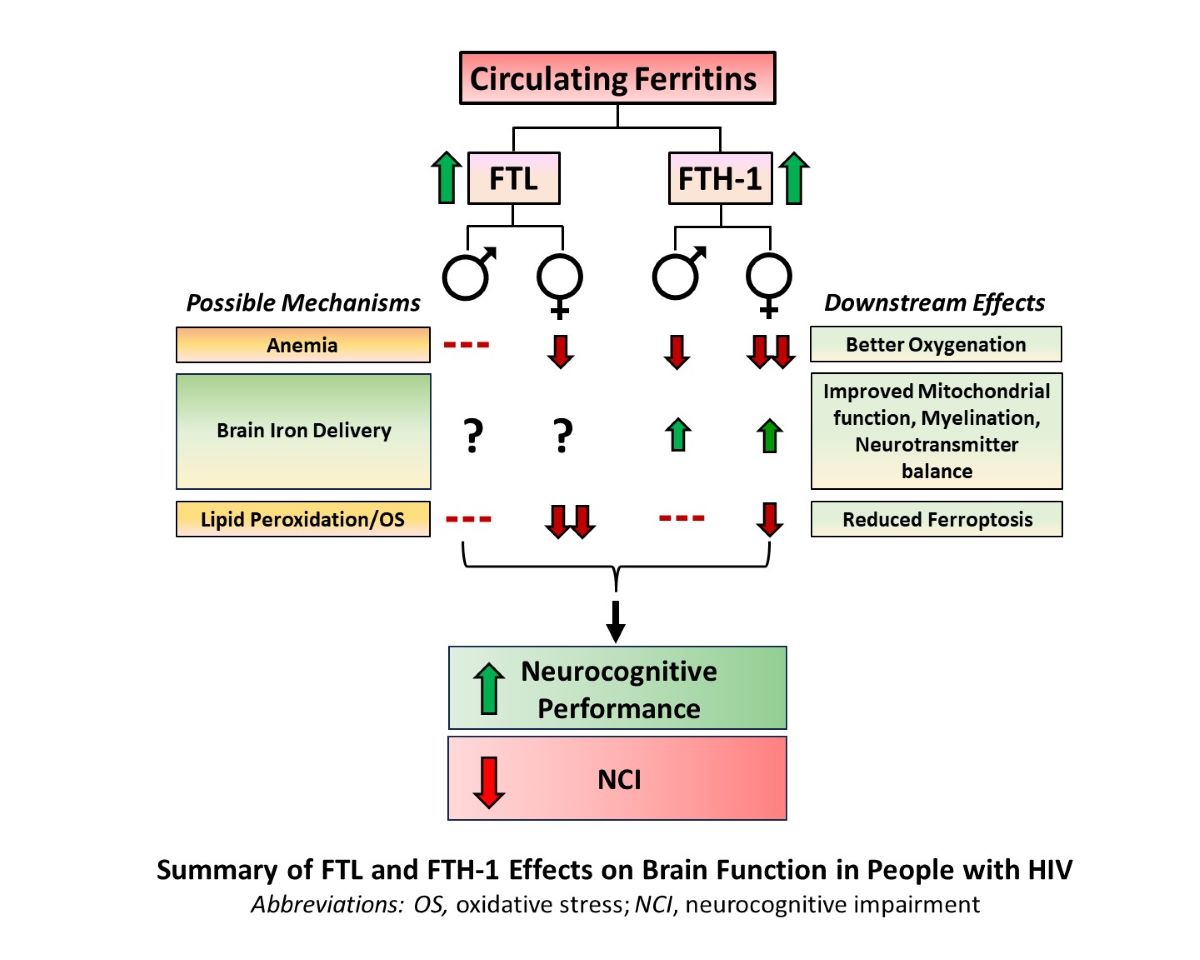
Ferroptosis is implicated in viral neuropathogenesis and may underlie HIV-associated neurocognitive impairment (NCI). Emerging data also suggest differences in brain iron transport by sex. We hypothesized that circulating ferritins that inhibit ferroptosis associate with neurocognitive function and NCI in people with HIV (PWH) in a sex-biased manner. Serum ferritin heavy-chain-1 (Fth-1), ferritin light-chain (Ftl), and urinary F2-isoprostanes (uF2-isoPs, specific lipid peroxidation marker) were quantified in 324 PWH (including 61 women) with serial global (NPZ4) and domain-specific neurocognitive testing. Biomarker associations with neurocognitive test scores and NCI were evaluated by multivariable regression; correlations with uF2-isoPs were also assessed. Higher Ftl and Fth-1 associated with less NCI in all PWH (adjusted odds ratios 0.53, 95% Confidence Interval (95% CI) 0.36-0.79 and 0.66, 95% CI 0.45-0.97, respectively). In women, higher Ftl and Fth-1 associated with better NPZ-4 (Ftl: adjusted beta (β)=0.15, 95% CI 0.02-0.29; Ftl-by-sex βinteraction=0.32, p=0.047) and domain-specific neurocognitive test scores. Effects on neurocognitive performance persisted for up to 5 years. Levels of both ferritins correlated inversely with uF2-isoPs in women (Ftl: rho=-0.47, p<0.001). Circulating Ftl and Fth-1 exert sustained, sex-biased neuroprotective effects in PWH, possibly by protecting against iron-mediated lipid peroxidation (ferroptosis). Larger studies are needed to confirm the observed sex differences and further delineate underlying mechanisms.
Keywords:
HIV
; ferritin heavy-chain
; ferritin light-chain
; ferroptosis
; lipid peroxidation
; iron
; oxidative stress
; neurocognitive impairment
; neurocognitive domains
1. Introduction
HIV-associated neurocognitive impairment (NCI) remains common among people with HIV (PWH), despite suppressive combination antiretroviral therapy (ART) [1]. The pathogenesis of NCI in PWH is multifactorial, with contributions from chronic inflammation, vascular risk factors, substance use, and an impaired gut mucosal barrier, as well other factors [2,3,4,5]. Despite extensive research, however, the mechanisms driving development of NCI, particularly in virally suppressed PWH, remain incompletely defined.
Ferroptosis, a regulated form of cell death due to iron-mediated lipid peroxidation, is an emerging pathophysiologic mechanism in a growing number of chronic inflammatory conditions, including neurocognitive disorders [6,7]. HIV and inflammation dysregulate iron transport, which is essential for mitochondrial energy production, myelination, and neurotransmitter homeostasis in the brain [8]. Ferritin, the primary iron storage protein in humans, is a macromolecule composed of heavy-chain (Fth-1) and light-chain (Ftl) subunits in proportions that vary by tissue type [9]. The proteins Fth-1 and Ftl are major antioxidants by virtue of their iron-binding functions; Fth-1 also has mitochondria-protective, immune-modulating, and tissue iron-delivery roles [9,10,11]. Both ferritin subunits inhibit ferroptosis by sequestering iron in a nonreactive form [12,13,14,15]. We recently showed that higher cerebrospinal fluid (CSF) levels of Fth-1, the ferroxidase-containing ferritin subunit, predict better neurocognitive performance in PWH over 3 years of follow-up [10]. Fth-1 is a major source of iron for mature oligodendrocytes via the oligodendrocyte receptor, T-cell Immunoglobulin and Mucin domain-1 (Tim-1) [16]. Recent studies suggest important differences in utilization of iron transport proteins such as Fth-1 for iron delivery to the brain between males and females [17,18]. The purpose of this study was to test the hypotheses that higher circulating Fth-1 and/or Ftl are associated with better neurocognitive function in PWH in a sex-specific manner, independent of inflammation and other known risk factors for NCI. In keeping with their iron-delivery and anti-ferroptosis roles, we also explored associations of Fth-1 and Ftl with anemia and isoprostanes (specific markers of in vivo oxidative stress) in PWH.
2. Participants and Methods
2.1. Study Design, Participants, and Outcome Measures
The Advancing Clinical Therapeutics Globally (ACTG) A5322 Study (HAILO, the Human Immunodeficiency Virus [HIV] Infection, Aging, Immune Function Long-Term Observational Study) was a prospective observational study, which enrolled 1035 middle-aged and older PWH (aged ≥40 years at enrollment) in 2013 and 2014 and followed them through 2021. The present analysis included a subset of 324 HAILO Study participants with serial neurocognitive assessments and pre-existing data on biomarkers of inflammation. Detailed information on clinical and HIV disease variables was collected, as described previously [19]. Data on participant gender identification were not available at the time of this study, though collected later. Neurocognitive function was assessed using the ACTG Study A5001 NeuroScreen at entry and every 48 weeks for up to 288 weeks [20]. The NeuroScreen included the following tests: Trail-Making A (TMA); Trail-Making B (TMB), the Wechsler Adult Intelligence Scale-Revised Digit Symbol (DSY) test, and Hopkins Verbal Learning test (HVLT); 5 participants lacked complete neurocognitive test score data and were not evaluable, since their NCI status could not be determined. Neurocognitive performance was summarized as a mean z-score, the NPZ-4, which incorporates normative adjustments for age, biological sex (referred to henceforth as men if male at birth, or women if female at birth), self-reported race, ethnicity, and education, as well as practice effects, and represents the average score across all 4 ability domains. NCI was defined as at least one z-score ≥2 standard deviations (SD) below the mean, or at least two z-scores ≥1 SD below the mean on separate tests within the NeuroScreen, which was previously validated against a comprehensive neuropsychological battery [21].
2.2. Biomarker Quantification
Fth-1 and Ftl levels were quantified in serum from baseline (entry) visits by enzyme-linked immunosorbent assay (ELISA). Commercially available ELISA kits from LSBioTM were used to measure serum Fth-1 and Ftl (Human Fth-1 ELISA kit, catalog #LS-F22867; Human Ftl ELISA kit and catalog #LS-F21824), according to the manufacturer’s protocol. In brief, 100 µl of standard, control, or diluted participant samples were added, in duplicate, to ELISA plates pre-coated with capture antibody (anti-Fth-1 or anti-Ftl) and incubated at 37oC for 90 minutes. Each well was aspirated and incubated with 100 µl of 1x biotinylated detection antibody for 1 hour at 37oC, followed by rinsing the wells 3 times with buffer. To each well, 100 µl of 1X HRP conjugate/1X ABC complex were added, followed by incubation at 37oC for 30 minutes and repeat rinsing of wells with buffer. Then, 90 µl of TMB (3,3',5,5'-Tetramethylbenzidine) substrate were added, and the wells were incubated at 37oC for 15 minutes; the reaction was stopped with 90 µl of stop solution and the optical density read at 450 nm.
Levels of IL-6, soluble tumor necrosis factor receptor-II (sTNFR-II), and soluble CD163 (sCD163) were previously quantified in entry plasma from A5322 Study participants using commercial ELISA assays (R&D Systems, Minneapolis, MN, US).
Isoprostanes are the most specific and sensitive markers of in vivo oxidative stress currently available, and their levels are more stable in urine compared to serum (or plasma), due to ex vivo autoxidation of arachidonic acid in the latter [22,23]. Among the various isoprostanes generated in vivo, the 5-series and particularly 15-series F2-isoprostanes are among the most abundant [24]. We therefore quantified 5-series and 15-series F2-isoprostanes in urine samples at entry by gas chromatography/mass spectrometry at the Vanderbilt University Eicosanoid Core Laboratory, using established protocols [24]. Since isoprostanes are renally excreted, urinary F2-isoprostane (uF2-IsoPs) levels were adjusted for renal function (serum creatinine) prior to analyses.
2.3. Statistical Methods
Biomarker levels were summarized as medians (interquartile ranges), if skewed, or as means (SD), when normally distributed. Variables at entry were compared using Pearson’s Chi-square test (if categorical) or the Wilcoxon Rank Sum test (if continuous). Biomarker values were log-transformed to improve normality. Fth-1 and Ftl relationships to biomarkers of inflammation were evaluated by non-parametric (Spearman’s) correlations. Multiple linear regression models were used to assess associations of Fth-1 and Ftl levels with NPZ-4, TMA, TMB, DSY and HVLT scores and estimate adjusted β-coefficients and their 95% confidence intervals (95% CIs). Logistic regression models were used to assess biomarker associations with global NCI at entry and estimate odds ratios and their 95% CIs, adjusting for influential covariates, as described below. Longitudinal associations of iron biomarkers with neurocognitive function over time were determined using generalized estimation equations, adjusting for the following factors at entry: age, nadir CD4<200/μl, inflammation (IL-6, sTNFR-II, sCD163), anemia, education, race, ethnicity, and comorbidity burden ± sex. To facilitate the plotting of results from these analyses, adjusted β-estimates for predicted adjusted changes in neurocognitive test scores over time were calculated for participants with ferritin biomarker levels above vs. below the median.
ACTG Study A5322 entry-visit variables evaluated as covariates included: age, biological sex, self-reported race (black or non-Hispanic white), Hispanic ethnicity, education, plasma HIV RNA (copies/mL), nadir CD4+ T-cell count, anemia (defined by sex-specific criteria as yes/no: hemoglobin (hgb)<12 mg/dL in females and hgb<13.5 mg/dL in males), hepatitis C virus (HCV) serostatus, current efavirenz use, and comorbidity burden (0 to ≥4). Previously measured IL-6, sTNFR-II, and sCD163 levels, converted to z-scores, were also evaluated. At least two of these markers (sTNFR-II and sCD163) were included in all regression models, since ferritins are acute-phase proteins and their levels generally increase with inflammation/immune activation [25,26,27]; IL-6 was included only if associated with the outcome variable with p<0.05 (i.e., for TMA and DSY scores). HIV-associated non-AIDS comorbidities included cancer, diabetes, cardiovascular or cerebrovascular disease (e.g., myocardial infarction, hypertension, transient ischemic attack, or stroke), chronic kidney disease, HCV status, and bone fractures. With the exception of age, inflammation, and anemia, which were forced into all models due to their particularly strong influence on iron metabolism and association with neurocognitive function in PWH [28,29], other covariates with p≤0.10 in univariate analyses were included in final multivariable models of neurocognitive outcomes in the entire sample or in men (81% of the sample). Models confined to women (N=61) included only age, ethnicity, comorbidity burden, anemia, and a single inflammation marker as covariates. (Additional adjustment for HCV status, nadir CD4, and efavirenz use did not alter results in women, so these variables were omitted to avoid overfitting and optimize power.) Interaction effects by sex (Ftl-by-sex and Fth-1-by-sex) were tested by including a multiplicative interaction term in multivariable models of global cognitive performance. A two-sided alpha of 0.05 was used to determine significance.
To evaluate effects at the extremes of biomarker distributions and account for possible threshold effects in vivo, Fth-1 and Ftl were also evaluated as quartiles (e.g., quartile 4 vs. quartile 1) [10]. Finally, potential mechanisms for observed associations were explored by testing univariate biomarker associations with anemia in logistic regression models, and with uF2-isoPs by linear regression or Spearman (nonparametric) correlations. Plots were generated with GraphPad Prism 9. STATA (version 17, StataCorp, College Station, TX, USA) was used to perform all statistical analyses.
3. Results
3.1. Study Participant Characteristics at Entry
Of 324 PWH, 81% (263) were men, and 19% (61) were women. Viral load data were available for all but 2 individuals, and 96% were virally suppressed (plasma HIV RNA<200 copies/mL). Neurocognitive assessments were complete in 319 PWH: 237 were categorized as neurocognitively normal, and 82 had NCI by the ACTG NeuroScreen [20]. Participant characteristics at entry are shown, stratified by NCI status, in Table 1. Proportions of PWH with 2 or more comorbidities and HCV seropositivity were higher among NCI cases. The prevalence of NCI in this sample differed by self-reported race and Hispanic ethnicity. Viral suppression in plasma was significantly less common, and current efavirenz use tended to be less common, in neurocognitively impaired PWH. Serum Fth-1, Ftl, and uF2-IsoP levels at entry are summarized in Table 2. Serum Fth-1 and Ftl levels were non-significantly lower in women. No sex differences were observed for uF2-IsoPs in this study sample, but uF2-IsoP levels were undetectable in 5.3% of men.
As shown in Supplementary Table S1, the prevalence of anemia and the proportion of non-Hispanic blacks were significantly higher in women than in men at entry; no other significant sex differences in demographic or HIV disease characteristics, or in the prevalence of neurocognitive impairment were observed.
3.2. Multivariable Ferritin Associations with Global NCI and Neurocognitive Performance, and Interaction Effects by Sex
Among all PWH, higher Ftl levels at entry were associated with significantly reduced odds of NCI, adjusting for age, sex, self-reported race and ethnicity, comorbidity burden, HCV status, anemia, efavirenz use, and inflammation or immune activation (adjusted odds ratio (adjOR) for NCI=0.53, 95% CI 0.36-0.79; Figure 1A). The association was also significant, with a stronger effect size, in women (adjOR=0.29, 95% CI 0.11-0.75), compared to men (adjOR=0.64, 95% CI 0.41-0.99). Higher Fth-1 levels were also associated with a lower likelihood of NCI in the entire sample (adjOR=0.66, 95% CI 0.45-0.97; Figure 1B), with similar results in women (women: adjOR=0.45, 95% CI 0.20-1.0; men: adjOR=0.77, 95% CI 0.48-1.23).
Results for NCI confined to 294 virally suppressed PWH with HIV RNA levels <200 copies/mL were similar. In this subset, both Ftl and Fth-1 levels at entry were associated with a reduced likelihood of NCI (adjORs=0.54, 95% CI 0.36-0.79, and 0.66, 95% CI 0.45-0.98, respectively, Figure S1). Results of sex-stratified analyses in this subset were similar: Ftl levels remained significantly associated with reduced NCI in 56 virally suppressed women and 239 virally suppressed men (adjOR=0.31, 95% CI 0.12-0.80 vs. 0.63, 95% CI 0.41-1.0). Fth-1 was associated with NCI in virally suppressed women only (adjOR=0.41, 95% CI 0.17-0.98).
Associations were also observed between higher serum Ftl levels at entry and better global neurocognitive (NPZ-4) test scores in all study participants (adjusted beta (β)=0.15, 95% CI 0.02-0.29), and particularly women (β=0.40, 95% CI 0.13-0.68, Figure 2A). For Fth-1, higher levels were significantly associated with better NPZ-4 scores at entry only in women (β=0.36, 95% CI 0.02-0.69, Figure 2B). Since threshold effects often occur in vivo, we further assessed associations of test scores with Fth-1 and Ftl levels in the highest (top) vs. lowest (bottom) quartiles of the biomarker distributions. Despite reduced power, Ftl levels in the top (≥22.1 ng/mL) vs. bottom quartile (≤7.7 ng/mL) remained associated with better NPZ-4 scores in women (β=1.20, 95% CI 0.58-1.7); Fth-1 levels in the top vs. bottom quartile (≥499.5 ng/mL vs. ≤192 ng/mL) also remained associated with better NPZ-4 scores in women (β=0.31, 95% CI 0.01-0.62). For both Ftl and Fth-1, multiplicative ferritin-by-sex interaction effects on the NPZ-4 score were statistically significant (Ftl: βinteraction=0.32, p=0.047; Fth-1: βinteraction=0.31, p=0.039), such that higher levels of each ferritin had a significantly greater impact on global neurocognitive function in women as compared to men.
3.3. Ftl and Fth-1 Associations with Neurocognitive Domain Test Scores
Ferritin associations with neurocognitive function in specific ability domains are shown in Figures S2 and S3. Higher Ftl was significantly associated with higher (better) TMA scores (β=0.67, 95% CI 0.33-1.0, Figure S2A; Ftl-by-sex βinteraction=0.59, p=0.005) and higher TMB scores (β=0.34, 95% CI 0.02-0.65, Figure S2B) only in women. Ftl levels were also associated with better DSY test scores in all PWH (β=0.29, 95% CI 0.11-0.47), with stronger effects in women (β=0.53, 95% CI 0.11-0.95) than in men (β=0.21, 95% CI 0.001-0.42, Figure S2C). As shown in Figure S3(A-C), associations of Fth-1 with neurocognitive domain test scores were generally weaker than for Ftl but still tended to be stronger in women; only the association with DSY score was statistically significant (women: TMA: β=0.38, 95% CI -0.05-0.79; TMB: β=0.32, 95% CI -0.03-0.67; DSY: β=0.48, 95% CI 0.01-0.96; Fth-1-by-sex βinteraction=0.47 for DSY, p=0.026). No associations were observed for either Ftl or Fth-1 with HVLT scores (Figures S2D and S3D).
3.4. Ftl and Fth-1 Associations with Longitudinal Neurocognitive Performance
Results of longitudinal regression analyses associating Ftl and Fth-1 levels at entry with neurocognitive test scores over time are shown in Figures S4 and S5. Participants with Ftl and Fth-1 levels above vs. below the median were compared across all visits, for up to 5 years of follow-up. Serum Ftl levels above the median (≥12.2 ng/ml) were associated with better NPZ-4 scores at each follow-up time-point up to 240 weeks in PWH (β=0.26, p=0.010, Figure S4A), and with significantly better NPZ-4 scores in women (β=0.47, p=0.032, Figure S4B) but not men (β=0.21, p=0.067, Figure S4C). Although Ftl (and to a lesser extent, Fth-1) levels predicted differences in neurocognitive performance over time, ferritin-by-time interaction terms were not statistically significant (i.e., biomarker levels at entry did not predict changes in neurocognitive scores over time). Higher Ftl was also associated with better TMA scores (β=0.84, p=0.002, Figure S4D) in women and with better DSY scores in all PWH (β=0.36, p=0.013, Figure S4E); associations with DSY scores over follow-up were close to statistical significance for both women and men (Figures S4F and S4G, respectively). For Fth-1, significant associations in women were observed for global neurocognitive function (NPZ-4: β=0.57, p<0.01), and the DSY test score (DSY: β=0.58, p=0.035; TMA: β=0.64, p=0.040, latter shown in Figure S5). No longitudinal associations with HVLT scores were detected.
3.5. Ferritin Associations with Inflammation, Lipid Peroxidation, and Anemia
Fth-1 and Ftl correlations with previously measured biomarkers of inflammation – sTNFR-II, sCD163, and IL-6 – and with both 5-series and 15-series uF2-IsoPs at entry were evaluated. Serum Fth-1 and Ftl were not significantly correlated with any biomarker of inflammation in either sex (Table 3). Levels of Fth-1, and particularly Ftl, were inversely correlated with 15-series uF2-IsoPs only in women (Fth-1: rho= -0.29, p=0.023; Ftl: rho= -0.47, p<0.001 for 15-series uF2-IsoPs). As shown, Ftl levels also correlated inversely with 5-series uF2-IsoPs in women and weakly but positively with 5-series uF2-IsoPs in men (rho=0.134, p=0.03).
Serum Fth-1, but not Ftl, levels were correlated positively with hemoglobin in all PWH and particularly in women (rho=0.39, p<0.01). Higher Fth-1 was strongly associated with a reduced prevalence of anemia, defined by sex-specific criteria, in unadjusted analyses at entry (Fth-1: β= -0.84, 95% CI: -1.1 to -0.58, p<0.001 for all PWH); this association was much stronger in women than in men but statistically significant in both sexes (Supplementary Table S2). A similar inverse relationship with anemia was observed for Ftl in women only (β= -0.46, 95% CI: -0.98 to 0.07 p=0.086).
4. Discussion
We previously reported that higher levels of the antioxidant iron-binding and iron-delivery protein Fth-1 in CSF were associated with better neurocognitive performance over time in PWH, independent of known contributors, such as comorbidity and CSF inflammation [10]. CSF sampling is invasive and impractical for disease monitoring in ambulatory PWH, however, so this study sought to determine whether similar relationships exist between neurocognitive function and circulating Fth-1, or the functionally distinct light-chain ferritin subunit, Ftl. This study confirms and extends our prior findings in a separate HIV population, by showing that higher serum Fth-1 and Ftl levels are associated with reduced NCI and better global neurocognitive function (NPZ-4 scores), particularly in women. Importantly, consistent findings were observed in virally suppressed PWH. Furthermore, scores on specific neurocognitive domain tests were better in study participants with higher serum Ftl and to a lesser extent, higher Fth-1 levels. Significant multiplicative Ftl-by-sex and Fth-1-by-sex interaction effects on overall neurocognitive performance and in some ability domains were also observed.
Iron is required for numerous biological processes in the brain and its homeostatic regulation is critical for brain health [10,30,31]. Moreover, iron metabolism is dysregulated by HIV infection [32]. These changes are accompanied by increases in nonspecific markers of inflammation, and increased levels of hepcidin, a pro-inflammatory iron-regulatory peptide hormone [32,33]; hence, careful adjustment for inflammation is key to establishing a role for iron transport in chronic inflammatory disorders. Biomarkers of inflammation (sTNFR-II and sCD163, with or without IL-6, measured in plasma at entry) were therefore included as covariates in our analyses. Inflammation has known adverse effects on neurocognitive function, whereas Fth-1 and Ftl were associated with better neurocognitive test scores and less NCI and were unrelated to inflammation in our study population, so the observed associations are not explained by the fact that they are acute-phase proteins. Persistent or stronger effects in the highest vs. lowest quartiles of both biomarkers with neurocognitive performance in PWH, and sustained differences over time in NPZ-4 scores between groups with biomarker levels higher vs. lower than the median, also indicate robust findings.
Neurocognitive impairment in PWH has been linked to thinning of the corpus callosum, reduced structural integrity of myelin, and white matter (or myelin) damage in the brain [34,35,36,37]. Preservation of oligodendrocyte differentiation and function is essential for normal myelination and re-myelination, processes which require a steady supply of iron; oligodendrocytes are the most iron-laden cell type in the CNS [38,39]. Mature oligodendrocytes lack transferrin receptors, however, and they are unable to import transferrin-bound iron as all other cells do; these cells take up iron via Fth-1, which binds the oligodendrocyte Tim-1 receptor [16]. Fth-1 also competes for binding to Tim-1 with an oligodendrocyte toxin released by HIV-infected T-cells, semaphorin 4a; hence, higher Fth-1 levels may also limit viral-mediated myelin damage by promoting oligodendrocyte survival [16]. Ferritin (particularly Fth-1) released from oligodendrocytes, microglia, and astrocytes also augments the antioxidant defenses of neighboring cells, and release of trophic Fth-1 may be reduced in HIV infection [40,41,42]. Glial activation by HIV may inhibit Fth-1 release, reducing iron bioavailability [42,43,44] and promoting functional iron deficiency in the brain, particularly in oligodendrocytes [45]. Reduced iron availability for glia and neurons, which have high metabolic demands for this micronutrient, could contribute to HIV neuropathogenesis and NCI. Previous studies have shown that deletion of Fth-1 in oligodendrocytes in mice results in diminished remyelination [46]. A similar role in iron delivery and oligodendrocyte health has not been identified for Ftl, but like Fth-1, this subunit is critical for antioxidant defense and may also have an immunomodulatory role [41,47,48].
Oxidative stress is a major contributor to neurodegenerative disorders, including HIV-associated NCI [49,50,51], and a significant proportion of oxidative brain injury in vivo may occur via the process of ferroptosis [52]. Ferroptosis, first described in 2012, is a highly regulated form of programmed cell death due to iron-mediated lipid peroxidation [53]. Ferroptosis has emerged as a novel mechanism in chronic human diseases and is implicated in viral neuropathogenesis [54,55,56]. Excess or non-protein-bound iron promotes the formation of highly reactive hydroxide radicals, lipid hydroperoxides, and other reactive oxygen/nitrogen species (ROS) via Fenton chemistry; these ROS directly damage biological macromolecules and cell-membrane constituents, including lipids, proteins and DNA [30,57,58]. The lipid-rich brain (including myelin) is especially susceptible to ferroptosis. Iron must therefore be tightly compartmentalized and maintained in a non-reactive state, primarily by sequestration within ferritins [9,14,30]. The synthesis of both Fth-1 and Ftl is upregulated under oxidative stress, and overexpression of Fth-1 and Ftl reduces accumulation of ROS in response to an oxidative challenge [30,59]. Recent studies have established Fth-1 and Ftl as important negative regulators of ferroptosis [60,61]. In this study, higher levels of Fth-1 and particularly Ftl were associated in women with lower levels of both 5- and 15-series uF2-isoPs, highly specific biomarkers of lipid peroxidation in vivo [22]. Although these relationships were only correlative, the results suggest that higher levels of these ferritins may prevent neurocognitive decline in PWH by suppressing lipid peroxidation and ferroptosis in vivo, which would be expected to protect oligodendrocytes and reduce white matter injury [39,62]. By contrast, higher Fth-1 levels in all PWH in our study were strongly associated with reduced anemia, another key contributor to NCI in this population [29,63,64]. We speculate that Ftl may be more important for suppressing lipid peroxidation, while Fth-1 primarily prevents anemia and preserves iron delivery to myelinating oligodendrocytes in the setting of functional iron deficiency [10,63,65]. Overall, our findings in PWH are consistent with a recent study by Kannan et al, which demonstrated that the HIV Tat protein increases intracellular labile iron, lipid oxidation, and ferroptosis in mouse primary microglia in vitro and induces Fth-1. In that study, expression of ferroptosis markers was significantly altered in the brains of HIV-1-transgenic rats, as well as in autopsy brain tissues from PWH [66].
The observed sex differences in Ftl and Fth-1 effects on neurocognitive function require deeper investigation. Recent studies suggest emerging sex differences in brain iron transport and accumulation, and in associations between brain iron and neurocognitive function [18,67,68,69,70]. Whether women with HIV differ in their vulnerability to HIV-mediated cognitive dysfunction remains controversial [71,72]. We found that neuroprotective associations of Ftl and Fth-1 were most prominent in women: stronger relationships of these biomarkers to reduced anemia and lower uF2-isoPs in women may partly explain the sex differences, particularly if oxidative-stress mechanisms are more important for HIV neuropathogenesis in women or specifically impact female reproductive fitness. Studies of other iron-metabolic markers, including the master iron-regulatory and pro-inflammatory hormone hepcidin and its regulator, erythroferrone, may provide further insights [73].
This study has several limitations worth noting, particularly the small number of women. Women have historically been underrepresented in HIV observational studies, however, and these findings require confirmation in more women with HIV [74]. Our studies of other iron-related biomarker associations with NCI among PWH have included too few women to explore sex differences [10,75,76]. Here, we detected strong associations in women despite limited power, while we were unable to detect any associations in men despite ample numbers of men in the study. The finding of sex-specific associations that were consistent in direction for both ferritins and across all of our analyses (of global NCI, domain-specific NCI, and continuous measures of neurocognitive function) also supports the validity of these results. Secondly, the interpretation of serum ferritin associations with NCI requires caution: both Fth-1 and Ftl, which cross the blood-brain barrier, have the potential to contribute to ferroptosis via dysregulated ferritinophagy (autophagy of ferritin) and release of intracellular iron [77], and it is unclear how serum ferritin levels relate to CSF or brain ferritin levels; simultaneous CSF Fth-1 and Ftl measurements for comparison to serum values were not possible in this sample. Another potential weakness is the lack of adjustment for substance use in our analyses. However, no participants reported current or active substance use, and we have not found substance use to be a significant confounder in our previous studies of these iron biomarkers; we would not expect past substance use to have altered measured iron biomarkers [10]. While iron-containing supplement or multivitamin use could potentially have affected our results, iron was not listed as a medication for any participants in this study. Finally, only later versions of the HAILO Study ascertained depression, and as our covariate adjustments were necessarily limited in women, our analyses did not adjust for depression as a covariate. Depression overlaps with NCI in PWH and may relate to iron status and/or ferroptosis [78,79]. We therefore cannot exclude the possibility of residual confounding by comorbidities such as depression and substance use.
5. Conclusions
Higher levels of the circulating antioxidant, iron-sequestering proteins and ferroptosis-inhibitors Ftl and Fth-1 were associated with less NCI and better neurocognitive performance globally and in specific ability domains in PWH, particularly in women. Similar results were observed in viral suppressed persons, and associations were sustained over several years of follow-up. These findings are distinct from, and expand upon, the prior associations limited to CSF Fth-1 and underscore the importance of iron dysregulation in HIV neuropathogenesis [80]. Levels of these proteins were also inversely correlated in women with a sensitive and specific marker of in vivo lipid peroxidation, a hallmark of ferroptosis, and with less anemia in both sexes. White matter (myelin) damage and anemia are established contributors to neurocognitive decline in people with and without HIV [29,34,37,64,81], and ferroptosis promoted by HIV-mediated iron dysregulation may be an important pathophysiologic mechanism underlying NCI in PWH. Circulating Fth-1 and Ftl appear to play neuroprotective roles that differentially impact each of these mechanisms. Larger studies with more women may help to determine the relative importance of lipid peroxidation vs. anemia in neuro-HIV complications and better understand the observed sex differences. Overall, these findings suggest a new role for ferroptosis in HIV neuropathogenesis and the potential utility of novel antioxidant interventions – e.g., oral supplementation with Ftl or Fth-1 – to mitigate ferroptosis and prevent neurocognitive decline in PWH.
Supplementary Materials
Figure S1: The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Forest plots summarizing multivariable associations of Ftl levels and Fth-1 levels at entry with NCI in virally suppressed people with HIV (subset of all participants, women, and men). Figure S2: Forest plots summarizing multivariable associations of Ftl levels at entry with domain-specific neurocognitive performance in all participants, women, and men, i.e., with scores on the following tests: Trail-Making-A (TMA), Trail-Making-B (TMB), Wechsler Adult Intelligence Scale-Revised Digit Symbol (DSY), and the Hopkins Verbal Learning Test (HVLT). Figure S3: Forest plots showing multivariable associations of Fth-1 levels at entry with neurocognitive test scores: TMA, TMB, DSY, and HVLT in all participants, women, and men. Figure S4: Longitudinal regression models of predicted adjusted changes over time in global (NPZ-4) and selected neurocognitive domain test scores in people with HIV for Ftl levels above versus below the median at entry. Figure S5: Longitudinal regression models of predicted adjusted changes over time in the TMA test score in women with HIV for serum Fth-1 levels above vs. below the median at entry. Table S1: Characteristics at study entry, stratified by sex. Table S2: Fth-1 and Ftl associations with anemia in people with HIV.
Author Contributions
Conceptualization, A.R.K.; Methodology, H.K., R.K.A.; Validation, H.K., R.K.A., A.R.K.; Formal Analysis, A.R.K., H.K., W.S.B.; Investigation, R.K.A., H.K.; Resources, K.W., S.L.K., R.C.K., K.M.E., R.J.B., B.O.T., K.M.E., K.K.T., R.J.E., F.J.P.; Data Curation, H.K., R.K.A., A.R.K.; Writing – Original Draft Preparation, R.K.A., A.R.K.; Writing – Review & Editing, A.R.K., R.K.A. H.K., K.M.E., K.K.T., R.J.E., R.J.B., B.O.T., F.J.P., R.C.K., C.O.H., S.L.K., J.R.S.; Visualization, R.K.A., H.K.; Supervision, A.R.K.; W.S.B.; Project Administration, A.R.K. Funding acquisition, A.R.K.
Funding
National Institutes of Health (NIH) grant 5R03 MH123291 (to ARK) and 5R01 MH124530 (to ARK), a Cleveland Clinic Catalyst Award (to ARK, RCK, WSB). Research reported in this publication was supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health under Award Numbers UM1 AI068634, UM1 AI068636 and UM1 AI106701. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Institutional Review Board Statement
The study was conducted in accord with the guidelines of the Declaration of Helsinki and approved by the Institutional Review Boards of all ACTG Study A5322 participating sites (IRB# 19-1423, approved 11, 6, 2019, considered exempt research).
Informed Consent Statement
All study participants provided written informed consent for the A5322 study and for use of stored biospecimens for these research purposes.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors upon request.
Acknowledgments
We are grateful to all A5322 (HAILO) Study participants, all ACTG study site investigators, and to Ginger Milne (Director, Vanderbilt University Eicosanoid Core).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Scanlan, A.; Zhang, Z.; Koneru, R.; Reece, M.; Gavegnano, C.; Anderson, A.M.; Tyor, W. A Rationale and Approach to the Development of Specific Treatments for HIV Associated Neurocognitive Impairment. Microorganisms 2022, 10, 2244. [Google Scholar] [CrossRef] [PubMed]
- Saloner, R.; Fields, J.A.; Marcondes, M.C.G.; Iudicello, J.E.; von Känel, S.; Cherner, M.; Letendre, S.L.; Kaul, M.; Grant, I.; Translational Methamphetamine AIDS Research Center (TMARC) Group. Methamphetamine and Cannabis: A Tale of Two Drugs and their Effects on HIV, Brain, and Behavior. J Neuroimmune Pharmacol 2020, 15, 743–764. [Google Scholar] [CrossRef]
- Nightingale, S.; Dreyer, A.J.; Saylor, D.; Gisslén, M.; Winston, A.; Joska, J.A. Moving on From HAND: Why We Need New Criteria for Cognitive Impairment in Persons Living With Human Immunodeficiency Virus and a Proposed Way Forward. Clin Infect Dis 2021, 73, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Hoenigl, M.; de Oliveira, M.F.; Pérez-Santiago, J.; Zhang, Y.; Morris, S.; McCutchan, A.J.; Finkelman, M.; Marcotte, T.D.; Ellis, R.J.; Gianella, S. (1→3)-β-D-Glucan Levels Correlate With Neurocognitive Functioning in HIV-Infected Persons on Suppressive Antiretroviral Therapy: A Cohort Study. Medicine (Baltimore) 2016, 95, e3162. [Google Scholar] [CrossRef] [PubMed]
- Hileman, C.O.; Kalayjian, R.C.; Azzam, S.; Schlatzer, D.; Wu, K.; Tassiopoulos, K.; Bedimo, R.; Ellis, R.J.; Erlandson, K.M.; Kallianpur, A.; Koletar, S.L.; Landay, A.L.; Palella, F.J.; Taiwo, B.; Pallaki, M.; Hoppel, C.L. Plasma Citrate and Succinate Are Associated With Neurocognitive Impairment in Older People With HIV. Clin Infect Dis 2021, 73, e765–e772. [Google Scholar] [CrossRef]
- Tang, D.; Chen, X.; Kang, R.; Kroemer, G. Ferroptosis: molecular mechanisms and health implications. Cell Res 2021, 31, 107–125. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Alhazmi, H.A.; Hassani, R.; Khuwaja, G.; Maheshkumar, V.P.; Aldahish, A.; Chidambaram, K. Role of ferroptosis pathways in neuroinflammation and neurological disorders: From pathogenesis to treatment. Heliyon 2024, 10, e24786. [Google Scholar] [CrossRef]
- Patton, S.M.; Wang, Q.; Hulgan, T.; Connor, J.R.; Jia, P.; Zhao, Z.; Letendre, S.L.; Ellis, R.J.; Bush, W.S.; Samuels, D.C.; Franklin, D.R.; Kaur, H.; Iudicello, J.; Grant, I.; Kallianpur, A.R. Cerebrospinal fluid (CSF) biomarkers of iron status are associated with CSF viral load, antiretroviral therapy, and demographic factors in HIV-infected adults. Fluids Barriers CNS 2017, 14, 11. [Google Scholar] [CrossRef]
- Chiou, B.; Connor, J.R. Emerging and Dynamic Biomedical Uses of Ferritin. Pharmaceuticals (Basel) 2018, 11, 124. [Google Scholar] [CrossRef]
- Kaur, H.; Bush, W.S.; Letendre, S.L.; Ellis, R.J.; Heaton, R.K.; Patton, S.M.; Connor, J.R.; Samuels, D.C.; Franklin, D.R.; Hulgan, T.; Kallianpur, A.R. Higher CSF Ferritin Heavy-Chain (Fth1) and Transferrin Predict Better Neurocognitive Performance in People with HIV. Mol Neurobiol 2021, 58, 4842–4855. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Arosio, P.; Poli, M.; Bou-Abdallah, F. A Novel Approach for the Synthesis of Human Heteropolymer Ferritins of Different H to L Subunit Ratios. J Mol Biol 2021, 433, 167198. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, S.; Wang, H.; Cui, J.; Tian, X.; Miao, Y.; Zhang, C.; Cao, L.; Ma, L.; Xu, X.; Qiao, Y.; Zhang, X. Transcriptional Repression of Ferritin Light Chain Increases Ferroptosis Sensitivity in Lung Adenocarcinoma. Front Cell Dev Biol 2021, 9, 719187. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Lu, J.; Hao, X.; Li, H.; Zhang, G.; Liu, X.; Li, X.; Zhao, C.; Kuang, W.; Chen, D.; Zhu, M. FTH1 Inhibits Ferroptosis Through Ferritinophagy in the 6-OHDA Model of Parkinson's Disease. Neurotherapeutics 2020, 17, 1796–1812. [Google Scholar] [CrossRef]
- Arosio, P.; Carmona, F.; Gozzelino, R.; Maccarinelli, F.; Poli, M. The importance of eukaryotic ferritins in iron handling and cytoprotection. Biochem J 2015, 472, 1–15. [Google Scholar] [CrossRef]
- Asensio-Lopez, M.C.; Sanchez-Mas, J.; Pascual-Figal, D.A.; de Torre, C.; Valdes, M.; Lax, A. Ferritin heavy chain as main mediator of preventive effect of metformin against mitochondrial damage induced by doxorubicin in cardiomyocytes. Free Radic Biol Med 2014, 67, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Chiou, B.; Lucassen, E.; Sather, M.; Kallianpur, A.; Connor, J. Semaphorin4A and H-ferritin utilize Tim-1 on human oligodendrocytes: A novel neuro-immune axis. Glia 2018, 66, 1317–1330. [Google Scholar] [CrossRef]
- Palsa, K.; Neely, E.B.; Baringer, S.L.; Helmuth, T.B.; Simpson, I.A.; Connor, J.R. Brain iron acquisition depends on age and sex in iron-deficient mice. FASEB J 2024, 38, e23331. [Google Scholar] [CrossRef]
- Baringer, S.L.; Neely, E.B.; Palsa, K.; Simpson, I.A.; Connor, J.R. Regulation of brain iron uptake by apo- and holo-transferrin is dependent on sex and delivery protein. Fluids Barriers CNS 2022, 19, 49. [Google Scholar] [CrossRef]
- Masters, M.C.; Perez, J.; Wu, K.; Ellis, R.J.; Goodkin, K.; Koletar, S.L.; Andrade, A.; Yang, J.; Brown, T.T.; Palella, F.J.; Sacktor, N.; Tassiopoulos, K.; Erlandson, K.M. Baseline Neurocognitive Impairment (NCI) Is Associated With Incident Frailty but Baseline Frailty Does Not Predict Incident NCI in Older Persons With Human Immunodeficiency Virus (HIV). Clin Infect Dis 2021, 73, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Robertson, K.R.; Smurzynski, M.; Parsons, T.D.; Wu, K.; Bosch, R.J.; Wu, J.; McArthur, J.C.; Collier, A.C.; Evans, S.R.; Ellis, R.J. The prevalence and incidence of neurocognitive impairment in the HAART era. AIDS 2007, 21, 1915–1921. [Google Scholar] [CrossRef]
- Ellis, R.J.; Evans, S.R.; Clifford, D.B.; Moo, L.R.; McArthur, J.C.; Collier, A.C.; Benson, C.; Bosch, R.; Simpson, D.; Yiannoutsos, C.T.; Yang, Y.; Robertson, K.; Neurological AIDS Research Consortium; AIDS Clinical Trials Group Study Teams A5001 and A362. Clinical validation of the NeuroScreen. J Neurovirol 2005, 11, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Milne, G.L.; Dai, Q.; Roberts, L.J., 2nd. The isoprostanes-25 years later. Biochim Biophys Acta 2015, 1851, 433–445. [Google Scholar] [CrossRef]
- Milne, G.L.; Nogueira, M.S.; Gao, B.; Sanchez, S.C.; Amin, W.; Thomas, S.; Oger, C.; Galano, J.M.; Murff, H.J.; Yang, G.; Durand, T. Identification of novel F2-isoprostane metabolites by specific UDP-glucuronosyltransferases. Redox Biol 2024, 70, 103020. [Google Scholar] [CrossRef]
- Milne, G.L.; Gao, B.; Terry, E.S.; Zackert, W.E.; Sanchez, S.C. Measurement of F2- isoprostanes and isofurans using gas chromatography-mass spectrometry. Free Radic Biol Med 2013, 59, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Knovich, M.A.; Coffman, L.G.; Torti, F.M.; Torti, S.V. Serum ferritin: Past.; present and future. Biochim Biophys Acta 2010, 1800, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. The Discovery of the Iron-Regulatory Hormone Hepcidin. Clin Chem 2019, 65, 1330–1331. [Google Scholar] [CrossRef] [PubMed]
- Kernan, K.F.; Carcillo, J.A. Hyperferritinemia and inflammation. Int Immunol 2017, 29, 401–409. [Google Scholar] [CrossRef]
- Guha, D.; Misra, V.; Yin, J.; Gabuzda, D. CSF Inflammation Markers Associated with Asymptomatic Viral Escape in Cerebrospinal Fluid of HIV-Positive Individuals on Antiretroviral Therapy. Viruses 2023, 15, 1829. [Google Scholar] [CrossRef]
- Okwuegbuna, O.K.; Kaur, H.; Jennifer, I.; Bush, W.S.; Bharti, A.; Umlauf, A.; Ellis, R.J.; Franklin, D.R.; Heaton, R.K.; McCutchan, J.A.; Kallianpur, A.R.; Letendre, S.L. Anemia and Erythrocyte Indices Are Associated With Neurocognitive Performance Across Multiple Ability Domains in Adults With HIV. J Acquir Immune Defic Syndr 2023, 92, 414–421. [Google Scholar] [CrossRef]
- Orino, K.; Lehman, L.; Tsuji, Y.; Ayaki, H.; Torti, S.V.; Torti, F.M. Ferritin and the response to oxidative stress. Biochem J 2001, 357 Pt 1, 241–247. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Armitage, A.E.; Stacey, A.R.; Giannoulatou, E.; Marshall, E.; Sturges, P.; Chatha, K.; Smith, N.M.; Huang, X.; Xu, X.; Pasricha, S.R.; Li, N.; Wu, H.; Webster, C.; Prentice, A.M.; Pellegrino, P.; Williams, I.; Norris, P.J.; Drakesmith, H.; Borrow, P. Distinct patterns of hepcidin and iron regulation during HIV-1, HBV, and HCV infections. Proc Natl Acad Sci U S A 2014, 111, 12187–12192. [Google Scholar] [CrossRef]
- Quiros-Roldan, E.; Castelli, F.; Lanza, P.; Pezzoli, C.; Vezzoli, M.; Biasiotto, G.; Zanella, I.; Inflammation in HIV Study Group. The impact of antiretroviral therapy on iron homeostasis and inflammation markers in HIV-infected patients with mild anemia. J Transl Med 2017, 15, 256. [Google Scholar] [CrossRef]
- Jensen, B.K.; Roth, L.M.; Grinspan, J.B.; Jordan-Sciutto, K.L. White matter loss and oligodendrocyte dysfunction in HIV: A consequence of the infection, the antiretroviral therapy or both? Brain Res 2019, 1724, 146397. [Google Scholar] [CrossRef]
- Kuhn, T.; Jin, Y.; Huang, C.; Kim, Y.; Nir, T.M.; Gullett, J.M.; Jones, J.D.; Sayegh, P.; Chung, C.; Dang, B.H.; Singer, E.J.; Shattuck, D.W.; Jahanshad, N.; Bookheimer, S.Y.; Hinkin, C.H.; Zhu, H.; Thompson, P.M.; Thames, A.D. The joint effect of aging and HIV infection on microstructure of white matter bundles. Hum Brain Mapp 2019, 40, 4370–4380. [Google Scholar] [CrossRef]
- Yoshihara, Y.; Kato, T.; Watanabe, D.; Fukumoto, M.; Wada, K.; Oishi, N.; Nakakura, T.; Kuriyama, K.; Shirasaka, T.; Murai, T. Altered white matter microstructure and neurocognitive function of HIV-infected patients with low nadir CD4. J Neurovirol 2022, 28, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Alakkas, A.; Ellis, R.J.; Watson, C.W.; Umlauf, A.; Heaton, R.K.; Letendre, S.; Collier, A.; Marra, C.; Clifford, D.B.; Gelman, B.; Sacktor, N.; Morgello, S.; Simpson, D.; McCutchan, J.A.; Kallianpur, A.; Gianella, S.; Marcotte, T.; Grant, I.; Fennema-Notestine, C.; CHARTER Group. White matter damage, neuroinflammation, and neuronal integrity in HAND. J Neurovirol 2019, 25, 32–41. [Google Scholar] [CrossRef]
- Todorich, B.; Zhang, X.; Connor, J.R. H-ferritin is the major source of iron for oligodendrocytes. Glia 2011, 59, 927–935. [Google Scholar] [CrossRef]
- Roth, A.D.; Núñez, M.T. Oligodendrocytes: Functioning in a Delicate Balance Between High Metabolic Requirements and Oxidative Damage. Adv Exp Med Biol 2016, 949, 167–181. [Google Scholar]
- Cheli, V.T.; Santiago González, D.A.; Wan, Q.; Denaroso, G.; Wan, R.; Rosenblum, S.L.; Paez, P.M. H-ferritin expression in astrocytes is necessary for proper oligodendrocyte development and myelination. Glia 2021, 12, 2981–2998. [Google Scholar] [CrossRef]
- Mukherjee, C.; Kling, T.; Russo, B.; Miebach, K.; Kess, E.; Schifferer, M.; Pedro, L.D.; Weikert, U.; Fard, M.K.; Kannaiyan, N.; Rossner, M.; Aicher, M.L.; Goebbels, S.; Nave, K.A.; Krämer-Albers, E.M.; Schneider, A.; Simons, M. Oligodendrocytes Provide Antioxidant Defense Function for Neurons by Secreting Ferritin Heavy Chain. Cell Metab 2020, 32, 259–272. [Google Scholar] [CrossRef]
- Zhang, X.; Surguladze, N.; Slagle-Webb, B.; Cozzi, A.; Connor, J.R. Cellular iron status influences the functional relationship between microglia and oligodendrocytes. Glia 2006, 54, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Saylor, D.; Dickens, A.M.; Sacktor, N.; Haughey, N.; Slusher, B.; Pletnikov, M.; Mankowski, J.L.; Brown, A.; Volsky, D.J.; McArthur, J.C. HIV-associated neurocognitive disorder-pathogenesis and prospects for treatment. Nat Rev Neurol 2016, 12, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Nnah, I.C.; Wessling-Resnick, M. Brain Iron Homeostasis: A Focus on Microglial Iron. Pharmaceuticals (Basel) 2018, 11, 129. [Google Scholar] [CrossRef]
- Garrido-Rodríguez, V.; Álvarez-Ríos, A.I.; Olivas-Martínez, I.; Pozo-Balado, M.D.M.; Bulnes-Ramos, Á.; Leal, M.; Pacheco, Y.M. Dysregulation of iron metabolism modulators in virologically suppressed HIV-infected patients. Front Immunol 2022, 13, 977316. [Google Scholar] [CrossRef]
- Wan, R.; Cheli, V.T.; Santiago-Gonzalez, D.A.; Rosenblum, S.L.; Wan, Q.; Paez, P.M. Impaired Postnatal Myelination in a Conditional Knockout Mouse for the Ferritin Heavy Chain in Oligodendroglial Cells. J Neurosci 2020, 40, 7609–7624. [Google Scholar] [CrossRef]
- Zarjou, A.; Black, L.M.; McCullough, K.R.; Hull, T.D.; Esman, S.K.; Boddu, R.; Varambally, S.; Chandrashekar, D.S.; Feng, W.; Arosio, P.; Poli, M.; Balla, J.; Bolisetty, S. Ferritin Light Chain Confers Protection Against Sepsis-Induced Inflammation and Organ Injury. Front Immunol 2019, 10, 131. [Google Scholar] [CrossRef]
- Finazzi, D.; Arosio, P. Biology of ferritin in mammals: an update on iron storage, oxidative damage and neurodegeneration. Arch Toxicol 2014, 88, 1787–1802. [Google Scholar] [CrossRef]
- Almer, G.; Teismann, P.; Stevic, Z.; Halaschek-Wiener, J.; Deecke, L.; Kostic, V.; Przedborski, S. Increased levels of the pro-inflammatory prostaglandin PGE2 in CSF from ALS patients. Neurology 2002, 58, 1277–1279. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.D.; Noren Hooten, N.; Trzeciak, A.R.; Evans, M.K. Markers of oxidant stress that are clinically relevant in aging and age-related disease. Mech Ageing 2013, 134, 139–157. [Google Scholar] [CrossRef]
- Ambegaokar, S.S.; Kolson, D.L. Heme oxygenase-1 dysregulation in the brain: implications for HIV-associated neurocognitive disorders. Curr HIV Res 2014, 12, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Sfera, A.; Bullock, K.; Price, A.; Inderias, L.; Osorio, C. Ferrosenescence: The iron age of neurodegeneration? Mech Ageing Dev 2018, 174, 63–75. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; Morrison, B., 3rd; Stockwell, B.R. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Yan, Q.; Zheng, W.; Jiang, Y.; Zhou, P.; Lai, Y.; Liu, C.; Wu, P.; Zhuang, H.; Huang, H.; Li, G.; Zhan, S.; Lao, Z.; Liu, X. Transcriptomic reveals the ferroptosis features of host response in a mouse model of Zika virus infection. J Med Virol 2023, 95, e28386. [Google Scholar] [CrossRef]
- Mitra, J.; Kodavati, M.; Provasek, V.E.; Rao, K.S.; Mitra, S.; Hamilton, D.J.; Horner, P.J.; Vahidy, F.S.; Britz, G.W.; Kent, T.A.; Hegde, M.L. SARS-CoV-2 and the central nervous system: Emerging insights into hemorrhage-associated neurological consequences and therapeutic considerations. Ageing Res Rev 2022, 80, 101687. [Google Scholar] [CrossRef]
- Zhang, R.; Sun, C.; Chen, X.; Han, Y.; Zang, W.; Jiang, C.; Wang, J.; Wang, J. COVID-19-Related Brain Injury: The Potential Role of Ferroptosis. J Inflamm Res 2022, 15, 2181–2198. [Google Scholar] [CrossRef]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron homeostasis and oxidative stress: An intimate relationship. Biochim Biophys Acta Mol Cell Res 2019, 1866, 118535. [Google Scholar] [CrossRef]
- Coradduzza, D.; Congiargiu, A.; Chen, Z.; Zinellu, A.; Carru, C.; Medici, S. Ferroptosis and Senescence: A Systematic Review. Int J Mol Sci 2023, 24, 3658. [Google Scholar] [CrossRef]
- Cairo, G.; Tacchini, L.; Pogliaghi, G.; Anzon, E.; Tomasi, A.; Bernelli-Zazzera, A. Induction of ferritin synthesis by oxidative stress. Transcriptional and post-transcriptional regulation by expansion of the “free” iron pool. J Biol Chem 1995, 270, 700–703. [Google Scholar] [CrossRef]
- Muhoberac, B.B.; Vidal, R. Iron.; Ferritin.; Hereditary Ferritinopathy, and Neurodegeneration. Front Neurosci 2019, 13, 1195. [Google Scholar] [CrossRef]
- Yang, X.; Ding, Y.; Sun, L.; Shi, M.; Zhang, P.; Huang, Z.; Wang, J.; He, A.; Wang, J.; Wei, J.; Liu, M.; Liu, J.; Wang, G.; Yang, X.; Li, R. Ferritin light chain deficiency-induced ferroptosis is involved in preeclampsia pathophysiology by disturbing uterine spiral artery remodelling. Redox Biol 2022, 58, 102555. [Google Scholar] [CrossRef]
- Sfera, A.; Thomas, K.G.; Andronescu, C.V.; Jafri, N.; Sfera, D.O.; Sasannia, S.; Zapata-Martín Del Campo, C.M.; Maldonado, J.C. Bromodomains in Human-Immunodeficiency Virus-Associated Neurocognitive Disorders: A Model of Ferroptosis-Induced Neurodegeneration. Front Neurosci 2022, 16, 904816. [Google Scholar] [CrossRef]
- Redig, A.J.; Berliner, N. Pathogenesis and clinical implications of HIV-related anemia in 2013. Hematology Am Soc Hematol Educ Program 2013, 2013, 377–381. [Google Scholar] [CrossRef]
- Kallianpur, A.R.; Wang, Q.; Jia, P.; Hulgan, T.; Zhao, Z.; Letendre, S.L.; Ellis, R.J.; Heaton, R.K.; Franklin, D.R.; Barnholtz-Sloan, J.; Collier, A.C.; Marra, C.M.; Clifford, D.B.; Gelman, B.B.; McArthur, J.C.; Morgello, S.; Simpson, D.M.; McCutchan, J.A.; Grant, I.; CHARTER Study Group. Anemia and Red Blood Cell Indices Predict HIV-Associated Neurocognitive Impairment in the Highly Active Antiretroviral Therapy Era. J Infect Dis 2016, 213, 1065–1073. [Google Scholar] [CrossRef]
- Drakesmith, H.; Prentice, A. Viral infection and iron metabolism. Nat Rev Microbiol 2008, 6, 541–552. [Google Scholar] [CrossRef]
- Kannan, M.; Sil, S.; Oladapo, A.; Thangaraj, A.; Periyasamy, P.; Buch, S. HIV-1 Tat-mediated microglial ferroptosis involves the miR-204-ACSL4 signaling axis. Redox Biol 2023, 62, 102689. [Google Scholar] [CrossRef]
- Duck, K.A.; Neely, E.B.; Simpson, I.A.; Connor, J.R. A role for sex and a common HFE gene variant in brain iron uptake. J Cereb Blood Flow Metab 2018, 38, 540–548. [Google Scholar] [CrossRef]
- Bartzokis, G.; Lu, P.H.; Tingus, K.; Peters, D.G.; Amar, C.P.; Tishler, T.A.; Finn, J.P.; Villablanca, P.; Altshuler, L.L.; Mintz, J.; Neely, E.; Connor, J.R. Gender and iron genes may modify associations between brain iron and memory in healthy aging. Neuropsychopharmacology 2011, 36, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Persson, N.; Wu, J.; Zhang, Q.; Liu, T.; Shen, J.; Bao, R.; Ni, M.; Liu, T.; Wang, Y.; Spincemaille, P. Age and sex related differences in subcortical brain iron concentrations among healthy adults. Neuroimage 2015, 122, 385–398. [Google Scholar] [CrossRef]
- Li, G.; Tong, R.; Zhang, M.; Gillen, K.M.; Jiang, W.; Du, Y.; Wang, Y.; Li, J. Age-dependent changes in brain iron deposition and volume in deep gray matter nuclei using quantitative susceptibility mapping. Neuroimage 2023, 269, 119923. [Google Scholar] [CrossRef]
- Rubin, L.H.; Maki, P.M. Neurocognitive Complications of HIV Infection in Women: Insights from the WIHS Cohort. Curr Top Behav Neurosci 2021, 50, 175–191. [Google Scholar]
- Dreyer, A.J.; Munsami, A.; Williams, T.; Andersen, L.S.; Nightingale, S.; Gouse, H.; Joska, J.; Thomas, K.G.F. Cognitive Differences between Men and Women with HIV: A Systematic Review and Meta-Analysis. Arch Clin Neuropsychol 2022, 37, 479–496. [Google Scholar] [CrossRef]
- Gao, G.; You, L.; Zhang, J.; Chang, Y.Z.; Yu, P. Brain Iron Metabolism, Redox Balance and Neurological Diseases. Antioxidants (Basel) 2023, 12, 1289. [Google Scholar] [CrossRef] [PubMed]
- Pond, R.A.; Collins, L.F.; Lahiri, C.D. Sex Differences in Non-AIDS Comorbidities Among People With Human Immunodeficiency Virus. Open Forum Infect Dis 2021, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Kallianpur, A.R.; Gittleman, H.; Letendre, S.; Ellis, R.; Barnholtz-Sloan, J.S.; Bush, W.S.; Heaton, R.; Samuels, D.C.; Franklin, D.R., Jr. Rosario-Cookson, D.; Clifford, D.B.; Collier, A.C.; Gelman, B.; Marra, C.M.; McArthur, J.C.; McCutchan, J.A.; Morgello, S.; Grant, I.; Simpson, D.; Connor, J.R.; Hulgan, T.; CHARTER Study Group. Cerebrospinal Fluid Ceruloplasmin, Haptoglobin, and Vascular Endothelial Growth Factor Are Associated with Neurocognitive Impairment in Adults with HIV Infection. Mol Neurobiol 2019, 56, 3808–3818. [Google Scholar]
- Mehta, S.R.; Pérez-Santiago, J.; Hulgan, T.; Day, T.R.; Barnholtz-Sloan, J.; Gittleman, H.; Letendre, S.; Ellis, R.; Heaton, R.; Patton, S.; Suben, J.D.; Franklin, D.; Rosario, D.; Clifford, D.B.; Collier, A.C.; Marra, C.M.; Gelman, B.B.; McArthur, J.; McCutchan, A.; Morgello, S.; Simpson, D.; Connor, J.; Grant, I.; Kallianpur, A. Cerebrospinal fluid cell-free mitochondrial DNA is associated with HIV replication, iron transport, and mild HIV-associated neurocognitive impairment. J Neuroinflammation 2017, 14, 72. [Google Scholar] [CrossRef]
- Chen, X.; Tsvetkov, A.S.; Shen, H.M.; Isidoro, C.; Ktistakis, N.T.; Linkermann, A.; Koopman, W.J.H.; Simon, H.U.; Galluzzi, L.; Luo, S.; Xu, D.; Gu, W.; Peulen, O.; Cai, Q.; Rubinsztein, D.C.; Chi, J.T.; Zhang, D.D.; Li, C.; Toyokuni, S.; Liu, J.; Roh, J.L.; Dai, E.; Juhasz, G.; Liu, W.; Zhang, J.; Yang, M.; Liu, J.; Zhu, L.Q.; Zou, W.; Piacentini, M.; Ding, W.X.; Yue, Z.; Xie, Y.; Petersen, M.; Gewirtz, D.A.; Mandell, M.A.; Chu, C.T.; Sinha, D.; Eftekharpour, E.; Zhivotovsky, B.; Besteiro, S.; Gabrilovich, D.I.; Kim, D.H.; Kagan, V.E.; Bayir, H.; Chen, G.C.; Ayton, S.; Lünemann, J.D.; Komatsu, M.; Krautwald, S.; Loos, B.; Baehrecke, E.H.; Wang, J.; Lane, J.D.; Sadoshima, J.; Yang, W.S.; Gao, M.; Münz, C.; Thumm, M.; Kampmann, M.; Yu, D.; Lipinski, M.M.; Jones, J.W.; Jiang, X.; Zeh, H.J.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Tang, D. International consensus guidelines for the definition, detection, and interpretation of autophagy-dependent ferroptosis. Autophagy 2024, 24, 1–34. [Google Scholar] [CrossRef]
- Kim, J.; Wessling-Resnick, M. Iron and mechanisms of emotional behavior. J Nutr Biochem 2014, 25, 1101–1107. [Google Scholar] [CrossRef]
- Xu, Y.; Jia, B.; Li, J.; Li, Q.; Luo, C. The Interplay between Ferroptosis and Neuroinflammation in Central Neurological Disorders. Antioxidants (Basel) 2024, 13, 395. [Google Scholar] [CrossRef]
- Minchella, P.A.; Armitage, A.E.; Darboe, B.; Jallow, M.W.; Drakesmith, H.; Jaye, A.; Prentice, A.M.; McDermid, J.M. Elevated Hepcidin Is Part of a Complex Relation That Links Mortality with Iron Homeostasis and Anemia in Men and Women with HIV Infection. J Nutr 2015 145, 1194–1201. [CrossRef]
- Wolters, F.J.; Zonneveld, H.I.; Licher, S.; Cremers, L.G.M.; Ikram, M.K.; Koudstaal, P.J.; Vernooij, M.W.; Ikram, M.A.; Heart Brain Connection Collaborative Research Group. Hemoglobin and anemia in relation to dementia risk and accompanying changes on brain MRI. Neurology. [CrossRef] [PubMed]
Figure 1.
Forest plot of multivariable associations of (A) ferritin-light-chain (Ftl) and (B) ferritin heavy-chain-1 (Fth-1) levels at entry with NCI in people with HIV (all study participants and stratified by sex). Adjusted odds ratios (ORs) and their 95% confidence intervals are shown. Lower ORs (<1.0) indicate a lower likelihood of NCI. See text for covariates included in regression models. *p<0.05, **p<0.005. All p-values<0.05 are considered statistically significant.
Figure 1.
Forest plot of multivariable associations of (A) ferritin-light-chain (Ftl) and (B) ferritin heavy-chain-1 (Fth-1) levels at entry with NCI in people with HIV (all study participants and stratified by sex). Adjusted odds ratios (ORs) and their 95% confidence intervals are shown. Lower ORs (<1.0) indicate a lower likelihood of NCI. See text for covariates included in regression models. *p<0.05, **p<0.005. All p-values<0.05 are considered statistically significant.
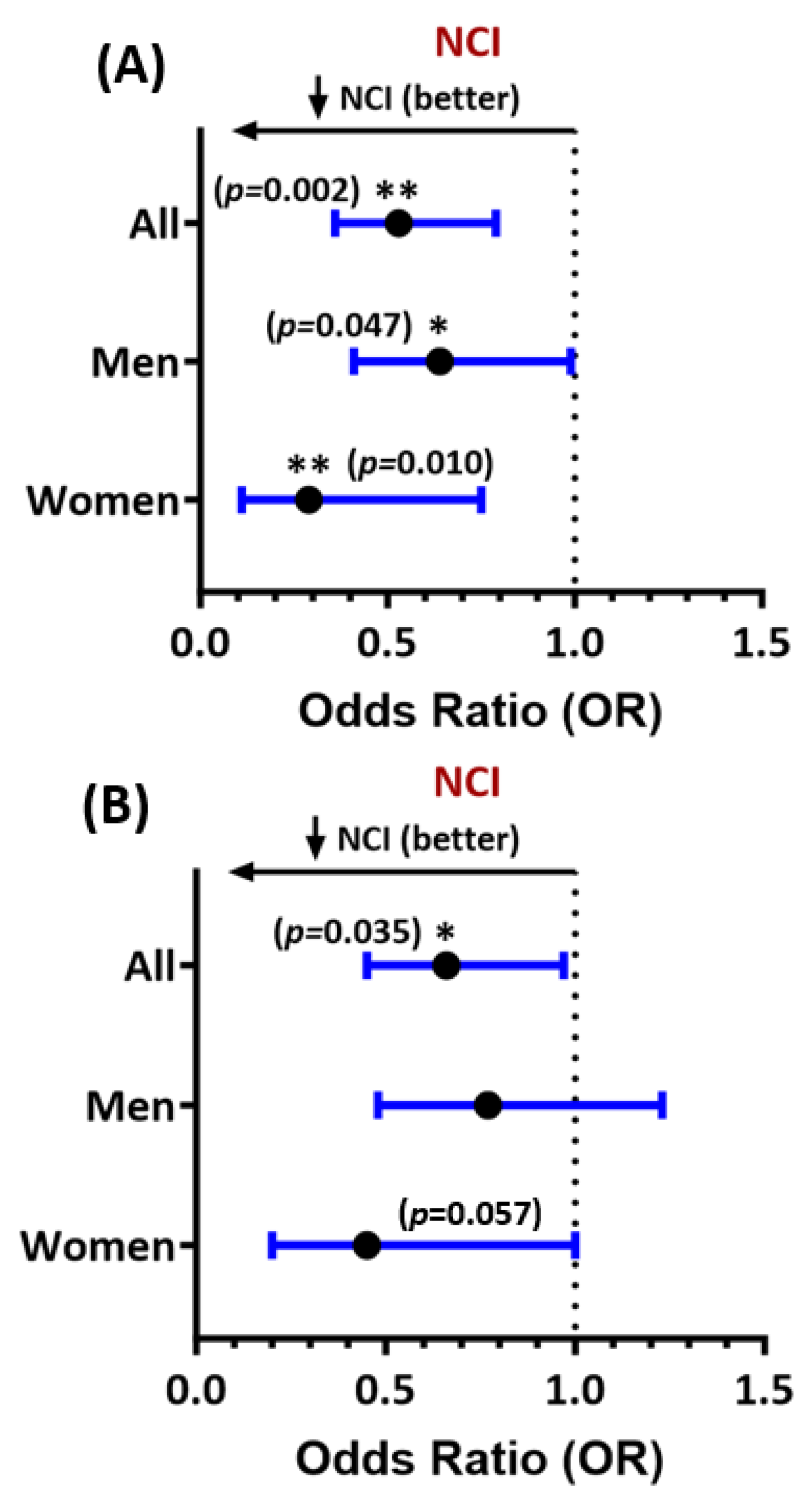
Figure 2.
Forest plot of adjusted associations of (A) Ftl levels at entry and global neurocognitive performance (NPZ-4 test score), and (B) Fth-1 levels at entry with the NPZ-4 test score. Results shown are adjusted beta estimates and their 95% confidence intervals for all study participants, men, and women. Higher adjusted beta estimates (values>0) indicate better neurocognitive function. See text for model-specific covariates. *p<0.05, **p<0.005. All p-values<0.05 are considered statistically significant.
Figure 2.
Forest plot of adjusted associations of (A) Ftl levels at entry and global neurocognitive performance (NPZ-4 test score), and (B) Fth-1 levels at entry with the NPZ-4 test score. Results shown are adjusted beta estimates and their 95% confidence intervals for all study participants, men, and women. Higher adjusted beta estimates (values>0) indicate better neurocognitive function. See text for model-specific covariates. *p<0.05, **p<0.005. All p-values<0.05 are considered statistically significant.
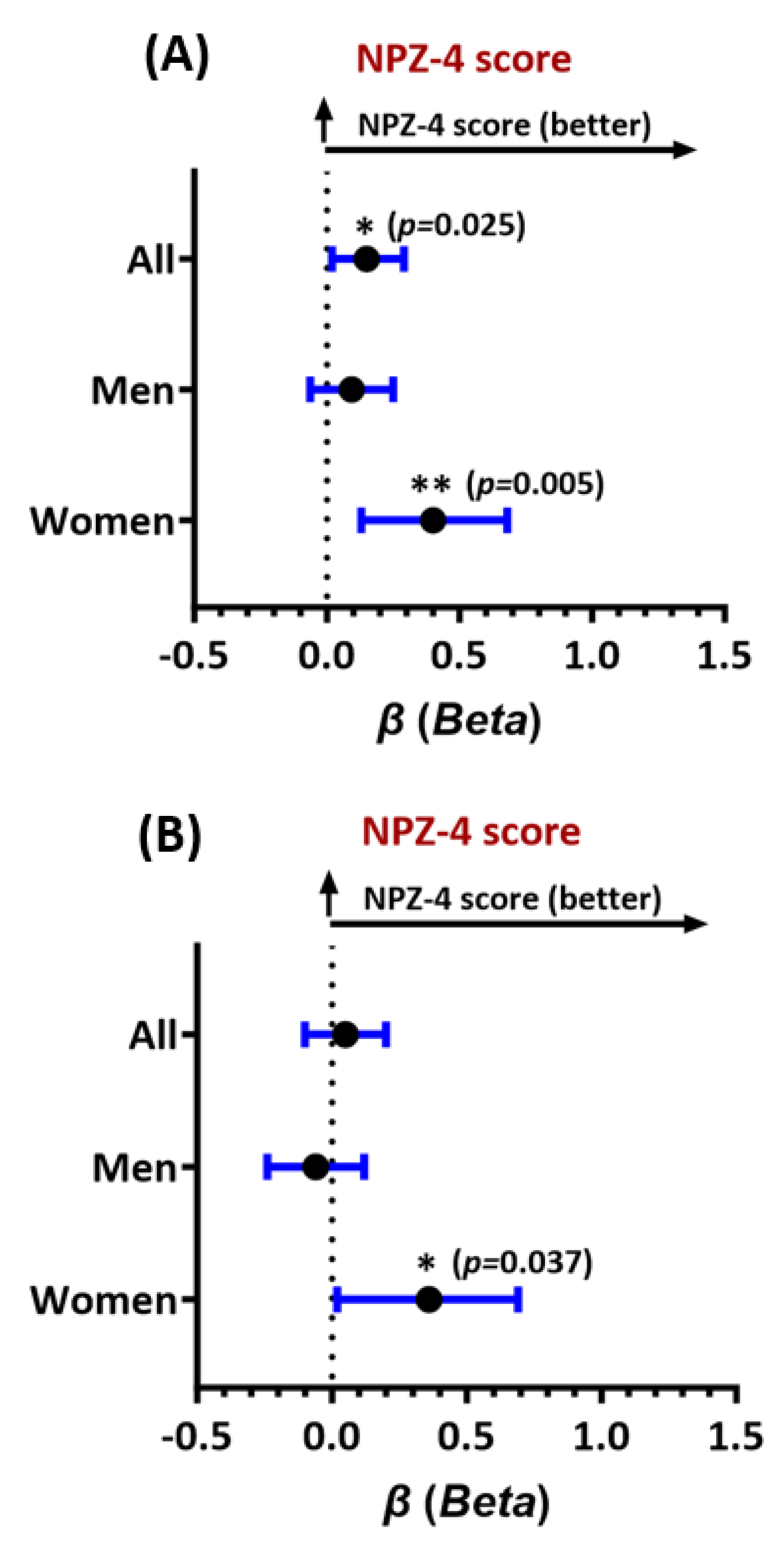

Table 1.
Characteristics of Study Participants at Entry.
| Variable | Neurocognitively Normal (N=237) |
Neurocognitively Impaired1 (N=82) |
|---|---|---|
| Age, mean (SD) | 51.4 (6.9) | 53.7 (8.3) |
| Sex, % female | 17.3 | 24.4 |
| Non-Hispanic Black2, % | 31.6 | 20.74 |
| Hispanic2, % | 20.7 | 36.6** |
| Efavirenz use, % | 51.5 | 39.05 |
|
Nadir CD4+ T-cells/µl, median (IQR) |
192 (67, 314) | 237 (80, 347) |
| HIV RNA <200 copies/ml, % | 95.7 | 96.3 |
| Anemia, % | 10.5 | 12.7 |
| HCV seropositive, % | 2.53 | 8.54* |
| ≥ 2 Comorbidities3, % | 32.5 | 47.6* |
Abbreviations: SD, Standard Deviation; IQR, Interquartile Range; HCV, Hepatitis C Virus; µl, microliter. 1Neurocognitive impairment status was missing for 5 people with HIV, due to incomplete test score data. 2Self-reported race or ethnicity. 3Clinically significant comorbidities, e.g., cancer, diabetes, cardiovascular disease, chronic kidney disease, chronic HCV infection, bone fractures, prior transient ischemic attack or stroke. No study participants reported active substance use. 4p=0.060; 5p=0.050; statistically significant p-values: *p<0.05; **p<0.01.
Table 2.
Summary of Serum and Urinary Biomarker Levels at Entry in People with HIV.
| Biomarker | All PWH (N=324) Median (IQR) |
Men (N=263) Median (IQR) |
Women (N=61) Median (IQR) |
|---|---|---|---|
| Serum Fth-1, ng/ml | 289 (199, 499) | 311 (197, 497) | 277 (136, 526) |
| Serum Ftl, ng/ml | 12.2 (7.7, 22.1) | 12.4 (7.7, 22.4) | 11 (7.2, 19.6) |
|
Urinary 15-series F2-IsoPs1, ng/mg Creatinine |
57 (32, 84) | 56 (31, 84) | 60 (43, 78) |
|
Urinary 5-series F2-IsoPs1, ng/mg Creatinine |
71 (34, 109) | 66 (33, 110) | 71 (38, 98) |
Abbreviations: Fth-1, ferritin heavy-chain-1; Ftl, ferritin light-chain; F2-IsoPs, urinary F2-isoprostanes (marker of in vivo lipid peroxidation and oxidative stress, corrected for renal function); IQR: Interquartile Range; ng, nanogram; ml, milliliter; mg, milligram; PWH, people with HIV. 1Urinary F2-IsoPs could not be quantified in 14 men (5.3% of men).
Table 3.
Biomarker Correlations with Inflammation and Lipid Peroxidation at Entry.
| Biomarker | IL-6 (N=324) rho (p) |
sTNFR-II (N=324) rho (p) |
sCD163 (N=324) rho (p) |
15-F2-IsoPs (N=310) rho (p) |
5-F2-IsoPs (N=310) rho (p) |
|---|---|---|---|---|---|
|
Fth-1 All PWH Women Men |
-0.007 (0.91) -0.085 (0.52) 0.013 (0.83) |
-0.043 (0.45) 0.128 (0.33) -0.085 (0.17) |
0.0005 (0.99) 0.157 (0.23) -0.041 (0.51) |
-0.061 (0.28) -0.290 (0.02) 0.016 (0.80) |
0.033 (0.55) -0.181 (0.16) 0.089 (0.15) |
|
Ftl All PWH Women Men |
-0.026 (0.64) 0.018 (0.89) -0.030 (0.63) |
-0.078 (0.16) 0.086 (0.51) -0.117 (0.06) |
-0.050 (0.37) 0.024 (0.85) -0.066 (0.29) |
-0.017 (0.77) -0.471 (<0.001) 0.094 (0.14) |
0.048 (0.39) -0.347 (<0.01) 0.134 (0.03) |
Abbreviations: IL-6, interleukin-6; sTNFR-II, soluble tumor necrosis factor receptor II; sCD163, soluble cluster differentiation antigen 163; Fth-1, ferritin heavy-chain-1; Ftl, ferritin light-chain; PWH, people with HIV; 5-F2-IsoPs and 15-F2-IsoPs, urinary 5- and 15-series F2-isoprostanes, corrected for serum creatinine. Rho values from Spearman’s correlations are shown, with corresponding p-values. p-values<0.05 (bolded) are considered statistically significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



