
Submitted:
23 August 2024
Posted:
26 August 2024
Read the latest preprint version here
Abstract
Although evolutionary transitions of individuality have been extensively theorized, little attention has been paid to the origin of levels of organization within organisms. How and why do specialized cells become organized into specialized tissues or organs? What spurs a transition in organizational level in cases where the function is already present in constituent cell types? We propose a model for this kind of evolutionary transition in terms of two key features of cellular self-maintenance: metabolic constraints on functional performance and the capacity for metabolic complementation between parenchymal and supporting cells. These features suggest a scenario whereby pre-existing specialized cell types are integrated into tissues when changes to the internal or external environment favor offloading metabolic burdens from a primary specialized cell type onto supporting cells. We illustrate this process of “supra-functionalization” using the nervous system and the pancreas as examples. The model reveals distinctive concerns for explaining this kind of evolutionary transition—instead of the suppression of “cheating” by components, the issue is how a tissue comes under modular genetic control as a distinct body part—and also points to a novel form of evolutionary complexity-increasing ratchet.
Keywords:
tissue
; cell metabolism
; subfunctionalization
1. A Different Kind of Evolutionary Transition
A central aim of evolutionary biology is to understand the origin and diversification of traits. Traditionally, it is assumed that the early establishment of a new trait is associated with the acquisition of a novel function [1]. Examples include the origin of implantation and eutherian viviparity [2], hypsodonty in ruminants [3], and the pharyngeal jaw apparatus of cichlids [4]. However, many organs perform functions that existed in their constituent cell types prior to the origin of the organ. For example, the endodermal cells that produce specific digestive and detoxifying enzymes in early branching chordates pre-date the origins of the vertebrate liver and pancreas, in which these cell types function as parenchymal cells [e.g., 5,6,7]. Additionally, neural cells pre-date the evolution of the central nervous system or even simpler ganglia and specific endocrine cells pre-date endocrine organs [e.g., 8]. These observations imply that the origins of at least some novelties are not associated with the origin of the function they perform. Instead, organs and tissues can arise by integration of an already functionally specialized cell type into a higher-level unit. Rather than co-opting a molecular pathway for a different function, cells can become integrated into a multicellular unit maintaining the original function. This immediately raises new questions: how and why do specialized cells become organized into a tissue or an organ? What spurs a transition in organizational level (e.g., from cells to tissues) when the function is already present?
Although there is an extensive literature on evolutionary transitions in individuality [9,10,11,12,13,14], especially with respect to the origin of multicellularity [e.g., 15,16,17,18], little attention has been paid to “organizational” transitions within organisms, such as the evolutionary origin of tissues in different multicellular lineages [19,20]. Recently, there has been a proliferation of work on animal tissues [see 21,22,23] that leverages new experimental tools (e.g., single cell transcriptomics). Because organisms in which functions are performed by distributed cells coexist today with organisms where those same functions are performed by tissues or organs, questions about the transition from one mode to another can be pursued empirically. Together these facts encourage a research program that can address the evolutionary mechanisms that underlie increases in the number of “levels of organization” above the cell but within the organism: tissues, organs, and anatomical parts.
Here we propose a model of tissue and organ origination that is motivated by the above observations and recent developments in tissue biology. Our model is based on the recognition that cells have a limited metabolic capacity for self-maintenance, but also have the ability to complement each other's metabolic needs. This creates the potential for cells to integrate into a higher-level unit through metabolic complementation that ultimately leads to interdependence among the cells and thus to the evolutionary stability of a new level of organization (i.e., a tissue). We present preliminary evidence for this scenario of an “organizational transition” to higher levels of integration and highlight several predictions it makes. Finally, we discuss the model in the context of existing ideas about major evolutionary transitions, outlining the contours of a research program that may illuminate tissue and organ origination.
2. Tissues, Organization, and Metabolism
Organization in a biological system is any configuration of biological entities that, through its internal structure and stereotypical interactions, takes on unit integrity over some temporal duration. Tissues are the epitome of organization because they integrate various cell types into a coherent, spatially localized functional unit. A tissue is conceptualized as a configuration of several cell types and the associated extracellular matrix (ECM). Most animal tissues include an epithelial cell type that is often also the parenchymal cell, dedicated to a specialized function, as well as tissue-specific fibroblasts and macrophages, endothelial cells, and a variable cast of immune and ancillary cells. (An exception is cartilage, with a single cell type, the chondrocyte, which also produces the ECM.) The hybrid epithelial-mesenchymal nature of many tissues and organs is also reflected in the fact that they largely develop from epithelial-mesenchymal interactions [24]. A major exception is the chordate central nervous system, which arises from extensive internal differentiation of an epithelial structure, the neural tube.
To ensure the proper proportion of different cell types in a tissue, constituent cells are engaged in continuous regulatory interactions via cell type-specific growth factors (for an example with fibroblast and macrophage, see [23]). The self-stabilizing nature of cell interactions within a tissue also explains the fact that tissues are typically locally exclusive (i.e., one tissue in a location excludes elements of other tissues from that same territory).
The traditional distinction between tissues and organs is not necessarily categorical and is consistent with a gradual transition between them. Some body parts can be understood as collections of appropriately arranged tissues, like a limb consisting of bone, connective tissue, muscles, nerves and vessels overlain by skin. Organs such as liver, lung or kidney largely consist of a highly organized “complex tissue.” The functionality of complex tissues depends on a spatially organized arrangement of parenchymal cells (e.g., glomeruli or loops of Henle). They also have additional scales of organization between the basic cell type and ECM complement and the whole, such as kidney nephrons or liver lobules [25]. The corresponding organ is not much more than the specialized complex tissue plus the “plumbing” that connects it to the rest of the organism via blood vessels, nerves, and ascending or descending tubes (bronchial tubes in the lung, ureter in the kidney). For this reason, we do not make a sharp distinction between “tissues” and “organs.”
Not all metazoans have tissues as characterized above. Cnidarians and ctenophores primarily consist of complex multifunctional epithelia with few mesenchymal cells. Most of our knowledge of tissue-level organization derives from vertebrate models. Among many other bilaterian animals, the nature of tissues is an underexplored topic and therefore we lack a firm understanding of the phylogenetic distribution of tissue-like levels of organization and their properties. Yet we can say with confidence that metazoan body plan evolution includes the segregation of specialized cell types into separate pockets of cells with associated mesenchymal support, conventionally called tissues and organs. How did this evolutionary transition occur?
2.1. Metabolism: Constraints and Complementation
There are two aspects of cellular metabolism that are crucial for our model of the origin of supracellular organization within multicellular organisms: the existence of metabolic constraints and the ability of cells to metabolically complement each other. Cellular metabolism refers to chemical reactions that provide free energy and biomass, thereby enabling cells to survive, grow, proliferate, and perform their functions. Each cell can only generate a finite amount of cellular energy by utilizing a limited number of metabolic pathways such as glycolysis, the Krebs cycle, the pentose phosphate pathway, and oxidative phosphorylation. Therefore, augmenting a function of the cell or introducing a new function reduces the amount of metabolic resources available for other pathways and cellular needs such as self-maintenance [26]. This is essentially a stoichiometric constraint—a molecule of glucose can only be used once. Such trade-offs are often invoked to explain the metabolic shifts in energy and biomass production pathways at high glucose and oxygen availability in unicellular organisms and tumor cells (Crabtree effect, Warburg effect; [27]).
A further characteristic of eukaryotic cells is that most of their metabolic reactions are employed in self-maintenance and performance of service functions, rather than the increase of biomass [28]. Consider the central metabolic process of carbohydrate metabolism. It begins with the uptake of glucose from the environment and its processing to generate either: (i) energy (ATP) via glycolysis and respiration, (ii) molecular building blocks (nucleotides, amino acid precursors), or (iii) reducing molecules like NADPH (e.g., via the pentose phosphate pathway) to counter the oxidative stress of ATP production during respiration. Importantly, the underlying metabolic pathways for these tasks partially overlap and thereby compete for resources and directly influence one another. This interconnectedness generates inherent trade-offs. For example, increased ATP production via oxidative phosphorylation results in an increased reactive oxygen species (ROS) load, which in turn increases the requirement for production of antioxidants. Similar trade-offs exist between ATP production and the synthesis of macromolecular building blocks needed for DNA repair, and between the replacement of damaged proteins and cellular proliferation. Each of the three core aspects of cellular metabolism are therefore intricately interdependent: ATP production for survival and the performance of service functions, redox homeostasis enabling chemical conversions and preventing oxidative damage (to DNA, lipids and proteins), and the synthesis of building blocks for repair and proliferation. Thus, the cell considered in isolation is inherently constrained in its ability to invest more in a specific functional role like producing and secreting enzymes or building and maintaining electrical membrane potential due to the interdependencies among its various metabolic pathways.
In addition to metabolic constraints, there is also the phenomenon of metabolic complementation. Like unicellular organisms, cells in multicellular organisms are capable of importing and exporting a great number of metabolic compounds (amino acids, sugars, small metabolites like lactate and acetate, etc.), provided that appropriate membrane transport mechanisms are in place. This includes compounds that the cell is principally able to synthesize itself, though perhaps at an insufficient rate. Moreover, many of these compounds feed into the above mentioned pathways of cellular metabolism: ATP production, redox homeostasis, and building block synthesis. For example, cells import lactate from the extracellular space, which, after oxidation to pyruvate, can enter respiration (the ATP producing citric cycle and electron transport chain), supplementing internal pyruvate production from glycolysis [29,30]. On the other hand, cells can exchange antioxidants and their precursors, such as cystine or cysteine, the rate-limiting amino acids for the synthesis of the antioxidant glutathione [31]. Similarly, the imported building blocks can serve in the synthesis of macromolecules.
3. A Model for the Origin of Tissues and Organs
We propose that the integration of pre-existing specialized cell types into higher-level units consisting of different cell types is driven by the offloading of metabolic burdens from a parenchymal cell type onto supporting cells. This would allow the augmentation of a specific “service” function carried out by the parenchymal cell type to benefit the organism. The selective demand for such an augmentation can arise due to a change in the external or internal environment. For example, a more toxic food source will favor a higher investment by the parenchymal cell into detoxification or a new prey type (or predator) may favor more metabolic investment in locomotion. Internal selection—functional demands arising from inside the organism—can have a similar effect: increases in body size and associated allometric changes in body volume may exceed the limits of diffusion processes, requiring some cell types to increase their investment into detoxifying waste products and for others to specialize in the transport of resources or metabolites into and out of the body. This not only establishes selective regimes for the augmented service functions, but it also selects for increased investment in other functions, such as transportation or skeletal support [32].
Figure 1.
An illustration of the metabolic profiles in the proposed model of supra-functionalization. A. Individual parenchymal cells are constrained by a metabolic trade-off between function performance and self-maintenance, represented by the red curve. B. Metabolic complementation by supporting cells displaces the trade-off curve (blue) for the parenchymal cell further away from the limit of minimal self-maintenance required for survival. This allows the parenchymal cell to augment its functional performance in response to changes in the external or internal selective environment. As a consequence, the parenchymal cell becomes dependent on the supporting cell for survival, and the function originally performed by the parenchymal cell is now performed by a “tissue.”.
Figure 1.
An illustration of the metabolic profiles in the proposed model of supra-functionalization. A. Individual parenchymal cells are constrained by a metabolic trade-off between function performance and self-maintenance, represented by the red curve. B. Metabolic complementation by supporting cells displaces the trade-off curve (blue) for the parenchymal cell further away from the limit of minimal self-maintenance required for survival. This allows the parenchymal cell to augment its functional performance in response to changes in the external or internal selective environment. As a consequence, the parenchymal cell becomes dependent on the supporting cell for survival, and the function originally performed by the parenchymal cell is now performed by a “tissue.”.
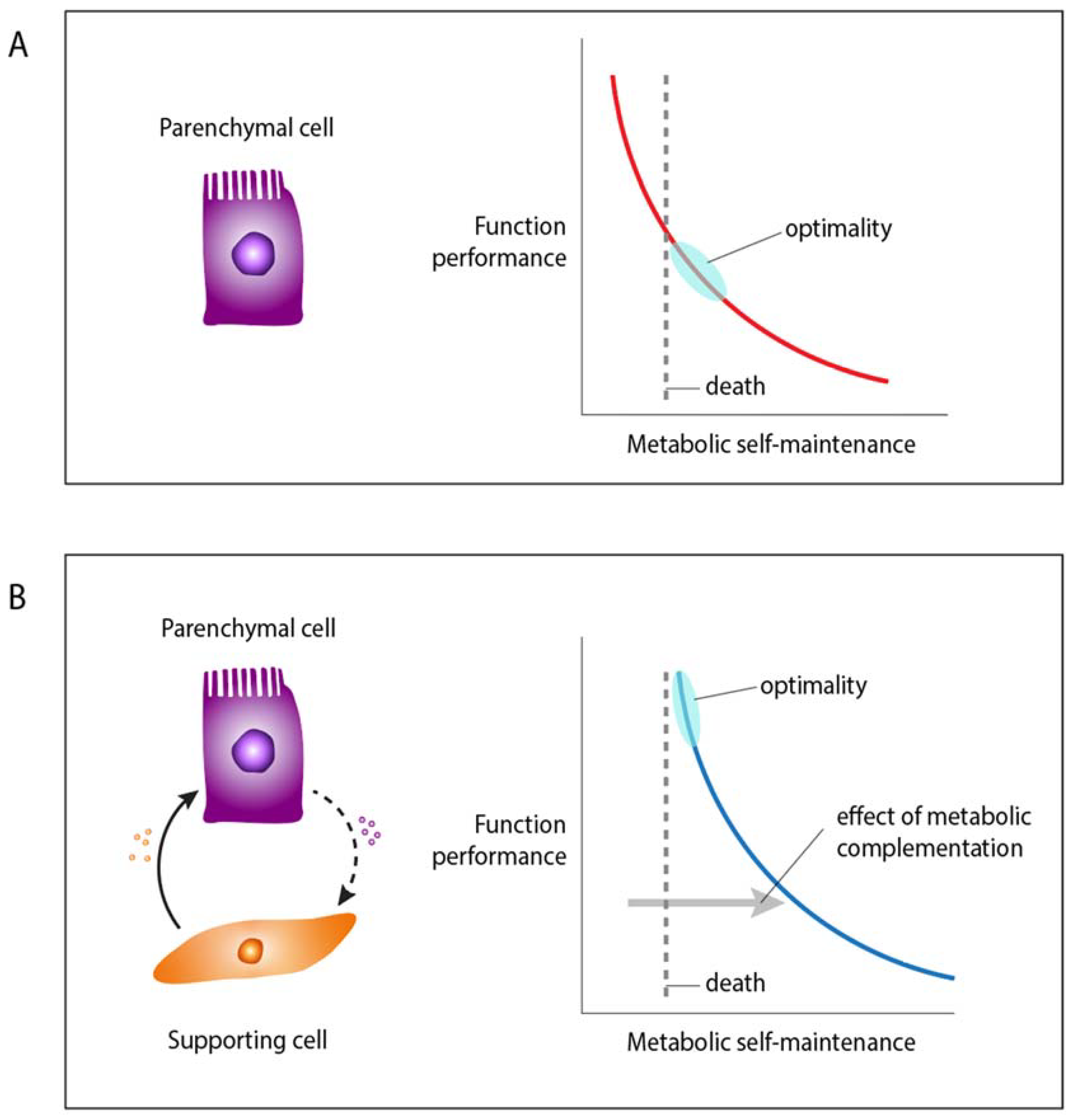
Because of metabolic constraints, the higher energy consumption of a parenchymal cell type will reduce resources available for its own maintenance. A compelling case of a metabolic trade-off between adaptive function and cell maintenance pertains to the generation of ROS. Metabolically high-performing cells require a large amount of energy. Since the oxidative processes of ATP production generate a high load of ROS, this increases the cost of maintenance. Augmentation of a focal cellular function is likely to decrease the ability to invest in the cell’s maintenance, while the need for repair increases, due to the damaging effects of ROS on DNA, lipids, and proteins. We propose that it is precisely these energy and redox trade-offs that have the potential to drive the origin of tissue-level organization: augmentation of function for parenchymal cells may be enabled by the recruitment of supporting cells [33]. Supporting cells are often found to provide substrates for ATP generation (e.g., lactate), antioxidants (e.g., cystine), or other critical resources (e.g., glutamate in the case of neurons), complementing the metabolism of the parenchyma and allowing an increased performance of their adaptive functions.
The resulting structure (i.e., tissue or organ) consists of specialized high-performance parenchymal cells that are dependent on supportive cells for their maintenance. Thus supporting cells also contribute indirectly to the function of the parenchymal cells. Once parenchymal cells rely on supporting cells for maintenance (and resources), the autonomy and primary function of focal cells is transferred to a new, supracellular unit. We propose the term “supra-functionalization” for this process (supra = ‘above or beyond’) to distinguish it from the more familiar notion of sub-functionalization [34].
The spatial organization of primary and supportive cells may initially be transient and facultative, occurring in response to transient functional needs, eventually becoming obligatory over evolutionary time [35]. Facultative spatial confinement may become permanent through the origin of a distinct “identity mechanism” for a new (tissue) level of organization [25]. Identity mechanisms are expected to regulate the abundance and spatial distribution of parenchymal and supporting cell types. They become stabilized and entrenched through increased interdependencies within the tissue. This specialization results in an augmentation of the focal function, achieved by tissue-level organization. Once interactions between the parts are entrenched, it is difficult to lose tissue-level identity without catastrophic failure of all tissue functions, giving rise to an evolutionary “ratchet” mechanism (see below, Section 4).
3.1. Examples of Tissue-Sustaining Metabolic Interactions
The importance of metabolic exchange between cells for tissue redox homeostasis has long been recognized. For example, Hermann [36] summarized observations on brain choroid plexus and eye ciliary body, both with their main function as aqueous humor production. In each case, the structures consist of humor-producing epithelium (parenchyme) and underlying stroma. It was found that epithelia show strongly positive oxidation-reduction potential and that stroma show strongly negative oxidation-reduction potential. This implies a highly oxidative ATP-generating metabolism of the parenchyme. Experimental work further suggested that lactate from the stroma is transported to the parenchyme, likely supplementing epithelial ATP production [36]. Similar differences between oxygen-dependent parenchyme and hypoxia-insensitive non-parenchymal cells were later described in many other tissues (e.g., kidney, liver; [37,38].
Recent research on metabolic exchange between specific cells provides more detailed evidence for the strong metabolic dependency of parenchymal cells on local support cells. We focus here on tissues with costly parenchymal function, brain- and exocrine pancreatic tissue. In particular the nervous system offers a paramount example, suggesting that other organs, such as liver, lung, and kidney, may also follow this pattern.
In the nervous system, various glial cells play supportive roles, providing nutritional and homeostatic support for neurons. Astrocytes of the vertebrate central nervous system (CNS) are a paradigmatic example of a support cell. Neuronal activity is costly. Astrocytes supply lactate to the extracellular space for the neurons to use in oxidation to yield ATP. Neurons are highly dependent on aerobic metabolism and this supplementation from astrocytes; they die very quickly without oxygen [39,40]. Astrocytes store glycogen to overcome periods of low glucose, uptake glutamate from synaptic space to limit its transmitter activity and extrude glutamine into extracellular space to be taken up and converted to glutamate by neurons. Glutamine is synthesized via an intermediate in the citric acid cycle (α-ketoglutarate). Finally, because neuronal activity is costly, it increases mitochondrial ROS production in neurons. Coupled to the activity of neurons, astrocytes release precursors of glutathione (GSH), a major antioxidant effector, which neurons use for GSH biosynthesis, strengthening their protection against ROS [41,42,43]. Together with endothelial cells providing oxygen and glucose, neurons and glia form what is sometimes referred to as metabolic unit [e.g., 40].
Another supportive glial cell type is the Schwann cell in the peripheral nervous system. It is mostly known for producing myelin sheets that enhance axonal conduction speed. However, glial supportive functions to neurons are evolutionarily ancient, pre-dating their specialized role in myelin production [44,45]. The abundance and diversity of glial cells increased with the complexity of the nervous system over evolutionary time. Although glia represent only 10–15% of cells in the Drosophila nervous system, they comprise 50% in mice and up to 90% of the cells in the human CNS [46]. More important than the changes in the abundance of supportive cells is the evolution of their metabolic contribution.
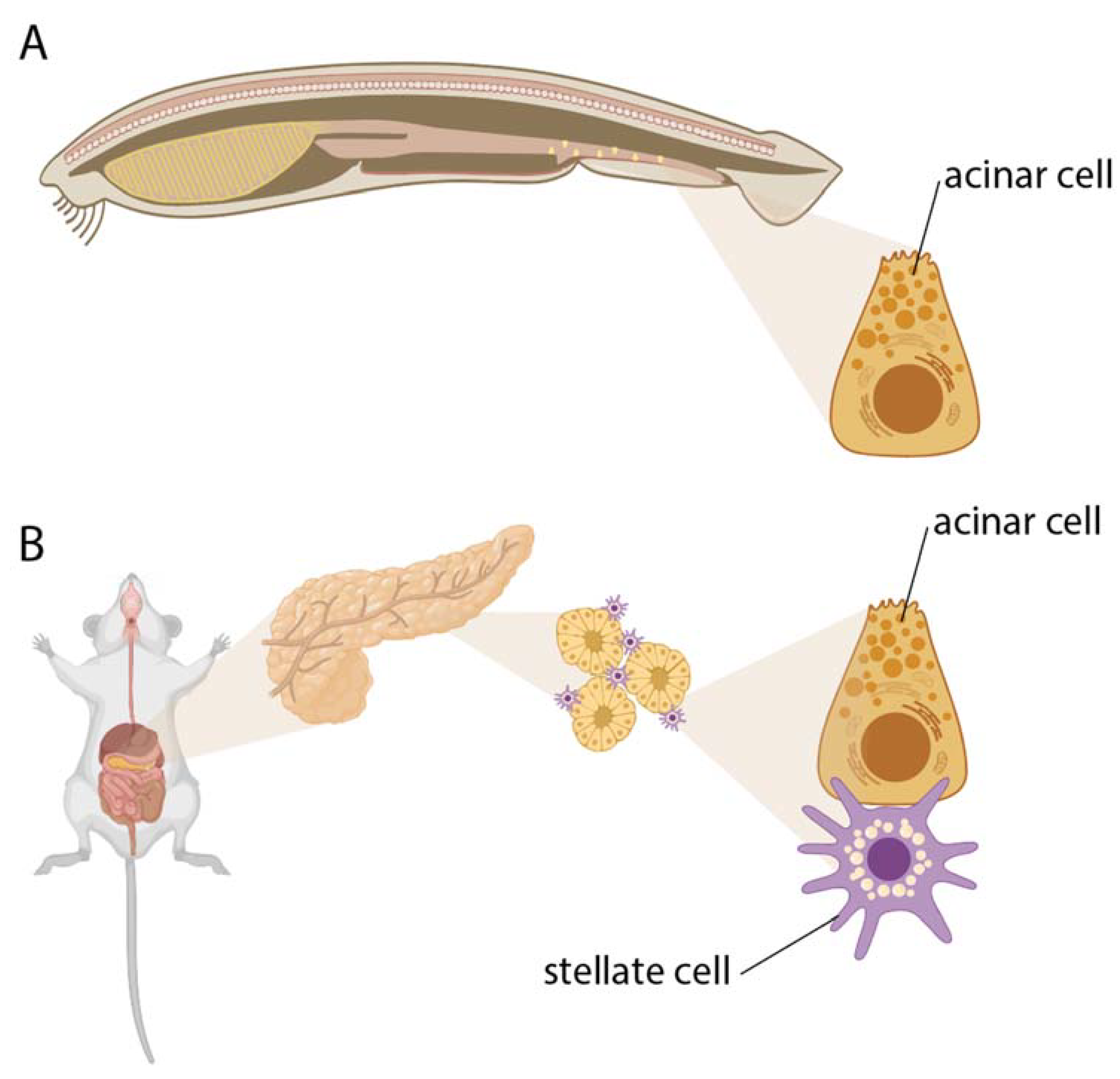
The parenchymal cells of exocrine pancreas are acinar cells, which are responsible for the production and secretion of digestive enzymes into the pancreatic ductal system and eventually the gut (Figure 2). Enzyme synthesis and secretion are energy intensive and require high glucose and amino acid uptake and a high rate of oxidative phosphorylation [47,48,49,50]. Correspondingly, culturing acinar cells requires special culture conditions with ECM components, high amino acid supplementation, and high oxygenation [51]. Reliance on mitochondrial oxidative processes is also reflected in high vulnerability of acinar cells to excessive stimulation. Normal stimulation triggers oscillatory intracellular Ca2+ signaling, synchronously mediating exocytosis and mitochondrial ATP production. Sustained stimulation, in contrast, causes mitochondrial failure resulting in necrosis [52,53]. Due to a high rate of ROS production during stimulation, acinar cells rely on the cystine/glutamate antiporter system, which imports extracellular antioxidant cystine in exchange for glutamate [54]. Metabolic complementation among healthy pancreatic cells and the sources of acinar amino acid and antioxidant uptake appears to not have been studied closely. A likely candidate for metabolic complementation is the pancreatic stellate cell (PaSC), a tissue-specific form of fibroblasts, similar to those of kidney, liver and lung. PaSCs are localized in close proximity to the receptor- and transporter-rich basal pole of acinar cells. The known contributions of healthy (“quiescent”) PaSCs to tissue homeostasis and acinar function are the ECM turnover and maintenance of basal membrane [55,56] and regulatory effects on acinar secretion [e.g., 57,58]. The metabolic role of quiescent PaSCs is unknown. However, it is well appreciated that activated PaSCs metabolically support pancreatic ductal cancer cells [59,60,61]. It remains to be determined whether a degree of metabolic support function is also performed by quiescent PaSCs towards healthy acinar cells.
As mentioned, specific stellate cells are present also in the liver, lung, and kidney. The contribution of hepatic stellate cells to the liver reaches back to the hepatic caecum of Amphioxus, a homolog of vertebrate liver [62]. Our proposed model suggests that their metabolic interaction with hepatocytes eventually became indispensable for the hepatocytes’ function (possibly already in Amphioxus).
The requirement for supportive cells also potentially explains the origin of small composite glands in vertebrates (e.g., thyroid, parathyroid), which consist of diverse types of endocrine cells. In some vertebrates, a main gland like the thyroid hosts nests of cells of different embryonic origin and different functions. In the case of the thyroid, one finds so-called parafollicular or C-cells between the thyroid follicles. These are endocrine cells that produce calcitonin. They originate from the fourth and fifth pharyngeal pouch of the embryo and migrate into the thyroid tissue in mammals, while the thyroid originates from the floor of the embryonic pharynx. In contrast, the C-cells of adult teleosts still reside near the posterior gill slits and not the thyroid. As metabolic requirements are generic across cells (i.e., energy, ROS, amino acids), these may have aggregated to share the supportive cells.
3.2. Why Was Metabolism Overlooked or Neglected in Evolutionary Biology?
Although well-appreciated in microbial ecology, as well as in general and pathological physiology, cell-level metabolism and cellular metabolic exchange have not featured prominently in developmental and evolutionary biology (but see [63]). This may be due to substantial metabolic plasticity plus the ancient origin and proverbial conservation of the central biochemical processes in multicellular organisms. However, the overall stability does not exclude changes in compartmentalization among the cells.
Accordingly, ideas about cell communication have also largely focused on the exchange of information—messages that inform the cell about its environment and elicit a reaction, such as change of movement, gene expression, or proliferation (but see [36,64]. This implicitly assumes a degree of autonomy of development and evolution from cellular metabolism. In contrast, our model emphasizes the exchange of metabolic compounds among animal cells. Even in the same organism, some cell types are “auxotroph” with respect to certain amino acids (e.g., they cannot synthesize a particular amino acid) and others “autotroph” with respect to a range of amino-acids, even though the genome includes genes for the enzymes necessary to synthesize these amino acids. Immune cells, for example, are auxotroph for a wide range of amino acids [e.g., 65]. Metabolic exchange is becoming recognized as a critical player in immune cell function [e.g., 66,67]. Beyond information, therefore, cells also exchange essential substances that allow—and thus can also limit—their function and survival. The need for the exchange of metabolic products may drive, in part, the evolutionary origin of higher-level organismal structures such as tissues and organs.
It is noteworthy that our model does not claim generality for the evolution of structural organization at all levels; it does not imply that all higher-level structural organization (e.g., the origin of organ systems) will be driven by the same principles [68]. Empirical work will decide whether aspects of this model also apply to other kinds of organ systems and at other levels of organization.
3.3. Implications of the Model
Our model precipitates expectations about the kind of interactions among cells in a tissue and organ that we should observe empirically. The most direct comparison that bears on why cells aggregate into tissues is between organisms where the relevant primary cell types have undergone the tissue transition (i.e., become tissue parenchyme) with organisms where homologous primary cell types are not integrated into a tissue. Specifically, we would expect detectable differences in the metabolic interactions of these cells. The model implies that parenchymal cells in tissue will: (1) have more metabolic exchange with surrounding cells; (2) be more existentially dependent on surrounding cells for self-maintenance, and thus more prone to death upon removal from the tissue; and (3) have detectable functional augmentation.
Although access to species that predate the tissue formation may be limited for many tissues, any assessment of the degree and kind of metabolic complementation among the cells of tissues will be important to test and further develop this model. The most abundant tissue or organ-wide cellular data is available at the transcriptomic level. Unlike in the study of microbial communities (e.g., [69]), the analysis of transcriptomic data in the study of multicellular organismal biology is seldom focused on the expression of metabolic pathway genes (but see [70,71,72]). Examples to focus on include genes for metabolic enzymes or signatures of the metabolic exchange, such as genes for peptide or amino acid transporters. This is perhaps the most immediate source of information that can illuminate cellular metabolic homeostasis in the context of tissue organization and its evolution, apart from the direct study of the metabolome.
4. Discussion: On the Origins of Within-Organism Levels of Organization
Our proposed model describes how an evolutionary shift of organizational level from cell type to tissue can occur without a qualitative change in functional role. Typically, the evolutionary emergence of a new level of organization is conceptualized within the “major transitions in individuality” framework [9,13,15]. However, the tissue transition within a multicellular organism requires a different approach.
“Evolutionary transitions in individuality” traditionally refer to the emergence of new units of selection or whole-organism levels, such as the aggregation of cells into a multicellular system that can only reproduce as a whole. A critical problem in understanding these transitions is to explain how these aggregates ensure cooperation and suppress “cheating” among component cells, such as through control of proliferation and germline sequestration [9,11,13,15]. By contrast, the cell-to-tissue transition creates a new body part rather than a new organism level or new unit of selection [see 19,20,73]. Instead of cooperation and conflict, the focal problem becomes understanding whether and how the tissue comes under modular genetic control, allowing it to vary and adapt quasi-independently of other body parts [25,74,75,76]. For example, if the supporting cells are generic to multiple tissues, this creates pleiotropic links. Such a case has been shown recently, where selection of skin fibroblasts is associated with gene expression changes in endometrial fibroblasts [77]. These links may need to be suppressed via supporting cell specialization or via tissue-specific compensation [78]. Such tissue-specific cell types belonging to the generic category of fibroblasts or macrophages are well known. Modularity at the tissue level, however, is not yet well understood.
The proposed model also differs from and complements existing ideas about complexity increase in evolution. Once the proposed metabolic interdependencies between primary and supporting cells have evolved, they will be difficult to reverse without dramatic loss of function. This may operate as a complexity-increasing “ratchet” mechanism—i.e., a process that only permits motion in one direction [79,80,81]. The interdependencies within a ratchet mechanism can be depicted as analogous to the subfunctionalization or neofunctionalization of paralogs following the duplication of genes [34], except with the duplicated parts being cells of similar type [20,21,82]. Although this may be valid for some cases of tissue and organ evolution, the tissue transition we propose requires a different conceptualization.
Because the parenchymal cell does not partition its functions with another, similar cell, the tissue transition is not a form of subfunctionalization and thus unlikely to be amenable to subfunctionalization models. Likewise, the transition does not involve the creation of a novel function (neofunctionalization). Instead, we suggest a more apt term for the transition from a single cell type to an integrated tissue performing the same function would be “supra-functionalization.”
At a very abstract level, supra-functionalization involves a division of (metabolic) labor between primary and supporting cells, but it also differs from existing division of labor theories in several key respects. Division of labor models picture an initially homogeneous set of components, each of which must perform the same set of adaptive tasks. The tasks cannot all be optimized simultaneously, giving rise to trade-offs. Trade-offs can be circumvented through partitioning tasks among components (i.e., a division of labor), thereby allowing the tasks to be performed more efficiently. In the proposed model, by contrast, primary and supportive cells already perform distinct tasks from the start. The central trade-off within primary cells is a metabolic trade-off between performance of adaptive function and self-maintenance. Crucially, in line with prevailing assumptions (see Section 3.2), division of labor models tend to represent components as persistent “atomic” units, and do not consider their self-maintenance as something that can be changed, constrained, and redistributed. All cells are dynamic, far-from-equilibrium systems that have to be maintained through metabolism against the physical tendency toward entropic degradation. Although this universal cellular condition gives rise to constraints on cellular adaptation, it also provides opportunities for overcoming these constraints through metabolic complementation via integration into tissues.
Author Contributions
MP drafted the core idea, all authors intensively discussed and developed a draft. Specifically, MP and GPW drafted the biological part, whereas JD and ACL developed its implications in the light of past and current discussions of evolution of biological complexity and organization. All authors contributed to the writing of the final manuscript.
Funding
The research for this paper was funded by the Austrian Science Foundation FWF grant #P33540 (to MP); GPW receives funding for single cell research from the Hagler Institute for Advanced Study at Texas A&M University. MP also acknowledges funding for the workshop, Evolvability: a unified concept in evolutionary biology, funded 2019–2020 by the Center of Advanced Science of the Norwegian Academy of Science and Letters, Oslo, where some of the ideas in this paper started to take shape. JD acknowledges funding from the Francis Crick Institute, which receives its core funding from Cancer Research UK, the UK Medical Research Council, and the Wellcome Trust, and thanks the Theoretical Biology Group at the Crick for helpful comments. ACL acknowledges funding from the John Templeton Foundation (Grant #62220). The opinions expressed in this paper are those of the authors and not those of the John Templeton Foundation.
Data Accessibility
The article presents no new data.
Competing Interests
Authors declare no competing interests
Ethics statement
The research involved no animal or cell experimentation, nor human trial.
References
- Mayr, E. The Emergence of Evolutionary Novelties.; Chicago University Press: Chicago, 1960. [Google Scholar]
- Chavan, A.R.; Griffith, O.W.; Stadtmauer, D.; Maziarz, J.; et al. Evolution of embryo implantation was enabled by the origin of decidual stromal cells in eutherian mammals. Mol Biol Evol. 2020. [Google Scholar] [CrossRef]
- DeMIGUEL, D.; Azanza, B.; Morales, J. Key innovations in ruminant evolution: a paleontological perspective. Integr Zool 2014, 9, 412–433. [Google Scholar] [CrossRef] [PubMed]
- Ronco, F.; Salzburger, W. Tracing evolutionary decoupling of oral and pharyngeal jaws in cichlid fishes. Evol Lett 2021, 5, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Leake, L.D. Comparative Histology. An introduction to the Microscopic Structure of Animals.; Academic Press: New York, 1975. [Google Scholar]
- Falkmer, S. Comparative morphology of pancreatic islets in animals. In The diabetic Pancreas; Volk, B.W., Arquille, E.R., Eds.; Plenum medical Book coMPANY: New York, 1985. [Google Scholar]
- Falkmer, S.; Dafgård, E.; el-Salhy, M.; Engström, W.; Grimelius, L.; Zetterberg, A. Phylogenetical aspects on islet hormone families: a minireview with particular reference to insulin as a growth factor and to the phylogeny of PYY and NPY immunoreactive cells and nerves in the endocrine and exocrine pancreas. Peptides 1985, 6 (Suppl. S3), 315–320. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, G. Origin of Sensory and Neurosecretory Cell Types: Vertebrate Cranial Placodes; CRC Press: 2021.
- Maynard Smith, J.; Szathmary, E. The Major Transitions in Evolution; Oxford University Press: Oxford, 1995. [Google Scholar]
- McShea, D.W. PERSPECTIVE METAZOAN COMPLEXITY AND EVOLUTION: IS THERE A TREND? Evolution 1996, 50, 477–492. [Google Scholar] [CrossRef]
- Michod, R. Darwinian Dynamics: evolutionary transitions in fitness and individuality.; Princeton University Press: Princeton, NJ, 2000. [Google Scholar]
- Michod, R.E. Evolution of individuality during the transition from unicellular to multicellular life. Proc Natl Acad Sci U S A 2007, 104 (Suppl. S1), 8613–8618. [Google Scholar] [CrossRef]
- Okasha S. 2006. Evolution and the levels of selection Oxford.
- Oxford; New York: Clarendon Press.
- Oxford University Press.
- West, S.A.; Fisher, R.M.; Gardner, A.; Kiers, E.T. Major evolutionary transitions in individuality. Proc Natl Acad Sci U S A 2015, 112, 10112–10119. [Google Scholar] [CrossRef]
- Buss, L.W. The Evolution of Individuality; Princeton University Press: 1987.
- Brunet, T.; King, N. The Origin of Animal Multicellularity and Cell Differentiation. Dev Cell 2017, 43, 124–140. [Google Scholar] [CrossRef]
- Herron, M.; Conlin, P.; Ratcliff, W. The Evolution of Multicellularity. 2021, 1.
- Niklas, K.J.; Newman, S. Multicellularity : origins and evolution; The MIT Press: Cambridge, MA, 2016. [Google Scholar]
- Wagner, G.P. Homology, Genes and Evolutionary Innovation; Princeton University Press: Princeton, 2014. [Google Scholar]
- Nejad Kourki, A. The evolution of complex multicellularity in animals. Biology & Philosophy 2022, 37, 43. [Google Scholar] [CrossRef]
- Adler, M.; Moriel, N.; Goeva, A.; Avraham-Davidi, I.; Mages, S.; Adams, T.S.; Kaminski, N.; Macosko, E.Z.; Regev, A.; Medzhitov, R.; et al. Emergence of division of labor in tissues through cell interactions and spatial cues. Cell Rep 2023, 42, 112412. [Google Scholar] [CrossRef]
- Adler, M.; Chavan, A.R.; Medzhitov, R. Tissue Biology: In Search of a New Paradigm. Annu Rev Cell Dev Biol 2023. [Google Scholar] [CrossRef]
- Zhou, X.; Franklin, R.A.; Adler, M.; Jacox, J.B.; Bailis, W.; Shyer, J.A.; Flavell, R.A.; Mayo, A.; Alon, U.; Medzhitov, R. Circuit Design Features of a Stable Two-Cell System. Cell 2018, 172, 744–757. [Google Scholar] [CrossRef]
- Hall, B.K. The embryonic development of bone. American Scientist 1988, 76, 174–181. [Google Scholar]
- DiFrisco, J.; Love, A.; Wagner, G.P. Character identity mechanisms: a conceptual model.
- for comparative-mechanistic biology. Biology & Philosophy 2020, 1-32.
- Weiße, A.Y.; Oyarzún, D.A.; Danos, V.; Swain, P.S. Mechanistic links between cellular trade-offs, gene expression, and growth. Proc Natl Acad Sci U S A 2015, 112, E1038–E1047. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, D.; van Berlo, R.; de Ridder, D.; Teusink, B. Shifts in growth strategies reflect tradeoffs in cellular economics. Mol Syst Biol 2009, 5, 323. [Google Scholar] [CrossRef]
- Kempes, C.P.; Dutkiewicz, S.; Follows, M.J. Growth, metabolic partitioning, and the size of microorganisms. Proc Natl Acad Sci U S A 2012, 109, 495–500. [Google Scholar] [CrossRef]
- Gladden, L.B. Lactate metabolism: a new paradigm for the third millennium. J Physiol 2004, 558, 5–30. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Cell-cell and intracellular lactate shuttles. J Physiol 2009, 587, 5591–5600. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Regulation of glutathione synthesis. Mol Aspects Med 2009, 30, 42–59. [Google Scholar] [CrossRef]
- DeLong, J.P.; Okie, J.G.; Moses, M.E.; Sibly, R.M.; Brown, J.H. Shifts in metabolic scaling, production, and efficiency across major evolutionary transitions of life. Proc Natl Acad Sci U S A 2010, 107, 12941–12945. [Google Scholar] [CrossRef]
- Pavlicev, M.; Wagner, G.P. Reading the palimpsest of cell interactions: What questions may we ask of the data? iScience 2024, 27, 109670. [Google Scholar] [CrossRef] [PubMed]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.L.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef] [PubMed]
- Love, A.C.; Wagner, G.P. Co-option of stress mechanisms in the origin of evolutionary novelties. Evolution 2022, 76, 394–413. [Google Scholar] [CrossRef] [PubMed]
- Hermann, H. Direct metabolic interactions between animal cells. Their role in tissue function and development is reconsidered. Science 1960, 132, 529–532. [Google Scholar] [CrossRef]
- Gasbarrini, A.; Borle, A.B.; Farghali, H.; Bender, C.; Francavilla, A.; Van Thiel, D. Effect of anoxia on intracellular ATP, Na+i, Ca2+i, Mg2+i, and cytotoxicity in rat hepatocytes. J Biol Chem 1992, 267, 6654–6663. [Google Scholar] [CrossRef]
- Brezis, M.; Epstein, F.H. Cellular mechanisms of acute ischemic injury in the kidney. Annu Rev Med 1993, 44, 27–37. [Google Scholar] [CrossRef]
- Gibbons, H.M.; Dragunow, M. Adult human brain cell culture for neuroscience research. Int J Biochem Cell Biol 2010, 42, 844–856. [Google Scholar] [CrossRef]
- Turner, D.A.; Adamson, D.C. Neuronal-astrocyte metabolic interactions: understanding the transition into abnormal astrocytoma metabolism. J Neuropathol Exp Neurol 2011, 70, 167–176. [Google Scholar] [CrossRef]
- Almeida, A.; Jimenez-Blasco, D.; Bolaños, J.P. Cross-talk between energy and redox metabolism in astrocyte-neuron functional cooperation. Essays Biochem 2023, 67, 17–26. [Google Scholar] [CrossRef]
- Bolaños, J.P. Bioenergetics and redox adaptations of astrocytes to neuronal activity. J Neurochem 2016, 139 (Suppl. S2), 115–125. [Google Scholar] [CrossRef]
- Bonvento, G.; Bolaños, J.P. Astrocyte-neuron metabolic cooperation shapes brain activity. Cell Metab 2021, 33, 1546–1564. [Google Scholar] [CrossRef]
- Kastriti, M.E.; Adameyko, I. Specification, plasticity and evolutionary origin of peripheral glial cells. Curr Opin Neurobiol 2017, 47, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Rey, S.; Zalc, B.; Klämbt, C. Evolution of glial wrapping: A new hypothesis. Dev Neurobiol 2021, 81, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Song, B.; Lee, I.S. Glia: Models for Human Neurodevelopmental and Neurodegenerative Disorders. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Van Venrooij, W.J.; Poort, C.; Kramer, M.F.; Jansen, M.T. Relationship between extracellular amino acids and protein synthesis in vitro in the rat pancreas. Eur J Biochem 1972, 30, 427–433. [Google Scholar] [CrossRef]
- Mann, G.E.; Peran, S. Basolateral amino acid transport systems in the perfused exocrine pancreas: sodium-dependency and kinetic interactions between influx and efflux mechanisms. Biochim Biophys Acta 1986, 858, 263–274. [Google Scholar] [CrossRef]
- Kosowski, H.; Schild, L.; Kunz, D.; Halangk, W. Energy metabolism in rat pancreatic acinar cells during anoxia and reoxygenation. Biochimica et Biophysica Acta 1989, 118–126. [Google Scholar] [CrossRef]
- Masamune, A.; Shimosegawa, T. Pancreatic stellate cells--multi-functional cells in the pancreas. Pancreatology 2013, 13, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Bläuer, M.; Nordback, I.; Sand, J.; Laukkarinen, J. A novel explant outgrowth culture model for mouse pancreatic acinar cells with long-term maintenance of secretory phenotype. Eur J Cell Biol 2011, 90, 1052–1060. [Google Scholar] [CrossRef]
- Petersen, O.H. The 2022 George E Palade Medal Lecture: Toxic Ca. Pancreatology 2023, 23, 1–8. [Google Scholar] [CrossRef]
- Criddle, D.N.; Tepkin, A.V. Bioenergetics of the Exocrine Pancreas: Physiology to Pathophysiology. In Pancreapedia: Exocrine Pancreas knowledge base, 2020.
- Pan, Z.; Van den Bossche, J.L.; Rodriguez-Aznar, E.; Janssen, P.; Lara, O.; Ates, G.; Massie, A.; De Paep, D.L.; Houbracken, I.; Mambretti, M.; et al. Pancreatic acinar cell fate relies on system x. Cell Death Dis 2023, 14, 536. [Google Scholar] [CrossRef]
- Phillips, P.A.; McCarroll, J.A.; Park, S.; Wu, M.J.; Pirola, R.; Korsten, M.; Wilson, J.S.; Apte, M.V. Rat pancreatic stellate cells secrete matrix metalloproteinases: implications for extracellular matrix turnover. Gut 2003, 52, 275–282. [Google Scholar] [CrossRef]
- Riopel, M.M.; Li, J.; Liu, S.; Leask, A.; Wang, R. β1 integrin-extracellular matrix interactions are essential for maintaining exocrine pancreas architecture and function. Lab Invest 2013, 93, 31–40. [Google Scholar] [CrossRef]
- Phillips, P.A.; Yang, L.; Shulkes, A.; Vonlaufen, A.; Poljak, A.; Bustamante, S.; Warren, A.; Xu, Z.; Guilhaus, M.; Pirola, R.; et al. Pancreatic stellate cells produce acetylcholine and may play a role in pancreatic exocrine secretion. Proc Natl Acad Sci U S A 2010, 107, 17397–17402. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.S.; Cui, Z.J. Pancreatic Stellate Cells Serve as a Brake Mechanism on Pancreatic Acinar Cell Calcium Signaling Modulated by Methionine Sulfoxide Reductase Expression. Cells 2019, 8. [Google Scholar] [CrossRef]
- Sousa, C.M.; Biancur, D.E.; Wang, X.; Halbrook, C.J.; Sherman, M.H.; Zhang, L.; Kremer, D.; Hwang, R.F.; Witkiewicz, A.K.; Ying, H.; et al. Pancreatic stellate cells support tumour metabolism through autophagic alanine secretion. Nature 2016, 536, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Vaziri-Gohar, A.; Zarei, M.; Brody, J.R.; Winter, J.M. Metabolic Dependencies in Pancreatic Cancer. Front Oncol 2018, 8, 617. [Google Scholar] [CrossRef]
- Hamada, S.; Matsumoto, R.; Masamune, A. Pancreatic Stellate Cells and Metabolic Alteration: Physiology and Pathophysiology. Front Physiol 2022, 13, 865105. [Google Scholar] [CrossRef]
- Wu, B.; Xu, W.; Wu, K.; Li, Y.; Hu, M.; Feng, C.; Zhu, C.; Zheng, J.; Cui, X.; Li, J.; et al. Single-cell analysis of the amphioxus hepatic caecum and vertebrate liver reveals genetic mechanisms of vertebrate liver evolution. Nat Ecol Evol 2024. [Google Scholar] [CrossRef] [PubMed]
- Krejci, A.; Tennessen, J.M. Metabolism in time and space - exploring the frontier of developmental biology. Development 2017, 144, 3193–3198. [Google Scholar] [CrossRef]
- Juliano, C.E. Metabolites play an underappreciated role in development. Proc Natl Acad Sci U S A 2024, 121, e2410810121. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Amino acid auxotrophy as a system of immunological control nodes. Nat Immunol 2016, 17, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Ganeshan, K.; Chawla, A. Metabolic regulation of immune responses. Annu Rev Immunol 2014, 32, 609–634. [Google Scholar] [CrossRef]
- Chi, H. Immunometabolism at the intersection of metabolic signaling, cell fate, and systems immunology. Cell Mol Immunol 2022, 19, 299–302. [Google Scholar] [CrossRef]
- Schmidt-Rhaesa, A. The evolution of organ systems; Oxford University Press: Oxford ; New York, 2007.
- Jiang, Y.; Xiong, X.; Danska, J.; Parkinson, J. Metatranscriptomic analysis of diverse microbial communities reveals core metabolic pathways and microbiome-specific functionality. Microbiome 2016, 4, 2. [Google Scholar] [CrossRef]
- Damiani, C.; Maspero, D.; Di Filippo, M.; Colombo, R.; Pescini, D.; Graudenzi, A.; Westerhoff, H.V.; Alberghina, L.; Vanoni, M.; Mauri, G. Integration of single-cell RNA-seq data into population models to characterize cancer metabolism. PLoS Comput Biol 2019, 15, e1006733. [Google Scholar] [CrossRef]
- Alghamdi, N.; Chang, W.; Dang, P.; Lu, X.; Wan, C.; Gampala, S.; Huang, Z.; Wang, J.; Ma, Q.; Zang, Y.; et al. A graph neural network model to estimate cell-wise metabolic flux using single-cell RNA-seq data. Genome Res 2021, 31, 1867–1884. [Google Scholar] [CrossRef]
- Jia, Z.; Wei, Y.; Zhang, Y.; Song, K.; Yuan, J. Metabolic reprogramming and heterogeneity during the decidualization process of endometrial stromal cells. Cell Commun Signal 2024, 22, 385. [Google Scholar] [CrossRef]
- McShea, D.; Venit, E.P. What is a part? In The Character Concept in Evolutionary Biology., Wagner, G.P., Ed. Academic Press: San Diego, 2001.
- Wagner, G.P.; Altenberg, L. Perspective: Complex Adaptations and the Evolution of Evolvability. Evolution 1996, 50, 967–976. [Google Scholar] [CrossRef]
- Raff, R.A. The shape of life : genes, development, and the evolution of animal form; University of Chicago Press: Chicago, 1996. [Google Scholar]
- Wagner, G.P.; Pavlicev, M.; Cheverud, J.M. The road to modularity. Nat Rev Genet 2007, 8, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Dighe, A.; Maziarz, J.; Ibrahim-Hashim, A.; Gatenby, R.A.; Kshitiz; Levchenko, A.; Wagner, G.P. Experimental and phylogenetic evidence for correlated gene expression evolution in endometrial and skin fibroblasts. iScience 2024, 27, 108593. [CrossRef]
- Pavlicev, M.; Wagner, G.P. A model of developmental evolution: selection, pleiotropy and compensation. Trends Ecol Evol 2012, 27, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Cairns-Smith, A. The Complexity ratchet. In Progress in Search for Extraterrestrial Life., Shostak, G., Ed. Astronomical Society of the Pacific.: San Francisco., 1995; pp. 31–36.
- Liard, V.; Parsons, D.P.; Rouzaud-Cornabas, J.; Beslon, G. The Complexity Ratchet: Stronger than Selection, Stronger than Evolvability, Weaker than Robustness. Artif Life 2020, 26, 38–57. [Google Scholar] [CrossRef]
- Lukeš, J.; Archibald, J.M.; Keeling, P.J.; Doolittle, W.F.; Gray, M.W. How a neutral evolutionary ratchet can build cellular complexity. IUBMB Life 2011, 63, 528–537. [Google Scholar] [CrossRef]
- Parker, J. Organ Evolution: Emergence of Multicellular Function. Annu Rev Cell Dev Biol 2024. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
The evolution of the exocrine pancreas illustrates the proposed transition from individual cells to tissues and organs. A. In early chordates, such as Amphioxus, acinar cells are individually dispersed throughout the intestine. B. Eventually, acinar cells were integrated into a tissue or organ structure, where they are dependent on metabolic complementation from supporting stellate cells. Figure drawn with BioRender.com.
Figure 2.
The evolution of the exocrine pancreas illustrates the proposed transition from individual cells to tissues and organs. A. In early chordates, such as Amphioxus, acinar cells are individually dispersed throughout the intestine. B. Eventually, acinar cells were integrated into a tissue or organ structure, where they are dependent on metabolic complementation from supporting stellate cells. Figure drawn with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



