
Submitted:
25 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Identification of the individuals having impaired kidney function is essential in preventing the complications of this disease. We measured 1 009 metabolites at the baseline study in 10 159 Finnish men of the METSIM cohort and associated the metabolites with estimated glomerular filtration rate (eGFR). A total of 7 090 men participated in the 12-year follow-up study. Non-targeted metabolomics profiling was performed at Metabolon, Inc. (Morrisville, NC, USA) on EDTA plasma samples obtained after overnight fasting. We applied Liquid chromatography-mass spectrometry (LC-MS/MS) to identify the metabolites (the Metabolon DiscoveryHD4 platform). We performed association analyses between the eGFR and metabolites using linear regression adjusted for confounding factors. We found 108 metabolites significantly associated with a decrease in eGFR, and 28 of them were novel including 12 amino acids, 8 xenobiotics, 5 lipids, 1 nucleotide, 1 peptide, and 1 partially characterized molecule. The most significant associations were with five amino acids, N-acetylmethionine, N-acetylvaline, gamma-carboxyglutamate, 3-methylglutaryl-carnitine, and pro-line. We identified 28 novel metabolites associated with decreased eGFR in the 12-year follow-up study of the METSIM cohort.
Keywords:
Genetics
; metabolomics
; type 2 diabetes
; glomerular filtration rate
1. Introduction
Chronic kidney disease (CKD) affects approximately 10% of the Western countries’ population [1]. Glomerular filtration rate (GFR) is accepted as the best marker of impaired kidney function, calculated as an estimated GFR (eGFR) [2]. Diabetes is a major risk factor for impaired kidney function [3] but also age, sex, hypertension, obesity, increased total triglycerides, and smoking are risk factors for CKD [4]. During the last few years, genome-wide association studies have identified hundreds of genetic variants for kidney diseases [5,6,7]. Interestingly, a recent study identified genetic variants in the individuals with and without diabetes and reported that a majority of eGFR loci were similar in individuals with and without diabetes [8].
The first studies aiming to identify metabolites associated with eGFR had a small size and included only a low number of metabolites [9,10,11,12,13]. Grams et al [14] included 587 participants in their study, and identified five metabolites (16-hydroxypalmitate, kynurenate, homovanillate sulphate, N2,N2-dimethylguanosine, hippurate) associated with CKD. Lin et al. performed a large metabolome-wide association study, including 640 metabolites in 3 906 participants of the Hispanic Community Health Study/Study of Latinos. They identified 404 eGFR-metabolite associations and found 79 novel associations [15], where amino acids and xenobiotics were the most frequent metabolites associated with eGFR. Recent two studies have reported substantially increased number of metabolites associated with CKD [16,17].
Early identification of the individuals having impaired kidney function is essential in the prevention of CKD and its complications. However, previous studies aiming to identify metabolites associated with a decrease in eGFR have been mainly cross-sectional and included relatively small number of participants and metabolites. Our population-based study included 10 159 participants having 1 009 metabolites measured at baseline. A total of 7 090 participated in a 12-year follow-up. Our study is the largest study identifying novel metabolites associated with a decline in eGFR during a long follow-up. Therefore, our study has a good statistical power to reveal new metabolic pathways for impaired kidney function.
2. Results
2.1. Baseline Characteristics
We included in our study 10 159 METSIM participants. Table 1 shows baseline characteristics of the participants according to their glucose tolerance. These groups differed significantly in age, systolic blood pressure, BMI, total triglycerides, fasting glucose, HbA1c, fasting plasma insulin, eGFR, urine albumin, and high-sensitivity C-reactive protein (hs-CPR). The difference between the three groups was statistically significant but small in eGFR (87.9 in the NGT group, 88.6 and 86.1 in the T2D group).
2.2. Metabolites in Participants with Decreased and Normal eGFR
We compared metabolite concentrations between the participants having eGFR < 80 and eGFR ≥ 80 ml/min/1.73 m2, and found statistically significant (p<5.0E-05) differences in 586 metabolites (Table S1). The most significant differences (p <1.1E-350) between the two groups were in 1-methylhistidine, 1-methyl-4-imidazoleacetate, 2,3-dihydroxy-5-methylthio-4-pentenoate (DMTPA), creatinine, hydroxy asparagine, N,N,N-trimethyl-alanylproline betaine, N-acetylalanine, and pseudouridine.
2.3. Effects of Glucose Tolerance on Metabolic Profile
We analysed the associations of eGFR with metabolites in different subgroups of glucose tolerance (n=1 057 in each group matched for age and BMI). Participants with NGT had 379 statistically significant associations with the metabolites, participants with prediabetes had 474 significant associations, and participants with T2D had 378 significant associations. Table S1 presents the 100 most significant metabolites in the participants with T2D, prediabetes and NGT. Independently of the glucose tolerance, all metabolites were associated with a decrease in eGFR. Venn diagram (Figure 1) shows that the participants in the different glucose tolerance groups shared 78% of the 100 most significant metabolite associations, 11 of the metabolites were found only in the NGT group, 7 in the prediabetes group, and 12 in the T2D group.
2.4. Metabolites Associated with a Decrease in eGFR
We performed linear regression to associate 1 009 metabolites with eGFR at baseline without adjustment for confounding factors, adjusted for baseline eGFR (Model 1), and adjusted for baseline eGFR, age, BMI, smoking, fasting glucose, total triglycerides, and systolic blood pressure (Model 2) (Table S2). All metabolites listed in Table S2 had p < 5x10-5 in all models. Adjustment for the baseline eGFR (Model 1) decreased substantially beta and p values. In Model 2 beta and p value further deceased but the decreases were relatively small.
We found 108 metabolites significantly associated with a decrease in eGFR, and 28 of them were novel (Table 2). The 10 most statistically significant metabolites associated with decreased eGFR were six amino acids, creatinine, hydroxyasparagine, N,N,N-trimethyl-alanylproline betaine, N-acetylalanine, N-acetylserine, C-glycosyltryptophan, and N-formylmethionine; a nucleotide pseudouridine; xenobiotics erythritol; and carbohydrate erythronate.
Among the novel 28 metabolites decreasing eGFR, 12 were amino acids, 5 lipids, 1 nucleotide, 1 peptide, 8 xenobiotics, and 1 partially characterized molecule. Among the amino acids the three most significant associations with a decrease in metabolites were for N-acetylmethionine (beta -0.087, p= 5.5E-24), N-acetylvaline (beta -0.082, p = 2.6E-21), and γ-carboxyglutamate (beta -0.065, p= 2.6E-14). Among the lipids the two most significant associations with a decrease in metabolites were for 11beta-hydroxyetiocholanolone (beta -0.050, p = 4.0E-7), and 2-methylmalonylcarnitive C4-DC (beta -0.042, p = 3.1.0E-6), and among the xenobiotics for 2,3-dihydroxyisovalerate (beta -0.048, p = 6.8E-06, and (S)-a-amino-omega-caprolactam (beta -0.050, p = 1.0E-8).
2.5. Genetic Variants Associated with Novel Metabolites
We identified nine genetic variants significantly associated with the novel metabolites (Table 3). The most significant associations were with 5-methyluridine, glycine, proline and N-acetylmethionine. Each of the nine genetic variants was associated with at least three different metabolites suggesting pleiotropy of these genes. Importantly, none of these genetic variants was significantly associated with a decrease in eGFR indicating that the effects of the metabolites on eGFR were not explained by genetic factors.
3. Discussion
We measured 1 009 metabolites with LC-MS/MS in 10188 participants of the METSIM study. Our study reports several novel findings. We found that glucose tolerance did not have a major effect on the metabolite profile at baseline. Among the top 100 metabolites associated with eGFR, 78 were identical in participants with normal glucose tolerance, pre-diabetes, and diabetes (Figure 1). Our results suggest that the metabolic pathways leading to a decrease in eGFR are largely independent of glucose tolerance. This observation agrees with a previous study reporting that the majority of the eGFR loci were similar in the individuals with and without diabetes [8].
We found several statistically significant associations of the metabolites with a decrease in eGFR in the 12-year follow-up of the METSIM cohort. Of the 108 metabolites associated with a decrease in eGFR 28 were novel (Table 2). We also replicated metabolite associations with decreased eGFR reported in previous studies [13,18,19,20,21,22,23,24,25,26,27].
We found three novel associations of N-acetylated amino acids (N-acetylmethionine, N-acetylvaline, and N-acetyltaurine) with a decrease in eGFR. N-acetylated amino acids are uremic toxins [28]. Aminoacylase-1 (ACY1) enzyme converts acetylated amino acids into free amino acids, and therefore the individuals having impaired activity of ACY1 or a mutation in the ACYL1 gene have increased concentrations of acetylated amino acids in blood and urine [29,30,31,32,33,34].
Amino acid γ-carboxyglutamate was significantly associated with a decrease in eGFR. γ-carboxyglutamate is a calcification inhibitor [35]. Atherosclerotic and vascular calcification are closely linked to the vitamin K–dependent protein matrix γ-carboxyglutamate. Vitamin K antagonists, including warfarin, are associated with increased calcification of renal and other arteries [35,36]. Coronary artery calcification has been previously associated with a decline in eGFR [37]. Our results suggest that γ-carboxyglutamate increases arterial damage leading to a decline in eGFR.
We report three novel associations of N-lactoyl-amino acids (N-lactoylvaline, N-lactoylisoleucine, N-lactoylphenylalanine) with a decrease in eGFR. N-lactoylphenylalanine concentrations are increased in patients with phenylketonuria [38]. These patients have increased oxidative stress leading to tubulointerstitial disease, impaired kidney function, proteinuria, and arterial hypertension [39,40]. N-lactoylvaline and N-lactoylisoleucine have been found in the urine of a patient with maple syrup urine disease [41], which is associated with nephrotic syndrome [42].
We also found that the nucleoside 5-methyluridine (ribothymidine), an endogenous methylated nucleoside, decreased eGFR. This finding has been previously reported in rats with CKD [43]. Altered DNA methylation modulates the expression of pro-inflammatory and pro-fibrotic genes, stimulating renal disease progression [44]. High concentrations of homocysteine, hypoxia, and inflammation alter the epigenetic regulation of gene expression in CKD, impacting eGFR [44].
Eight of the 28 novel metabolites impairing eGFR were xenobiotics, chemical substances within an organism that are not naturally produced. Xenobiotics are food components, plant constituents, pesticides, industrial chemicals, environmental pollutants, or benzoate metabolites. An organic compound (S)-a-amino-omega-caprolactam is a uremic solute previously shown to impair kidney function [45]. 3-methyl catechol sulphate, a marker of current smoking and coffee consumption [46], decreased eGFR in our study. We also showed that genetic variants were not associated with xenobiotics suggesting that decreased eGFR is largely regulated also by lifestyle and environmental factors.
Our findings highlight multiple metabolic pathways associated with a decrease in eGFR. We identified 28 novel metabolites among amino acids, lipids, nucleotides, peptides, and xenobiotics associated with decreased eGFR. Eight xenobiotics were associated with a decrease in eGFR show that non-genetic factors, including benzoate pathway, food components, and plants play an important role in kidney dysfunction demonstrating the influence of environmental factors on eGFR. Additionally, the effects of N-lactoyl-amino acids and 5-methyluridine show a potential for epigenetic regulation of kidney function. Overall, our novel findings provide valuable insights into the complex biochemical interactions affecting kidney function and pave the way for future studies to explore metabolic pathways on kidney function in diverse populations.
The strength of our study is that the METSIM study is the largest randomly selected population-based cohort identifying metabolites associated with a decrease in eGFR applying the LC-MS/MS analysis method. Additionally, we followed our cohort for 12 years, and at baseline and at follow-up the metabolites identified were inversely associations with eGFR increasing the credibility of our findings. We applied a conservative statistical significance threshold in all analyses to obtain reliable conclusions. The limitations of our study are that our study included only middle-aged and elderly Finnish men. Therefore, the replication of our findings in non-European populations, including both men and women, is needed.
4. Materials and Methods
4.1. Study Population and Laboratory Measurements
The METSIM (METabolic Syndrome In Men) study includes 10 197 men, aged from 45 to 73 years at baseline, and randomly selected from the population register of Kuopio, Eastern Finland. The METSIM study was approved by the Ethics Committee of the Kuopio University Hospital, Finland. All participants provided written informed consent.
The design and methods of the METSIM study have been previously described in detail [47,48]. A total of 10 159 men were included in the current study, 3034 had normal glucose tolerance (NGT, fasting glucose < 6.1 mmol/l, 2-hour glucose < 7.8 mmol/l), 5 715 prediabetes [impaired fasting glucose (6.1-6.9 mmol/l) or impaired glucose tolerance (7.8 to 11.0 mmol/l) or both], and 1 410 T2D, [fasting glucose ≥ 7.0 mmol/l, or 2-hour glucose ≥ 11.1 mmol/l or glycated hemoglobin A1c (HbA1c) ≥ 6.5 %] according to the American Diabetes Association classification [49]. BMI was calculated as weight divided by height squared. Smoking status was defined as current smoking (yes/no). All participants excluding participants with T2D at baseline underwent a 2-hour oral glucose tolerance test (75 g of glucose), and samples for plasma glucose and insulin were drawn at 0, 30, and 120 minutes. Other laboratory measurements have been previously explained [47]. eGFR was calculated using the CKD-Epi equation [50].
4.2. Metabolomics
Non-targeted metabolomics profiling was performed at Metabolon, Inc. (Morrisville, NC, USA) on EDTA plasma samples obtained after overnight fasting, as previously described in detail [48,51]. We applied Liquid chromatography-mass spectrometry (LC-MS/MS) to identify the metabolites (the Metabolon DiscoveryHD4 platform). All samples were processed together for peak quantification and data scaling. We quantified raw mass spectrometry peaks for each metabolite using the area under the curve, and evaluated overall process variability by the median relative standard deviation for endogenous metabolites present in all 20 technical replicates in each batch. We adjusted for variation caused by day-to-day instrument tuning differences and columns used for biochemical extraction by scaling the raw peak quantifications to the median for each metabolite by the Metabolon batch.
4.3. Selection of genetic variants decreasing glomerular filtration rate
We identified genetic variants associated with a decrease in eGFR from previously published studies and from the GWAS Catalog (The NHGRI-EBI Catalog of human genome-wide association studies (https://www.ebi.ac.uk/gwas/) in the individuals of European ancestry. Altogether 117 genes were found to be associated with impaired eGFR.
4.4. Statistical Analysis
We conducted statistical analyses using IBM SPSS Statistics, version 29. We log-transformed all continuous variables except for age and follow-up time to correct for their skewed distribution. We performed association analyses between the eGFR and metabolites using linear regression adjusted for confounding factors (Model 1, adjustment for eGFR at baseline, Model 2, adjustment for eGFR at baseline, age, BMI, smoking, systolic blood pressure, fasting glucose, and total triglycerides). We give the results as standardized beta coefficients and p values with the metabolite as a dependent variable. We used one-way ANOVA to assess the differences in clinical traits and metabolites between the two groups at baseline. We applied the Bonferroni correction to determine statistical significance for the metabolites identified (p<5.0 x 10-5).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1, and Table S2.
Author Contributions
Conceptualization: LFS and ML; Methodology: AO, JV, LFS and ML; Investigation: LFS, JV, AO and ML; Visualization: LFS and AO; Funding acquisition: ML; Project administration: ML; Supervision: ML; Writing – original draft: LFS, JV, AO, and ML.
Funding
The research leading to these results has received support from the Innovative Medicines Initiative Joint Undertaking under grant agreements no. 115372 EMIF (to M.L), and no. 115974 BEAt-DKD (to M.L.). This Joint Undertaking received support from the European Union’s 7th Framework (EMIF) resp. Horizon 2020 (BEAt-DKD) research and innovation programmes and EFPIA, with JDRF (BEAt-DKD). Academy of Finland grant no. 321428 (ML). Centre of Excellence of Cardiovascular and Metabolic Diseases, the Academy of Finland grant no. 271961 (ML). Sigrid Juselius Foundation grant (ML). Finnish Foundation for Cardiovascular Research grant (ML). Kuopio University Hospital grant (ML).
Institutional Review Board Statement
The METSIM study was approved by the Ethics Committee of the Kuopio University Hospital, Finland. All participants provided written informed consent.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors, [ML] upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sekula, P.; Goek, O.-N.; Quaye, L.; Barrios, C.; Levey, A.S.; Römisch-Margl, W.; et al. A metabolome-wide association study of kidney function and disease in the general population. J. Am. Soc. Nephrol. 2016, 27, 1175-1188. [CrossRef]
- Levey, A.S.; Stevens, L.A.; Schmid, C.H.; Zhang, Y.; Castro, A.F.; Feldman, H.I.; et al. A new equation to estimate glomerular filtration rate. Ann. Intern. Med. 2009, 150, 604. [CrossRef]
- Baumeister, S.E.; Böger, C.A.; Krämer, B.K.; Döring, A.; Eheberg, D.; Fischer, B.; et al. Effect of chronic kidney disease and comorbid conditions on health care costs: A 10-year observational study in a general population. Am. J. Nephrol. 2010, 31, 222-229. [CrossRef]
- Kazancioğlu, R. Risk factors for chronic kidney disease: an update. Kidney Int. Suppl. 2013, 3, 368-371. [CrossRef]
- Wuttke, M.; Li, Y.; Li, M.; Sieber, K.B.; Feitosa, M.F.; Gorski, M.; et al. A catalog of genetic loci associated with kidney function from analyses of a million individuals. Nat. Genet. 2019, 51, 957-972.
- Wuttke, M.; König, E.; Katsara, M.-A.; Kirsten, H.; Farahani, S.K.; Teumer, A.; et al. Imputation-powered whole-exome analysis identifies genes associated with kidney function and disease in the UK Biobank. Nat. Commun. 2023, 14, 1287. [CrossRef]
- Gorski, M.; Rasheed, H.; Teumer, A.; Thomas, L.F.; Graham, S.E.; Sveinbjornsson, G.; et al. Genetic loci and prioritization of genes for kidney function decline derived from a meta-analysis of 62 longitudinal genome-wide association studies. Kidney Int. 2022, 102, 624-639. [CrossRef]
- Winkler, T.W.; Rasheed, H.; Teumer, A.; Gorski, M.; Rowan, B.X.; Stanzick, K.J.; et al. Differential and shared genetic effects on kidney function between diabetic and non-diabetic individuals. Commun. Biol. 2022, 5, 580. [CrossRef]
- Titan, S.M.; Venturini, G.; Padilha, K.; Tavares, G.; Zatz, R.; Bensenor, I.; et al. Metabolites related to eGFR: Evaluation of candidate molecules for GFR estimation using untargeted metabolomics. Clin. Chim. Acta 2019, 489, 242-248. [CrossRef]
- Lee, H.; Jang, H.B.; Yoo, M.-G.; Park, S.I.; Lee, H.-J. Amino acid metabolites associated with chronic kidney disease: An eight-year follow-up Korean epidemiology study. Biomedicines 2020, 8, 222. [CrossRef]
- Lee, S.; Han, M.; Moon, S.; Kim, K.; An, W.J.; Ryu, H.; et al. Identifying genetic variants and metabolites associated with rapid estimated glomerular filtration rate decline in Korea based on genome-metabolomic integrative analysis. Metabolites 2022, 12, 1139. [CrossRef]
- Peng, H.; Liu, X.; Aoieong, C.; Tou, T.; Tsai, T.; Ngai, K.; et al. Identification of metabolite markers associated with kidney function. J. Immunol. Res. 2022, 2022, 1-9. [CrossRef]
- Peng, H.; Liu, X.; Ieong, C.A.; Tou, T.; Tsai, T.; Zhu, H.; et al. A metabolomics study of metabolites associated with the glomerular filtration rate. BMC Nephrol. 2023, 24, 105. [CrossRef]
- Grams, M.E.; Tin, A.; Rebholz, C.M.; Shafi, T.; Köttgen, A.; Perrone, R.D.; et al. Metabolomic alterations associated with cause of CKD. Clin. J. Am. Soc. Nephrol. 2017, 12, 1787-1794. [CrossRef]
- Lin, B.M.; Zhang, Y.; Yu, B.; Boerwinkle, E.; Thygarajan, B.; Yunes, M.; et al. Metabolome-wide association study of estimated glomerular filtration rates in Hispanics. Kidney Int. 2022, 101, 144-151. [CrossRef]
- Schlosser, P.; Scherer, N.; Grundner-Culemann, F.; Monteiro-Martins, S.; Haug, S.; Steinbrenner, I.; et al. Genetic studies of paired metabolomes reveal enzymatic and transport processes at the interface of plasma and urine. Nat. Genet. 2023, 55, 995-1008. [CrossRef]
- Nierenberg, J.L.; He, J.; Li, C.; Gu, X.; Shi, M.; Razavi, A.C. Novel associations between blood metabolites and kidney function among Bogalusa Heart Study and Multi-Ethnic Study of Atherosclerosis participants. Metabolomics 2019, 15, 149. [CrossRef]
- Wang, F.; Sun, L.; Sun, Q.; Liang, L.; Gao, X.; Li, R.; et al. Associations of plasma amino acid and acylcarnitine profiles with incident reduced glomerular filtration rate. Clin. J. Am. Soc. Nephrol. 2018, 13, 560-568. [CrossRef]
- Kwan, B.; Fuhrer, T.; Zhang, J.; Darshi, M.; Van Espen, B.; Montemayor, D.; et al. Metabolomic markers of kidney function decline in patients with diabetes: evidence from the Chronic Renal Insufficiency Cohort (CRIC) Study. Am. J. Kidney Dis. 2020, 76, 511-520. [CrossRef]
- Lee, S.; Han, M.; Moon, S.; Kim, K.; An, W.J.; Ryu, H.; et al. Identifying genetic variants and metabolites associated with rapid estimated glomerular filtration rate decline in Korea based on genome-metabolomic integrative analysis. Metabolites 2022, 12, 1139. [CrossRef]
- Wen, D.; Zheng, Z.; Surapaneni, A.; Yu, B.; Zhou, L.; Zhou, W.; et al. Metabolite profiling of CKD progression in the chronic renal insufficiency cohort study. JCI Insight 2022, 7, e161696. [CrossRef]
- Bernard, L.; Zhou, L.; Surapaneni, A.; Chen, J.; Rebholz, C.M.; Coresh, J.; et al. Serum metabolites and kidney outcomes: The Atherosclerosis Risk in Communities Study. Kidney Med. 2022, 4, 100522. [CrossRef]
- Guo, X.; Peng, H.; Liu, P.; Tang, L.; Fang, J.; Aoieong, C.; et al. Novel metabolites to improve glomerular filtration rate estimation. Kidney Blood Press Res. 2023, 48, 287-296. [CrossRef]
- Au, A.Y.M.; Mantik, K.; Bahadory, F.; Stathakis, P.; Guiney, H.; Erlich, J.E.; et al. Plasma arginine metabolites in health and chronic kidney disease. Nephrol. Dial. Transplant. 2023, 38, 2767-2775. [CrossRef]
- Liu, J.-J.; Ching, J.; Wee, H.N.; Liu, S.; Gurung, R.L.; Lee, J.; et al. Plasma tryptophan-kynurenine pathway metabolites and risk for progression to end-stage kidney disease in patients with type 2 diabetes. Diabetes Care 2023, 46, 2223-2231. [CrossRef]
- Hou, Y.; Xiao, Z.; Zhu, Y.; Li, Y.; Liu, Q. Blood metabolites and chronic kidney disease: a Mendelian randomization study. BMC Med. Genomics 2024, 17, 147. [CrossRef]
- Luo, Y.; Zhang, W.; Qin, G. Metabolomics in diabetic nephropathy: Unveiling novel biomarkers for diagnosis. Mol. Med. Rep. 2024, 30, 156. [CrossRef]
- Wishart, D.S.; Guo, A.C.; Oler, E.; et al. HMDB 5.0: the Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622-D631. [CrossRef]
- Stelzer, G.; Rosen, R.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; et al. The GeneCards Suite: From gene data mining to disease genome sequence analysis. Curr. Protoc. Bioinform. 2016, 54, 1.30.1-1.30.33.
- Jellum, E.; Horn, L.; Thoresen, O.; Kvittingen, E.A.; Stokke, O. Urinary excretion of N-acetyl amino acids in patients with some inborn errors of amino acid metabolism. Scand. J. Clin. Lab. Investig. 1986, 184, 21-26.
- Engelke, U.F.; Sass, O.; Van Coster, R.N.; Gerlo, E.; Olbrich, H.; Krywawych, S.; et al. NMR spectroscopy of aminoacylase 1 deficiency, a novel inborn error of metabolism. NMR Biomed. 2008, 21, 138-147. [CrossRef]
- Okajima, K.; Inoue, M.; Morino, Y. Studies on the mechanism for renal elimination of N-acetylphenylalanine: Its pathophysiologic significance in phenylketonuria. J. Lab. Clin. Med. 1985, 105, 132-138.
- Sass, J.O.; Mohr, V.; Olbrich, H.; Engelke, U.; Horvath, J.; Fliegauf, M.; et al. Mutations in ACY1, the gene encoding aminoacylase 1, cause a novel inborn error of metabolism. Am. J. Hum. Genet. 2006, 78, 401-409. [CrossRef]
- Van Coster, R.N.; Gerlo, E.A.; Giardina, T.G.; Engelke, U.F.; Smet, J.E.; De Praeter, C.M.; et al. Aminoacylase I deficiency: A novel inborn error of metabolism. Biochem. Biophys. Res. Commun. 2005, 338, 1322-1326. [CrossRef]
- Luo, G.; Ducy, P.; McKee, M.D.; et al. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78-81. [CrossRef]
- Chatrou, M.L.; Winckers, K.; Hackeng, T.M.; Reutelingsperger, C.P.; Schurgers, L.J. Vascular calcification: the price to pay for anticoagulation therapy with vitamin K-antagonists. Blood Rev. 2012, 26, 155-166. [CrossRef]
- Budoff, M.J.; Rader, D.J.; Reilly, M.P.; Mohler, E.R., 3rd; Lash, J.; Yang, W.; et al. Relationship of estimated GFR and coronary artery calcification in the CRIC (Chronic Renal Insufficiency Cohort) Study. Am. J. Kidney Dis. 2011, 58, 519-526.
- Jansen, R.S.; Addie, R.; Merkx, R.; et al. N-lactoyl-amino acids are ubiquitous metabolites that originate from CNDP2-mediated reverse proteolysis of lactate and amino acids. Proc. Natl. Acad. Sci. USA 2015, 112, 6601-6606. [CrossRef]
- Hennermann, J.B.; Roloff, S.; Gellermann, J.; Vollmer, I.; Windt, E.; Vetter, B.; et al. Chronic kidney disease in adolescent and adult patients with phenylketonuria. J. Inherit. Metab. Dis. 2013, 36, 747-756. [CrossRef]
- Burton, B.K.; Bradford Jones, K.; Cederbaum, S.; Rohr, F. Prevalence of comorbid conditions among adult patients diagnosed with phenylketonuria. Mol. Genet. Metab. 2018, 125, 228-234. [CrossRef]
- Hagenfeldt, L.; Naglo, A.S. New conjugated urinary metabolites in intermediate type maple syrup urine disease. Clin. Chim. Acta 1987, 169, 77-83. [CrossRef]
- Maceda, E.B.G.; Abadingo, M.E.; Magbanua-Calalo, C.J.; Dator, M.A.; Resontoc, L.P.R.; Castro-Hamoy, L.; et al. Maple syrup urine disease associated with nephrotic syndrome in a Filipino child. BMJ Case Rep. 2021, 14, e242689. [CrossRef]
- Zhang, Z.H.; He, J.Q.; Qin, W.W.; Zhao, Y.Y.; Tan, N.H. Biomarkers of obstructive nephropathy using a metabolomics approach in rat. Chem. Biol. Interact. 2018, 296, 229-239. [CrossRef]
- Rysz, J.; Franczyk, B.; Rysz-Górzyńska, M.; Gluba-Brzózka, A. Are alterations in DNA methylation related to CKD development? Int. J. Mol. Sci. 2022, 23, 7108.
- Ganesan, L.L.; O'Brien, F.J.; Sirich, T.L.; Plummer, N.S.; Sheth, R.; Fajardo, C.; et al. Association of plasma uremic solute levels with residual kidney function in children on peritoneal dialysis. Clin. J. Am. Soc. Nephrol. 2021, 16, 1531-1538. [CrossRef]
- He, W.J.; Chen, J.; Razavi, A.C.; Hu, E.A.; Grams, M.E.; Yu, B.; et al. Metabolites associated with coffee consumption and incident chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2021, 16, 1620-1629. [CrossRef]
- Laakso, M.; Kuusisto, J.; Stančáková, A.; Kuulasmaa, T.; Pajukanta, P.; Lusis, A.J.; et al. The Metabolic Syndrome in Men study: a resource for studies of metabolic and cardiovascular diseases. J. Lipid Res. 2017, 58, 481-493. [CrossRef]
- Yin, X.; Chan, L.S.; Bose, D.; Jackson, A.U.; VandeHaar, P.; Locke, A.E.; et al. Genome-wide association studies of metabolites in Finnish men identify disease-relevant loci. Nat. Commun. 2022, 13, 1644. [CrossRef]
- Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2010, 33, S62-S69.
- Inker, L.A.; Eneanya, N.D.; Coresh, J.; Tighiouart, H.; Wang, D.; Sang, Y.; et al. New creatinine- and cystatin C-based equations to estimate GFR without race. N. Engl. J. Med. 2021, 385, 1737-1749. [CrossRef]
- Fernandes Silva, L.; Vangipurapu, J.; Kuulasmaa, T.; Laakso, M. An intronic variant in the GCKR gene is associated with multiple lipids. Sci. Rep. 2019, 9, 10240. [CrossRef]
Figure 1.
Venn diagram showing the number of the 100 most significant metabolites associated with eGFR, shared between the groups of normal glucose tolerance, prediabetes, and type 2 diabetes. .
Figure 1.
Venn diagram showing the number of the 100 most significant metabolites associated with eGFR, shared between the groups of normal glucose tolerance, prediabetes, and type 2 diabetes. .
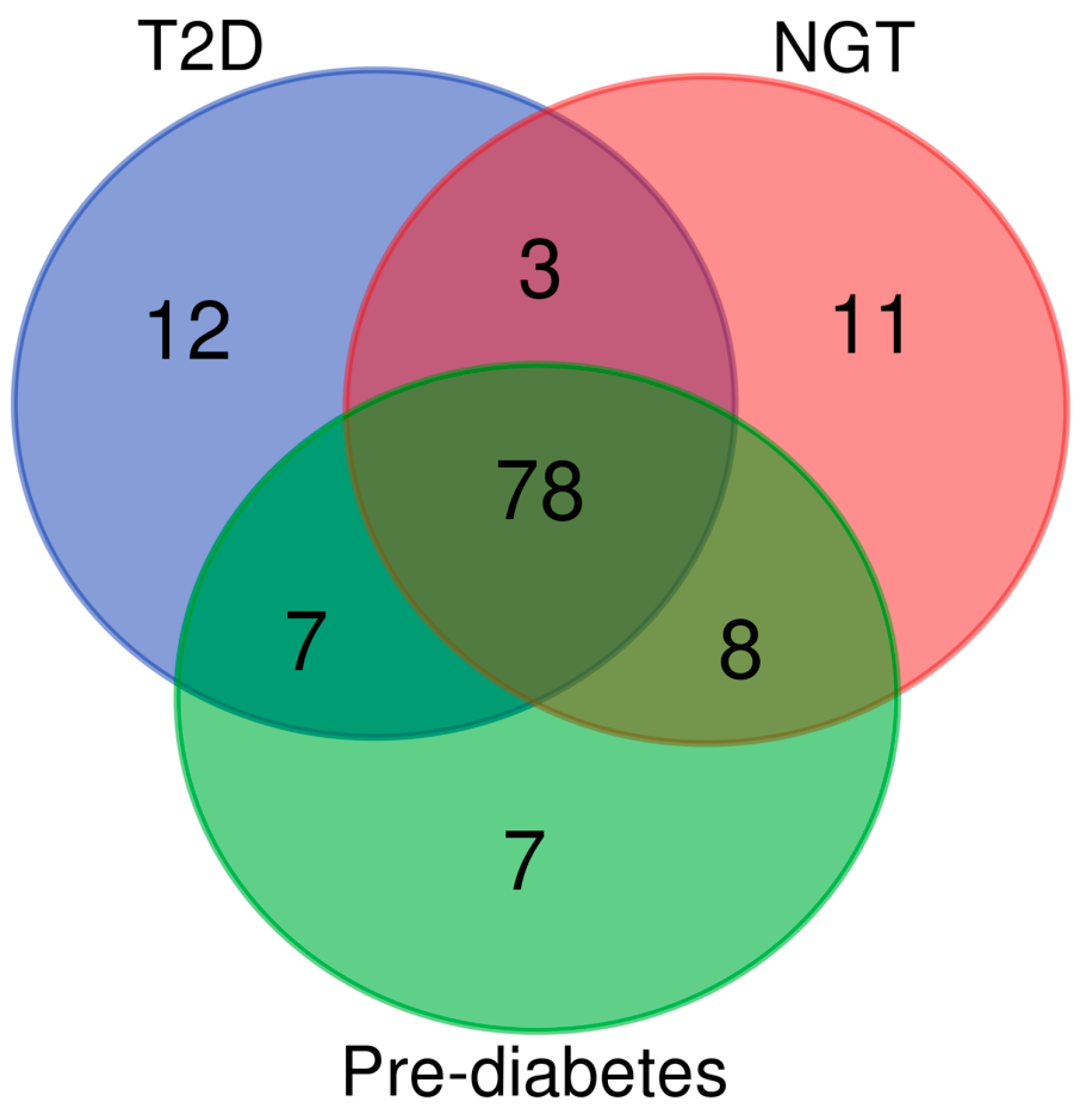

Table 1.
Baseline characteristics of the participants according to glucose tolerance.
| Measurements | NGT (n=3034) |
Prediabetes (n=5715) |
T2D (n=1410) |
p |
|---|---|---|---|---|
| Age (years) | 56.8 ± 6.9 | 57.4 ± 7.2 | 60.6 ± 6.7 | 1.1E-63 |
| Systolic blood pressure (mmHg) | 134.3 ± 15.9 | 138.7 ± 16.2 | 145.2 ± 18.1 | 2.1E-93 |
| Body mass index (kg/m2) | 25.8 ± 3.38 | 27.4 ± 3.9 | 30.2 ± 5.2 | 1.1E-247 |
| Current smoking (%) | 18.0 | 18.4 | 17.2 | 0.606 |
| Total triglycerides (mmol/l) | 1.22 ± 0.65 | 1.49 ± 1.08 | 1.90 ± 1.21 | 1.2E-143 |
| Fasting glucose (mmol/l) | 5.24 ± 0.24 | 5.97 ± 0.37 | 7.51 ± 2.01 | < 1E-250 |
| HbA1C (%) | 5.59 ± 0.31 | 5.71 ± 0.34 | 6.58 ± 1.13 | < 1E-250 |
| Fasting plasma insulin (mU/l) | 6.25 ± 4.11 | 9.32 ± 6.4 | 19.6 ± 28.5 | < 1E-250 |
| Creatinine (umol/l) | 84.6 ± 15.9 | 83.4 ± 12.8 | 84. 6 ± 22.3 | 0.0003 |
| eGFR (ml/min/1.73 m2) | 87.9 ± 12.3 | 88.6 ± 12.2 | 86.1 ± 14.5 | 4.5E-10 |
| Urine albumin (mg/l) | 18.4 ± 110.9 | 20.6 ± 82.5 | 93.5 ± 380.1 | 7.2E-181 |
| hs-CRP (mg/l) | 1.82 ± 2.96 | 2.13 ± 4.5 | 3.22 ± 6.07 | 3.4E-40 |
|
Abbreviations: NGT, normal glucose tolerance; T2D, type 2 diabetes; HbA1C, hemoglobin A1C; eGFR, estimated glomerular filtration rate; hs-CRP, high sensitivity C-reactive protein | ||||
Table 2.
Novel metabolites associated with a decrease in eGFR.
| Metabolite | Sub-class | N | Beta | p * | Beta | p ** |
|---|---|---|---|---|---|---|
| Amino acids | ||||||
| N-acetylmethionine |
Methionine, cysteine, taurine metab. | 7080 | -0.334 | 1.4E-183 | -0.087 | 5.5E-24 |
| N-acetylvaline | Leucine, isoleucine, valine metab. | 7082 | -0.343 | 1.0E-194 | -0.082 | 2.6E-21 |
| γ-carboxyglutamate | Glutamate metab. | 6929 | -0.295 | 1.1E-138 | -0.065 | 2.6E-14 |
| 3-methylglutaryl- carnitine (2) |
Leucine, isoleucine, valine metab. | 7001 | -0.257 | 1.1E-105 | -0.058 | 5.8E-12 |
| Proline | Urea cycle; arginine proline metab. | 7081 | -0.107 | 1.3E-19 | -0.048 | 3.9E-9 |
| Pro-hydroxy-pro | Urea cycle; arginine proline metab. | 7079 | -0.155 | 1.9E-39 | -0.047 | 5.2E-9 |
| 4-guanidinobutanoate | Guanidino acetamido metab. | 7049 | -0.158 | 1.7E-40 | -0.049 | 2.3E-9 |
| N-acetyltaurine | Methionine, cysteine, taurine metab. | 7048 | -0.208 | 1.4E-69 | -0.041 | 7.6E-7 |
| Hydantoin-5-propionate | Histidine metab. | 6154 | -0.211 | 3.6E-63 | -0.043 | 1.1E-6 |
| N-lactoyl valine | Lactoyl amino acid | 6781 | -0.182 | 2.5E-51 | -0.043 | 3.1E-6 |
| N-lactoylisoleucine | Lactoyl amino acid | 5437 | -0.189 | 4.4E-45 | -0.043 | 1.6E-5 |
| N-lactoyl phenylalanine | Lactoyl amino acid | 7033 | -0.233 | 2.7E-87 | -0.037 | 4.4E-5 |
| Lipids | ||||||
| 11beta-hydroxy etiocholanolone glucuronide |
Androgenic steroids | 4891 | -0.204 | 2.9E-47 | -0.050 | 4.0E-7 |
| 3-decenoylcarnitine | Fatty acid metab. | 5395 | -0.217 | 2.9E-58 | -0.042 | 9.2E-6 |
| Cis-3,4-methylene heptanoylglycine | Fatty acid metab. | 6825 | -0.161 | 5.2E-41 | -0.038 | 4.8E-6 |
| 2-methylmalonyl carnitine (C4-DC) |
Fatty acid metab. | 5827 | -0.235 | 8.0E-74 | -0.042 | 3.1E-6 |
| Propionylglycine | Fatty acid metab | 3960 | -0.119 | 4.9E-14 | -0.049 | 1.3E-5 |
| Nucleotide | ||||||
| 5-methyluridine(ribothymidine) | Pyrimidine metab. | 7082 | -0.134 | 6.8E-30 | -0.038 | 3.1E-6 |
| Peptide | ||||||
| Pyroglutamylvaline | Modified peptides | 6398 | -0.202 | 7.7E-60 | -0.051 | 2.6E-9 |
| Xenobiotics | ||||||
| 2,3-dihydroxyisovalerate | Food component/plant | 6998 | -0.206 | 3.8E-68 | -0.048 | 6.8E-9 |
| (S)-a-amino-omega-caprolactam | Food component/plant | 7007 | -0.296 | 1.3E-141 | -0.050 | 1.0E-8 |
| 3-methoxycatechol sulfate (2) | Benzoate metab. | 5379 | -0.185 | 2.0E-42 | -0.044 | 1.9E-6 |
| 3-methyl catechol sulfate (1) | Benzoate metab. | 7065 | -0.209 | 3.0E-70 | -0.040 | 2.1E-6 |
| 3-methoxycatechol sulfate (1) | Benzoate metab. | 6318 | -0.174 | 4.0E-44 | -0.039 | 5.5E-6 |
| 2-acetamidophenol sulfate | Food component/plant | 5939 | -0.153 | 2.9E-32 | -0.042 | 3.6E-6 |
| N-(2-furoyl)glycine | Food component/plant | 5025 | -0.235 | 5.0E-64 | -0.042 | 2.4E-5 |
| 2-aminophenol sulfate | Food component/plant | 7066 | -0.147 | 2.8E-35 | -0.036 | 1.1E-5 |
| Other metabolite | ||||||
| Glutamine_degradant | Partially characterized molecules | 7060 | -0.222 | 7.3E-80 | -0.071 | 2.2E-17 |
| p*: non-adjusted; p**: adjusted for eGFR at baseline, age, BMI, smoking, fasting glucose, total triglycerides and systolic blood pressure. | ||||||
Table 3.
The association of genetic variants of nine genes with novel metabolites association with a decline in eGFR the METSIM cohort.
Table 3.
The association of genetic variants of nine genes with novel metabolites association with a decline in eGFR the METSIM cohort.
| Gene-variant | Metabolite | p |
|---|---|---|
| KLHDC7B-rs470118 | 5-methyluridine | 9.9E-199 |
| CPS1-rs715 | Glycine | 8.1E-90 |
| AC007326.4-rs5992344 | Proline | 2.0E -63 |
| DOCK3- rs138144932 | N-acetylmethionine | 1.3E -44 |
| AOX1-rs7562507 | Hydantoin-5-propionate | 1.4E-17 |
| COLEC10-rs13264172 | Pro-hydroxy-pro | 3.5E-10 |
| MAGI1-rs264676 | 2.3-dihydroxy-5-methylthio-4-penenoate | 2.9E-8 |
| DCBLD2- rs192423025 | Pyroglutamylvaline | 3.4E-8 |
| CNTNAP2-rs533473709 | γ-carboxyglutamate | 5.3 E-8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



